

Abstract
Intracellular metabolites can cause critical changes in biological functions. Itaconate is perhaps the most fascinating substance in macrophages. Lipopolysaccharide can activate aconitate decarboxylase 1 and induces the generation of itaconate from the tricarboxylic acid cycle by decarboxylation of cis-aconitate. It has been reported that itaconate has beneficial effects on inflammation and oxidation. The mechanisms involved in these effects include the suppression of succinate dehydrogenase, the activation of nuclear factor E2-related factor 2 by alkylation of Kelch-like ECH-associated protein 1, suppression of aerobic glycolysis through regulation of glyceraldehyde-3-phosphate dehydrogenase and fructose-bisphosphate aldolase A, and suppression of IκBζ translation through activating transcription factor 3 activation. All of these findings elucidated the possible therapeutic implications of itaconate in inflammation-related diseases. In this review, we highlight that itaconate is a crucial molecule of the immunomodulatory response in macrophages and can regulate between immune response and cardiovascular metabolism. Furthermore, these discoveries suggest that itaconate is a very novel therapeutic molecule for the treatment of inflammation-related heart diseases.
Keywords: Activating transcription factor 3, Inflammation, Itaconate, Nuclear factor erythroid 2-related factor 2, Succinate dehydrogenase
INTRODUCTION
Intermediary metabolites are responsible for maintaining organ homeostasis, and their signaling functions are involved in the modulation of cell function [1]. Numerous studies have found that internal metabolites perform an important part in the progress of several diseases, such as Type 2 diabetes [2] cancers [3,4] atherosclerosis [5,6] cardiac disease [7,8] chronic kidney disease [9] and Alzheimer disease [10]. These natural chemical compounds are rather cheap and simple to manufacture in huge quantities. The most intriguing example of a metabolite with specific immunologic functions is itaconate, which was initially determined as an object during the distilled process of citric acid in 1836 and has been used broadly in the polymer industry for several years. Previous studies have found that itaconate acts as an antibacterial metabolite by inhibiting the activity of isocitrate lyase involved in the maintenance of the bacterial growth amid infection [11,12,13]. In addition, itaconate alleviates reperfusion injury through the suppression of succinate dehydrogenase (SDH) [14]. Itaconate increased the survival rate in mice models with traumatic brain injury and hemorrhagic shock [14]. It has been reported that itaconate inhibited the formation of abdominal aortic aneurysm (AAA) induced by angiotensin II in apolipoprotein E-deficient mice [15]. In addition, inhaled itaconate improves bleomycin-induced pulmonary fibrosis in mice [16]. Ho et al. found that the combination of antibiotic tobramycin with itaconate increases the Pseudomonas aeruginosa biofilm eradicating efficiency [17]. Based on the previous findings, suggesting that itaconate has potentials in the therapeutic treatment of various diseases in mice models. In this review, we emphasize on itaconate metabolism and itaconate regulation of inflammation-related cardiovascular diseases, giving a rationale for therapeutic applications in future.
ITACONATE SYNTHESIS AND METABOLISM
Itaconate displays a similar structure to succinate and malonate (SDH inhibitor). The metabolic process of itaconate is linked with the tricarboxylic acid (TCA) cycle. Itaconate is produced from cis-aconitate in the TCA cycle in macrophages activated with lipopolysaccharide (LPS), Toll-like receptor (TLR) ligands, and type I and type II interferons [18,19,20,21]. The expression of aconitate decarboxylase 1 (ACOD1), named immune-responsive gene 1 (IRG1) at first is upregulated by these stimuli. It was found that the increase in the production of itaconate is through upregulation of ACOD1 expression [Figure 1] [21]. These findings remarkably widen our comprehension of itaconate. In addition, the crystal structures of cis-ACOD (CAD, also known as ACOD1 or IRG1) were determined, with eight active sites and were important for CAD function [22]. Despite the fact that rare mutations were observed in the active center of CAD in humans, elucidation of the structure of CAD would provide insight into the investigation of CAD mutations and their relation to the pathological mechanism of disease and therapeutic strategy [22]. Furthermore, pyruvate dehydrogenase (PDH) is partially related to itaconate biosynthesis. PDH kinase 1 (PDK1) upregulates the phosphorylation of PDH and subsequently inhibits its activity [23]. LPS suppresses PDK1 activity, which results in increased switch of pyruvate to acetyl-CoA through activating PDH [24]. In addition, acetyl-CoA is a crucial precursor for citrate production, and enough citrate is necessary for itaconate biosynthesis. These biological reactions represent the metabolic cycle in macrophages. Other metabolic approaches for itaconate need further investigation.
Figure 1.

Itaconate biogenesis and metabolism. (a) Inflammatory stimuli activate aconitate decarboxylase 1 expression, which induces the production of itaconate by decarboxylation of cis-aconitate in the tricarboxylic acid cycle of the mitochondrial matrix. Citrate lyase subunit beta-like catalyzies citramalyl-CoA to pyruvate and acetyl-CoA. (b) Chemical structures of itaconate, dimethyl itaconate and 4-octyl itaconate
ITACONATE MODULATES INFLAMMATION BY SUPPRESSION OF SUCCINATE DEHYDROGENASE
SDH is a vital enzyme that converts succinate to fumarate in the TCA cycle. In addition, SDH oxidizes cumulative succinate to produce superfluous reduced coenzyme Q for the production of superoxide anion in mitochondrial [25]. Furthermore, the reactive oxygen species (ROS) triggers the inflammasomes, resulting in the secretion of pro-inflammatory modulators [26]. A previous study had found that itaconate was a competitive inhibitor of SDH [27]. The structural similarity between them accounts for its ability to inhibit SDH. It had been reported that SDH oxidizes succinate to generate ROS, which increases hypoxia-inducible factor 1α and finally, the interleukin (IL)-1β transcription in macrophages [28]. Moreover, pretreatment with dimethyl itaconate (DI) significantly reduces SDH activity, which consecutively inhibits ROS production, suppresses the activation of nod-like receptor protein 3 inflammasome, and decreases proinflammatory cytokines in mouse bone-marrow-derived macrophage (BMDM) cells [29]. In addition, ACOD1 overexpression in macrophages induces itaconate production that results in succinate accumulation, due to itaconate straightly suppresses SDH activity [Figure 2] [29,30]. In summary, inhibition of SDH is partially involved in itaconate-regulated inflammation.
Figure 2.

Itaconate suppresses inflammatory and oxidative signaling pathways. Itaconate is produced in macrophages activated by lipopolysaccharide through upregulating aconitate decarboxylase 1 expression. Increased itaconate activates the nuclear factor erythroid 2-related factor 2 signaling through alkylation of Kelch-like ECH-associated protein 1, which activates the transcription of HO-1 and glutathione. In addition, itaconate can suppress succinate dehydrogenase and decrease reactive oxygen species generation and interleukin-1β secretion. Itaconate increases activating transcription factor 3 expression, which directly suppresses IκBζ expression and results in reducing interleukin interleukin-6. Furthermore, itaconate promotes alkylation of glyceraldehyde-3-phosphate dehydrogenase and aldolase A to suppress glycolysis, thus alleviating the inflammation
ITACONATE REGULATES INFLAMMATION VIA THE NUCLEAR FACTOR ERYTHROID 2-RELATED FACTOR 2 PATHWAY
It is well-known that the nuclear factor erythroid 2-related factor 2 (NRF2) transcription factor plays an important role in the modulation of inflammation and oxidative stress [31,32,33]. NRF2 can bind to the promoters of IL-6 and IL-1β, and inhibit their transcription [34]. In addition, NRF2 activation upregulates HO-1 and modulates glutathione (GSH) production, which results in protecting against oxidative stress [35,36]. Under quiescent conditions, Kelch-like ECH-associated protein 1 (KEAP1) suppresses NRF2 activity in the cytoplasm, while NRF2 is dissociated from KEAP1 under the stimulus. NRF2 can then shift to the nucleus to trigger the anti-inflammatory and antioxidative pathways [37,38,39,40,41]. In addition, the alkylation of KEAP1 fails to suppress NRF2 [42]. Itaconate was found to enhance the alkylation of cysteine residues on KEAP1, which promotes KEAP1 degradation to further activate NRF2 [37]. 4-Octyl itaconate (4-OI) alleviated H2O2-activated ROS generation, cell death, and lipid oxidation in SH-SY5Y cells through the KEAP1-NRF2 pathway [43]. Itaconate was found to reduce cerebral ischemia/reperfusion injury through activation of the NRF2 signaling and inhibition of SDH activity [44]. In addition, DI decreased ROS production and malondialdehyde levels through NRF2/HO-1 signaling in doxorubicin-induced cardiotoxicity in mice [45]. 4-OI treatment ameliorated the survival rate and decreased the expression level of proinflammatory cytokines in the mouse model of LPS-induced sepsis [37]. Furthermore, Song et al. found that itaconate inhibited the formation of AAA induced by angiotensin II in apolipoprotein E-deficient mice. Mechanistically, itaconate suppressed vascular inflammation by allowing NRF2 to act as a repressor of downstream inflammation-related genes through KEAP1 alkylation. This suggested that therapeutic strategies to increase itaconate are feasibly valuable for the prevention of AAA formation [15]. These observations suggest that itaconate is attributed to activating the NRF2 signaling and the transcription of downstream antioxidant genes [Figure 2].
ITACONATE MODULATES INFLAMMATION THROUGH THE ACTIVATING TRANSCRIPTION FACTOR 3 AND IκBζ PATHWAY
Exposure of macrophages to LPS has been widely utilized as a model of inflammation. LPS activates the inflammatory pathways by binding to TLRs on the surfaces of macrophages. In addition, LPS stimulates the production of tumour necrosis factor-alpha (TNF-α) and IL-6 [46]. TNF-α is induced by nuclear factor-κB during LPS treatment [47] while IL-6 is induced by IκBζ pathway [48]. The transcription factor IκBζ is an ankyrin-repeat-containing nuclear protein [48]. It was found that knockout of Nfkbiz decreases IL-6 production in peritoneal macrophages after the treatment with TLR ligands and IL-1 [49]. The ablation of IκBζ caused a reduction in IL-6 production in mouse macrophages treated with LPS [50]. Kim et al. had found that activating transcription factor 3 (ATF3) deficiency in mouse embryonic fibroblasts increases IκBζ expression and stimulates the secretion of proinflammatory cytokines [51]. Furthermore, Bambouskova et al. had found that DI was less capable to downregulate LPS-induced IκBζ expression in BMDMs of ATF3 KO mice [52]. Furthermore, the upregulation of ATF3 by DI treatment inactivated α subunit of eukaryotic initiation factor 2α, which further suppressed IκBζ transcription [52]. Furthermore, treatment with antioxidant or GSH could abolish the effect of DI on IL-6 expression [52]. This suggested that GSH/ROS pathway is involved in the modulation of DI-regulated inflammation [52]. In addition, it was noted that NRF2 is involved in the suppression of macrophage inflammation by 4-OI [37]. These findings suggest that targeting the itaconate/IκBζ modulatory axis could be a novel strategy for the treatment of inflammatory diseases [Figure 2].
ITACONATE MODULATES INFLAMMATION BY SUPPRESSING GLYCOLYSIS VIA TARGETING GLYCERALDEHYDE-3-PHOSPHATE DEHYDROGENASE
Glycolytic pathway acts as an important role in cell growth, differentiation, and phenotype shifts in macrophages [53]. LPS treatment results in the increment of glycolysis. The immune phenotypes of macrophages are related to their metabolic statuses [53]. Macrophages can be classified into two types: The M1 phenotype activated by LPS has proinflammatory property, and the M2 phenotype activated by IL-4 and IL-10 displays anti-inflammatory features [54]. In addition, itaconate regulation of macrophage polarization is yet controversial. Qin et al. had found that the anti-inflammatory effect of itaconate is mediated by subsiding glycolysis through fructose-bisphosphate aldolase A (ALDOA, a glycolytic enzyme) suppression [55]. 4-OI induced alkylation of glyceraldehyde-3-phosphate dehydrogenase (GAPDH), which suppressed glycolysis and the generation of cytokines in LPS-stimulated macrophages [56]. Furthermore, the anti-inflammatory effect exerted by 4-OI was suppressed by a high concentration of glucose, thus implying that 4-OI suppresses inflammation via the inhibition of glycolysis [56]. In contrast, one study found that microRNA-93 downregulates ACOD1expression and itaconate production to trigger M2 polarization, probably due to less itaconate for enhancing oxidative phosphorylation [57]. In summary, these findings suggest that itaconate modulates the inflammation and cell polarization of macrophage in distinct metabolic statuses [Figure 2].
ITACONATE MAY ACT AS A POSSIBLE THERAPEUTIC MOLECULE IN HEART DISEASE VIA ACTIVATING TRANSCRIPTION FACTOR 3 SIGNALING
DI has been reported to induce ATF3 expression [52]. ATF3 modulation of cardiac function varies depending on the stress patterns. ATF3 deficient mice show decreased cardiac remodeling and hypertrophy after phenylephrine treatment [58,59]. Ectopic ATF3 expression in cardiomyocytes induces cardiac dysfunction in transgenic mice [58,60]. On the contrary, our previous study showed that ATF3 knockout showed a loss of normal hypertrophic remodeling after transaortic banding treatment [61]. Some studies showed that ATF3 plays a beneficial role in the mice model of transverse aortic constriction [62,63]. The effect of ATF3 in cardiac remodeling is still inconclusive and worth further exploring [64,65]. Cardiac ATF3 deficient mice show worse cardiac remodeling and cardiac dysfunction after a high-fat diet [66]. Furthermore, Song et al. found that itaconate inhibited the AAA formation induced by angiotensin II in apolipoprotein E-deficient mice. This suggested that therapeutic strategies to increase itaconate are feasibly valuable for the prevention of AAA formation [15]. Further studies are needed to elucidate whether itaconate is a therapeutic molecule in heart diseases via ATF3 signaling pathways.
FUTURE APPLICATION OF ITACONATE
Itaconate was reported to regulate several signaling pathways, such as NRF2 and ATF3 for its beneficial role in anti-inflammation and anti-oxidation. It has been known that NRF2 activator has a therapeutic effect in the treatment of several inflammation-related diseases. Dimethyl fumarate had been applied in the clinical treatment of multiple sclerosis via NRF2 activation [67]. Furthermore, itaconate is less toxic for therapeutic application in treating inflammation-related diseases. However, there is still no obvious proof of the outcomes of the in vivo eradication of itaconate. In addition, other pathways may be involved in itaconate-regulating cell function. Citrate lyase subunit beta-like (CLYBL) is localized in the mitochondria. Mutations in CLYBL cause decreased circulating levels of vitamin B12 [68,69]. Itaconate is converted to itaconyl-CoA and then to citramalyl-CoA catabolized by CLYBL to produce pyruvate and acetyl-CoA [70]. Knockout of CLYBL increases the accumulation of itaconyl-CoA, which results in vitamin B12 degradation. It is still obscure whether Vitamin B12 inactivation influences the activity of itaconate and whether CLYBL modulates itaconate biosynthesis during the inflammation response [71]; therefore, the utilization of itaconate as a treatment agent needs a cautious investigation. Furthermore, it is interesting to analyze whether circulating levels of itaconate could be utilized as a biomarker for inflammation-related diseases. In addition, treatment of inhaled itaconate improves bleomycin-induced pulmonary fibrosis in mice, whereas pulmonary fibrosis worsening in the ACOD1 KO mice, implying that directly targeting ACOD1/itaconate and pharmacological applications using itaconate may potentially act as anti-fibrotic agents for preclinical use in pulmonary fibrosis [16]. Ho et al. found that itaconate increases the efficacy of tobramycin against P. aeruginosa biofilms. Combination of antibiotic tobramycin with an anti-inflammatory compound itaconate (molar ratio [tobramycin]: [itaconate] of 1:5) increases the P. aeruginosa biofilm eradicating efficiency of tobramycin four-fold compared to the usage of tobramycin alone, suggesting that combination of tobramycin and itaconate may be plausible in preclinical models of P. aeruginosa biofilm infections [17].
CONCLUSION
Itaconate has received considerable attention due to its anti-inflammatory and anti-oxidative effects. In addition, itaconate acts as an interesting link between metabolism and immune response in the cells for elucidating the pathogenesis of inflammation-related diseases. At present, the molecular mechanisms of itaconate for anti-inflammation and anti-oxidation have been elucidated, including the suppression of SDH, activation of NRF2 by liberating from KEAP1, upregulation of ATF3 to inhibit the IκBζ activation, and inhibition of glycolysis through GAPDH alkylation and suppression of ALDOA. In addition, itaconate can reprogram macrophages into the M2 phenotype. Itaconate and its derivatives also can induce electrophilic stress in the immunomodulating process. These new findings suggest that itaconate acts as a very suitable therapeutic molecule for inflammation-related heart diseases. The immunomodulatory mechanisms of itaconate need to be fully determined before clinical experiments. A summary of the related pathways involved in itaconate regulation of inflammation is depicted in Figure 3. It is concluded that itaconate is a very novel therapeutic molecule for the treatment of inflammation-related diseases and has the potential for use as a preclinical drug in inflammation-related heart disease based on the in vitro and in vivo findings.
Figure 3.
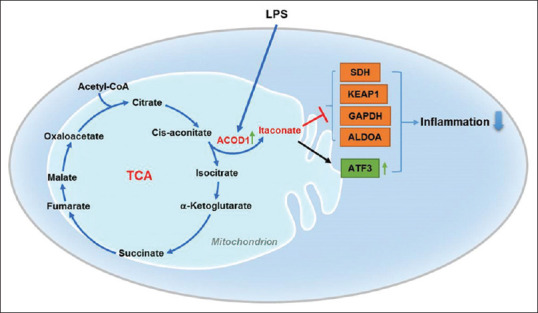
The summarized graphical abstract of related pathways involved in itaconate regulation of inflammation. Lipopolysaccharide can activate aconitate decarboxylase 1 and induces the production of itaconate from the tricarboxylic acid cycle. Itaconate exerts anti-inflammatory effects by suppression of succinate dehydrogenase, Kelch-like ECH-associated protein 1, glyceraldehyde-3-phosphate dehydrogenase, and aldolase A, but activation of activating transcription factor 3
Financial support and sponsorship
This work was support by Ministry of Science and Technology (MOST 107-2314-B-303-015-MY3 to C-FC), Buddhist Tzu Chi Medical Foundation (TCMMP108-04-02 to C-FC), and Tzu Chi and Academia Sinica cooperation (TCAS-108-01 translational research grants to C-FC).
Conflicts of interest
Dr. Ching-Feng Cheng, an editorial board member at Tzu Chi Medical Journal, had no role in the peer review process of or decision to publish this article. The other authors declared no conflicts of interest in writing this paper.
REFERENCES
- 1.Ferreira LM, Li AM, Serafim TL, Sobral MC, Alpoim MC, Urbano AM. Intermediary metabolism: An intricate network at the crossroads of cell fate and function. Biochim Biophys Acta Mol Basis Dis. 2020;1866:165887. doi: 10.1016/j.bbadis.2020.165887. [DOI] [PubMed] [Google Scholar]
- 2.Moraru A, Wiederstein J, Pfaff D, Fleming T, Miller AK, Nawroth P, et al. Elevated levels of the reactive metabolite methylglyoxal recapitulate progression of type 2 diabetes. Cell Metab. 2018;27:926–34. doi: 10.1016/j.cmet.2018.02.003. e8. [DOI] [PubMed] [Google Scholar]
- 3.Lee N, Spears ME, Carlisle AE, Kim D. Endogenous toxic metabolites and implications in cancer therapy. Oncogene. 2020;39:5709–20. doi: 10.1038/s41388-020-01395-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wei Q, Qian Y, Yu J, Wong CC. Metabolic rewiring in the promotion of cancer metastasis: Mechanisms and therapeutic implications. Oncogene. 2020;39:6139–56. doi: 10.1038/s41388-020-01432-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Goonewardena SN, Prevette LE, Desai AA. Metabolomics and atherosclerosis. Curr Atheroscler Rep. 2010;12:267–72. doi: 10.1007/s11883-010-0112-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Iida M, Harada S, Takebayashi T. Application of metabolomics to epidemiological studies of atherosclerosis and cardiovascular disease. J Atheroscler Thromb. 2019;26:747–57. doi: 10.5551/jat.RV17036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mervaala E, Biala A, Merasto S, Lempiäinen J, Mattila I, Martonen E, et al. Metabolomics in angiotensin II-induced cardiac hypertrophy. Hypertension. 2010;55:508–15. doi: 10.1161/HYPERTENSIONAHA.109.145490. [DOI] [PubMed] [Google Scholar]
- 8.Griffin JL, Atherton H, Shockcor J, Atzori L. Metabolomics as a tool for cardiac research. Nat Rev Cardiol. 2011;8:630–43. doi: 10.1038/nrcardio.2011.138. [DOI] [PubMed] [Google Scholar]
- 9.Chen DQ, Cao G, Chen H, Argyopoulos CP, Yu H, Su W, et al. Identification of serum metabolites associating with chronic kidney disease progression and anti-fibrotic effect of 5-methoxytryptophan. Nat Commun. 2019;10:1476. doi: 10.1038/s41467-019-09329-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Varma VR, Oommen AM, Varma S, Casanova R, An Y, Andrews RM, et al. Brain and blood metabolite signatures of pathology and progression in Alzheimer disease: A targeted metabolomics study. PLoS Med. 2018;15:e1002482. doi: 10.1371/journal.pmed.1002482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.McFadden BA, Purohit S. Itaconate, an isocitrate lyase-directed inhibitor in Pseudomonas indigofera. J Bacteriol. 1977;131:136–44. doi: 10.1128/jb.131.1.136-144.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rittenhouse JW, McFadden BA. Inhibition of isocitrate lyase from Pseudomonas indigofera by itaconate. Arch Biochem Biophys. 1974;163:79–86. doi: 10.1016/0003-9861(74)90456-1. [DOI] [PubMed] [Google Scholar]
- 13.Stocks CJ, Schembri MA, Sweet MJ, Kapetanovic R. For when bacterial infections persist: Toll-like receptor-inducible direct antimicrobial pathways in macrophages. J Leukoc Biol. 2018;103:35–51. doi: 10.1002/JLB.4RI0917-358R. [DOI] [PubMed] [Google Scholar]
- 14.Cordes T, Lucas A, Divakaruni AS, Murphy AN, Cabrales P, Metallo CM. Itaconate modulates tricarboxylic acid and redox metabolism to mitigate reperfusion injury. Mol Metab. 2020;32:122–35. doi: 10.1016/j.molmet.2019.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Song H, Xu T, Feng X, Lai Y, Yang Y, Zheng H, et al. Itaconate prevents abdominal aortic aneurysm formation through inhibiting inflammation via activation of Nrf2. EBioMedicine. 2020;57:102832. doi: 10.1016/j.ebiom.2020.102832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ogger PP, Albers GJ, Hewitt RJ, O'Sullivan BJ, Powell JE, Calamita E, et al. Itaconate controls the severity of pulmonary fibrosis. Sci Immunol. 2020;5:eabc1884. doi: 10.1126/sciimmunol.abc1884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ho DK, De Rossi C, Loretz B, Murgia X, Lehr CM. Itaconic Acid Increases the Efficacy of Tobramycin against Pseudomonas aeruginosa Biofilms. Pharmaceutics. 2020;12:691. doi: 10.3390/pharmaceutics12080691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Shin JH, Yang JY, Jeon BY, Yoon YJ, Cho SN, Kang YH, et al. (1) H NMR-based metabolomic profiling in mice infected with Mycobacterium tuberculosis. J Proteome Res. 2011;10:2238–47. doi: 10.1021/pr101054m. [DOI] [PubMed] [Google Scholar]
- 19.Strelko CL, Lu W, Dufort FJ, Seyfried TN, Chiles TC, Rabinowitz JD, et al. Itaconic acid is a mammalian metabolite induced during macrophage activation. J Am Chem Soc. 2011;133:16386–9. doi: 10.1021/ja2070889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sugimoto M, Sakagami H, Yokote Y, Onuma H, Kaneko M, Mori M, et al. Non-targeted metabolite profiling in activated macrophage secretion. Metabolomics. 2012;8:624–33. [Google Scholar]
- 21.Michelucci A, Cordes T, Ghelfi J, Pailot A, Reiling N, Goldmann O, et al. Immune-responsive gene 1 protein links metabolism to immunity by catalyzing itaconic acid production. Proc Natl Acad Sci U S A. 2013;110:7820–5. doi: 10.1073/pnas.1218599110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chen F, Lukat P, Iqbal AA, Saile K, Kaever V, van den Heuvel J, et al. Crystal structure of cis-aconitate decarboxylase reveals the impact of naturally occurring human mutations on itaconate synthesis. Proc Natl Acad Sci U S A. 2019;116:20644–54. doi: 10.1073/pnas.1908770116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yu XH, Zhang DW, Zheng XL, Tang CK. Itaconate: An emerging determinant of inflammation in activated macrophages. Immunol Cell Biol. 2019;97:134–41. doi: 10.1111/imcb.12218. [DOI] [PubMed] [Google Scholar]
- 24.Meiser J, Krämer L, Sapcariu SC, Battello N, Ghelfi J, D'Herouel AF, et al. Pro-inflammatory macrophages sustain pyruvate oxidation through pyruvate dehydrogenase for the synthesis of itaconate and to enable cytokine expression. J Biol Chem. 2016;291:3932–46. doi: 10.1074/jbc.M115.676817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Murphy MP. How mitochondria produce reactive oxygen species. Biochem J. 2009;417:1–13. doi: 10.1042/BJ20081386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bauernfeind F, Ablasser A, Bartok E, Kim S, Schmid-Burgk J, Cavlar T, et al. Inflammasomes: Current understanding and open questions. Cell Mol Life Sci. 2011;68:765–83. doi: 10.1007/s00018-010-0567-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ackermann WW, Potter VR. Enzyme inhibition in relation to chemotherapy. Proc Soc Exp Biol Med. 1949;72:1–9. doi: 10.3181/00379727-72-17313. [DOI] [PubMed] [Google Scholar]
- 28.Tannahill GM, Curtis AM, Adamik J, Palsson-McDermott EM, McGettrick AF, Goel G, et al. Succinate is an inflammatory signal that induces IL-1β through HIF-1α. Nature. 2013;496:238–42. doi: 10.1038/nature11986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lampropoulou V, Sergushichev A, Bambouskova M, Nair S, Vincent EE, Loginicheva E, et al. Itaconate links inhibition of succinate dehydrogenase with macrophage metabolic remodeling and regulation of inflammation. Cell Metab. 2016;24:158–66. doi: 10.1016/j.cmet.2016.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Cordes T, Wallace M, Michelucci A, Divakaruni AS, Sapcariu SC, Sousa C, et al. Immunoresponsive gene 1 and itaconate inhibit succinate dehydrogenase to modulate intracellular succinate levels. J Biol Chem. 2016;291:14274–84. doi: 10.1074/jbc.M115.685792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Tocmo R, Parkin K. S-1-propenylmercaptocysteine protects murine hepatocytes against oxidative stress via persulfidation of Keap1 and activation of Nrf2. Free Radic Biol Med. 2019;143:164–75. doi: 10.1016/j.freeradbiomed.2019.07.022. [DOI] [PubMed] [Google Scholar]
- 32.Shao Y, Yu H, Yang Y, Li M, Hang L, Xu X. A solid dispersion of quercetin shows enhanced nrf2 activation and protective effects against oxidative injury in a mouse model of dry age-related macular degeneration. Oxid Med Cell Longev. 2019;2019:1479571. doi: 10.1155/2019/1479571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wang R, Luo Y, Lu Y, Wang D, Wang T, Pu W, et al. Maggot extracts alleviate inflammation and oxidative stress in acute experimental colitis via the activation of nrf2. Oxid Med Cell Longev. 2019;2019:4703253. doi: 10.1155/2019/4703253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kobayashi EH, Suzuki T, Funayama R, Nagashima T, Hayashi M, Sekine H, et al. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat Commun. 2016;7:11624. doi: 10.1038/ncomms11624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Yang YC, Lii CK, Lin AH, Yeh YW, Yao HT, Li CC, et al. Induction of glutathione synthesis and heme oxygenase 1 by the flavonoids butein and phloretin is mediated through the ERK/Nrf2 pathway and protects against oxidative stress. Free Radic Biol Med. 2011;51:2073–81. doi: 10.1016/j.freeradbiomed.2011.09.007. [DOI] [PubMed] [Google Scholar]
- 36.Ma Q. Role of nrf2 in oxidative stress and toxicity. Annu Rev Pharmacol Toxicol. 2013;53:401–26. doi: 10.1146/annurev-pharmtox-011112-140320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Mills EL, Ryan DG, Prag HA, Dikovskaya D, Menon D, Zaslona Z, et al. Itaconate is an anti-inflammatory metabolite that activates Nrf2 via alkylation of KEAP1. Nature. 2018;556:113–7. doi: 10.1038/nature25986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Cui G, Li L, Xu W, Wang M, Jiao D, Yao B, et al. Astaxanthin protects ochratoxin A-induced oxidative stress and apoptosis in the heart via the nrf2 pathway. Oxid Med Cell Longev. 2020;2020:7639109. doi: 10.1155/2020/7639109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Han D, Gu X, Gao J, Wang Z, Liu G, Barkema HW, et al. Chlorogenic acid promotes the Nrf2/HO-1 anti-oxidative pathway by activating p21Waf1/Cip1 to resist dexamethasone-induced apoptosis in osteoblastic cells. Free Radic Biol Med. 2019;137:1–12. doi: 10.1016/j.freeradbiomed.2019.04.014. [DOI] [PubMed] [Google Scholar]
- 40.Bellezza I, Giambanco I, Minelli A, Donato R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim Biophys Acta Mol Cell Res. 2018;1865:721–33. doi: 10.1016/j.bbamcr.2018.02.010. [DOI] [PubMed] [Google Scholar]
- 41.Yamamoto M, Kensler TW, Motohashi H. The KEAP1-NRF2 system: A thiol-based sensor-effector apparatus for maintaining redox homeostasis. Physiol Rev. 2018;98:1169–203. doi: 10.1152/physrev.00023.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hayes JD, Dinkova-Kostova AT. The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem Sci. 2014;39:199–218. doi: 10.1016/j.tibs.2014.02.002. [DOI] [PubMed] [Google Scholar]
- 43.Liu H, Feng Y, Xu M, Yang J, Wang Z, Di G. Four-octyl itaconate activates Keap1-Nrf2 signaling to protect neuronal cells from hydrogen peroxide. Cell Commun Signal. 2018;16:81. doi: 10.1186/s12964-018-0294-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Michaličková D, Hrnčíř T, Canová NK, Slanař O. Targeting Keap1/Nrf2/ARE signaling pathway in multiple sclerosis. Eur J Pharmacol. 2020;873:172973. doi: 10.1016/j.ejphar.2020.172973. [DOI] [PubMed] [Google Scholar]
- 45.Shan Q, Li X, Zheng M, Lin X, Lu G, Su D, et al. Protective effects of dimethyl itaconate in mice acute cardiotoxicity induced by doxorubicin. Biochem Biophys Res Commun. 2019;517:538–44. doi: 10.1016/j.bbrc.2019.07.046. [DOI] [PubMed] [Google Scholar]
- 46.Ye Y, Wang Y, Yang Y, Tao L. Aloperine suppresses LPS-induced macrophage activation through inhibiting the TLR4/NF-κB pathway. Inflamm Res. 2020;69:375–83. doi: 10.1007/s00011-019-01313-0. [DOI] [PubMed] [Google Scholar]
- 47.Mazgaeen L, Gurung P. Recent advances in lipopolysaccharide recognition systems. Int J Mol Sci. 2020;21:379. doi: 10.3390/ijms21020379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Annemann M, Plaza-Sirvent C, Schuster M, Katsoulis-Dimitriou K, Kliche S, Schraven B, et al. Atypical IκB proteins in immune cell differentiation and function. Immunol Lett. 2016;171:26–35. doi: 10.1016/j.imlet.2016.01.006. [DOI] [PubMed] [Google Scholar]
- 49.Yamamoto M, Yamazaki S, Uematsu S, Sato S, Hemmi H, Hoshino K, et al. Regulation of Toll/IL-1-receptor-mediated gene expression by the inducible nuclear protein IkappaBzeta. Nature. 2004;430:218–22. doi: 10.1038/nature02738. [DOI] [PubMed] [Google Scholar]
- 50.Kitamura H, Kanehira K, Okita K, Morimatsu M, Saito M. MAIL, a novel nuclear I kappa B protein that potentiates LPS-induced IL-6 production. FEBS Lett. 2000;485:53–6. doi: 10.1016/s0014-5793(00)02185-2. [DOI] [PubMed] [Google Scholar]
- 51.Kim EY, Shin HY, Kim JY, Kim DG, Choi YM, Kwon HK, et al. ATF3 plays a key role in Kdo2-lipid A-induced TLR4-dependent gene expression via NF-κB activation. PLoS One. 2010;5:e14181. doi: 10.1371/journal.pone.0014181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Bambouskova M, Gorvel L, Lampropoulou V, Sergushichev A, Loginicheva E, Johnson K, et al. Electrophilic properties of itaconate and derivatives regulate the IκBζ-ATF3 inflammatory axis. Nature. 2018;556:501–4. doi: 10.1038/s41586-018-0052-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Van den Bossche J, Baardman J, de Winther MP. Metabolic characterization of polarized M1 and M2 bone marrow-derived macrophages using real-time extracellular flux analysis. J Vis Exp. 2015;105:53424. doi: 10.3791/53424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Mills CD. Anatomy of a discovery: M1 and m2 macrophages. Front Immunol. 2015;6:212. doi: 10.3389/fimmu.2015.00212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Qin W, Qin K, Zhang Y, Jia W, Chen Y, Cheng B, et al. S-glycosylation-based cysteine profiling reveals regulation of glycolysis by itaconate. Nat Chem Biol. 2019;15:983–91. doi: 10.1038/s41589-019-0323-5. [DOI] [PubMed] [Google Scholar]
- 56.Liao ST, Han C, Xu DQ, Fu XW, Wang JS, Kong LY. 4-Octyl itaconate inhibits aerobic glycolysis by targeting GAPDH to exert anti-inflammatory effects. Nat Commun. 2019;10:5091. doi: 10.1038/s41467-019-13078-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ganta VC, Choi MH, Kutateladze A, Fox TE, Farber CR, Annex BH. A MicroRNA93-interferon regulatory factor-9-immunoresponsive gene-1-itaconic acid pathway modulates M2-like macrophage polarization to revascularize ischemic muscle. Circulation. 2017;135:2403–25. doi: 10.1161/CIRCULATIONAHA.116.025490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Koren L, Elhanani O, Kehat I, Hai T, Aronheim A. Adult cardiac expression of the activating transcription factor 3, ATF3, promotes ventricular hypertrophy. PLoS One. 2013;8:e68396. doi: 10.1371/journal.pone.0068396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Koren L, Alishekevitz D, Elhanani O, Nevelsky A, Hai T, Kehat I, et al. ATF3-dependent cross-talk between cardiomyocytes and macrophages promotes cardiac maladaptive remodeling. Int J Cardiol. 2015;198:232–40. doi: 10.1016/j.ijcard.2015.06.099. [DOI] [PubMed] [Google Scholar]
- 60.Okamoto Y, Chaves A, Chen J, Kelley R, Jones K, Weed HG, et al. Transgenic mice with cardiac-specific expression of activating transcription factor 3, a stress-inducible gene, have conduction abnormalities and contractile dysfunction. Am J Pathol. 2001;159:639–50. doi: 10.1016/S0002-9440(10)61735-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Lin H, Li HF, Chen HH, Lai PF, Juan SH, Chen JJ, et al. Activating transcription factor 3 protects against pressure-overload heart failure via the autophagy molecule Beclin-1 pathway. Mol Pharmacol. 2014;85:682–91. doi: 10.1124/mol.113.090092. [DOI] [PubMed] [Google Scholar]
- 62.Zhou H, Shen DF, Bian ZY, Zong J, Deng W, Zhang Y, et al. Activating transcription factor 3 deficiency promotes cardiac hypertrophy, dysfunction, and fibrosis induced by pressure overload. PLoS One. 2011;6:e26744. doi: 10.1371/journal.pone.0026744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Zhou H, Guo H, Zong J, Dai J, Yuan Y, Bian ZY, et al. ATF3 regulates multiple targets and may play a dual role in cardiac hypertrophy and injury. Int J Cardiol. 2014;174:838–9. doi: 10.1016/j.ijcard.2014.04.160. [DOI] [PubMed] [Google Scholar]
- 64.Yang J, Yang CJ, Yang J, Fan ZX. ATF3: A promotion effect or a inhibition effect in cardiac maladaptive remodeling. Int J Cardiol. 2015;201:245–6. doi: 10.1016/j.ijcard.2015.08.079. [DOI] [PubMed] [Google Scholar]
- 65.Koren L, Shaked Y, Aronheim A. Response letter: “ATF3: A promoter or inhibitor of cardiac maladaptive remodeling”. Int J Cardiol. 2015;201:692. doi: 10.1016/j.ijcard.2015.08.169. [DOI] [PubMed] [Google Scholar]
- 66.Kalfon R, Koren L, Aviram S, Schwartz O, Hai T, Aronheim A. ATF3 expression in cardiomyocytes preserves homeostasis in the heart and controls peripheral glucose tolerance. Cardiovasc Res. 2017;113:134–46. doi: 10.1093/cvr/cvw228. [DOI] [PubMed] [Google Scholar]
- 67.Gold R, Kappos L, Arnold DL, Bar-Or A, Giovannoni G, Selmaj K, et al. Placebo-controlled phase 3 study of oral BG-12 for relapsing multiple sclerosis. N Engl J Med. 2012;367:1098–107. doi: 10.1056/NEJMoa1114287. [DOI] [PubMed] [Google Scholar]
- 68.Grarup N, Sulem P, Sandholt CH, Thorleifsson G, Ahluwalia TS, Steinthorsdottir V, et al. Genetic architecture of vitamin B12 and folate levels uncovered applying deeply sequenced large datasets. PLoS Genet. 2013;9:e1003530. doi: 10.1371/journal.pgen.1003530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Lin X, Lu D, Gao Y, Tao S, Yang X, Feng J, et al. Genome-wide association study identifies novel loci associated with serum level of vitamin B12 in Chinese men. Hum Mol Genet. 2012;21:2610–7. doi: 10.1093/hmg/dds062. [DOI] [PubMed] [Google Scholar]
- 70.Shen H, Campanello GC, Flicker D, Grabarek Z, Hu J, Luo C, et al. The human knockout gene CLYBL connects itaconate to vitamin B12. Cell. 2017;171:771–82. doi: 10.1016/j.cell.2017.09.051. e11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Lek M, Karczewski KJ, Minikel EV, Samocha KE, Banks E, Fennell T, et al. Analysis of protein-coding genetic variation in 60,706 humans. Nature. 2016;536:285–91. doi: 10.1038/nature19057. [DOI] [PMC free article] [PubMed] [Google Scholar]