

Abstract
Gene drives can be highly effective in controlling a target population by disrupting a female fertility gene. To spread across a population, these drives require that disrupted alleles be largely recessive so as not to impose too high of a fitness penalty. We argue that this restriction may be relaxed by using a double gene drive design to spread a split binary expression system. One drive carries a dominant lethal/toxic effector alone and the other a transactivator factor, without which the effector will not act. Only after the drives reach sufficiently high frequencies would individuals have the chance to inherit both system components and the effector be expressed. We explore through mathematical modeling the potential of this design to spread dominant lethal/toxic alleles and suppress populations. We show that this system could be implemented to spread engineered seminal proteins designed to kill females, making it highly effective against polyandrous populations.
Subject terms: Biotechnology, Computational biology and bioinformatics, Genetics
Introduction
There are, in most parts of the world, insect species that inflict significant harm to the human condition. These insect species, usually invasive, cause or transmit diseases, reduce agricultural production, or threaten endemic biodiversity (reviewed in1). However, only a few of these species can be effectively controlled by current methods, in many cases by using unsustainable or ecologically damaging strategies such as broad-spectrum pesticides. More recently, integrative pest management (IPM) has focused on minimizing the use of pesticides, while promoting other less ecologically damaging strategies, such as biological control2. The great efficiency and versatility of CRISPR-Cas9 nucleases for genome editing and the super-Mendelian inheritance of self-propagating gene drives have recently made engineered CRISPR-Cas9-mediated (gene) drives one of the most promising—though controversial—technologies for biological control3–10. This young technology could be used, for instance, to quickly spread desired genetics variants (e.g., payload genes) through a target population after a single release of transgenic individuals carrying the engineered drive.
During meiosis in sexually reproducing species, heterozygous individuals containing a single drive allele can have it copied onto the homolog wild-type (wt) chromosome in a process known as drive conversion or homing3. Homing can increase the inheritance rate of gene drive alleles beyond the Mendelian rule of gene segregation, pushing it towards 100%. Therefore, a drive can spread through a population even if it confers a fitness cost3. Based on this mechanism, fertility or viability suppression drives are intended to introduce a considerable genetic load into the target population by homing and disrupting a gene essential for fertility or viability. The potential of this approach has been shown in silico through mathematical modeling (e.g.,11–17). A suppression drive targeting the doublesex gene, which controls sex determination and is essential for female fertility in mosquitoes18, is among the most effective gene drive systems recently developed in the laboratory (e.g.,7,10,19,20). This doublesex gene drive was able to collapse cage populations of the Anopheles gambiae mosquito in as few as seven generations18. However, a number of technical limitations still call into question the versatility of this approach in the field21–28.
One of the biggest limitations to the spread of a single gene drive system (affecting fertility or viability) is the requirement that the disrupted allele effects be largely recessive and preferentially target only females, i.e., the target gene should be haplosufficient for females while, for the best outcome, nonessential for males12,15. If the drive carries a dominant or semidominant lethal gene, which may avoid the need of targeting haplosufficient endogenous genes, it imposes a high fitness penalty and, consequently, it may not be able to invade the population. In many species, however, finding haplosufficient endogenous genes may be challenging, not only because it requires a deep and comprehensive knowledge of female and male fertility/viability genetics but also because many of the essential genes involved in female fertility/viability may affect pleiotropically other life-history traits and/or may not be largely haplosufficient (i.e., mutant heterozygotes may experience reduced fertility/viability), both of which impede drive spread.
We propose that the haplosufficiency limitation of suppression drives may be significantly reduced by deploying a double CRISPR-based gene drive design to spread a split binary expression system. One drive would carry a transactivator factor (Fig. 1a) and the other would carry a dominant or semidominant lethal/toxic effector (Fig. 1b). As part of a binary expression system, the effector would only be expressed in the presence of the transactivator protein, so the system would impose a low fitness cost at its introduction and may allow each drive alone to rapidly spread from rare. After some generations, only when each drive reaches sufficiently high frequencies, both system components would have meaningful chances to co-occur within an individual. At this point the effector would be expressed killing or sterilizing the individual, eventually leading to a decline in the target population. We refer to this double gene drive design as the Binary Expression Drive (BED). The concept is similar to the ‘interacting drives’ approach proposed by Esvelt et al.6. However, while the ‘interacting drives’ approach achieves suppression effects by disrupting native, essential genes, BED aims to cause the expression of non-native, dominant toxins.
Figure 1.

Binary expression drive constructs. Left: the transactivator construct (a). Right: the effector construct (b). The promoter of the transactivator gene is only active in the target tissue/organ, for instance, the ovaries or the male accessory glands. The transactivator protein will activate the expression of the effector gene, which encodes a dominant or semidominant toxin, in the target tissue/organ. In both constructs, the expression of Cas9 and the gRNA is under the regulation of a germline promoter that allows drive conversion during meiosis. Ideally, target loci are highly conserved genes that cannot tolerate mutations so resistance alleles are selected out of the population while each drive allele alone, carrying a re-coded target gene, will not impose an intended fitness cost.
BED could be used, for instance, to spread intracellular toxic proteins to be specifically expressed in the female reproductive organs so adult females carrying both the transactivator and the effector constructs would be sterilized. We will refer to this design as ‘female fertility BED’ (ffBED). The BED concept could be modified to spread engineered seminal proteins designed to be activated in the uterus and kill or sterilize females. We will refer to this design as a ‘seminal BED’ (sBED). Males carrying a sBED will behave as "terminators" of females, transferring into them lethal toxins. Nonetheless, such artificial seminal toxins are currently hypothetical. In polyandrous species, where females mate with several males, terminator males will be able to kill more than one female, thus making the intervention more efficient than it would be in monogamous species.
Here, we developed a model for stochastic simulations in the R programming language to study the behavior of BEDs and compare it to female fertility single drives (ffSD) targeting a haplosufficient but essential female fertility gene. In both BED designs described here (female fertility or seminal), the transactivator and effector constructs are inserted into unlinked autosomal loci that are essential for embryonic development (so resistance alleles are selected out of the population). In the ffBEDs, the transactivator construct is exclusively expressed in the ovary, and the effector construct encodes a toxin that sterilizes the female. In the sBEDs, the transactivator construct is only expressed in the male accessory gland, and the effector construct encodes a seminal toxin that is innocuous to the male that produces it but lethal to the female that receives it via the ejaculate during copulation. In ffSDs, the target gene is essential for female fertility but has no effect on male fitness.
The model assumes a panmictic population with discrete, non-overlapping generations. Each generation is composed of four stages: (1) eggs, (2) larvae (or juveniles), (3) virgin adults (females and males), and (4) mothers (i.e., inseminated females) which produce the next generation eggs (Fig. 2). Given that polyandry is generally common among insects29–31, the modeled population is, by default, polyandrous such that females may copulate with up to three males before producing offspring. A single release of transgenic males carrying each drive takes place at generation zero. Drives efficiency, intended effects, and unintended fitness costs, among other intrinsic drives and population features, are adjustable (see Tables 1 and 2 for a complete description of main model parameters).
Figure 2.

Model stages and steps across a generation. Major invoked parameters are shown per step. (1) Female and male virgin adults mate randomly giving rise to mothers, i.e., inseminated females. Females may mate more than once. Male mating success of drive-carrying males is reduced by unintended drive effects, and terminator males may kill the female during mating. (2) For simplicity, meiosis and homing conversion are only modeled for surviving mothers and mating males. (3) Each mother produces a clutch of eggs where the expected number of eggs may be reduced for drive-carrying mothers. (4) Eggs have a probability of dying before reaching the larval stage, which may be higher for drive-carrying eggs. (5) Larvae have a probability of dying right after hatching. (6) Then, larvae are subject to density-dependent mortality. (7) Surviving larvae result in virgin adults, and sex is randomly determined.
Table 1.
Main population parameters.
| Name | Description | Default value |
|---|---|---|
| Fecundity | Expected number of fertile eggs produced by a mother. For each mother, the number of eggs is generated from a Poisson distribution with lambda equals Fecundity | 48 |
| Larval survival |
Density-independent larval-to-adult survival rate. Fecundity and Larval survival together define the expected number of adult female offspring produced per adult female in the absence of intraspecific competition (Rm), i.e., the growth rate at low-density, far from the carrying capacity
In turn, Rm defines the critical genetic load (Lc = 1—1/Rm), over which population is expected to crash in the absence of stochastic effects14,15 |
0.5 (Rm = 12) (Lc = 0.92) |
| Critical population size |
Number of virgin adult males below which mate-finding Allee effect occurs. In such a case, the chances that virgin females achieve mating (CM) and the number of mates per female (NM) are modeled as positive sigmoid functions of the population size yielding a 50% reduction at the Critical population size
|
250 |
| Polyandry degree | Maximum and most likely number of mates per adult female. For each female, the number of mates is generated from a Poisson distribution truncated between 1 and Polyandry degree, with lambda equals two times Polyandry degree | 3 |
A description and default value are shown for each parameter.
Table 2.
Main system parameters.
| Name | Description | Default value | ||
|---|---|---|---|---|
| ffBED | sBED | ffSD | ||
| Intended fecundity cost | Relative reduction of mother’s fertility (total number of fertile eggs a mother is expected to produce) caused by drive system activity. In BED designs, this reduction is experienced by mothers homozygous for both drive system components (AABB mothers, whereas A and B are each of the gene drive components). In SD designs, it is experienced by mothers homozygous for the drive (CC, where C is the gene drive). If Dominance degree is higher than 0 (see below), this cost will also be experienced, at least partially, by non-homozygous mothers carrying at least one allele of each system component in BED designs (AABb, AaBB, or AaBb mothers) or by drive-carrying heterozygous mothers in SD designs (Cc mothers) (lower case letters designate native alleles) | 1 | 0 | 1 |
| Terminator efficiency | Probability that a female that copulates with an AABB terminator is killed following mating due to the action of the drive system. It only applies to BED designs. If Dominance degree is higher than 0 (see below), Terminator efficiency will also apply, at least partially, for non-AABB terminators (AABb, AaBB, or AaBb males) | 0 | 0.95 | 0 |
| Dominance degree | Proportion of the drive system effect (Terminator efficiency or Intended fecundity cost) experienced by double heterozygotes (AaBb) carrying one allele of each drive in BED designs or by drive-carrying heterozygotes (Cc) in SD designs. Drive-carrying homozygotes experience full intended effect (see above). In BED designs, individuals homozygous for one drive but heterozygous for the other will experience an intermediate effect between the effect experienced by AaBb and AABB individuals. In SD designs, this parameter also affects the expected fecundity of heterozygous mothers carrying a resistance allele (see Resistance allele functionality below) | 0.6 | 0.2 | |
| Unintended reproductive cost | Relative reduction of female fertility and male mating success due to unintended drive activity. For drive-carrying mothers, it is the reduction of the expected number of fertile eggs. For drive-carrying males, it is the reduction in mating success. These costs are dominant (i.e., mothers and males carrying one and two drive alleles experience the same reduction) and, in BED designs, are imposed by each drive separately 1 − (1 − Unintended reproductive cost)2 | 0 (IS) or 0.025 (RS) | ||
| Unintended viability cost | Probability that a drive-carrying egg perishes due to unintended drive activity. This cost is dominant (i.e., eggs carrying one and two drive alleles experience the same viability reduction) and, in BED designs, is imposed by each drive separately 1 − (1 − Unintended viability cost)2 | 0 (IS) or 0.025 (RS) | ||
| Homing efficiency | Probability that a drive converts the wild target locus into a drive allele during gametogenesis in drive-carrying heterozygotes | 1 (IS) or 0.9 (RS) | ||
| Resistance allele formation | Probability that a drive converts the wild target locus into a cleavage-resistant allele (resistance allele) during gametogenesis in drive-carrying heterozygotes | 0 (IS) or 0.05 (RS) | ||
| Resistance allele functionality | Relative functionality of resistance alleles. Ranging from 0 to 1, in BED designs (where target genes are essential for development), it is the probability that viable eggs homozygous for resistant alleles of any drive develop into a larva. In SD designs, where the target gene is essential for female fertility, it is the fraction of Fecundity preserved in mothers homozygous for resistant alleles. To be conservative in BED designs, the deleterious effect of resistance alleles was assumed to be recessive (i.e., resistant-wt-heterozygotes, resistant-drive-heterozygotes, and wt-homozygotes are equally functional for the target genes). In ffSD, the expected fecundity of resistant-wt-heterozygous females (or resistant-drive-heterozygous females) is intermediate between resistance-homozygous females (or drive-homozygous females, respectively) and cc females, whereas Dominance degree determines linearly how close fecundity of resistant-wt-heterozygotes (or resistant-drive-heterozygotes) is to resistance-homozygotes' (or drive-homozygotes', respectively) | 0 | ||
A description and the default value(s) are shown for each parameter. For the three parameters related to intended drive effects, the default value may vary between designs (ffBED, sBED, and ffSD). For the four parameters related to drive endonuclease activity and unintended fitness cost, two default values are included: one for a system with ideal drive activity [Ideal System (IS)], that has perfect conversion rate and does not impose unintended fitness costs, and the other for a system with reference drive activity [Reference System (RS)], with imperfect conversion and unintended fitness costs.
Results
Inspired by the performance of gene drive systems recently established in flies and mosquitoes7,10,18,20,26,32,33, and previous studies on modeling of gene drives for suppression of insects (e.g.,12,16,34–36), we chose realistic—yet optimistic—parameters values as default (Tables 1 and 2). Default values for population parameters are not intended to reflect any particular species but a hypothetical ordinary insect with reasonable attributes matching those previously observed in insect species (e.g.,15,37). For each modeled design, two classes of drive activity were considered: ideal and reference. In ideal systems, drive conversion rate (i.e., homing efficiency) is 100% (which implies no resistance allele formation) and drive(s) activity imposes neither reproductive nor viability fitness cost. In reference systems, homing efficiency is 90%, resistance allele formation rate is 5%, unintended reproductive cost (imposed on male mating success and female fecundity by each drive separately) is 2.5%, and unintended viability cost (imposed on eggs also by each drive separately) is also 2.5%. As both Unintended reproductive cost and Unintended viability cost were kept equal in all the performed simulations, we refer to these parameters together as Unintended fitness costs. As other studies (e.g.11,12,14) only modeled one or the other fitness cost (viability or female fecundity/male mating success), the value of Unintended fitness costs in this study corresponds to larger fitness costs in most other papers.
To test the performance of our model we first set our parameters to their defaults (ideal or reference) and then explored parameter space for one or two parameters at a time. For every considered scenario, we performed 100 simulations, each of which comprised up to 36 generations starting from 50,000 virgin adults and 250 adult transgenic males released per drive (0.5% of the population carrying capacity) in BED designs or 500 in SD designs. In all cases, we recorded the adult population size, genetic load (i.e., reduction of the expected number of adult female offspring produced per adult female in the absence of intraspecific competition), and allele frequencies (drives, wt, and resistance alleles) in every generation, and the rate of elimination within 36 generations, which we termed ‘fast elimination rate’. The number of replicate simulations was set to 100 since, at least with default parameters, the dispersion (difference between the minimum and maximum) of the final adult population size always stabilizes within 50 simulations.
Homing efficiency and unintended drive effects
Figure 3 and Fig. S1 summarize the performance of the three gene drive designs with ideal and reference drive activity. Using the ideal drive parameters, we observed a fast and complete elimination rate for all three designs. However, using reference drive parameters (i.e. assuming imperfect drive activity), performance differed between designs. Overall, ffBED showed the worst performance. It was not able to eliminate the target population in any simulation, although it reduced the population by ~ 51% in 16 generations; and genetic load stabilized at around 0.84 after generation 14. Irrespective of how low the drive costs on fitness were, the population was only eliminated under perfect homing efficiency (Fig. 3a). Furthermore, compared to sBED or ffSD, genetic load imposed by ffBED was remarkably lower across any simulated combination of Homing efficiency and Unintended fitness costs (Fig. S1). In contrast, sBED showed great suppression potential, tolerating a wide range of Homing efficiency and Unintended fitness costs (Fig. 3b, Fig. S1). Therefore, further exploration of alternative scenarios and parameter space is only shown for sBED alone or together with ffSD for comparison purposes. To explore and compare the suppression potential of these designs, we opted to show the fast elimination rate and the progression of the population size, which strongly correlates to the imposed genetic load (Fig. S2).
Figure 3.

Suppression potential of ffBED (a), sBED (b), and ffSD (c). Left: population size (relative to the initial population size), drive(s) allele frequency, and genetic load are shown across the first 25 generations for each design using ideal and reference drive parameters. Lines represent the 100-simulations mean value while the colored envelopes stand for the maximum and minimum values. For BED designs, frequencies of the two drives were averaged. Right: fast elimination rate (the percentage of simulations where the population was eliminated within 36 generations) is shown for a combined space of Homing efficiency and Unintended fitness costs (Unintended reproductive cost and Unintended viability cost). Resistance allele formation was assumed to be half the complement of Homing efficiency (e.g., if Homing efficiency is 0.8, Resistance allele formation is 0.1).
In general, sBED was superior to ffSD when comparing the fast elimination rate (within 36 generations) across a range of Homing efficiency and Unintended fitness costs (Fig. 3, right panels). With reference drive parameters, for instance, while sBED always eliminated the population in less than 36 generations, ffSD did it in 97% of the simulations taking, on average, 6.7 extra generations. Seminal BED also succeeded in suppressing the population under conservative levels of Homing efficiency and Unintended fitness costs. For instance, when efficiency was reduced to 0.8 and Unintended fitness costs raised to 0.05, the intervention resulted in a population collapse in 82% of the cases in less than 24 generations. Under the same conditions, ffSD did not result in the population collapsing before generation 36 in any simulation. However, when Homing efficiency and Unintended fitness costs were set at 0.95 and 0.125, respectively (or very similar values), the fast elimination rate was slightly higher for ffSD (100%) than for sBED (94%).
System dominance
While the Terminator efficiency parameter (in sBED designs) or the Intended fecundity cost parameter (in ffSD designs) determines the system intended effect expected for drive-carrying homozygotes, the Dominance degree parameter determines the extent to which this intended effect manifests in drive-carrying heterozygotes (Table 2). In sBED designs, a certain degree of system dominance (Dominance degree > 0.5) may be expected given the deleterious effects of having a male-derived toxin that efficiently kills the female within the reproductive tract of females. In other words, the consequence of having even an intermediate amount of toxin is likely severe enough and would not scale linearly. Thus, the phenotypic consequence of mating with a terminator male heterozygous for both drive system components (AaBb) will be closer to mating with a double homozygous terminator male (AABB) than to mating with a wt male (aabb). On the other hand, homozygous transgenic organisms will typically yield a higher level of transgene expression than heterozygotes (e.g.,38–40). Consequently, AABB males, relative to AaBb heterozygotes, would likely produce higher amounts of the toxin and, in turn, are expected to cause a stronger deleterious effect on females (Dominance degree < 1). Therefore, for sBED, we arbitrarily chose 0.6, a value between 0.5 and 1, as the default Dominance degree value (i.e., the transgenic toxin was considered semidominant). This means that adult males carrying one copy of each drive exhibit, relative to double homozygous drive-carrying males, 60% of the ability to kill females during mating. In ffSD designs, the default value was 0 (i.e., the target was supposed to be completely haplosufficient), which means that heterozygous mothers experience no intended fecundity reduction.
To analyze how sBED may deal with more dominant or recessive expression systems and how ffSD may deal with more dominant targets, we further explored the performance of both designs with ideal and reference drive parameters at several Dominance degree values. In this way, we determined for each design the range of Dominance degree for which the drive preserves a fast elimination rate higher than 80% (which we dubbed ‘efficient suppression potential’). For sBED, Dominance degree did not notably affect the performance using ideal drive parameters (Fig. 4a). With imperfect drive activity (reference drive parameters), the sBED preserved efficient suppression potential except under very low or very high Dominance degree values (< 0.15 or > 0.93, respectively). sBED potential for suppression was maximized at ~ 0.7. For ffSD, Dominance degree strongly affected the drive performance using both ideal and reference drive parameters (Fig. 4b). The ideal and reference ffSDs showed efficient suppression potential with Dominance degree values lower than 0.85 and 0.68, respectively. In both cases, suppression potential was optimized at around 0.25.
Figure 4.
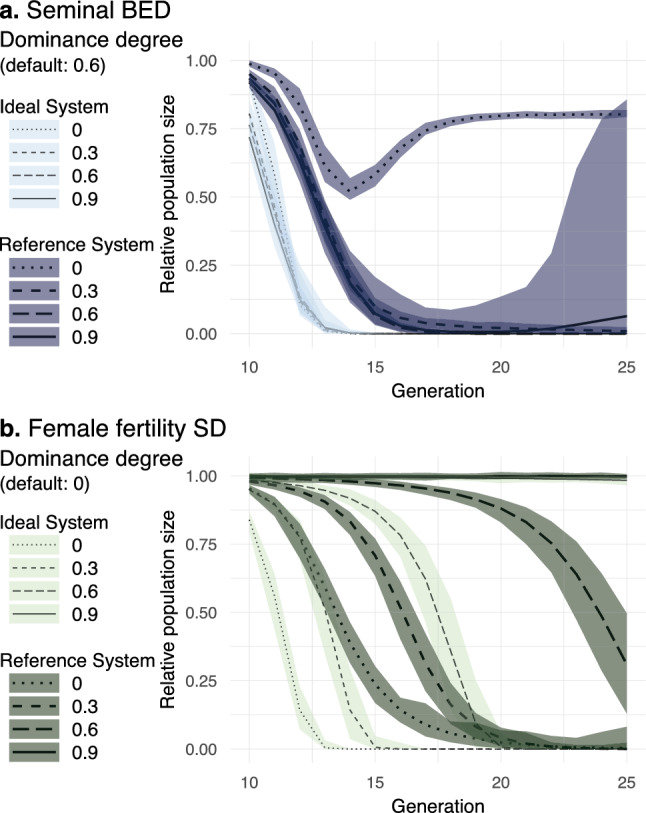
Impact of Dominance degree. Relative population size from generation 10 to 25 is shown with ideal and reference drive parameters for sBED (a) and ffSD (b) using in each case different values of Dominance degree. Lines represent the 100-simulations mean value while the colored envelopes stand for the maximum and minimum values.
Low-density growth rate
We refer to low-density growth rate (or Rm) as the expected number of adult female offspring produced per adult female in the absence of intraspecific competition. Higher Rm values indicate the production of greater offspring numbers per female so the population is harder to suppress and can quickly recover from the deleterious effects of the drive system. Therefore, the fast elimination rate is expected to decrease with Rm.
In our model, Rm can be adjusted with the Fecundity parameter (Table 1). Therefore, to evaluate the impact of Rm we explored the Fecundity parameter space. Our result is in accordance with previous findings on ffSD (e.g.,12) and also reveals that sBED may deal better with high-Rm populations (Fig. 5) which may represent the case of some pests (e.g.,16,37,41). While with ideal drive parameters both designs could in all cases eliminate populations with Rm of 24 in less than 36 generations, with reference drive parameters the fast elimination rate was still 100% for sBED but 0% for ffSD.
Figure 5.
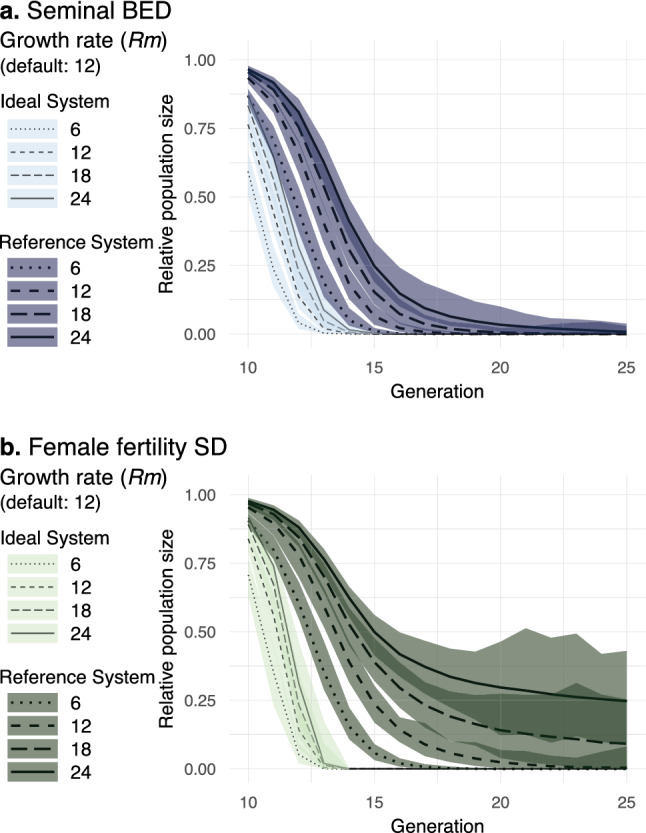
Impact of low-density growth rate. Relative population size from generation 10 to 25 is shown with ideal and reference drive parameters for sBED (a) and ffSD (b) using in each case different values of Rm. Rm was adjusted with the Fecundity parameter. Lines represent the 100-simulations mean value while the colored envelopes stand for the maximum and minimum values.
Resistance allele fitness
For homing to occur in CRISPR-Cas9-mediated gene drives, the double-stranded breaks induced by Cas9 nuclease must be resolved by the homology-directed repair pathway (HDR), which predominates specifically during early meiosis42. When the breaks are produced and resolved before or after the window optimal for HDR, nonhomologous end joining (NHEJ) prevails and the sequence of the target site is often changed so it is no longer recognized by the drive (e.g.,24). Frequently, NHEJ results in cleavage-resistant alleles (so-called resistance alleles) with disrupted gene function (also known as nonfunctional or r2 resistance alleles) that tend not to significantly affect the propagation of the drive allele since they will be selected out of the population13. Alternatively, if the resistance allele preserves the function of the target, it will possess an adaptive advantage over the drive allele and thus represents a great obstacle to the spread of the drive. The formation rate of these functional resistance alleles (also known as r1 resistance alleles) is rare and can be further reduced, for instance, by using multiple gRNAs (e.g.,13) or a conserved target gene that cannot tolerate mutations (e.g.,18).
In our models, resistance alleles were considered nonfunctional (r2) by default. However, given that r1 resistance alleles can restrict gene drive propagation43, we also assessed how our design is affected by the level of fitness reduction imposed by resistance alleles. To do so, we explored the performance of the systems assuming different degrees of Resistance allele functionality (see the parameter description in Table 2). Our results show that with reference drive parameters (which admits the formation of resistance alleles) sBED is more tolerant than ffSD to functional resistant alleles. While ffSD did not tolerate a Resistance allele functionality of 10%, sBED showed a fast elimination rate of 85% with as high as 40% functional resistance alleles (Fig. 6).
Figure 6.
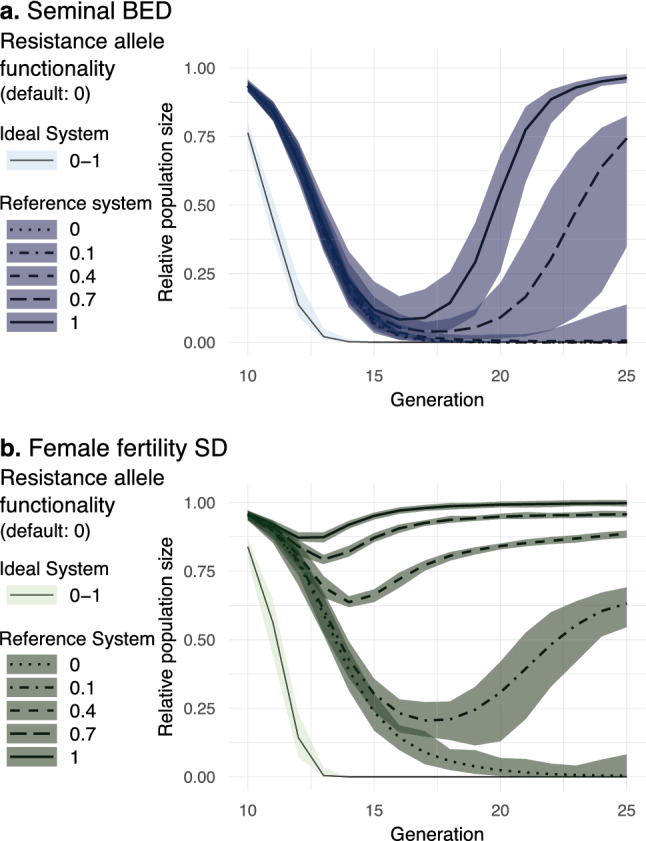
Impact of Resistance allele functionality. Relative population size from generation 10 to 25 is shown with ideal and reference drive parameters for sBED (a) and ffSD (b) using in each case different values of Resistance allele functionality. Lines represent the 100-simulations mean value while the colored envelopes stand for the maximum and minimum values.
Polyandry and terminator efficiency
By default, in our model, Polyandry degree was set to 3, which means that each adult female copulates with one, two, or, more likely, three males (Table 1). Each adult male can also copulate multiple times, but the number of times he copulates is randomly determined by panmixia. By reducing Polyandry degree in sBED designs with ideal drive parameters, the fast elimination rate was not affected, yet elimination was slightly delayed (Fig. 7a). In reference sBED designs, changing Polyandry degree from 3 to 2 resulted in a severe reduction of the fast elimination rate, from 100 to 1%. Nonetheless, in 78% of the cases, the population was eliminated between generations 36 and 60 (we did not simulate more generations). In the cases where the population was not eliminated by generation 60, it remained highly suppressed at a low size until the end of the simulated period (Fig. 7a). When Polyandry degree was reduced to 1, the fast elimination rate fell to 0%, which indicates that sBED interventions may not be able to completely suppress populations with monogamous females. However, the population size remained reduced at ~ 53% from generation 16 (Fig. 7, a), resembling the output of ffBED (Fig. 3a). In contrast, Polyandry degree did not affect the intervention outcome in ideal or reference ffSD designs (Fig. 7b). This is not surprising because ffBED affects female fertility irrespective of the number of mates.
Figure 7.
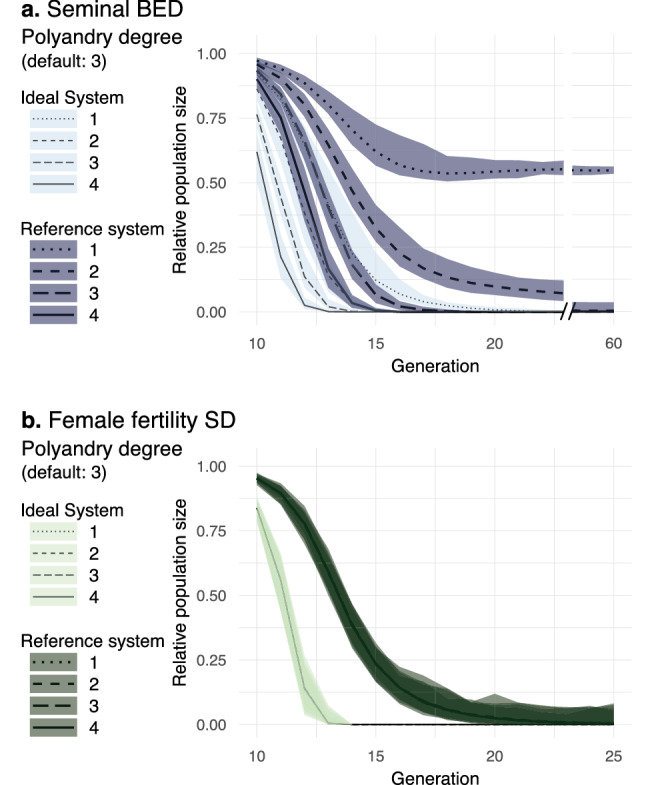
Impact of polyandry. Relative population size from generation 10 to 25 or 60 is shown with ideal and reference drive parameters for sBED (a) and ffSD (b) using in each case different values of Polyandry degree. Lines represent the 100-simulations mean value while the colored envelopes stand for the maximum and minimum values.
It can be expected that highly efficient transferred seminal toxins, whose action is modeled through the Terminator efficiency parameter (Table 2), may compensate for the effect of reduced polyandry. To evaluate this possibility, we estimated the fast elimination rate of the sBED with reference drive parameters under different combinations of Terminator efficiency and Polyandry degree (Fig. S3). We also repeated the simulations using different combinations of Critical population size and varying the number of released males per drive. Our results showed a poor compensation capacity of Terminator efficiency on the effect of Polyandry degree. Overall, a high fast elimination rate was only achieved when Terminator efficiency was 0.8 (or higher) and Polyandry degree was 3 (or higher) (Fig. S3).
Release size and critical population size
The number of released males per drive (release size) did not notably affect the simulation outcome of sBED (Fig. S3). Using reference parameters, when the number of released transgenic males was reduced from 500 to 100 (250–50 per drive), the fast elimination rate remained at 100%, though population elimination occurred, on average, 3.6 generations later. Furthermore, increasing the release size from 500 to 1000 accelerated elimination only by 1.1 generations on average. For the reference ffSD, the impact of the release size on the drive performance was also tenuous (Table S1). Diminishing the release size from 500 to 100 reduced the fast elimination rate from 97 to 91% and delayed elimination by 2.9 generations on average. Increasing the release size from 500 to 1000 did not impact the fast elimination rate and accelerated elimination only by 0.6 generations on average.
On the other hand, the impact of Critical population size (i.e., the population size of virgin adult males below which mate-finding Allee effect occurs) on suppression potential was higher for ffSD (Table S1) than for sBED (Fig. S3). For instance, varying Critical population size from 250 to 0 or 500 did not impact the fast elimination rate of the reference sBED (100%). For the reference ffSD, the fast elimination rate moved from 97 to 99% when Critical population size was raised from 250 to 500; but it fell to 85% when the parameter was reduced to 0, revealing a higher sensitivity of this design to Allee effects.
Discussion
Our results reveal that the use of BEDs to spread dominant or semidominant toxins has the potential for effective population suppression. Particularly, we show that the sBED design may be highly efficient and work relatively fast against polyandrous target populations. We identified, however, four key elements that may affect the feasibility of this approach (yet other restrictive elements may also have an important impact): (1) the challenge of generating effective seminal bioinsecticides, (2) the formation of functional resistance alleles, (3) the requirement that target females be polyandrous, and (4) the lack of panmixia in the target population.
The development of a sBED implies the design and generation of seminal toxins that do not affect the mating success of the male that produces it but kill or sterilize the target female. Currently, many effective bioinsecticides are bacteria-derived protoxin proteins known to be proteolytically activated upon ingestion by the insect midgut lumen proteases44. Interestingly, the female reproductive tract resembles the insect midgut as both tracts have a rich repertoire of secreted proteases, especially serine proteases. In the insect uterus, these proteases are responsible for processing male-derived peptides, degrading the mating plug, and protecting the female from microbes (e.g.,45–52). Though no protoxin has ever been evaluated inside the female reproductive tract, some insecticidal proteins may exhibit similar toxic effects therein. Also, given the cellular and physiological differences between the male accessory glands, where most seminal proteins are produced, and the uterus, it might be possible to design insecticidal proteins that will be activated solely within female tissues. Given the recent advances in knowledge of biological control agents53–56, the development of in silico tools to predict and scan molecular targets for pest control57, and the advances in computational protein design58–61, the engineering of seminal toxins may be attainable in the short term. However, the evolution of resistance to the toxins in the target population may always restrict insecticide-based approaches (reviewed in62).
Since population suppression mediated by BEDs relies on effector genes, another vulnerability of the system is the generation of loss-of-function mutation of the effector gene during HDR. While the mutated alleles will be innocuous, they will preserve the target function and be cleavage-resistant. Therefore, if the mutation rate is sufficiently high, these functional resistance alleles would spread rapidly, restricting the gene drive propagation.
Concerning polyandry in our model, the success of the sBED intervention requires that most females mate with more than one male before oviposition (Fig. 7) to allow each terminator male the opportunity to kill (or sterilize), on average, more than one female. Although multiple paternity is common among insects, the level of polyandry is highly variable between species29,30, with many species rarely remating (e.g.,63), therefore reducing the range of targets for which this control strategy would be effective. Moreover, even in polyandrous populations, sequential polyandry would reduce sBED effectiveness because females could mate with wt males and produce some offspring before mating with a terminator. Nonetheless, our model showed that with ideal drive parameters, which implies that terminator males do not undergo mating success reduction, even a monandrous population can be rapidly eliminated (Fig. 7). This suggests that low polyandrous populations can be efficiently suppressed as long as drive-carrying males are highly successful in achieving mating or drives' frequencies are kept high likely by releasing drive-carrying males in multiple bouts across several generations.
Mathematical modeling has shown that deviations from random mating may allow the clustering of free-drive individuals which, consequently, may thwart gene-drive-mediated extinction12,64. Population structure may have a particularly strong impact on the outcome of BEDs because they require that both drive components spread evenly across the population. For example, if one of the drive components is introduced in a given subpopulation several generations before the other, without imposing the intended fitness cost, it will probably spread rapidly. However, the second component will not have the chance of spreading alone because by the time it arrives in the subpopulation too many individuals will already carry the first component. Therefore the second component, and consequently the BED system, will not be able to fully invade that particular subpopulation. Resembling population dynamics of underdominance gene drive systems65, this scenario would potentially produce divergence among subpopulations in which different drives establish first because hybrid males will be terminators, incapable of producing any offspring. More complex simulations are necessary to help predict the outcome of BEDs in structured populations and how repeated releases of transgenic males may enhance BEDs' propagation.
The concept we present combines three biological tools that are versatile and have been proven to act in a wide range of taxa: artificial CRISPR-Cas9 nucleases66, artificial gene expression systems such as GAL4-UAS67, and insecticidal proteins such as Bt-toxins or spider venom peptides68. Therefore, transferring a BED design from one species to another would likely not require major adjustments in protocols. In addition, the development of insecticidal seminal proteins will provide a significant improvement to the Sterile Insect Technique, one common approach used to control insect pest populations69. The strategy consists of regular releases of sterile males unable to produce viable offspring so the target population is severely reduced or eliminated. However, female remating with wt males restricts the efficacy of this approach. Furthermore, females mated to sterile males may still attempt oviposition, which could lead to crop damage. But if instead, released males were not only sterile but also delivered seminal toxins to females during mating, target females could be killed right after copulation enhancing the efficacy of the control.
There is an ever-increasing need for ecologically conscious control of pest species that have a negative impact on health, agriculture, or the environment. Although future research on seminal protein engineering is warranted, our modeling indicates that BED designs might be an efficient strategy for biological control.
Methods
Model steps
Every generation, the number of mates for each adult virgin female is generated from a Poisson distribution truncated between 1 and Polyandry degree, with lambda equals two times Polyandry degree. Therefore, the Polyandry degree parameter defines the maximum and most likely number of mates per adult female. After these numbers are settled, male mates of each female are randomly sampled with replacement so a given male may mate with different females.
In sBED designs, if an adult male carrying both the transactivator and effector constructs (i.e., a terminator) successfully mates, the female then has a probability of dying after mating. This probability equals the Terminator efficiency if the terminator male is homozygous for both drives. However, an unintended dominant reproductive cost, which is adjusted for each drive, reduces the terminators' chances of successfully mating (see Unintended reproductive cost in Table 2). The Dominance degree parameter determines the fraction of the Terminator efficiency exhibited by heterozygous terminator males (see Table 2). Each surviving female, which is assumed to carry equal amounts of sperm from all her mates, will oviposit eggs. For simplicity, meiosis is only modeled for mothers and mating males. Both the probability of homing conversion and formation of cleavage-resistant alleles (so-called resistance alleles) are adjustable and restricted to the meiosis stage of drive-wt heterozygotes (see Homing efficiency and Resistance allele formation in Table 2).
Giving rise to the next generation, each mother produces a clutch of eggs, whose size is generated from a Poisson distribution with lambda equals Fecundity. However, according to the Unintended reproductive cost parameter, the fecundity of drive-carrying mothers is reduced by each drive separately. In ffBED designs, the fecundity of mothers that are homozygous for both drives (i.e., AABB mothers, where A and B are each of the gene drive loci) is further reduced according to the Intended fecundity cost parameter. The same occurs to homozygous drive-carrying mothers in ffSD designs (i.e., CC mothers, where C is the gene drive locus). The Dominance degree parameter determines the fraction of the intended fecundity cost that affects heterozygotes.
Eggs' genotypes are stochastically generated by sampling with replacement the haplotypes of the mother's gametes and the sperm she stores. According to the Unintended viability cost parameter, which defines a dominant viability cost imposed by each drive, drive-carrying eggs may have an added probability of dying before reaching the larval stage. Surviving eggs develop into larvae which are subject to density-independent and density-dependent mortality according to the Beverton-Holt model70, such that the probability of surviving is (Larval survival × N50)/(N × N50), where Larval survival is the density-independent larval-to-adult survival rate, N is the larval population size, and N50 is a constant defined as Carrying capacity/(Larval survival − 2/Fecundity). Each surviving larva results in a virgin adult female or male with equal probability.
Simulation process
At generation zero, the population size is in equilibrium at its carrying capacity with an equal number of females and males. Released males are homozygous (AAbb and aaBB) in BED designs while heterozygous (Cc) in SD designs, given that large amounts of homozygotes may be difficult to produce. At the virgin adult stage of every generation, if the number of males falls below a critical number (see Critical population size in Table 1), the population experiences a mate-finding Allee effect71, making mating opportunities less likely. Specifically, we modeled both the probability that virgin females achieve mating (and become mothers) and the number of mates per mother as positive sigmoid functions of the population size (for more details see Table 1). The population simulation ends (i.e. the population collapses) when no female becomes a mother, no mother produces viable eggs, or no larva survives to become an adult virgin female.
We created a tool in R62, binXdrives, to run the simulations. The tool is available at https://github.com/santiagorevale/binXdrives. See software versions in Table S2.
Supplementary Information
Acknowledgements
We thank Prof. Fred Gould and members of his lab for assistance during the initial stages of model designing. We also thank Prof. Jackson Champer and Silvina Belliard for their critical insights on earlier drafts of the manuscript. This work was supported by The National Scientific and Technical Research Council (Argentina). Computational procedures of this research were supported by the Wellcome Trust Core Award Grant Number 203141/Z/16/Z with additional support from the NIHR Oxford BRC. The views expressed are those of the authors and not necessarily those of the National Health Service, the NIHR, or the Department of Health.
Author contributions
J.H. conceived the original idea, designed the model, and took the lead in writing the manuscript. J.H. and S.R. wrote the code. S.R. optimized the code and run the simulations. L.M.M. was involved in planning the work and supervised the project. J.H. and L.M.M. wrote the manuscript. All authors discussed the results, and read and approved the final manuscript.
Data availability
The source code of binXdrives is available at https://github.com/santiagorevale/binXdrives and all simulation data generated in support of this research is available at https://github.com/santiagorevale/binXdrives/tree/main/simulations.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Juan Hurtado and Santiago Revale.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-022-10327-4.
References
- 1.Pyšek P, et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020;95:1511–1534. doi: 10.1111/brv.12627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Dara SK. The new integrated pest management paradigm for the modern age. J. Integr. Pest Manage. 2019;10:25. doi: 10.1093/jipm/pmz010. [DOI] [Google Scholar]
- 3.Burt A. Site-specific selfish genes as tools for the control and genetic engineering of natural populations. Proc. R. Soc. B Biol. Sci. 2003;270:921–928. doi: 10.1098/rspb.2002.2319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Burt A. Heritable strategies for controlling insect vectors of disease. Philos. Trans. R. Soc. B Biol. Sci. 2014;369:25. doi: 10.1098/rstb.2014.0217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Champer J, Buchman A, Akbari OS. Cheating evolution: Engineering gene drives to manipulate the fate of wild populations. Nat. Rev. Genet. 2016;17:146–159. doi: 10.1038/nrg.2015.34. [DOI] [PubMed] [Google Scholar]
- 6.Esvelt KM, Smidler AL, Catteruccia F, Church GM. Concerning RNA-guided gene drives for the alteration of wild populations. Elife. 2014;3:1–21. doi: 10.7554/eLife.03401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gantz VM, Bier E. The mutagenic chain reaction: A method for converting heterozygous to homozygous mutations. Science. 2015;348:442–444. doi: 10.1126/science.aaa5945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Godfray HCJ, North A, Burt A. How driving endonuclease genes can be used to combat pests and disease vectors. BMC Biol. 2017;15:1–12. doi: 10.1186/s12915-017-0420-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rode NO, Estoup A, Bourguet D, Courtier-Orgogozo V, Débarre F. Population management using gene drive: Molecular design, models of spread dynamics and assessment of ecological risks. Conserv. Genet. 2019;20:671–690. doi: 10.1007/s10592-019-01165-5. [DOI] [Google Scholar]
- 10.Gantz VM, et al. Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. Proc. Natl. Acad. Sci. USA. 2015;112:E6736–E6743. doi: 10.1073/pnas.1521077112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Beaghton AK, Hammond A, Nolan T, Crisanti A, Burt A. Gene drive for population genetic control: Non-functional resistance and parental effects. Proc. R. Soc. B Biol. Sci. 2019;286:20191586. doi: 10.1098/rspb.2019.1586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Champer J, Kim IK, Champer SE, Clark AG, Messer PW. Suppression gene drive in continuous space can result in unstable persistence of both drive and wild-type alleles. Mol. Ecol. 2021;30:1086–1101. doi: 10.1111/mec.15788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Champer SE, et al. Computational and experimental performance of CRISPR homing gene drive strategies with multiplexed gRNAs. Sci. Adv. 2020;6:eaaz0525. doi: 10.1126/sciadv.aaz0525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Deredec A, Burt A, Godfray HCJ. The population genetics of using homing endonuclease genes in vector and pest management. Genetics. 2008;179:2013–2026. doi: 10.1534/genetics.108.089037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Deredec A, Godfray HCJ, Burt A. Requirements for effective malaria control with homing endonuclease genes. Proc. Natl. Acad. Sci. USA. 2011;108:E874–E880. doi: 10.1073/pnas.1110717108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.North AR, Burt A, Godfray HCJ. Modelling the suppression of a malaria vector using a CRISPR-Cas9 gene drive to reduce female fertility. BMC Biol. 2020;18:1–14. doi: 10.1186/s12915-020-00834-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Prowse TAA, et al. Dodging silver bullets: Good CRISPR gene-drive design is critical for eradicating exotic vertebrates. Proc. R. Soc. B Biol. Sci. 2017;284:20170799. doi: 10.1098/rspb.2017.0799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kyrou K, et al. A CRISPR–Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nat. Biotechnol. 2018;36:1062–1071. doi: 10.1038/nbt.4245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Grunwald HA, et al. Super-Mendelian inheritance mediated by CRISPR–Cas9 in the female mouse germline. Nature. 2019;566:105–109. doi: 10.1038/s41586-019-0875-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hammond A, et al. A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nat. Biotechnol. 2016;34:78–83. doi: 10.1038/nbt.3439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Adelman Z, et al. Rules of the road for insect gene drive research and testing. Nat. Biotechnol. 2017;35:716–718. doi: 10.1038/nbt.3926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Akbari OS, et al. Safeguarding gene drive experiments in the laboratory. Science. 2015;349:927–929. doi: 10.1126/science.aac7932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Brossard D, Belluck P, Gould F, Wirz CD. Promises and perils of gene drives: Navigating the communication of complex, post-normal science. Proc. Natl. Acad. Sci. USA. 2019;116:7692–7697. doi: 10.1073/pnas.1805874115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Champer J, et al. Novel CRISPR/Cas9 gene drive constructs reveal insights into mechanisms of resistance allele formation and drive efficiency in genetically diverse populations. PLoS Genet. 2017;13:1–18. doi: 10.1371/journal.pgen.1006796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hammond A, et al. The creation and selection of mutations resistant to a gene drive over multiple generations in the malaria mosquito. PLoS Genet. 2017;13:1–16. doi: 10.1371/journal.pgen.1007039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hammond A, et al. Regulating the expression of gene drives is key to increasing their invasive potential and the mitigation of resistance. PLoS Genet. 2021;17:1–21. doi: 10.1371/journal.pgen.1009321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.KaramiNejadRanjbar M, et al. Consequences of resistance evolution in a Cas9-based sex conversion-suppression gene drive for insect pest management. Proc. Natl. Acad. Sci. USA. 2018;115:6189–6194. doi: 10.1073/pnas.1713825115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Price TAR, et al. Resistance to natural and synthetic gene drive systems. J. Evol. Biol. 2021;33:1345–1360. doi: 10.1111/jeb.13693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Taylor ML, Price TAR, Wedell N. Polyandry in nature: A global analysis. Trends Ecol. Evol. 2014;29:376–383. doi: 10.1016/j.tree.2014.04.005. [DOI] [PubMed] [Google Scholar]
- 30.Arnqvist G, Nilsson T. The evolution of polyandry: Multiple mating and female fitness in insects. Anim. Behav. 2000;60:145–164. doi: 10.1006/anbe.2000.1446. [DOI] [PubMed] [Google Scholar]
- 31.Hurtado J, Iglesias PP, Lipko P, Hasson E. Multiple paternity and sperm competition in the sibling species Drosophila buzzatii and Drosophila koepferae. Mol. Ecol. 2013;22:5016–5026. doi: 10.1111/mec.12436. [DOI] [PubMed] [Google Scholar]
- 32.Champer J, et al. Reducing resistance allele formation in CRISPR gene drive. Proc. Natl. Acad. Sci. USA. 2018;115:5522–5527. doi: 10.1073/pnas.1720354115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Terradas G, et al. Inherently confinable split-drive systems in Drosophila. Nat. Commun. 2021;12:1480. doi: 10.1038/s41467-021-21771-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Burt A, Deredec A. Self-limiting population genetic control with sex-linked genome editors. Proc. R. Soc. B Biol. Sci. 2018;285:25. doi: 10.1098/rspb.2018.0776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Willis K, Burt A. Double drives and private alleles for localised population genetic control. PLoS Genet. 2021;17:1–20. doi: 10.1371/journal.pgen.1009333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Beaghton A, Beaghton PJ, Burt A. Gene drive through a landscape: Reaction–diffusion models of population suppression and elimination by a sex ratio distorter. Theor. Popul. Biol. 2016;108:51–69. doi: 10.1016/j.tpb.2015.11.005. [DOI] [PubMed] [Google Scholar]
- 37.Rendon D, Buser J, Tait G, Lee JC, Walton VM. Survival and fecundity parameters of two Drosophila suzukii (Diptera: Drosophilidae) morphs on variable diet under suboptimal temperatures. J. Insect Sci. 2018;18:25. doi: 10.1093/jisesa/iey113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Buchman A, et al. Engineered resistance to Zika virus in transgenic Aedes aegypti expressing a polycistronic cluster of synthetic small RNAs. Proc. Natl. Acad. Sci. USA. 2019;116:3656–3661. doi: 10.1073/pnas.1810771116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Halfhill MD, Millwood RJ, Weissinger AK, Warwick SI, Stewart CN. Additive transgene expression and genetic introgression in multiple green-fluorescent protein transgenic crop x weed hybrid generations. Theor. Appl. Genet. 2003;107:1533–1540. doi: 10.1007/s00122-003-1397-7. [DOI] [PubMed] [Google Scholar]
- 40.Myers JL, et al. Mutants of the white ABCG transporter in Drosophila melanogaster have deficient olfactory learning and cholesterol homeostasis. Int. J. Mol. Sci. 2021;22:25. doi: 10.3390/ijms222312967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Hosseini-Tabesh B, Sahragard A, Karimi-Malati A. A laboratory and field condition comparison of life table parameters of Aphis gossypii Glover (Hemiptera: Aphididae) J. Plant Prot. Res. 2015;55:1–7. doi: 10.1515/jppr-2015-0001. [DOI] [Google Scholar]
- 42.Ranjha L, Howard SM, Cejka P. Main steps in DNA double-strand break repair: An introduction to homologous recombination and related processes. Chromosoma. 2018;127:187–214. doi: 10.1007/s00412-017-0658-1. [DOI] [PubMed] [Google Scholar]
- 43.Unckless RL, Clark AG, Messer PW. Evolution of resistance against CRISPR/Cas9 gene drive. Genetics. 2017;205:827–841. doi: 10.1534/genetics.116.197285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Palma L, et al. Bacillus thuringiensis toxins: An overview of their biocidal activity. Toxins (Basel) 2014;6:3296–3325. doi: 10.3390/toxins6123296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Al-Wathiqui N, Lewis SM, Dopman EB. Using RNA sequencing to characterize female reproductive genes between Z and E Strains of European Corn Borer moth (Ostrinia nubilalis) BMC Genom. 2014;15:1–13. doi: 10.1186/1471-2164-15-189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kelleher ES, Swanson WJ, Markow TA. Gene duplication and adaptive evolution of digestive proteases in Drosophila arizonae female reproductive tracts. PLoS Genet. 2007;3:1541–1549. doi: 10.1371/journal.pgen.0030148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Lawniczak MKN, et al. Mating and immunity in invertebrates. Trends Ecol. Evol. 2007;22:48–55. doi: 10.1016/j.tree.2006.09.012. [DOI] [PubMed] [Google Scholar]
- 48.Lawniczak MKN, Begun DJ. Molecular population genetics of female-expressed mating-induced serine proteases in Drosophila melanogaster. Mol. Biol. Evol. 2007;24:1944–1951. doi: 10.1093/molbev/msm122. [DOI] [PubMed] [Google Scholar]
- 49.McGraw LA, Gibson G, Clark AG, Wolfner MF. Genes regulated by mating, sperm, or seminal proteins in mated female Drosophila melanogaster. Curr. Biol. 2004;14:1509–1514. doi: 10.1016/j.cub.2004.08.028. [DOI] [PubMed] [Google Scholar]
- 50.Meslin C, et al. Digestive organ in the female reproductive tract borrows genes from multiple organ systems to adopt critical functions. Mol. Biol. Evol. 2015;32:1567–1580. doi: 10.1093/molbev/msv048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Plakke MS, Deutsch AB, Meslin C, Clark NL, Morehouse NI. Dynamic digestive physiology of a female reproductive organ in a polyandrous butterfly. J. Exp. Biol. 2015;218:1548–1555. doi: 10.1242/jeb.118323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Plakke MS, et al. Characterization of female reproductive proteases in a butterfly from functional and evolutionary perspectives. Physiol. Biochem. Zool. 2019;92:579–590. doi: 10.1086/705722. [DOI] [PubMed] [Google Scholar]
- 53.Paul S, Das S. Natural insecticidal proteins, the promising bio-control compounds for future crop protection. Nucleous. 2021;64:7–20. doi: 10.1007/s13237-020-00316-1. [DOI] [Google Scholar]
- 54.Liu L, et al. Identification and evaluations of novel insecticidal proteins from plants of the class polypodiopsida for crop protection against key lepidopteran pests. Toxins (Basel) 2019;11:1–25. doi: 10.3390/toxins11070383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.King GF. Tying pest insects in knots: The deployment of spider-venom-derived knottins as bioinsecticides. Pest Manage. Sci. 2019;75:2437–2445. doi: 10.1002/ps.5452. [DOI] [PubMed] [Google Scholar]
- 56.Kumar J, Ramlal A, Mallick D, Mishra V. An overview of some biopesticides and their importance in plant protection for commercial acceptance. Plants. 2021;10:1–15. doi: 10.3390/plants10061185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Agrawal P, Kumar S, Singh A, Raghava GPS, Singh IK. NeuroPIpred: A tool to predict, design and scan insect neuropeptides. Sci. Rep. 2019;9:1–12. doi: 10.1038/s41598-019-41538-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Simonson T, et al. Physics-based computational protein design: An update. J. Phys. Chem. A. 2020;124:10637–10648. doi: 10.1021/acs.jpca.0c07605. [DOI] [PubMed] [Google Scholar]
- 59.Zhou J, Panaitiu AE, Grigoryan G. A general-purpose protein design framework based on mining sequence-structure relationships in known protein structures. Proc. Natl. Acad. Sci. USA. 2020;117:1059–1068. doi: 10.1073/pnas.1908723117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Leman JK, et al. Macromolecular modeling and design in Rosetta: Recent methods and frameworks. Nat. Methods. 2020;17:665–680. doi: 10.1038/s41592-020-0848-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Lee ACL, Harris JL, Khanna KK, Hong JH. A comprehensive review on current advances in peptide drug development and design. Int. J. Mol. Sci. 2019;20:1–21. doi: 10.3390/ijms20102383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Siegwart M, et al. Resistance to bio-insecticides or how to enhance their sustainability: A review. Front. Plant Sci. 2015;6:1–19. doi: 10.3389/fpls.2015.00381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Richardson JB, Jameson SB, Gloria-Soria A, Wesson DM, Powell J. Evidence of limited polyandry in a natural population of Aedes aegypti. Am. J. Trop. Med. Hyg. 2015;93:189–193. doi: 10.4269/ajtmh.14-0718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Bull JJ, Remien CH, Krone SM. Gene-drive-mediated extinction is thwarted by population structure and evolution of sib mating. Evol. Med. Public Health. 2019;2019:66–81. doi: 10.1093/emph/eoz014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Champer J, Zhao J, Champer SE, Liu J, Messer PW. Population dynamics of underdominance gene drive systems in continuous space. ACS Synth. Biol. 2020;9:779–792. doi: 10.1021/acssynbio.9b00452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Adli M. The CRISPR tool kit for genome editing and beyond. Nat. Commun. 2018;9:1911. doi: 10.1038/s41467-018-04252-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. doi: 10.1242/dev.118.2.401. [DOI] [PubMed] [Google Scholar]
- 68.Saez NJ, Herzig V. Versatile spider venom peptides and their medical and agricultural applications. Toxicon. 2019;158:109–126. doi: 10.1016/j.toxicon.2018.11.298. [DOI] [PubMed] [Google Scholar]
- 69.McGraw EA, O’Neill SL. Beyond insecticides: New thinking on an ancient problem. Nat. Rev. Microbiol. 2013;11:181–193. doi: 10.1038/nrmicro2968. [DOI] [PubMed] [Google Scholar]
- 70.Beverton RJH, Holt SJ. On the dynamics of exploited fish populations. Fish. Investig. 1957;19:1–533. [Google Scholar]
- 71.Allee WC. Animal Aggregations. University of Chicago Press; 1931. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The source code of binXdrives is available at https://github.com/santiagorevale/binXdrives and all simulation data generated in support of this research is available at https://github.com/santiagorevale/binXdrives/tree/main/simulations.