

Abstract
K-RAS is the most frequently mutated oncogene in solid tumors, such as pancreatic, colon or lung cancer. The GTPase K-RAS can either be in an active (GTP-loaded) or inactive (GDP-loaded) form. In its active form K-RAS forwards signals from growth factors, cytokines or hormones to the nucleus, regulating essential pathways, such as cell proliferation and differentiation. In turn, activating somatic mutations of this proto-oncogene deregulate the complex interplay between GAP (GTPase-activating) - and GEF (Guanine nucleotide exchange factor) - proteins, driving neoplastic transformation. Due to a rather shallow surface, K-RAS lacks proper binding pockets for small molecules, hindering drug development over the past thirty years. This review summarizes recent progress in the development of low molecular antagonists and further shows insights of a newly described interaction between mutant K-RAS signaling and PD-L1 induced immunosuppression, giving new hope for future treatments of K-RAS mutated cancer.
Keywords: K-RAS, small molecules, immune checkpoints, PD-1, PD-L1
Introduction
During evolution, cells have acquired the ability to respond to external stimuli through fine-tuned signaling cascades. To this extent, rat sarcoma (RAS)-proteins are small monomeric G-proteins that utilize various signal transduction pathways to regulate cell growth, differentiation, and apoptosis[1]. Due to their proto-oncogenic character, RAS proteins are a popular target in the field of cancer research[2]. The most prominent members of the RAS family are the isoforms N-RAS, H-RAS, K-RAS 4A and K-RAS 4B. These proteins emerge from proto-oncogenes, as their point-mutations are frequently present in different types of cancer[3]. The amino acid sequence of all four isoforms is shown in Figure 1. The first 166 amino acids constitute the catalytic G-domain and share a sequence identity of 89%. The subsequent region spanning residues 167 to 188 and 189, respectively, differ among the isoforms. Hence, this C-terminal domain of RAS proteins is denoted as the hypervariable region.
Figure 1.
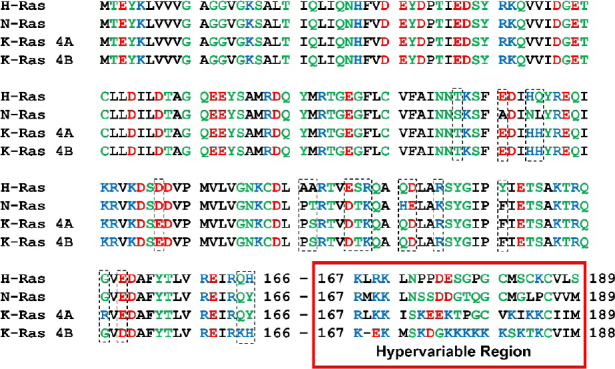
Amino acid sequences of RAS isoforms. The sequences of the RAS isoforms are compared and non-identical areas are marked by dashed rectangles. Amino acids 1-166 constitute the catalytic domain and amino acids 167-188/189 represent the hypervariable region. The amino acids are coloured based on their physico-chemical properties. Hydrophobic (black), polar/neutral (green), acidic (red) und basic (blue)
K-RAS 4A and 4B represent two splicing variants of the exon 4 of the K-RAS pre-mRNA and all four isoforms possess the C-terminal CaaX-Box. The cysteine at position 185 or 186 respectively is followed by two aliphatic amino acids and it is this cysteine that plays a crucial role in the post-translational processing of RAS. All RAS isoforms are farnesylated to enable their anchoring to the membrane. Additionally, K-RAS 4A can be palmitoylated at cysteine 180, N-RAS at cysteine 181, and H-RAS at cysteine 181 and 184. This additional palmitoylation increases the hydrophobicity of the protein. In contrast to this, K-RAS 4B is not palmitoylated but contains a poly lysine stretch (K175-K180) in the hypervariable region instead. This positively charged sequence facilitates a more transient anchorage to the negatively charged plasma membrane.
RAS - function and structure
RAS proteins function as binary molecular switches in signal transduction. They exist in an active GTP-bound “ON”-state and an inactive GDP-bound “OFF”-state[4]. In the active state, the G-protein is able to bind to different effector proteins and transfer the signal. The intrinsic GTPase property enables the hydrolysis of GTP to GDP and hence the transition from the active to the inactive state. For the reactivation of RAS, the GDP nucleotide has to dissociate and the GTP nucleotide, which is present in a 10-fold excess, has to be incorporated. The intrinsic GDP-dissociation rate is 10-5 s-1[5] and thus too low to be biologically significant. This rate is enhanced 104-fold by guanine nucleotide exchange factors (GEFs)[6]. These GEFs are positive regulators of the RAS activity. Hitherto, the protein encoded by the cell division cycle gene 25 (Cdc25) and the son of sevenless (SOS) are the best understood GEFs for RAS. GTPase activating proteins (GAPs) function as counterparts to GEFs. They are essential for the hydrolysis of GTP as the inherent GTPase activity of RAS is only 10-4 s-1, which is amplified by GAPs by 103-times.
The three-dimensional structure of RAS proteins explains the molecular mechanism of their biological function as cellular protein switches [Figure 2]. The guanosine nucleotide binds to the G-protein conserved G-domain. The G-domain constitutes of six β-sheets and five surrounding α-helices which are connected with ten loops[6].
Figure 2.
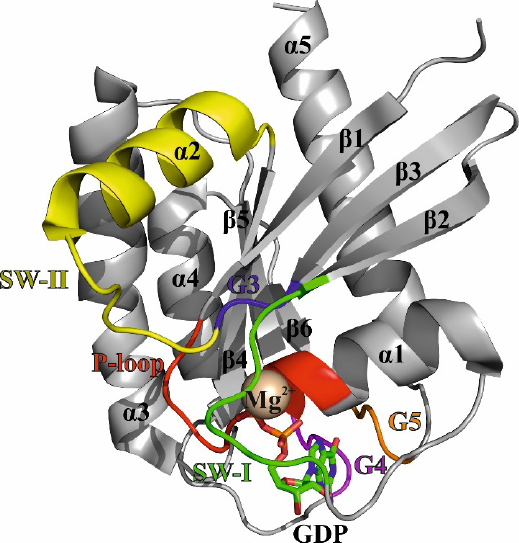
Crystal structure of K-RAS 4B GDP in ribbon representation (4EPY). The α-helices (α1-α5), β-sheets (β1-β6), the magnesium-ion (Mg2+), and the GDP nucleotide are labeled in black. The loops and switch regions are color-coded as follows: P-loop (red); switch I - SW-I (green); G3-loop (blue); switch II - SW-II (yellow); G4-loop (magenta); G5-loop (orange)
Five of the ten loops that can be found between the secondary structural motifs are denoted as G1 to G5 motifs and participate in nucleotide binding. The G1-loop is glycine-rich region and utilizes the positive charge of lysine 16 to bind the β-phosphate of the nucleotide. The β-phosphate is additionally coordinated by serine 17 and a magnesium ion. The positive charge of magnesium is essential to stabilize the three negative phosphate groups of the nucleotide and, therefore, the G1-loop has also been described as the P-loop (GxxxGKS/T). The amino acids 32 to 38 form the G2-loop, also known as switch I. The threonine at position 35 is crucial in the binding of the γ-phosphate and the magnesium ion. In the presence of the γ-phosphate, the switch I region adopts the active conformation and participates in the effector binding[4]. The G3-loop includes the motif DxxG and shares glycine 60 with the neighboring switch II region, which comprises amino acids 60 to 76. The glycine 60 is the second amino acid to bind the γ-phosphate through a hydrogen bond. Like switch I, switch II changes its conformation upon binding of GTP and does not only take part in effector binding but also acts as an interaction site for GEFs and GAPs[7]. The binding of the guanosine nucleotide is established by the G4-loop (N/TKxD aa 116 to 119) as well as the G5-loop (SAK aa 145 to 147)[8].
The conformational change of the protein from the inactive, GDP-bound state to the active, GTP-bound state has been coined as the loaded-spring mechanism[8]. In the active state, amino groups of threonine 35 (located in switch I) and glycine 60 (located in switch II) coordinate one oxygen atom of the γ-phosphate via hydrogen bonds. Upon hydrolysis of the γ-phosphate, the hydrogen bonds are broken and the flexible switch-loops spring back into their initial positions. Noteworthy, the loaded spring of the active state is not simply a rigid construct. The active state contains further two (sub-) states[9]. The difference between these two active states is the flexibility of the switches regions and the results in different affinities towards the effector proteins. In state 1, the switch regions exhibit a higher conformational fluctuation compared to state 2, which results in impaired binding to the effector RAF[10]. These conformational changes between the active and inactive conformation as well as the subdivision in active state 1 and 2 further emphasizes the dynamic nature of these switch regions of the RAS protein, an attribute referred to as polysterism[11].
K-RAS in cancer
K-RAS signaling
K-RAS acts as a cellular switch to regulate essential intrinsic pathways[12-14]. Extracellular stimuli, e.g., binding of growth factors, hormones or cytokines, are leading to a dimerization of receptor tyrosine kinases, which in turn activate GEFs, such as SOS, which leads to a nucleotide exchange and an activation of K-RAS. In its wild-type (wt) state K-RAS forwards signals from growth factors, cytokines or hormones to the nucleus, thereby regulating crucial pathways such as proliferation, differentiation and cell growth[15,16]. GAPs are responsible to enhance the catalytic rate of K-RAS GTPase function, leading to the hydrolyzation of GTP, the replacement with GDP, resulting in the inactivation of K-RAS[6,17]. Activating somatic mutations of this proto-oncogene disrupt this complex interplay between GEFs and GAPs[18,19]. GAPs can no longer increase the GTP hydrolyzation of oncogenic K-RAS, leading to a constitutive active mutant (mut) form driving neoplastic transformations in many cancer entities[20-22]. With a frequency of 21.6% somatic point mutations in K-RAS are the most common mutations in the RAS gene of all human cancers, followed by an 8.0% incidence of N-RAS mutations and a 3.3% rate of H-RAS mutations[23].
The mitogen-activated protein kinase (MAPK) signaling cascade is crucial for the regulation of cellular functions like differentiation and cell growth of normal cells. Constitutive activation of this pathway leads to uncontrolled cell proliferation, transformation, dissemination of cancer cells and is one of the major effector pathways deregulated in K-RAS mutant cancer[24,25]. The pathway consists of the kinases RAF, mitogen-activated protein kinase/extracellular receptor-stimulated kinase 1/2 (MEK), and extracellular receptor-stimulated kinase 1/2 (ERK), that are part of a phosphorylation cascade, downstream of activated GTP-bound RAS [Figure 3]. Once activated in the cytoplasm, ERK can activate proteins of the dynamic cytoskeletal complex that affect cell adhesion, trafficking, and movement[26,27]. In addition, it can also enter the nucleus and regulate various mitogenic transcription factors involved in stimulating cell proliferation[28,29]. The phosphoinositide 3-kinase (PI3K)/AKT pathway is the second major effector pathway downstream of mut K-RAS signaling. GTP-bound K-RAS can recruit and phosphorylate PI3K. Activated PI3K can phosphorylate the downstream serine/threonine kinase AKT, which in turn translocate from the plasma membrane to other cellular compartments, phosphorylating target proteins responsible for regulating cell growth, cell survival and entry of the cell cycle[28,30]. An important downstream target of AKT is the mammalian target of rapamycin (mTOR) complex 1, which results in lipid or nucleotide synthesis, as well as in protein translation[31].
Figure 3.
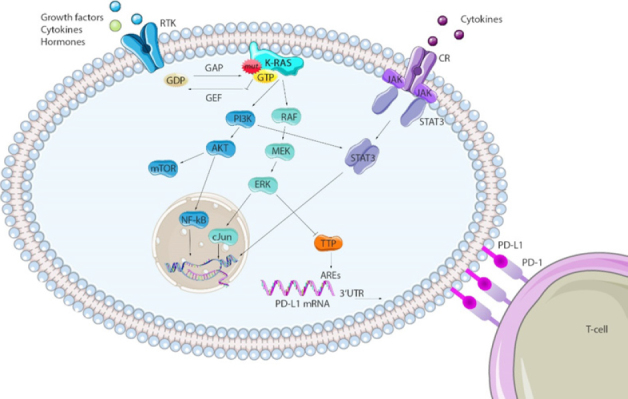
Major effector pathways of K-RAS and potential interactors. Mutations in K-RAS genes lead to constitutive GTP bound K-RAS proteins that activate downstream phosphorylation cascades. Major effector pathways dysregulated in K-RAS mut cancers are the MAPK (RAF/MEK/ERK) and the PI3K/AKT pathway. Transcription factors regulated by these pathways, as well as STAT3, can bind to the PD-L1 gene, enhancing the transcription rate. TPP destabilizes PD-L1 mRNA at AREs on the 3’UTR region and can be inhibited via MEK signaling in K-RAS mut cancers, prolonging the PD-L1 mRNA’s half-life. This figure was partly created using SMART Servier Medical Art. ARE: AU-rich elements; CR: cytokine receptor; RTK: receptor tyrosine kinase; TTP: tristetraprolin
Nowadays, cancer therapy is able to inhibit both effector pathways downstream of K-RAS via selective MEK- (Binimetinib, Cobimetinib, Selumetinib, Trametinib)[32-36], B-RAF- (Dabrafenib, Vemurafenib, Sorafenib, Encorafenib)[32,37-40], or PI3K- (Idealisib, Buparlisib)[41-44] inhibitors. However, continuous efforts over the past decades failed to develop novel therapies for K-RAS mutant cancer.
K-RAS mutations as a key molecule driving chemoresistance
As there is no targeted therapy against K-RAS mut cancers available today, chemotherapy is still standard care in the treatment of cancer patients harboring mutations in this very oncogene. Pancreatic cancer, associated with a 90% chance of mutations in the K-RAS gene, is very poorly responsive to standard care first and second line gemcitabine/fluoropyrimidine chemotherapy due to metabolism-dependent drug resistances[45,46]. Metabolic reprogramming of K-RAS mut cancers is characterized by boosted glycolysis, glutaminolysis and pentose phosphate pathway, among others. Enhanced metabolism is not only providing more energy to the tumor, but has further impact on cell growth, rapid proliferation, invasion and drug resistance[47]. K-RAS and MYC oncogenes, as well as tumor suppressors like tumor protein p53, or phosphatase and tensin homolog (PTEN) are directly reprogramming a cancer cell’s metabolism in order to sustain unrestricted tumor growth, thereby creating a metabolism addiction[47-49]. The correlation between the strong addiction of cells to this oncogene and its impact on drug resistance highlights the big need to find specific K-RAS antagonists for more efficient future therapies.
Very recent advances in developing low molecular weight antagonists of RAS
RAS proteins already were in the focus of medicinal chemical endeavors in the last millennium but unfortunately proved to be difficult targets. This previously earned RAS the inglorious title of being “undruggable”. During the past decade, tremendous progress has been made in modulating RAS by low molecular weight compounds, rendering RAS now indeed “druggable”. In principle, there are three common approaches to address RAS directly. These include to target the CaaX-Box, the nucleotide binding domain (NBD) or a shallow hydrophobic pocket including the switches I and II.
The CaaX-Box of RAS as a conceptionally new target site
The first attempts to target the C-terminal CaaX-Box were based on the idea of interrupting the membrane binding and hence abolishing the signal transduction of RAS. The farnesyltransferase (FTase) is responsible for the farnesylation of all RAS isoforms at cysteine 185/186. The development of FTase inhibitors led to various products, of which some have entered clinical trials. Probably the most promising one is Tipifarnib that is currently in clinical trials[50]. However, studies have shown that Tipifarnib and other FTase inhibitors only affect the isoform H-RAS. The other isoforms, N-RAS as well as K-RAS 4A and 4B, are still prenylated by geranylgeranyl transferase[51].
The G12C RAS mutant as a special target
Targeting the NBD is difficult due to the high picomolar affinity of RAS towards the nucleotide GDP and GTP[52], as well as the high micromolar abundance of intracellular GDP and GTP[53]. One opportunity to compete with these conditions unveils itself in form of the oncogenic mutant K-RAS G12C. The mutation of glycine 12 to cysteine establishes a nucleophilic target in proximity to the NBD for covalent binding of electrophilic inhibitors. SML-8-73-1 is a GDP-analogue which utilizes an electrophilic warhead to tether cysteine 12 and bind to the nucleotide binding pocket of K-RAS[54]. As a GDP-analogue it is able to lock K-RAS G12C in the inactive state. Another way to employ the solvent exposed cysteine anchor is based on the combination of an electrophile with a compound which is able to bind to the neighboring switch II region[55,56]. The crucial aspect of the interaction with the switch II is that the binding pocket is only present in the inactive GDP-bound state. In case of the covalent inhibitor 12 developed by Ostrem et al.[56] the binding to switch II results in disorder of both switch regions and enhances the affinity of K-RAS G12C towards GDP, hence rendering the protein inactive and decreasing the affinity towards the effector RAF. As the covalent tethering only works for the G12C oncogene mutation but not for the other oncogene mutants such as G12V, G12D and Q61H the demand for allosteric based inhibitors binding to the RAS-surface becomes evident.
Targeting the hydrophobic ligand binding site located next to the switch regions of RAS
Pharmaceuticals aiming at the surface of RAS have been extensively studied and different compounds have been synthesized that target the GDP or GTP form. Various peptidomimetics[57] and antibody fragments[58] have been characterized as possible antagonists for RAS based tumors. This review however, will focus on low-molecular weight compounds for RAS. The ligands discussed in the following chapter are presented in Figure 4 and are numbered according to the order in this figure.
Figure 4.

Low-molecular weight compounds targeting the RAS hydrophobic binding pocket
In 2012, two groups, one from Genentech and the other from Vanderbilt University, independently published the discovery of ligands that bind reversibly to a shallow hydrophobic pocket located next to switch I and II in the GDP-bound state of K-RAS 4B[59,60]. For both compounds, (1) and (2) the interaction site overlaps with the GEF binding site and hence interferes with the RAS-GEF complex formation. In addition, Schöpel et al.[61,66] reported bisphenol derivatives to bind to the same hydrophobic binding pocket in the sub-millimolar to micromolar range. Bisphenol A (BPA) is widely used as a plasticizer best known for its endocrine disrupting character[67]. Notably, while BPA antagonizes the interaction between RAS and SOS, the fluorinated bisphenols BPAF and BPNH2 (3) even induce an allosteric activation of the GDP-bound K-RAS 4B. This induces a transition of RAS from the inactive, i.e., OFF-state, to a state in-between the OFF- and ON-state that is active[61].
In 2017, Xie et al.[62] reported on another ligand (4), that interacts with the hydrophobic binding pocket with the potential to hamper the binding of GTP. Further studies with non-small cell lung cancer (NSCLC) cell lines have shown that these compounds induce apoptosis in cell lines with oncogenic mutants of K-RAS 4B but not in wt K-RAS 4B cell lines.
Modulating the transition from inactive to active RAS is a reasonable strategy in disrupting the signal transition, but so is the interruption of the RAS-Effector complex of the active GTP-bound form. Molecular Dynamic simulations of the switch regions have identified a cluster of small binding domains which are present in all RAS isoforms[63]. Welsch et al.[63] developed a so-called Pan-RAS inhibitor (5), which shows micromolar affinity in vitro for the RAS isoforms K-RAS, H-RAS, and N-RAS in the active state. This compound was tested in in vivo experiments using xenograft mouse tumor models and was shown to inhibit tumor growth over a period of 15 days of treatment. Notably, this study also reports certain levels of toxicity as well as off-target activity, which could hamper the indicated tumor growth inhibition.
As mentioned above, antibody fragment-based therapeutics have already been tested (Antibody 1998), but prove to be challenging regarding their permeability into the cell. Based on the structural information of the interaction between the antibody and its target site, Quevedo et al.[64] have generated a low-molecular weight compound, Abd-7(6), which targets the same region with nanomolar affinity. Abd-7 binds to the switch regions and antagonizes the protein-protein interaction between RAS and its effectors. Abd-7 interacts with K-RAS G12V, which was bound to a non-hydrolysable GTP analogue, with an affinity of 51 nmol/L. However, its IC50 in cells is only 8 µmol/L that could suggest off-target effects and, once again, highlights the difference between molecular affinity in vitro and biological potency in vivo.
Finally, a novel inhibitor (7), by McCarthy et al.[65] selectively interacts with both the GTP-bound wt and oncogenic mutants of K-RAS with sub-micromolar affinity but neither binds to the isoform H-RAS nor N-RAS[65]. The binding region of this pyrazolopyrimidine-inhibitor overlaps with the switch regions and thereby enables the inhibition of the RAS-RAF complex. The differentiation between the active and inactive state of RAS targets presents a promising opportunity for the future development of much needed selective inhibitors of activating oncogenic RAS-mutations, such as G12V, G12D, and Q61H.
As described in the previous chapter, the presented ligands all bind to a hydrophobic binding pocket adjacent to the switch I and switch II region. To illustrate the binding orientation of the ligands the co-crystal structure of (2), [4EPY] was used in a surface projection. The co-crystal structures of (1) [4DST] and (6) [6FA4] as well as (3) NMR data-based docked onto 4DSO[61] were then aligned to 4EPY [Figure 5].
Figure 5.
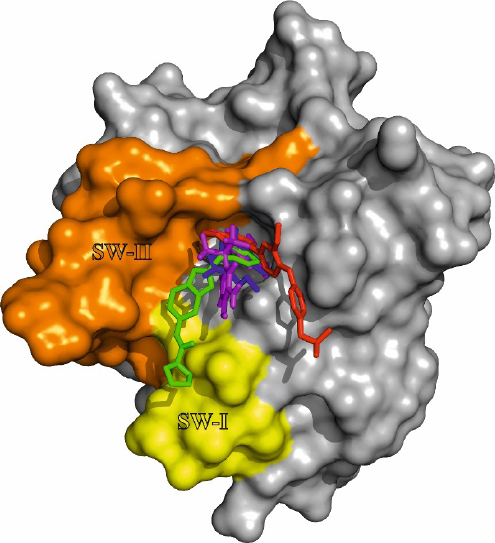
Ligand orientation in the hydrophobic binding pocket of K-RAS 4B. Co-crystal structure of (2) with K-RAS G12V (aa 1-169) [4EPY] in surface projection. The co-crystal structure of (1) [4DST] and (6) [6FA4] as well as (3) docked to [4DSO] have been aligned to 4EPY. The compounds are color-coded as follows: (1) blue; (2) green; (3) magenta; (6) red. The switch regions adjacent to the hydrophobic binding pocket are labeled switch I SW-I (yellow) and switch II SW-II (orange)
Influence of oncogenic K-RAS on cancer immunotherpy
The programmed cell death protein 1 and its ligands
Evidence that inflammatory triggers play a critical role in the development and progression of cancer was already emerging at the beginning of the 20th century. Cancer patients were infected with bacteria to provoke inflammation which also affected the tumor cells[68]. For example, until today non-muscle invasive bladder cancer patients are receiving a therapy based on the infection of the bladder with bacillus-calmette-guérin[69].
In 2018, the Nobel Prize for medicine and physiology was dedicated to the US American immunologist James Allison and the Japanese immunologist Tasku Honjo for their discovery of so called “immune checkpoints” and their therapeutic “mode of action” as negative immune regulators. Programmed cell death protein 1 (PD-1), also known as CD279, discovered in 1992 by Honjo et al.[70] is a transmembrane immunoinhibitory receptor of the CD28 family, mostly expressed on active T-cells. Binding of PD-1 with its ligands (PD-L1/L2; known as CD274 and CD273) transmits a co-inhibitory signal in activated T-cells that promotes T-cell exhaustion, leading to tumor immune evasion[71]. However, contradictory results have suggested that PDL2 can serve as a negative and a positive regulator of T cell function. In 2004, Honjo et al.[72] also discovered expression of PD-L1 on tumor cells, a potential strategy to suppress the host immune response and escape immune destruction. By using different preclinical models for melanoma and colon cancer he could show that a genetically or antibody-based PD-1 depletion leads to diminished hematogenous dissemination[72]. These experiments were the basis of the second immunotherapy approach of blocking PD-1/PD-L1 interactions. The anti-PD-1 antibody Nivolumab, in 2014 FDA approved for the treatment of advanced melanoma patients, is by the end of 2017and after more than 500 conducted clinical trials now approved for the treatment of nine different tumor entities including e.g., NSCLC, hodgkin lymphoma, squamous head and neck cancer or urothelial cancer[73]. Further FDA approved agents targeting the PD-1/PD-L1 axis e.g., Pembrolizumab (PD-1 inhibitor), Atezolizumab and Avelumab (both targeting PD-L1) are additionally as mono- or combination therapy in clinical use.
Regulation of PD-L1 expression via oncogenic RAS signaling
Hanahan and Weinberg[74] defined the capability of cancer cells to suppress the immune system, and to evade immune responses/destruction as a hallmark of cancer[74]. Moreover, the gain of mutations in cancer cells leads to changes in expression of surface molecules on the plasma membrane, so called neoantigens. The immune system is capable to recognize these neoantigens driving cytotoxic T-cells to destroy cancer cells. The immune system has certain breaks to dim or block this immune reaction, a mechanism preventing severe autoimmune responses, known as immune exhaustion[75]. As K-RAS mutant cancer is characterized by a high presentation of neoantigens that should activate T-cell response, these tumors have the potential to escape from immune destruction. Since therapeutic monoclonal antibodies targeting the immune checkpoints was a breakthrough by demonstrating long-term survival benefit for some patients with solid cancer, recent evidence suggest that there might be a connection between the regulation of the PD1/PD-L1 axis and K-RAS signaling.
Recent progress in developing small molecules against the formerly deemed “undruggable” K-RAS gives hope to patients suffering from K-RAS mutant cancer. However, there are still no clinically active drugs against mutant K-RAS variants available. Thus, understanding the complex interplay of oncogenic signaling and the immune system may help to circumvent mechanisms of resistance to immune checkpoint inhibitors. Preclinical and clinical data assumed that upregulation of PD-L1 on the cell surface of tumor cells seems to be a major driver of immune evasion. It was reported, that K-RAS mut NSCLC patients showed a prolonged overall survival after a Nivolumab treatment compared to K-RAS wt patients, who did not benefit from a PD-1 blockade[76]. Moreover, Garon et al.[76] recently reported increased PD-L1 expression in K-RAS mut vs. K-RAS wt NSCLC tumors[77]. Therefore, it is of major importance to unravel the molecular mechanisms involved in the reciprocal interaction of activated K-RAS and presentation of PD-L1.
The best characterized stimulus from the tumor microenvironment that leads to up-regulation of PD-L1 expression is the inflammatory cytokine interferon (IFN) γ, acting via activation of the Janus kinase (JAK)/STAT pathway[78]. It has been shown that STAT3 binds to the PD-L1 promotor region, enhancing its transcription in vitro and in vivo[79]. This mechanism was proven in anaplastic large-cell lymphoma, T-cell lymphoma and in K-RAS mut NSCLC[79-81]. Recent evidence suggests that oncogenic RAS signaling can directly increase PD-L1 mRNA stability, which leads to enhanced PD-L1 surface expression and impaired tumor immune surveillance[82]. Whereas PD-L1 protein level were significantly increased in BRAF V600E mut melanoma cell lines resistant to BRAF inhibition, this effect was decreased both after using small interfering RNA (siRNA) mediated knockdown of ERK1/2, and after pharmacologic inhibition of MEK[83,84]. Jiang et al.[83] further showed that depletion of c-Jun (transcription factor downstream of ERK) and STAT3 resulted in a synergistic decrease of PD-L1 surface expression, indicating a cooperation of RAS and STAT3 signaling in controlling PD-L1 expression in melanoma. MEK and ERK abrogation was further effective to minimize ectopic PD-L1 expression in K-RAS mut NSCLC cell lines[81].
Moreover, the second major effector pathway of RAS, the PI3K/AKT pathway, also seems to play a role in regulating PD-L1 expression, as inhibition of either AKT or mTOR resulted in decreased PD-L1 levels in triple negative breast cancer (TNBC)[85]. This is in line with the observation that a knockdown of the PTEN (negative regulator of P13K) leads to elevated PD-L1 expression level in the same TNBC cell model, as well as in a murine lung squamous cell carcinoma model[86]. Inhibition of PI3K resulted in PD-L1 down-regulation in many tumor entities including K-RAS or EGFR mut NSCLC, renal cell carcinoma and melanoma[83,87,88].
Coelho et al.[82] were the first describing a novel translational control mechanism of PD-L1 expression via direct mut K-RAS signaling. By using a tamoxifen-inducible ER-KRASG12V fusion protein they could show a profound up-regulation of PD-L1 mRNA and surface protein level upon activation of oncogenic RAS. In contrast to IFN-γ dependent PD-L1 regulation, induction of PD-L1 via activated RAS is mediated by an increased half-life of the PD-L1 mRNA[82]. AU-rich elements (AREs) of 3’ untranslated regions (UTR) of the PD-L1 mRNA are targets of the ARE-binding protein tristetraprolin (TTP). Once TTP is physically bound to AREs, it destabilizes the PD-L1 mRNA. It was proven in lung cancer cell lines, that mut K-RAS driven MEK signaling reduces intracellular TTP levels, thereby prolonging PD-L1mRNA’s half-life and stability, which in turn increased the cells PD-L1 surface expression. Additionally, TTP restoration in K-RAS mut tumors resulted in enhanced anti-tumor immunity in murine colon cancer models[82]. Curiously, a TTP deficiency in macrophages and cytotoxic T-cells increases T-cell induced anti-tumor activities via modified cytokine expression[89].
Interestingly, genomic alterations in the 3’UTR region of the PD-L1 gene itself have an impact on the PD-L1 expression in some cancers[90], which may indicate a shared TTP regulation mechanism of the PD-L1 mRNA, as it is the case in K-RAS mut cancers.
Clinical implication of K-RAS status and PD-L1 expression in cancer
Continuous efforts in the past three decades failed to develop approved therapies targeting activated K-RAS. Although it is too early to claim clinical benefits, new therapeutic strategies can evolve from the knowledge about the interaction of these key molecules in tumor biology. So, the strategy to combine immunotherapies has brought new hope for K-RAS-dependent tumors. A more recent clinical study analyzed the impact of the individual K-RAS mutations on PD-L1 expression and clinical outcome in lung adenocarcinoma patients. They showed that exclusively G12V, and none of the other mutations in K-RAS, leads to a significant increase in PD-L1 expression[91]. This study further states a correlation between high PD-L1 expression and improved OS only in K-RAS mut cancer. Additionally, Falk et al.[91] reported that high PD-L1 expressing immune cells in these patients correlated with poor OS, independent on the tumor’s K-RAS mutation. These findings show how heterogeneous K-RAS mutational status affects the immune system in lung adenocarcinomas, indicating a K-RAS dependent patient stratification for immunotherapy in future.
Combination of small molecules and PD-1 inhibition in patients with RAS mutant tumors
Currently there are many preclinical analyses and clinical trials ongoing to target K-RAS-driven cancer. A broad range of novel therapeutic strategies, e.g., the usage of siRNA and exosome siRNA, monoclonal antibodies targeting the GTP-bound K-RAS, use of oligonucleotides to prevent membrane embedding and farnesyltransferase inhibitors implicate new hope. However, it will take some time before we can draw conclusions about the effectiveness of these new therapeutic approaches. Nevertheless, a currently recruiting phase 1b clinical trial, initiated by Targovax, is evaluating the effect of the multi-kinase inhibitor TG02 in combination with the anti-PD-1 antibody Pembrolizumab in patients with locally advanced primary tumors in RAS mutated colorectal cancers (ID: CT TG02-01). TG02, a second-generation RAS neo-antigen vaccine, selectively targets tyrosine kinase receptors (e.g., fms-like tyrosine kinase receptor-3), mitotic and transcriptional cyclin dependent kinases 1, 2, 3, 5, as well as lymphocyte-specific protein tyrosine kinase, proto-oncogene tyrosine-protein kinase Fyn, non-receptor tyrosine-protein kinase 2 and JAK 2[92,93]. TG02 is reported to induce apoptosis in cell lines and primary cells of acute myeloid leukemia, and lead to tumor regression in xenograft models[93]. In 2018, Su et al.[94] reported induction of apoptosis, reduced tumor cell proliferation and a prolonged OS of mice harboring glioblastoma after TG02 treatment. Early exploratory clinical data indicate that TG02 induces immune response in patients with so called “hot tumors” which are rich in numbers of activated tumor-infiltrating T-cells compared to controls. Moreover, PD-1 expression was observed in both circulating and tumor-infiltrating T-cells. This further strengthens the rationale for combining TG02 with PD-1 checkpoint blockade. Whether this dual targeting would be advantageous compared with the previous therapeutic strategies still await clinical validation.
Conclusion
RAS oncogenes are often activated by point mutations in a wide range of different cancer entities, thereby identifying RAS as an important clinical target. There are three RAS genes (H-RAS, K-RAS and N-RAS) whereas K-RAS is the most frequently mutated oncogene in cancer. Activating somatic mutations of this proto-oncogene occur in approximately 30% of human cancers driving neoplastic transformation. The K-RAS protein is lacking proper binding pockets for small molecules, resulting in no progress in drug development over the past thirty years. Recently new findings e.g., the identification of a new binding pocket interfering with the SOS-K-RAS interaction has aroused the interest to re-visit the “formerly undruggable” target. In addition, more recent insights into the interplay between mutant RAS and the PD-1/PD-L1 signaling provide a portfolio of synergistic treatment options through the additional use of inhibitors targeting the immune checkpoints. Nevertheless, although novel therapeutic directions are currently under investigation, the risk for induction of acquired resistance is also in clinically effective drugs expected given by the strong selective pressure applied to the genetically unstable cancer cells.
Declarations
Authors’ contributions
Writing and original draft preparation: Mörchen B, Shkura O, Stoll R, Helfrich I
Review and editing: Stoll R, Helfrich I
Availability of data and materials
Not applicable.
Financial support and sponsorship
This work was partially supported by the European Fond for Regional Development (EFRD) (EFRE-0800951) to Mörchen B and Helfrich I; and (EFRE-0800947) to Shkura O and Stoll R.
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2019.
References
- 1.Feig LA, Buchsbaum RJ. Cell signaling: life or death decisions of Ras proteins. Curr Biol. 2002;12:259–61. doi: 10.1016/s0960-9822(02)00787-x. [DOI] [PubMed] [Google Scholar]
- 2.Shields JM, Pruitt K, Shaub A, Der CJ. Understanding Ras : ‘it ain’t over ‘til it’s over’. Trends Cell Biol. 2000;10:147–54. doi: 10.1016/s0962-8924(00)01740-2. [DOI] [PubMed] [Google Scholar]
- 3.Bos JL. ras oncogenes in human cancer : a review. Cancer Res. 1989;49:4682–9. [PubMed] [Google Scholar]
- 4.Vetter IR, Wittinghofer A. The guanine nucleotide - binding switch in three dimensions. Science. 2001;294:1299–304. doi: 10.1126/science.1062023. [DOI] [PubMed] [Google Scholar]
- 5.Lenzen C, Cool RH, Prinz H, Wittinghofer A. Kinetic analysis by fluorescence of the interaction between Ras and the catalytic domain of the guanine nucleotide exchange factor Cdc25Mm. Biochemistry. 1998;37:7420–30. doi: 10.1021/bi972621j. [DOI] [PubMed] [Google Scholar]
- 6.Karnoub AE, Weinberg RA. Ras oncogenes: split personalities. Nat Rev Mol Cell Biol. 2008;9:517–31. doi: 10.1038/nrm2438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Vetter IR. The structure of the G domain of the Ras superfamily. Ras Superfamily Small G Proteins: Biology and Mechanisms 1. Wien: Springer Verlag; 2014. pp. 25–51. [Google Scholar]
- 8.Wittinghofer A, Vetter IR. Structure-function relationships of the G domain, a canonical switch motif. Ann Rev Biochem. 2011;80:943–71. doi: 10.1146/annurev-biochem-062708-134043. [DOI] [PubMed] [Google Scholar]
- 9.Spoerner M, Nuehs A, Herrmann C, Steiner G, Kalbitzer HR. Slow conformational dynamics of the guanine nucleotide-binding protein Ras complexed with the GTP analogue GTPγS. FEBS J. 2007;274:1419–33. doi: 10.1111/j.1742-4658.2007.05681.x. [DOI] [PubMed] [Google Scholar]
- 10.Spoerner M, Hozsa C, Poetzl JA, Reiss K, Ganser P, et al. Conformational states of human rat sarcoma (Ras) protein complexed with its natural ligand GTP and their role for effector interaction and GTP hydrolysis. J Biol Chem. 2010;285:39768–78. doi: 10.1074/jbc.M110.145235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ito Y, Yamasaki K, Iwahara J, Terada T, Kamiya A, et al. Regional polysterism in the GTP-bound form of the human c-Ha-Ras protein. Biochemistry. 1997;36:9109–19. doi: 10.1021/bi970296u. [DOI] [PubMed] [Google Scholar]
- 12.Barbacid M. ras genes. Annu Rev Biochem. 1987;56:779–827. doi: 10.1146/annurev.bi.56.070187.004023. [DOI] [PubMed] [Google Scholar]
- 13.Malumbres M, Barbacid M. RAS oncogenes: the first 30 years. Nat Rev Cancer. 2003;3:459–65. doi: 10.1038/nrc1097. [DOI] [PubMed] [Google Scholar]
- 14.Vetter IR, Wittinghofer A. The guanine nucleotide-binding switch in three dimensions. Science. 2001;294:1299–304. doi: 10.1126/science.1062023. [DOI] [PubMed] [Google Scholar]
- 15.Hancock JF. Ras proteins: different signals from different locations. Nat Rev Mol Cell Biol. 2003;4:373–84. doi: 10.1038/nrm1105. [DOI] [PubMed] [Google Scholar]
- 16.Lemmon MA, Schlessinger J. Cell signaling by receptor tyrosine kinases. Cell. 2010;141:1117–34. doi: 10.1016/j.cell.2010.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cox AD, Der CJ. Ras history: the saga continues. Small GTPases. 2010;1:2–27. doi: 10.4161/sgtp.1.1.12178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gibbs JB, Sigal IS, Poe M, Scolnick EM. Intrinsic GTPase activity distinguishes normal and oncogenic ras p21 molecules. Proc Natl Acad Sci U S A. 1984;81:5704–8. doi: 10.1073/pnas.81.18.5704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.McGrath JP, Capon DJ, Goeddel DV, Levinson AD. Comparative biochemical properties of normal and activated human ras p21 protein. Nature. 1984;310:644–9. doi: 10.1038/310644a0. [DOI] [PubMed] [Google Scholar]
- 20.Sweet RW, Yokoyama S, Kamata T, Feramisco JR, Rosenberg M, et al. The product of ras is a GTPase and the T24 oncogenic mutant is deficient in this activity. Nature. 1984;311:273–5. doi: 10.1038/311273a0. [DOI] [PubMed] [Google Scholar]
- 21.Scheffzek K, Lautwein A, Kabsch W, Ahmadian MR, Wittinghofer A. Crystal structure of the GTPase-activating domain of human p120GAP and implications for the interaction with Ras. Nature. 1996;384:591–6. doi: 10.1038/384591a0. [DOI] [PubMed] [Google Scholar]
- 22.Schubbert S, Shannon K, Bollag G. Hyperactive Ras in developmental disorders and cancer. Nat Rev Cancer. 2007;7:295–308. doi: 10.1038/nrc2109. [DOI] [PubMed] [Google Scholar]
- 23.Jemal A, Siegel R, Xu J, Ward E. Cancer statistics, 2010. CA Cancer J Clin. 2010;60:277–300. doi: 10.3322/caac.20073. [DOI] [PubMed] [Google Scholar]
- 24.Dhillon AS, Hagan S, Rath O, Kolch W. MAP kinase signalling pathways in cancer. Oncogene. 2007;26:3279–90. doi: 10.1038/sj.onc.1210421. [DOI] [PubMed] [Google Scholar]
- 25.De Luca A, Maiello MR, D'Alessio A, Pergameno M, Normanno N. The RAS/RAF/MEK/ERK and the PI3K/AKT signalling pathways: role in cancer pathogenesis and implications for therapeutic approaches. Expert Opin Ther Targets. 2012;16 Suppl 2:S17–27. doi: 10.1517/14728222.2011.639361. [DOI] [PubMed] [Google Scholar]
- 26.Pullikuth AK, Catling AD. Scaffold mediated regulation of MAPK signaling and cytoskeletal dynamics: a perspective. Cell Signal. 2007;19:1621–32. doi: 10.1016/j.cellsig.2007.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ma L, Chen Z, Erdjument-Bromage H, Tempst P, Pandolfi PP. Phosphorylation and functional inactivation of TSC2 by Erk implications for tuberous sclerosis and cancer pathogenesis. Cell. 2005;121:179–93. doi: 10.1016/j.cell.2005.02.031. [DOI] [PubMed] [Google Scholar]
- 28.Choi M, Bien H, Mofunanya A, Powers S. Challenges in Ras therapeutics in pancreatic cancer. Semin Cancer Biol. 2019;54:101–8. doi: 10.1016/j.semcancer.2017.11.015. [DOI] [PubMed] [Google Scholar]
- 29.Yang SH, Sharrocks AD, Whitmarsh AJ. MAP kinase signalling cascades and transcriptional regulation. Gene. 2013;513:1–13. doi: 10.1016/j.gene.2012.10.033. [DOI] [PubMed] [Google Scholar]
- 30.Andjelkovic M, Alessi DR, Meier R, Fernandez A, Lamb NJ, et al. Role of translocation in the activation and function of protein kinase B. J Biol Chem. 1997;272:31515–24. doi: 10.1074/jbc.272.50.31515. [DOI] [PubMed] [Google Scholar]
- 31.Cai SL, Tee AR, Short JD, Bergeron JM, Kim J, et al. Activity of TSC2 is inhibited by AKT-mediated phosphorylation and membrane partitioning. J Cell Biol. 2006;173:279–89. doi: 10.1083/jcb.200507119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Long GV, Hauschild A, Santinami M, Atkinson V, Mandala M, et al. Adjuvant Dabrafenib plus Trametinib in stage III BRAF-mutated melanoma. N Engl J Med. 2017;377:1813–23. doi: 10.1056/NEJMoa1708539. [DOI] [PubMed] [Google Scholar]
- 33.Janne PA, van den Heuvel MM, Barlesi F, Cobo M, Mazieres J, et al. Selumetinib plus docetaxel compared with docetaxel alone and progression-free survival in patients with KRAS-mutant advanced non-small cell lung cancer: the SELECT-1 randomized clinical trial. JAMA. 2017;317:1844–53. doi: 10.1001/jama.2017.3438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Pulido JS, Heier JS, Marmorstein AD. Selumetinib in plexiform neurofibromas. N Engl J Med. 2017;376:1195. doi: 10.1056/NEJMc1701029. [DOI] [PubMed] [Google Scholar]
- 35.Signorelli J, Shah Gandhi A. Cobimetinib. Ann Pharmacother. 2017;51:146–53. doi: 10.1177/1060028016672037. [DOI] [PubMed] [Google Scholar]
- 36.Dummer R, Schadendorf D, Ascierto PA, Arance A, Dutriaux C, et al. Binimetinib versus dacarbazine in patients with advanced NRAS-mutant melanoma (NEMO): a multicentre, open-label, randomised, phase 3 trial. Lancet Oncol. 2017;18:435–45. doi: 10.1016/S1470-2045(17)30180-8. [DOI] [PubMed] [Google Scholar]
- 37.Nijenhuis CM, Huitema AD, Blank C, Haanen JB, van Thienen JV, et al. Clinical Pharmacokinetics of Vemurafenib in BRAF-mutated melanoma patients. J Clin Pharmacol. 2017;57:125–8. doi: 10.1002/jcph.788. [DOI] [PubMed] [Google Scholar]
- 38.Zhang W, Heinzmann D, Grippo JF. Clinical pharmacokinetics of vemurafenib. Clin Pharmacokinet. 2017;56:1033–43. doi: 10.1007/s40262-017-0523-7. [DOI] [PubMed] [Google Scholar]
- 39.Delord JP, Robert C, Nyakas M, McArthur GA, Kudchakar R, et al. Phase I dose-escalation and -expansion study of the BRAF inhibitor encorafenib (LGX818) in metastatic BRAF-mutant melanoma. Clin Cancer Res. 2017;23:5339–48. doi: 10.1158/1078-0432.CCR-16-2923. [DOI] [PubMed] [Google Scholar]
- 40.Gounder MM, Mahoney MR, Van Tine BA, Ravi V, Attia S, et al. Sorafenib for advanced and refractory desmoid tumors. N Engl J Med. 2018;379:2417–28. doi: 10.1056/NEJMoa1805052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Di Leo A, Johnston S, Lee KS, Ciruelos E, Lonning PE, et al. Buparlisib plus fulvestrant in postmenopausal women with hormone-receptor-positive, HER2-negative, advanced breast cancer progressing on or after mTOR inhibition (BELLE-3): a randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Oncol. 2018;19:87–100. doi: 10.1016/S1470-2045(17)30688-5. [DOI] [PubMed] [Google Scholar]
- 42.Furman RR, Sharman JP, Coutre SE, Cheson BD, Pagel JM, et al. Idelalisib and rituximab in relapsed chronic lymphocytic leukemia. N Engl J Med. 2014;370:997–1007. doi: 10.1056/NEJMoa1315226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gopal AK, Kahl BS, de Vos S, Wagner-Johnston ND, Schuster SJ, et al. PI3Kdelta inhibition by idelalisib in patients with relapsed indolent lymphoma. N Engl J Med. 2014;370:1008–18. doi: 10.1056/NEJMoa1314583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Younes A, Salles G, Martinelli G, Bociek RG, Barrigon DC, et al. Pan-phosphatidylinositol 3-kinase inhibition with buparlisib in patients with relapsed or refractory non-Hodgkin lymphoma. Haematologica. 2017;102:2104–12. doi: 10.3324/haematol.2017.169656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wang C, Liu B, Xu X, Zhuang B, Li H, et al. Toward targeted therapy in chemotherapy-resistant pancreatic cancer with a smart triptolide nanomedicine. Oncotarget. 2016;7:8360–72. doi: 10.18632/oncotarget.7073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Takai E, Yachida S. Genomic alterations in pancreatic cancer and their relevance to therapy. World J Gastrointest Oncol. 2015;7:250–8. doi: 10.4251/wjgo.v7.i10.250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Grasso C, Jansen G, Giovannetti E. Drug resistance in pancreatic cancer: Impact of altered energy metabolism. Crit Rev Oncol Hematol. 2017;114:139–52. doi: 10.1016/j.critrevonc.2017.03.026. [DOI] [PubMed] [Google Scholar]
- 48.Hezel AF, Kimmelman AC, Stanger BZ, Bardeesy N, Depinho RA. Genetics and biology of pancreatic ductal adenocarcinoma. Genes Dev. 2006;20:1218–49. doi: 10.1101/gad.1415606. [DOI] [PubMed] [Google Scholar]
- 49.Cetinbas NM, Sudderth J, Harris RC, Cebeci A, Negri GL, et al. Glucose-dependent anaplerosis in cancer cells is required for cellular redox balance in the absence of glutamine. Sci Rep. 2016;6:32606. doi: 10.1038/srep32606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Soleimani A, Amirinejad M, Rahsepar S, Vazirian F, Bahrami A, et al. Therapeutic potential of RAS prenylation pharmacological inhibitors in the treatment of breast cancer, recent progress, and prospective. J Cell Biochem. 2019;120:6860–7. doi: 10.1002/jcb.27992. [DOI] [PubMed] [Google Scholar]
- 51.Whyte DB, Kirschmeier P, Hockenberry TN, Nunez-Oliva I, James L, et al. K- and N-Ras are geranylgeranylated in cells treated with farnesyl protein transferase inhibitors. J Biol Chem. 1997;272:14459–64. doi: 10.1074/jbc.272.22.14459. [DOI] [PubMed] [Google Scholar]
- 52.Feuerstein J, Kalbitzer HR, John J, Goody RS, Wittinghofer A. Characterisation of the metal-ion-GDP complex at the active sites of transforming and nontransforming p21 proteins by observation of the 17O-Mn superhyperfine coupling and by kinetic methods. Eur J Biochem. 1987;162:49–55. doi: 10.1111/j.1432-1033.1987.tb10540.x. [DOI] [PubMed] [Google Scholar]
- 53.Traut TW. Physiological concentrations of purines and pyrimidines. Mol Cell Biochem. 1994;140:1–22. doi: 10.1007/BF00928361. [DOI] [PubMed] [Google Scholar]
- 54.Lim SM, Westover KD, Ficarro SB, Harrison RA, Choi HG, et al. Therapeutic targeting of oncogenic K-Ras by a covalent catalytic site inhibitor. Angew Chem Int Ed Engl. 2014;53:199–204. doi: 10.1002/anie.201307387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Patricelli MP, Janes MR, Li LS, Hansen R, Peters U, et al. Selective inhibition of oncogenic KRAS output with small molecules targeting the inactive state. Cancer Discov. 2016;6:316. doi: 10.1158/2159-8290.CD-15-1105. [DOI] [PubMed] [Google Scholar]
- 56.Ostrem JM, Peters U, Sos ML, Wells JA, Shokat KM. K-Ras(G12C) inhibitors allosterically control GTP affinity and effector interactions. Nature. 2013;503:548–51. doi: 10.1038/nature12796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Patgiri A, Yadav KK, Arora PS, Bar-sagi D. An orthosteric inhibitor of Ras-Sos interaction. Nat Chem Biol. 2011;7:585–7. doi: 10.1038/nchembio.612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Cochet O, Kenigsberg M, Delumeau I, Virone-Oddos A, Multon MC, et al. Intracellular Expression of an Antibody Fragment-neutralizing p21 Ras Promotes Tumor Regression. Cancer Research. 1998;58:1170. [PubMed] [Google Scholar]
- 59.Maurer T, Garrenton LS, Oh A, Pitts K, Anderson DJ, et al. Small-molecule ligands bind to a distinct pocket in Ras and inhibit SOS-mediated nucleotide exchange activity. Proc Natl Acad Sci U S A. 2012;109:5299–304. doi: 10.1073/pnas.1116510109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Sun Q, Burke JP, Phan J, Burns MC, Olejniczak ET, et al. Discovery of small molecules that bind to K-Ras and inhibit Sos-mediated activation. Angew Chem Int Ed Engl. 2012;51:6140–3. doi: 10.1002/anie.201201358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Schöpel M, Shkura O, Seidel J, Kock K, Zhong X, et al. Allosteric activation of GDP-bound ras isoforms by bisphenol derivative plasticisers. Int J Mol Sci. 2018;19:1133. doi: 10.3390/ijms19041133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Xie C, Li Y, Li LL, Fan XX, Wang YW, et al. Identification of a new potent inhibitor targeting KRAS in non-small cell lung cancer cells. Front Pharmacol. 2017;8:823. doi: 10.3389/fphar.2017.00823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Welsch ME, Kaplan A, Chambers JM, Stokes ME, Bos PH, et al. Multivalent small-molecule Pan-RAS inhibitors. Cell. 2017;168:878–89.e29. doi: 10.1016/j.cell.2017.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Quevedo CE, Cruz-Migoni A, Bery N, Miller A, Tanaka T, et al. Small molecule inhibitors of RAS-effector protein interactions derived using an intracellular antibody fragment. Nature communications. 2018;9:3169. doi: 10.1038/s41467-018-05707-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.McCarthy MJ, Pagba CV, Prakash P, Naji AK, van der Hoeven D, et al. Discovery of high-affinity noncovalent allosteric KRAS inhibitors that disrupt effector binding. ACS Omega. 2019;4:2921–30. doi: 10.1021/acsomega.8b03308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Schöpel M, Jockers KFG, Düppe PM, Autzen J, Potheraveedu VN, et al. Bisphenol a binds to Ras proteins and competes with guanine nucleotide exchange: implications for GTPase-selective antagonists. J Med Chem. 2013;56:9664–72. doi: 10.1021/jm401291q. [DOI] [PubMed] [Google Scholar]
- 67.Rogers JA, Metz L, Yong VW. Review: endocrine disrupting chemicals and immune responses: a focus on bisphenol-A and its potential mechanisms. Mol Immunol. 2013;53:421–30. doi: 10.1016/j.molimm.2012.09.013. [DOI] [PubMed] [Google Scholar]
- 68.Herr HW, Morales A. History of bacillus calmette-guerin and bladder cancer: an immunotherapy success story. J Urol. 2008;179:53–6. doi: 10.1016/j.juro.2007.08.122. [DOI] [PubMed] [Google Scholar]
- 69.Balakrishnan AS, Washington SL, 3rd, Meng MV, Porten SP. Determinants of guideline-based treatment in patients With cT1 bladder cancer. Clin Genitourin Cancer. 2019;17:e461–71. doi: 10.1016/j.clgc.2019.01.007. [DOI] [PubMed] [Google Scholar]
- 70.Ishida Y, Agata Y, Shibahara K, Honjo T. Induced expression of PD-1, a novel member of the immunoglobulin gene superfamily, upon programmed cell death. EMBO J. 1992;11:3887–95. doi: 10.1002/j.1460-2075.1992.tb05481.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Xia Y, Jeffrey Medeiros L, Young KH. Signaling pathway and dysregulation of PD1 and its ligands in lymphoid malignancies. Biochim Biophys Acta. 2016;1865:58–71. doi: 10.1016/j.bbcan.2015.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Iwai Y, Terawaki S, Honjo T. PD-1 blockade inhibits hematogenous spread of poorly immunogenic tumor cells by enhanced recruitment of effector T cells. Int Immunol. 2005;17:133–44. doi: 10.1093/intimm/dxh194. [DOI] [PubMed] [Google Scholar]
- 73.Gong J, Chehrazi-Raffle A, Reddi S, Salgia R. Development of PD-1 and PD-L1 inhibitors as a form of cancer immunotherapy: a comprehensive review of registration trials and future considerations. J Immunother Cancer. 2018;6:8. doi: 10.1186/s40425-018-0316-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–74. doi: 10.1016/j.cell.2011.02.013. [DOI] [PubMed] [Google Scholar]
- 75.McKinney EF, Smith KG. T cell exhaustion and immune-mediated disease-the potential for therapeutic exhaustion. Curr Opin Immunol. 2016;43:74–80. doi: 10.1016/j.coi.2016.09.005. [DOI] [PubMed] [Google Scholar]
- 76.Garon EB, Rizvi NA, Hui R, Leighl N, Balmanoukian AS, et al. Pembrolizumab for the treatment of non-small-cell lung cancer. N Engl J Med. 2015;372:2018–28. doi: 10.1056/NEJMoa1501824. [DOI] [PubMed] [Google Scholar]
- 77.Borghaei H, Paz-Ares L, Horn L, Spigel DR, Steins M, et al. Nivolumab versus docetaxel in advanced nonsquamous non-small-cell lung cancer. N Engl J Med. 2015;373:1627–39. doi: 10.1056/NEJMoa1507643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Song TL, Nairismagi ML, Laurensia Y, Lim JQ, Tan J, et al. Oncogenic activation of the STAT3 pathway drives PD-L1 expression in natural killer/T-cell lymphoma. Blood. 2018;132:1146–58. doi: 10.1182/blood-2018-01-829424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Marzec M, Zhang Q, Goradia A, Raghunath PN, Liu X, et al. Oncogenic kinase NPM/ALK induces through STAT3 expression of immunosuppressive protein CD274 (PD-L1, B7-H1). Proc Natl Acad Sci U S A. 2008;105:20852–7. doi: 10.1073/pnas.0810958105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Atsaves V, Tsesmetzis N, Chioureas D, Kis L, Leventaki V, et al. PD-L1 is commonly expressed and transcriptionally regulated by STAT3 and MYC in ALK-negative anaplastic large-cell lymphoma. Leukemia. 2017;31:1633–7. doi: 10.1038/leu.2017.103. [DOI] [PubMed] [Google Scholar]
- 81.Sumimoto H, Takano A, Teramoto K, Daigo Y. RAS-mitogen-activated protein kinase signal is required for enhanced PD-L1 expression in human lung cancers. PLoS One. 2016;11:e0166626. doi: 10.1371/journal.pone.0166626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Coelho MA, de Carne Trecesson S, Rana S, Zecchin D, Moore C, et al. Oncogenic RAS signaling promotes tumor immunoresistance by stabilizing PD-L1 mRNA. Immunity. 2017;47:1083–99.e6. doi: 10.1016/j.immuni.2017.11.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Jiang X, Zhou J, Giobbie-Hurder A, Wargo J, Hodi FS. The activation of MAPK in melanoma cells resistant to BRAF inhibition promotes PD-L1 expression that is reversible by MEK and PI3K inhibition. Clin Cancer Res. 2013;19:598–609. doi: 10.1158/1078-0432.CCR-12-2731. [DOI] [PubMed] [Google Scholar]
- 84.Sanlorenzo M, Vujic I, Floris A, Novelli M, Gammaitoni L, et al. BRAF and MEK inhibitors increase PD-1-positive melanoma cells leading to a potential lymphocyte-independent synergism with anti-PD-1 antibody. Clin Cancer Res. 2018;24:3377–85. doi: 10.1158/1078-0432.CCR-17-1914. [DOI] [PubMed] [Google Scholar]
- 85.Mittendorf EA, Philips AV, Meric-Bernstam F, Qiao N, Wu Y, et al. PD-L1 expression in triple-negative breast cancer. Cancer Immunol Res. 2014;2:361–70. doi: 10.1158/2326-6066.CIR-13-0127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Xu C, Fillmore CM, Koyama S, Wu H, Zhao Y, et al. Loss of Lkb1 and Pten leads to lung squamous cell carcinoma with elevated PD-L1 expression. Cancer Cell. 2014;25:590–604. doi: 10.1016/j.ccr.2014.03.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Lastwika KJ, Wilson W, 3rd, Li QK, Norris J, Xu H, et al. Control of PD-L1 Expression by oncogenic activation of the AKT-mTOR pathway in non-small cell lung cancer. Cancer Res. 2016;76:227–38. doi: 10.1158/0008-5472.CAN-14-3362. [DOI] [PubMed] [Google Scholar]
- 88.Balan M, Miery Teran E, Waaga-Gasser AM, Gasser M, Choueiri TK, et al. Novel roles of c-Met in the survival of renal cancer cells through the regulation of HO-1 and PD-L1 expression. J Biol Chem. 2015;290:8110–20. doi: 10.1074/jbc.M114.612689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Wang Q, Ning H, Peng H, Wei L, Hou R, et al. Tristetraprolin inhibits macrophage IL-27-induced activation of antitumour cytotoxic T cell responses. Nat Commun. 2017;8:867. doi: 10.1038/s41467-017-00892-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kataoka K, Shiraishi Y, Takeda Y, Sakata S, Matsumoto M, et al. Aberrant PD-L1 expression through 3'-UTR disruption in multiple cancers. Nature. 2016;534:402–6. doi: 10.1038/nature18294. [DOI] [PubMed] [Google Scholar]
- 91.Falk AT, Yazbeck N, Guibert N, Chamorey E, Paquet A, et al. Effect of mutant variants of the KRAS gene on PD-L1 expression and on the immune microenvironment and association with clinical outcome in lung adenocarcinoma patients. Lung Cancer. 2018;121:70–5. doi: 10.1016/j.lungcan.2018.05.009. [DOI] [PubMed] [Google Scholar]
- 92.Pallis M, Abdul-Aziz A, Burrows F, Seedhouse C, Grundy M, et al. The multi-kinase inhibitor TG02 overcomes signalling activation by survival factors to deplete MCL1 and XIAP and induce cell death in primary acute myeloid leukaemia cells. Br J Haematol. 2012;159:191–203. doi: 10.1111/bjh.12018. [DOI] [PubMed] [Google Scholar]
- 93.Goh KC, Novotny-Diermayr V, Hart S, Ong LC, Loh YK, et al. TG02, a novel oral multi-kinase inhibitor of CDKs, JAK2 and FLT3 with potent anti-leukemic properties. Leukemia. 2012;26:236–43. doi: 10.1038/leu.2011.218. [DOI] [PubMed] [Google Scholar]
- 94.Su YT, Chen R, Wang H, Song H, Zhang Q, et al. Novel targeting of transcription and metabolism in glioblastoma. Clin Cancer Res. 2018;24:1124–37. doi: 10.1158/1078-0432.CCR-17-2032. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.