

Abstract
Successful de novo protein design ideally targets specific folding kinetics, stability thermodynamics, and biochemical functionality, and the simultaneous achievement of all these criteria in a single step design is challenging. Protein design is potentially simplified by separating the problem into two steps: (a) an initial design of a protein “scaffold” having appropriate folding kinetics and stability thermodynamics, followed by (b) appropriate functional mutation—possibly involving insertion of a peptide functional “cassette.” This stepwise approach can also separate the orthogonal effects of the “stability/function” and “foldability/function” tradeoffs commonly observed in protein design. If the scaffold is a protein architecture having an exact rotational symmetry, then there is the potential for redundant folding nuclei and multiple equivalent sites of functionalization; thereby enabling broader functional adaptation. We describe such a “scaffold” and functional “cassette” design strategy applied to a β‐trefoil threefold symmetric architecture and a heparin ligand functionality. The results support the availability of redundant folding nuclei within this symmetric architecture, and also identify a minimal peptide cassette conferring heparin affinity. The results also identify an energy barrier of destabilization that switches the protein folding pathway from monomeric to trimeric, thereby identifying another potential advantage of symmetric protein architecture in de novo design.
Keywords: de novo design, heparin affinity, oligomerization, protein folding, protein stability
1. INTRODUCTION
Protein design offers an infinite possibility of novel protein structures and functionalities with diverse application in the areas of novel therapeutics, diagnostics, sensors, materials, etc. One common approach is computational protein design targeting enzyme engineering, protein specificity, cellular pathway control, and higher‐order protein assembly. 1 , 2 , 3 , 4 , 5 , 6 However, as a single step, computational design has the challenging requirement of solving at least three design problems simultaneously, including (a) protein foldability (i.e., folding kinetics requirements), (b) protein stability (i.e., thermodynamic requirements), and (c) the accommodation of specific function (with potential structural dynamics requirements). These discrete design requirements are interconnected and likely orthogonal, as there is evidence for both a “function/stability tradeoff,” 7 , 8 and “function/foldability tradeoff.” 9 , 10 , 11 Protein design is arguably in a nascent stage, and can therefore benefit from the investigation of alternative strategies to improve successful design outcomes. In the present report, we describe a protein design strategy that separates the overall design process into two discrete steps, involving the initial development of an efficiently folding and thermostable (but functionless) “scaffold,” followed by functionalization (which occurs at the expense of folding and thermostability). The selected scaffold is a symmetric protein architecture having an exact primary structure repeating motif, and an associated tertiary structure having a corresponding rotational symmetry. Such symmetry is postulated to provide redundant definitions of the critical folding nucleus, potentially enabling robust foldability upon subsequent deleterious functional mutation.
De novo protein design utilizing the targeted functionalization of a benign scaffold has a long history but is still being formalized as a design strategy. 12 In the present report, we pursue a functionalization design strategy using a symmetric protein scaffold, testing the hypothesis that symmetric protein architectures may contain redundant instances of the folding nucleus. Symmetric protein scaffolds may therefore exhibit a unique tolerance to destabilizing mutations by utilizing alternative folding pathways made possible by the intrinsic structural symmetry. Symmetric tertiary structure is a common feature in nature, 13 and a number of de novo designed symmetric protein architectures have been reported and present a wide variety of potential design applications. 14 , 15 , 16 , 17 , 18 , 19 , 20 , 21 , 22 , 23
The model system utilized as the protein scaffold in the present report is the de novo designed Symfoil‐4T protein, a β‐trefoil protein. 17 , 18 The β‐trefoil is a common protein architecture that comprises approximately 1% of the proteome, involving 10 different superfamilies. 24 β‐trefoil proteins share a common architecture involving a six‐stranded anti‐parallel β‐barrel, closed at one end via three β‐hairpins. 25 , 26 , 27 This architecture has threefold (i.e., C3) rotational symmetry of a repeating “trefoil” motif comprised of four anti‐parallel β‐strands, with the simplest instance being ~40 amino acids in length. The β‐trefoil family encompasses diverse functionalities, including toxins, protease inhibitors, cytokines, growth factors, agglutinins, lectins, and ligands (SCOP database 28 ), and such functionalities are associated with structural heterogeneity within specific loop regions 29 (Figure 1).
FIGURE 1.

Conserved and variable tertiary structure in β‐trefoil proteins. “Ribbon” diagrams of representative β‐trefoil proteins clostridium neurotoxin (1EPW), inositol 1,4,5P3 receptor (1N4K), and C. cinereal protease inhibitor (3VWC). The conserved fundamental β‐trefoil structural region is indicated by gray shading, and the variable regions are colored. Also shown is the structure of the Symfoil‐4T de novo designed symmetric protein (3O4B). The variable regions of the different β‐trefoil proteins describe surface loops conferring functionality; in contrast, Symfoil‐4T is an essential β‐trefoil “scaffold” devoid of function. The C3 axis of rotational symmetry, characteristic of the β‐trefoil architecture, is aligned vertically in this view. Also indicated are the locations of the N‐ and C‐termini, which are in proximity to each other and define a “discontinuous surface turn” in the overall symmetric architecture
The Symfoil‐4T protein has a number of favorable characteristics as a scaffold relevant to the proposed design strategy: (a) Its tertiary structure has C3 rotational symmetry and the three repeating “trefoil” motifs (42 amino acids) share the same primary structure (Figure S1); (b) It has a β‐trefoil architecture that is devoid of any extended loop regions or specific functionality, and can therefore be considered as a “minimalist” architecture common to the β‐trefoil family 17 , 18 ; (c) The repeating trefoil motif, as an isolated polypeptide, has been shown to contain an effective folding nucleus 17 , 18 , 30 and Symfoil‐4T therefore has the potential for redundant folding nuclei; (d) The protein is a hyperthermophile, having cooperative two‐state folding properties, with no evidence of kinetic trapping in the folding/unfolding pathway 17 , 18 , 30 ; (e) As evidenced by major structural perturbation studies, including circular permutation and extensive regional substitution, Symfoil‐4T is capable of folding despite substantial deleterious mutation. 30 , 31
Given the folding/function and stability/function tradeoff hypotheses, for the purpose of undertaking functionalization design a generally destabilizing mutation could serve as a relevant proxy; however, we have attempted to introduce a specific functionality into Symfoil‐4T. The β‐trefoil family of proteins can be characterized as having an overall ligand functionality with no known enzymes. A number of growth factor and cytokine members of the β‐trefoil family have heparin sulfate (HS) binding functionality; thus, a rudimentary HS‐binding polypeptide “cassette” was designed for the purpose of insertion into, and functionalization of, Symfoil‐4T. This cassette was inserted into Symfoil‐4T at three equivalent positions related by the C3 symmetry. Subsequent biophysical analyses show that the resultant mutants, although significantly destabilized, are folded proteins. Heparin affinity chromatography demonstrates that a rudimentary heparin‐binding functionality was successfully introduced by the inserted peptide cassette. The results also show that foldability was achieved by an alternative folding pathway involving oligomerization (i.e., trimerization). Analysis of mutants involving smaller regions of the HS‐binding cassette show that oligomeric assembly appears critically dependent upon the magnitude of the functional destabilization, and the mutant proteins remain monomeric below a certain destabilization energy threshold. The results indicate that the two‐step strategy of scaffold functionalization in protein design is a potentially viable approach to achieve foldable, stable, and functional proteins; additionally, symmetric protein architecture can provide a unique folding robustness utilizing alternative folding nuclei as well as oligomeric pathways.
2. RESULTS
All heparin sulfate mutants (HSM) expressed as soluble proteins with yields of between 10 and 30 mg/L of culture. Purified proteins were >90% homogenous as determined by densitometry of Coomassie blue stained sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS PAGE).
2.1. Analytical size exclusion chromatography of HSM proteins
Symfoil‐4T comprises 142 amino acids and has a theoretical mass of 15.9 kDa. When resolved on Superdex 200 size exclusion chromatography (SEC) in 50 mM Na2PO4, 100 mM NaCl, 10 mM (NH4)2SO4 pH 7.5 (“phosphate buffer”). Symfoil‐4T elutes with an apparent mass of 28 kDa calibrated against a protein mass standard comprising hen egg white lysozyme (HEW; 14.3 kDa), carbonic anhydrase (31.0 kDa), ovalbumin (45.0 kDa), bovine serum albumin (66.2 kDa), and immunoglobulin G (IgG; 153 kDa). HSM1, HSM2, and HSM3 mutant proteins are each 144 amino acids in length and have a calculated mass of 16.1 kDa. HSM1 resolves on SEC as a single peak with an apparent mass of 91 kDa (Figure 2). HSM2 resolves on SEC as an apparent heterogeneous mixture having a major peak at 56 kDa and a secondary peak at 28 kDa. Isolation of the HSM2 56 kDa major peak, followed by overnight equilibration and repeated SEC, demonstrated a redistribution into the characteristic heterogenous profile observed in the initial chromatogram. HSM3 resolves on SEC with four peaks having an apparent mass of 600, 67, 35, and 28 kDa. Isolation of the HSM3 67 kDa major peak, followed overnight equilibration and repeated SEC, also demonstrated a redistribution into the characteristic heterogenous profile observed in the initial chromatogram. Analysis of HSM1, 2, and 3 SEC fractions on SDS PAGE confirm that all the peaks having different apparent masses on SEC resolve with the same apparent 16 kDa mass on SDS PAGE.
FIGURE 2.

Size exclusion chromatography (SEC) of HSM1, 2, and 3 proteins in phosphate buffer. The chromatograms of individual HSM1, 2, and 3 proteins resolved on Superdex 200 SEC are provided; also shown are mass standards (see text for details) and the indicated mass is kDa
The left half (LH) HS cassette HSM1 mutant protein resolved on SEC with an essentially identical elution volume to that of the HSM1 mutant protein (Figure 3), suggesting the same molecular assembly. The right half (RH) cassette HSM1 mutant protein, however, resolved essentially identical to that of the Symfoil‐4T protein, indicating a monomer form and loss of oligomerization. In the case of the quarter cassette mutations, only the left quarter 2 (LQ2) mutant resolved on SEC similar to the HSM1 mutant protein, while the left quarter 1 (LQ1), right quarter 1 (RQ1), and right quarter 2 (RQ2) mutants resolved essentially identical to the Symfoil‐4T protein as monomeric forms.
FIGURE 3.

Size exclusion chromatography (SEC) chromatograms of half and quarter HSM1 proteins. Mutant proteins were resolved on Superdex 200 SEC in phosphate buffer. HSM1 and Symfoil‐4T reference chromatograms are also shown. Mass standards are omitted for clarity, but are identical to those provided in Figure 2
2.2. Analytical ultracentrifugation of HSM proteins
The G(s) distributions derived from analytical ultracentrifugation (AUC) data for the Symfoil‐4T, HSM1, 2, and 3, LH and RH mutant proteins are provided in Figure 4. The G(s) distribution profile of Symfoil‐4T is consistent with previous crystallographic and biophysical data confirming it as a monomer. 17 , 18 AUC data for HSM1 correlates with a single distribution indicative of one ideal population at a mass ratio of 3:1 in comparison to Symfoil‐4T. AUC data for HSM2 indicates a mass distribution consistent with a dominant higher order mass species essentially trimeric in relation to Symfoil‐4T. AUC data for HSM3 is consistent with an equilibrium distribution of various sized species ranging from monomer, through trimer, and higher‐order oligomeric species. The LH mutant G(s) distribution profile is similar to that of HSM1, while the RH mutant G(s) distribution profile is similar to Symfoil‐4T.
FIGURE 4.

Analytical ultracentrifugation (AUC) data and derived sedimentation coefficients for Symfoil‐4T and heparin sulfate mutants (HSM) proteins. AUC analysis of HSM proteins was performed in phosphate buffer pH 7.0, 293 K, and with protein concentrations of 10–16 μM
2.3. Isothermal equilibrium denaturation of HSM proteins
Symfoil‐4T exhibits cooperative unfolding in GuHCl with a midpoint of denaturation of 2.60 M in phosphate buffer (in good agreement with previously published data collected at pH 6.6 17 , 18 ). All HSM proteins exhibit cooperative unfolding, albeit with substantial destabilization compared to Symfoil‐4T (Figure 5). HSM1 and HSM2 proteins exhibit a clear native state baseline and a similar magnitude of destabilization; in contrast, HSM3 has an incomplete pre‐transition state baseline and appears partially unfolded in 0 M GuHCl (i.e., in phosphate buffer). Since the SEC and AUC data are consistent with trimeric assembly, the isothermal equilibrium denaturation (IED) data were fit with a trimer model (we note the heterogeneous nature of the HSM2 and HSM3 proteins, but fit all data to a trimer model for comparative purposes). The fitted parameters for the HSM1, HSM2, and HSM3 IED data are provided in Table 1; this table also includes analysis of the Symfoil‐4T IED data in phosphate buffer. All half and quarter HS cassette mutants exhibited cooperative folding with a clear pre‐transition baseline (Figure 6). Based upon the SEC and AUC results, the HSM1, LH, and LQ2 mutants were analyzed using a trimer two‐state model, while the RH, LQ1, RQ1, and RQ2 mutants were analyzed using a monomer two‐state model (Table 1).
FIGURE 5.

Isothermal equilibrium denaturation (IED) analysis of HSM1, 2, and 3 mutants and Symfoil‐4T. IED analysis of heparin sulfate mutants proteins was performed in phosphate buffer pH 7.0, 298 K, and with protein concentrations of 5.0 μM
TABLE 1.
IED data for HSM proteins and Symfoil‐4T in phosphate buffer at 298 K
| Protein | IED model | Co (μM) a | ΔG0 (kJ/mol) | m‐Value (kJ mol−1M−1) | C m b (M) | ΔΔG c (kJ/mol) |
|---|---|---|---|---|---|---|
| Symfoil‐4T | Monomer | 5.00 | 44.1 ± 2.5 | 17.1 ± 0.2 | 2.60 | ‐ |
| HSM1 | Trimer | 1.67 | 80.3 ± 1.1 | 19.5 ± 0.9 | 1.20 | 25.6 |
| HSM2 | Trimer | 1.67 | 80.7 ± 1.1 | 22.1 ± 0.9 | 1.10 | 29.4 |
| HSM3 | Trimer | 1.67 | 76.2 ± 0.5 | 30.0 ± 1.6 | 0.60 | 47.1 |
| LH | Trimer | 1.67 | 33.1 ± 1.5 | 23.4 ± 1.4 | 1.20 | 28.4 |
| RH | Monomer | 5.0 | 29.9 ± 2.7 | 17.5 ± 1.6 | 1.70 | 15.6 |
| LQ1 | Monomer | 5.0 | 27.7 ± 1.9 | 19.1 ± 1.3 | 1.45 | 20.8 |
| LQ2 | Trimer | 1.67 | 22.7 ± 4.6 | 32.7 ± 4.0 | 1.15 | 36.1 |
| RQ1 | Monomer | 5.0 | 28.4 ± 1.3 | 19.4 ± 0.9 | 1.45 | 21.0 |
| RQ2 | Monomer | 5.0 | 42.3 ± 3.3 | 17.4 ± 1.4 | 2.55 | 0.9 |
Abbreviations: HSM, heparin sulfate mutants; IED, isothermal equilibrium denaturation; LH, left half; LQ, left quarter; RH, right half; RQ, right quarter.
Monomer concentration.
[GuHCl] X U = 0.5.
(Avg m‐value) × (C m Symfoil‐4T − C m Mutant). A positive value indicates mutant is destabilized.
FIGURE 6.

Isothermal equilibrium denaturation (IED) of half and quarter cassette HSM1 proteins. IED analysis of heparin sulfate mutants proteins was performed in phosphate buffer pH 7.0, 298 K, and with protein concentrations of 5.0 μM
2.4. Differential scanning calorimetry of HSM1 protein
Symfoil‐4T and the HSM1 protein exhibited a temperature‐dependent phase transition having an excess heat capacity (Figure S2). Both proteins exhibited a similar heat capacity for the folded (N) and unfolded (D) states (i.e., CpN and CpD, respectively) and a positive value for ΔCp (attributable to exposure of a hydrophobic core during unfolding 32 ). The Symfoil‐4T differential scanning calorimetry (DSC) data were fit to a monomer model, while the HSM1 data were fit to a trimer model. Compared to Symfoil‐4T, the HSM1 mutant protein exhibits ~25% reduction in folding enthalpy, and a 13 K reduction in T m (Table 2).
TABLE 2.
DSC parameters for Symfoil‐4T and HSM1 in phosphate buffer
| Protein | Model | T m (K) a | ∆H (kJ/mol) | ∆Cp (kJ mol−1 K−1) |
|---|---|---|---|---|
| Symfoil‐4T | Monomer | 346.6 ± 0.1 | 501 ± 3.0 | 8.5 ± 0.7 |
| HSM1 | Trimer | 333.1 ± 0.1 | 371 ± 2.3 | 6.7 ± 0.1 |
Abbreviations: DSC, differential scanning calorimetry; HSM, heparin sulfate mutants.
X U = 0.5.
2.5. Heparin Sepharose affinity chromatography of HSM proteins
The Symfoil‐4T protein eluted in the flow through on analytical heparin Sepharose chromatography with no apparent affinity. The HSM1 protein eluted as a single peak at a [NaCl] of 0.73 M. The HSM2 protein eluted as a single peak, albeit with a leading‐edge shoulder, at a [NaCl] of 0.70 M. The HSM3 protein exhibited two apparent elution peaks, at a [NaCl] of 0.53 and 0.98 M, respectively. The isolated HS cassette 24‐mer peptide eluted as a single peak at a [NaCl] of 0.59 M. Fibroblast growth factor‐1 (FGF‐1) was included as a control, and eluted at a [NaCl] of 1.38 M. The LH mutant eluted as a single peak at a [NaCl] of 0.71 M, while the LQ2 mutant eluted as a single peak at a [NaCl] of 0.57 M (Figure 7, Table 3). The LQ1, RH, RQ1 and RQ2 cassette mutants exhibited no apparent HS affinity and were present in the column loading/wash flow‐through.
FIGURE 7.

Analytical heparin Sepharose chromatography of heparin sulfate mutants proteins and HS peptide cassette. Upper panel: HSM1, 2, and 3 proteins. The chromatogram covers the [NaCl] gradient subsequent to loading and washing with phosphate buffer. The Symfoil‐4T protein does not bind and elutes in the column wash. The [NaCl] gradient used to elute bound proteins is indicated by the dotted line. Also included with the set of HS mutants is the 24‐mer HS cassette peptide, and the fibroblast growth factor‐1 (FGF‐1) protein. Lower panel: Half and quarter HS cassette mutant proteins. Only the LH cassette and LQ2 cassette mutants bind to Heparin Sepharose; the RH, LQ1, RQ1, and RQ2 mutants are present in the column load flow through. Also included for reference in this figure are the elution profiles of the 24‐mer HS cassette peptide and the HSM1 protein
TABLE 3.
NaCl concentration required for elution of HS mutant proteins from heparin Sepharose affinity resin
| Protein | [NaCl] (M) |
|---|---|
| Symfoil‐4T | N/A a |
| HSM1 | 0.73 |
| HSM2 | 0.70 |
| HSM3 | 0.53, 0.98 |
| LH | 0.71 |
| RH | N/A a |
| LQ1 | N/A a |
| LQ2 | 0.57 |
| RQ1 | N/A a |
| RQ2 | N/A a |
| HS cassette peptide | 0.59 |
| FGF‐1 | 1.38 |
Abbreviations: FGF, fibroblast growth factor; HSM, heparin sulfate mutants; LH, left half; LQ, left quarter; RH, right half; RQ, right quarter.
Present in flow‐through (no affinity).
3. DISCUSSION
The HSM1, 2, and 3 proteins exhibit SEC profiles having an unexpectedly large apparent molecular mass, suggesting the possibility of either an oligomeric state or an unfolded protein (SEC of Symfoil‐4T unfolded and resolved in 4.5 M GuHCl running buffer exhibits an apparent mass of 98 kDa); thus, an initial question was whether these mutant proteins were actually folded. The IED data for these mutants show a cooperative transition of fluorescence signal in response to increasing denaturant concentration, a characteristic of folded proteins. The AUC data are consistent with a trimer oligomeric assembly (or major trimer component) for the HSM1, 2, and 3 proteins. DSC provides a direct measurement of unfolding enthalpy, and the DSC data show that HSM1 (the most homogenous mutant by SEC) exhibits excess enthalpy associated with a temperature‐induced phase transition; furthermore, the endotherm exhibits a positive ΔCp value, with native and denatured state baseline functions essentially identical to Symfoil‐4T. Overall, these data indicate that the HSM proteins are cooperatively folded oligomeric assemblies most consistent with a trimeric quaternary structure. Under the conditions and protein concentrations utilized, the SEC of HSM2 and HSM3 mutants demonstrate a minor population of apparent monomer. In this regard, the HSM3 mutant is notably heterogenous, also exhibiting evidence of an apparent oligomer greater than trimeric. Structural heterogeneity of HSM3 is supported by the AUC data, and the IED data show evidence of a minor unfolded population even in the absence of denaturant. Overall, however, the data indicate the HSM1, HSM2, and HSM3 mutant proteins are principally cooperatively folding trimeric assemblies.
The Symfoil‐4T protein has no heparin Sepharose affinity, while the FGF‐1 protein (a heparin‐binding growth factor 33 , 34 ) binds heparin Sepharose and requires 1.38 M [NaCl] for elution. As an isolated peptide, the designed 24‐mer peptide binds to heparin Sepharose and elutes at 0.59 M [NaCl], indicating that the unstructured peptide has an intrinsic electrostatic affinity for polyanions due to the seven basic amino acids (i.e., Lys and Arg). HSM1 and HSM2 proteins both elute from heparin Sepharose at a higher 0.73–0.74 M [NaCl]. These data indicate that structuring of the 24‐mer peptide by incorporation into the Symfoil‐4T scaffold enhances HS‐binding affinity. Elution of the HSM3 mutant from heparin Sepharose suggests an affinity associated both with an unstructured HS‐binding peptide region ([NaCl] of 0.53 M), but also an affinity requiring [NaCl] of 0.98 M for elution. HSM3 shows evidence of higher‐order oligomers in SEC, and it is possible that this apparent higher affinity is due to polyvalent HS binding present in such oligomers. Overall, the data support successful, structure‐based, HS‐binding functionalization of the functionless Symfoil‐4T scaffold. The HS‐affinity of the HSM proteins is intermediate between the unstructured 24‐mer peptide, and the naturally‐evolved heparin‐binding protein FGF‐1. The HS‐binding functionalization of the Symfoil‐4T scaffold can therefore be described as rudimentary. We note that this heparin Sepharose chromatography data provide ancillary supporting evidence of folding (i.e., formation of structure compatible with enhance heparin binding affinity compared to the unstructured 24‐mer peptide cassette).
Characterization of the HS‐binding affinity of the half‐ and quarter‐HS peptide mutants also helps identify the minimum requirements of a rudimentary HS‐binding functionality. The RH mutant exhibits no HS affinity, while the LH mutant exhibits an affinity indistinguishable from the intact 24‐mer; thus, the LH region is the primary determinant of the 24‐mer HS‐binding affinity. Of the seven basic amino acids in the 24‐mer peptide, five are located in the LH and two in the RH region (Figure 8). The LQ1 region exhibits no HS‐affinity, while the LQ2 region binds heparin Sepharose and elutes with 0.57 M [NaCl]. The LQ1 region contains two basic residues, while the LQ2 region contains three. Thus, there may be a direct correlation between HS‐affinity and net basic charge. The LQ2 mutant is the most destabilizing of the quarter peptide mutants, although it is folded in phosphate buffer (Figure 6, Table 1). Basic amino acid mutations (possibly as few as three) in the T3/T7/T11 turn region may therefore be sufficient to generate rudimentary heparin‐binding functionality in the β‐trefoil (with subsequent mutations optimizing affinity and stability).
FIGURE 8.

Design of a heparin sulfate (HS)‐binding 24‐mer peptide “cassette” derived from bovine and human FGF‐1 and human FGF‐2. The sequences of human/bovine FGF‐1 and human FGF‐2 in the region 108–131 (using the numbering scheme of the 140 amino acid form of human FGF‐1) which form the majority of the HS ligand binding interactions are shown color coded by protein. Boxed positions indicate conserved amino acid positions between all three proteins. Blue positions in the designed HS binding cassette indicate conserved hydrophobic core residues in the Symfoil‐4T scaffold protein, and were therefore retained. The magenta Ala117 positions is an optimized mutation designed to eliminate the reactive Cys thiol at this position. Half‐ and quarter‐cassette designs are also indicated. The origin of residues in the HS binding cassette based upon a consensus or composite of sequences is indicated by color code
The accommodation of the 24‐mer HS‐binding functional cassette, as represented by the HSM1, 2, and 3 proteins, is associated with significant destabilization of the Symfoil‐4T scaffold; however, the destabilization is not identical despite the three structurally equivalent sites of insertion. While the HSM1 and HSM2 proteins are destabilized by a similar 25.6 and 29.4 kJ/mol, respectively, the HSM3 protein is destabilized by a significantly greater 47.1 kJ/mol. The site of HSM3 functional insertion is in the centrally located trefoil motif repeat (Figure 9). The repeating trefoil motif in Symfoil‐4T, expressed as an isolated polypeptide (i.e., the “Monofoil” 42‐mer), is an autonomously folding motif, and therefore contains an effective folding nucleus. 17 , 18 , 35 However, as a concatenated trimer, the folding nucleus of Symfoil‐4T is not distributed within each of the repeating trefoil motifs, rather, it spans the region from Turn IV through Turn IX (i.e., the central trefoil motif, plus preceding and following β‐hairpin turns). 30 A recent mutational study, disrupting a conserved buried water within each of the three repeating trefoil motifs, showed that β‐hairpin structures preceding and following the central motif provide additional folding nucleation structure. 36 Such hairpins are not present in the first and third motif due to the structural “discontinuity” created by the N‐ and C‐termini. The HSM1 and HSM2 proteins do not disrupt the preferred centrally‐located folding nucleus, but the HSM3 mutation does. However, while significantly destabilized, the HSM3 mutant nonetheless is substantially folded, indicating that the first or third motif are providing a serviceable (albeit less efficient) folding nucleus. Thus, the Symfoil‐4T symmetric protein architecture is able to provide alternative folding nuclei in response to deleterious mutation; however, such tolerance appears modulated by fused termini effects of adjacent motifs which can augment a specific folding nucleus region. In the threefold symmetric β‐trefoil architecture it is only the central motif (and not the N‐ and C‐terminus motifs) that benefits from novel hairpin structures created by termini fusion of the repeating motifs. In symmetric protein architectures with higher rotational symmetry (e.g., C4–C8) multiple instances of optimized folding nuclei may therefore exist; such multiple instances are not available with C3 symmetry (as is present in the β‐trefoil).
FIGURE 9.

24‐mer heparin sulfate (HS)‐binding “cassette” and sites of insertion in the Symfoil‐4T β‐trefoil “scaffold.” The designed HS‐binding cassette 24‐mer peptide is indicated in the shaded red box. The equivalent structural location in FGF‐1 is indicated in top panel (“Heparin binding mutant #1”). The center and lower panel indicate two equivalent positions related by the threefold rotational symmetry intrinsic to the Symfoil‐4T protein, generating the HSM2 and HSM3 mutants, respectively
A notable feature of the HSM proteins is their trimeric quaternary structure. Several previous mutations of Symfoil‐4T (or closely related forms) have also yielded oligomer assemblies. As mentioned above, expression of the 42‐mer “Monofoil” trefoil motif spontaneously assembles as a homotrimer, generating an intact β‐trefoil architecture. A tandem repeat of Monofoil yields the “Difoil” protein, which also spontaneously assembles as a homotrimer, generating two complete β‐trefoil architectures. In the case of Difoil, two polypeptides each fold to yield two‐thirds of a β‐trefoil, while the remaining motif adopts a unique “domain‐swapped” 37 structure that effectively completes both of these partial β‐trefoils. In this latter motif, the domain‐swapping occurs due to a structural divergence within the T11 extended loop region, near residue position 119 (see Figure 10). In yet another example, a Symfoil‐4T mutant where the majority of the central trefoil motif was substituted by a “blade” motif from a β‐propeller protein, a trimeric quaternary structure was also formed. 31 Thus, common themes resulting in oligomerization are mutations that destabilize the central folding nucleus or the T3, T7, T11 region(s).
FIGURE 10.
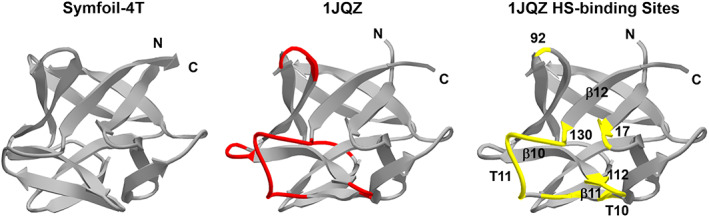
Structurally heterogenous surface loops and heparin sulfate (HS)‐binding positions in FGF‐1/FGF‐2. The structurally heterogenous surface loops in FGF‐1 (1JQZ) compared to Symfoil‐4T are indicated in red (center image). The positions associated with HS‐binding interactions in human/bovine FGF‐1 and FGF‐2 (Table S1) are shown in yellow (right image). Most of the surface loops unique to FGF‐1/FGF‐2 compared to the fundamental β‐trefoil architecture represented by Symfoil‐4T are associated with HS‐binding functionality
When the separate 12‐mer LH and RH mutations are expressed, it is only the LH mutant that oligomerizes, while the RH is monomeric. The LH mutant is almost twice as destabilizing as the RH mutant (28.4 kJ/mol vs. 15.6 kJ/mol, respectively). When the 6‐mer LQ1 and LQ2 mutations are expressed (i.e., splitting the LH mutant) it is only the LQ2 mutant that oligomerizes, while the LQ1 is monomeric. The LQ2 is substantially more destabilizing than the LQ1 mutant (36.1 kJ/mol vs. 20.8 kJ/mol, respectively). The RQ1 and RQ2 mutants are both monomeric and destabilize by 21.0 and 0.9 kJ/mol, respectively. Thus, the energetics of destabilization, and tendency to oligomerize, appear directly related, with the key monomer to trimer transition occurring between 21.0 and 25.6 kJ/mol destabilization (Figure 11). With a ΔG0 of 44.1 kJ/mol, the Symfoil‐4T should be able to tolerate a destabilization of 21.0–25.6 kJ/mol and still fold as a monomer. The formation of trimeric quaternary structure therefore indicates that an alternative folding pathway of oligomerization is accessible to the Symfoil‐4T protein, and the net balance of energetics associated with the entropic penalty and enthalpic gain for trimeric assembly appears to be overall favorable upon a monomeric destabilization of 21.0–25.6 kJ/mol. Is the monomer folding pathway still accessible to the HSM proteins? To address this question, the HSM1 and HSM2 proteins were subject to analytical SEC (Superdex 75 in phosphate buffer) with dilute protein concentrations varying between 5, 1.6, and 0.5 μM. The chromatograms indicated that there is a corresponding diminution of the high mass trimer peak (55–60 kDa on Superdex 75), and an increasing appearance of a low mass monomer peak (14.5–14.9 kDa on Superdex 75), with protein dilution (Figure S3). These results suggest that the monomer folding pathway is still accessible, and is utilized when oligomerization is disfavored under conditions of dilute protein concentration. From a different viewpoint, it is apparent that the available folding pathway(s) of the monomer β‐trefoil includes an intrinsic potential for trimeric assembly. The accessibility of such alternative folding pathways involving oligomerization may be a survival advantage intrinsic to symmetric protein architecture, providing alternative practical means to fold despite diverse deleterious mutation. We note that oligomers are reportedly more common in nature than monomers, 3 and so such folding adaptability may be a common feature of extant protein folds (especially symmetric folds).
FIGURE 11.
Relationship between oligomeric state and stability for heparin sulfate (HS) mutants. A transition from monomeric to trimeric oligomerization for the set of HS mutants occurs upon a destabilization greater than ~25 kJ/mol
4. MATERIALS AND METHODS
4.1. Design of a HS binding peptide “cassette”
HS binding functionality is shared by the majority of the FGF family of proteins. 38 A total of 10 X‐ray and NMR structures in the RCSB structural databank of human FGF‐1, FGF‐2, and bovine FGF‐1 with bound HS (or analogue) were queried to identify all protein contacts within 3.4 Å. This set of HS/analog ligands includes heparin tetramer, pentamer and hexamer, sucrose octasulfate (SOS), 5‐amino‐2‐naphthalenesulfonate, 1,3,6‐naphthlalenetrisulfonate, inositol hexaphosphate, dobelisate, and disaccharide S312 (2‐deoxy‐3‐O‐sulfo‐2‐(sulfoamino)‐alpha‐D‐glucopyranose‐(1–4)‐1‐O‐methyl‐2‐O‐sulfo‐alpha‐L‐idopyranuronic acid). A compilation of the FGF‐1 and FGF‐2 residue positions interacting with HS and analogues is provided in Table S1. HS binding sites in FGF‐1/FGF‐2 include residues 17,18, 92, and an essentially contiguous region spanning residues 112–130 (referencing the 140 amino acid form of FGF‐1). Residues 17 and 18 are located in turn 1 (T1), residue 92 is located in T8, and residues 112–130 correspond to the C‐terminal of β‐strand 10 (β10) and continuing through T10, β11, and T11 regions (Figure 10). The human and bovine FGF‐1 and human FGF‐2 sequences spanning positions 112–130 contain a total of seven basic residues which form various salt‐bridges and charged H‐bonds with sulfonate groups of bound HS/analogue. Additionally, previous studies have reported that an Arg119Gly mutation combined with deletion of residue positions 120–122 (including Arg122) in FGF‐1 can effectively abolish HS affinity. 39 These data suggest that the region 112–130 in FGF‐1/FGF‐2 likely contributes a majority of the binding enthalpy with HS, and is therefore likely sufficient to confer at least a rudimentary HS binding functionality if inserted into the appropriate location in a β‐trefoil “scaffold” protein. We note that Kumar and coworkers increased the heparin affinity of FGF‐1 by introducing an Asp68Arg mutation in the adjacent T6 turn region, thereby extending the HS‐binding patch on the surface of FGF‐1. 40 However, in the present case, we focus upon a short contiguous region that might confer HS‐affinity.
Design of a peptide “cassette” conferring HS binding functionality was initially pursued focusing upon the region of FGF‐1 primary structure spanning residues 112–130 with the following modifications: (a) An Arg residue was substituted for Lys at position 113. The Arg guanidino group provides for additional H‐bonding possibilities compared to Lys, and is present at the equivalent position in FGF‐2. (b) An Arg residue was substituted for Ser at position 116. The Arg side chain provides for potential charge interactions with HS sulfonates, is present at this position in bovine FGF‐1, and has been shown experimentally to increase the HS affinity of human FGF‐1. 41 (c) An Ala residue was substituted for Cys at position 117. This Cys is a free reactive thiol that can cause unwanted dimer formation, and an Ala substitution has been reported as the most stable substitution at this position. 42 These substitutions were modeled into the 19‐mer region of residues 112–130 of the FGF‐1 crystal structure (RCSB accession 1JQZ) to generate a putative HS‐binding peptide cassette. This modified FGF‐1 structure was then overlaid onto the symmetric designed β‐trefoil protein Symfoil‐4T (RCSB accession 3O4B) using the following regions of backbone atoms: 12–16, 19–47, 54–67, 70–76, 81–90, 94–102, 107–112, 129–136, yielding a 0.63 Å rmsd for the overlaid atoms. Symfoil‐4T was selected as an appropriate β‐trefoil scaffold due to the following properties: (a) it is a purely‐symmetric β‐trefoil whose threefold rotational symmetry provides a potential for redundant folding nuclei and three equivalent positions for mutation (i.e., functional insertion); (b) It is a thermophilic protein whose enhanced thermostability should permit potentially deleterious functional mutation while retaining foldability; (c) It is a “functionally benign” protein scaffold devoid of any known function and adoption of novel functionality should therefore be readily detectable. The Symfoil‐4T structure was then analyzed for any close contacts (i.e., less than van der Waals) involving the 112–130 region of the substituted HS‐binding peptide. Nine positions within the 19‐mer HS‐binding peptide exhibited close contacts with the Symfoil‐4T host structure. Three of these close contacts involved Arg110 in Symfoil‐4T. Position 110 in FGF‐1 is Gly; thus, including mutation Arg110Gly in Symfoil‐4T potentially eliminates three close contacts. A close contact involving Tyr108 in Symfoil‐4T is eliminated if this residue is mutated to Phe as in FGF‐1. Another close contact is eliminated if Gln131 in Symfoil‐4T is mutated to Leu as in FGF‐1. Thus, five of the nine identified close contacts would potentially be eliminated if the 19‐mer HS‐binding peptide was extended to a 24‐mer comprising residue positions 108–131 and with the additional five positions adopting the FGF‐1 sequence. Of additional concern, residues Leu109 and Ile111 form part of the symmetric core packing group in the Symfoil‐4T host protein and were therefore retained. The overall design of the resulting 24‐mer putative HS‐binding cassette peptide is provided in Figure 8.
4.2. Mutant construction, expression, and purification
Introduction of the HS binding cassette at the structurally equivalent 108–131 position in a β‐trefoil protein can potentially confer HS binding functionality, yielding an HS mutant (“HSM”). The Symfoil‐4T protein is a purely‐symmetric, thermophile, designed β‐trefoil protein that is devoid of any specific functionality, 17 , 18 and was selected as an appropriate “scaffold” for functional mutation. The 24‐mer HS‐binding peptide substitution involves an insertion of three amino acids, a deletion of one amino acid, and a substitution of 17 amino acids, compared to the corresponding region in Symfoil‐4T. Residue positions 108–131 in Symfoil‐4T (i.e., spanning the T10‐β11‐T11 region) are located within the third repeating “trefoil” motif, and is the native location of HS ligand binding (referencing FGF‐1 and FGF‐2 structures). Due to the threefold (C3) rotational symmetry of primary structure present in Symfoil‐4T, two other structurally equivalent locations are possible, including residue positions 22–43 (i.e., the T2‐β3‐T3 region in the first trefoil motif), and residue positions 64–84 (i.e., the T6‐β7‐T7 region in the second trefoil motif). Insertion of the HS‐binding peptide at positions 108–131 yields the HSM1 mutant, insertion at positions 22–43 yields the HSM2 mutant, and insertion at positions 64–84 yields the HSM3 mutant (Figure 9). In addition to this 24‐mer, two 12‐mer half‐cassettes, and four 6‐mer quarter‐cassette mutants were also constructed at the 108–131 region (i.e., using the HSM1 mutant location in the third trefoil motif). All mutant expression vectors were constructed by gene synthesis (GenScript Inc., Piscataway, NJ) and insertion into the pet21a(+) expression vector (and included an N‐terminal 6x His‐tag). Sequences were confirmed by nucleic acid sequence analysis (Biomolecular Analysis Synthesis and Sequencing Laboratory, Florida State University). The HS binding cassette was also synthesized as a 24‐mer peptide (GenScript, Piscataway, NJ).
Protein expression followed previously described procedures 43 utilizing the BL21 (DE3) E. coli expression host. Briefly, initial chromatographic purification of expressed proteins was performed using Ni‐NTA affinity resin (GE healthcare, Marlborough, MA). Cleared cell lysate was loaded directly on a 2.5 × 2.5 cm column, washed with 50 mM sodium phosphate, 500 mM NaCl and eluted using a step‐wise gradient of 5, 25, 50, 100, and 200 mM imidazole in the same buffer. Elution fractions containing recombinant protein were pooled. The Ni‐NTA elution pool was subsequently loaded directly onto a Superdex 200 26/60 size exclusion column (GE Life Sciences, Marlborough, MA) equilibrated to 50 mM Na2PO4, 100 mM NaCl, 10 mM (NH4)2SO4 pH 7.5 (“phosphate buffer”). Fractions containing protein were pooled based upon the A280 chromatogram and corresponding Coomassie brilliant blue stained 15% SDS‐PAGE of individual fractions.
4.3. Analytical SEC
Analytical SEC was performed using Superdex 200 resin in a 26/60 column for all proteins except dilution experiments with HSM1 protein which utilized Superdex 75 resin (to better separate potential subunits). All proteins were run in phosphate buffer and with nominal loading volume of 10 ml and concentrations of 10 μM.
4.4. Determination of heparin affinity of HSM proteins
Heparin affinity (i.e., HS‐binding functionality) of the HSM proteins was evaluated by chromatography on Heparin Sepharose CL‐6B resin (GE Life Sciences, Marlborough, MA). Purified HSM proteins, as well as Symfoil‐4T and FGF‐1 control proteins, were dialyzed (2× 1:100 vol:vol) in phosphate buffer. Proteins were diluted to 0.25–0.50 mg/ml and a 2 ml volume was loaded onto a 1.0 cm × 10 cm analytical column (~8 ml column volume). The column was subsequently washed with two column volumes of loading buffer and the flow through was evaluated for any proteins lacking heparin affinity. Bound proteins were eluted with a 0–2.0 M NaCl gradient in loading buffer over 18 column volumes. Fractions containing HSM protein were pooled based upon the A280 chromatogram and corresponding SDS‐PAGE of individual fractions, and the NaCl concentration required for elution (i.e., elution peak maximum) determined. The 24‐mer peptide was dissolved in phosphate buffer to a concentration of 0.10 mg/ml and a 2 ml volume was similarly chromatographed; however, in this case the A254 was monitored.
4.5. Isothermal equilibrium denaturation
Fluorescence data associated with protein unfolding under isothermal conditions were collected as previously described. 35 , 44 Briefly, 5 μM protein samples were equilibrated by dialysis at 298 K for 24 h with 0.0–4.5 M guanidinium hydrochloride (GuHCl) in increments of 0.05 M GuHCl in phosphate buffer. The Symfoil‐4T protein contains three buried Phe residues useful as a spectroscopic probe of unfolding. 17 , 18 The Phe fluorophores were excited using λ ex = 277 nm and λ em was recorded between 284 to 410 nm using a Cary Eclipse fluorescence spectrophotometer (Agilent Technology, Santa Clara, CA). Samples were scanned in triplicate and averaged, buffer scans were subtracted, and the resulting spectra were integrated. Analysis of IED data utilized either a monomer two‐state unfolding model, or a trimer two‐state unfolding model as previously described. 31 Nonlinear least squares fitting of IED data were performed using the DataFit software package (Oakdale Engineering, Oakdale, PA).
4.6. Analytical ultracentrifugation
AUC velocity data were collected in a Beckman XL‐I centrifuge (Beckman Coulter, Indianapolis, IN) using absorbance optics with a standard two‐channel Epon centerpiece and an AN60 Ti rotor at 58,000 rpm. Intensity scans were recorded at 280 nm and 293 K. Protein samples were run at an optical density A280 between 0.50 and 0.80 (0.010–0.016 mM) in phosphate buffer. Data were analyzed using the Ultra Scan III version 4.0 software suite. 45 A time derivative 2‐D spectrum analysis was performed on all data with removal of time invariant noise. Data were processed using the Alamo E cluster at the University of Texas Health Science Center at San Antonio and the Texas Advanced Computing Center at the University of Texas in Austin. A Monte Carlo genetic algorithm analysis followed by an enhanced van Holde–Weischet analysis was applied to the data. The G(s) plots were normalized to the average sedimentation coefficient of the Symfoil‐4T protein.
4.7. Differential scanning calorimetry
All calorimetric data were collected on a Nano‐Differential Scanning Calorimeter (TA Instruments, New Castle, DE) pressurized to 40 psi with nitrogen gas and all samples were degassed for 15 min prior to loading. Protein endotherms were collected from 290 to 370 K at a scan rate of 0.25 K min−1. 46 Data were averaged and reference buffer/buffer scans were subtracted, and the resulting scans normalized to molar concentration. Analysis of DSC endotherms utilized either a monomer two‐state unfolding model, 47 or a trimer two‐state unfolding model as previously described. 31
CONFLICT OF INTEREST
Michael Blaber is a cofounder and has equity ownership in Trefoil Therapeutics Inc.
AUTHOR CONTRIBUTIONS
Connie A. Tenorio: Formal analysis (equal); investigation (equal); methodology (equal); writing – original draft (equal). Joseph B. Parker: Investigation (equal). Michael Blaber: Conceptualization (lead); formal analysis (equal); project administration (lead); supervision (lead); writing – original draft (equal); writing – review and editing (lead).
Supporting information
Appendix S1: Supporting Information
ACKNOWLEDGMENTS
This work was supported in part by a research support agreement from Trefoil Therapeutics Inc. Support from the FSU department of Biomedical Sciences for Connie A. Tenorio is acknowledged. Joseph B. Parker was supported by a summer research fellowship by the FSU College of Medicine.
Tenorio CA, Parker JB, Blaber M. Functionalization of a symmetric protein scaffold: Redundant folding nuclei and alternative oligomeric folding pathways. Protein Science. 2022;31(5):e4301. 10.1002/pro.4301
Review Editor: Aitziber Cortajarena
Funding information Trefoil Therapeutics, Grant/Award Number: RF02251
REFERENCES
- 1. Sawyer N, Speltz EB, Regan L. NextGen protein design. Biochem Soc Trans. 2013;41:1131–1136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Chevalier A, Silva D‐A, Rocklin GJ, et al. Massively parallel de novo protein design for targeted therapeutics. Nature. 2017;550:74–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Gwyther REA, Jones DD, Worthy HL. Better together: Building protein oligomers naturally and by design. Biochem Soc Trans. 2019;47:1773–1780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Zhou W, Šmidlehner T, Jerala R. Synthetic biology principles for the design of protein with novel structures and functions. FEBS Lett. 2020;594:2199–2212. [DOI] [PubMed] [Google Scholar]
- 5. Zhang Y, Chen Y, Wang C, et al. ProDCoNN: Protein design using a convolutional neural network. Proteins. 2020;88:819–829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Surpeta B, Sequeiros‐Borja CE, Brezovsky J. Dynamics, a powerful component of current and future in silico approaches for protein design and engineering. Int J Mol Sci. 2020;21:2713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Beadle BM, Shoichet BK. Structural basis of stability ‐ function tradeoffs in enzymes. J Mol Biol. 2002;321:285–296. [DOI] [PubMed] [Google Scholar]
- 8. Tokuriki N, Stricher F, Serrano L, Tawfik DS. How protein stability and new functions trade off. PLoS Comput Biol. 2008;4:e1000002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Rubini M, Lepthien S, Golbik R, Budisa N. Aminotryptophan‐containing barstar: Structure‐function tradeoff in protein design and engineering with an expanded genetic code. Biochim Biophys Acta. 2006;1764:1147–1158. [DOI] [PubMed] [Google Scholar]
- 10. Longo L, Lee J, Blaber M. Experimental support for the foldability‐function tradeoff hypothesis: Segregation of the folding nucleus and functional regions in FGF‐1. Protein Sci. 2012;21:1911–1920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Gosavi S. Understanding the folding‐function tradeoff in proteins. PLoS ONE. 2013;8:e61222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Webster K, Sasso L, Domigan LJ. Adding function to protein scaffolds. In: Gerrard JA, Domigan LJ, editors. Protein nanotechnology: Protocols, instrumentation, and applications. New York, NY: Springer US, 2020; p. 119–147. [DOI] [PubMed] [Google Scholar]
- 13. Guerler A, Wang C, Knapp E‐W. Symmetric structures in the universe of protein folds. J Chem Inform Model. 2009;49:2147–2151. [DOI] [PubMed] [Google Scholar]
- 14. Hocker B, Claren J, Sterner R. Mimicking enzyme evolution by generating new (beta‐alpha)8‐barrels from (beta‐alpha)4‐half‐barrels. Proc Natl Acad Sci U S A. 2004;101:16448–16453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Nikkhah M, Jawad‐Alami Z, Demydchuk M, Ribbons D, Paoli M. Engineering of β‐propellor protein scaffolds by multiple gene duplication and fusion of an idealized WD repeat. Biomol Eng. 2006;23:185–194. [DOI] [PubMed] [Google Scholar]
- 16. Yadid I, Tawfik DS. Functional β‐propeller lectins by tandem duplications of repetitive units. Prot Eng Des Sel. 2011;24:185–195. [DOI] [PubMed] [Google Scholar]
- 17. Lee J, Blaber M. Experimental support for the evolution of symmetric protein architecture from a simple peptide motif. Proc Natl Acad Sci U S A. 2011;108:126–130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Lee J, Blaber SI, Dubey VK, Blaber M. A polypeptide “building block” for the ß‐trefoil fold identified by “top‐down symmetric deconstruction”. J Mol Biol. 2011;407:744–763. [DOI] [PubMed] [Google Scholar]
- 19. Broom A, Doxey AC, Lobsanov YD, et al. Modular evolution and the origins of symmetry: Reconstruction of a three‐fold symmetric globular protein. Structure. 2012;20:1–11. [DOI] [PubMed] [Google Scholar]
- 20. Aksel T, Barrick D. Direct observation of parallel folding pathways revealed using a symmetric repeat protein system. Biophys J. 2014;107:220–232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Voet ARD, Noguchi H, Addy C, et al. Computational design of a self‐assembling symmetrical β‐propeller protein. Proc Natl Acad Sci U S A. 2014;111:15102–15107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Huang P‐S, Feldmeier K, Parmeggiani F, Fernandez Velasco DA, Hocker B, Baker D. De novo design of a four‐fold symmetric TIM‐barrel protein with atomic‐level accuracy. Nat Chem Biol. 2016;12:29–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Terada D, Voet ARD, Noguchi H, et al. Computational design of a symmetrical β‐trefoil lectin with cancer cell binding activity. Sci Rep. 2017;7:5943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Andreeva A, Howorth D, Chothia C, Kulesha E, Murzin AG. SCOP2 prototype: A new approach to protein structure mining. Nucleic Acids Res. 2013;42:D310–D314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Blow DM, Janin J, Sweet RM. Mode of action of soybean trypsin inhibitor (Kunitz) as a model for specific protein‐protein interactions. Nature. 1974;249:54–57. [DOI] [PubMed] [Google Scholar]
- 26. McLachlan AD. Three‐fold structural pattern in the soybean trypsin inhibitor (Kunitz). J Mol Biol. 1979;133:557–563. [DOI] [PubMed] [Google Scholar]
- 27. Murzin AG, Lesk AM, Chothia C. β‐Trefoil fold. Patterns of structure and sequence in the kunitz inhibitors interleukins‐1β and 1α and fibroblast growth factors. J Mol Biol. 1992;223:531–543. [DOI] [PubMed] [Google Scholar]
- 28. Andreeva A, Kulesha E, Gough J, Murzin AG. The SCOP database in 2020: Expanded classification of representative family and superfamily domains of known protein structures. Nucleic Acids Res. 2019;48:D376–D382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Blaber M. Conserved buried water molecules enable the β‐trefoil architecture. Protein Sci. 2020;29:1794–1802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Xia X, Longo LM, Sutherland MA, Blaber M. Evolution of a protein folding nucleus. Protein Sci. 2016;25:1227–1240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Tenorio CA, Parker JB, Blaber M. Oligomerization of a symmetric β‐trefoil protein in response to folding nucleus perturbation. Protein Sci. 2020;29:1629–1640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Myers JK, Pace CN, Scholtz JM. Denaturant m values and heat capacity changes: Relation to changes in accessible surface areas of protein unfolding. Protein Sci. 1995;4:2138–2148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Gospodarowicz D, Cheng J, Lui G‐M, Baird A, Bohlen P. Isolation by heparin‐sepharose affinity chromatography of brain fibroblast growth factor: Identity with pituitary fibroblast growth factor. Proc Natl Acad Sci U S A. 1984;81:6963–6967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Lobb RR, Fett JW. Purification of two distinct growth factors from bovine neural tissue by heparin affinity chromatography. Biochemistry. 1984;23:6295–6299. [DOI] [PubMed] [Google Scholar]
- 35. Tenorio CA, Longo LM, Parker JB, Lee J, Blaber M. Ab initio folding of a trefoil‐fold motif reveals structural similarity with a β‐propeller blade motif. Protein Sci. 2020;29:1172–1185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Parker JB, Tenorio CA, Blaber M. The ubiquitous buried water in the beta‐trefoil architecture contributes to the folding nucleus and ~20% of the folding enthalpy. Protein Sci. 2021;30:2287–2297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Bennett MJ, Schlunegger MP, Eisenberg D. 3D domain swapping: A mechanism for oligomer assembly. Protein Sci. 1995;4:2455–2468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Whittington K, Patrick M, Roberts JL. A national study of pressure ulcer prevalence and incidence in acute care hospitals. J Wound Ostomy Cont Nurs. 2000;27:209–215. [DOI] [PubMed] [Google Scholar]
- 39. Brych SR, Dubey VK, Bienkiewicz E, Lee J, Logan TM, Blaber M. Symmetric primary and tertiary structure mutations within a symmetric superfold: A solution, not a constraint, to achieve a foldable polypeptide. J Mol Biol. 2004;344:769–780. [DOI] [PubMed] [Google Scholar]
- 40. Davis JE, Gundampati RK, Jayanthi S, et al. Effect of extension of the heparin binding pocket on the structure, stability, and cell proliferation activity of the human acidic fibroblast growth factor. Biochem Biophys Rep. 2017;13:45–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Xia X, Kumru OS, Blaber SI, et al. An S116R phosphorylation site mutation in human fibroblast growth factor‐1 differentially affects mitogenic and glucose‐lowering activities. J Pharm Sci. 2016;105:3507–3519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Xia X, Kumru OS, Blaber SI, et al. Engineering a cysteine‐free form of human fibroblast growth factor‐1 for “second generation” therapeutic application. J Pharm Sci. 2016;105:1444–1453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Brych SR, Blaber SI, Logan TM, Blaber M. Structure and stability effects of mutations designed to increase the primary sequence symmetry within the core region of a β‐trefoil. Protein Sci. 2001;10:2587–2599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Longo LM, Lee J, Tenorio CA, Blaber M. Alternative folding nuclei definitions facilitate the evolution of a symmetric protein fold from a smaller peptide motif. Cell Struct. 2013;21:1–9. [DOI] [PubMed] [Google Scholar]
- 45. Demeler B. Methods for the design and analysis of sedimentation velocity and sedimentation equilibrium experiments with proteins. Curr Protoc Protein Sci. 2010;60:7.13.11–17.13.24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Blaber SI, Culajay JF, Khurana A, Blaber M. Reversible thermal denaturation of human FGF‐1 induced by low concentrations of guanidine hydrochloride. Biophys J. 1999;77:470–477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Grek SB, Davis JK, Blaber M. An efficient, flexible‐model program for the analyis of differential scanning calorimetry protein denaturation data. Protein Pept Lett. 2001;8:429–436. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix S1: Supporting Information