

Summary
Adoptive therapies with genetically modified somatic T cells rendered HIV resistance have shown promise for AIDS therapy. A renewable source of HIV-resistant human T cells from induced pluripotent stem cells (iPSCs) would further facilitate and broaden the applicability of these therapies. Here, we report successful targeting of the CCR5 locus in iPSCs generated from T cells (T-iPSCs) or fibroblasts (fib-iPSCs) from Mauritian cynomolgus macaques (MCM), using CRISPR-Cas9 technology. We found that CCR5 editing does not affect hematopoietic and T cell differentiation potentials of fib-iPSCs. However, T-iPSCs with edited CCR5 lost their capacity to differentiate into CD4+CD8+ T cells while maintaining myeloid differentiation potential. T cells and macrophages produced from CCR5-edited MCM iPSCs did not support replication of the CCR5-tropic simian immunodeficiency viruses SIVmac239 (T cell tropic) and SIVmac316 (macrophage-tropic). Overall, these studies provide a platform for further exploration of AIDS therapies based on gene-edited iPSCs in a nonhuman primate model.
Keywords: nonhuman primates, induced pluripotent stem cells, CCR5, HIV, SIV, T cells, macrophages, CRISPR-Cas9 gene editing
Graphical abstract
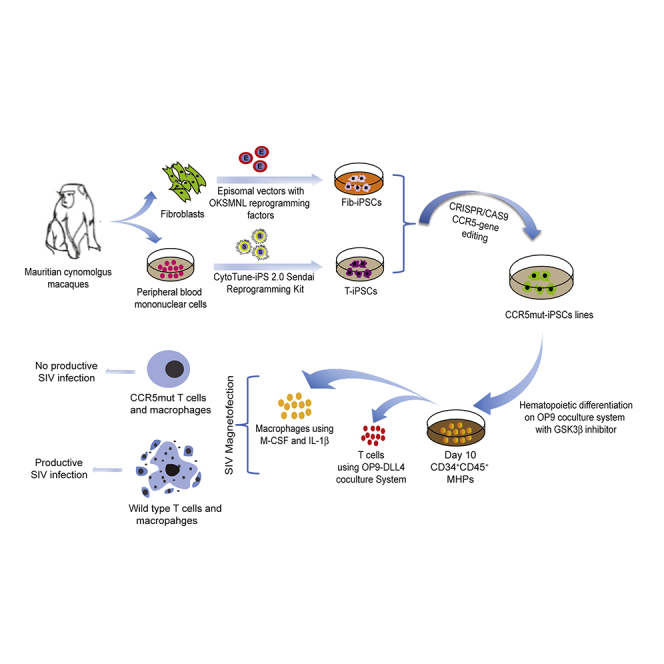
Highlights
-
•
iPSCs with edited CCR5 locus are generated from NHP fibroblasts and T cells
-
•
CCR5-edited T-iPSCs demonstrated impaired differentiation into CD4+CD8+ T cells
-
•
T cells and macrophages from CCR5-edited iPSCs do not support SIV replication
To enable evaluation of iPSC-based technologies in a preclinical HIV infection model, D’Souza et al. established iPSC lines with edited CCR5 locus from Mauritian cynomolgus macaques with MHC-defined genotype. They demonstrated that T cells and macrophages produced from CCR5-edited iPSCs did not to support replication of the CCR5-tropic SIVmac239 (T cell tropic) and SIVmac316 (macrophage-tropic) simian immunodeficiency viruses.
Introduction
Adoptive T cell therapies with in vitro expanded genetically modified T cells have been considered a valuable strategy to treat and cure HIV (Hale et al., 2017; Lam and Bollard, 2013; Patel et al., 2018; Sung et al., 2018). However, T cell exhaustion along with complicated logistics for generation and delivery of genetically modified T cells hampers the broader application of these technologies. Genetic modification of induced pluripotent stem cells (iPSCs) to introduce HIV-resistance and/or anti-HIV molecules, can serve as a versatile and scalable source for off-the-shelf adoptive T cell therapies. In addition, reprogramming of HIV-specific cytotoxic T lymphocytes (CTLs) from HIV-infected patients allows for capturing the specific T cell receptors (TCRs) within the iPSC genome and generating “rejuvenated” antigen-specific CTLs from these iPSCs (Ando and Nakauchi, 2017; Nishimura et al., 2013).
To enable evaluation of iPSC-based technologies in a preclinical HIV infection model, we explored the feasibility of interrupting the CCR5 locus in iPSCs from nonhuman primate (NHP) sources and de novo generating simian immunodeficiency (SIV)-resistant T cells and macrophages from these modified iPSCs. In these studies, we used Mauritian cynomolgus macaques (MCMs) that have a limited major histocompatibility complex (MHC) diversity (Budde et al., 2010; Wiseman et al., 2007, 2013), and could be used to assess adoptive cellular therapies, including T cell therapies in an MHC-defined setting (Greene et al., 2013). iPSCs were generated from fibroblasts and peripheral blood T cells. To successfully disrupt CCR5, we designed two CCR5 synthetic guide RNAs (gRNAs) to target sequences within exon 2, including a 24-base pair (bp) deletion region that was previously found to prevent functional CCR5 expression in NHPs (Chen et al., 1998). Using this approach, we generated SIV-resistant T cells and macrophages from NHP iPSCs, thus laying a foundation for further exploration of iPSC technology for AIDS treatment in an NHP preclinical model. In addition, we noted an impaired capacity of iPSCs generated from T cells (T-iPSCs) to re-differentiate into T cells, especially following biallelic CCR5 disruption. This finding should be taken into consideration when designing strategies for HIV immunotherapies using rejuvenated T cells.
Results
Editing of CCR5 locus in iPSCs from MCM fibroblasts and T cells
Using oriP/EBNA-1 episomal plasmids expressing the six reprogramming factors OCT4, KLF4, SOX2, MYC, NANOG, and LIN28 (OKSMNL), we successfully established an iPSC line from fibroblasts of MHC homozygous MCMs with M3/M3 (fib-iPSC-M3/M3). T-iPSC lines from MCMs with an MHC M3/M3 (T-iPSC-M3/M3) and an MHC M1/M3 (T-iPSC-M1/M3) genotype were generated from peripheral blood T cells using the CytoTune-iPS 2.0 Sendai Reprogramming Kit (Figures 1A and S1A). Generated fib-iPSCs and T-iPSCs exhibited typical NHP embryonic stem cell (ESC) morphology and expressed pluripotency markers OCT4, SOX2, and NANOG (Figures S2A and S2B).
Figure 1.

Generation of CCR5mut iPSCs from MCM fibroblasts and T cells
(A) Schematic diagram of the experimental design.
(B) Schematic representation showing the target site, sequences of the two gRNAs used to delete the 198-bp CCR5 fragment, and position of PCR primers (P1 and P2) used to detect deletion.
(C) Genomic PCR to detect deletion within CCR5 locus. ∗Denotes clones selected in these studies.
See also Figures S1, S2, and S4.
To disrupt CCR5, we used two gRNAs to target sequences within exon 2, including a 24-bp deletion region known to be essential for expressing functional CCR5 in NHPs (Chen et al., 1998) (Figure 1B). We have shown in prior studies with human iPSCs that dual single guide RNAs (sgRNAs) were more efficient in introducing CCR5 gene editing in human iPSCs as compared with a single sgRNA (Kang et al., 2015). By genomic PCR, we found that the dual gRNAs resulted in 66% of CCR5 mutations (CCR5mut) in fib-iPSC-M3/M3, 50% in T-iPSC-M3/M3, and 83% in T-iPSC M1/M3. Of these, 19% of the clones demonstrated biallelic mutation in the fib-iPSC-M3/M3 and 33% in T-iPSC M3/M3 and 15% in T-iPSC M1/M3 (Figures 1C and S1B). Following CCR5 editing, we established CCR5mut iPSC lines from each wild-type (WT) iPSC:CCR5mutC6 from fib-iPSC-M3/M3, CCR5mutC2 from T-iPSC-M3/M3, and CCR5mutC2 from T-iPSC-M1/M3 lines. CCR5mut iPSCs retained pluripotent morphology and expression of pluripotency markers OCT4, SOX2, and NANOG (Figures S2A and S2B). Karyotyping revealed a normal karyotype for CCR5mutC6-fib-iPSC-M3/M3 and CCR5mutC2-T-iPSC-M1/M3. However, CCR5mutC2-T-iPSC-M3/M3 demonstrated a balanced translocation between the long (q) arms of chromosomes 2 and 7 (Figure S2C). This translocation was detected in several CCR5 mutated clones, suggesting that it was introduced during reprogramming, rather than during CCR5 editing.
Generation of T cells and macrophages from CCR5mut iPSCs
To induce hematopoietic differentiation, we used the OP9 co-culture system with CHIR99021 and vascular endothelial growth factor (VEGF) to efficiently induce mesoderm and definitive hematopoiesis (D'Souza et al., 2016) (Figure 2A). Floating cells collected from day 10 of iPSCs/OP9 co-culture were analyzed by flow cytometry. All iPSCs, including WT and CCR5mut fib- and T-iPSCs efficiently produced multipotent hematopoietic progenitors (MHPs) with more than 90% of floating cells expressing CD34 and CD45 (Figure 2B).
Figure 2.

Generation of T cells from CCR5mut MCM iPSCs
(A) Schematic representation of hematopoietic differentiation of fib- and T-iPSCs to MHPs and their further differentiation to T cells.
(B) Both WT and CCR5mut iPSCs efficiently differentiated into MHPs as analyzed by flow cytometry of floating cells collected from iPSC/OP9 co-cultures on day 10 of differentiation.
(C) Day10 MHPs from WT and CCR5mut iPSCs were cultured on OP9-DLL4 in the presence of SCF, IL-7, and FLT3L for 2 weeks, and the floating cells were analyzed by flow cytometry after gating CD45+ cells. ∗Intracellular CD3 was analyzed in WT and CCR5mut fib-iPSCs differentiation cultures.
In (B) and (C), control staining with the appropriate isotype matched mouse monoclonal antibody were included to establish a threshold for positive staining. The graphs are representative of at least three independent experiments.
See also Figures S3 and S4.
To induce T cell differentiation, we collected day 10 CD34+CD45+ MHPs and cultured them on OP9-DLL4 in the presence of interleukin (IL)-7, FLT3 ligand, and stem cell factor (SCF) according to our protocol, which generates functional T cells with rearranged TCR (D'Souza et al., 2016; Kumar et al., 2019). In these cultures, MHPs gave rise to CD5+CD7+ lymphoid progenitors and eventually to CD4+CD8+ T cells (Figure 2C). As reported in prior studies with human T-iPSCs (Nishimura et al., 2013), MCM T-iPSCs show surface CD3 expression very early during differentiation. As shown in Figure 2C, surface CD3 expression was already detected in T-iPSC cultures at 2 weeks of T cell differentiation. However, T cells from fib-iPSCs demonstrated mostly intracellular CD3 expression with negligible surface CD3 expression at this stage of differentiation (Figure 2C). No differences in T cell differentiation were observed between WT fib-iPSC-M3/M3 and CCR5mutC6-fib-iPSC-M3/M3. In contrast, CD4+CD8+ T cell differentiation of MHPs from T-iPSCs was less efficient as compared with fib-iPSCs and both CCR5mutC2-T-iPSC-M1/M3 and CCR5mutC2-T-iPSC-M3/M3 iPSCs failed to produce CD4+CD8+ T cells (Figure 2C). To ensure that these differences were not clone- and individual-dependent, we established three additional CCR5mut iPSCs from the WT fib- and T-iPSC lines described above: CCR5mutC17-fib-iPSC-M3/M3, CCR5mutC10-T-iPSC-M3/M3-, and CCR5mutC23-T-iPSC-M1/M3; and generated five additional WT and CCR5mut fibroblast and T-iPSCs from monkeys with the MHC M6/M6 genotype: fib-iPSC-M6/M6, T-iPSC-M6/M6, CCR5mutC7-fib-iPSC-M6/M6, CCR5mutC3-T-iPSC-M6/M6, and CCR5mutC4-T-iPSC-M6/M6. As shown in Figures S3 and S4, all these lines demonstrated a similar pattern of T cell differentiation, thus confirming that CCR5 knockout in T-iPSCs impairs their differentiation into CD4+CD8+ T cells.
For macrophage differentiation, the day 10 MHPs were collected and plated on ultralow attachment plates in Iscove’s modified Dulbecco’s medium (IMDM) with 10% fetal bovine serum (FBS), macrophage colony-stimulating factor (M-CSF), and IL-1β (Figure 3A). Cells collected after 5 to 7 days displayed typical macrophage morphology and phenotype (Figure 3B). We did not observe significant differences in the macrophage differentiation between WT and CCR5mut-fib-iPSCs and CCR5mut-T-iPSCs from multiple CCR5mut iPSC lines (Figures 3B and S3B). Genomic PCR analysis confirmed that biallelic CCR5 mutations were maintained in both the fib-iPSC and T-iPSC derivatives following hematopoietic differentiation, including multipotent CD34+CD45+ MHPs, macrophages, and T cells (Figures 3C and 3D). In addition, the presence of native CCR5 mRNA in macrophages and T cells from WT iPSCs and the lack of CCR5 mRNA expression in these cells from CCR5mut iPSCs was confirmed by RT-PCR (Figure 3E).
Figure 3.

Generation of macrophages from CCR5mut iPSCs
(A) Schematic representation of hematopoietic differentiation of iPSCs to MHPs and their further differentiation into macrophages.
(B) Day10 MHPs were cultured for 5 to 6 days in the presence of M-CSF and IL-1β to generate macrophages. The phenotype and morphology of the cells was confirmed by flow cytometry and Wright stain. Representative graphs and images of three independent experiments are shown. Scale bar, 50 μm.
(C and D) Genomic PCR to confirm CCR5 mutation in Fib-iPSCs (C) and T-iPSCs (D). Biallelic CCR5 mutation was maintained in the iPSCs, MHPs, macrophages, and T cells following differentiation as checked by genomic PCR.
(E) Loss of CCR5 expression was confirmed in macrophages and T cells from both WT and mutant clones by RT-PCR. β-actin was used as an internal control.
See also Figure S3.
Resistance of CCR5mut macrophages and T cells to SIV infection
The aim of our study was to generate CCR5 gene-disrupted NHP iPSCs that when differentiated to T cells and macrophages are resistant to CCR5-tropic SIV infection. Since CCR5mut-T-iPSCs failed to differentiate into CD4+CD8+ T cells, we only used fib-iPSC-M3/M3 for these studies. To evaluate protection from infection, we challenged T cells and macrophages from WT and CCR5mut-fib-iPSCs with the T cell-tropic SIVmac239 and macrophage-tropic SIVmac316 open SpX (1) virus isolates. As shown in Figure 4, SIV replication, as judged by SIV Gag p27 production, was observed at 4 days in T cells and macrophage cultures from WT fib-iPSCs. In contrast, CCR5mut T cell and macrophage cultures were resistant to productive SIV infection (Figure 4). The productive SIVmac239 infections in our WT fib-iPSC-derived CD4+CD8+ cultures were lower than productive infections reported in peripheral blood T cells (Gautam et al., 2007; Naidu et al., 1988; Sacha and Watkins, 2010). These findings are consistent with a previous report of thymocytes supporting low levels of CCR5 tropic HIV replications (Pedroza-Martins et al., 1998). Nevertheless, we observed distinct differences in the susceptibility of WT and CCR5mut iPSC-derived CD4+CD8+ T cells to SIVmac239 infections. Although we detected ∼100 pg of p27/mL of supernatant 1-day post-magnetofection in the WT fib-iPSC-derived T cell cultures, we believe this reflects residual virus bound to cell surfaces rather than significant levels of de novo SIVmac239 production. Overall, these results demonstrate that CCR5 gene-disrupted T cells and macrophages successfully generated from CCR5 gene altered MCM iPSCs were protected from CCR5-tropic SIV challenge.
Figure 4.

CCR5mut T cells and macrophages resist SIV infection
(A and B) Fib-iPSC-derived mutant and WT T cells (A) and macrophages (B) were incubated with the T-cell-tropic SIVmac239 and macrophage-tropic SIVmac316, respectively. Virus production was measured by collecting cell culture supernatant on days 1, 4, and 7 post-infection and performing SIV Gag p27 ELISAs. The graph is representative of three independent experiments performed in triplicate. The error bars represent the SE of each time point. ∗Denotes p < 0.05.
Discussion
iPSCs derived from somatic cells offer an attractive strategy for the generation of HIV-resistant immune cells for adoptive immunotherapies. In this study, we demonstrated the feasibility of reprogramming T cells from NHPs to pluripotency and generation of SIV-resistant CCR5-mutated T cells and macrophages from fib-iPSCs obtained from MCMs. MCMs are descended from a small founder population and have a very limited MHC diversity consisting of only seven common haplotypes M1-M7 (Budde et al., 2010; Wiseman et al., 2007), therefore making it possible to rapidly select MHC identical, MHC homozygous, and MHC heterozygous animals to explore the utility of MHC homozygous iPSC banking for allogeneic immunotherapies using cells with beneficial MHC match in a preclinical NHP model.
Gene editing to inactivate the CCR5 gene using nucleases has shown promising results toward developing a functional HIV cure (Mock et al., 2015; Mussolino et al., 2014; Tebas et al., 2014; Xu et al., 2019). Three main classes of nucleases, including ZFNs, TALENs, and CRISPR-Cas9 are routinely used to precisely delete or insert specific DNA sequences into the genome. Of these nucleases, the CRISPR-Cas9 system has demonstrated higher cleavage efficiency compared with ZFNs and TALENs (Chen et al., 2013; Fu et al., 2014; Tycko et al., 2016; Zhang et al., 2016). In this study, we employed dual gRNA-guided Cas9 systems to specifically target the CCR5 region affected by Δ24 mutation, which prevents R5 lentiviral infections in macaques (Chen et al., 1998) and is functionally equivalent to the CCR5Δ32 mutation in humans (Liu et al., 1996; Samson et al., 1996). Using the dual gRNA CRISPR/Cas9 system, we achieved up to a 33% biallelic mutation in NHP iPSCs. Our results are consistent with several other reports showing improved editing efficiency using dual gRNA in primary human CD4+ T cells, CD34+ hematopoietic progenitor cells, and iPSCs (Kang et al., 2015; Mandal et al., 2014; Xiao et al., 2019). T cells and macrophages produced from CCR5mut-fib-iPSCs were resistant to SIV challenge, implying that immune cells derived from the NHP iPSCs are useful for continued studies of novel immunotherapies for HIV in a preclinical NHP model. In addition, gene-editing technologies can be used to modify other genes in iPSCs that are essential for HIV/SIV replication, even producing NHP cells that support HIV replication. For example, a recent study knocked out TRIM5 from NHP iPSCs (Iwamoto et al., 2021), a gene encoding a restriction factor that blocks cross-species retrovirus infections, yielding NHP macrophages that are permissive to HIV infection, further demonstrating iPSC technologies' utility in modeling HIV infections.
Generating T cells from T-iPSCs opens opportunities for T cell rejuvenation and unlimited manufacturing of T cells with an antigen-specific monoclonal TCR (Iriguchi et al., 2021; Karagiannis et al., 2016; Nishimura et al., 2013; Trotman-Grant et al., 2021; Vizcardo et al., 2013). The feasibility of this approach for generating therapeutic cells has been demonstrated in murine and human studies, where rejuvenated T cells from tumor- or virus-specific targets were generated (Maeda et al., 2016; Saito et al., 2016; Vizcardo et al., 2013), including HIV (Nishimura et al., 2013). To advance this strategy in a preclinical NHP model, we successfully generated T-iPSCs from MCM peripheral blood T cells, and demonstrated that T-iPSC can be successfully re-differentiated into T cells. However, we found that CD34+ MHP generated from T-iPSC produced T cells less efficiently and CCR5mut-T-iPSCs did not generate CD4+CD8+ T cells, despite no apparent loss in the efficiency of hematopoietic differentiation and macrophage production from WT and CCR5mut-T-iPSCs. This defect in hematopoietic differentiation affected the T cell differentiation step only from T-iPSCs, but not fib-iPSCs, which reproducibly produced CD4+CD8+ T cells regardless of the presence or absence of the CCR5 mutation. The reason for these differences remains unclear. It has been shown that following differentiation, T-iPSCs express TCR complex prematurely before the CD4+CD8+ double-positive stage (Maeda et al., 2016; Nagano et al., 2020), which may lead to strong TCR signaling and eventually death of T cell progenitors. In addition, previous studies have revealed that CCR5 expression promotes IL-2-dependent events during T cell activation, including IL-2R expression, STAT5 phosphorylation, and T cell proliferation (Camargo et al., 2009). Thus, it is possible that CCR5 deficiency in an environment of premature TCR expression may further contribute to the demise of T cell differentiation potential. Further focused studies are required to determine the exact mechanism(s) responsible for selective loss of CD4+CD8+ differentiation potential of T-iPSCs with edited CCR5.
In summary, we have shown that genomic editing of CCR5 can be easily and effectively attained in NHP iPSCs that can be clonally selected to ensure homogeneous CRISPR-Cas9 gene editing. These lines can be useful for understanding the role of CCR5 in HIV pathogenesis and further advancement of iPSC-based technologies for an HIV cure in NHP preclinical models. We also noted that introduction of CCR5 mutation into T-iPSCs affected their T cell redifferentiation potential. This unexpected finding presents an additional challenge to applying T cell rejuvenation technologies for HIV therapies using CCR5-edited HIV-resistant T-iPSCs.
Experimental procedures
NHP iPSC culture and generation of CCR5mut iPSC lines
All animal procedures were approved by the University of Wisconsin Medical School’s Animal Care and Use Committee. MCM iPSCs were harvested using Collagenase IV followed by TrypLE to make a single cell suspension; 1 × 105 singularized cells were resuspended in 100 μL of nucleofector solution (Lonza) containing 10 μg of each sgRNA (#1 and #2 modified sgRNAs, Synthego) and 15 μg Cas9 protein (PNA Bio) and were electroporated using program A23 on the Nucleofector 2b Device (Lonza). After transfection, cells were replated onto MEFs (mouse embryonic fibroblasts) in Primate Embryonic Stem cell medium (ReproCELL) supplemented with 4 ng/mL bFGF (basic fibroblast growth factor) (154 amino acids.) (Peprotech). 15–20 days later, colonies were picked and expanded. Single cell derived knockout cell lines were obtained by single colony picking method with low-density iPSCs culture on MEFs. Genomic DNA from iPSC colonies was extracted using the Quick-DNA Miniprep kit (Zymo Research) and analyzed by PCR. The targeting genomic PCR in CCR5-mutated clones was performed using Q5 Hot Start High Fidelity DNA polymerase (NEB) with the following primers: P1 (TCAATGTGAAACAAATCGCAGC) and P2 (TCGTTTCGACACCGAAGCAG) CCR5-specific primers. Primers specific to β-actin (forward: 5′-GCAGGAGATGGCCACGGCGCC-3′, reverse: 5′-TCTCCTTCTGCATCCTGTCGGC-3′) were used for internal controls.
Differentiation to T lymphoid cells and macrophages
Hematopoietic differentiation of NHP iPSCs was performed on OP9 in the presence of CHIR99021, as previously described (D'Souza et al., 2016). Briefly, small cell aggregates of iPSCs were added to a prolonged culture of OP9 feeder in medium supplemented with 10% HyClone FBS (Cytiva) and 50 μM β-mercaptoethanol (MilliporeSigma); 4 μM of CHIR99021 (Peprotech) and 50 ng/mL VEGF (Peprotech) were added on day 1, for 2 days. The medium was changed and fresh medium was supplemented with 50 ng/mL VEGF. On day 6, an additional 5 mL of medium along with a hematopoietic cytokine cocktail consisting of 50 ng/mL SCF (Peprotech), 50 ng/mL of VEGF (Peprotech), 20 ng/mL of TPO (Thrombopoietin) (Peprotech), 20 ng/mL of IL-3 (Peprotech), and 20 ng/mL of IL-6 (Peprotech) was added to the co-culture. The co-culture was incubated for 10 days in standard conditions of 37°C and 5% CO2. The phenotype of the cells was confirmed by flow cytometry using antibodies against CD45 (Miltenyi) and CD34 (BD Biosciences).
For lymphoid differentiation, the floating CD45+CD34+ MHPs were collected from day 10 of NHP iPSC/OP9 co-cultures, strained through a 70 μm cell strainer (ThermoFisher Scientific) and resuspended in a T cell differentiation medium consisting of αMEM (Gibco) supplemented with 20% HyClone FBS, 5 ng/mL IL-7 (Peprotech), 5 ng/mL Flt3-Ligand (Peprotech), and 10 ng/mL SCF (Peprotech). The cells were cultured on OP9-DLL4 for 2 weeks with weekly passage. The floating cells from T cell cultures were analyzed by flow cytometry using antibodies against CD3, CD4, CD7 (BD Biosciences), CD5, CD8, and TCRαβ (Biolegend) and used for subsequent SIV challenge. For intracellular staining, cells were fixed and permeabilized by resuspending the cell pellet in BD cytofix/cytoperm buffer (BD Biosciences) for 30 min on ice. The cells were then washed with 1xPerm Buffer (BD Biosciences) and stained with CD3 antibody for 30 min in the dark, washed, and analyzed using the MACSQuant Analyzer (Miltenyi Biotec) and FlowJo software (Tree star). Control staining with the appropriate isotype matched mouse monoclonal antibody and unstained controls were included to establish a threshold for positive staining.
For macrophage differentiation, day 10 MHPs from OP9/iPSC co-culture were suspended in IMDM (Gibco) with 10% HyClone FBS supplemented with 20 ng/mL of M-CSF (Peprotech) and 10 ng/mL of IL-1β for 5 to 7 days. Cell phenotype was confirmed by Wright-Giemsa staining and flow cytometry using antibodies against CD45 (Miltenyi), CD14 (BD Biosciences), and HLA-DR (BD Biosciences). Cells from day 6 of macrophage culture were used for SIV challenge. See Table S1 for the complete list and description of antibodies used in this study.
SIV challenge of iPSC-derived T cells and macrophages
To determine whether the CCR5mut immune cells were resistant to infection, we challenged T cells and macrophages derived from WT fib-iPSC-M3/M3 and CCR5mutC6-fib-iPSC-M3/M3 with the CCR5-tropic SIV isolates SIVmac239 and SIVmac316 open SpX, previously shown to be T cell or macrophage-tropic, respectfully (Mori et al., 2000). The SIV stocks were purified by overlaying 127 ng (SIVmac316 open SpX) or 87 ng (SIVmac239) of Gag p27 on 100 μL of a 20% sucrose cushion and centrifuging at 21,000 × g for 1 h at 4°C. Media and sucrose were removed, and the virus pellet was resuspended in 70 μL of PBS. Cell infection was performed using magnetofection. Briefly, 30 μL of ViroMag R/L beads (OZ Biosciences) were added and incubated at room temperature for 15 min (Sacha and Watkins, 2010). During incubation 3 × 105 WT or CCR5-mut cells were placed in a single well of a 24-well plate and centrifuged at 530 × g for 5 min. The virus/bead mixture was then added dropwise to the cells and placed on a magnet for 1 h at 37°C. When the incubation was finished, the cells were pelleted and washed five times with 1 mL PBS. T cells from iPSCs were treated with 0.05% or 0.25% trypsin for 2 min at 37°C to remove bound but noninternalized virions. Then 1 × 105 of each virus/cell combination was placed into three wells of a 48-well plate containing growth media and incubated for 7 days at 37°C, 5% CO2. Culture supernatants were sampled at days 1, 4, and 7 post-magnetofection. A p27 ELISA (Zeptometrix) was performed on each time point according to the manufacturer’s instructions to determine the amount of virus produced in each well. See supplemental materials for additional details of the experimental procedure.
Author contributions
S.S.D. generated and characterized CCR5mut T-iPSCs and fib-iPSCs, analyzed iPSC hematopoietic differentiation potential, produced macrophages, interpreted experimental data, made figures and wrote the manuscript. A.K. generated iPSCs from T cells, characterized CCR5mut T-iPSCs, and generated T cells from iPSCs. J.W. performed SIV infection studies. M.A.P. and L.T. generated and characterized CCR5mut T-iPSCs. J.M. generated iPSCs from fibroblasts. H.J.K. designed CCR5 targeting gRNAs. S.T.D. assisted in generation and characterization of hematopoietic differentiation of iPSCs from M6/M6 animals. T.G. and J.A.T. advised on iPSC generation and characterization. M.R. and I.S. developed the concept, led and supervised studies, analyzed and interpreted data, and wrote the manuscript.
Conflicts of interests
The authors declare no competing interests.
Acknowledgments
This work is supported by funds from the National Institutes of Health, United States (R24OD021322, R01HL132891 and P51OD011106). The following reagent was obtained through the NIH AIDS Reagent Program, Division of AIDS, NIAID, NIH: SIVmac316 open SpX from Dr. Ronald C. Desrosiers.
Published: March 31, 2022
Footnotes
Supplemental information can be found online at https://doi.org/10.1016/j.stemcr.2022.03.003.
Supplemental information
References
- Ando M., Nakauchi H. 'Off-the-shelf' immunotherapy with iPSC-derived rejuvenated cytotoxic T lymphocytes. Exp. Hematol. 2017;47:2–12. doi: 10.1016/j.exphem.2016.10.009. [DOI] [PubMed] [Google Scholar]
- Budde M.L., Wiseman R.W., Karl J.A., Hanczaruk B., Simen B.B., O'Connor D.H. Characterization of Mauritian cynomolgus macaque major histocompatibility complex class I haplotypes by high-resolution pyrosequencing. Immunogenetics. 2010;62:773–780. doi: 10.1007/s00251-010-0481-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Camargo J.F., Quinones M.P., Mummidi S., Srinivas S., Gaitan A.A., Begum K., Jimenez F., VanCompernolle S., Unutmaz D., Ahuja S.S., Ahuja S.K. CCR5 expression levels influence NFAT translocation, IL-2 production, and subsequent signaling events during T lymphocyte activation. J. Immunol. 2009;182:171–182. doi: 10.4049/jimmunol.182.1.171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen B., Gilbert L.A., Cimini B.A., Schnitzbauer J., Zhang W., Li G.W., Park J., Blackburn E.H., Weissman J.S., Qi L.S., Huang B. Dynamic imaging of genomic loci in living human cells by an optimized CRISPR/Cas system. Cell. 2013;155:1479–1491. doi: 10.1016/j.cell.2013.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Z., Kwon D., Jin Z., Monard S., Telfer P., Jones M.S., Lu C.Y., Aguilar R.F., Ho D.D., Marx P.A. Natural infection of a homozygous delta24 CCR5 red-capped mangabey with an R2b-tropic simian immunodeficiency virus. J. Exp. Med. 1998;188:2057–2065. doi: 10.1084/jem.188.11.2057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Souza S.S., Maufort J., Kumar A., Zhang J., Smuga-Otto K., Thomson J.A., Slukvin GSK3beta inhibition promotes efficient myeloid and lymphoid hematopoiesis from non-human primate-induced pluripotent stem cells. Stem Cell Rep. 2016;6:243–256. doi: 10.1016/j.stemcr.2015.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu Y., Sander J.D., Reyon D., Cascio V.M., Joung J.K. Improving CRISPR-Cas nuclease specificity using truncated guide RNAs. Nat. Biotechnol. 2014;32:279–284. doi: 10.1038/nbt.2808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gautam R., Carter A.C., Katz N., Butler I.F., Barnes M., Hasegawa A., Ratterree M., Silvestri G., Marx P.A., Hirsch V.M., et al. In vitro characterization of primary SIVsmm isolates belonging to different lineages. In vitro growth on rhesus macaque cells is not predictive for in vivo replication in rhesus macaques. Virology. 2007;362:257–270. doi: 10.1016/j.virol.2006.12.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greene J.M., Lhost J.J., Hines P.J., Scarlotta M., Harris M., Burwitz B.J., Budde M.L., Dudley D.M., Pham N., Cain B., et al. Adoptive transfer of lymphocytes isolated from simian immunodeficiency virus SIVmac239Deltanef-vaccinated macaques does not affect acute-phase viral loads but may reduce chronic-phase viral loads in major histocompatibility complex-matched recipients. J. Virol. 2013;87:7382–7392. doi: 10.1128/JVI.00348-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hale M., Mesojednik T., Romano Ibarra G.S., Sahni J., Bernard A., Sommer K., Scharenberg A.M., Rawlings D.J., Wagner T.A. Engineering HIV-resistant, anti-HIV chimeric antigen receptor T cells. Mol. Ther. 2017;25:570–579. doi: 10.1016/j.ymthe.2016.12.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mori K., Rosenzweig M., Desrosiers R.C. Mechanisms for adaptation of simian immunodeficiency virus to replication in alveolar macrophages. J Virol. 2000;74:10852–10859. doi: 10.1128/jvi.74.22.10852-10859.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iriguchi S., Yasui Y., Kawai Y., Arima S., Kunitomo M., Sato T., Ueda T., Minagawa A., Mishima Y., Yanagawa N., et al. A clinically applicable and scalable method to regenerate T-cells from iPSCs for off-the-shelf T-cell immunotherapy. Nat. Commun. 2021;12:430. doi: 10.1038/s41467-020-20658-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iwamoto Y., Seki Y., Taya K., Tanaka M., Iriguchi S., Miyake Y., Nakayama E.E., Miura T., Shioda T., Akari H., et al. Generation of macrophages with altered viral sensitivity from genome-edited rhesus macaque iPSCs to model human disease. Mol. Ther. Methods Clin. Dev. 2021;21:262–273. doi: 10.1016/j.omtm.2021.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang H., Minder P., Park M.A., Mesquitta W.T., Torbett B.E., Slukvin CCR5 disruption in induced pluripotent stem cells using CRISPR/Cas9 provides selective resistance of immune cells to CCR5-tropic HIV-1 virus. Mol. Ther. Nucleic Acids. 2015;4:e268. doi: 10.1038/mtna.2015.42. [DOI] [PubMed] [Google Scholar]
- Karagiannis P., Iriguchi S., Kaneko S. Reprogramming away from the exhausted T cell state. Semin. Immunol. 2016;28:35–44. doi: 10.1016/j.smim.2015.10.007. [DOI] [PubMed] [Google Scholar]
- Kumar A., Lee J.H., Suknuntha K., D'Souza S.S., Thakur A.S., Slukvin NOTCH activation at the hematovascular mesoderm stage facilitates efficient generation of T cells with high proliferation potential from human pluripotent stem cells. J. Immunol. 2019;202:770–776. doi: 10.4049/jimmunol.1801027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lam S., Bollard C. T-cell therapies for HIV. Immunotherapy. 2013;5:407–414. doi: 10.2217/imt.13.23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu R., Paxton W.A., Choe S., Ceradini D., Martin S.R., Horuk R., MacDonald M.E., Stuhlmann H., Koup R.A., Landau N.R. Homozygous defect in HIV-1 coreceptor accounts for resistance of some multiply-exposed individuals to HIV-1 infection. Cell. 1996;86:367–377. doi: 10.1016/s0092-8674(00)80110-5. [DOI] [PubMed] [Google Scholar]
- Maeda T., Nagano S., Ichise H., Kataoka K., Yamada D., Ogawa S., Koseki H., Kitawaki T., Kadowaki N., Takaori-Kondo A., et al. Regeneration of CD8alphabeta T cells from T-cell-derived iPSC imparts potent tumor antigen-specific cytotoxicity. Cancer Res. 2016;76:6839–6850. doi: 10.1158/0008-5472.CAN-16-1149. [DOI] [PubMed] [Google Scholar]
- Mandal P.K., Ferreira L.M., Collins R., Meissner T.B., Boutwell C.L., Friesen M., Vrbanac V., Garrison B.S., Stortchevoi A., Bryder D., et al. Efficient ablation of genes in human hematopoietic stem and effector cells using CRISPR/Cas9. Cell Stem Cell. 2014;15:643–652. doi: 10.1016/j.stem.2014.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mock U., Machowicz R., Hauber I., Horn S., Abramowski P., Berdien B., Hauber J., Fehse B. mRNA transfection of a novel TAL effector nuclease (TALEN) facilitates efficient knockout of HIV co-receptor CCR5. Nucleic Acids Res. 2015;43:5560–5571. doi: 10.1093/nar/gkv469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mussolino C., Alzubi J., Fine E.J., Morbitzer R., Cradick T.J., Lahaye T., Bao G., Cathomen T. TALENs facilitate targeted genome editing in human cells with high specificity and low cytotoxicity. Nucleic Acids Res. 2014;42:6762–6773. doi: 10.1093/nar/gku305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagano S., Maeda T., Ichise H., Kashima S., Ohtaka M., Nakanishi M., Kitawaki T., Kadowaki N., Takaori-Kondo A., Masuda K., Kawamoto H. High frequency production of T cell-derived iPSC clones capable of generating potent cytotoxic T cells. Mol. Ther. Methods Clin. Dev. 2020;16:126–135. doi: 10.1016/j.omtm.2019.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naidu Y.M., Kestler H.W., 3rd, Li Y., Butler C.V., Silva D.P., Schmidt D.K., Troup C.D., Sehgal P.K., Sonigo P., Daniel M.D., et al. Characterization of infectious molecular clones of simian immunodeficiency virus (SIVmac) and human immunodeficiency virus type 2: persistent infection of rhesus monkeys with molecularly cloned SIVmac. J. Virol. 1988;62:4691–4696. doi: 10.1128/JVI.62.12.4691-4696.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishimura T., Kaneko S., Kawana-Tachikawa A., Tajima Y., Goto H., Zhu D., Nakayama-Hosoya K., Iriguchi S., Uemura Y., Shimizu T., et al. Generation of rejuvenated antigen-specific T cells by reprogramming to pluripotency and redifferentiation. Cell Stem Cell. 2013;12:114–126. doi: 10.1016/j.stem.2012.11.002. [DOI] [PubMed] [Google Scholar]
- Patel S., Chorvinsky E., Albihani S., Cruz C.R., Jones R.B., Shpall E.J., Margolis D.M., Ambinder R.F., Bollard C.M. HIV-specific T cells generated from naive T cells suppress HIV in vitro and recognize wide epitope breadths. Mol. Ther. 2018;26:1435–1446. doi: 10.1016/j.ymthe.2018.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pedroza-Martins L., Gurney K.B., Torbett B.E., Uittenbogaart C.H. Differential tropism and replication kinetics of human immunodeficiency virus type 1 isolates in thymocytes: coreceptor expression allows viral entry, but productive infection of distinct subsets is determined at the postentry level. J. Virol. 1998;72:9441–9452. doi: 10.1128/JVI.72.12.9441-9452.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sacha J.B., Watkins D.I. Synchronous infection of SIV and HIV in vitro for virology, immunology and vaccine-related studies. Nat. Protoc. 2010;5:239–246. doi: 10.1038/nprot.2009.227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saito H., Okita K., Chang A.E., Ito F. Adoptive transfer of CD8+ T cells generated from induced pluripotent stem cells triggers regressions of large tumors along with immunological memory. Cancer Res. 2016;76:3473–3483. doi: 10.1158/0008-5472.CAN-15-1742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samson M., Libert F., Doranz B.J., Rucker J., Liesnard C., Farber C.M., Saragosti S., Lapoumeroulie C., Cognaux J., Forceille C., et al. Resistance to HIV-1 infection in caucasian individuals bearing mutant alleles of the CCR-5 chemokine receptor gene. Nature. 1996;382:722–725. doi: 10.1038/382722a0. [DOI] [PubMed] [Google Scholar]
- Sung J.A., Patel S., Clohosey M.L., Roesch L., Tripic T., Kuruc J.D., Archin N., Hanley P.J., Cruz C.R., Goonetilleke N., et al. HIV-specific, ex vivo expanded T cell therapy: feasibility, safety, and efficacy in ART-suppressed HIV-infected individuals. Mol. Ther. 2018;26:2496–2506. doi: 10.1016/j.ymthe.2018.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tebas P., Stein D., Tang W.W., Frank I., Wang S.Q., Lee G., Spratt S.K., Surosky R.T., Giedlin M.A., Nichol G., et al. Gene editing of CCR5 in autologous CD4 T cells of persons infected with HIV. N. Engl. J. Med. 2014;370:901–910. doi: 10.1056/NEJMoa1300662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trotman-Grant A.C., Mohtashami M., De Sousa Casal J., Martinez E.C., Lee D., Teichman S., Brauer P.M., Han J., Anderson M.K., Zuniga-Pflucker J.C. DL4-mubeads induce T cell lineage differentiation from stem cells in a stromal cell-free system. Nat. Commun. 2021;12:5023. doi: 10.1038/s41467-021-25245-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tycko J., Myer V.E., Hsu P.D. Methods for optimizing CRISPR-cas9 genome editing specificity. Mol. Cell. 2016;63:355–370. doi: 10.1016/j.molcel.2016.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vizcardo R., Masuda K., Yamada D., Ikawa T., Shimizu K., Fujii S., Koseki H., Kawamoto H. Regeneration of human tumor antigen-specific T cells from iPSCs derived from mature CD8(+) T cells. Cell Stem Cell. 2013;12:31–36. doi: 10.1016/j.stem.2012.12.006. [DOI] [PubMed] [Google Scholar]
- Wiseman R.W., Karl J.A., Bohn P.S., Nimityongskul F.A., Starrett G.J., O'Connor D.H. Haplessly hoping: macaque major histocompatibility complex made easy. ILAR J. 2013;54:196–210. doi: 10.1093/ilar/ilt036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiseman R.W., Wojcechowskyj J.A., Greene J.M., Blasky A.J., Gopon T., Soma T., Friedrich T.C., O'Connor S.L., O'Connor D.H. Simian immunodeficiency virus SIVmac239 infection of major histocompatibility complex-identical cynomolgus macaques from Mauritius. J. Virol. 2007;81:349–361. doi: 10.1128/JVI.01841-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao Q., Chen S., Wang Q., Liu Z., Liu S., Deng H., Hou W., Wu D., Xiong Y., Li J., Guo D. CCR5 editing by Staphylococcus aureus Cas9 in human primary CD4(+) T cells and hematopoietic stem/progenitor cells promotes HIV-1 resistance and CD4(+) T cell enrichment in humanized mice. Retrovirology. 2019;16:15. doi: 10.1186/s12977-019-0477-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu L., Wang J., Liu Y., Xie L., Su B., Mou D., Wang L., Liu T., Wang X., Zhang B., et al. CRISPR-edited stem cells in a patient with HIV and acute lymphocytic leukemia. N. Engl. J. Med. 2019;381:1240–1247. doi: 10.1056/NEJMoa1817426. [DOI] [PubMed] [Google Scholar]
- Zhang J.P., Li X.L., Neises A., Chen W., Hu L.P., Ji G.Z., Yu J.Y., Xu J., Yuan W.P., Cheng T., Zhang X.B. Different effects of sgRNA length on CRISPR-mediated gene knockout efficiency. Sci. Rep. 2016;6:28566. doi: 10.1038/srep28566. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.