

Highlights
-
•
The endothelial glycocalyx thins with age and cardiovascular comorbidities.
-
•
Endothelial glycocalyx is affected by and integral to severe pediatric illnesses.
-
•
Mechanistic insight into cause/effect of endothelial glycocalyx injury is paramount.
-
•
Vascular glycocalyx damage in pediatric critical illness warrants further study.
Abbreviations: ACE2, angiotensin-converting enzyme 2; CD, cell differentiation marker; COVID-19, coronavirus disease 2019; CPB, cardiopulmonary bypass; CT, component therapy; DENV NS1, dengue virus nonstructural protein 1; DM, diabetes mellitus; ECLS, extracorporeal life support; ECMO, extracorporeal membrane oxygenation; EG, endothelial glycocalyx; eNOS, endothelial nitric oxide synthase; FFP, fresh frozen plasma; GAG, glycosaminoglycan; GPC, glypican; HPSE, heparanase; HSV, herpes simplex virus; IV, intravenous; MIS-C, multisystem inflammatory syndrome in children; MMP, matrix metalloproteinase; Pragmatic, Randomized Optimal Platelet and Plasma Ratios; RHAMM, receptor for hyaluronan-mediated motility; S protein, spike protein; SAFE, Saline versus Albumin Fluid Evaluation; SARS-CoV-2, severe acute respiratory syndrome coronavirus 2; SDC, syndecan; SDF, sidestream darkfield; SIRT1, sirtuin 1; TBI, traumatic brain injury; TBSA, total body surface area; Th2, type 2 helper T cell; TMPRSS2, transmembrane protease serine 2; VSMC, vascular smooth muscle cell; WB+CT, whole blood and component therapy
Keywords: Children, Critical illness, Endothelial glycocalyx, Vascular biology
Abstract
The vascular endothelium is the interface between circulating blood and end organs and thus has a critical role in preserving organ function. The endothelium is lined by a glycan-rich glycocalyx that uniquely contributes to endothelial function through its regulation of leukocyte and platelet interactions with the vessel wall, vascular permeability, coagulation, and vasoreactivity. Degradation of the endothelial glycocalyx can thus promote vascular dysfunction, inflammation propagation, and organ injury. The endothelial glycocalyx and its role in vascular pathophysiology has gained increasing attention over the last decade. While studies characterizing vascular glycocalyx injury and its downstream consequences in a host of adult human diseases and in animal models has burgeoned, studies evaluating glycocalyx damage in pediatric diseases are relatively few. As children have unique physiology that differs from adults, significant knowledge gaps remain in our understanding of the causes and effects of endothelial glycocalyx disintegrity in pediatric critical illness. In this narrative literature overview, we offer a unique perspective on the role of the endothelial glycocalyx in pediatric critical illness, drawing from adult and preclinical data in addition to pediatric clinical experience to elucidate how marked derangement of the endothelial surface layer may contribute to aberrant vascular biology in children. By calling attention to this nascent field, we hope to increase research efforts to address important knowledge gaps in pediatric vascular biology that may inform the development of novel therapeutic strategies.
Introduction
Children who present with critical illness have life-threatening single or multi-organ injury from bodily insults ranging from congenital anomalies to severe infection to trauma. The vascular endothelium has an integral role in mediating immune system interactions with end-organs during severe diseases by regulating immune cell trafficking, vascular permeability, and coagulation activation. The glycan-rich glycocalyx lining the luminal surface of blood vessels is the body’s first line of defense against circulating pathogens and inflammatory mediators while also coordinating ligand-endothelial receptor interactions, leukocyte/platelet-endothelial interactions, vasoreactivity, and coagulation. Though the endothelial glycocalyx (EG) is receiving greater attention as an integral player in a host of adult and pediatric pathophysiologies [1], the processes driving EG injury and the role of EG damage in varying diseases remain unresolved.
Relative to adults, very little is known about the EG in pediatric critical illness. A recent review has nicely summarized the current state of the literature characterizing glycocalyx damage in pediatric diseases [2]. Arguably, each of the 14 studies reported in that review are, or can be, related to pediatric critical illness. However, the relatively low number of studies and limited scope of EG characterization in children highlights the nascency of this topic of study. Given the significant role that the EG likely has in the development of life-threatening diseases in children, a more thorough understanding of the interplay between EG injury and organ dysfunction occurring during severe pediatric illnesses may inform novel treatment approaches that could substantially improve outcomes.
The factors that dictate EG integrity during pediatric critical illness include the baseline glycocalyx expression, severity of the inflammatory response, endothelial cell response, and treatments deployed (Fig. 1). In this narrative review of the literature, we first highlight the impact of age maturation, immune system development, and pediatric comorbidities on baseline EG health. Next, we delve into the role of EG injury in particular broad categories of pediatric critical care medicine and, conversely, how current treatment strategies for each category may impact EG integrity. Finally, we touch on the challenges faced in studying the EG in critically ill children and identify areas for future translational research. It is our hope that bringing more attention to this area of pediatric health will encourage research efforts to fill important knowledge gaps and foster the discovery of successful treatments for severely ill children.
Fig. 1.

Impact of blood vessel aging, immune system development, and comorbidities on endothelial glycocalyx (EG) integrity in children with critical illness. As children develop, their vasculature adapts to progressively more elevated blood pressures and a cumulative exposure to inflammatory insults that may thin the EG. The immune system similarly matures and becomes more responsive to a broader range of pathogens; however, this may increase the threat to EG integrity at baseline and during acute systemic inflammation. Children may also develop comorbidities (e.g., chronic hypoxemia from a congenital heart or lung defect, diabetes mellitus, chronic kidney disease) that can increase basal levels of inflammation or alter the vascular stiffness that further threaten EG health at baseline. Then, during a severe systemic illness, the host inflammatory response (and potentially the treatment employed) acutely perturbs the child’s EG. Therefore, EG integrity in critically ill children is affected by the complex interaction of baseline blood vessel and immune system maturity, presence of comorbidities, the acute inflammatory response, and treatment strategies. The degree of EG disintegrity during the child’s critical illness then contributes to the vascular pathobiology that leads to organ injury and adverse outcomes.
The endothelial glycocalyx in children
Endothelial glycocalyx structure and function
We provide a brief synopsis of what is currently known about glycocalyx composition and physiology to set the stage for discussions around EG pathobiology in critically ill children. Extrapolating from rodent studies, we have learned that glycocalyx size varies along the vascular tree with thickness increasing proportionally to vessel diameter [3], [4], [5]. The EG is comprised mainly of highly sulfated, negatively charged glycosaminoglycans, or GAGs, (e.g., heparan sulfate and chondroitin sulfate) attached to endothelial surface-anchored proteoglycans. The two major families of proteoglycans found at the endothelial surface are syndecans (SDC) and glypicans (GPC). There are four SDC isoforms (i.e., SDC1-4), each a transmembrane molecule with an endodomain attached to the cellular cytoskeleton and an ectodomain to which 3–5 heparan sulfate molecules (all SDCs) attach near the N-terminus (furthest from cell surface) and 2 chondroitin sulfate molecules (SDC1 only) attach near the cell membrane [6]. The GPC family consists of six molecules (i.e., GPC1-6) with GPC1 widely accepted as the only member represented on the endothelial surface [6], [7]. GPCs are anchored to the cell membrane by a C-terminal glycosylphosphatidylinositol linkage and have three to four heparan sulfate molecules attached to the core ectodomain [7]. Hyaluronan is the largest EG GAG and remains in close association with the glycocalyx by binding to cell surface hyaladherins (e.g., cell differentiation marker, or CD, 44 and receptor for hyaluronan-mediated motility, or RHAMM) and intercalating between heparan sulfate proteoglycans [8], [9]. The anionic charge and coiled structure of hyaluronan molecules facilitates water molecule association with the glycocalyx that maintains its viscosity.
The glycocalyx has an important role in a myriad of endothelial cell functions. It serves as a physical barrier to circulating cell interactions with the endothelial surface [10] and protects surface glycoproteins such as cell adhesion molecules and selectins from indiscriminate leukocyte and platelet attachment to the endothelial surface. Heparan sulfate proteoglycans serve as co-factors for cognate receptor activation by local and circulating cytokines such as fibroblast growth factor [11], [12]. The viscous mesh-like structure supports the storage of growth factors and cytokines secreted from the endothelium, which are liberated to serve as signaling molecules primarily following EG injury. Coagulation mediators such as antithrombin, tissue factor pathway inhibitor, and heparin cofactor III are also sequestered in the EG and contribute to the maintenance of an anticoagulant phenotype of the endothelial surface. The negatively charged heparan sulfate molecules within the glycocalyx deflect circulating proteins from the endothelial surface, helping maintain an oncotic gradient between intra- and extravascular spaces that limits fluid extravasation [13]. Finally, proteoglycans and GAGs play a critical role in mechanotransduction of surface shear stress to intracellular signaling pathways within the endothelium that help maintain endothelial homeostasis and vascular tone [14], [15], [16].
Impact of age maturation
The EG likely undergoes important changes throughout childhood that parallel other vascular developments and could impact disease manifestation in a critically ill child. In the fetus, the EG is likely present at the onset of blood flow and is instrumental in directing normal angiogenesis [17]. Though the mechanisms regulating EG expression in fetu are unknown, it is doubtless that this intricate process is as fine-tuned as vasculogenesis and angiogenesis [18], [19]. Observations by Puchwein-Schwepcke et al. [20] suggest that fetal EG thickness gradually decreases as gestation advances, most especially before 30 weeks gestation. The authors also found that EG thicknesses measured in extremely premature infants at a corrected gestational age of 33 weeks were significantly less than the EG of infants born at 33 weeks gestation, pointing to postnatal environmental factors as important modifiers of EG health that may predispose extremely premature infants to later cardiovascular disease [21], [22] and endothelial dysfunction [23], [24].
By the time a child reaches school age, the vasculature begins maturing toward an adult phenotype [25] of increased arterial stiffness [26], decreased vascular reactivity [27], and attenuated microvascular density [28], [29]. While it is currently unknown how EG expression evolves over childhood, adult and preclinical data suggest that age-related EG expression parallels other changes in blood vessel characteristics such as arterial stiffness. Indirect measures of human EG thickness using sublingual videomicroscopy demonstrate that older adults have a significantly thinner microvascular EG compared to younger adults (gender-matched comparison) [28]. Similarly, serum SDC1 levels decline with advanced age in adult humans [30]. Preclinical studies support these findings, demonstrating age-dependent EG thinning in male C57BL/6 mice [28] and diminished GPC1 expression on the aortae of older (28–32 weeks) wild-type mice compared to young (6–8 weeks) mice [31]. Human blood outgrowth endothelial cells harvested from 20 to 30 year olds enzymatically stripped of surface heparan sulfate produced a chemotactic phenotype similar to that of untreated endothelial cells from 50 to 70 year olds [32], suggesting that, like SDC1 and GPC1, heparan sulfate expression in the EG appears to diminish with age. Finally, gene expression of hyaluronan synthase isoform 2, associated with EG expression of high molecular weight hyaluronan [33], is significantly less in the aortae of old (23–27 months) C57BL/6 mice compared to younger (4–6 months) C57BL/6 mice [28], pointing to an attenuated expression of high molecular weight hyaluronan in the EG with advancing age.
Though the mechanisms linking aging and EG expression remain unclear, several pathways have been implicated in this complex relationship. As humans advance in age, endothelial cells cumulate exposure to reactive oxygen species and other harmful stimuli that drive them toward premature senescence [34]. In vitro studies demonstrate that senescent endothelial cells express a thinner glycocalyx and a more contractile phenotype [35], both of which can impair endothelial barrier function. Moreover, as endothelial cells become senescent, expression of the nicotinamide adenine dinucleotide-dependent histone deacetylase sirtuin 1 (SIRT1) falls [36], which may potentiate glycocalyx thinning [37]. In a vicious positive feedback loop, loss of EG volume from reduced SIRT1-mediated synthesis may then further contribute to premature endothelial senescence [38] and vascular dysfunction [39] observed in the aged population [40].
A recent study by Mahmoud et al. [31] brings to light another potential mechanism contributing to age-related EG changes. In this ground-breaking work, surface expression of heparan sulfate and GPC1 on human endothelial cells was inversely related to basolateral surface membrane stiffness. Concomitantly, increased basement membrane stiffness of human endothelial cells decreased the phosphorylation of endothelial nitric oxide synthase (eNOS), suggesting that stiffer blood vessels promote decreased eNOS activity. They further observed that young mice with GPC1 knockout demonstrated substantial decreases in en face phosphorylated-eNOS staining of the aortic endothelium to levels similar to those observed in older wild-type mice. In light of the fact that eNOS activity is integral in vasoreactivity through the generation of nitric oxide with downstream activation of vascular smooth muscle cell (VSMC) guanylate cyclase, this work links vascular stiffness to GPC1 expression in the EG and vasoreactivity. As vascular stiffness increases with advancing age [41], [42], these findings suggest that the more compliant vasculature in young children promotes a more robust baseline EG compared to adolescents and young adults that could have important implications for basal EG health.
Evolution of the inflammatory response
The immune system undergoes drastic changes throughout childhood, ranging from immune-tolerance and dependence on innate immunity in the neonate to a rapidly adaptive immune system in the school-aged child [43]. The maturity and responsiveness of the immune system likely have a significant impact on EG integrity during the systemic inflammatory response in critical illness in children. Neonates and young infants are vitally dependent upon circulating maternal antibodies acquired through transplacental adsorption as natural killer cells [44], [45], neutrophils [46], [47], and monocytes/macrophages [48], [49] demonstrate decreased cell responsiveness. CD4+ T cells preferentially differentiate toward anti-inflammatory type 2 helper T (Th2) cell [50] and regulatory T cell [51] phenotypes. More B cells are of B1 lineage that secrete more low-specificity immunoglobulin M, have limited class-switching capabilities [52], and support an anti-inflammatory Th2 response [43]. Thus, neonates and young infants generally have a more muted immune response to pathogen- and damage-associated molecular patterns. Though the immune profile of this age group increases the child’s susceptibility to invasive organisms, it may better preserve EG integrity by limiting sheddase activity and free radical generation. As children develop a more responsive immune system with memory to an increasing repertoire of pathogens [53], [54], their bodies can mount a more robust, protective reaction to local and systemic insults that may come at a cost to EG integrity.
Influence of comorbidities
Comorbid conditions can substantially affect baseline EG integrity and may therefore lower the threshold for aberrant vascular biology during acute childhood illnesses. Diseases such as congenital cyanotic heart defects and lung vascular malformations, bronchopulmonary dysplasia, and hepatopulmonary syndrome expose affected patients to chronic hypoxemia. Homeostatic bioenergy pathways in the child with chronic hypoxemia are forced to adapt to a lower than normal oxygen tension to avoid tissue hypoxia. Hypoxemia triggers production and release of pro-inflammatory interleukin-6 [55] and mitochondria-derived reactive oxygen species [56] from endothelial cells. These oxygen free radicals go on to promote hypoxia-inducible transcription factor expression that may sustain pro-inflammatory cytokine production and signaling [57]. Together, the higher presence of reactive oxygen species and circulating pro-inflammatory milieu [58] during chronic hypoxemia increase the risk of EG thinning [59], though the impact of chronic hypoxemia on pediatric EG expression has not been systematically evaluated.
Children with chronic kidney disease are also at risk for EG thinning through mechanisms that remain unclear [60]. Chronic renal insufficiency places affected patients at risk for developing hypertension and hyperhomocysteinemia. Persistently elevated blood pressure and circulating homocysteine levels promote vascular stiffness [61], [62] that, as demonstrated by Mahmoud et al. [31], can result in attenuated expression of heparan sulfate proteoglycans on the vascular surface. Further, exposure to chronic venovenous dialysis is associated with EG shedding [63].
Children with type I diabetes mellitus (DM) can manifest EG thinning prior to overt signs of vasculopathy [64]. Nussbaum et al. [64] observed that microvascular EG thickness inversely correlated with blood glucose levels in children with type I DM. Similarly, EG thinning was observed in adults with type II DM [65], suggesting that children with poorly controlled type II DM may also manifest EG thinning. EG injury may occur in children with persistent hyperglycemia [66] by promoting increased expression of EG-targeting sheddases [67], [68], reactive oxygen species [69], and advanced glycation end products [70].
Childhood obesity may also threaten baseline EG health secondary to increased basal inflammation [71], [72]. Throughout childhood, macrophages accumulate in adipose tissue to moderate excess macronutrients and adipocyte turn-over [73], [74]. This cellular response leads to complex adipocyte-macrophage interactions that result in diminished expression of the protective adipokine adiponectin and increased expression of pro-inflammatory leptin and tumor necrosis factor-α [75], [76], [77]. The circulating pro-inflammatory milieu may therefore increase expression of EG-targeting sheddases and reactive oxygen species. The denuded vascular surface then permits lipid extravasation and leukocyte adhesion at sites of EG damage [78], [79]. Atherosclerotic plaques begin to develop as monocytes/macrophages infiltrate the vascular wall, promoting VSMC recruitment [80] with resultant vessel stiffening that may exacerbate EG thinning [81], [82]. These plaques also alter the shear stress profile at the endothelial surface, leading to downstream changes in endothelial gene expression and cell conformation [83], [84]. The architectural modifications to the vasculature make the endothelial surface more prone to leukocyte adhesion [85], creating a vicious cycle that can further threaten EG integrity and propagate vascular derangements.
Children with hematologic or immunologic diseases such as sickle cell disease or thrombotic microangiopathy are at particular risk of EG thinning [86]. In sickle cell disease, circulating levels of sickled, poorly deformable red blood cells adhere to the endothelial surface and occlude microcirculation [87], promoting local tissue hypoxia. Additionally, the endothelium is locally activated, promoting a pro-inflammatory and pro-coagulant milieu [88]. Together, these events likely drive the release of sheddases at the vascular surface that thin the EG. Children who develop thrombotic microangiopathies in diseases such as atypical hemolytic-uremic syndrome have aberrant complement pathway activation that precipitates leukocyte priming and endothelial secretion of von Willebrand factor. These processes generate microthrombi and promote leukocyte-endothelial adhesion that culminate in EG disruption and microvascular occlusions that further drive hypoxia-related inflammation [89] that threaten EG health.
Collectively, children with any of the above comorbidities may have a less robust EG either systemically or in particular diseased organs at baseline that has important implications in a child’s predisposition to leukocyte-endothelial adhesion, vascular leakage, and impaired vascular responsiveness during critical illness. Informed by these predispositions, when presented with a child who has an underlying comorbidity known to impact EG integrity, a critical care provider may then better anticipate signs or symptoms of vascular leak that may impair organ function and may employ therapeutic strategies to protect and restore the EG.
The endothelial glycocalyx in specific pediatric critical illnesses
Severe infection and sepsis
Severe infection leading to sepsis remains a significant cause of pediatric mortality globally [90], [91], prompting the World Health Organization to pass a resolution to make the identification and treatment of sepsis a high priority for the 194 members of the United Nations [92]. Sepsis, recently clarified as the dysregulated host response to an infection resulting in life-threatening organ dysfunction [93], continues to have incompletely understood pathophysiology with limited successful treatments. It is logical that a clearer understanding of the etiology and implications of endothelial derangements, including EG disintegrity, caused by invasive pathogens may shed light on novel therapeutic strategies for this pervasive disease impacting children and adults alike.
Several recent reviews provide lengthy descriptions of the impact of sepsis-induced inflammatory response on EG integrity [94], [95], [96]. Here, we would like to highlight several important mechanisms implicated in sepsis-mediated EG degradation and discuss specific diseases relevant to children. Oxygen free radicals generated and released by circulating leukocytes in defense against invading pathogens promote glycocalyx disruption [97], [98]. Leukocytes, platelets, and endothelial cells release proteases, endoglucuronidases, and hyaluronoglucosidases that respectively cleave proteoglycans, GAGs, and hyaladherins from the vascular endothelial surface [99], [100]. Finally, low flow states in large and small vessels during septic shock can disrupt endothelial surface shear stress, promoting EG thinning [101] and inhibiting EG restoration [102] that can exacerbate pathological endothelial cell signaling [103].
At present, little is known about the signature of circulating EG-degrading enzymes or soluble glycocalyx components in children with sepsis. We observed in a secondary analysis of a small, single-center observational study [104] that plasma levels of SDC1 increase in children with sepsis (Fig. 2A). This was concordant with elevations in plasma neutrophil elastase levels (Fig. 2B), a protease with known activity on SDC1 [105], measured at the time of admission to the intensive care unit. This cohort included a host of viral, bacterial, and/or fungal infections in children with a wide array of comorbidities. Further, Pudjiadi et al. [106] observed that SDC1 levels in children admitted to the hospital with sepsis correlated with measures of organ dysfunction as indicated by Pediatric Logistic Organ Dysfunction-2 scores [107] determined on the first day of hospitalization. Together, these data suggest that the EG damage is present in children with sepsis and may play a significant role in the development of organ injury as put forward by Fernandez-Sarmiento et al. [108].
Fig. 2.
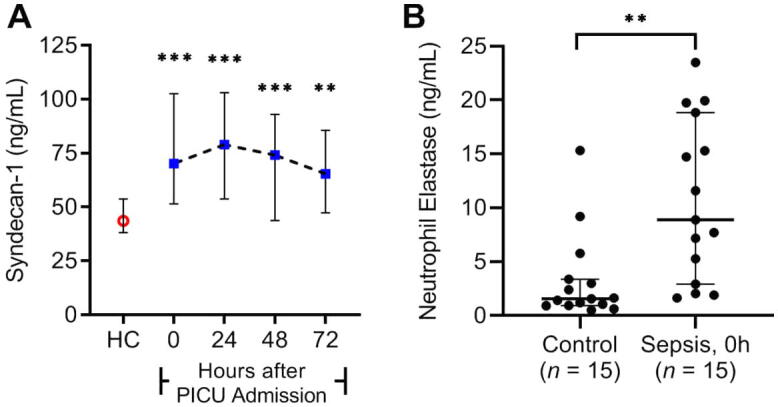
Plasma levels of syndecan-1 and neutrophil elastase in children with sepsis. A, Syndecan-1 levels are significantly elevated in children admitted to the pediatric intensive care unit with sepsis (n = 39) compared to healthy controls (HC) (n = 39). **P < 0.01, ***P < 0.001 versus control after Kruskal-Wallis one-way ANOVA corrected by Dunn’s multiple comparisons test. B, Neutrophil elastase levels in fifteen randomly selected children with sepsis were significantly elevated upon admission to the pediatric intensive care unit compared to controls (n = 15). Data presented as median (interquartile range). **P < 0.01 versus control after Mann-Whitney U test.
Preclinical and clinical studies characterizing EG injury during particular infections, including herpes simplex viral (HSV), dengue viral, and malarial diseases, may shed light into the pathophysiology occurring at the endothelial surface during pediatric sepsis. HSV infection is a serious perinatal disease with high rates of morbidity and mortality [109]. Recent work has demonstrated that HSV promotes heparanase (HPSE) 1, matrix metalloproteinase (MMP)-3, and MMP-7 upregulation in infected cells, resulting in heparan sulfate and SDC1 cleavage from the cell surface that thereby liberates virions from the host cell to infect other host cells [110]. Although these studies were performed using human corneal epithelial cells, these findings likely have direct implications in the pathophysiology of disseminated neonatal HSV driving endothelial surface layer degradation, downstream endotheliopathy, and eventual organ dysfunction that commonly portends death.
Dengue viral infection is a severe tropical illness that has the highest prevalence in children <15 years of age [111]. Plasma levels of SDC1 and hyaluronan are significantly higher in patients with dengue-related hemorrhagic fever or severe plasma leakage than those with the milder dengue fever alone [112]. Intriguingly, plasma heparan sulfate levels were not significantly different between dengue severity groups despite in vitro work demonstrating that dengue virus nonstructural protein 1 (DENV NS1) promoted endothelial cell upregulation of HPSE1 with resultant increased transcellular permeability [113]. Treatment of human endothelial cells with DENV NS1 was also associated with an increased surface expression of SDC1, suggesting that dengue viral infection may increase EG turn over to permit viral escape from the endothelium.
Children with severe Plasmodium falciparum infection, one of the five protozoan species that cause human malaria, demonstrate higher circulating levels of SDC1, GPC1, heparan sulfate, and hyaluronan than healthy controls [114], [115]. Further, using videomicroscopy, children with severe malaria were observed to have EG thinning [114]. In light of the pathogenesis of this parasite (i.e., preferential infection of circulating erythrocytes with phases of erythrocyte rupture and associated parasitemia, malarial toxin release, and leukocyte activation [116], [117]), these findings suggest that the host inflammatory response to plasmodia is responsible for indiscriminant EG erosion. P. falciparum infection in particular is responsible for cerebral malaria in children. Infected erythrocytes adhere to the cerebral vascular endothelium, leading to microvascular obstruction and local inflammation [118]. In this process, the EG is likely eroded, promoting coagulation, leukocyte adhesion, and loss of endothelial barrier function that culminates in cerebral edema [119], [120], [121]. Acute kidney injury is also common with P. falciparum through microvascular occlusion, local inflammation, and EG erosion [122], [123], resulting in filtration and tubular absorption defects [124].
The resuscitation strategy for pediatric sepsis [125] has important differences from adult sepsis [126] with potential ramifications on EG health. First, fluid resuscitation tends to be more liberal in children than adults as children typically receive at least 40–60 mL/kg of intravenous (IV) crystalloid before starting vasoactive medications compared to adults who tend to receive approximately 30 mL/kg of crystalloid before initiating vasoactive support. As high volume IV crystalloid administration can damage the EG [127], resuscitation strategies that incorporate higher volumes of fluid resuscitation may promote greater EG disruption and, paradoxically, worsen vascular leak and organ function. Therefore, a more restrictive fluid resuscitation strategy with earlier initiation of vasoactive medication may protect the vasculature. However, high circulating levels of epinephrine may similarly promote EG disintegrity [128]. Additionally, vasoconstricting regions of microcirculation in need of ongoing volume repletion may actually worsen regional perfusion, increasing local vascular inflammation that could further threaten EG integrity. Thus, the balance between circulating catecholamines, crystalloid resuscitation, and microcirculatory perfusion likely has significant impact on EG health during pediatric sepsis. The relative contribution of each of these competing variables requires further interrogation.
Choice of resuscitative fluid for pediatric sepsis may also impact EG recovery and clinical outcomes. Preclinical data suggest that resuscitation with albumin restores microcirculatory flow to a greater degree than 0.9% saline and may support EG restoration [129], [130]. In a small randomized controlled trial in adults with sepsis, IV colloid resuscitation resulted in higher cardiac indices and plasma volumes compared to IV crystalloids with the benefit of lower volumes of fluid administration to achieve hemodynamic goals [131]. This study notably included gelatin- and hydroxyethyl starch-based colloids, both of which may increase the risk of acute kidney injury [132], [133], and did not evaluate the specific interaction between albumin-based resuscitation and later organ function or markers of EG integrity. In the Saline versus Albumin Fluid Evaluation (SAFE) trial, a subgroup analysis of adults with severe sepsis demonstrated a trend toward improved outcomes with the use of 4% albumin as primary resuscitation fluid compared to crystalloid [134]. However, targeting a specific serum albumin level using 20% albumin infusions did not improve survival rates in adults with severe sepsis [135]. Similarly, two meta-analyses examining the effect of albumin versus crystalloid resuscitation on clinical outcomes in critically ill adults (not limited to sepsis) were unable to detect differences in mortality [136], [137]. Informed by these large studies and the potential for worsening extravasation of plasma proteins, international guidelines suggest the use of crystalloid over albumin for initial pediatric and adult sepsis resuscitation [125], [126]. Nevertheless, a clinical trial evaluating the impact of albumin-based resuscitation on EG integrity and organ function in pediatric sepsis could have merit.
Fresh frozen plasma (FFP) has an even greater ability to protect the EG compared to albumin [138] and thus may reduce vascular permeability and sequential organ failure if used early during pediatric sepsis resuscitation. FFP is not considered a first-line fluid for sepsis in either children or adults and inherently presents the unpredictable risk of transfusion related events (e.g., acute lung injury, worsened fever, alloimmunization, and infectious disease transmission). In light of its potential EG-protective effects with the ability to better preserve oncotic pressure relative to crystalloid, evaluation of the use of FFP in the resuscitation of pediatric sepsis is also rational.
SARS-CoV-2 infection
Infection by the novel severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) appears to impact humans differently from other respiratory viruses, particularly in children [139]. Thus, SARS-CoV-2 infection warrants its own discussion. When the alpha and beta SARS-CoV-2 variants circulated in early-to-mid-2020, elderly were disproportionately affected while most children remained asymptomatic or mildly symptomatic outside of those with a comorbid history of prematurity, chronic lung disease, or obesity [140]. It was not until weeks to months after the first wave of SARS-CoV-2 infection that children began manifesting symptoms similar to Kawasaki disease with myocardial dysfunction that was not observed in adults, a non-specific inflammatory condition called multisystem inflammatory syndrome in children (MIS-C). We are emerging from the global wave of the highly contagious delta variant wave with greater symptomatic disease in children. Regardless of the variations in disease presentation in children and adults with coronavirus disease 2019 (COVID-19), the vascular endothelium seems to be center stage.
SARS-CoV-2 infection is caused by viral spike (S) protein interaction with angiotensin-converting enzyme 2 (ACE2) receptors on the nasopharyngeal epithelium [141]. Transmembrane protease, serine 2 (TMPRSS2) expressed on the epithelial surface then cleaves the S protein, permitting viral fusion with the host cell membrane and intracellular infection [141]. Heparan sulfate in the epithelial surface glycocalyx enhances SARS-CoV-2 attachment to ACE2 receptors by binding region-binding domains of the S protein, promoting conformational changes to the S protein that favor ACE2 receptor interaction [142]. The innate immune system is typically the child’s primary host defense (unless born to a vaccinated or previously infected mother). As discussed above, a child’s immune system has particular features unique from the adult immune system that include a quicker antiviral interferon response [143], more rapid immunoglobulin production with broad reactivity [144], and a tempered natural killer cell response [44]. Therefore, the pediatric immune system may be able to more quickly and quietly handle acute SARS-CoV-2 infection without a hyperinflammatory response. However, the idiosyncrasies of the pediatric immune system that permit acute SARS-CoV-2 viral infection to transition to MIS-C are unknown.
In severe COVID-19, SARS-CoV-2 virions cross from the respiratory epithelium into circulation, infecting the endothelium of multiple organs [145]. Adults with COVID-19 are at risk of micro- and macrovascular thrombosis due to EG injury, endothelial cell activation, neutrophil extracellular traps [146], [147], and monocyte tissue factor release with subsequent increase in monocyte-platelet aggregates [148]. Consistent with this pathophysiology, several studies have shown that SDC1 [149], heparan sulfate [150], [151], and hyaluronan levels [150] are elevated in adults with severe COVID-19 along with soluble levels of P-selectin [152], thrombomodulin [152], intercellular adhesion molecule 1 [153], and vascular cell adhesion molecule 1 [153] (all proteins commonly expressed on the endothelial surface), suggesting significant EG erosion. Similarly, Buijsers et al. [154] observed that plasma levels of heparanase activity correlated with illness severity in adults with COVID-19. Our understanding of the precise mechanisms by which SARS-CoV-2 promotes EG damage, however, remains incomplete.
At present, there are no glycocalyx biomarker data available in children with acute COVID-19, and only two case reports of children with MIS-C have reported plasma levels of glycocalyx biomarkers [155], [156]. Fraser et al. [155] observed that hyaluronan and soluble P-selectin, but not heparan sulfate, levels were elevated in an adolescent female with MIS-C compared to 20 healthy controls. Fernandez-Sarmiento et al. [156] reported elevated SDC1 levels and microvascular EG thinning on videomicroscopy in two patients with MIS-C relative to a healthy child. Though preliminary, these data shed light into COVID-19-related vascular disease in children and suggest that plasma biomarkers of glycocalyx shedding and videomicroscopy may serve as important evidences of therapeutic efficacy for debated treatments [157].
Trauma/hemorrhage
Despite modern advancements in the prevention and management of severe trauma, traumatic injuries continue to be a leading cause of morbidity and mortality for children and adults worldwide [158]. The traumatically injured child can have marked differences in mechanism and pattern of injury compared to adults [159]. The child’s higher metabolic needs and lower hepatic glycogen reserve make relatively short periods of asphyxiation less tolerated by the brain and the heart, resulting in higher risk of hypoxic-induced encephalopathy and cardiac arrest than their adult counterparts. Each of these age-specific factors may heavily influence the injury pattern to the vascular surface following trauma. Recent preclinical work and studies in adults have shed light on the role that trauma-related glycocalyx disintegrity has on post-injury coagulation abnormalities, inflammation propagation, and clinical outcomes. However, little is known about EG injury in pediatric trauma and its impact on vascular functions, organ injury, and outcomes.
Endotheliopathy of trauma, driven principally by the pathological activation of the endothelium and associated damage to the EG, has been extensively characterized in adults [160], [161], [162]. Only a few studies have reported biomarkers of EG degradation and pathologic activation of the endothelium following pediatric trauma [163], [164]. Glycocalyx shedding has been reported within minutes of trauma in adults [165] and is associated with early organ dysfunction [166]. Glycocalyx damage also appears to be an early event after pediatric trauma as suggested by elevated plasma SDC1 levels in injured children measured upon hospital arrival [163]. Non-survivors of adult [160], [161] and pediatric [163] trauma have significantly higher circulating markers of glycocalyx damage compared to survivors. Trauma-induced EG degradation is also associated with postinjury coagulopathy in adults [160], [167], [168] and children [163], [164], implicating a role for glycocalyx shedding in post-traumatic coagulation abnormalities.
Following the initial direct insult to the vascular surface and the ensuing inflammatory response, persistent derangements in EG integrity have been found to propagate vascular dysfunction in traumatically injured adults, culminating in organ dysfunction [165]. However, the associations between ongoing EG damage and organ injury have not been evaluated in the pediatric trauma population. Overall, children have a lower risk of developing postinjury multiorgan failure [169]. This may be related to trauma epidemiology (e.g., younger children have a higher proportion of isolated traumatic brain injury (TBI) compared to a higher incidence of polytrauma in the adolescent [170]), differences in the pediatric post-traumatic immune response [169], and presence of relatively fewer comorbidities that may confer protection to children. However, the impact of age and injury characteristics on EG integrity following pediatric trauma have not been reported. Data from the Pragmatic, Randomized Optimal Platelet and Plasma Ratios (PROPPR) clinical trial [171] indicate an inverse relationship between age and plasma levels of SDC1 as a marker of vascular glycocalyx damage in patients with similar injury characteristics and degree of shock (Fig. 3). We posit that the age-dependent reduction in circulating SDC1 is a function of reduced glycocalyx expression with increasing age (as described above) in addition to age-related changes in the post-traumatic immune response [172]. A more robust characterization of the inflammatory response and other glycocalyx markers is needed to more clearly define the impact of age on post-traumatic EG damage.
Fig. 3.

Effect of age on trauma-induced glycocalyx shedding. Seventy-four traumatically injured adults who were enrolled in the PROPPR trial [171] and had available injury severity data, admission base excess levels, and admission plasma syndecan-1 levels were stratified by age quartile for analysis. No differences were observed in injury severity (A) or degree of hemorrhagic shock as measured by blood levels of base excess (B) between age groups. Syndecan-1 levels, indicative of glycocalyx damage, declined with increasing age (C). Data presented as median (middle line in box) with interquartile range (IQR) (upper and lower limit of boxes) +/- 1.5 × IQR (Tukey whiskers). ***P < 0.001 after Kruskal-Wallis one-way ANOVA.
Hemorrhagic shock compounds the post-traumatic inflammatory insult and promotes greater EG damage [173]. Informed by the PROPPR trial [171], resuscitation of traumatically injured adults with hemorrhagic shock typically involves balanced component blood product transfusion (administration of equal volumes of FFP and platelets with the transfused red cells). Children, on the other hand, do not commonly present in hemorrhagic shock following severe trauma, likely related to trauma epidemiology and pattern of injury (i.e., more isolated head injury). Further, those children who present with trauma-related hemorrhagic shock that does not warrant massive transfusion are more commonly resuscitated with crystalloid fluid and packed red blood cells without a balanced transfusion approach. However, this strategy may lead to worse clinical outcomes and is thus being challenged [174].
At present, it is unknown how choice of resuscitative fluid affects EG integrity and clinical outcomes in traumatically injured children. Preclinical data from hemorrhagic shock models in rats demonstrated that resuscitation with 5% albumin partly restored EG thickness and attenuated vascular leak [175], [176]. Pati et al. [177] did not observe the same improvement in vascular leak when using an albumin-based resuscitation in hemorrhaged mice compared to saline, though a substantially more dilute albumin product (approximately 0.03%) was used. The mechanisms by which albumin contributes to EG protection are not entirely clear but may relate to inhibition of leukocyte rolling and endothelial adhesion [178] in addition to facilitating sphingosine-1-phosphate interactions with endothelial surface receptors to inhibit EG degradation and promote EG reconstitution [179], [180]. Results of albumin-based resuscitation in adult trauma have been less favorable. Post-hoc analysis of the SAFE trial [181] demonstrated that critically ill adults who suffered TBI and were resuscitated with 4% albumin had significantly higher mortality than those resuscitated with saline 0.9% [181]. A recent review [182] brings to light important limitations to this study that may have influenced outcomes, particularly the use of a hypotonic albumin solution that may have contributed to worsening cerebral edema and increased intracranial pressure [183]. Despite the limitations of the SAFE trial and any promising preclinical data that would support albumin use in trauma/hemorrhage resuscitation, pediatric intensivists, surgeons, and neurosurgeons tend to avoid albumin in the management of trauma. It is unlikely that interest in studying an albumin-based resuscitation strategy in children will pique without more compelling preclinical and adult clinical evidence of its efficacy on vascular health and outcomes.
Blood product transfusion may confer greater EG protection than crystalloid or albumin resuscitation following pediatric trauma. In a rat model of hemorrhage-resuscitation, Torres Filho et al. [175] observed that resuscitation with FFP or fresh whole blood following hemorrhage resulted in significantly greater EG preservation compared to crystalloid or albumin as indicated by lower plasma levels of heparan sulfate and SDC1, reduced vascular leak, and greater microvascular EG thickness on intravital microscopy. Other groups have similarly demonstrated the benefits of FFP resuscitation on EG integrity in various in vitro and in vivo models [184], [185]. Clinically, traumatically injured adults at risk of hemorrhagic shock who were administered FFP in the prehospital setting showed a reduction in coagulation abnormalities and improved mortality [186]. In a recent propensity-matched retrospective cohort study of the American College of Surgeons 2017 Trauma Quality Improvement Program national database [187], children resuscitated with whole blood and component blood product therapy (WB + CT) in the first 4 h post-trauma required less blood product by 4 and 24 h and experienced fewer days of mechanical ventilation compared to children who received component therapy (CT) alone. Total crystalloid fluid resuscitation in the first 24 h was not reported. The volume of FFP and whole blood accounted for 68% of the total blood volume in the WB + CT group, whereas FFP accounted for only 25% of the total blood volume administered in the CT group. If whole blood is considered to have similar EG-protective effects as FFP [175], [188], then it is plausible that the higher total FFP volume in the WB + CT group could have sustained or improved EG integrity, thereby limiting lung microvascular damage and systemic coagulation abnormalities. Despite the limitations of a retrospective analysis, this study provides evidence that a balanced blood product resuscitation approach is both feasible and safe in children.
Severe burns
Unintentional injuries were the leading cause of death in the United States in 2019 in children above 12 months of age [189]. This statistic included burn injury with a peak incidence in the 5–9 year old age group, a particular at-risk population due to their mobility without capacity to anticipate fire or thermal hazards. Because of the relatively higher body surface area-to-weight ratio, thermally injured pediatric patients suffer higher intravascular volume losses [190]. Given the smaller circulating blood volumes in children, this places the critically ill child with burn injury at significant risk for severe hypovolemia, resulting in hypoperfusion and multiorgan dysfunction.
The EG is integral to the pathophysiology of burned children, regulating vascular leak, platelet and leukocyte adhesion/activation, and inflammatory endothelial cell signaling. In thermally burned rodents, plasma levels of SDC1 abruptly rise proportional to the total body surface area (TBSA) burned [191], [192], corresponding with a significant increase in vascular leak [192]. This finding was corroborated in a small, single-center adult study in which plasma SDC1 levels were significantly higher than control levels at hospital admission and remained elevated for up to 30 days following hospitalization [193]. Interestingly, admission SDC1 levels did not correlate with TBSA percentage but instead correlated with total fluid resuscitation administered over the first 24 h. Fluid resuscitation strategy was dictated by both the percent TBSA burned and a targeted urine output, which may explain the discrepancy with the correlation observed in the rodent models described above. Plasma levels of SDC1 at hospital admission were found to be a helpful predictor of 30-day mortality following severe burns in adults [194]. Together, these data support a derangement of the EG as a likely key driver of vascular leak and clinical outcomes following severe thermal injury that may similarly apply to children.
The mainstay therapy for severe pediatric burns (traditionally defined as second- or third-degree thermal injury of TBSA ≥ 10%) is judicious, algorithmic IV crystalloid resuscitation [190]. Though life-saving, administration of high volume crystalloid in a patient with an inflamed endothelium may exacerbate vascular leak by increasing EG shedding [195]. Albumin supplementation is commonly considered in burn resuscitation to boost oncotic pressure in the vascular lumen, improve microcirculatory perfusion, and reduce the amount of crystalloid administered [190]. Albumin may also protect the EG from ongoing injury by reinforcing its fragile structure, buying the vasculature time to restore the glycocalyx. FFP transfusion may also have a role in the resuscitation of the severely burned child. In thermally injured rats with TBSA 40% burn, FFP administration improved SDC1 expression in lung tissue and normalized vascular leak [192]. FFP is not a typical therapy employed in pediatric burn management outside of the massively hemorrhaging patient and comes with inherent risks [196]. Nonetheless, research into the mechanisms driving vascular injury after severe burns and evaluation of the merits of more aggressive albumin or FFP administration in thermally injured children warrants consideration.
Extracorporeal life support
The availability of extracorporeal membrane oxygenation (ECMO) and cardiopulmonary bypass (CPB) technologies has revolutionized modern medicine by allowing life-sustaining support for cardiovascular and/or pulmonary system failure or for surgical intervention of life-threatening cardiothoracic diseases. Deployment of these forms of extracorporeal life support (ECLS) in the neonatal and pediatric populations has grown substantially over the last 10 years (1,955 neonatal runs in 2020 compared to 1,312 runs in 2010; 2,914 pediatric ECMO runs in 2020 compared to 1,068 in 2010 [197]) as more medical centers become facile with ECLS management and surgical sophistication burgeons for once lethal diseases. As important as these technologies have been in improving outcomes of otherwise fatal disease processes, their use is not without consequence. Organs remote to the respiratory and cardiovascular systems are at substantial risk for injury from ECLS. Direct and indirect EG injury is central to this process.
Generally the immune system is already systemically active in critically ill children requiring ECMO prior to cannulation [93], [198]. The vascular endothelium is thereby pathologically activated to upregulate EG-targeting sheddases and is primed for leukocyte adhesion, resulting in ongoing EG injury [199]. After ECMO initiation, circulating components of the immune and coagulation systems interact with artificial surfaces, increasing inflammation and coagulation activation. To maintain hemostasis, patients commonly receive component blood product transfusion, which further exposes the child to alloantigens and increases the risk for inflammation and coagulation activation. Over time, as the ECMO circuit longevity wanes due to increased thrombosis of circuit components, fibrin deposits shear red blood cell membranes with resultant hemolysis and an increase in plasma levels of free hemoglobin and iron. Increased circulating levels of plasma free hemoglobin may overwhelm the body’s anti-oxidant reserves, leading to increased oxygen and nitrogen free radicals with deleterious effects on the EG [200].
Children who require CPB generally have a congenital heart defect requiring corrective or palliative surgical intervention or need heart and/or lung transplantation. Depending on the type of congenital heart defect or degree of lung disease requiring transplantation, some children live with chronic hypoxia prior to palliative surgery that, as discussed above, can promote pro-inflammatory endothelial cell signaling [201] and endothelial surface layer thinning [59]. Then, with transition to CPB at the time of surgical intervention, regions of the body experience variable states of hypoperfusion and ischemia. Following completion of surgery and liberation from CPB, areas of hypoperfusion and ischemia are flooded with oxygen-rich blood, leading to reperfusion injury from mitochondrial dysfunction and depletion of anti-oxidant reserves. Ischemia-reperfusion injury then drives the activation of complement [202] and upregulation of pro-inflammatory cytokines [203] that promote the release of EG-eroding sheddases [204].
Puchwein-Schwepcke et al. [2] have nicely summarized the literature available characterizing EG injury in children undergoing congenital heart surgery. As with adults [205], [206], exposure to CPB results in increased circulating levels of SDC1, heparan sulfate, and hyaluronan in children [207], [208], [209], [210], corresponding to decreased EG thickness on videomicroscopy [211]. While measures of EG injury are unknown during neonatal or pediatric ECMO, we can deduce from pre-clinical studies that a similar phenomenon likely occurs after initiation of ECMO in children [212].
There are major differences between management of CPB and ECMO that may translate in more prolonged EG injury with ECMO use. ECMO duration is typically much longer than CPB, exposing the patient to a pro-inflammatory milieu for a greater duration with untoward effects on the endothelial surface layer discussed above. Many children who undergo CPB receive FFP or human albumin during the course of their surgery that may have glycocalyx-protective effects. Upon completion of CPB, many centers are using modified ultrafiltration that provides some cytokine adsorption and limits the degree of fluid overload to the patient after surgery, which may similarly protect the EG. On the other hand, crystalloid resuscitation is the norm during ECMO to sustain adequate flows. The benefit of temporarily improved flows is then counter-balanced with fluid overload that potentially worsens EG injury and microcirculation [213]. Finally, corticosteroids are often used in the management of critically ill children requiring CPB or ECMO to attenuate the pro-inflammatory response, treat absolute or relative adrenal insufficiency, and promote convalescence [214], [215]. Corticosteroid use may also confer protection to the vascular EG through unresolved mechanisms [216], [217]. However, pediatric intensivists must pay particular attention to the development of steroid-induced hyperglycemia or secondary infections related to immunosuppression, both of which can compromise the EG [66], [218].
Challenges and opportunities
Studying vascular glycobiology is difficult and nuanced, particularly in children. The microscopic, fragile nature of the EG makes this structure challenging to directly image and quantify [219]. Special techniques are used to attempt tissue EG preservation for electron microscopy [220] but may not consistently prepare the endothelium for high quality imaging [221]. Others have studied tissue-level EG expression using immunohistochemistry and plasma measures of glycocalyx components to characterize EG injury in both humans and animal models. These techniques are unfortunately susceptible to inconsistent detection of sugar moieties due to inherently poor protein–sugar interactions by commercial antibodies. To overcome these challenges, some groups are using liquid chromatography-tandem mass spectrometry to quantify shed GAG fragments in biological fluids [222]. While this technique provides more granular detail of shed GAG size and sulfation pattern, it requires a highly specialized laboratory. For real time visualization of the EG in animal models, investigators use intravital microscopy with or without fluorescence [223]. However, this technique is limited to surface-layer evaluation of microcirculation due to its poor tissue penetration. For human studies, orthogonal polarization spectroscopy or sidestream darkfield (SDF) imaging can be used to determine EG thickness of microcirculatory beds. Like intravital microscopy, videomicroscopy of human microcirculation is limited to accessible surface-layer vascular beds, namely the sublingual [224], buccal [114], auricular [20], and extremity [225] microvasculature. Moreover, videomicroscopy currently lacks the capability of detecting granular detail of the EG such as the expression patterns of specific proteoglycans or GAGs and is only able to provide indirect measures of EG thickness.
Biomarker evaluation and SDF videomicroscopy have been most commonly used to characterize EG damage in children [2]. However, performing even these limited assessments can be difficult to perform and interpret in critically ill children. First, there are relatively fewer children admitted to the hospital for critical illness compared to adults, making it challenging to recruit large cohorts for comprehensive research efforts. Next, in vivo EG characterization using videomicroscopy has not yet achieved wide-spread popularity. Therefore, most clinical investigations into pediatric endothelial health rely upon plasma and urine biomarker evaluations. These too have significant limitations related to the developmental status of the child, which may limit the cooperativity in obtaining specimens, and access to a free-flowing sample. Due to the smaller circulating blood volumes in children and concern for iatrogenic anemia, most hospitals have policies limiting the blood volume sampled in a 24-hour period [226]. This limitation may further hinder sampling for research. Blood and urine levels of EG biomarkers can also be influenced by renal function [227], complicating the interpretation of analyte results. Finally, it can be challenging to approach parents/guardians of critically ill children for research purposes. Given the high stress environment and lack of direct benefit to their child, many families elect to withhold from participating in research. Multi-center collaborations are thus needed to overcome these limitations and help move the field of pediatric vascular biology forward.
In vitro modeling of the endothelium is employed to help resolve gaps in our understanding of the mechanisms driving pediatric vascular disease and EG damage. However, this work is similarly challenging. First, unlike commercially available adult-derived endothelial cells commonly used for mechanistic studies, pediatric-derived organ-specific endothelial cells are not currently commercially available outside of human umbilical vein endothelial cells, which may limit the translatability of laboratory observations to critically ill children. Further, endothelial cells are commonly cultured on hard plastic in a static environment away from their natural habitat with surrounding pericytes and VSMCs, which significantly hinders EG expression [31]. Subjecting endothelial cells surface layer shear stress using parallel flow chambers or cone-and-plate viscometers generates a model that mimics a healthy, quiescent human endothelium [228], [229] with more robust EG expression relative to statically cultured cells [102], [230]. However, employing in vitro flow modeling to more aptly study the role of the EG in endothelial pathobiology can be technically challenging and have low through-put. Finally, EG-targeting enzymes used for in vitro studies may not perfectly simulate human enzymatic activity [231], further limiting the translation of bench work to bedside understanding of pediatric vascular biology.
Despite these challenges, improving our understanding of the mechanisms up- and downstream of EG injury during severe pediatric diseases holds great promise for the discovery of novel therapeutic strategies that mitigate endothelial dysfunction and organ injury. Elucidating the sheddases involved in acute inflammation may point to particular small molecule inhibitors that confer EG protection [232], [233]. Uncovering the endothelial signaling aberrancies downstream of EG injury that promote vascular leak and propagate inflammation could suggest targets for signaling modulation that restore endothelial homeostasis. Finally, identifying methods to restore EG integrity through extrinsic repletion (using FFP [184], [185] or a purified GAG admixture such as sulodexide [65], [234], [235]) or intrinsic reconstitution [236], [237], [238] may provide synergy with other treatments to bolster the endothelium against inflammatory insults.
Conclusions
The pediatric critical care community is gaining a greater appreciation for the importance of endothelial glycobiology in the pathogenesis of severe childhood illnesses. A child’s age, immune status, and comorbidities can dictate baseline EG expression and significantly influence the severity of vasculitis from systemic insults like sepsis or trauma. As the EG and downstream aberrant vascular biology are central to many of the pathophysiologies observed in the pediatric intensive care unit, treatment strategies aimed at restoring EG integrity thus hold promise in reversing vascular dysfunction and improving organ injury during severe illnesses in children.
Funding sources
This work was supported by the National Institutes of Health (NIH) grants K08 GM144788 to RPR; U01 HL133536 to NA; R01 HL102371 to AG; and R35 GM137958 to JRR. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- 1.Kolarova H., Ambruzova B., Svihalkova Sindlerova L., Klinke A., Kubala L. Modulation Of Endothelial Glycocalyx Structure Under Inflammatory Conditions. Mediators Inflamm. 2014;2014 doi: 10.1155/2014/694312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Puchwein-Schwepcke A., Genzel-Boroviczeny O., Nussbaum C. The Endothelial Glycocalyx: Physiology And Pathology In Neonates, Infants And Children. Front. Cell Dev. Biol. 2021;9:1–11. doi: 10.3389/fcell.2021.733557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Van Den Berg B.M., Vink H., Spaan J.A. The Endothelial Glycocalyx Protects Against Myocardial Edema. Circ. Res. 2003;92:592–594. doi: 10.1161/01.RES.0000065917.53950.75. [DOI] [PubMed] [Google Scholar]
- 4.Van Haaren P.M., Vanbavel E., Vink H., Spaan J.A. Localization Of The Permeability Barrier To Solutes In Isolated Arteries By Confocal Microscopy. Am. J. Physiol. Heart Circ. Physiol. 2003;285:H2848–2856. doi: 10.1152/ajpheart.00117.2003. [DOI] [PubMed] [Google Scholar]
- 5.Megens R.T., Reitsma S., Schiffers P.H., Hilgers R.H., De Mey J.G., Slaaf D.W., et al. Two-Photon Microscopy Of Vital Murine Elastic And Muscular Arteries. Combined Structural And Functional Imaging With Subcellular Resolution. J. Vasc. Res. 2007;44:87–98. doi: 10.1159/000098259. [DOI] [PubMed] [Google Scholar]
- 6.Rosenberg R.D., Shworak N.W., Liu J., Schwartz J.J., Zhang L. Heparan Sulfate Proteoglycans Of The Cardiovascular System. Specific Structures Emerge But How Is Synthesis Regulated? J. Clin. Invest. 1997;99:2062–2070. doi: 10.1172/JCI119377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Fransson L.A., Belting M., Cheng F., Jonsson M., Mani K., Sandgren S. Novel Aspects Of Glypican Glycobiology. Cell. Mol. Life Sci. 2004;61:1016–1024. doi: 10.1007/s00018-004-3445-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Culty M., Miyake K., Kincade P.W., Sikorski E., Butcher E.C., Underhill C. The Hyaluronate Receptor Is A Member Of The CD44 (H-CAM) Family Of Cell Surface Glycoproteins. J. Cell Biol. 1990;111:2765–2774. doi: 10.1083/jcb.111.6.2765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Queisser K.A., Mellema R.A., Petrey A.C. Hyaluronan And Its Receptors As Regulatory Molecules Of The Endothelial Interface. J. Histochem. Cytochem. 2021;69:25–34. doi: 10.1369/0022155420954296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Henry C.B., Duling B.R. Permeation Of The Luminal Capillary Glycocalyx Is Determined By Hyaluronan. Am. J. Physiol. 1999;277:H508–514. doi: 10.1152/ajpheart.1999.277.2.H508. [DOI] [PubMed] [Google Scholar]
- 11.Rapraeger A.C. In The Clutches Of Proteoglycans: How Does Heparan Sulfate Regulate FGF Binding? Chem. Biol. 1995;2:645–649. doi: 10.1016/1074-5521(95)90025-x. [DOI] [PubMed] [Google Scholar]
- 12.Sarrazin S., Lamanna W.C., Esko J.D. Heparan Sulfate Proteoglycans. Cold Spring Harb. Perspect. Biol. 2011;3 doi: 10.1101/cshperspect.a004952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Woodcock T.E., Woodcock T.M. Revised Starling Equation And The Glycocalyx Model Of Transvascular Fluid Exchange: An Improved Paradigm For Prescribing Intravenous Fluid Therapy. Br. J. Anaesth. 2012;108:384–394. doi: 10.1093/bja/aer515. [DOI] [PubMed] [Google Scholar]
- 14.Florian J.A., Kosky J.R., Ainslie K., Pang Z., Dull R.O., Tarbell J.M. Heparan Sulfate Proteoglycan Is A Mechanosensor On Endothelial Cells. Circ. Res. 2003;93:E136–142. doi: 10.1161/01.RES.0000101744.47866.D5. [DOI] [PubMed] [Google Scholar]
- 15.Mochizuki S., Vink H., Hiramatsu O., Kajita T., Shigeto F., Spaan J.A., et al. Role Of Hyaluronic Acid Glycosaminoglycans In Shear-Induced Endothelium-Derived Nitric Oxide Release. Am. J. Physiol. Heart Circ. Physiol. 2003;285:H722–726. doi: 10.1152/ajpheart.00691.2002. [DOI] [PubMed] [Google Scholar]
- 16.Pahakis M.Y., Kosky J.R., Dull R.O., Tarbell J.M. The Role Of Endothelial Glycocalyx Components In Mechanotransduction Of Fluid Shear Stress. Biochem. Biophys. Res. Commun. 2007;355:228–233. doi: 10.1016/j.bbrc.2007.01.137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Henderson-Toth C.E., Jahnsen E.D., Jamarani R., Al-Roubaie S., Jones E.A. The Glycocalyx Is Present As Soon As Blood Flow Is Initiated And Is Required For Normal Vascular Development. Dev. Biol. 2012;369:330–339. doi: 10.1016/j.ydbio.2012.07.009. [DOI] [PubMed] [Google Scholar]
- 18.Conway E.M., Collen D., Carmeliet P. Molecular Mechanisms Of Blood Vessel Growth. Cardiovasc. Res. 2001;49:507–521. doi: 10.1016/s0008-6363(00)00281-9. [DOI] [PubMed] [Google Scholar]
- 19.Lee H.S., Han J., Bai H.J., Kim K.W. Brain Angiogenesis In Developmental And Pathological Processes: Regulation, Molecular And Cellular Communication At The Neurovascular Interface. FEBS J. 2009;276:4622–4635. doi: 10.1111/j.1742-4658.2009.07174.x. [DOI] [PubMed] [Google Scholar]
- 20.Puchwein-Schwepcke A., Artmann S., Rajwich L., Genzel-Boroviczeny O., Nussbaum C. Effect Of Gestational Age And Postnatal Age On The Endothelial Glycocalyx In Neonates. Sci. Rep. 2021;11:3133. doi: 10.1038/s41598-021-81847-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Markopoulou P., Papanikolaou E., Analytis A., Zoumakis E., Siahanidou T. Preterm Birth As A Risk Factor For Metabolic Syndrome And Cardiovascular Disease In Adult Life: A Systematic Review And Meta-Analysis. J. Pediatr. 2019;210(69–80):E65. doi: 10.1016/j.jpeds.2019.02.041. [DOI] [PubMed] [Google Scholar]
- 22.Lee H., Dichtl S., Mormanova Z., Dalla Pozza R., Genzel-Boroviczeny O. In Adolescence, Extreme Prematurity Is Associated With Significant Changes In The Microvasculature, Elevated Blood Pressure And Increased Carotid Intima-Media Thickness. Arch. Dis. Child. 2014;99:907–911. doi: 10.1136/archdischild-2013-304074. [DOI] [PubMed] [Google Scholar]
- 23.Bassareo P.P., Fanos V., Puddu M., Demuru P., Cadeddu F., Balzarini M., et al. Reduced Brachial Flow-Mediated Vasodilation In Young Adult Ex Extremely Low Birth Weight Preterm: A Condition Predictive Of Increased Cardiovascular Risk? J. Matern Fetal Neonatal. Med. 2010;23(Suppl 3):121–124. doi: 10.3109/14767058.2010.506811. [DOI] [PubMed] [Google Scholar]
- 24.Engan B., Engan M., Greve G., Vollsaeter M., Hufthammer K.O., Leirgul E. Vascular Endothelial Function Assessed By Flow-Mediated Vasodilatation In Young Adults Born Very Preterm Or With Extremely Low Birthweight: A Regional Cohort Study. Front. Pediatr. 2021;9 doi: 10.3389/fped.2021.734082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Short K.R., Blackett P.R., Gardner A.W., Copeland K.C. Vascular Health In Children And Adolescents: Effects Of Obesity And Diabetes. Vasc. Health Risk Manag. 2009;5:973–990. doi: 10.2147/vhrm.s7116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Majerczak J., Grandys M., Frolow M., Szkutnik Z., Zakrzewska A., Nizankowski R., et al. Age-Dependent Impairment In Endothelial Function And Arterial Stiffness In Former High Class Male Athletes Is No Different To That In Men With No History Of Physical Training. J. Am. Heart Assoc. 2019;8:E012670. doi: 10.1161/JAHA.119.012670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Mitchell G.F., Parise H., Vita J.A., Larson M.G., Warner E., Keaney J.F., et al. Local Shear Stress And Brachial Artery Flow-Mediated Dilation: The Framingham Heart Study. Hypertension. 2004;44:134–139. doi: 10.1161/01.HYP.0000137305.77635.68. [DOI] [PubMed] [Google Scholar]
- 28.Machin D.R., Bloom S.I., Campbell R.A., Phuong T.T., Gates P.E., Lesniewski L.A., et al. Advanced Age Results In A Diminished Endothelial Glycocalyx. Am. J. Physiol. Heart Circ. Physiol. 2018;315:H531–H539. doi: 10.1152/ajpheart.00104.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Groen B.B., Hamer H.M., Snijders T., Van Kranenburg J., Frijns D., Vink H., et al. Skeletal Muscle Capillary Density And Microvascular Function Are Compromised With Aging And Type 2 Diabetes. J. Appl. Physiol. 2014;1985(116):998–1005. doi: 10.1152/japplphysiol.00919.2013. [DOI] [PubMed] [Google Scholar]
- 30.Oda K., Okada H., Suzuki A., Tomita H., Kobayashi R., Sumi K., et al. Factors Enhancing Serum Syndecan-1 Concentrations: A Large-Scale Comprehensive Medical Examination. J. Clin. Med. 2019;8 doi: 10.3390/jcm8091320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mahmoud M., Mayer M., Cancel L.M., Bartosch A.M., Mathews R., Tarbell J.M. The Glycocalyx Core Protein Glypican 1 Protects Vessel Wall Endothelial Cells From Stiffness-Mediated Dysfunction And Disease. Cardiovasc. Res. 2021;117:1592–1605. doi: 10.1093/cvr/cvaa201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Williamson K.A., Hamilton A., Reynolds J.A., Sipos P., Crocker I., Stringer S.E., et al. Age-Related Impairment Of Endothelial Progenitor Cell Migration Correlates With Structural Alterations Of Heparan Sulfate Proteoglycans. Aging Cell. 2013;12:139–147. doi: 10.1111/acel.12031. [DOI] [PubMed] [Google Scholar]
- 33.Liu Y.Y., Lee C.H., Dedaj R., Zhao H., Mrabat H., Sheidlin A., et al. High-Molecular-Weight Hyaluronan–A Possible New Treatment For Sepsis-Induced Lung Injury: A Preclinical Study In Mechanically Ventilated Rats. Crit. Care. 2008;12:R102. doi: 10.1186/cc6982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Erusalimsky J.D., Skene C. Mechanisms Of Endothelial Senescence. Exp. Physiol. 2009;94:299–304. doi: 10.1113/expphysiol.2008.043133. [DOI] [PubMed] [Google Scholar]
- 35.Cheung T.M., Yan J.B., Fu J.J., Huang J., Yuan F., Truskey G.A. Endothelial Cell Senescence Increases Traction Forces Due To Age-Associated Changes In The Glycocalyx And Sirt1. Cell. Mol. Bioeng. 2015;8:63–75. doi: 10.1007/s12195-014-0371-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Zu Y., Liu L., Lee M.Y., Xu C., Liang Y., Man R.Y., et al. Sirt1 Promotes Proliferation And Prevents Senescence Through Targeting LKB1 In Primary Porcine Aortic Endothelial Cells. Circ. Res. 2010;106:1384–1393. doi: 10.1161/CIRCRESAHA.109.215483. [DOI] [PubMed] [Google Scholar]
- 37.Lipphardt M., Song J.W., Goligorsky M.S. Sirtuin 1 And Endothelial Glycocalyx. Pflugers Arch. 2020;472:991–1002. doi: 10.1007/s00424-020-02407-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sosinska-Zawierucha P., Mackowiak B., Staniszewski R., Suminska-Jasinska K., Maj M., Krasinski Z., et al. Sulodexide Slows Down The Senescence Of Aortic Endothelial Cells Exposed To Serum From Patients With Peripheral Artery Diseases. Cell. Physiol. Biochem. 2018;45:2225–2232. doi: 10.1159/000488167. [DOI] [PubMed] [Google Scholar]
- 39.Zhang X., Sun D., Song J.W., Zullo J., Lipphardt M., Coneh-Gould L., et al. Endothelial Cell Dysfunction And Glycocalyx - A Vicious Circle. Matrix Biol. 2018;71–72:421–431. doi: 10.1016/j.matbio.2018.01.026. [DOI] [PubMed] [Google Scholar]
- 40.Lahteenvuo J., Rosenzweig A. Effects Of Aging On Angiogenesis. Circ. Res. 2012;110:1252–1264. doi: 10.1161/CIRCRESAHA.111.246116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Alecu C., Gueguen R., Aubry C., Salvi P., Perret-Guillaume C., Ducrocq X., et al. Determinants Of Arterial Stiffness In An Apparently Healthy Population Over 60 Years. J. Hum. Hypertens. 2006;20:749–756. doi: 10.1038/sj.jhh.1002072. [DOI] [PubMed] [Google Scholar]
- 42.Lin L.Y., Liao Y.C., Lin H.F., Lee Y.S., Lin R.T., Hsu C.Y., et al. Determinants Of Arterial Stiffness Progression In A Han-Chinese Population In Taiwan: A 4-Year Longitudinal Follow-Up. BMC Cardiovasc. Disord. 2015;15:100. doi: 10.1186/s12872-015-0093-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Simon A.K., Hollander G.A., Mcmichael A. Evolution Of The Immune System In Humans From Infancy To Old Age. Proc. Biol. Sci. 2015;282:20143085. doi: 10.1098/rspb.2014.3085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lee Y.C., Lin S.J. Neonatal Natural Killer Cell Function: Relevance To Antiviral Immune Defense. Clin. Dev. Immunol. 2013;2013 doi: 10.1155/2013/427696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Ivarsson M.A., Loh L., Marquardt N., Kekalainen E., Berglin L., Bjorkstrom N.K., et al. Differentiation And Functional Regulation Of Human Fetal NK Cells. J. Clin. Invest. 2013;123:3889–3901. doi: 10.1172/JCI68989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Filias A., Theodorou G.L., Mouzopoulou S., Varvarigou A.A., Mantagos S., Karakantza M. Phagocytic Ability Of Neutrophils And Monocytes In Neonates. Bmc Pediatr. 2011;11:29. doi: 10.1186/1471-2431-11-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Nussbaum C., Gloning A., Pruenster M., Frommhold D., Bierschenk S., Genzel-Boroviczeny O., et al. Neutrophil And Endothelial Adhesive Function During Human Fetal Ontogeny. J. Leukoc. Biol. 2013;93:175–184. doi: 10.1189/jlb.0912468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Forster-Waldl E., Sadeghi K., Tamandl D., Gerhold B., Hallwirth U., Rohrmeister K., et al. Monocyte Toll-Like Receptor 4 Expression And LPS-Induced Cytokine Production Increase During Gestational Aging. Pediatr. Res. 2005;58:121–124. doi: 10.1203/01.PDR.0000163397.53466.0F. [DOI] [PubMed] [Google Scholar]
- 49.Sadeghi K., Berger A., Langgartner M., Prusa A.R., Hayde M., Herkner K., et al. Immaturity Of Infection Control In Preterm And Term Newborns Is Associated With Impaired Toll-Like Receptor Signaling. J. Infect. Dis. 2007;195:296–302. doi: 10.1086/509892. [DOI] [PubMed] [Google Scholar]
- 50.P.G. Holt, The Role Of Genetic And Environmental Factors In The Development Of T-Cell Mediated Allergic Disease In Early Life. Paediatr. Respir. Rev. (Suppl 5A) (2004), S27-30. [DOI] [PubMed]
- 51.Mold J.E., Venkatasubrahmanyam S., Burt T.D., Michaelsson J., Rivera J.M., Galkina S.A., et al. Fetal And Adult Hematopoietic Stem Cells Give Rise To Distinct T Cell Lineages In Humans. Science. 2010;330:1695–1699. doi: 10.1126/science.1196509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hannet I., Erkeller-Yuksel F., Lydyard P., Deneys V., Debruyere M. Developmental And Maturational Changes In Human Blood Lymphocyte Subpopulations. Immunol. Today. 1992;13(215):218. doi: 10.1016/0167-5699(92)90157-3. [DOI] [PubMed] [Google Scholar]
- 53.Lesniewski L.A., Durrant J.R., Connell M.L., Folian B.J., Donato A.J., Seals D.R. Salicylate Treatment Improves Age-Associated Vascular Endothelial Dysfunction: Potential Role Of Nuclear Factor KappaB And Forkhead Box O Phosphorylation. J. Gerontol. A Biol. Sci. Med. Sci. 2011;66:409–418. doi: 10.1093/gerona/glq233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Schouten L.R., Van Kaam A.H., Kohse F., Veltkamp F., Bos L.D., De Beer F.M., et al. Age-Dependent Differences In Pulmonary Host Responses In ARDS: A Prospective Observational Cohort Study. Ann. Intensive Care. 2019;9:55. doi: 10.1186/s13613-019-0529-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Yan S.F., Tritto I., Pinsky D., Liao H., Huang J., Fuller G., et al. Induction Of Interleukin 6 (IL-6) By Hypoxia In Vascular Cells. Central Role Of The Binding Site For Nuclear Factor-IL-6. J. Biol. Chem. 1995;270:11463–11471. doi: 10.1074/jbc.270.19.11463. [DOI] [PubMed] [Google Scholar]
- 56.Lee P., Chandel N.S., Simon M.C. Cellular Adaptation To Hypoxia Through Hypoxia Inducible Factors And Beyond. Nat. Rev. Mol. Cell Biol. 2020;21:268–283. doi: 10.1038/s41580-020-0227-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Imtiyaz H.Z., Williams E.P., Hickey M.M., Patel S.A., Durham A.C., Yuan L.J., et al. Hypoxia-Inducible Factor 2alpha Regulates Macrophage Function In Mouse Models Of Acute And Tumor Inflammation. J. Clin. Invest. 2010;120:2699–2714. doi: 10.1172/JCI39506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Yilmaz E., Ustundag B., Sen Y., Akarsu S., Kurt A.N., Dogan Y. The Levels Of Ghrelin, TNF-Alpha, And IL-6 In Children With Cyanotic And Acyanotic Congenital Heart Disease. Mediators Inflamm. 2007;2007:32403. doi: 10.1155/2007/32403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Jackson-Weaver O., Friedman J.K., Rodriguez L.A., Hoof M.A., Drury R.H., Packer J.T., et al. Hypoxia/Reoxygenation Decreases Endothelial Glycocalyx Via Reactive Oxygen Species And Calcium Signaling In A Cellular Model For Shock. J. Trauma Acute Care Surg. 2019;87:1070–1076. doi: 10.1097/TA.0000000000002427. [DOI] [PubMed] [Google Scholar]
- 60.Padberg J.S., Wiesinger A., Di Marco G.S., Reuter S., Grabner A., Kentrup D., et al. Damage Of The Endothelial Glycocalyx In Chronic Kidney Disease. Atherosclerosis. 2014;234:335–343. doi: 10.1016/j.atherosclerosis.2014.03.016. [DOI] [PubMed] [Google Scholar]
- 61.Yasmin, Falzone R., Brown M.J. Determinants Of Arterial Stiffness In Offspring Of Families With Essential Hypertension. Am. J. Hypertens. 2004;17:292–298. doi: 10.1016/j.amjhyper.2003.12.002. [DOI] [PubMed] [Google Scholar]
- 62.Elias M.F., Crichton G.E., Abhayaratna W.P. Interactions Between Plasma Homocysteine And Arterial Stiffness In Chronic Kidney Disease In Community-Dwelling Individuals: The Maine-Syracuse Study. J. Hum. Hypertens. 2015;29:726–731. doi: 10.1038/jhh.2015.17. [DOI] [PubMed] [Google Scholar]
- 63.Vlahu C.A., Lemkes B.A., Struijk D.G., Koopman M.G., Krediet R.T., Vink H. Damage Of The Endothelial Glycocalyx In Dialysis Patients. J. Am. Soc. Nephrol. 2012;23:1900–1908. doi: 10.1681/ASN.2011121181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Nussbaum C., Cavalcanti Fernandes Heringa A., Mormanova Z., Puchwein-Schwepcke A.F., Bechtold-Dalla Pozza S., Genzel-Boroviczeny O. Early Microvascular Changes With Loss Of The Glycocalyx In Children With Type 1 Diabetes. J. Pediatr. 2014;164(584–589):E581. doi: 10.1016/j.jpeds.2013.11.016. [DOI] [PubMed] [Google Scholar]
- 65.Broekhuizen L.N., Lemkes B.A., Mooij H.L., Meuwese M.C., Verberne H., Holleman F., et al. Effect Of Sulodexide On Endothelial Glycocalyx And Vascular Permeability In Patients With Type 2 Diabetes Mellitus. Diabetologia. 2010;53:2646–2655. doi: 10.1007/s00125-010-1910-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Nieuwdorp M., Van Haeften T.W., Gouverneur M.C., Mooij H.L., Van Lieshout M.H., Levi M., et al. Loss Of Endothelial Glycocalyx During Acute Hyperglycemia Coincides With Endothelial Dysfunction And Coagulation Activation In Vivo. Diabetes. 2006;55:480–486. doi: 10.2337/diabetes.55.02.06.db05-1103. [DOI] [PubMed] [Google Scholar]
- 67.Wang F., Wang Y., Kim M.S., Puthanveetil P., Ghosh S., Luciani D.S., et al. Glucose-Induced Endothelial Heparanase Secretion Requires Cortical And Stress Actin Reorganization. Cardiovasc. Res. 2010;87:127–136. doi: 10.1093/cvr/cvq051. [DOI] [PubMed] [Google Scholar]
- 68.Lopez-Quintero S.V., Cancel L.M., Pierides A., Antonetti D., Spray D.C., Tarbell J.M. High Glucose Attenuates Shear-Induced Changes In Endothelial Hydraulic Conductivity By Degrading The Glycocalyx. PLoS ONE. 2013;8:E78954. doi: 10.1371/journal.pone.0078954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Brownlee M. Biochemistry And Molecular Cell Biology Of Diabetic Complications. Nature. 2001;414:813–820. doi: 10.1038/414813a. [DOI] [PubMed] [Google Scholar]
- 70.Katsumura C., Sugiyama T., Nakamura K., Obayashi H., Hasegawa G., Oku H., et al. Effects Of Advanced Glycation End Products On Hyaluronan Photolysis: A New Mechanism Of Diabetic Vitreopathy. Ophthalmic Res. 2004;36:327–331. doi: 10.1159/000081635. [DOI] [PubMed] [Google Scholar]
- 71.Wu H., Ballantyne C.M. Metabolic Inflammation And Insulin Resistance In Obesity. Circ. Res. 2020;126:1549–1564. doi: 10.1161/CIRCRESAHA.119.315896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Eskens B.J., Leurgans T.M., Vink H., Vanteeffelen J.W. Early Impairment Of Skeletal Muscle Endothelial Glycocalyx Barrier Properties In Diet-Induced Obesity In Mice. Physiol. Rep. 2014;2:E00194. doi: 10.1002/phy2.194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Cinti S., Mitchell G., Barbatelli G., Murano I., Ceresi E., Faloia E., et al. Adipocyte Death Defines Macrophage Localization And Function In Adipose Tissue Of Obese Mice And Humans. J. Lipid Res. 2005;46:2347–2355. doi: 10.1194/jlr.M500294-JLR200. [DOI] [PubMed] [Google Scholar]
- 74.Ellulu M.S., Patimah I., Khaza'ai H., Rahmat A., Abed Y. Obesity And Inflammation: The Linking Mechanism And The Complications. Arch. Med. Sci. 2017;13:851–863. doi: 10.5114/aoms.2016.58928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Lafontan M. Fat Cells: Afferent And Efferent Messages Define New Approaches To Treat Obesity. Annu. Rev. Pharmacol. Toxicol. 2005;45:119–146. doi: 10.1146/annurev.pharmtox.45.120403.095843. [DOI] [PubMed] [Google Scholar]
- 76.Payne G.A., Kohr M.C., Tune J.D. Epicardial Perivascular Adipose Tissue As A Therapeutic Target In Obesity-Related Coronary Artery Disease. Br. J. Pharmacol. 2012;165:659–669. doi: 10.1111/j.1476-5381.2011.01370.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Hill D.A., Lim H.W., Kim Y.H., Ho W.Y., Foong Y.H., Nelson V.L., et al. Distinct Macrophage Populations Direct Inflammatory Versus Physiological Changes In Adipose Tissue. Proc. Natl. Acad. Sci. U.S.A. 2018;115:E5096–E5105. doi: 10.1073/pnas.1802611115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Gordts P., Esko J.D. The Heparan Sulfate Proteoglycan Grip On Hyperlipidemia And Atherosclerosis. Matrix Biol. 2018;71–72:262–282. doi: 10.1016/j.matbio.2018.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Van Den Berg B.M., Spaan J.A., Vink H. Impaired Glycocalyx Barrier Properties Contribute To Enhanced Intimal Low-Density Lipoprotein Accumulation At The Carotid Artery Bifurcation In Mice. Pflugers Arch. 2009;457:1199–1206. doi: 10.1007/s00424-008-0590-6. [DOI] [PubMed] [Google Scholar]
- 80.Grootaert M.O.J., Bennett M.R. Vascular Smooth Muscle Cells In Atherosclerosis: Time For A Re-Assessment. Cardiovasc. Res. 2021;117:2326–2339. doi: 10.1093/cvr/cvab046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Xue X.J., Jiang Y., Chen L., Chen S.L. Relationship Between The Endothelial Glycocalyx And The Extent Of Coronary Atherosclerosis. Microcirculation. 2018;25:E12504. doi: 10.1111/micc.12504. [DOI] [PubMed] [Google Scholar]
- 82.Gorshkov A.Y., Klimushina M.V., Boytsov S.A., Kots A.Y., Gumanova N.G. Increase In Perfused Boundary Region Of Endothelial Glycocalyx Is Associated With Higher Prevalence Of Ischemic Heart Disease And Lesions Of Microcirculation And Vascular Wall. Microcirculation. 2018;25:E12454. doi: 10.1111/micc.12454. [DOI] [PubMed] [Google Scholar]
- 83.Cunningham K.S., Gotlieb A.I. The Role Of Shear Stress In The Pathogenesis Of Atherosclerosis. Lab. Invest. 2005;85:9–23. doi: 10.1038/labinvest.3700215. [DOI] [PubMed] [Google Scholar]
- 84.Chatterjee S. Endothelial Mechanotransduction, Redox Signaling And The Regulation Of Vascular Inflammatory Pathways. Front. Physiol. 2018;9:524. doi: 10.3389/fphys.2018.00524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Mensah S.A., Nersesyan A.A., Harding I.C., Lee C.I., Tan X., Banerjee S., et al. Flow-Regulated Endothelial Glycocalyx Determines Metastatic Cancer Cell Activity. FASEB J. 2020;34:6166–6184. doi: 10.1096/fj.201901920R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Van Beers E.J., Nieuwdorp M., Duits A.J., Evers L.M., Schnog J.J., Biemond B.J., et al. Sickle Cell Patients Are Characterized By A Reduced Glycocalyx Volume. Haematologica. 2008;93:307–308. doi: 10.3324/haematol.12027. [DOI] [PubMed] [Google Scholar]
- 87.Sparkenbaugh E., Pawlinski R. Interplay Between Coagulation And Vascular Inflammation In Sickle Cell Disease. Br. J. Haematol. 2013;162:3–14. doi: 10.1111/bjh.12336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Nasimuzzaman M., Malik P. Role Of The Coagulation System In The Pathogenesis Of Sickle Cell Disease. Blood Adv. 2019;3:3170–3180. doi: 10.1182/bloodadvances.2019000193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Hofer J., Rosales A., Fischer C., Giner T. Extra-Renal Manifestations Of Complement-Mediated Thrombotic Microangiopathies. Front. Pediatr. 2014;2:97. doi: 10.3389/fped.2014.00097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Weiss S.L., Fitzgerald J.C., Pappachan J., Wheeler D., Jaramillo-Bustamante J.C., Salloo A., et al. Global Epidemiology Of Pediatric Severe Sepsis: The Sepsis Prevalence, Outcomes, And Therapies Study. Am. J. Respir. Crit. Care Med. 2015;191:1147–1157. doi: 10.1164/rccm.201412-2323OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Fleischmann-Struzek C., Goldfarb D.M., Schlattmann P., Schlapbach L.J., Reinhart K., Kissoon N. The Global Burden Of Paediatric And Neonatal Sepsis: A Systematic Review. Lancet Respir. Med. 2018;6:223–230. doi: 10.1016/S2213-2600(18)30063-8. [DOI] [PubMed] [Google Scholar]
- 92.Kissoon N., Reinhart K., Daniels R., Machado M.F.R., Schachter R.D., Finfer S. Sepsis In Children: Global Implications Of The World Health Assembly Resolution On Sepsis. Pediatr Crit. Care Med. 2017;18:E625–E627. doi: 10.1097/PCC.0000000000001340. [DOI] [PubMed] [Google Scholar]
- 93.Singer M., Deutschman C.S., Seymour C.W., Shankar-Hari M., Annane D., Bauer M., et al. The Third International Consensus Definitions For Sepsis And Septic Shock (Sepsis-3) JAMA. 2016;315:801–810. doi: 10.1001/jama.2016.0287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Iba T., Levy J.H. Derangement Of The Endothelial Glycocalyx In Sepsis. J. Thromb. Haemost. 2019;17:283–294. doi: 10.1111/jth.14371. [DOI] [PubMed] [Google Scholar]
- 95.Uchimido R., Schmidt E.P., Shapiro N.I. The Glycocalyx: A Novel Diagnostic And Therapeutic Target In Sepsis. Crit. Care. 2019;23:16. doi: 10.1186/s13054-018-2292-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Goligorsky M.S., Sun D. Glycocalyx In Endotoxemia And Sepsis. Am. J. Pathol. 2020;190:791–798. doi: 10.1016/j.ajpath.2019.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Macdonald J., Galley H.F., Webster N.R. Oxidative Stress And Gene Expression In Sepsis. Br. J. Anaesth. 2003;90:221–232. doi: 10.1093/bja/aeg034. [DOI] [PubMed] [Google Scholar]
- 98.Van Golen R.F., Van Gulik T.M., Heger M. Mechanistic Overview Of Reactive Species-Induced Degradation Of The Endothelial Glycocalyx During Hepatic Ischemia/Reperfusion Injury. Free Radic. Biol. Med. 2012;52:1382–1402. doi: 10.1016/j.freeradbiomed.2012.01.013. [DOI] [PubMed] [Google Scholar]
- 99.Sun C., Beard R.S., Mclean D.L., Rigor R.R., Konia T., Wu M.H., et al. ADAM15 Deficiency Attenuates Pulmonary Hyperpermeability And Acute Lung Injury In Lipopolysaccharide-Treated Mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013;304:L135–142. doi: 10.1152/ajplung.00133.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Yang X., Meegan J.E., Jannaway M., Coleman D.C., Yuan S.Y. A Disintegrin And Metalloproteinase 15-Mediated Glycocalyx Shedding Contributes To Vascular Leakage During Inflammation. Cardiovasc. Res. 2018;114:1752–1763. doi: 10.1093/cvr/cvy167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Koo A., Dewey C.F., Jr., Garcia-Cardena G. Hemodynamic Shear Stress Characteristic Of Atherosclerosis-Resistant Regions Promotes Glycocalyx Formation In Cultured Endothelial Cells. Am. J. Physiol. Cell Physiol. 2013;304:C137–146. doi: 10.1152/ajpcell.00187.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Wang G., Kostidis S., Tiemeier G.L., Sol W., De Vries M.R., Giera M., et al. Shear Stress Regulation Of Endothelial Glycocalyx Structure Is Determined By Glucobiosynthesis. Arterioscler. Thromb. Vasc. Biol. 2020;40:350–364. doi: 10.1161/ATVBAHA.119.313399. [DOI] [PubMed] [Google Scholar]
- 103.Lupu F., Kinasewitz G., Dormer K. The Role Of Endothelial Shear Stress On Haemodynamics, Inflammation, Coagulation And Glycocalyx During Sepsis. J. Cell Mol. Med. 2020;24:12258–12271. doi: 10.1111/jcmm.15895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Richter R.P., Zheng L., Ashtekar A.R., Walker S.C., Pittet J.F., Richter J.R. Associations Of Plasma Angiopoietins-1 And -2 And Angiopoietin-2/-1 Ratios With Measures Of Organ Injury And Clinical Outcomes In Children With Sepsis: A Preliminary Report. Pediatr. Crit. Care Med. 2020;21:E874–E878. doi: 10.1097/PCC.0000000000002508. [DOI] [PubMed] [Google Scholar]
- 105.Suzuki K., Okada H., Takemura G., Takada C., Kuroda A., Yano H., et al. Neutrophil Elastase Damages The Pulmonary Endothelial Glycocalyx In Lipopolysaccharide-Induced Experimental Endotoxemia. Am. J. Pathol. 2019;189:1526–1535. doi: 10.1016/j.ajpath.2019.05.002. [DOI] [PubMed] [Google Scholar]
- 106.Pudjiadi A.H., Saidah F., Alatas F.S. Correlation Between Syndecan-1 Level And PELOD-2 Score And Mortality In Pediatric Sepsis. Rev. Bras. Ter. Intensiva. 2022;33:549–556. doi: 10.5935/0103-507X.20210083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Leteurtre S., Duhamel A., Salleron J., Grandbastien B., Lacroix J., Leclerc F., et al. PELOD-2: An Update Of The Pediatric Logistic Organ Dysfunction Score. Crit. Care Med. 2013;41:1761–1773. doi: 10.1097/CCM.0b013e31828a2bbd. [DOI] [PubMed] [Google Scholar]
- 108.Fernandez-Sarmiento J., Salazar-Pelaez L.M., Carcillo J.A. The Endothelial Glycocalyx: A Fundamental Determinant Of Vascular Permeability In Sepsis. Pediatr. Crit. Care Med. 2020;21:E291–E300. doi: 10.1097/PCC.0000000000002266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Samies N.L., James S.H., Kimberlin D.W. Neonatal Herpes Simplex Virus Disease: Updates And Continued Challenges. Clin. Perinatol. 2021;48:263–274. doi: 10.1016/j.clp.2021.03.003. [DOI] [PubMed] [Google Scholar]
- 110.Hadigal S., Koganti R., Yadavalli T., Agelidis A., Suryawanshi R., Shukla D. Heparanase-Regulated Syndecan-1 Shedding Facilitates Herpes Simplex Virus 1 Egress. J. Virol. 2020;94 doi: 10.1128/JVI.01672-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Verhagen L.M., De Groot R. Dengue In Children. J. Infect. 2014;69(Suppl 1):S77–86. doi: 10.1016/j.jinf.2014.07.020. [DOI] [PubMed] [Google Scholar]
- 112.Suwarto S., Sasmono R.T., Sinto R., Ibrahim E., Suryamin M. Association Of Endothelial Glycocalyx And Tight And Adherens Junctions With Severity Of Plasma Leakage In Dengue Infection. J. Infect. Dis. 2017;215:992–999. doi: 10.1093/infdis/jix041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Puerta-Guardo H., Glasner D.R., Harris E. Dengue Virus NS1 Disrupts The Endothelial Glycocalyx, Leading To Hyperpermeability. PLoS Pathog. 2016;12:E1005738. doi: 10.1371/journal.ppat.1005738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.E. Lyimo, L.E. Haslund, T. Ramsing, C.W. Wang, A.M. Efunshile, A. Manjurano, et al. In Vivo Imaging Of The Buccal Mucosa Shows Loss Of The Endothelial Glycocalyx And Perivascular Hemorrhages In Pediatric Plasmodium Falciparum Malaria. Infect. Immun. 88, (2020). [DOI] [PMC free article] [PubMed]
- 115.Bush M.A., Florence S.M., Yeo T.W., Kalingonji A.R., Chen Y., Granger D.L., et al. Degradation Of Endothelial Glycocalyx In Tanzanian Children With Falciparum Malaria. FASEB J. 2021;35:E21805. doi: 10.1096/fj.202100277RR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Mawson A.R. The Pathogenesis Of Malaria: A New Perspective. Pathog. Glob. Health. 2013;107:122–129. doi: 10.1179/2047773213Y.0000000084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Milner D.A. Malaria Pathogenesis. Cold Spring Harb. Perspect Med. 2018;8 doi: 10.1101/cshperspect.a025569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Plewes K., Turner G.D.H., Dondorp A.M. Pathophysiology, Clinical Presentation, And Treatment Of Coma And Acute Kidney Injury Complicating Falciparum Malaria. Curr. Opin Infect. Dis. 2018;31:69–77. doi: 10.1097/QCO.0000000000000419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Viebig N.K., Wulbrand U., Forster R., Andrews K.T., Lanzer M., Knolle P.A. Direct Activation Of Human Endothelial Cells By Plasmodium Falciparum-Infected Erythrocytes. Infect. Immun. 2005;73:3271–3277. doi: 10.1128/IAI.73.6.3271-3277.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Moxon C.A., Wassmer S.C., Milner D.A., Chisala N.V., Taylor T.E., Seydel K.B., et al. Loss Of Endothelial Protein C Receptors Links Coagulation And Inflammation To Parasite Sequestration In Cerebral Malaria In African Children. Blood. 2013;122:842–851. doi: 10.1182/blood-2013-03-490219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Milner D.A., Whitten R.O., Kamiza S., Carr R., Liomba G., Dzamalala C., et al. The Systemic Pathology Of Cerebral Malaria In African Children. Front. Cell. Infect. Microbiol. 2014;4:104. doi: 10.3389/fcimb.2014.00104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Yeo T.W., Bush P.A., Chen Y., Young S.P., Zhang H., Millington D.S., et al. Glycocalyx Breakdown Is Increased In African Children With Cerebral And Uncomplicated Falciparum Malaria. FASEB J. 2019;33:14185–14193. doi: 10.1096/fj.201901048RR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Yeo T.W., Weinberg J.B., Lampah D.A., Kenangalem E., Bush P., Chen Y., et al. Glycocalyx Breakdown Is Associated With Severe Disease And Fatal Outcome In Plasmodium Falciparum Malaria. Clin. Infect. Dis. 2019;69:1712–1720. doi: 10.1093/cid/ciz038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Katsoulis O., Georgiadou A., Cunnington A.J. Immunopathology Of Acute Kidney Injury In Severe Malaria. Front. Immunol. 2021;12 doi: 10.3389/fimmu.2021.651739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Weiss S.L., Peters M.J., Alhazzani W., Agus M.S.D., Flori H.R., Inwald D.P., et al. Surviving Sepsis Campaign International Guidelines For The Management Of Septic Shock And Sepsis-Associated Organ Dysfunction In Children. Intensive Care Med. 2020;46:10–67. doi: 10.1007/s00134-019-05878-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Evans L., Rhodes A., Alhazzani W., Antonelli M., Coopersmith C.M., French C., et al. Surviving Sepsis Campaign: International Guidelines For Management Of Sepsis And Septic Shock 2021. Intensive Care Med. 2021;47:1181–1247. doi: 10.1007/s00134-021-06506-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Smart L., Hughes D. The Effects Of Resuscitative Fluid Therapy On The Endothelial Surface Layer. Front. Vet. Sci. 2021;8 doi: 10.3389/fvets.2021.661660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Johansson P.I., Stensballe J., Ostrowski S.R. Shock Induced Endotheliopathy (SHINE) In Acute Critical Illness - A Unifying Pathophysiologic Mechanism. Crit. Care. 2017;21:25. doi: 10.1186/s13054-017-1605-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Cosenza L., Donati A., Cecero E., Carlucci M., Adembri C., Fusi F. Effects Of Albumin Infusion On LPS-Induced Damage Of Mesenteric Microcirculation. Eur. J. Anaesthesiol. 2012;29:176–177. [Google Scholar]
- 130.Damiani E., Ince C., Orlando F., Pierpaoli E., Cirioni O., Giacometti A., et al. Effects Of The Infusion Of 4% Or 20% Human Serum Albumin On The Skeletal Muscle Microcirculation In Endotoxemic Rats. PLoS ONE. 2016;11:E0151005. doi: 10.1371/journal.pone.0151005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Van Der Heijden M., Verheij J., Van Nieuw Amerongen G.P., Groeneveld A.B. Crystalloid Or Colloid Fluid Loading And Pulmonary Permeability, Edema, And Injury In Septic And Nonseptic Critically Ill Patients With Hypovolemia. Crit. Care Med. 2009;37:1275–1281. doi: 10.1097/CCM.0b013e31819cedfd. [DOI] [PubMed] [Google Scholar]
- 132.Haase N., Perner A., Hennings L.I., Siegemund M., Lauridsen B., Wetterslev M., et al. Hydroxyethyl Starch 130/0.38-0.45 Versus Crystalloid Or Albumin In Patients With Sepsis: Systematic Review With Meta-Analysis And Trial Sequential Analysis. BMJ. 2013;346:F839. doi: 10.1136/bmj.f839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Rochwerg B., Alhazzani W., Gibson A., Ribic C.M., Sindi A., Heels-Ansdell D., et al. Fluid Type And The Use Of Renal Replacement Therapy In Sepsis: A Systematic Review And Network Meta-Analysis. Intensive Care Med. 2015;41:1561–1571. doi: 10.1007/s00134-015-3794-1. [DOI] [PubMed] [Google Scholar]
- 134.Finfer S., Bellomo R., Boyce N., French J., Myburgh J., Norton R., et al. A Comparison Of Albumin And Saline For Fluid Resuscitation In The Intensive Care Unit. N. Engl. J. Med. 2004;350:2247–2256. doi: 10.1056/NEJMoa040232. [DOI] [PubMed] [Google Scholar]
- 135.Caironi P., Tognoni G., Masson S., Fumagalli R., Pesenti A., Romero M., et al. Albumin Replacement In Patients With Severe Sepsis Or Septic Shock. N. Engl. J. Med. 2014;370:1412–1421. doi: 10.1056/NEJMoa1305727. [DOI] [PubMed] [Google Scholar]
- 136.Lewis S.R., Pritchard M.W., Evans D.J., Butler A.R., Alderson P., Smith A.F., et al. Colloids Versus Crystalloids For Fluid Resuscitation In Critically Ill People. Cochrane Database Syst. Rev. 2018;8:Cd000567. doi: 10.1002/14651858.CD000567.pub7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Martin G.S., Bassett P. Crystalloids vs. Colloids For Fluid Resuscitation In The Intensive Care Unit: A Systematic Review And Meta-Analysis. J. Crit. Care. 2019;50:144–154. doi: 10.1016/j.jcrc.2018.11.031. [DOI] [PubMed] [Google Scholar]
- 138.Torres L.N., Sondeen J.L., Ji L., Dubick M.A., Torres Filho I. Evaluation Of Resuscitation Fluids On Endothelial Glycocalyx, Venular Blood Flow, And Coagulation Function After Hemorrhagic Shock In Rats. J. Trauma Acute Care Surg. 2013;75:759–766. doi: 10.1097/TA.0b013e3182a92514. [DOI] [PubMed] [Google Scholar]
- 139.Fialkowski A., Gernez Y., Arya P., Weinacht K.G., Kinane T.B., Yonker L.M. Insight Into The Pediatric And Adult Dichotomy Of COVID-19: Age-Related Differences In The Immune Response To SARS-CoV-2 Infection. Pediatr. Pulmonol. 2020;55:2556–2564. doi: 10.1002/ppul.24981. [DOI] [PubMed] [Google Scholar]
- 140.Shekerdemian L.S., Mahmood N.R., Wolfe K.K., Riggs B.J., Ross C.E., Mckiernan C.A., et al. Characteristics And Outcomes Of Children With Coronavirus Disease 2019 (COVID-19) Infection Admitted To US And Canadian Pediatric Intensive Care Units. JAMA Pediatr. 2020;174:868–873. doi: 10.1001/jamapediatrics.2020.1948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Hoffmann M., Kleine-Weber H., Schroeder S., Kruger N., Herrler T., Erichsen S., et al. SARS-CoV-2 Cell Entry Depends On ACE2 And TMPRSS2 And Is Blocked By A Clinically Proven Protease Inhibitor. Cell. 2020;181(271–280):E278. doi: 10.1016/j.cell.2020.02.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Clausen T.M., Sandoval D.R., Spliid C.B., Pihl J., Perrett H.R., Painter C.D., et al. SARS-CoV-2 Infection Depends On Cellular Heparan Sulfate And ACE2. Cell. 2020;183(1043–1057):E1015. doi: 10.1016/j.cell.2020.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Loske J., Rohmel J., Lukassen S., Stricker S., Magalhaes V.G., Liebig J., et al. Pre-Activated Antiviral Innate Immunity In The Upper Airways Controls Early SARS-CoV-2 Infection In Children. Nat. Biotechnol. 2021 doi: 10.1038/s41587-021-01037-9. [DOI] [PubMed] [Google Scholar]
- 144.Carsetti R., Quintarelli C., Quinti I., Piano Mortari E., Zumla A., Ippolito G., et al. The Immune System Of Children: The Key To Understanding SARS-CoV-2 Susceptibility? Lancet Child Adolesc. Health. 2020;4:414–416. doi: 10.1016/S2352-4642(20)30135-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Varga Z., Flammer A.J., Steiger P., Haberecker M., Andermatt R., Zinkernagel A.S., et al. Endothelial Cell Infection And Endotheliitis In COVID-19. Lancet. 2020;395:1417–1418. doi: 10.1016/S0140-6736(20)30937-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Zuo Y., Yalavarthi S., Shi H., Gockman K., Zuo M., Madison J.A., et al. Neutrophil Extracellular Traps In COVID-19. JCI Insight. 2020:5. doi: 10.1172/jci.insight.138999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Zuo Y., Zuo M., Yalavarthi S., Gockman K., Madison J.A., Shi H., et al. Neutrophil Extracellular Traps And Thrombosis In COVID-19. J. Thromb. Thrombolysis. 2021;51:446–453. doi: 10.1007/s11239-020-02324-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Hottz E.D., Azevedo-Quintanilha I.G., Palhinha L., Teixeira L., Barreto E.A., Pao C.R.R., et al. Platelet Activation And Platelet-Monocyte Aggregate Formation Trigger Tissue Factor Expression In Patients With Severe COVID-19. Blood. 2020;136:1330–1341. doi: 10.1182/blood.2020007252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Stahl K., Gronski P.A., Kiyan Y., Seeliger B., Bertram A., Pape T., et al. Injury To The Endothelial Glycocalyx In Critically Ill Patients With COVID-19. Am. J. Respir. Crit. Care Med. 2020;202:1178–1181. doi: 10.1164/rccm.202007-2676LE. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Queisser K.A., Mellema R.A., Middleton E.A., Portier I., Manne B.K., Denorme F., et al. COVID-19 Generates Hyaluronan Fragments That Directly Induce Endothelial Barrier Dysfunction. JCI Insight. 2021;6 doi: 10.1172/jci.insight.147472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Fernandez S., Moreno-Castano A.B., Palomo M., Martinez-Sanchez J., Torramade-Moix S., Tellez A., et al. Distinctive Biomarker Features In The Endotheliopathy Of COVID-19 And Septic Syndromes. Shock. 2021 doi: 10.1097/SHK.0000000000001823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Goshua G., Pine A.B., Meizlish M.L., Chang C.H., Zhang H., Bahel P., et al. Endotheliopathy In COVID-19-Associated Coagulopathy: Evidence From A Single-Centre, Cross-Sectional Study. Lancet Haematol. 2020;7:E575–E582. doi: 10.1016/S2352-3026(20)30216-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Tong M., Jiang Y., Xia D., Xiong Y., Zheng Q., Chen F., et al. Elevated Expression Of Serum Endothelial Cell Adhesion Molecules In COVID-19 Patients. J. Infect. Dis. 2020;222:894–898. doi: 10.1093/infdis/jiaa349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Buijsers B., Yanginlar C., De Nooijer A., Grondman I., Maciej-Hulme M.L., Jonkman I., et al. Increased Plasma Heparanase Activity In COVID-19 Patients. Front. Immunol. 2020;11 doi: 10.3389/fimmu.2020.575047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Fraser D.D., Patterson E.K., Daley M., Cepinskas G. Case Report: Inflammation And Endothelial Injury Profiling Of COVID-19 Pediatric Multisystem Inflammatory Syndrome (MIS-C) Front. Pediatr. 2021;9 doi: 10.3389/fped.2021.597926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Fernandez-Sarmiento J., Florez S., Alarcon-Forero L.C., Salazar-Pelaez L.M., Garcia-Casallas J., Mulett H., et al. Case Report: Endothelial Glycocalyx Damage In Critically Ill Patients With SARS-CoV-2-Related Multisystem Inflammatory Syndrome (MIS-C) Front. Pediatr. 2021;9 doi: 10.3389/fped.2021.726949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Borrelli M., Corcione A., Castellano F., Fiori Nastro F., Santamaria F. Coronavirus Disease 2019 In Children. Front. Pediatr. 2021;9 doi: 10.3389/fped.2021.668484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Global Health Estimates 2020: Deaths By Cause, Age, Sex, By Country And By Region, 2000-2019. Geneva, World Health Organization; 2020. [Available from: https://www.who.int/data/gho/data/themes/mortality-and-global-health-estimates/ghe-leading-causes-of-death].
- 159.Kissoon N., Dreyer J., Walia M. Pediatric Trauma: Differences In Pathophysiology, Injury Patterns And Treatment Compared With Adult Trauma. CMAJ. 1990;142:27–34. [PMC free article] [PubMed] [Google Scholar]
- 160.Johansson P.I., Stensballe J., Rasmussen L.S., Ostrowski S.R. A High Admission Syndecan-1 Level, A Marker Of Endothelial Glycocalyx Degradation, Is Associated With Inflammation, Protein C Depletion, Fibrinolysis, And Increased Mortality In Trauma Patients. Ann. Surg. 2011;254:194–200. doi: 10.1097/SLA.0b013e318226113d. [DOI] [PubMed] [Google Scholar]
- 161.Gonzalez Rodriguez E., Ostrowski S.R., Cardenas J.C., Baer L.A., Tomasek J.S., Henriksen H.H., et al. Syndecan-1: A Quantitative Marker For The Endotheliopathy Of Trauma. J. Am. Coll. Surg. 2017;225:419–427. doi: 10.1016/j.jamcollsurg.2017.05.012. [DOI] [PubMed] [Google Scholar]
- 162.Johansson P.I., Henriksen H.H., Stensballe J., Gybel-Brask M., Cardenas J.C., Baer L.A., et al. Traumatic Endotheliopathy: A Prospective Observational Study Of 424 Severely Injured Patients. Ann. Surg. 2017;265:597–603. doi: 10.1097/SLA.0000000000001751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Russell R.T., Christiaans S.C., Nice T.R., Banks M., Mortellaro V.E., Morgan C., et al. Histone-Complexed Dna Fragments Levels Are Associated With Coagulopathy, Endothelial Cell Damage, And Increased Mortality After Severe Pediatric Trauma. Shock. 2018;49:44–52. doi: 10.1097/SHK.0000000000000902. [DOI] [PubMed] [Google Scholar]
- 164.Richter R.P., Russell R.T., Hu P.J., Uhlich R.M., Swain T.A., Kerby J.D., et al. Plasma Angiopoietin-2/-1 Ratio Is Elevated And Angiopoietin-2 Levels Correlate With Plasma Syndecan-1 Following Pediatric Trauma. Shock. 2019;52:340–346. doi: 10.1097/SHK.0000000000001267. [DOI] [PubMed] [Google Scholar]
- 165.Naumann D.N., Hazeldine J., Davies D.J., Bishop J., Midwinter M.J., Belli A., et al. Endotheliopathy Of Trauma Is An On-Scene Phenomenon, And Is Associated With Multiple Organ Dysfunction Syndrome: A Prospective Observational Study. Shock. 2018;49:420–428. doi: 10.1097/SHK.0000000000000999. [DOI] [PubMed] [Google Scholar]
- 166.Qi F., Zhou H., Gu P., Tang Z.H., Zhu B.F., Chen J.R., et al. Endothelial Glycocalyx Degradation Is Associated With Early Organ Impairment In Polytrauma Patients. BMC Emerg. Med. 2021;21:52. doi: 10.1186/s12873-021-00446-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Rahbar E., Cardenas J.C., Baimukanova G., Usadi B., Bruhn R., Pati S., et al. Endothelial Glycocalyx Shedding And Vascular Permeability In Severely Injured Trauma Patients. J. Transl. Med. 2015;13:117. doi: 10.1186/s12967-015-0481-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Walker S.C., Richter R.P., Zheng L., Ashtekar A.R., Jansen J.O., Kerby J.D., et al. Increased Plasma Hyaluronan Levels Are Associated With Acute Traumatic Coagulopathy. Shock. 2021 doi: 10.1097/SHK.0000000000001867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Calkins C.M., Bensard D.D., Moore E.E., Mcintyre R.C., Silliman C.C., Biffl W., et al. The Injured Child Is Resistant To Multiple Organ Failure: A Different Inflammatory Response? J. Trauma. 2002;53:1058–1063. doi: 10.1097/00005373-200212000-00005. [DOI] [PubMed] [Google Scholar]
- 170.Bayreuther J., Wagener S., Woodford M., Edwards A., Lecky F., Bouamra O., et al. Paediatric Trauma: Injury Pattern And Mortality In The UK. Arch. Dis. Child Educ. Pract. Ed. 2009;94:37–41. doi: 10.1136/adc.2007.132787. [DOI] [PubMed] [Google Scholar]
- 171.Holcomb J.B., Tilley B.C., Baraniuk S., Fox E.E., Wade C.E., Podbielski J.M., et al. Transfusion Of Plasma, Platelets, And Red Blood Cells In A 1:1:1 vs A 1:1:2 Ratio And Mortality In Patients With Severe Trauma: The PROPPR Randomized Clinical Trial. JAMA. 2015;313:471–482. doi: 10.1001/jama.2015.12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Vanzant E.L., Hilton R.E., Lopez C.M., Zhang J., Ungaro R.F., Gentile L.F., et al. Advanced Age Is Associated With Worsened Outcomes And A Unique Genomic Response In Severely Injured Patients With Hemorrhagic Shock. Crit. Care. 2015;19:77. doi: 10.1186/s13054-015-0788-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Halbgebauer R., Braun C.K., Denk S., Mayer B., Cinelli P., Radermacher P., et al. Hemorrhagic Shock Drives Glycocalyx, Barrier And Organ Dysfunction Early After Polytrauma. J. Crit. Care. 2018;44:229–237. doi: 10.1016/j.jcrc.2017.11.025. [DOI] [PubMed] [Google Scholar]
- 174.Polites S.F., Moody S., Williams R.F., Kayton M.L., Alberto E.C., Burd R.S., et al. Timing And Volume Of Crystalloid And Blood Products In Pediatric Trauma: An Eastern Association For The Surgery Of Trauma Multicenter Prospective Observational Study. J. Trauma Acute Care Surg. 2020;89:36–42. doi: 10.1097/TA.0000000000002702. [DOI] [PubMed] [Google Scholar]
- 175.Torres Filho I.P., Torres L.N., Salgado C., Dubick M.A. Plasma Syndecan-1 And Heparan Sulfate Correlate With Microvascular Glycocalyx Degradation In Hemorrhaged Rats After Different Resuscitation Fluids. Am. J. Physiol. Heart Circ. Physiol. 2016;310:H1468–1478. doi: 10.1152/ajpheart.00006.2016. [DOI] [PubMed] [Google Scholar]
- 176.Torres L.N., Chung K.K., Salgado C.L., Dubick M.A., Torres Filho I.P. Low-Volume Resuscitation With Normal Saline Is Associated With Microvascular Endothelial Dysfunction After Hemorrhage In Rats, Compared To Colloids And Balanced Crystalloids. Crit. Care. 2017;21:160. doi: 10.1186/s13054-017-1745-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Pati S., Potter D.R., Baimukanova G., Farrel D.H., Holcomb J.B., Schreiber M.A. Modulating The Endotheliopathy Of Trauma: Factor Concentrate Versus Fresh Frozen Plasma. J. Trauma Acute Care Surg. 2016;80:576–584. doi: 10.1097/TA.0000000000000961. [DOI] [PubMed] [Google Scholar]
- 178.Horstick G., Lauterbach M., Kempf T., Bhakdi S., Heimann A., Horstick M., et al. Early Albumin Infusion Improves Global And Local Hemodynamics And Reduces Inflammatory Response In Hemorrhagic Shock. Crit. Care Med. 2002;30:851–855. doi: 10.1097/00003246-200204000-00023. [DOI] [PubMed] [Google Scholar]
- 179.Zeng Y., Adamson R.H., Curry F.R., Tarbell J.M. Sphingosine-1-Phosphate Protects Endothelial Glycocalyx By Inhibiting Syndecan-1 Shedding. Am. J. Physiol. Heart Circ. Physiol. 2014;306:H363–372. doi: 10.1152/ajpheart.00687.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Zeng Y., Liu X.H., Tarbell J., Fu B. Sphingosine 1-Phosphate Induced Synthesis Of Glycocalyx On Endothelial Cells. Exp. Cell Res. 2015;339:90–95. doi: 10.1016/j.yexcr.2015.08.013. [DOI] [PubMed] [Google Scholar]
- 181.Myburgh J., Cooper D.J., Finfer S., Bellomo R., Norton R., Bishop N., et al. Saline Or Albumin For Fluid Resuscitation In Patients With Traumatic Brain Injury. N. Engl. J. Med. 2007;357:874–884. doi: 10.1056/NEJMoa067514. [DOI] [PubMed] [Google Scholar]
- 182.Ma H.K., Bebawy J.F. Albumin Use In Brain-Injured And Neurosurgical Patients: Concepts, Indications, And Controversies. J. Neurosurg. Anesthesiol. 2021;33:293–299. doi: 10.1097/ANA.0000000000000674. [DOI] [PubMed] [Google Scholar]
- 183.Cooper D.J., Myburgh J., Heritier S., Finfer S., Bellomo R., Billot L., et al. Albumin Resuscitation For Traumatic Brain Injury: Is Intracranial Hypertension The Cause Of Increased Mortality? J. Neurotrauma. 2013;30:512–518. doi: 10.1089/neu.2012.2573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Haywood-Watson R.J., Holcomb J.B., Gonzalez E.A., Peng Z., Pati S., Park P.W., et al. Modulation Of Syndecan-1 Shedding After Hemorrhagic Shock And Resuscitation. PLoS ONE. 2011;6:E23530. doi: 10.1371/journal.pone.0023530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Peng Z., Pati S., Potter D., Brown R., Holcomb J.B., Grill R., et al. Fresh Frozen Plasma Lessens Pulmonary Endothelial Inflammation And Hyperpermeability After Hemorrhagic Shock And Is Associated With Loss Of Syndecan 1. Shock. 2013;40:195–202. doi: 10.1097/SHK.0b013e31829f91fc. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Sperry J.L., Guyette F.X., Brown J.B., Yazer M.H., Triulzi D.J., Early-Young B.J., et al. Prehospital Plasma During Air Medical Transport In Trauma Patients At Risk For Hemorrhagic Shock. N. Engl. J. Med. 2018;379:315–326. doi: 10.1056/NEJMoa1802345. [DOI] [PubMed] [Google Scholar]
- 187.Anand T., Obaid O., Nelson A., Chehab M., Ditillo M., Hammad A., et al. Whole Blood Hemostatic Resuscitation In Pediatric Trauma: A Nationwide Propensity-Matched Analysis. J. Trauma Acute Care Surg. 2021;91:573–578. doi: 10.1097/TA.0000000000003306. [DOI] [PubMed] [Google Scholar]
- 188.Torres L.N., Sondeen J.L., Dubick M.A., Torres Filho I. Systemic And Microvascular Effects Of Resuscitation With Blood Products After Severe Hemorrhage In Rats. J. Trauma Acute Care Surg. 2014;77:716–723. doi: 10.1097/TA.0000000000000448. [DOI] [PubMed] [Google Scholar]
- 189.Kochanek K.D., Xu J., Arias E. Mortality In The United States, NCHS Data Brief No 395. National Center For Health Statistics; Hyattsville, Md: 2019. pp. 1–7. [Google Scholar]
- 190.Romanowski K.S., Palmieri T.L. Pediatric Burn Resuscitation: Past, Present, And Future. Burns Trauma. 2017;5:26. doi: 10.1186/s41038-017-0091-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Luker J.N., Vigiola Cruz M., Carney B.C., Day A., Moffatt L.T., Johnson L.S., et al. Shedding Of The Endothelial Glycocalyx Is Quantitatively Proportional To Burn Injury Severity. Ann. Burns Fire Disasters. 2018;31:17–22. [PMC free article] [PubMed] [Google Scholar]
- 192.Vigiola Cruz M., Carney B.C., Luker J.N., Monger K.W., Vazquez J.S., Moffatt L.T., et al. Plasma Ameliorates Endothelial Dysfunction In Burn Injury. J. Surg. Res. 2019;233:459–466. doi: 10.1016/j.jss.2018.08.027. [DOI] [PubMed] [Google Scholar]
- 193.Osuka A., Kusuki H., Yoneda K., Matsuura H., Matsumoto H., Ogura H., et al. Glycocalyx Shedding Is Enhanced By Age And Correlates With Increased Fluid Requirement In Patients With Major Burns. Shock. 2018;50:60–65. doi: 10.1097/SHK.0000000000001028. [DOI] [PubMed] [Google Scholar]
- 194.Keyloun J.W., Le T.D., Pusateri A.E., Ball R.L., Carney B.C., Orfeo T., et al. Circulating Syndecan-1 And Tissue Factor Pathway Inhibitor, Biomarkers Of Endothelial Dysfunction, Predict Mortality In Burn Patients. Shock. 2021;56:237–244. doi: 10.1097/SHK.0000000000001709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Hippensteel J.A., Uchimido R., Tyler P.D., Burke R.C., Han X., Zhang F., et al. Intravenous Fluid Resuscitation Is Associated With Septic Endothelial Glycocalyx Degradation. Crit. Care. 2019;23:259. doi: 10.1186/s13054-019-2534-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Cote C.J., Drop L.J., Hoaglin D.C., Daniels A.L., Young E.T. Ionized Hypocalcemia After Fresh Frozen Plasma Administration To Thermally Injured Children: Effects Of Infusion Rate, Duration, And Treatment With Calcium Chloride. Anesth. Analg. 1988;67:152–160. [PubMed] [Google Scholar]
- 197.ECMO Registry Of The Extracorporeal Life Support Organization, ELSO, (September 2021).
- 198.Orwoll B.E., Sapru A. Biomarkers In Pediatric ARDS: Future Directions. Front. Pediatr. 2016;4:55. doi: 10.3389/fped.2016.00055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Grundmann S., Fink K., Rabadzhieva L., Bourgeois N., Schwab T., Moser M., et al. Perturbation Of The Endothelial Glycocalyx In Post Cardiac Arrest Syndrome. Resuscitation. 2012;83:715–720. doi: 10.1016/j.resuscitation.2012.01.028. [DOI] [PubMed] [Google Scholar]
- 200.Ali M.M., Mahmoud A.M., Le Master E., Levitan I., Phillips S.A. Role Of Matrix Metalloproteinases And Histone Deacetylase In Oxidative Stress-Induced Degradation Of The Endothelial Glycocalyx. Am. J. Physiol. Heart Circ. Physiol. 2019;316:H647–H663. doi: 10.1152/ajpheart.00090.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Pham K., Parikh K., Heinrich E.C. Hypoxia And Inflammation: Insights From High-Altitude Physiology. Front. Physiol. 2021;12 doi: 10.3389/fphys.2021.676782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Bongoni A.K., Lu B., Mcrae J.L., Salvaris E.J., Toonen E.J.M., Vikstrom I., et al. Complement-Mediated Damage To The Glycocalyx Plays A Role In Renal Ischemia-Reperfusion Injury In Mice. Transplant. Direct. 2019;5:E341. doi: 10.1097/TXD.0000000000000881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Selim J., Hamzaoui M., Boukhalfa I., Djerada Z., Chevalier L., Piton N., et al. Cardiopulmonary Bypass Increases Endothelial Dysfunction After Pulmonary Ischaemia-Reperfusion In An Animal Model. Eur. J. Cardiothorac. Surg. 2021;59:1037–1047. doi: 10.1093/ejcts/ezaa412. [DOI] [PubMed] [Google Scholar]
- 204.Annecke T., Fischer J., Hartmann H., Tschoep J., Rehm M., Conzen P., et al. Shedding Of The Coronary Endothelial Glycocalyx: Effects Of Hypoxia/Reoxygenation vs Ischaemia/Reperfusion. Br. J. Anaesth. 2011;107:679–686. doi: 10.1093/bja/aer269. [DOI] [PubMed] [Google Scholar]
- 205.Rehm M., Bruegger D., Christ F., Conzen P., Thiel M., Jacob M., et al. Shedding Of The Endothelial Glycocalyx In Patients Undergoing Major Vascular Surgery With Global And Regional Ischemia. Circulation. 2007;116:1896–1906. doi: 10.1161/CIRCULATIONAHA.106.684852. [DOI] [PubMed] [Google Scholar]
- 206.Koning N.J., Vonk A.B., Vink H., Boer C. Side-By-Side Alterations In Glycocalyx Thickness And Perfused Microvascular Density During Acute Microcirculatory Alterations In Cardiac Surgery. Microcirculation. 2016;23:69–74. doi: 10.1111/micc.12260. [DOI] [PubMed] [Google Scholar]
- 207.Bruegger D., Brettner F., Rossberg I., Nussbaum C., Kowalski C., Januszewska K., et al. Acute Degradation Of The Endothelial Glycocalyx In Infants Undergoing Cardiac Surgical Procedures. Ann. Thorac. Surg. 2015;99:926–931. doi: 10.1016/j.athoracsur.2014.10.013. [DOI] [PubMed] [Google Scholar]
- 208.Pesonen E., Keski-Nisula J., Andersson S., Palo R., Salminen J., Suominen P.K. High-Dose Methylprednisolone And Endothelial Glycocalyx In Paediatric Heart Surgery. Acta Anaesthesiol. Scand. 2016;60:1386–1394. doi: 10.1111/aas.12785. [DOI] [PubMed] [Google Scholar]
- 209.De Melo Bezerra Cavalcante C.T., Castelo Branco K.M., Pinto Junior V.C., Meneses G.C., De Oliveira Neves F.M., De Souza N.M., et al. Syndecan-1 Improves Severe Acute Kidney Injury Prediction After Pediatric Cardiac Surgery. J. Thorac. Cardiovasc. Surg. 2016;152:178–186. doi: 10.1016/j.jtcvs.2016.03.079. [DOI] [PubMed] [Google Scholar]
- 210.Bangalore H., Carter M.J., Parmar K., Austin C., Shankar-Hari M., Hunt B.J., et al. Degradation Of The Endothelial Glycocalyx Contributes To Metabolic Acidosis In Children Following Cardiopulmonary Bypass Surgery. Pediatr. Crit Care Med. 2021 doi: 10.1097/PCC.0000000000002746. [DOI] [PubMed] [Google Scholar]
- 211.Nussbaum C., Haberer A., Tiefenthaller A., Januszewska K., Chappell D., Brettner F., et al. Perturbation Of The Microvascular Glycocalyx And Perfusion In Infants After Cardiopulmonary Bypass. J. Thorac. Cardiovasc. Surg. 2015;150:1474–1481. doi: 10.1016/j.jtcvs.2015.08.050. [DOI] [PubMed] [Google Scholar]
- 212.Zhang Y., Zeng J., He X., Cao W., Peng X., Li G. Pulsatility Protects The Endothelial Glycocalyx During Extracorporeal Membrane Oxygenation. Microcirculation. 2021 doi: 10.1111/micc.12722. E12722. [DOI] [PubMed] [Google Scholar]
- 213.Wei T.J., Wang C.H., Chan W.S., Huang C.H., Lai C.H., Wang M.J., et al. Microcirculatory Response To Changes In Venoarterial Extracorporeal Membrane Oxygenation Pump Flow: A Prospective Observational Study. Front Med (Lausanne) 2021;8 doi: 10.3389/fmed.2021.649263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Kilger E., Weis F., Briegel J., Frey L., Goetz A.E., Reuter D., et al. Stress Doses Of Hydrocortisone Reduce Severe Systemic Inflammatory Response Syndrome And Improve Early Outcome In A Risk Group Of Patients After Cardiac Surgery. Crit. Care Med. 2003;31:1068–1074. doi: 10.1097/01.CCM.0000059646.89546.98. [DOI] [PubMed] [Google Scholar]
- 215.Menon K., Mcnally J.D. Corticosteroids In Pediatric Septic Shock Are Helpful. Crit. Care Med. 2018;46:635–636. doi: 10.1097/CCM.0000000000002979. [DOI] [PubMed] [Google Scholar]
- 216.Chappell D., Jacob M., Hofmann-Kiefer K., Bruegger D., Rehm M., Conzen P., et al. Hydrocortisone Preserves The Vascular Barrier By Protecting The Endothelial Glycocalyx. Anesthesiology. 2007;107:776–784. doi: 10.1097/01.anes.0000286984.39328.96. [DOI] [PubMed] [Google Scholar]
- 217.Chappell D., Hofmann-Kiefer K., Jacob M., Rehm M., Briegel J., Welsch U., et al. TNF-Alpha Induced Shedding Of The Endothelial Glycocalyx Is Prevented By Hydrocortisone And Antithrombin. Basic Res. Cardiol. 2009;104:78–89. doi: 10.1007/s00395-008-0749-5. [DOI] [PubMed] [Google Scholar]
- 218.Zimmerman J.J. Corticosteroids In Pediatric Septic Shock Are Not Helpful. Crit. Care Med. 2018;46:637–639. doi: 10.1097/CCM.0000000000002980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Haymet A.B., Bartnikowski N., Wood E.S., Vallely M.P., Mcbride A., Yacoub S., et al. Studying The Endothelial Glycocalyx In Vitro: What Is Missing? Front Cardiovasc. Med. 2021;8 doi: 10.3389/fcvm.2021.647086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Ebong E.E., Macaluso F.P., Spray D.C., Tarbell J.M. Imaging The Endothelial Glycocalyx In Vitro By Rapid Freezing/Freeze Substitution Transmission Electron Microscopy. Arterioscler. Thromb. Vasc. Biol. 2011;31:1908–1915. doi: 10.1161/ATVBAHA.111.225268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Hempel C., Kapishnikov S., Perez-Berna A.J., Werner S., Guttmann P., Pereiro E., et al. The Need To Freeze-Dehydration During Specimen Preparation For Electron Microscopy Collapses The Endothelial Glycocalyx Regardless Of Fixation Method. Microcirculation. 2020;27:E12643. doi: 10.1111/micc.12643. [DOI] [PubMed] [Google Scholar]
- 222.Sun X., Li L., Overdier K.H., Ammons L.A., Douglas I.S., Burlew C.C., et al. Analysis Of Total Human Urinary Glycosaminoglycan Disaccharides By Liquid Chromatography-Tandem Mass Spectrometry. Anal. Chem. 2015;87:6220–6227. doi: 10.1021/acs.analchem.5b00913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Haeren R.H., Van De Ven S.E., Van Zandvoort M.A., Vink H., Van Overbeeke J.J., Hoogland G., et al. Assessment And Imaging Of The Cerebrovascular Glycocalyx. Curr. Neurovasc. Res. 2016;13:249–260. doi: 10.2174/1567202613666160504104434. [DOI] [PubMed] [Google Scholar]
- 224.Donati A., Damiani E., Domizi R., Romano R., Adrario E., Pelaia P., et al. Alteration Of The Sublingual Microvascular Glycocalyx In Critically Ill Patients. Microvasc. Res. 2013;90:86–89. doi: 10.1016/j.mvr.2013.08.007. [DOI] [PubMed] [Google Scholar]
- 225.Hiedl S., Schwepcke A., Weber F., Genzel-Boroviczeny O. Microcirculation In Preterm Infants: Profound Effects Of Patent Ductus Arteriosus. J. Pediatr. 2010;156:191–196. doi: 10.1016/j.jpeds.2009.08.034. [DOI] [PubMed] [Google Scholar]
- 226.Howie S.R. Blood Sample Volumes In Child Health Research: Review Of Safe Limits. Bull. World Health Organ. 2011;89:46–53. doi: 10.2471/BLT.10.080010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Hahn R.G., Hasselgren E., Bjorne H., Zdolsek M., Zdolsek J. Biomarkers Of Endothelial Injury In Plasma Are Dependent On Kidney Function. Clin. Hemorheol. Microcirc. 2019;72:161–168. doi: 10.3233/CH-180444. [DOI] [PubMed] [Google Scholar]
- 228.Zhang Y., Lee T.S., Kolb E.M., Sun K., Lu X., Sladek F.M., et al. AMP-Activated Protein Kinase Is Involved In Endothelial NO Synthase Activation In Response To Shear Stress. Arterioscler. Thromb. Vasc. Biol. 2006;26:1281–1287. doi: 10.1161/01.ATV.0000221230.08596.98. [DOI] [PubMed] [Google Scholar]
- 229.Dixit M., Bess E., Fisslthaler B., Hartel F.V., Noll T., Busse R., et al. Shear Stress-Induced Activation Of The AMP-Activated Protein Kinase Regulates FoxO1a And Angiopoietin-2 In Endothelial Cells. Cardiovasc. Res. 2008;77:160–168. doi: 10.1093/cvr/cvm017. [DOI] [PubMed] [Google Scholar]
- 230.Zeng Y., Liu J. Role Of Glypican-1 In Endothelial NOS Activation Under Various Steady Shear Stress Magnitudes. Exp. Cell Res. 2016;348:184–189. doi: 10.1016/j.yexcr.2016.09.017. [DOI] [PubMed] [Google Scholar]
- 231.Fux L., Ilan N., Sanderson R.D., Vlodavsky I. Heparanase: Busy At The Cell Surface. Trends Biochem. Sci. 2009;34:511–519. doi: 10.1016/j.tibs.2009.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Becker B.F., Jacob M., Leipert S., Salmon A.H., Chappell D. Degradation Of The Endothelial Glycocalyx In Clinical Settings: Searching For The Sheddases. Br. J. Clin. Pharmacol. 2015;80:389–402. doi: 10.1111/bcp.12629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Ramnath R.D., Butler M.J., Newman G., Desideri S., Russell A., Lay A.C., et al. Blocking Matrix Metalloproteinase-Mediated Syndecan-4 Shedding Restores The Endothelial Glycocalyx And Glomerular Filtration Barrier Function In Early Diabetic Kidney Disease. Kidney Int. 2020;97:951–965. doi: 10.1016/j.kint.2019.09.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Li T., Liu X., Zhao Z., Ni L., Liu C. Sulodexide Recovers Endothelial Function Through Reconstructing Glycocalyx In The Balloon-Injury Rat Carotid Artery Model. Oncotarget. 2017;8:91350–91361. doi: 10.18632/oncotarget.20518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Song J.W., Zullo J.A., Liveris D., Dragovich M., Zhang X.F., Goligorsky M.S. Therapeutic Restoration Of Endothelial Glycocalyx In Sepsis. J. Pharmacol. Exp. Ther. 2017;361:115–121. doi: 10.1124/jpet.116.239509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Yang Y., Haeger S.M., Suflita M.A., Zhang F., Dailey K.L., Colbert J.F., et al. Fibroblast Growth Factor Signaling Mediates Pulmonary Endothelial Glycocalyx Reconstitution. Am. J. Respir. Cell Mol. Biol. 2017;56:727–737. doi: 10.1165/rcmb.2016-0338OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Desideri S., Onions K.L., Baker S.L., Gamez M., El Hegni E.H.H., Russell A., et al. Endothelial Glycocalyx Restoration By Growth Factors In Diabetic Nephropathy. Biorheology. 2019;56:163–179. doi: 10.3233/BIR-180199. [DOI] [PubMed] [Google Scholar]
- 238.Drost C.C., Rovas A., Kusche-Vihrog K., Van Slyke P., Kim H., Hoang V.C., et al. Tie2 Activation Promotes Protection And Reconstitution Of The Endothelial Glycocalyx In Human Sepsis. Thromb. Haemost. 2019;119:1827–1838. doi: 10.1055/s-0039-1695768. [DOI] [PubMed] [Google Scholar]