

Abstract
Associative learning enables animals to predict rewards or punishments by their associations with predictive stimuli, while non-associative learning occurs without reinforcement. The latter includes latent inhibition (LI), whereby animals learn to ignore an inconsequential ‘familiar’ stimulus. Individual honey bees display heritable differences in expression of LI. We examined the behavioral and neuronal responses between honey bee genetic lines exhibiting high and low LI. We observed, as in previous studies, that high LI lines learned a familiar odor more slowly than low LI bees. By measuring gustatory responses to sucrose, we determined that perception of sucrose reward was similar between both lines, thereby not contributing to the LI phenotype. We then used extracellular electrophysiology to determine differences in neural responses of the antennal lobe (AL) to familiar and novel odors between the lines. Low LI bees responded significantly more strongly to both familiar and novel odors than the high LI bees, but the lines showed equivalent differences in response to the novel and familiar odors. This work suggests that some effects of genotype are present in early olfactory processing, and those effects could complement how LI is manifested at later stages of processing in brains of bees in the different lines.
Keywords: latent inhibition, Apis mellifera, antennal lobe, proboscis extension reflex (PER), extracellular recording
Introduction
Understanding the link between genetics and learning is crucial for uncovering the sources of individual variation in behavior. The interactions between genetics, stimulus integration and expression of behavior are complex. Learning behaviorally relevant stimuli is important as animals can then utilize their environment more effectively. In particular learning to pay less attention to irrelevant stimuli, known as Latent Inhibition (LI) (Lubow and Moore 1959; Lubow 1973), is also an important form of plasticity. LI occurs when familiarization to a repeated, unreinforced stimulus reduces subsequent learning of the familiar stimulus. One theoretical interpretation of LI proposes that it could be due to loss of attention to familiar, unimportant stimuli. Alternatively, acquisition could occur normally, but that association is poorly expressed in behavior (reviewed in (Pearce and Bouton 2001)). In either interpretation, expression of LI can differ among individuals of any species. Variation in expression of LI has been documented in animal taxa as diverse as mice, cats, rabbits and honey bees, indicating that inter-individual variation in this trait may be ubiquitous across most animal taxa (Ginton et al. 1975; Lubow and Gewirtz 1995; Chandra et al. 2000).
In the present study, we use honey bees as a model to examine the potential sources of variation in LI phenotypes. Honey bees occupy a special role in the history of behavioral and neurobiological studies, and in many ways, they are ideal for studying the variation of LI expression in sensory systems due to their rich behavioral repertoire. Variation in LI exists among individual foragers in a honey bee colony (Chandra et al. 2001), and the ecological relevance of this variation has recently been examined (Cook et al. 2020). On average, foraging scouts exhibit high LI and recruits exhibit low LI (Cook et al. 2018). Thus, it may be adaptive for a colony to have phenotypic variation in LI to maximize efficiency of foraging. This variation in LI is at least partially explained by heritable factors and sensory context (Chandra et al. 2001; Chandra et al. 2010), but it is unknown how early sensory processing contributes to the LI phenotype. It is possible that variation in LI expression could be due to differences in gustatory and olfactory senses. Studies have shown that sucrose perception (gustatory response) can be shaped by many factors, for example, including foraging experience and genotype (Page Jr et al. 1998; Pankiw et al. 2001; Moauro et al. 2018). A recent study showed that waggle-dance followers showed increased gustatory responses versus non-followers (Moauro et al. 2018). Furthermore, pollen foragers (Page Jr et al. 1998) and strains of pollen-hoarding genetic lines (Pankiw et al. 2001) have weaker gustatory responses than nectar foragers. It is as yet unknown if low or high LI bees differ in sucrose perception.
Neural mechanisms underlying LI have been studied using a variety of techniques such as pharmacological manipulation, direct lesions or cytochrome oxidase histochemistry (Clark et al. 1992; Puga et al. 2007; Weiner and Feldon 1997). These studies have identified parts of the brain that display neural activities correlating with the expression of LI in vertebrates. In honey bees, there is evidence that LI may be manifested in early olfactory processing in the AL of honey bees (Locatelli et al. 2013), which consists of 162 spherical neuropils called glomeruli (Winnington et al. 1996). Glomeruli are sites of synaptic interactions among various types of neurons including the output neurons or projection neurons (PNs), local neurons (LNs) and sensory neurons. One recent study using calcium imaging on PNs from glomeruli revealed that familiarization to a particular odor – a procedure that is known to generate latent inhibition to that odor behaviorally (Chandra et al. 2001; Chandra et al. 2010) - indeed altered the processing of a mixture containing the familiar odor and another novel odor (Locatelli et al. 2013). Here, we use lines of honey bees artificially selected for high and low LI behavior to further investigate how the genetic background affects odor-driven AL responses and sucrose-driven gustatory responses.
Methods
Rearing LI lines
In order to generate forager honey bees with specific heritable LI phenotypes, queens and drones were tested for LI (Chandra et al. 2001). Queens were then inseminated by sperm from a single drone exhibiting the same high or low LI phenotype as the queen. LI phenotype was assessed using the LI behavioral assay described below. If queens or drones showed slower learning of the familiar odor, they were designated as high LI bees. Alternatively, if acquisition was unaffected by the familiarization, they were designated low LI (as in (Chandra et al. 2001)). Colonies were reared from these queens thereby producing workers of known LI phenotypes. Emerging bees were marked on their thorax with paint to distinguish their line of origin, so they could be collected and identified approximately 3 weeks later when they became foragers. Lines were maintained on the Arizona State University campus in Tempe, AZ and all experiments were conducted in the March-April 2018.
LI behavioral assay
Marked non-pollen foragers (n= 88 high LI; n= 70 low LI) were collected by placing a wire mesh over the entrance of the hive and capturing incoming foragers with a glass vial. While inside the vials, bees were placed on ice for 3–5 minutes to immobilize them. Using a thin strip of duct tape, bees were strapped between the head and thorax to a plastic harness. Bees were then fed 5μl of 1 M sucrose solution and allowed to acclimate for 1 hour before the assay. Similar to previous studies testing LI in honey bees (Chandra et al. 2010), the LI assay consisted of a familiarization phase which contains 40 4-second unreinforced repeated-exposures to an odor, occurring with an inter-trial interval of 5 minutes. Odors were delivered using a custom built PER system (Smith and Burden 2014); a solenoid-controlled, flow-meter-regulated air stream through a 5 cc glass syringe containing a strip of filter paper with 3 μl of pure odor. The familiarization phase is then followed by a conditioning phase using PER (proboscis extension reflex) to evaluate responses to the familiar odor and a novel odor (Chandra et al. 2010; Smith and Burden 2014; Cook et al. 2018). The repeated odor that was used in the familiarization phase serves as the familiar stimulus in the conditioning phase, and the second odor is considered as the novel odor. During the conditioning trials, a 2 M sucrose reward was paired with odor stimulation. To ensure the repeated odor to be perceived as familiar odor regardless of its chemical identity, we counterbalanced 1-hexanol (Hex) or 2-octanone (Oct) as the novel or familiar odor, and switched odors between experiments to control for any existing preferences to either odor. In other words, the expressed differences during the conditioning trials are only based on the familiarity to the odor. The inter-trial interval during the conditioning phase was approximately ten minutes.
Gustatory response
To determine if LI lines differed in their gustatory responsiveness to sucrose (Pankiw et al. 2001; Page Jr et al. 1998), we measured PER to different concentrations of sucrose touching their antennae (n= 87 high LI, n= 90 low LI). We controlled for forager type by not selecting pollen foragers, because it is known that pollen foragers have lower gustatory responses than nectar foragers (Page Jr et al. 1998). We followed the gustatory response protocol described by (Scheiner et al. 2013), by diluting sucrose solutions with distilled water to a series of molar concentrations: 0.01, 0.1, 1, 3, 10 and 30 M. Between sucrose exposures, antennae were touched with water to control for sensitization of antennae and inhibiting appetitive learning from the antennae making contact with the tongue (Scheiner et al. 2013). Gustatory response data were binomial, either the bee responded by exhibiting PER or they did not (1 or 0). The inter-trial interval was approximately 10 minutes between sucrose or water exposures.
Electrophysiology in the AL
Multichannel extracellular-recording methods (Lei et al. 2004; Strube-Bloss et al. 2012) were used to measure responses of the AL in LI assays. The LI assay took approximately two hours, and we achieved eight stable recordings from high and low LI bees (n= 25 units from 3 high LI bees and 5 low LI bees). Bees were harnessed in a similar way as described in (Strube-Bloss et al. 2012). Briefly, bees were cold-anaesthetized before being harnessed on a custom-made Plexiglass block. The bee was immobilized on the block with low-melting-temperature utility wax, along with the bee’s antennae at the joints using Eicosane. A rectangular window was cut open between the two compound eyes and between the ocelli and the base of the two antennae. The glandular material and some trachea membrane were removed to expose the two ALs. To prevent the bee’s head from pumping hemolymph and proboscis from moving, the clypeus was punctured and the muscle at the base of proboscis was severed. Eicosane was used to reseal the clypeus. The bee was positioned on a recording stage in such a way that the anterior side of the brain was facing upward.
A constant saline flow was introduced into the head capsule to ensure the brain tissue was immersed in an adequate ionic environment. A standard bee saline recipe was adopted from literature (Galizia and Vetter 2004), which contains 130 mM NaCl, 6 mM KCl, 4 mM MgCl2 × 6H2O, 5 mM CaCl2 × 2H2O, 160 mM Sucrose, 25 mM D-Glucose × H2O, 10 mM HEPES. The pH was adjusted to 6.7 and the final osmolarity was 500 mOsm. A reference electrode, made of a thin silver wire, was placed behind the brain in the saline water.
A Tucker-Davis Technologies (TDT) RZ2 microprocessor system, in conjunction with a PZ2 preamplifier from the same manufacturer (TDT®, Alachua, FL 32615 USA) was used to digitize neural signals, which was sampled with a Neuronexus A2×2 multichannel probe (Neuronexus®, Ann Arbor, MI 48108 USA), as described in (Lei et al. 2004). Briefly, the probe was carefully inserted into the central neuropil of the AL aided with a Leica micromanipulator. After initial contact, the probe was slowly moved deeper, a few microns each step, until spikes appeared. Care was taken to insert probes approximately in the same location each time. The acquisition software was configured to acquire the spike waveforms at 25 kHz sampling rate, and the 16 recording channels were grouped in the software to form 4 tetrodes (Gray et al. 1995), facilitating the separation of units in later analysis.
To quantify the neural responses, the tetrode waveforms were exported from the TDT acquisition software to the Offline Sorter program (Plexon® Inc., Dallas, TX USA), which allows automatic as well as manual sorting of the waveforms. In Offline Sorter, each set of tetrode waveforms, composed of 32 (A/D points) × 4 (recording sites) thus 128 dimensions, was reduced to 3 dimensions using principal component analysis, and each concatenated waveform was projected onto a 3D space consisting of the 3 principal components for visualization (Fig. 3A, B). An advantage of the tetrode setting arises in its expanded power to discriminate subtle differences among waveforms (Gray et al. 1995). In general, K-means automatic sorting or semi-automatic sorting using templates was first applied, then visual inspection and adjustment were taken to yield multiple units from each tetrode. Offline Sorter assumes stable waveforms but see (Rey et al. 2015) for exceptions. The quality of spike sorting was statistically verified within Offline Sorter. The time stamps of all waveforms were then exported to a spike analysis program, Neuroexplorer ® (Nex Technologies, Dallas, TX USA) or Matlab ® (Mathworks, Natick, MA USA) for further analysis. The example in Fig. 3a–b shows separations of 4 units from a tetrode verified with multivariate ANOVA (In the PCA 3D space, F (18, 197965) = 4.149, p=6.95e-6). Units are treated as individual neurons in the antennal lobe. We did not attempt to classify the neuron types as described in (Meyer et al. 2013) due to relatively small sample size and consideration of unknown effect of LI lines on spiking physiology.
Fig. 3.
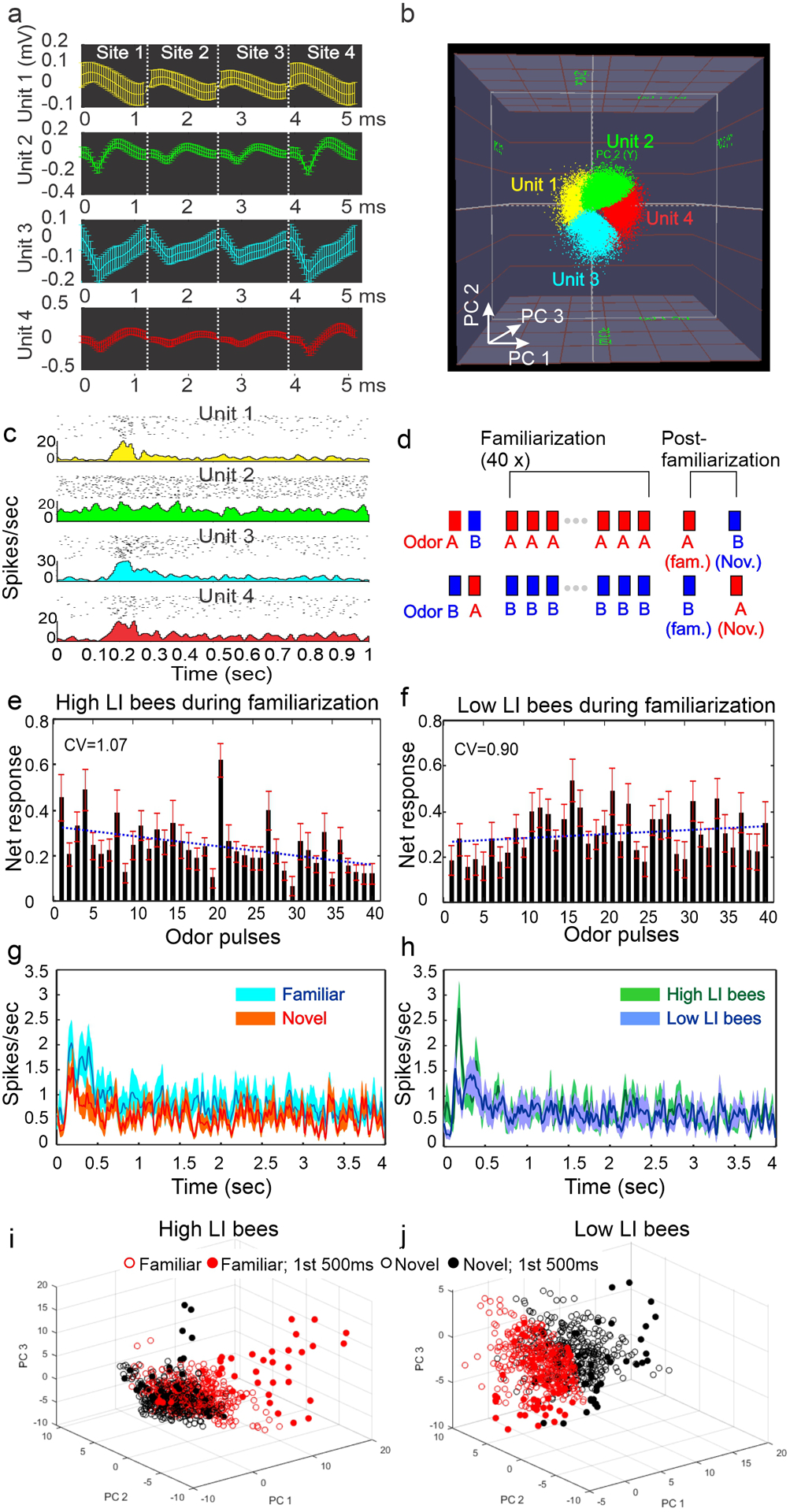
Neural responses to familiar and novel odors. Individual units were separated from extracellularly-recorded waveforms (a) based on their distribution in the 3D principle-component space (b). The responses of the 4 units in (a) are shown with their raster plots (40 stimulations) and peristimulus time histograms (PSTH) (c). Unit 2 did not produce an obvious response. Familiarity (or novelty) to odor A or B was introduced by stimulating the animal repeatedly 40 times with odor A or B (inter-pulse interval = 5 min; 4 sec duration) without any reward or punishment during the familiarization phase, and later tested during the post-familiarization phase (d). The averaged net responses (mean ± SE) over the 40x repeated stimulation in the high LI bees (e) and low LI bees (f) show a large amount of trial-to-trial variation. The net response within a unit (n=13 in high LI bees; n=12 in low LI bees) was normalized to the maximum of the 40 responses. The blue dotted lines represent 1st degree polynomial regression. CV: coefficient of variation. In Panel (g), the PSTHs (mean ± SE) from all AL units regardless the type of bees show stronger response to the familiar odor (cyan) than to the novel odor (red) after familiarization. Similarly, the PSTHs from the high LI bees (green) show a higher transient response shortly after odor stimulation than the low LI bees (blue) (h). In both panels, odor onset was at time zero and stimulation lasted for 4 sec; the spike traces were binned at 10 msec resolution and the curves were Gaussian filtered with a moving window of 3 bins. Principle component analysis revealed that the ensemble dynamics to the familiar and novel odor stimulation in high LI bees were largely overlapped in the entire 4 sec of stimulation window, but showed apparent separation during the first 500 msec (filled red circles vs. filled black circles) (i). In low LI bees, the separation was less pronounced (j).
Similarly, to the behavioral protocol, we familiarized bees to either Hex or Oct with 40 repeated stimulations during the familiarization phase without any reward. Prior to and after familiarization, bees were briefly presented with two 4-sec pulses of both odors with 5 min interval between the two pulses. Thus, both odors were new to the bees prior to familiarization, but one odor was considered as novel after the familiarization to the other odor. During the familiarization phase, the inter-pulse interval was 5 min. The entire procedure was performed while a multichannel probe was inserted in the AL.
Statistical Methods
Behavioral assays were analyzed using R (R Core Team 2019) and RStudio (Version 1.1.463). We performed a logistic regression using a generalized linear model (glm) with a binomial error distribution and a logit link to compare how the LI lines differed in LI learning behavior and gustatory response. For the LI assay, we compared how the different lines respond to familiar or novel odors across trials. Our response variable was whether the bee responded, quantified as a 1 or a 0. In the LI assay, our predictor variables were the LI line, the odor (familiar or novel), the trial (1–4), as well as the secondary and tertiary interactions of each of these variables. To assess gustatory response, we compared how the high and low LI bees responded to sucrose concentration. We used a reverse-model selection approach by removing the most complex, insignificant interactions to identify the simplest but most explanatory model. To further explore these effects, we used a Tukey posthoc test using the CRAN package emmeans (estimated marginal means) function.
After spike sorting as described above, based on the peristimulus time histograms of all odor-evoked responses, a 2 sec response window was chosen, which was sufficiently wide to cover the rising phase and recovery phase of all responses, and was consistently used for all units. The response window starts 150 ms after the onset of solenoid valve opening (i.e. Time = 0 sec), taking into account the odor traveling time in the delivery tube. A net response was calculated as: net response = (number of spikes in response window – number of spikes in pre-response window of the same length) / the latter quantity. This ratio metric was chosen in order to normalize the quantification across individual units, which have variable levels of baseline activities. The data was then transformed to Gaussian distribution using Box-Cox power transformation. The results of transformation were visually inspected using histograms and Q-Q plots.
To examine how neural responses are affected by genetic lines and familiar/novel odors, we performed general linear mixed-effect (glme) modeling analysis with normal distribution, identify link function and MPL fitting method, and with the response term being the net response after familiarization (i.e. post-netresp) and a predictor being the net response before familiarization (i.e. pre-netresp). Additionally, the high/low LI line (i.e. bee type), familiar/novel odor (i.e. odors) and their interaction (i.e. bee type × odors) are also categorical predictors. This model also accounts for possible non-independence among the units recorded from the same brain by factoring in random-effect terms that group the predictors per animal (i.e. prep id). Coefficient of each term is tested against the null hypothesis: coef = 0 (Table 1).
Table 1.
Fixed-effects coefficients (95% confidence intervals) derived from generalized linear mixed-effect modeling
| Model terms | Estimate | SE | t Statistic | DF | p Value | Lower | Upper |
|---|---|---|---|---|---|---|---|
| Intercept | 1.1751 | 0.3056 | 3.8451 | 42 | 0.0004 | 0.5584 | 1.7918 |
| Odors_novel | −0.6147 | 0.3520 | −1.7466 | 42 | 0.0880 | −1.3250 | 0.0955 |
| Bee type_low LI | 1.0279 | 0.4390 | 2.3417 | 42 | 0.0240 | 0.1421 | 1.9137 |
| Transformed pre-net-response | 0.2271 | 0.1517 | 1.4972 | 42 | 0.1418 | −0.0790 | 0.5332 |
| Odors x Bee type | −0.7258 | 0.4927 | −1.4731 | 42 | 0.1481 | −1.7202 | 0.2685 |
Principle component analysis was performed on the units from high LI and low LI bees in the ensemble responses (Fig.3i and Fig.3j). The principle components integrate information from peri-stimulus time histograms of all units in each LI line, thus capturing the characteristics of ensemble evolution in response to familiar and novel odor in the two LI lines.
Results
LI behavioral assay
We constructed the learning curves of both LI lines to the familiar and novel odors after the familiarization phase (Fig. 1). Less than 10% of bees exhibited PER on the 1st trial, but up to 80% showed the behavioral response to novel odor on the 4th trial (dotted lines, Fig. 1). The two LI lines (different symbols, Fig. 1) displayed very similar learning curves to the novel odor (dotted lines, Fig. 1), but they differed in responsiveness to the familiar odor (solid lines, Fig. 1), which shows LI.
Fig. 1.
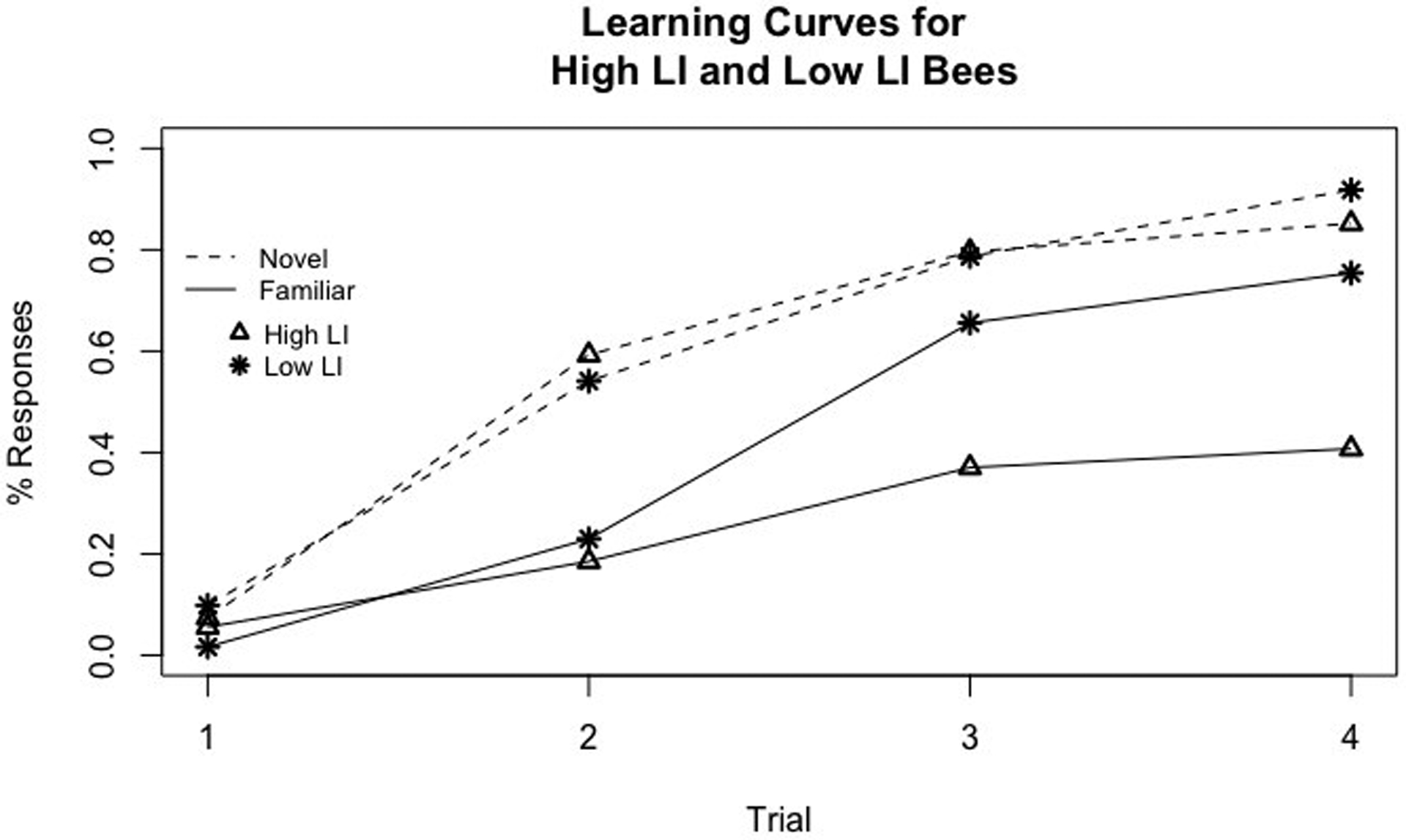
Learning curves of LI phenotypes to a familiar and novel odor after the LI assay. Percent PER response over four trials are shown. Dashed lines represent the novel odor, while solid lines represent the familiar odor. Stars indicate low LI responses and triangles represent high LI responses
A multi-factor analysis using the generalized linear model showed that honey bees from the different lines learned the novel and familiar odors differently, as evidenced by the significant ‘line X odor’ interaction term (n=88 high LI, 70 low LI, LI line X odor, χ2 = 4.12, df=1, p=0.04). Consistent with previous studies (Chandra et al. 2000; Cook et al. 2018), high LI individuals responded significantly less overall to the familiar odor (effect size= 1.41, Z=6.63, p < 0.0001). Even though low LI lines, too, learned the odors differently, the high LI line treated the novel and familiar odor more differently than the low LI line (effect size = 0.08, z=4.13, p < 0.0001).
Gustatory response
We tested the hypothesis that LI lines differed in their response to concentrations of sucrose using a generalized linear model (Fig. 2). We found no statistical difference in gustatory responsiveness between the LI lines at any sucrose concentration (χ2= 2.7, df=6, p= 0.84). Although, both LI lines had higher responses with increasing concentration of sucrose (χ2= 335.09, df= 6, p< 0.0001). At the lowest concentration, gustatory response was approximately 10% and at the highest concentration gustatory response was approximately 60% (Fig. 2).
Fig. 2.
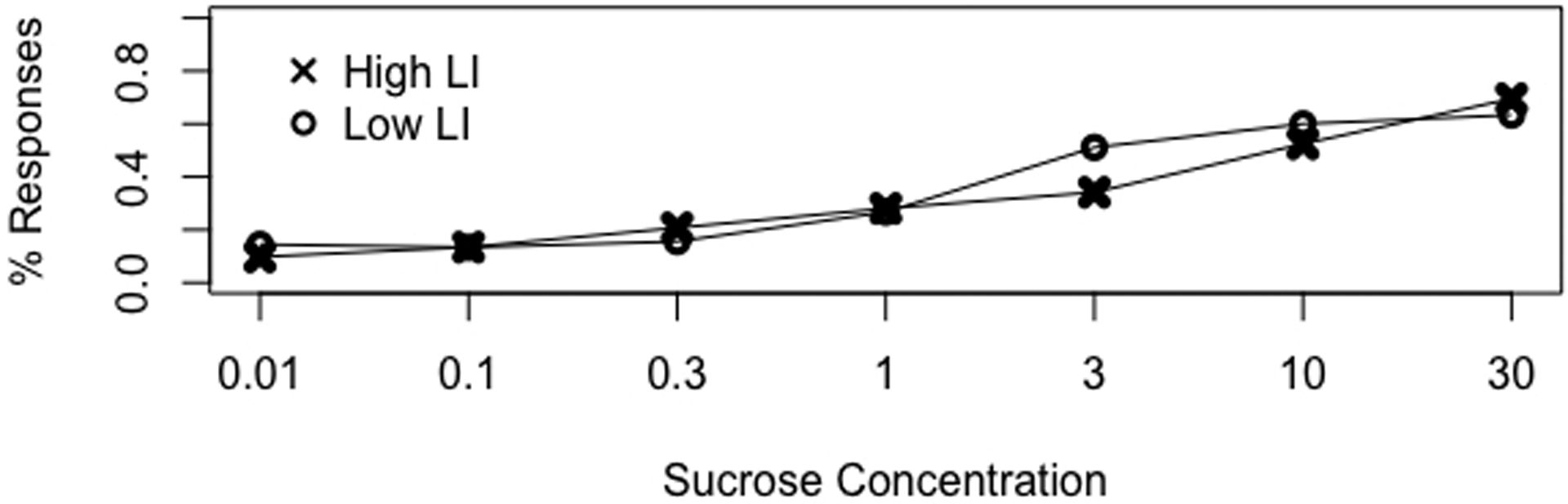
Responses to sucrose concentration between LI lines. Proportion of PER response to concentrations of sucrose from 0.01 molar to 30 molar solutions are shown. High LI responses are represented by “X” and low LI responses by “O”
Neural responses in the AL
We first asked whether the spontaneous activities changed before and after the process of familiarization for both LI lines. We quantified the number of spikes in the 2 sec window in response to solvent-control stimulation. In low LI bees, 10 out of 13 units decreased the number of spikes after familiarization with an averaged decrement of 56%. In high LI bees, only 7 out of 12 showed a decrease and the rest showed an increase with an averaged increment of 6%. This result indicated that the spontaneous activities were not stable over the course of the experiment. Thus, the net response (the difference between just prior to response onset and just after the response onset expressed as a ratio) was calculated to quantify the odor-evoked responses.
We then examined response changes over the 40 trials during the familiarization phase (Fig. 3d). In both genetic lines, the 40x repeated stimulation did not result in a consistent change in response across trials, in terms of either a consistent decrease or an increase (Fig. 3e–f). The response magnitude fluctuated from trial to trial, although there was a slight decreasing tendency for the high LI bees (y=−0.004*x+0.33; R2=0.20) and slight increasing tendency for the low LI bees (y=0.002*x+0.27; R2=0.04). Additionally, the coefficient of variation was slightly higher in the high LI bees than in the low LI bees (CV=1.07 vs. 0.90), suggesting the responses in the low LI bees were more consistent over the 40 consecutive odor pulses.
We next asked whether the responses to familiar and novel odors before and after familiarization were different in the AL of high and low LI lines. The familiarization protocol (Fig. 3d) allowed us to compare the responses to familiar and novel odors in the two genetic lines once the familiarization phase was complete. To visualize the responses of AL units after familiarization, we plotted the peristimulus time histograms (PSTH) (mean ± S.E.) using instantaneous frequencies derived from the responses to familiar and novel odors of all units (n=25) regardless of the high and low genetic line (Fig. 3g). The familiar odor generally produced stronger overall responses than the novel odor through the stimulation period. This result inversely correlates with the observation from the PER assay, especially at Trial 2, where both LI lines showed stronger responses to the novel odor (Fig. 1). We then replotted the same data for high versus low genetic line, now regardless of odor identity. The AL responses are similar for most of the stimulation period, except for a brief transient increase in the high LI bees within the first few hundred milliseconds after onset of odor stimulation (Fig. 3h).
To capture the characteristics of ensemble evolution in response to familiar and novel odor in both LI lines, we performed principle component (PC) analysis on the PSTHs derived from each unit from each LI line to both odors. The number of units constituted the original dimensions, which were reduced to three PCs for visualization (Fig. 3i–j). In this PC space, the time axis is represented by the trajectories of data points. In the high LI bees (Fig. 3i), the ensemble dynamics evoked by familiar and novel odors were indistinguishable for most of the 4 sec window except for the first 500 msec (as in Fig. 3h), when the ensemble dynamics of the familiar odor (filled red circles) were apparently separate from that of the novel odors (filled black circles). A similar phenomenon was observed in the low LI bees (Fig. 3j), although the corresponding time sections occupied different regions in the PC space. Therefore, the largest difference in ensemble responses to novel and familiar odor occurred in the early phase of the AL responses in both LI lines.
Next, we compared the change of responses to familiar and novel odors caused by the familiarization process in both genetic lines through general linear mixed-effect modeling (Table 1; Fig. 4). Among the fixed-effect terms, the genetic lines contribute significantly to the model output (t statistic = 2.3417, p = 0.024, Table 1). Its estimated coefficient of 1.0279 suggests that the low LI bees have higher averaged net response by a factor of 1.0279 compared to the high LI bees (see Fig.4a left two columns for group averages), assuming all other terms are kept constant. Odors (the term ‘Odors_novel’) have a marginal effect on the model output (t statistic = −1.7466, p = 0.088, Table 1; see Fig.4a right two columns for group averages). Interestingly, this term has a negative coefficient (estimate = −0.6147, Table 1), suggesting stimulation with novel odor decreases the net response by a factor of 0.6147 on average, compared to stimulation with familiar odor. No significant interaction between the genetic lines and type of odors was detected in the model (t statistic = −1.4731, p = 0.1482, Table 1; Fig.4b), indicating a lack of dependence of odor response on the type of bees. This observation differs from the behavioral observation where the odor-associated difference does depend on bee lines. In the random-effect terms, none of their coefficients is significantly different from zero (t test, p>0.05), suggesting that there is no evidence for a significant correlation among the units that were recorded from the same brain.
Fig. 4.
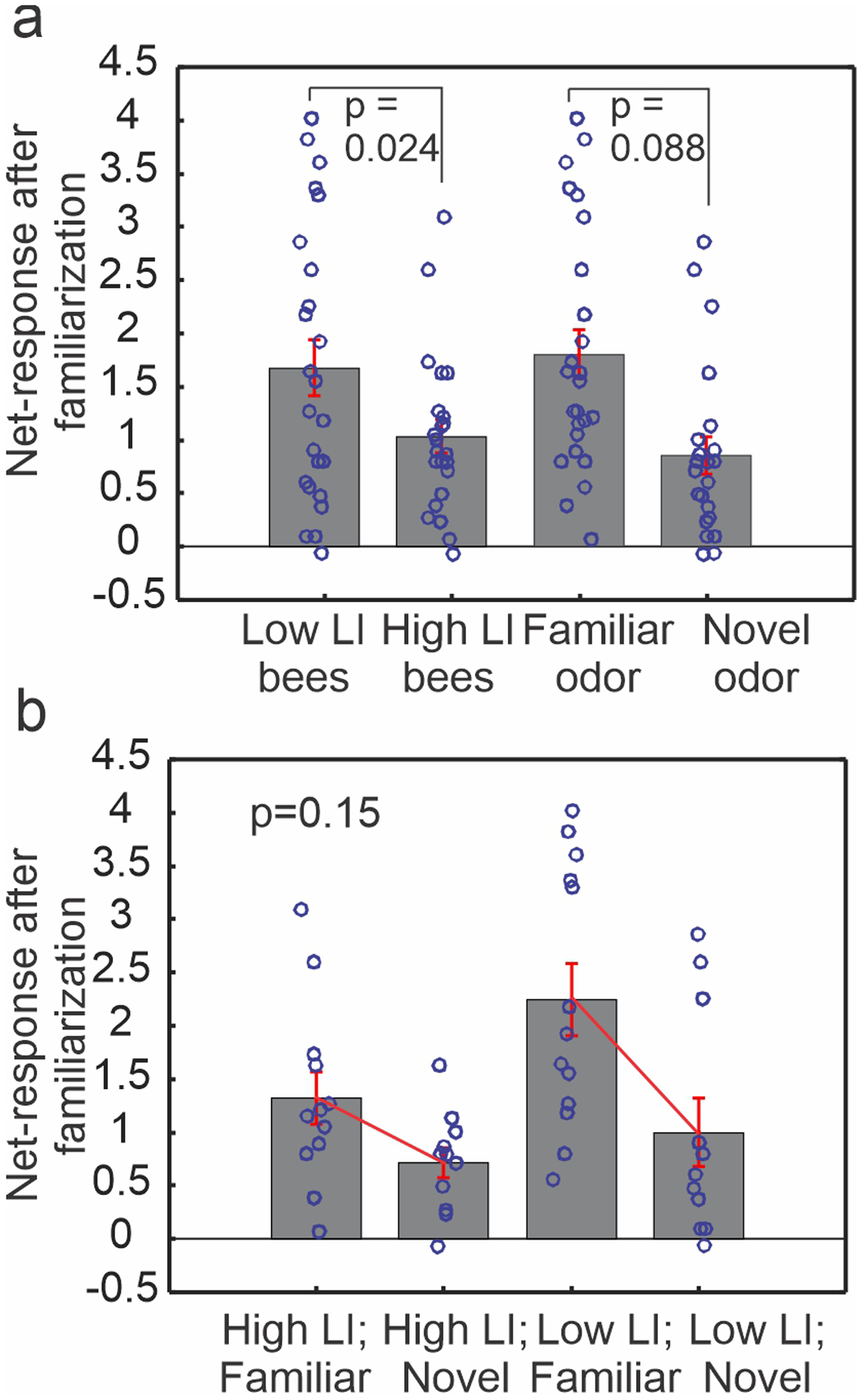
Familiarization-induced responses to familiar and novel odors in high and low LI bees, analyzed with general linear mixed-effect modeling. The averaged post-net-responses (mean ± SE; Cox-Box transformed) are plotted to compare the contributions of the fixed-effect terms to the model output. The p values indicate whether the coefficient of that term is significantly different from zero in the generalized linear model (see Methods). (a) Type of bees is a significant predictor (p=0.024, t test) with the low LI bees responding more strongly than the high LI bees; type of odors is a marginal predictor (p=0.088, t test) with the familiar odor eliciting stronger response than the novel odor. (b) The contribution of the interaction term (Odors X Bees) to the model is not significant (p=0.15, t test), indicating the response difference between familiar and novel odor was not dependent on the type of bees, evident from the nearly parallel lines across the bar graphs (mean ± SE).
Discussion
In this study, we examined how heritable differences in LI affect sensory processing in honey bee foragers. Behaviorally, we observed high LI bees to be more strongly inhibited in learning about a familiar odor than bees from the low LI line. These results are consistent with other studies where the LI phenotypes are screened by artificially training the bees to familiar and novel odors (Chandra et al. 2000). We hypothesized that both gustatory and olfactory sensory-processing could contribute to the LI phenotypes. However, our data show these two sensory modalities differ in whether they contribute to LI.
High and low LI lines do not show difference in sucrose response
Gustatory responses were not different between the high and low LI lines. Studies have shown that foraging experience and genetics can affect sucrose perception. For example, pollen foragers have lower sucrose response thresholds than nectar foragers (Page Jr et al. 1998). This was also true in pollen-hoarding strains of honey bees (Pankiw et al. 2001). Responses to lower concentrations of sucrose are hypothesized to hold a higher value to that type of bee. Because low and high LI bees differ in their acquisition of familiar odors, we wanted to determine if sucrose perception could be contributing to this effect. We found no difference in sucrose perception between the line; high and low LI bees responded similarly to low (0.01 mol/l) through high (30 mol/l) concentrations of sucrose (Fig. 2). Our results therefore suggest it is unlikely high and low LI bees perceive sucrose differently; instead, odor-based neuronal responses in AL could contribute to LI behavior.
Differences between neural and behavioral responses
We found the response in the AL was significantly different between genetic lines, however not in a way that would account completely for LI. After familiarization, bees from the low LI line showed stronger responses on average to both the novel and familiar odors across the entire stimulation period than bees from the high LI line. There was a difference in the temporal properties of the responses, with a short spike in responsiveness in the high LI bees early in the stimulation period. It is possible that this short transient increase is meaningful for the way the brain process odor information. However, this short spike was not prolonged enough to overcome the more consistent higher responsiveness in the low LI bees during odor stimulation. There was a non-significant trend (p<0.088) for differences in responses to the novel and familiar odor when pooling bees from the two lines, and there was not a significant interaction between the lines in these responses. Thus, whatever difference in response there was to the two types of odors, there was no difference in this effect across lines (Fig. 4A–B).
In contrast, behaviorally there is a significant interaction between line and odor, indicating that differences in learning about familiar and novel odors depended on the genetic lines – high LI bees learned the familiar odor slower than the low LI bees (Fig. 1). This discrepancy between neurophysiology and behavior could potentially be explained by different stages of olfactory information processing. LI expressed in behavior is the final output of a series of neural mechanisms that most likely involve circuitries in the AL, mushroom bodies, lateral horn and other high-order olfactory centers in the protocerebrum. The AL neurons may only contribute to the early coding of the familiarity and novelty, emphasizing after experience, for example, the presence of an odor, and whether it is familiar or novel. This response is then interpreted by higher processing centers to yield different behavioral responses in high and low LI bees. When the behavioral conditions in which LI was encoded arise, including contexts from other sensory modalities such as vision (Chandra et al. 2010), they may interact to fine tune responses in the different lines. Perhaps then, the heritable differences in responsiveness in the high and low lines to familiar and novel odors arise at a stage of processing after the AL.
Potential neural mechanisms underlying the LI in the antennal lobes
The changes in responsiveness to the two odors, with higher responses to the novel than to the familiar odor (Fig 4a right two bars), may be due to interactions among different types of interneurons in the AL. In spite of not being able to identify units as projection neurons (PN) or local neurons (LN), there is marginal difference between the responses to the familiar and novel odor regardless of LI line. Furthermore, when tested with the familiar odor in the PER assay, the low LI bees learned the new association (i.e. familiar odor paired with sucrose reward) more efficiently than the high LI bees. This result is also consistent with the electrophysiological data where the AL units in the low LI line produced a stronger response to the both the novel and familiar odors (Fig. 4B). The stronger response to the latter in low vs high LI bees could trigger stronger acquisition of the odor sucrose association at later processing stages in the brain.
Although neuron types are not classified in this study, an earlier study from our group used calcium imaging to record activity specifically from PNs (Locatelli et al. 2013). They showed that PN’s responsiveness to the familiar and novel odors did not change after familiarization when the two odors were tested separately (Locatelli et al. 2013). However, PN responses to the mixture of the two odors became more similar to the responses to novel odor, suggesting that a novel odor can more strongly overshadow a familiar odor in a mixture. This model predicted that a population of inhibitory LN-PN synapses is strengthened by the familiarization process but do not exert their effect on PNs until these LNs are activated by the presence of the novel odor. Experience dependent plasticity at the synapses implemented in the model has been shown in work on odor habituation in fruit flies (Das et al. 2011). Therefore, the LNs that interconnect a large array of glomeruli are probably more subjected to modulation by familiarization, allowing the novel odor to outcompete the familiar odor in the mixture representation.
In this study, although we did not test mixtures, we observed marginally significant differences between familiar and novel odor-evoked responses when they were tested separately (t test, p=0.088, Fig. 4A), which initially seems to contradict imaging study (Locatelli et al. 2013). However, we should note here two caveats. First, the temporal resolution of the calcium imaging method may not pick up slight but behaviorally significant changes in PN responses. Furthermore, that method images glomeruli, which contain up to 5 PNs, which further limits the resolution. Electrophysiology can pick up slight changes because of the faster temporal resolution of the signal. Therefore, changes in PN responses might still be resolved via faster imaging methods or through electrophysiology of identified cell types. Second, Locatelli et al. specifically sampled pollen foragers for their study (Locatelli et al. 2013). Pollen foragers could express higher LI than nectar foragers (Latshaw and Smith 2005), but they may not do so under all conditions (Drezner-Levy et al. 2009). Further studies, perhaps combining imaging with electrophysiology using LI phenotypes, are needed to now resolve these questions.
As we have noted above, it is possible that LI could be more effectively engaged at a later stage of processing, such as the mushroom body. Hatorri et al. reported in the fruit fly that a subset of mushroom body output neurons (MBONs) in the α′3 compartment of the mushroom body suppress responses to familiar odor but maintain their ability to respond to novel odor. This transition in neural activity upon familiarization requires odor-evoked activity in the dopaminergic neuron innervating this compartment (Hattori et al. 2017). It is possible that similar mechanisms operate in the honeybee mushroom bodies. The LI expression level may affect how dopaminergic neurons regulate the MBONs. In high LI bees, for example, the response sensitivity to familiar odor may be kept low for a longer period of time compared with in the low LI bees.
While LI is most likely a property of central circuits, our experiments cannot exclude the possibility that the selected LI lines may exhibit differential peripheral responses. In fact, the observed differences of AL responses between the two LI lines could be derived from the periphery. Worth noting, however, is that the familiarization process itself has no effect on the electroantennogram response (Locatelli et al. 2013).
In summary, our data confirmed the LI behavioral phenotypes in learning to respond to novel and familiar odors, and we found a lack of contribution from the gustatory system. The selection of high and low LI genetic lines resulted in different responsiveness in the AL to familiarization. However, it seems likely that the stereotypical learning behaviors in high and low LI lines cannot be completely explained by differences in plasticity in the AL. Other higher-order olfactory circuits most likely play important roles in driving the LI learning behaviors.
Acknowledgements
We would like to thank undergraduates Eda Sezen and Alexa Phillips for their help collecting behavioral data; and Drs. Patel, Iaci and Navas Zuloaga for assistance in statistics. Furthermore, we greatly appreciate the work by Cahit Ozturk for doing the artificial insemination of queens and maintaining the learning lines. This research is funded by National Science Foundation (1556337) and National Institute of Health (NIGMS) (GM113967) to BHS, NIH NIGMS (F32GM126728) to CNC, and a subaward to BHS as part of the NSF/CIHR/DFG/FRQ/UKRI-MRC Next Generation Networks for Neuroscience Program.
Funding
This research is funded by National Science Foundation (1556337) and National Institute of Health (NIGMS) (GM113967) to BHS; NIH NIGMS (F32GM126728) to CNC; a subaward to BHS as part of the NSF/CIHR/DFG/FRQ/UKRI-MRC Next Generation Networks for Neuroscience Program.
Footnotes
Conflicts of interest/Competing interests
The authors report no competing interests.
Availability of data and material
References
- Chandra SBC, Hosler JS, Smith BH (2000) Heritable variation for latent inhibition and its correlation with reversal learning in honeybees (Apis mellifera). J Comp Psychol 114 (1):86–97. doi: 10.1037/0735-7036.114.1.86 [DOI] [PubMed] [Google Scholar]
- Chandra SBC, Hunt GJ, Cobey S, Smith BH (2001) Quantitative trait loci associated with reversal learning and latent inhibition in honeybees (Apis mellifera). Behav Genet 31 (3):275–285. doi: 10.1023/A:1012227308783 [DOI] [PubMed] [Google Scholar]
- Chandra SBC, Wright GA, Smith BH (2010) Latent inhibition in the honey bee, Apis mellifera: Is it a unitary phenomenon? Anim Cogn 13 (6):805–815. doi: 10.1007/s10071-010-0329-6 [DOI] [PubMed] [Google Scholar]
- Clark AJM, Feldon J, Rawlins JNP (1992) Aspiration lesions of rat ventral hippocampus disinhibit responding in conditioned suppression or extinction, but spare latent inhibition and the partial reinforcement extinction effect. Neuroscience 48 (4):821–829. doi: 10.1016/0306-4522(92)90270-C [DOI] [PubMed] [Google Scholar]
- Cook CN, Lemanski NJ, Mosqueiro T, Ozturk C, Gadau J, Pinter-Wollman N, Smith BH (2020) Individual learning phenotypes drive collective behavior. Proc Natl Acad Sci USA:201920554. doi: 10.1073/pnas.1920554117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cook CN, Mosqueiro T, Brent CS, Ozturk C, Gadau J, Pinter‐Wollman N, Smith BH (2018) Individual differences in learning and biogenic amine levels influence the behavioural division between foraging honeybee scouts and recruits. J Anim Ecol 88 (2):9. doi: https://doi-org.ezproxy1.lib.asu.edu/10.1111/1365-2656.12911 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das S, Sadanandappa MK, Dervan A, Larkin A, Lee JA, Sudhakaran IP, Priya R, Heidari R, Holohan EE, Pimentel A, Gandhi A, Ito K, Sanyal S, Wang JW, Rodrigues V, Ramaswami M (2011) Plasticity of local GABAergic interneurons drives olfactory habituation. Proc Natl Acad Sci USA 108 (36):E646–E654. doi: 10.1073/pnas.1106411108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drezner-Levy T, Smith BH, Shafir S (2009) The effect of foraging specialization on various learning tasks in the honey bee (Apis mellifera). Behav Ecol Sociobiol 64 (1):135. doi: 10.1007/s00265-009-0829-z [DOI] [Google Scholar]
- Galizia CG, Vetter R (2004) Optical methods for analyzing odor-evoked activity in the insect brain. In: Christensen TA (ed) Advances in Insect Sensory Neuroscience. CRC Press, Boca Raton, pp 349–392 [Google Scholar]
- Ginton A, Urca G, Lubow RE (1975) The effects of preexposure to a nonattended stimulus on subsequent learning: Latent inhibition in adults. Bull Psych Soc 5 (1):5–8. doi: 10.3758/BF03336682 [DOI] [Google Scholar]
- Gray CM, Maldonado PE, Wilson M, McNaughton B (1995) Tetrodes markedly improve the reliability and yield of multiple single-unit isolation from multi-unit recordings in cat striate cortex. J Neurosci Met 63 (1):43–54. doi: 10.1016/0165-0270(95)00085-2 [DOI] [PubMed] [Google Scholar]
- Hattori D, Aso Y, Swartz KJ, Rubin GM, Abbott LF, Axel R (2017) Representations of novelty and familiarity in a mushroom body compartment. Cell 169 (5):956–969.e917. doi: 10.1016/j.cell.2017.04.028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Latshaw JS, Smith BH (2005) Heritable variation in learning performance affects foraging preferences in the honey bee (Apis mellifera). Behav Ecol Sociobiol 58 (2):200–207. doi: 10.1007/s00265-004-0904-4 [DOI] [Google Scholar]
- Lei H, Christensen TA, Hildebrand JG (2004) Spatial and temporal organization of ensemble representations for different odor classes in the moth antennal lobe. J Neurosci 24:11108–11119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Locatelli FF, Fernandez PC, Villareal F, Muezzinoglu K, Huerta R, Galizia CG, Smith BH (2013) Nonassociative plasticity alters competitive interactions among mixture components in early olfactory processing. Eur J Neurosci 37 (1):63–79 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lubow RE (1973) Latent inhibition. Psychol Bull 79 (6):398–407. doi: 10.1037/h0034425 [DOI] [PubMed] [Google Scholar]
- Lubow RE, Gewirtz JC (1995) Latent inhibition in humans: Data, theory, and implications for schizophrenia. Psychol Bull 117 (1):87–103. doi: 10.1037/0033-2909.117.1.87 [DOI] [PubMed] [Google Scholar]
- Lubow RE, Moore AU (1959) Latent inhibition: The effect of nonreinforced pre-exposure to the conditional stimulus. J Comp Physiol Psychol 52 (4):415–419. doi: 10.1037/h0046700 [DOI] [PubMed] [Google Scholar]
- Meyer A, Galizia CG, Nawrot MP (2013) Local interneurons and projection neurons in the antennal lobe from a spiking point of view. J Neurophysiol 110 (10):2465–2474. doi: 10.1152/jn.00260.2013 [DOI] [PubMed] [Google Scholar]
- Moauro MA, Balbuena MS, Farina WM (2018) Assessment of appetitive behavior in honey bee dance followers. Front Behav Neurosci 12 (74). doi: 10.3389/fnbeh.2018.00074 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Page RE Jr, Erber J, Fondrk MK (1998) The effect of genotype on response thresholds to sucrose and foraging behavior of honey bees (Apis mellifera L.). J Comp Physiol A 182 (4):489–500. doi: 10.1007/s003590050196 [DOI] [PubMed] [Google Scholar]
- Pankiw T, Waddington KD, Page RE (2001) Modulation of sucrose response thresholds in honey bees (Apis mellifera L.): influence of genotype, feeding, and foraging experience. J Comp Physiol A 187 (4):293–301. doi: 10.1007/s003590100201 [DOI] [PubMed] [Google Scholar]
- Pearce JM, Bouton ME (2001) Theories of associative learning in animals. Ann Rev Psychol 52 (1):111–139. doi: 10.1146/annurev.psych.52.1.111 [DOI] [PubMed] [Google Scholar]
- Puga F, Barrett DW, Bastida CC, Gonzalez-Lima F (2007) Functional networks underlying latent inhibition learning in the mouse brain. NeuroImage 38 (1):171–183. doi: 10.1016/j.neuroimage.2007.06.031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- R Core Team (2019) R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria [Google Scholar]
- Rey HG, Pedreira C, Quian Quiroga R (2015) Past, present and future of spike sorting techniques. Brain Research Bulletin 119:106–117. doi: 10.1016/j.brainresbull.2015.04.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheiner R, Abramson CI, Brodschneider R, Crailsheim K, Farina WM, Fuchs S, Grünewald B, Hahshold S, Karrer M, Koeniger G, Koeniger N, Menzel R, Mujagić S, Radspieler G, Schmickl T, Schneider CW, Siegel AJ, Szopek M, Thenius R (2013) Standard methods for behavioural studies of Apis mellifera. J Api Res 52:58 [Google Scholar]
- Smith BH, Burden CM (2014) A proboscis extension response protocol for investigating behavioral plasticity in insects: Application to basic, biomedical, and agricultural research. JoVE (91):51057. doi: 10.3791/51057 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strube-Bloss MF, Herrera-Valdez MA, Smith BH (2012) Ensemble response in mushroom body output neurons of the honey bee outpaces spatiotemporal odor processing two synapses earlier in the antennal lobe. PLoS ONE 7 (11):e50322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiner I, Feldon J (1997) The switching model of latent inhibition: an update of neural substrates. Behav Brain Res 88 (1):11–25. doi: 10.1016/S0166-4328(97)02314-0 [DOI] [PubMed] [Google Scholar]
- Winnington AP, Napper RM, Mercer A (1996) Structural plasticity of identified glomeruli in the antennal lobes of the adult worker honey bee. J Comp Neurol 365:479–490 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.