

Abstract
Intracellular compartments make up roughly two thirds of the body, but delivery of molecular imaging probes to these spaces can be challenging. This situation is particularly true for probes designed for detection by magnetic resonance imaging (MRI), a high resolution but relatively insensitive modality. Most MRI contrast agents are polar and membrane impermeant, making it difficult to deliver them in sufficient quantities for measurement of intracellular analytes. Here we address this problem by introducing a new class of planar tetradentate Mn(III) chelates assembled from a 1,2-phenylenediamido (PDA) backbone. Mn(III)-PDA complexes display T1 relaxivity comparable to that of Gd(III)-based contrast agents and undergo spontaneous cytosolic localization via defined mechanisms. Probe variants incorporating enzyme-cleavable acetomethoxy ester groups are processed by intracellular esterases and accumulate in cells. Probes modified with ethyl esters preferentially label genetically modified cells that express a substrate-selective esterase. In each case, the contrast agents gives rise to robust T1-weighted MRI enhancements, providing precedents for the detection of intracellular targets by Mn(III)-PDA complexes. These compounds therefore constitute a platform from which to develop reagents for molecular MRI of diverse processes inside cells.
The ability to map intracellular processes in space and time by noninvasive molecular imaging is highly desirable for studying organismic-level phenomena in the lab and clinic. MRI provides a combination of high spatiotemporal resolution and unlimited tissue penetration that makes it an ideal platform for application of cytosolic imaging probes, but few such probes currently exist. Most MRI contrast agents are complexes of Mn(II) or Gd(III) with aminopolycarboxylic acid chelating ligands that stabilize bound ions and reduce toxicity, but inherently limit membrane permeability. Nanoparticle-based MRI probes are more potent than paramagnetic complexes, but because of their size are even less likely to permeate membranes.
Strategies to deliver MRI agents to cells have therefore made use of cell permeable carrier vehicles,1,2 but contrast agents delivered using such methods do not necessarily localize to the cytosol, limiting applicability for molecular imaging of many intracellular targets. A further strategy has been to shuttle agents into cells via active transport mechanisms under genetic control.3–5 This task requires genetic manipulation of the recipient cell and so far has proved effective for delivery of relatively few probes. Additional receptor targeting approaches are suitable for promoting cell uptake of MRI contrast agents, but not cytosolic probe localization.6,7
A mechanistically simpler strategy that could enable efficient delivery of intracellular molecular imaging agents is to design probes around paramagnetic platforms that are intrinsically membrane permeable.8,9 To implement such a strategy, we prepared complexes of Mn(III) with tetradentate N2O2 and N3O ligand systems derived from 1,2-phenylenediamine (PDA).10,11 These ligands convey significant stability and magnetic properties of Mn(III) complexes arising from their high-spin d4 electronic configurations, but have not previously been explored as MRI contrast agents. Based on the structures of homologous compounds, Mn(III)-PDA complexes are likely to accommodate one or more exchangeable inner-sphere solvent or co-ligand molecules,11,12 an ideal characteristic for T1-weighted MRI contrast agents. These complexes differ from open-chain Mn-based MRI agents, which incorporate Mn(II) in d5 configuration, which requires ligands with greater denticity to achieve thermodynamic and kinetic stabilization.13 In Mn(III)-PDA complexes, we expect the combination of a lipophilic aryl backbone with a hydrophilic metal binding site to afford a balance that facilitates crossing of cell membranes. PDA ligands should also provide greater rigidity than ethylenediamino ligands, and they are more stable to hydrolysis than o-phenylenediimine derivatives.14
Candidate PDA ligands L1-L3 and their corresponding Mn complexes (Figure 1a) were synthesized as described in Supplemental Information. Spectra of the ligands and their complexes are presented in Figure S1. The absorbance signatures were used to measure Mn3+ dissociation constants of 1.2 ± 0.1 μM, 610 ± 110 nM for L2, and 500 ± 80 nM for L3 by titration with Mn(OAc)3 (Figure S2). These values are inferior to the stabilities of commercial Gd-based MRI agents, but comparable to the clinically approved Mn(II) agent Mn-N,N′-dipyridoxylethylenediamine-N,N′-diacetate 5,5′-bis(phosphate).15 Octanol-water partition coefficients (Poct/wat)16 and solubilities of the three Mn-PDA complexes are presented in Table S1. The neutral Mn(III) complex MnL3 displays highest lipophilicity, but all complexes have higher log Poct/wat values than the clinical contrast agent Gd-diethylaminetriamine pentaacetic acid (DTPA). The T1 relaxivity (r1) of each complex, a measure of its strength as an MRI contrast agent, was determined at 7 T and 22 °C. Values of r1 for MnL1, MnL2, and MnL3 are 4.1 ± 0.1, 5.4 ± 0.2, and 5.1 ± 0.1 mM−1s−1, respectively, comparable to Gd-DTPA17. Relaxivity values were also determined at 3 T (Table S1) and for MnL3 at 7 T in the presence of serum albumin, bicarbonate, and phosphate (Table S2). These measurements indicate that Mn-PDA agents combine robust performance as contrast agents with physicochemical properties conducive to membrane permeability.
Figure 1.
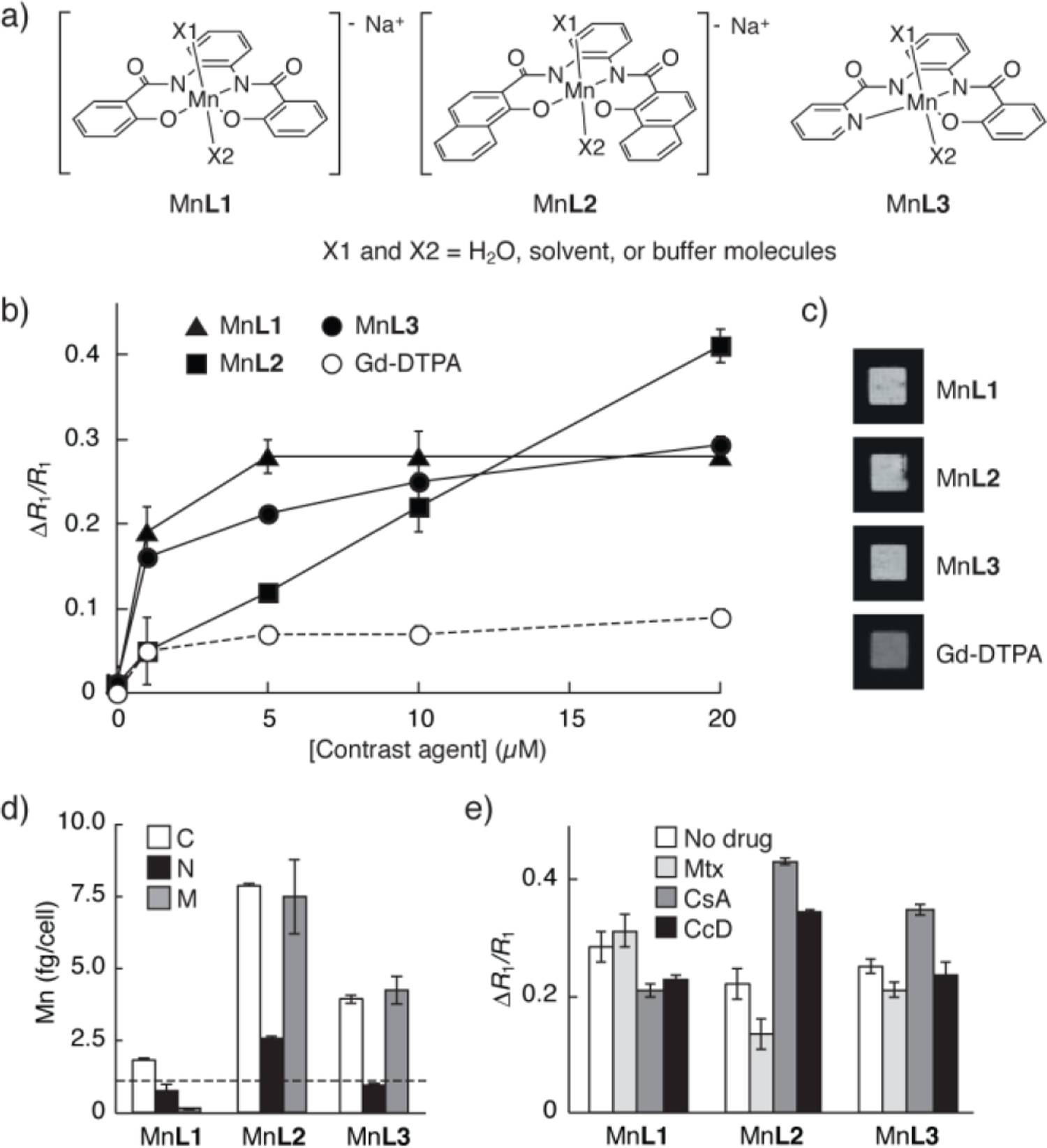
Cell-permeable Mn-PDA contrast agents. (a) Structures of three Mn-PDA complexes. (b) Relative change in T1 relaxation rates (ΔR1/R1) measured from HEK293 cells incubated with MnL1-L3 or with Gd-DTPA. (c) Images of cell pellets corresponding to the 10 μM incubation condition in (b). (d) Mn quantification by ICP-MS in cytosolic (C), nuclear (N), and membranous (M) fractions obtained from cells incubated with 10 μM MnL1-L3. Horizontal line indicates cytosolic Mn quantified from unlabeled cells. (e) Effects of treatment with 50 μM Mtx, 20 μM CsA, or 5 μM CcD on cell labeling by 10 μM MnL1-L3. Error bars denote s.d., n ≥ 3.
To assess the cell labeling characteristics of the new contrast agents, HEK293 cells were incubated with up to 20 μM of the Mn-PDA compounds for 30 min., followed by washing and then MRI analysis. Negligible toxicity was observed under these conditions (Figure S3). All three Mn-PDA agents appeared to label cells readily, producing enhancements of 28% ± 3%, 41% ± 4% and 29% ± 1% in the T1 relaxation rates (R1) measured from cells incubated with 20 μM MnL1, L2, and L3, respectively (Figure 1b). These changes reflect substantial T1-weighted contrast enhancements observed in cell pellet images (Figure 1c). The more hydrophilic Gd-DTPA produced smaller R1 changes with minimal visible contrast, and the cell-permeable ion Mn2+ produced a concentration-dependent labeling profile similar to MnL3 (Figure S4).
To explain the enhancements in cell pellet R1 in terms of subcellular localization patterns, cells were fractionated after 30 min labeling with 10 μM of each the contrast agents. These extracts were analyzed by inductively coupled plasma mass spectrometry (ICP-MS). Total manganese levels of 2.62 ± 0.05, 18.3 ± 1.2, and 9.3 ± 0.8 fg/cells were observed among fractions labeled with MnL1, MnL2, and MnL3, respectively. Fractionation results (Figure 1d) indicated that both MnL2 and MnL3 display strong cytosolic localization combined with roughly equal membranous concentrations, consistent with their hydrophobicity. In contrast, MnL1 shows considerably lower cytosolic localization, and negligible levels in membranous fractions. The findings thus indicate that MnL2 and MnL3 penetrate cells more effectively than MnL1. The fact that MnL1 gives rise to R1 enhancements comparable to MnL2 and MnL3 argues that MnL1 may have higher cell-associated r1, or that it could be bound to the cell exterior and partially lost during fractionation. The second explanation is supported by ICP-MS analysis of minimally treated cell pellets, which shows ~30% more total Mn in MnL1 treated cells than those prepared with MnL3.
To probe the contributions of selected active import or export mechanisms to cell labeling by the Mn-PDA complexes, we repeated the cell labeling studies of Figure 1b in the presence of methotrexate (Mtx), which inhibits drug import by folate transporters RFC1 and PCFT, and cyclosporin A (CsA), which blocks drug export via the multidrug resistance protein MDR1; both the folate and multidrug transport pathways have been specifically shown to process planar aromatic compounds similar to our Mn-PDA complexes.18,19 We also tested the affects of cytochalasin D (CcD), which blocks actin polymerization required for endocytosis. Results summarized in Figures 1e and S5 indicate that MnL2 cell labeling is significantly affected by all three drugs (t-test p < 0.006, n = 3), indicating susceptibility of MnL2 to active transport mechanisms, despite its apparently non-saturating labeling profile in Figure 1b. In contrast, MnL3 cell labeling is enhanced by CsA (p = 0.008), but unaffected by Mtx or CcD (p > 0.1). MnL1 labels cells somewhat more poorly in the presence of CsA and CcD. The fact that cell labeling by MnL2 and MnL3, but not MnL1, is promoted by CsA supports evidence that these two compounds localize more effectively to the cytosol, where MDR1 is active.
The cell labeling and fractionation studies show that MnL3 displays the clearest evidence of innate membrane permeability of the three Mn-PDA agents, consistent with its lipophilicity. Tests of resistance to dissociation (Figure S6) and transmetallation (Figure S7) revealed the stability of this compound in the presence of excess ethylenediamine tetraacetic acid (EDTA) and biologically relevant transition metals for up to 48 hr. Based on these results and its other properties, MnL3 was deemed most suitable as a platform for development of further membrane-permeant agents.
We next sought to determine whether MnL3 could be incorporated into probes for MRI-based detection of intracellular species. Intracellular esterases represent an important class of targets for imaging agents that can reach the cell interior. Endogenous or ectopically expressed esterases can trap appropriately esterified probes.20–22 Previous studies have introduced esterase-sensitive MRI contrast agents, but none has demonstrated intracellular cleavage or cytosolic trapping.23–26 To test the ability of MnL3-based MRI contrast agents to undergo esterase-dependent capture and accumulation in cells, we synthesized an acetoxymethyl (AM) modified MnL3 analog, MnL3-OAc-AM. AM esters are well known substrates for intracellular esterases,20 so we anticipated that MnL3-OAc-AM would enter cells and be processed into a charged and membrane-impermeant product, MnL3-OAc-OH (Figure 2a).
Figure 2.
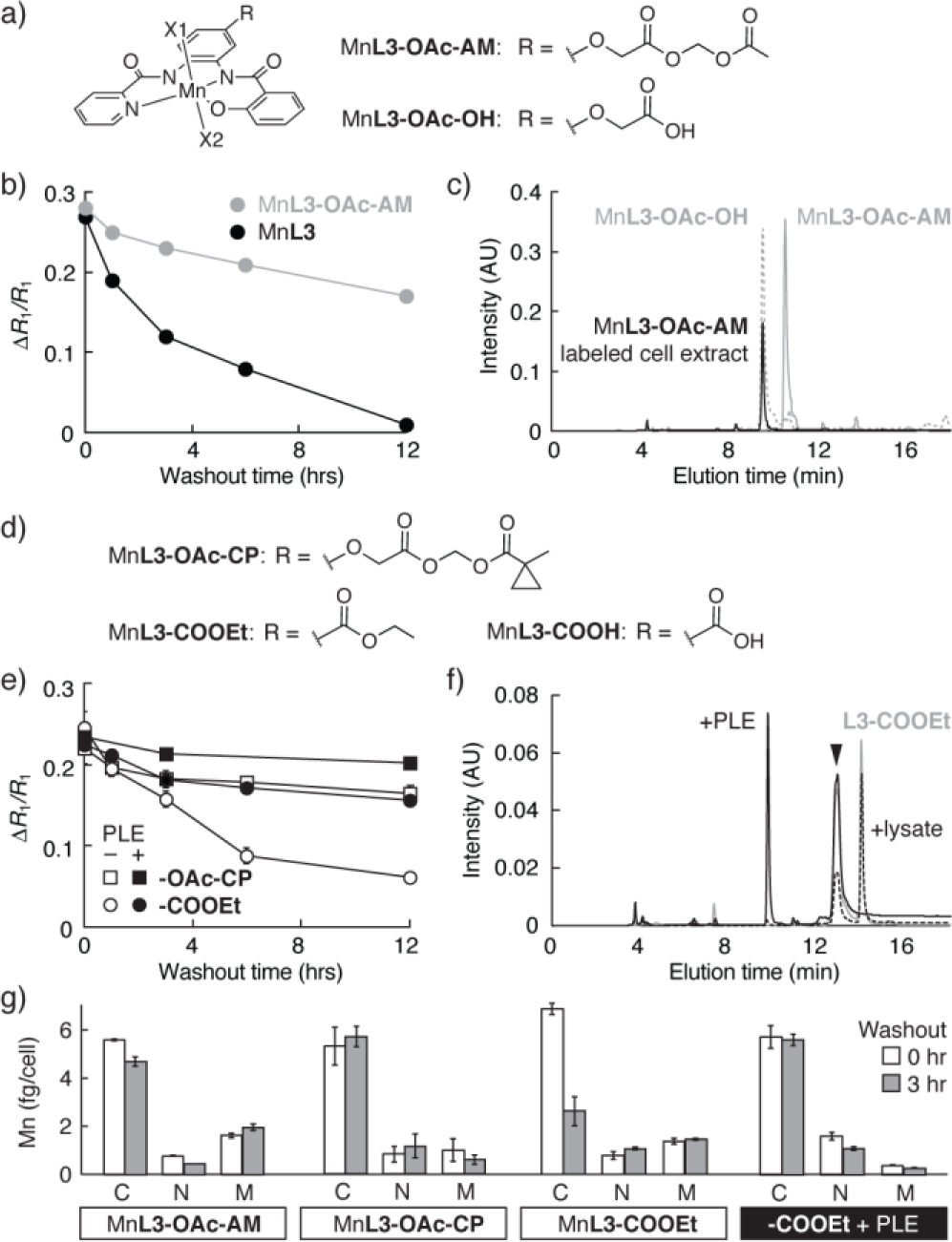
Targeting intracellular esterases with Mn-PDA contrast agents. (a) Esterase substrate MnL3-OAc-AM and its hydrolysis product. (b) Washout time courses from cells preincubated with 10 μM MnL3-OAc-AM or MnL3. (c) HPLC analysis of extracts from cells labeled with MnL3-OAc-AM (black). The elution peak corresponds to MnL3-OAc-OH (dashed), rather than MnL3-OAc-AM (solid gray), indicating complete hydrolysis in cells. (d) Sidechain structures of MnL3-OAc-CP MnL3-COOEt, and MnL3-COOH. (e) Washout time courses from cells incubated with 10 μM MnL3-OAc-CP or MnL3-COOEt, following transfection with genes encoding either PLE or green fluorescent protein. (f) HPLC traces obtained from 5 μM L3-COOEt either alone (gray), and following treatment with cell lysate (black dotted) or 15 units PLE (solid line), showing selective cleavage. (g) Quantification of Mn in fractions from cells incubated with the agents, at washout times t = 0 and 3 hr. All measurements n ≥ 3, error bars s.d.
Cells were incubated for 30 min with 10 μM MnL3-OAc-AM or with MnL3 control, rinsed, incubated for a variable wash-out delay period, rinsed again to remove extracellular contrast agent, and then imaged by MRI to measure contrast arising from remaining intracellular probe. Consistent with an esterase-mediated trapping mechanism, MnL3 and MnL3-OAc-AM displayed striking differences in wash-out kinetics (Figure 2b). Cells incubated with each compound showed similar initial R1 values, indicating equivalent labeling efficiency. Cells labeled with MnL3 displayed a complete return to baseline R1 over 12 hr, with a half-life of ~3 hr. In contrast, cells preloaded with MnL3-OAc-AM displayed only a 37% reduction in R1 over 12 hr, with a half-life longer than the experimental duration. Fractionation and analytical HPLC analysis of cells incubated with MnL3-OAc-AM revealed complete conversion of this compound into the expected product MnL3-OAc-OH (Figure 2c). These results confirm that complete hydrolysis of the esterase-targeted contrast agent MnL3-OAc-AM takes place in labeled cells.
Given the evidence for endogenous esterase-mediated cell trapping of MnL3-OAc-AM, we wondered whether selective esterase-mediated cell trapping22 could be implemented as part of a strategy for genetically targeting MnL3 derivatives. Two candidates for selective esterase processing were therefore synthesized, one bearing a cyclopropyl ester moiety (MnL3-OAc-CP) and the other bearing a simple ethyl ester (MnL3-COOEt) (Figure 2d).
These further MnL3-based esters were coincubated with HEK293 cells transfected with constructs encoding either green fluorescent protein (GFP, control) or a porcine liver esterase (PLE) known to exhibit distinct specificity from the repertoire of esterases normally present in these cells.22 As expected based on their similarity to MnL3-OAc-AM, both MnL3-OAc-CP and MnL3-COOEt were effectively taken up by PLE-expressing cells as well as control cells, and neither showed evidence of acute cell toxicity (Figure S8). Retention of the contrast agents differed significantly however. MRI results suggested that, although both MnL3-OAc-CP and MnL3-COOEt were retained in PLE-expressing cells, the ethyl ester MnL3-COOEt showed substantially greater selectivity for PLE cells compared with control cells not transfected with the PLE gene (Figure 2e). After 12 hr, the R1 enhancement of cells incubated with MnL3-COOEt was reduced by 75%, whereas the R1 increase of cells loaded with MnL3-OAc-CP was reduced by 32%, only 7% more than the reduction observed for MnL3-OAc-AM.
The cell retention results suggest that MnL3-COOEt is cleaved more effectively in the presence of expressed PLE than by endogenous intracellular esterases alone.27 To evaluate this possibility, we monitored L3-COOEt in buffer following addition of 20 mg HEK293 cell extract or 15 units of purified PLE. As predicted from the cell labeling MRI result, the contrast agent was hydrolyzed much more effectively in the presence of PLE (Figure 2f ). In contrast, the L3-OAc-CP ligand was found to be hydrolyzed completely by addition of cell extract alone, with no requirement for PLE, explaining the nonspecific retention of MnL3-OAc-CP in untransfected cells (Figure S9). To obtain further evidence for the esterase specificity of these selective retention effects, we performed a fractionation and ICP-MS analysis of HEK293 cells incubated with 10 μM MnL3-OAc-AM, MnL3-OAc-CP, or MnL3-COOEt, followed by either 0 or 3 hr washout periods. Only MnL3-COOEt showed substantial reduction in cytosolic content during this period, and this reduction was absent in cells expressing PLE.
Our data thus indicate that Mn-PDA agents have the ability to enter cells and interact with distinct cytosolic enzymes, demonstrating their suitability for molecular imaging of intracellular targets. The basic architecture of the Mn-PDA agents allows for facile synthesis, confers stability, and provides MRI T1 relaxivity comparable to commercial contrast agents. Of the compounds surveyed, the neutral complex MnL3 seemed to be the best candidate for further modification. Functionalization of MnL3 with different ester groups did not diminish cell uptake, showing that MnL3 can indeed act as a building block for molecular imaging agents.
Although we present Mn-PDA-based esterase substrates as a proof of concept for MRI detection of intracellular analytes, these contrast agents could also be used for molecular imaging themselves. In particular, ester derivatives of MnL3 are likely to be effective reagents for cell labeling and in vivo cell tracking. Specific labeling of cells expressing PLE or related enzymes could also provide a pathway to reporter gene mapping by MRI. Although extracellular esterases might interfere under some circumstances,28 extracellularly hydrolyzed agents would not tend to be trapped, and might therefore not contribute heavily to background signal. Additional targeted or responsive agents could be constructed by modifying the Mn-PDA platform. This new family of contrast agents would be most applicable for molecular imaging applications in animals, but the stability and relaxivity of Mn-PDA derivatives might eventually enable applications in human subjects.
Supplementary Material
ACKNOWLEDGMENT
The authors thank Atushi Takahashi, Koli Taghizadeh, Emily Young, and Emily Loucks. Funding came from the MIT Simons Center and NIH grants R21-MH102470 and U01-NS090451 to AJ, as well as grant P30-ES002109 for the CEHS Bioanalytical Core.
Footnotes
ASSOCIATED CONTENT
Supporting Information. Detailed methods and further characterization of synthetic compounds. This material is available free of charge at http://pubs.acs.org.
REFERENCES
- (1).Bhorade R; Weissleder R; Nakakoshi T; Moore A; Tung C-H Bioconjug. Chem. 2000, 11, 301. [DOI] [PubMed] [Google Scholar]
- (2).Hung AH; Holbrook RJ; Rotz MW; Glasscock CJ; Mansukhani ND; MacRenaris KW; Manus LM; Duch MC; Dam KT; Hersam MC; Meade TJ ACS Nano 2014, 8, 10168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (3).Weissleder R; Moore A; Mahmood U; Bhorade R; Benveniste H; Chiocca EA; Basilion JP Nat. Med. 2000, 6, 351. [DOI] [PubMed] [Google Scholar]
- (4).Bartelle BB; Szulc KU; Suero-Abreu GA; Rodriguez JJ; Turnbull DH Magn. Reson. Med. 2013, 70, 842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (5).Patrick PS; Hammersley J; Loizou L; Kettunen MI; Rodrigues TB; Hu D-E; Tee S-S; Hesketh R; Lyons SK; Soloviev D; Lewis DY; Aime S; Fulton SM; Brindle KM Proc. Natl. Acad. Sci. 2014, 111, 415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (6).Aime S; Barge A; Cabella C; Gianolio SGC and Targeting E Cells with MR Imaging Probes Based on Paramagnetic Gd(III) Chelates. Curr. Pharm. Biotechnol. 2004, 5, 509–518. [DOI] [PubMed] [Google Scholar]
- (7).McCarthy JR; Kelly KA; Sun EY; Weissleder R Nanomedicine 2007, 2, 153. [DOI] [PubMed] [Google Scholar]
- (8).Lee T; Zhang X; Dhar S; Faas H; Lippard SJ; Jasanoff A Chem. Biol. 2010, 17, 665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (9).Zhang X; Lovejoy KS; Jasanoff A; Lippard SJ Proc. Natl. Acad. Sci. 2007, 104, 10780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (10).Bertoncello K; Fallon GD; Murray KS Inorganica Chim. Acta 1990, 174, 57. [Google Scholar]
- (11).Sunatsuki Y; Shimada H; Matsuo T; Nakamura M; Kai F; Matsumoto N; Re N Inorg. Chem. 1998, 37, 5566. [DOI] [PubMed] [Google Scholar]
- (12).Yang C; Wang Q-L; Ma Y; Tang G-T; Liao D-Z; Yan S-P; Yang G-M; Cheng P Inorg. Chem. 2010, 49, 2047. [DOI] [PubMed] [Google Scholar]
- (13).Gale EM; Atanasova IP; Blasi F; Ay I; Caravan PJ Am. Chem. Soc. 2015, 137, 15548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (14).Venkataramanan NS; Kuppuraj G; Rajagopal S Coord. Chem. Rev. 2005, 249, 1249. [Google Scholar]
- (15).Rocklage SM; Cacheris WP; Quay SC; Hahn FE; Raymond KN Inorg. Chem. 1989, 28, 477. [Google Scholar]
- (16).Leo A; Hansch C; Elkins D Chem. Rev. 1971, 71, 525. [Google Scholar]
- (17).Shen Y; Goerner FL; Snyder C; Morelli JN; Hao D; Hu D; Li X; Runge VM Invest. Radiol. 2015, 50, 330. [DOI] [PubMed] [Google Scholar]
- (18).Hou Z; Matherly LH In Exchangers; Membranes, T. MOBBT-C in, Ed.; Academic Press, 2014; Vol. Volume 73, pp. 175–204. [Google Scholar]
- (19).Leslie EM; Deeley RG; Cole SPC Toxicol. Appl. Pharmacol. 2005, 204, 216. [DOI] [PubMed] [Google Scholar]
- (20).Tsien RY Nature 1981, 290, 527. [DOI] [PubMed] [Google Scholar]
- (21).Rehberg M; Lepier, Solchenberger Alexandra, Barbara Osten P; Blum R Cell Calcium 2008, 44, 386. [DOI] [PubMed] [Google Scholar]
- (22).Tian L; Yang Y; Wysocki LM; Arnold AC; Hu A; Ravichandran B; Sternson SM; Looger LL; Lavis LD Proc. Natl. Acad. Sci. U. S. A. 2012, 109, 4756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (23).Himmelreich U; Aime S; Hieronymus T; Justicia C; Uggeri F; Zenke M; Hoehn M Neuroimage 2006, 32, 1142. [DOI] [PubMed] [Google Scholar]
- (24).Giardiello M; Lowe MP; Botta M Chem. Commun. 2007, 4044. [DOI] [PubMed] [Google Scholar]
- (25).Li Y; Sheth VR; Liu G; Pagel MD Contrast Media Mol. Imaging 2011, 6, 219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (26).MacRenaris KW; Ma Z; Krueger RL; Carney CE; Meade TJ Bioconjug. Chem. 2016, 27, 465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (27).Lind PA; Daniel RM; Monk C; Dunn RV Biochim. Biophys. Acta - Proteins Proteomics 2004, 1702, 103. [DOI] [PubMed] [Google Scholar]
- (28).Jobsis PD; Rothstein EC; Balaban RS J. Microsc. 2007, 226, 74. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.