

Abstract
The membrane fluidity of reduced-amphotericin B (AmB)-sensitivity Kluyveromyces lactis mutant strain is higher than that of the wild-type K. lactis strain. After culture of the K. lactis and K. lactis mutant cells in the presence of subinhibitory doses of AmB (10 and 125 mg/liter, respectively), the plasma membranes of both yeast strains also showed a higher fluidity than did those of control cells. High membrane fluidity was associated with changes in the structural properties of the membranes. Culture of the K. lactis and K. lactis mutant cells in the presence of AmB induced changes in membrane lipid contents. In particular, phospholipid contents were increased in both strains treated with AmB, compared with their corresponding counterparts. As a result, the sterol/phospholipid ratio decreased. The relative proportion of monounsaturated fatty acids also increased after AmB treatment. The saturated fatty acid/monounsaturated fatty acid ratio decreased in K. lactis and K. lactis mutant cells treated with AmB but also in K. lactis mutant control cells compared to that in the K. lactis wild strain. These changes in lipid composition explain the higher fluidity, which could represent a process of metabolic resistance of the yeasts to AmB.
Antifungal heptaen amphotericin B (AmB) is presently considered the “gold standard” for the intravenous chemotherapy of deep fungal infections in spite of its secondary effects (24). This antibiotic has been a therapeutic agent since 1956, but its mode of action at the molecular level has not been totally elucidated. Various interaction models with membrane sterols have been proposed. However, the most recent models are based on the formation of stable complexes not only with sterols but also with membrane phospholipids whose interactions induce pore formation (1, 6).
AmB was isolated from Streptomyces nodosus cultures by Gold et al. (9). It is a polyene belonging to the macrolide family, and its molecular conformation is closed to phospholipid molecules. This antifungal agent possesses a hydrophobic part (hydrocarbon chain) and a hydrophilic part (polyhydroxyl chain); its amphipathic properties allow interactions with the membrane after diffusion through the yeast cell wall. Its interaction with membrane constituents (14, 16, 18) provokes the formation of pores responsible for several reactions, such as lipid oxidations and peroxidations, inhibition of membrane enzymes (3, 4), osmotic shocks, and, finally, cell death (32).
Yeast resistance to antifungal agents, mainly azol derivatives, is a well-known phenomenon. Yeast cells with reduced sensitivity to AmB have been isolated from patients (24, 29). Horsburgh et al. (13) and Hakkou et al. (11) also isolated a Candida lusitaniae strain and a Kluyveromyces lactis strain, respectively, with reduced sensitivity to AmB. The reduced sensitivity of the strains either may stem from changes in the membrane chemical properties which may result from mutations or may be induced by the antifungal agent itself. The alterations of the membrane may also result in changes in its fluidity and stability.
Following up on earlier studies (11, 27), the aim of this work was to determine the relationship between membrane stability and fluidity in reduced-AmB-sensitivity cells and in cells grown in the presence of subinhibitory doses of AmB, as well as the membrane chemical composition of the K. lactis wild strain and a reduced-AmB-sensitivity K. lactis strain (K. lactis mutant).
The membrane fluidity of spheroplasts from K. lactis mutant cells and K. lactis and K. lactis mutant cells grown in the presence of AmB is significantly higher than the fluidity of spheroplasts from K. lactis cells.
Moreover, besides the well-known alteration in ergosterol induced by AmB, changes occur in phospholipid content and fatty acid profile. These changes could explain the modifications in membrane fluidity.
MATERIALS AND METHODS
Yeast strains and culture conditions.
K. lactis (ATCC 96897) was isolated from dairy products, and the reduced-AmB-sensitivity K. lactis strain (ATCC 96896) was obtained by culture of the wild strain in the presence of subinhibitory doses of AmB (11).
The isolation of K. lactis from human sources is rarely reported, and previous reports of human infection are unavailable in the literature. An immunosuppressed heart transplant patient with pulmonary infection due to the yeast Kluyveromyces fragilis was described by Lutwick et al. (20).
The cells were grown aerobically in Sabouraud liquid medium (2% glucose, 1% Primatone [Sigma]) at 25°C in the absence or presence of subinhibitory doses of AmB (10 and 125 mg/liter for K. lactis and K. lactis mutant, respectively).
AmB (Squibb) was solubilized in a mixture of dimethyl sulfoxide and ethanol (1:1, vol/vol).
The growth of the cells was estimated by measuring the absorbance of the culture at 620 nm on a Perkin-Elmer spectrophotometer. No difference in morphology was observed by optical microscopy.
Spheroplast preparation.
After 10 h of growth, both Kluyveromyces strains were harvested at 4°C by centrifugation at 1,500 × g for 10 min and washed twice with distilled water and then twice with 1.2 M KCl. One gram of cells was suspended in 4 ml of 0.02 M phosphate buffer (25 mM Na2HPO4 · 12H2O, 15 mM KH2PO4) (pH 7.4) with 80 mg of pronase E (Merck) (20 mg/ml) and 0.01 M dithiothreitol (Sigma). It was incubated for 60 min at 30°C with slight stirring.
After centrifugation at 1,500 × g, the cells were washed three times in an isotonic solution (0.15 M Na2HPO4, 0.08 M citric acid, 1.4 M KCl) (pH 5.8) and resuspended in 4 ml of the isotonic solution.
The suspension was then incubated at 30°C for 3 h with 30 mg of “lysing enzyme” from Trichoderma harzianum (Sigma) and 5 mM dithiothreitol (Sigma) with slight stirring. Once centrifuged at 1,500 × g, the spheroplasts were washed twice with 1.2 M KCl and resuspended in the isotonic solution (pH 5.8). All experiments were then performed at a temperature of 30°C.
The kinetics of spheroplast formation was monitored by enumerating the remaining intact yeast cells after dilution of the suspension with distilled water on a Malassez cell with a binocular optic microscope.
To study the influence of the ionic strength of the buffer on spheroplast stability, several dilutions of the isotonic (pH 5.8) and 1.8-ionic-strength solution were achieved. Dilutions of 10, 13, 16, 20, 25, 30, 40, and 50% of the 1.8-ionic-strength solution in distilled water corresponded, respectively, to ionic strengths of 1.6, 1.53, 1.5, 1.4, 1.3, 1.2, 1.0, and 0.8.
To study the influence of incubation time on spheroplast stability, samples were collected at intervals of 0, 20, 40, 60, 80, and 100 min in the isotonic solution (pH 5.8). The number of spheroplasts was determined, for each assay, by the difference between the total number of intact cells and the number of intact cells remaining after spheroplast lysis in a nonosmotic solution.
Spheroplast labeling with a fluorescent probe.
Lipidic fluidity was assessed by steady-state fluorescence polarization using a lipophilic fluorescent probe: 1-[4-(trimethylamino)phenyl]-6-phenyl-1,3,5-hexatriene (TMA-DPH; Molecular Probes, Eugene, Oreg.), which is incorporated into the plasma membrane, remaining anchored at the lipid-water interface with the DPH moiety intercalated between the upper parts of the fatty acyl chains (26).
Stock solutions of TMA-DPH in N-N′-dimethylformamide at a concentration of 2 × 10−3 M were kept in the dark at 4°C.
For fluorescence polarization measurements, 1 μl of probe solution (final probe concentration, 2 × 10−6 M) was added to 1 ml of membrane suspension (spheroplast concentration, 4 × 106 cells per ml) and the labeled suspension was incubated at room temperature in the dark for 5 min.
Fluorescence was measured at 25°C immediately after labeling, without further washing.
Fluorescence polarization measurements.
The fluorescence anisotropy was determined at 25°C using a continuous-excitation apparatus (Fluofluidimeter; Regulest, Florange, France). The excitation wavelength was selected with a band-pass filter centered on 365 nm. The emission wavelength was 425 nm.
The intensities of the emitted light oriented parallel (III) and perpendicular (II) to the plane of the excitation beam were measured. For each sample, five determinations were done and signals from an unlabeled membrane suspension “blank” (IbII and IbI) were also determined to eliminate the influence of the light scattered by the membrane suspension. The fluorescence anisotropy, <r> (1/membrane fluidity), was calculated using the following relationship:
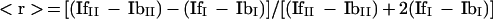 |
The parameter <r> is inversely related to lipid fluidity.
Plasma membrane isolation and purification.
By following the protocol of Rose and Veasey (30), the spheroplasts were suspended in a pH 7.2 Tris-mannitol buffer (50 mM Tris and 0.3 M mannitol adjusted to pH 7.2 with HCl). After 10 min at room temperature, the suspension was incubated for 20 min in an ice bath to make the membranes more rigid. Lysis was then carried out by gentle stirring at low speed with an Ika-Werk homogenizer to mechanically favor osmotic lysis. To ensure the isolation of the plasma membranes, the crushed material was checked with a binocular optic microscope to confirm the absence of cytoplasmic organelles.
The spheroplast lysate was suspended in 3 ml of a 62% (p/vol) sucrose solution then supplemented with a discontinuous gradient of sucrose constituted of 2 ml of 7, 6, 5, 4, 3, 2, and 1% (p/vol) sucrose in mannitol buffer. After centrifugation of this suspension at 100,000 × g, the pellet contained the rough membrane fraction. This preparation was suspended in 4 ml of mannitol buffer in a 40-ml Ultraclear centrifugation tube (Beckman). Then a discontinuous gradient of sucrose was prepared by successively introducing 5 ml of 62 and 50% (p/vol) sucrose and 3 ml of 45, 40, 35, 30, 20, and 10% (p/vol) sucrose in mannitol buffer. After centrifugation at 24,000 × g for 90 min at 4°C, the eight fractions were collected and submitted to a protein concentration determination by the method of Lowry et al. (19). The fractions containing the plasma membranes were also submitted to lipid concentration determination.
Extraction and analysis of membrane constituents.
The membrane lipids were extracted by use of a chloroform-methanol (2:1, vol/vol) mixture for 18 h with gentle stirring at 37°C under nitrogen by following the method of Folch et al. (7), with a slight modification (18 h instead of 3 min).
After filtration on Whatman no. 2 filter paper, a washing with the Folch solvent (chloroform-methanol-water, 200:100:75, vol/vol/vol), and decantation, the chloroform extract was evaporated and dried in a vacuum. The dry extract contained the membrane total lipids and was submitted to saponification in 30% (p/vol) KOH-methanol for 4 h at 100°C under reflux. After cooling, the mixture was filtered on Whatman filter paper. After the addition of 30 ml of distilled water, the filtrate was submitted to three successive extractions with petroleum ether.
The petroleum ether extract containing the sterols was washed with water, dried, evaporated to dryness in a vacuum, and then acetylated with a pyridine-acetic anhydride (1:1, vol/vol) mixture, purified, and finally identified according to the method of Belrhiti et al. (2). The total sterol content was analyzed by the method of Zak (33), with a slight modification (ergosterol was used as the standard instead of cholesterol). The different acetylated sterols were determined by gas-liquid chromatography using a PE 1701 (15 by 0.25 m; Perkin-Elmer) capillary column at 260°C. The nitrogen flow rate was 20 ml/min, and the injector and detector temperatures were 250 and 300°C, respectively.
The aqueous filtrate containing the fatty acid salts was acidified. The free fatty acids liberated were methylated according to the protocol of Belrhiti et al. (2) and identified by gas-liquid chromatography using a PE 225 (30 m by 0.25 mm; Perkin-Elmer) capillary column at 220°C. The nitrogen flow rate was 20 ml/min; the injector and detector temperatures were 250 and 300°C, respectively. Saturated (SFA) (C10 to C18), monounsaturated (MUFA) (C14:1, C16:1, and C18:1), and polyunsaturated (C18:2 and C18:3) fatty acids in methylester form were purchased from Sigma and used as standards.
From the dry extract containing the membrane total lipids, the amount of phosphorus was estimated by the method of MacClare (21) and the phospholipid amount in the sample was estimated by multiplying the phosphorus level of each extract by 25, which is the mean of the phosphorus level in yeast membrane phospholipids (23).
RESULTS
Kinetics of spheroplast formation.
In the present study, in order to obtain a high yield of spheroplasts of all yeast strains, the experimental conditions were 80 mg of pronase E and 30 mg of lysing enzyme. The yeast cells were harvested during their exponential growth phase, as they were most sensitive to lysing enzymes at this stage. Spheroplasts were then prepared under the conditions described in Materials and Methods. As shown in Fig. 1, spheroplast yield was nearly 95% from all the yeast samples after incubation for 2.5 h. Under these conditions, the K. lactis and K. lactis mutant control strains and cells grown in the presence of subinhibitory doses of AmB (10 and 125 mg/liter for K. lactis and the K. lactis mutant, respectively) showed the same spheroplast formation kinetics; therefore, further experiments were carried out with an incubation time of 3 h with lysing enzyme.
FIG. 1.

Kinetics of spheroplast formation from K. lactis and K. lactis mutant strains. The curves correspond to the percentage of spheroplasts obtained during the incubation period with lysing enzyme. The K. lactis and K. lactis mutant strains were grown in a Sabouraud liquid medium without (control) or with AmB and harvested during their exponential growth stage. ■, K. lactis control; □, K. lactis with 10 mg of AmB, ●, K. lactis mutant control, ○, K. lactis mutant with 125 mg of AmB.
Measurement of spheroplast stability. (i) Spheroplast stability versus buffer ionic strength.
Spheroplasts of the K. lactis and K. lactis mutant strains (control cells and cells cultured in the presence of the respective subinhibitory doses of AmB) were suspended in an isotonic solution (pH 5.8) at 30°C. Because of the progressive addition of distilled water until a final volume corresponding to a dilution of 50% was reached, the ionic strength of the solution decreased from 1.8 to 0.8 and the pH increased from 5.8 to 5.96. This change of pH had no effect on spheroplast stability. The incubation period was 5 min, which corresponded to the time necessary for optimal spheroplast lysis.
As shown in Fig. 2, spheroplasts of K. lactis and K. lactis mutant control cells showed 98% stability in an ionic strength range between 1.8 and 1.4 for K. lactis and between 1.8 and 1.6 for the K. lactis mutant. Then the percentage of spheroplasts diminished in relation to the decrease of buffer ionic strength. The residual percentage corresponded to spheroplasts which were not lysed. As shown by the curves, K. lactis mutant spheroplasts were less stable during dilution than K. lactis spheroplasts. The stability of spheroplasts of K. lactis and the K. lactis mutant cultured in the presence of their AmB subinhibitory doses was also diminished by 8% in a buffer ionic strength of 1.4 for K. lactis and by 16% in a buffer ionic strength of 1.6 for the K. lactis mutant. These assays demonstrated that spheroplasts of cells grown in the presence of AmB and spheroplasts of reduced-AmB-sensitivity cells were less stable than spheroplasts of the control strain.
FIG. 2.

Spheroplast stability versus buffer ionic strength. The curves correspond to the percentage of stable spheroplasts in relation to the decrease of buffer ionic strength. ■, K. lactis control, □, K. lactis with 10 mg of AmB, ●, K. lactis mutant control, ○, K. lactis mutant with 125 mg of AmB.
(ii) Spheroplast stability versus incubation time.
When spheroplasts of K. lactis and K. lactis mutant control cells and cells cultured in the presence of AmB were incubated with slight stirring in the isotonic buffer (pH 5.8) at 30°C, an autolytic phenomenon was observed as a function of time.
As shown in Fig. 3, spheroplasts of K. lactis control cells were stable for 20 min and then a constant drop appeared in the time function, while spheroplasts of K. lactis mutant control cells showed a fast and constant drop during the incubation. After 100 min of incubation, 40 and 60% of K. lactis and K. lactis mutant spheroplasts were disrupted.
FIG. 3.

Spheroplast stability versus incubation time. The curves correspond to the percentage of stable spheroplasts in relation to the incubation period. ■, K. lactis control, □, K. lactis with 10 mg of AmB, ●, K. lactis mutant control, ○, K. lactis mutant with 125 mg of AmB.
When K. lactis cells were grown in the presence of AmB, their spheroplasts were less stable than those of control cells. Indeed, after 60 min of incubation, 18% of K. lactis control spheroplasts were lysed while 40% of K. lactis spheroplasts grown in the presence of AmB were disrupted.
The spheroplast stability of the K. lactis mutant was similar after growth of the yeasts in the absence or presence of AmB, but its spheroplasts were more sensitive than those of the K. lactis wild strain cultured under the same conditions.
These assays of stability also showed that at 25°C, an interval no longer than 30 min between obtaining spheroplasts and further experiments such as the determinations of membrane fluidity was sufficient to ensure conditions and allow comparisons between the different spheroplasts.
(iii) Membrane fluidity.
Mean anisotropy values measured after a 5-min incubation at 25°C with TMA-DPH are reported below for the different spheroplasts (n = 18).
The anisotropy value of TMA-DPH embedded in the spheroplasts of the K. lactis mutant strain (<r> = 0.269 ± 0.005 [standard deviation]) was significantly lower than that of the K. lactis control strain (<r> = 0.292 ± 0.004) (P < 0.001 versus K. lactis control). After growth in the presence of AmB, the anisotropy values of the K. lactis and K. lactis mutant strains were lower than that of the corresponding control strains (0.268 ± 0.003 and 0.229 ± 0.003 for K. lactis and K. lactis mutant strains, respectively) (P < 0.001 versus corresponding controls). Moreover, spheroplasts obtained from the K. lactis mutant control strain had an anisotropy value similar to that of spheroplasts obtained from the K. lactis strain cultured in the presence of a subinhibitory dose of AmB.
Membrane lipid relative composition.
The effect of sterols on the plasma membrane osmotic stability of Saccharomyces cerevisiae wild-type and nystatin-resistant spheroplasts has been studied. These strains exhibited differences in membrane fluidity (22). Also, a study of the lipidic constitution of K. lactis and K. lactis mutant spheroplast membranes (control cells and cells cultured in the presence of AmB) was carried out.
As summarized in Table 1, K. lactis and K. lactis mutant spheroplast membranes showed chemical composition differences after culture of the yeasts in the absence or presence of AmB.
TABLE 1.
Membrane lipid levels in K. lactis and K. lactis mutant strains grown in the presence or absence of AmBa
| Strain | Content (mean ± SD) of:
|
Sterol/phospholipid ratio | |||
|---|---|---|---|---|---|
| Membrane lipidsb | Total fatty acidsc | Sterolsc | Phospholipidsc | ||
| K. lactis | 30 ± 0.4 | 18 ± 0.17 | 12.5 ± 0.3 | 12 ± 0.32 | 1.04 |
| K. lactis + AmB | 33 ± 0.5 | 21 ± 0.18 | 16 ± 0.4 | 17 ± 0.94 | 0.94 |
| K. lactis mutant | 36 ± 0.45 | 24 ± 0.21 | 14.5 ± 0.35 | 16 ± 0.90 | 0.90 |
| K. lactis mutant + AmB | 34 ± 0.47 | 23 ± 0.20 | 12.5 ± 0.32 | 19 ± 0.65 | 0.65 |
AmB was used at subinhibitory doses (10 and 125 mg/liter for K. lactis and the K. lactis mutant, respectively).
Results are expressed as percent dry weight.
Results are expressed as percent weight of membrane total lipids.
Membrane total lipid content was 33% ± 1.1% in K. lactis grown with AmB and 30% ± 0.8% in the K. lactis control strain. It was 34% ± 0.9% in the K. lactis mutant grown with AmB and 36% ± 0.95% in the K. lactis mutant control strain (dry weight).
Sterol content was also higher (+28%) in the K. lactis strain grown with AmB and lower (−13.7%) in K. lactis mutant grown with AmB than in their control counterparts. In contrast, phospholipid content (expressed as a percentage by weight of membrane total lipids) increased in both K. lactis and K. lactis mutant cells grown in the presence of AmB (+29.4 and +18.7%, respectively).
The sterol/phospholipid ratio was lower in K. lactis mutant control membranes (0.90) than in K. lactis control membranes (1.04). The sterol/phospholipid ratio was also lower in K. lactis and the K. lactis mutant grown with AmB (0.94 and 0.65, respectively) than in K. lactis and K. lactis mutant control membranes. Moreover, it should be noted that K. lactis cells grown with AmB and K. lactis mutant control cells had similar or nearly identical sterol/phospholipid ratios.
Total fatty acid content (expressed as a percentage by weight of membrane total lipids) was 21% ± 0.6% in K. lactis cells grown with AmB and 23% ± 0.51% in K. lactis mutant cells grown in the presence of AmB; it was 18% ± 0.3% and 24% ± 0.4% of membrane total lipid contents in K. lactis and K. lactis mutant control cells, respectively (Table 1). Analysis of the fatty acid profile showed marked modifications in the relative proportions of several fatty acids (Table 2). The presence of AmB in K. lactis cultures decreased palmitic acid (C16:0) and increased oleic acid (C18:1) levels. In K. lactis mutant cells treated with AmB, oleic acid and linolenic acid (C18:3) levels increased, whereas linoleic acid (C18:2) levels decreased.
TABLE 2.
Fatty acid levels in K. lactis and K. lactis mutant strains grown in the presence or abscence of AmBa
| Fatty acid | % of total fatty acid content in straina
|
|||
|---|---|---|---|---|
| K. lactis | K. lactis + AmB | K. lactis mutant | K. lactis mutant + AmB | |
| C15:0 | tr | tr | 4.2 ± 0.2 | |
| C16:0 | 35 ± 1.1 | 15 ± 0.8 | 25 ± 0.8 | 26.5 ± 0.9 |
| C16:1 | 6 ± 0.4 | 11 ± 0.6 | 5 ± 0.2 | 4.5 ± 0.2 |
| C18:0 | 4 ± 0.2 | 2 ± 0.1 | 3 ± 0.1 | 4 ± 0.2 |
| C18:1 | 29 ± 3 | 45 ± 4.0 | 27 ± 3 | 44 ± 5 |
| C18:2 | 19 ± 0.5 | 21 ± 0.8 | 36 ± 1.1 | 7 ± 0.5 |
| C18:3 | 3 ± 0.1 | 1 ± 0.2 | 1.5 ± 0.1 | 5 ± 0.3 |
| C20:0 | 4 ± 0.2 | 4 ± 0.3 | tr | 3 ± 0.2 |
| SFA | 43 | 21 | 28 | 37.7 |
| MUFA | 35 | 56 | 32 | 48.1 |
| SFA/MUFA ratio | 1.23 | 0.38 | 0.87 | 0.78 |
The mean values of fatty acid chain lengths were estimated by the formula Σ (n × %)/100, in which n is the number of carbons in the chain and % is the percentage of the derivatives. Values are means ± standard deviations. AmB was used at subinhibitory doses (10 and 125 mg/liter for K. lactis and the K. lactis mutant, respectively).
It follows that the relative proportions of MUFA increased by 60 and 51.5% in K. lactis and K. lactis mutant cells grown in the presence of AmB. The relative proportion of palmitic acid (C16:0) in K. lactis mutant cells was also 28.5% lower than in K. lactis wild cells.
Therefore, the SFA/MUFA ratio decreased in cells treated with AmB or in K. lactis mutant cells compared to the K. lactis wild strain.
It is noteworthy that the sterol compositions (Table 3) in both control yeasts and yeasts grown with AmB were in agreement with those found in previous studies (11). Ergosterol content decreased by 55 and 32%, respectively, in K. lactis and the K. lactis mutant grown in the presence of AmB, while the content of precursors (zymosterol, dehydroergosterol, dihydroergosterol, and lanosterol) increased under these growth conditions.
TABLE 3.
Membrane sterol levels in K. lactis and K. lactis mutant strains grown in the presence or absence of AmBa
| Compound | Relative retention timeb | % of sterol content in strain
|
|||
|---|---|---|---|---|---|
| K. lactis | K. lactis + AmB | K. lactis mutant | K. lactis mutant + AmB | ||
| Cholesterol | 1 | 1 ± 0.2 | 3 ± 0.4 | 5 ± 0.4 | 6 ± 0.5 |
| Dehydroergosterol | 1.08 | 2 ± 0.3 | 5 ± 0.5 | 2 ± 0.4 | 8 ± 0.7 |
| Zymosterol | 1.12 | 3 ± 0.6 | 12 ± 1.0 | 9 ± 0.8 | 13 ± 1.1 |
| Dihydroergosterol | 1.20 | 5 ± 0.5 | 17 ± 1.5 | 3 ± 0.4 | 5 ± 0.6 |
| Ergosterol | 1.23 | 81 ± 4 | 37 ± 4 | 65 ± 3 | 44 ± 5 |
| 4-Methylzymosterol | 1.27 | 2 ± 0.4 | tr | 12 ± 0.9 | 7 ± 0.7 |
| Fecosterol | 1.28 | tr | 3 ± 0.3 | tr | 2 ± 0.3 |
| Stigmasterol | 1.35 | tr | 2 ± 0.3 | tr | tr |
| Episterol | 1.37 | 2 ± 0.2 | 5 ± 0.3 | 1 ± 0.1 | 7 ± 0.6 |
| Ergosta-7-ene-3-ol | 1.40 | tr | tr | tr | 2 ± 0.2 |
| Lanosterol | 1.45 | 2 ± 0.3 | 9 ± 1.3 | 1 ± 0.2 | 5 ± 0.5 |
| 4,4-Dimethylzymosterol | 1.51 | tr | 6 ± 0.5 | 1 ± 0.2 | tr |
| 4,4-Dimethyl-ergosta-8,24(28)diene-3-ol | 1.73 | 1 ± 0.2 | tr | tr | tr |
AmB was used at subinhibitory doses (10 and 125 mg/liter for K. lactis and the K. lactis mutant, respectively).
Retention times are expressed as ratio to cholesterol retention time (1).
DISCUSSION
The anisotropy value of the fluorescence emitted by TMA-DPH embedded in membranes of the K. lactis mutant strain was lower than that recorded for the membrane of the K. lactis wild strain.
After culture of the K. lactis and K. lactis mutant yeasts in the presence of subinhibitory doses of AmB (10 and 125 mg/liter, respectively), the anisotropy value of the probe in the plasma membranes was lower than that of the control cells. This decrease in anisotropy value was not due to interference of AmB with the spectroscopic characterization of the fluorescent probe; indeed, AmB was totally eliminated from yeast membranes during the exponential phase of the cells, as shown by spectroscopic analysis of yeast and spheroplast membranes. The decreased anisotropy value in samples treated with AmB was also not due to a different stability of the spheroplasts from yeasts cultured in the presence of AmB. Indeed, studies of stability versus the buffer ionic strength and versus the incubation time showed that the spheroplasts were stable for 20 min at 25°C, the period required for fluorescence measurements. Therefore, the decreased anisotropy value truly expressed an increase in membrane fluidity induced by AmB.
Several hypotheses may be put forward to explain these results.
First, AmB binding to the fungal cell membrane is a spontaneous and relatively fast process. The increased membrane fluidity may result from a local rearrangement of the membrane components, which is due to a delocalization of ergosterol from phospholipid-ergosterol interactions. Delocalization induces in turn an increase in phospholipid-phospholipid interactions and the emergence of phospholipid-enriched domains with a higher fluidity. Furthermore, the interaction of AmB with the membrane components provokes, inside the membrane layers, the formation of several types of channels differentiated by their diameter (1, 6, 12). At low AmB concentrations, similar to those used in our study, the formation of channels with a low diameter induces a transitory high permeability for water and a loss of K+ and Mg2+ (17). Divalent cations such as Mg2+ and Ca2+ play an important role in stabilizing biological membranes. They form ionic bonds with phospholipid phosphate groups, which lead to lipid binding and decreased fluidity (5). Following channel formation, these divalent cations leak out. Phospholipids are no longer bound, and membrane fluidity may be increased.
Second, higher fluidity may also result from changes in structural properties of the membrane, such as a decrease in sterol/phospholipid ratios (31) or an increase in the degree of fatty acid unsaturation (15). In our study, we showed a decrease in sterol/phospholipid ratios in K. lactis mutant control cells as well as in K. lactis and in the K. lactis mutant grown in the presence of subinhibitory doses of AmB. This decrease in sterol/phospholipid ratios might be due in both yeasts to an excess of phospholipid synthesis and might play an important role in the resistance of yeasts to AmB. Indeed, it has been reported that phospholipid-enriched cells could selectively protect S. cerevisiae cells from the action of AmB, leading to acquired resistance against different polyenes (28).
AmB treatment also induced an increase in the relative proportion of MUFA in K. lactis and K. lactis mutant cells. It is well known that an overall decrease in the membrane molecular order is related to increased MUFA content in the membrane phospholipids (8, 10). The nature of the fatty acyl chain of phospholipids is one of the most important factors modulating the fluidity and integrity of the membrane. The SFA/MUFA ratio may greatly influence membrane properties. A lower ratio can give rise to better membrane permeability (25).
Thus, changes in the structural properties of the membrane of K. lactis mutant cells as well as that of K. lactis and K. lactis mutant cells treated with AmB, the decreased sterol/phospholipid ratio, the increased relative proportion of MUFA, and the decreased SFA/MUFA ratio explain the higher membrane fluidity, which would represent a metabolic resistance of the yeast to AmB.
REFERENCES
- 1.Anachi R B, Bansal M, Easwaran K R K, Namboodri K, Gaber B P. Molecular modeling studies on amphotericin B and its complex with phospholipid. J Biomol Struct Dyn. 1995;12:957–970. doi: 10.1080/07391102.1995.10508790. [DOI] [PubMed] [Google Scholar]
- 2.Belrhiti A, Naji B, Bonaly R. Membrane lipid and sterol distribution in Saccharomyces bayanus. Process Biochem. 1995;30:427–433. [Google Scholar]
- 3.Bolard J, Joly B, Yeni P. Mechanism of action of amphotericin B at the cellular level. Its modulation by delivery systems. J Liposome Res. 1993;3:409–428. [Google Scholar]
- 4.Brajtburg J, Powderly W G, Kobayashi G S, Medoff G. Amphotericin B: current understanding of mechanisms of action. Antimicrob Agents Chemother. 1990;34:183–188. doi: 10.1128/aac.34.2.183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Carratu L, Franceschelli S, Pardini C L, Kobayashi G S, Horvath I, Vigh L, Maresca B. Membrane lipid perturbation modifies the set point of the temperature of heat shock response in yeast. Proc Natl Acad Sci USA. 1996;93:3870–3875. doi: 10.1073/pnas.93.9.3870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cohen B. A sequential mechanism for the formulation of aqueous channels by amphotericin B in liposomes. The effect of sterols and phospholipid composition. Biochim Biophys Acta. 1992;1108:49–58. doi: 10.1016/0005-2736(92)90113-z. [DOI] [PubMed] [Google Scholar]
- 7.Folch J, Lees M, Sloane Stanley G H S. A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem. 1957;226:497–509. [PubMed] [Google Scholar]
- 8.Gargano S, Di Lallo G, Kobayashi G S, Maresca B. A temperature sensitive strain of Histoplasma capsulatum has an altered delta-9 fatty acid desaturase gene. Lipids. 1995;30:899–906. doi: 10.1007/BF02537480. [DOI] [PubMed] [Google Scholar]
- 9.Gold W, Stout H A, Pagona J F, Donovick R. Amphotericin A and B: antifungal antibiotics produced by a streptomycete. Antibiot Annu. 1956;1956:579–586. [PubMed] [Google Scholar]
- 10.Gyorfy Z, Benko S, Kusz E, Maresca B, Vight L, Ouda E. Highly increased TNF sensitivity of tumor cells expressing the yeast delta-9 desaturase gene. Biochem Biophys Res Commun. 1997;241:465–470. doi: 10.1006/bbrc.1997.7835. [DOI] [PubMed] [Google Scholar]
- 11.Hakkou A, Coulon J, Mpona-Monga M, Bonaly R. Variations des stérols chez Kluyveromyces lactis chez un mutant résistant par culture en présence de doses sublétales d'amphotéricine B. Can J Microbiol. 1988;34:787–792. [PubMed] [Google Scholar]
- 12.Hartsel S C, Benz S K, Ayenew W, Bolard J. Na+, K+ and Cl− selectivity of the permeability pathways induced through sterol-containing membrane vesicles by amphotericin B and other polyenes. Eur Biophys J. 1994;23:125–132. doi: 10.1007/BF00208866. [DOI] [PubMed] [Google Scholar]
- 13.Horsburgh C R, Jr, Kirkpatrick C H, Teutsch C B. Ketoconazole and the liver. Lancet. 1982;i:860. doi: 10.1016/s0140-6736(82)91915-8. [DOI] [PubMed] [Google Scholar]
- 14.Iannitelli R C, Ikawa M. Effect of fatty acids on action of polyene antibiotics. Antimicrob Agents Chemother. 1980;17:861–864. doi: 10.1128/aac.17.5.861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kataoka R, Mitaku S, Kinosita K, Ikegami A. Interaction between Ca2+ and dipalmitoylphosphatidylcholine membranes. II. Fluorescence anisotropy study. Biophys Chem. 1985;21:277–284. doi: 10.1016/0301-4622(85)80015-6. [DOI] [PubMed] [Google Scholar]
- 16.Khutorsky V E. Structures of amphotericin B-cholesterol complex. Biochim Biophys Acta. 1992;1108:123–127. doi: 10.1016/0005-2736(92)90015-e. [DOI] [PubMed] [Google Scholar]
- 17.Kotler-Brajtburg J, Medoff G, Kobayashi G S, Boggs S, Schlessinger D, Pandey R C, Rinehart K L., Jr Classification of polyene antibiotics according to chemical structure and biological effects. Antimicrob Agents Chemother. 1979;15:716–722. doi: 10.1128/aac.15.5.716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Langlet J, Berges J, Caillet J, Demaret J P. Theoretical study of amphotericin B with sterols. Biochim Biophys Acta. 1994;1191:79–93. doi: 10.1016/0005-2736(94)90235-6. [DOI] [PubMed] [Google Scholar]
- 19.Lowry O H, Rosebrough N J, Farr A L, Randall R J. Protein measurement with the Folin phenol reagent. J Biol Chem. 1951;193:265–275. [PubMed] [Google Scholar]
- 20.Lutwick L I, Phaff H J, Stevens D A. Kluyveromyces fragilis as an opportunistic fungal pathogen in man. Sabouraudia. 1980;18:69–73. [PubMed] [Google Scholar]
- 21.MacClare C W F. An accurate and convenient organic phosphorus assay. Anal Biochem. 1971;39:527–535. doi: 10.1016/0003-2697(71)90443-x. [DOI] [PubMed] [Google Scholar]
- 22.MacLean-Bowen C A, Parks L W. Effect of altered sterol composition on osmotic behavior of spheroplasts and mitochondria of Saccharomyces cerevisiae. Lipids. 1982;17:662–665. doi: 10.1007/BF02535375. [DOI] [PubMed] [Google Scholar]
- 23.Nagar-Leyman R, Magalith L. A comparative study of the lipid composition of yeasts with different fermentation capacities. Appl Microbiol Biotechnol. 1987;26:49–54. [Google Scholar]
- 24.Odds F C. Resistance of yeasts to azole-derivative antifungals. J Antimicrob Chemother. 1993;31:463–471. doi: 10.1093/jac/31.4.463. [DOI] [PubMed] [Google Scholar]
- 25.Panchal C J, Stewart G G. Regulatory factors in alcohol fermentation. In: Stewart G G, Russel I, editors. Current developments in yeast research. Toronto, Canada: Pergamon Press; 1980. pp. 9–15. [Google Scholar]
- 26.Prendergast F G, Haugland R P, Callahan P J. 1-[4(Trimethylamino)phenyl]-6-phenylhexa-1,3,5-triene: synthesis, fluorescence properties, and use as a fluorescence probe of lipid bilayers. Biochemistry. 1981;20:7333–7338. doi: 10.1021/bi00529a002. [DOI] [PubMed] [Google Scholar]
- 27.Ramanandraibe E, Younsi M, Coulon J, Loppinet V, Hakkou A, Bonaly R. Implication of cell wall constituents in the sensitivity of Kluyveromyces lactis strains to amphotericin B. Res Microbiol. 1998;149:109–118. doi: 10.1016/s0923-2508(98)80026-0. [DOI] [PubMed] [Google Scholar]
- 28.Rao T V, Trivedi A, Prasad R. Phospholipid enrichment of Saccharomyces cerevisiae and its effect on polyene sensitivity. Can J Microbiol. 1985;31:322–326. doi: 10.1139/m85-061. [DOI] [PubMed] [Google Scholar]
- 29.Rex J H, Rinaldi M G, Pfaller M A. Resistance of Candida species to fluconazole. Antimicrob Agents Chemother. 1995;39:1–8. doi: 10.1128/aac.39.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rose A H, Veasey F J. Membranes and lipid of yeast. In: Campbell I, Duffus J H, editors. Yeast: a practical approach. Oxford, England: IRL Press; 1988. pp. 255–275. [Google Scholar]
- 31.Van Blitterswijk W J, Van Hoeven R P, Van der Meer B W. Lipid structural order parameter (reciprocal of fluidity) in membranes derived from steady-state fluorescence polarisation measurement. Biochim Biophys Acta. 1981;644:323–332. doi: 10.1016/0005-2736(81)90390-4. [DOI] [PubMed] [Google Scholar]
- 32.Warnock D W. Amphotericin B: an introduction. J Antimicrob Chemother. 1991;28:27–38. doi: 10.1093/jac/28.suppl_b.27. [DOI] [PubMed] [Google Scholar]
- 33.Zak B. Simple rapid microtechnic for serum total cholesterol. Am J Clin Pathol. 1967;27:583. doi: 10.1093/ajcp/27.5_ts.583. [DOI] [PubMed] [Google Scholar]