

Abstract
Background
The rise in Plasmodium falciparum resistance to dihydroartemisinin–piperaquine (DHA–PPQ) treatment has been documented in the Greater Mekong Subregion with associations with mutations in the P. falciparum chloroquine resistance transporter (pfcrt) and plasmepsin 2 (pfpm2) genes. However, it is unclear whether other genes also play a role with PPQ resistance, such as the E415G mutation in the exonuclease (pfexo) gene. The aim of this study was to investigate the role of this mutation in PPQ resistance by generating transgenic parasites expressing the pfexo-E415G mutant allele.
Methods
Transgenic parasite clones carrying the E415G mutation in PfEXO of the B5 isolate were derived by CRISPR-Cas9 gene editing and verified using PCR and gene sequencing. Polymorphisms of pfkelch-13, pfcrt, and pfexo were examined by PCR while the copy number variations of pfpm2 were examined by both relative quantitative real-time PCR and the duplication breakpoint assay. Drug sensitivity against a panel of antimalarials, the ring-stage survival assay (RSA), the PPQ survival assay (PSA), and bimodal dose-response curves were used to evaluate antimalarial susceptibility.
Results
The transgenic line, B5-rexo-E415G-B8, was successfully generated. The PPQ-IC90, %PPQ survival, and the bimodal dose-response clearly showed that E415G mutation in PfEXO of B5 isolate remained fully susceptible to PPQ. Furthermore, growth assays demonstrated that the engineered parasites grew slightly faster than the unmodified parental isolates whereas P. falciparum isolates harbouring pfkelch-13, pfcrt, and pfexo mutations with multiple copies of pfpm2 grew much more slowly.
Conclusions
Insertion of the E415G mutation in PfEXO did not lead to increased PPQ-IC90 and %PPQ survival, suggesting that this mutation alone may not be associated with PPQ resistance, but could still be an important marker if used in conjunction with other markers for monitoring PPQ-resistant parasites. The results also highlight the importance of monitoring and evaluating suspected genetic mutations with regard to parasite fitness and resistance.
Supplementary information
The online version contains supplementary material available at 10.1186/s12936-022-04148-z.
Keywords: Exonuclease, Gene editing, Malaria, Piperaquine resistance, Plasmodium falciparum
Background
Artemisinin-based combination therapy (ACT) remains the recommended first-line antimalarial therapy for uncomplicated Plasmodium falciparum infections in almost all endemic countries [1]. ACT consists of two drugs: a semisynthetic artemisinin (ART) derivative and a longer-lasting partner drug. Currently, six artemisinin-based combinations are available including artemether + lumefantrine (AL), artesunate + amodiaquine (AS + AQ), artesunate + mefloquine (AS + MQ), artesunate + sulfadoxine–pyrimethamine (AS + SP), dihydroartemisinin + piperaquine (DHA + PPQ) and most recently artesunate + pyronaridine (AS + PND) [1]. However, the effectiveness of ACT is threatened by the emergence of parasites with decreased susceptibility to the ART derivatives and/or resistance to ACT partner drugs [2–4]. Soon after DHA–PPQ was recommended by the World Health Organization (WHO) as a first-line therapy for uncomplicated P. falciparum malaria [5], the emergence of DHA–PPQ resistance was reported within the Greater Mekong subregion including Cambodia [6–9], Vietnam [10], and more recently in Thailand [11].
ART resistance is characterized by the presence of mutations in the Pfkelch13 (pfk13) propeller domain gene, which is associated with delayed in vivo parasite clearance times [2, 3] and with increased survival in the in vitro ring-stage survival assay (RSA) [12]. There are three molecular marker candidates for PPQ resistance: multiple copies of P. falciparum plasmepsin 2 (pfpm2) (PF3D7_1408000) [13–16], novel mutations in P. falciparum chloroquine resistance transporter (PfCRT) (PF3D7_0709000) [17–20], and a single mutation in P. falciparum exonuclease (PfEXO) (PF3D7_1362500) [13, 14, 21]. There have been eight previous publications reporting on the E415G mutation in the pfexo gene [13, 22–28]. In addition to the amplification of pfpm2 and pfpm3, Amato et al. [13] identified a non-synonymous single nucleotide polymorphism (SNP) E415G substitution on an exonuclease-encoding gene from recrudescence isolates from DHA–PPQ treatment failures in Cambodia. The prevalence of DHA–PPQ treatment failures was at 2% in Ratanakiri, 16% in Preah Vihear, and 46% in Pursat. In Vietnam, neither PfEXO-E415G mutations nor pfpm2 amplification was observed in parasites from south-central Vietnam between 2015 and 2016 [27], but P. falciparum samples collected from 2018 to 2019 in Vietnam’s Central Highlands [25] were found to harbour the PfEXO-E415G mutation with a prevalence of 66.7% and 85.5% in Dak Nong and Dak Lak, respectively. Si et al. [26] have recently reported that no parasites from the China-Myanmar border carried PfEXO-E415G, novel PfCRT mutations, or pfpm2 amplification, even though some parasites with higher PPQ survival assay (PSA) values were detected, indicating different mechanisms of reduced PPQ susceptibility. In Africa, Robert et al. [23] first investigated the presence of both the PfEXO-E415G mutation and multiple copies of pfpm2 in Senegalese parasites; however, no isolated parasites carried both molecular markers. A similar trend was observed in Sudan where no PfEXO-E415G mutations were observed [28]. However, in Mali, of 214 P. falciparum samples from Dangassa, two isolates exhibited the PfEXO-E415G mutation [22].
While the implication of multiple copies of pfpm2 and novel PfCRT mutations on PPQ resistance have been elucidated [19, 20, 29], and the PPQ-resistance phenotype can be observed in the presence of novel PfCRT mutations with the PfEXO-E415G mutation [24], there is still a lack of confirmation as to whether the PfEXO-E415G mutation alone causes PPQ-resistance. To investigate whether the PfEXO-E415G is directly associated with PPQ resistance, in this study, CRISPR-Cas9 genome editing was used to generate P. falciparum Cambodian parasites harbouring PfEXO-E415G. The in vitro susceptibility to piperaquine and other frontline antimalarials of these gene-edited parasites was determined and compared to those of validated PPQ-resistant P. falciparum clinical isolates of Cambodian origin.
Methods
pfexo gene and amino acid prediction
The nucleotide sequence of pfexo from 3D7 was obtained from PlasmoDB with the accession number PF3D7_1362500. The P. falciparum B5 line was obtained from the cloning of a Cambodian P. falciparum isolate [24]. This isolate is ART- and PPQ-sensitive, but CQ-resistant. The full-length pfexo gene of B5 was amplified from P. falciparum B5 genomic DNA using polymerase chain reaction (PCR) with primers PA_exon1 and Screening_3’UTR_Rev (Additional file 1: Table S1). The encoded amino acid sequences and the nucleotide sequences were aligned using Clustal Omega [30]. The signal peptide region was predicted using Signa-P.5.0 [31] and the InterPro program was used for protein family classification [32].
Plasmodium falciparum culture
Asexual blood-stages of P. falciparum were maintained in fresh human erythrocytes (O+) in RPMI1640 (Sigma-Aldrich, USA) supplemented with 5.94 gL−1 HEPES, 2.1 gL−1 sodium bicarbonate, 0.1 gL−1 gentamycin sulphate, 0.5% (w/v) Albumax II, 4 gL−1 dextrose 0.05 gL−1 hypoxanthine, and 10–15% of human serum (complete medium) [33]. Human blood products (erythrocytes and serum) were obtained from the Thai Red Cross. Culture flasks were gassed with 5% CO2, 5% O2, 90% N2 gas and incubated at 37 °C. For synchronisation, mature schizont stage parasites were isolated on cushions of 75% (v/v) isotonic Percoll cushion (GE Healthcare Life Science) as previously described [34, 35] whereas enrichment for ring stages following invasion was performed using 5% (w/v) d-sorbitol [36] The final ring cultures were washed and returned to culture or used as required.
Construction of plasmid constructs to genetically modify P. falciparum
PCR amplicons used in plasmid cloning were generated using Fusion, High Fidelity DNA polymerase (New England Biolabs) and purified using Qiagen PCR purification or Qiagen Gel extraction kits. For diagnostic PCR amplification, GoTaq (Promega) DNA master mix was used. All constructed plasmids were sequenced to verify authenticity (Biobasic, Canada). For parasite genomic DNA extraction, total cell pellets were first treated with 0.15% saponin in PBS for 10 min, then washed with PBS before DNA was extracted using a DNeasy Blood & Tissue Kit (Qiagen).
Construct for donor plasmids
A fragment of exo sequence with E415G mutation (1441 bp) was commercially synthesised (GenScript, USA), comprising a stretch of native P. falciparum 3D7 exo sequence (covering residues Lys283, exo intron, and Asn408) followed by a stretch of recodonized exo gene sequence encoding the 3D7 amino acid residues Met409 to Glu514 with the point mutation at residue 415 from Glu to Gly, but using a different codon usage, and finally a stretch of native 3D7 exo sequence covering amino acid residues 515 to the stop codon. This fragment was cloned into pUC57 by EcoRV generated pUC57-Exo-E415G plasmid. To generate a donor plasmid to introduce Glu415Glu wild-type, pUC57-Exo-E415E plasmid was derived from pUC57-Exo-E415G by Q5 site-directed mutagenesis (New England Biolabs) to alter the codon encoding at amino acid position 415 from GGC to GAA primers Q5SDM_G415E_F and Q5SDM_G415E_R were used (Additional file 1: Table S1).
Construct for CRISPR-Cas9 plasmids
pDC2-Cas9-hDHFR-yFCU, containing a Cas9 expression cassette and the drug selection marker human dihydrofolate reductase (hdhfr) [37] was used as a template vector to generate pDC2-Cas9-bsd-yFCU. Since the P. falciparum B5 strain is resistant to pyrimethamine, the hdhfr gene was replaced with the blasticidin gene (bsd). The bsd gene (399 bp) was amplified from PkpSKIP_Pk47 plasmid [38] using primers bsd_NcoI_F and bsd_SacII_R (Additional file 1: Table S1) and was cloned into the NcoI/SacII-digested pDC2-Cas9-hDHFR-yFCU, giving rise to pDC2-Cas9-bsd-yFCU.
Guide RNA sequences specific for targeting at amino acid position 415 were identified using Benchling (https://www.benchling.com/crispr/) (Additional file 1: Fig. S1). A pair of complementary oligonucleotides (sgE415G-1F and sgE415G-1R) corresponding to the 19 nucleotides adjacent to the identified PAM sequences were phosphorylated using T4 polynucleotide kinase, annealed and ligated into pDC2-Cas9-bsd-yFCU predigested with BbsI, resulting in the guide vector pGuide1.4-bsd.
Generation of P. falciparum lines expressing exo-E415G mutant
The donor plasmid pUC57-Exo-E415G was linearized with ScaI prior to electroporation. Percoll-enriched mature schizonts of P. falciparum B5 were electroporated with 20 µg of pGuide1.4-bsd and 60 µg of linearized pUC57-Exo-E415G using either Amaxa P3 primary cell 4D Nucleofector X Kit L (Lonza) or Amaxa Basic Parasite Nucleofector Starter Kit as described [39, 40]. Twenty-four hours post-transfection, the electroporated parasites were treated with 5.45 µM blasticidin-S-hydrochloride (Sigma-Aldrich, USA) to select for transfectants harbouring pGuide1.4-bsd before returning the cultures to medium without drug. Detection of the exo-E415G modified locus was carried out by diagnostic PCR using primer pairs ExonI_K283_F and Screen_WT_Rev_V616, ExonI_K283_F and Recodon_R, and Recodon_F and Screen_3′UTR_R. The wild-type pfexo locus was detected by diagnostic PCR using primers ExonI_K283_F and Screen_WT_Rev_V616, with a PCR product of 766 bp. Transgenic parasite clones were obtained by limiting dilution cloning by plating a calculated 0.3 parasite per well (200 µL and 1% haematocrit) in flat-bottomed 96-well microplate wells as described [41]. Wells containing single plaques were identified after 10–14 days using an inverted microscope and the parasites subsequently expanded into round-bottomed wells for further analysis. Transgenic parasite clones were finally checked by diagnostic PCR for integration and modification of the endogenous pfexoE415G gene. A pfexoE415E transgenic line expressing the wild-type Glu415 codon was generated in the same manner of that for a pfexoE415G transgenic line by transfecting P. falciparum B5 with 20 µg of pGuide1.4-bsd and 60 µg of linearized pUC57-Exo-E415E. Once established, all transgenic clones were maintained in medium without any drug.
pfk13, pfcrt, and pfexo genotyping
Master Cycler Nexus Gradient (Eppendorf) was employed to evaluate the propeller domain of the P. falciparum kelch13 (pfk13) (amino acid residues 442–727) [42, 43], P. falciparum exonuclease (pfexo) SNP at a codon corresponding to the amino acid position 415 [13], and the P. falciparum chloroquine resistant transporter (pfcrt) SNPs at codons corresponding to amino acid positions 93, 97, 145, 218, 343, 350, and 353 [19, 44]. Primers used to identify pfk13, pfexo, and pfcrt SNPs are shown in Additional file 1: Table S1. Plasmodium falciparum reference DNAs from 3D7 and W2 clones (Malaria Research & Reference Reagent Resource, Manassas, VA) were used as positive controls, and all samples were performed in duplicate.
Plasmepsin (pfpm) 1, 2 and 3 copy number variation assay
To determine copy numbers of pfpm1 (PF3D7_1407900), pfpm2 (PF3D7_1408000), and pfpm3 (PF3D7_1408100) gene, real-time quantitative PCR (qPCR) was performed on genomic DNA as previously described [29]. The amplification reactions were performed according to Luna® Universal qPCR master mix kit (New England Biolabs) with 200 nM of each forward and reverse primer (Additional file 1: Table S1) and 2 ng of DNA template using Rotor-Gene Q (QIAGEN, Valencia, CA). For the housekeeping gene, β-tubulin (PF3D7_1008700), β-tubulin forward and reverse primers were designed and used as a reference control for all experiments with the same validated PCR conditions as target primers. P. falciparum 3D7 was used as a reference clone. All samples including the references clones were performed in triplicate. The average copy number values for each gene were calculated using 2−ΔΔCt method where ΔΔCt is [Ctpfpm − Ctpf β-tubulin]sample − [Ctpfpm2 − Ctpf β-tubulin]3D7. Parasites with copy number greater than 1.6 copies for pfpm2 and pfpm2 [15, 16] were interpreted to contain multiple copies.
Plasmepsin 2/3 (pfpm2/3)duplication breakpoint PCR assay
The pfpm2/3 breakpoint PCR assay was performed as previously described [45]. Three pairs of primers (Additional file 1: Table S1) were used in this assay. Primers AF_for and AR_rev amplified a 623 bp product surrounding the breakpoint located at the 3′ end of pfpm1. Primers BF_for and BR_rev amplified a 484 bp product surrounding the breakpoint at the 3′ end of pfpm3. Primers BF_for and AR_rev amplified the junction between the breakpoint, producing a 497 bp product in parasite isolates with pfpm2/3 amplifications. A pfpm2/3 single copy isolate is not expected to have the PCR product with these primers. One copy isolate was only noted when the control primer sets amplified a product, and the duplication PCR was negative. Two or more copies were annotated as > 1 copy of pfpm2/3 only when both the control and duplication primer sets generated a product. PCR reactions contained 12.5 µL GoTaq® Green Master Mix (Promega), 1 µL of each primer (10 µM stocks), 3 µL of DNA up to 25 µL final volume with water. PCR conditions were as follow: 95 °C for 2 min, followed by 30 cycles of 95 °C for 45 s, 50 °C for 30 s, 72 °C for 1 min, followed by a 5-min extension at 72 °C.
In vitro drug susceptibility
Drug susceptibility testing used HRP-2 ELISA to measure 50% or 90% inhibitory concentration (IC50 and IC90) performed as previously published [46, 47]. In vitro drug susceptibility testing was carried out for control reference clones (W2, D6, C2B) (Malaria Research & Reference Reagent Resource, Manassas, Vermont, USA) as described previously [48]. IC50s and IC90s were estimated by nonlinear regression analysis using GraphPad Prism version 6.0 program. Samples having poor growth rate, as perceived by obtaining an OD ratio of < 1.7 between the no-drug test wells and the maximum tested drug concentration, were excluded from data analysis.
Bimodal dose response curve
To determine a bimodal-dose response curve, the PPQ concentration (2.44 to 100,000 ng/mL) and the dilution series were increased from 8 to 24 points, according to previously published reports [16, 24]. Culture-adapted clinical isolates or engineered parasites were prepared in the similar manner as in in vitro drug susceptibility testing. The synchronized rings were grown for 72 h in the presence of different concentrations of PPQ (24-point dilution) in 96-well plates at 1.5% haematocrit, 0.5% starting parasitaemia in 0.5% Albumax RPMI 1640. Growth at 72 h was measured by HRP-2 ELISA. Assays were carried out in three biological replicates and the control reference clone W2 was tested along with each culture-adapted clinical isolate. The area under the curve (AUC) for the dose response curve at 0.01–100 µM was calculated using GraphPad Prism 6.0.
Ring-stage survival assay (RSA)
In vitro RSA0–3 h was performed on 0–3 h post-invasion rings obtained from selected culture-adapted clinical isolates following published methods [12] with slight modifications. Briefly, parasite cultures were tightly synchronized using 5% (w/v) d-sorbitol and 75% Percoll to obtain 0 to 3-h post-invasion rings which were adjusted to 0.5–1% starting parasitaemia with a 2% haematocrit in culture media (0.5% Albumax RPMI 1640 with 2.5% AB serum) and cultured in a 48-well microplate with 700 nM DHA and 0.1% DMSO in separate wells for growth control. The culture plate was then incubated for 6 h at 37 °C in modular incubator chambers and gassed with 5% CO2, 5% O2 and 90% N2 gas. Cells were then washed once, resuspended in drug-free medium, and cultured further for 66 h. Susceptibility to DHA was assessed microscopically on thin films by estimating the percentage of viable parasites, relative to control (% survival rate). For the controls, the RSA0–3 h was also performed on P. falciparum reference clones W2 (ART-sensitive control), IPC-4884 and IPC-5202 (BEI Resources, NIAID, NIH, USA) as ART-resistant control lines. A survival rate > 1% was deemed resistant for RSA.
Piperaquine survival assay (PSA)
PSA0–3 h was performed on culture-adapted clinical isolates with 0–3-h ring-stage parasite cultures following a previously published method [17]. Briefly, parasite cultures were tightly synchronized using 5% (w/v) d-sorbitol and 75% Percoll to obtain 0 to 3-h post-invasion. Synchronized ring parasites at 0.5–1% starting parasitaemia and 2% haematocrit were incubated with 200 nM PPQ or 0.5% lactic acid in water at 37 °C for 48 h in a 48-well microplate. The cultures were then washed once, resuspended in drug-free medium, and cultured further for 24 h. Susceptibility to PPQ was assessed microscopically on thin films by estimating the percentage of viable parasites in the similar manner as RSA. A survival > 10% was deemed resistant to PPQ.
Growth assays
For longer-term replication assays, cultures were synchronized as described and resulting in ring stage cultures. The ring stage parasites were synchronized using 5% (w/v) d-sorbitol and parasitaemia levels were calculated, and cultures adjusted to 0.1% parasitaemia, 2% haematocrit in a final volume of 1 mL per well of a 12-well plate. Samples were then taken at t = 0, 24, 72, 120, and 168 h, fixed in 0.8% glutaraldehyde in PBS and stored at 4 °C for flow cytometry analysis. Culture media was replaced at 48, 71, and 120 h. Giemsa-stained thin films were also prepared as required for microscopic analysis.
Flow cytometry for parasite quantification
Parasite samples were fixed in 0.8% glutaraldehyde in PBS and stored at 4 °C. Cells were prepared for analysis by staining with 2× SYBR Green I nucleic acid gel stain (Invitrogen, Thermo Fisher Scientific) for 30 min at 37 °C. Labelling was stopped with an equal volume of PBS and samples acquired using a CytoFlex (Beckman Coulter, USA) with CyEXpert software. Total RBC numbers were calculated using forward- and side-scatter while fluorescence was detected using the 530/630 blue detection laser. Fluorescence intensity was used to distinguish uninfected from infected RBCs, low fluorescence indicating uninfected cells and gating fixed accordingly. Data were analysed using FlowJo.
Statistical analysis
Statistical analysis was performed using GraphPad Prism version 6.0 (GraphPad Software, Inc., San Diego, CA, USA). The difference of the data between groups was assessed by nonparametric Mann–Whitney U test, as appropriate. Statistical significance was defined as a P value of < 0.05.
Results
CRISPR-Cas9-mediated editing to introduce pfexoE415G into P. falciparum parasites
The pfexo gene from P. falciparum 3D7 (PF3D7_1362500) consists of 2274 bp with 2 exons and 1 intron. Exon 1 comprises nucleotides 1-949, while the exon 2 starts from nucleotides 1082–2274 (Additional file 1: Fig. S1). PfEXO consists of 713 amino acids with a molecular mass of 86.7 kDa. The enzyme contains a peptide signal region (residues 1–70) and a 3′ to 5′ exonuclease domain (residues 445–618). The DNA sequence of the pfexo gene from P. falciparum B5 [24], the parental line for genome editing in this study, showed a single non-synonymous mutation, leading to a missense mutation from Lys to Asn at residue 614 (Additional file 1: Fig. S2).
To evaluate the impact of the PfEXO-E415G mutation on parasite susceptibility to piperaquine and other front-line chemotherapies, CRISPR-Cas9 editing was used to introduce the E415G mutation into the native pfexo gene (Fig. 1A). A pfexoE415G was efficiently installed onto the P. falciparum B5 background [24]. In parallel, a pfexoE415E transgenic line expressing wild-type E415 codon in the context of recodonization abating the Cas9 protospacer adjacent motif (PAM) site was also attempted to rule out a role for silent mutations in drug susceptibility. Eleven transfections were performed to introduce pfexoE415G mutation and 3 transfections for pfexoE415E mutation but only three transfections provided revived parasites. Two of the three transfections gave rise to the modified pfexo gene, and one transfection did not provide the modified parasites. Successful modification of the pfexo gene in the transfected population following the introduction of the targeting vector was observed around 21 days after transfection and confirmed by diagnostic PCR (Fig. 1B, pre-cloning). Limiting dilution cloning of the modified parasites resulted in the isolation of parasite clone, B5-rexo-E415G-B8. It is noted that several clones obtained from the cloning did not contain the modified region of the pfexo gene. Modification of the native pfexo locus was then confirmed in the transgenic line by diagnostic PCR (Fig. 1B, B5-rexo-E415G-B8) and genomic sequences of transgenic lines (Fig. 1C). It is noted that the band of 828 bp appearing on lane 2 (Fig. 1B) arose from the non-specificity of primer 3 which can anneal to both parental and transgenic parasite lines. Unfortunately, the pfexoE415E transgenic line could not be obtained from this study.
Fig. 1.

Gene editing of pfexo. (A) CRISPR-Cas9 editing was used to install the pfexoE415G variant codon into the endogenous locus. Using a donor plasmid and Cas9-aided homologous recombination, the region of the pfexo encoding Met409 to Glu514 was replaced with a recodonized pfexo gene (navy blue) with the point mutation at residue 415 (a cyan star) from Glu to Gly. Stop codon is represented as a red hexagon. Position of hybridisation of primers used for confirmation of the integration event by diagnostic PCR are shown as coloured arrows. A schematic of the co-transfected plasmid for expression of Cas9 nuclease and the guide RNAs is also shown. (B) Diagnostic PCR analysis of genomic DNA of the control parental B5 line, the transfected parasites prior to limiting dilution cloning and clone parasite lines expressing pfexoE415G (clone B8). Expected sizes of the various PCR amplicons are indicated on the right of the gel, whilst the left-hand lane contains double-stranded DNA ladder marker fragment (Quick-load Purple 1 kb Plus DNA ladder (New England Biolabs)). (C) Chromatograms of a region of exon 2 obtained from the PCR product amplified pfexo gene of B5 and B5-rexo-E415G-B8 lines. The codon “GAG” encoding Glu in B5 parasites is changed to “GGC” encoding Gly in B5-rexo-E415G-B8 clone
Molecular genotyping of modified pfexo transgenic lines and clinical isolates with PfEXO-E415G mutation
The genotypic profiling for piperaquine and artemisinin molecular markers is outlined in Table 1. In addition to the reference clones and engineered parasite lines, two Cambodian isolates from a previous P. falciparum clinical efficacy study (ASAP-21 and ASAP-168) [49] were included as they harboured PfEXO-E415G with and without novel PfCRT mutations. As expected, the transgenic B5-rexo-E415G-B8 line carried a similar genotypic profile as the parental B5 line except for the E415G mutation in the pfexo gene. Regarding the two clinical isolates, both carried the PfK13-C580Y and PfEXO-E415G mutations. The ASAP-21 isolate harboured the novel PfCRT-F145I mutation while ASAP-168 did not. Genetic studies have identified copy number variations in the pfpm2 and pfpm3 genes that associate with clinical and in vitro PPQ resistance [13, 15]. For pfpm2/3 amplification, the putative breakpoints lie near the 3′ end of both pfpm1 and pfpm3 so each amplification produces an intact extra copy of pfpm2 together with a new chimeric pfpm3 with its 3′ end replaced by the 3′ end of pfpm1 (Additional file 1: Fig. S3). This result showed that the copy number variation determined by qPCR assay showed that all parasite lines harboured a single copy of pfpm1. Two methods were then carried out to determine the copy number variation of the pfpm2 and pfpm3 gene: the SYBR-green qPCR and breakpoint assays [45]. By using the qPCR assay, ASAP-21 was found to contain multiple copies of pfpm2 and ASAP-168 carried a single copy of pfpm2, whereas B5 parental line and transgenic B5-rexo-E415G-B8 were found to have multiple copies of pfpm3. On the contrary, both ASAP-21 and ASAP-168 isolates were positive for pfpm2/3 amplification (>1 copy) while B5 and B5-rexo-E415G-B8 lines showed no pfpm2/3 amplification as detected by the breakpoint assay (Additional file 1: Fig. S3). The non-concordant detection between the two methods were also reported by Ansbro et al. [45] and it was suggested that the breakpoint PCR assay is more sensitive than the qPCR assay for detecting minor clones containing the duplication in field isolates. Therefore, in this study, ASAP-21 and ASAP-168 isolates were treated as having multiple copies of pfpm2/3 amplification, while B5 and B5-rexo-E415G-B8 lines contained a single copy of pfpm2 and pfpm3.
Table 1.
Molecular genotyping of pfexo-modified, parental and clinical P. falciparum parasites
| Sample | PfK13 C580Y |
PfEXO E415G |
PfCRT | Copy number variation | pfpm2/3 break point | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T93S | H97Y | F145I | I218F | M343L | C350R | G353V | pfpm1 | pfpm2 | pfpm3 | ||||
| 3D7 | C | E | T | H | F | I | M | C | G | 1.00 | 1.00 | 1.00 | 1 |
| B5 | C | E | T | H | F | I | M | C | G | 1.37 | 0.93 | 1.99 | 1 |
| B5-rexo-E415G-B8 | C | G | T | H | F | I | M | C | G | 1.19 | 0.74 | 2.08 | 1 |
| ASAP-21 | Y | G | T | H | I | I | M | C | G | 1.25 | 2.31 | 2.60 | > 1 |
| ASAP-168 | Y | G | T | H | F | I | M | C | G | 1.18 | 1.11 | 1.88 | > 1 |
Underline and bold letters indicate either mutations or multiple copy number. A cut-off copy number of 1.6 is used to define pfpm2 and pfpm3 multiple copy number
Drug sensitivity and survival assay of modified pfexo transgenic lines and clinical isolates
The transgenic parasites expressing E415G mutant of PfEXO and the clinical isolates were tested for their PPQ susceptibility (Table 2). No significant difference in IC50 values of B5-rexo-E415G-B8 and ASAP-168 parasites compared to B5 parasites was observed, while PPQ-IC50 value of ASAP-21 parasites was significantly higher than that of B5 parasites. The same trend was observed for PPQ-IC90 values in that there was no significant difference in the IC90 values for B5-rexo-E415G-B8 and ASAP-168 parasites compared to B5 parasites. The PPQ-IC90 of ASAP-21 exceeded the highest PPQ concentration used in this study, so the IC90 could not be reported.
Table 2.
Piperaquine (PPQ) susceptibility (mean ± SD)
| Sample | PPQ-IC50 (nM) | PPQ-IC90 (nM) |
|---|---|---|
| B5 | 112 ± 54 | 172 ± 9 |
| B5-rexo-E415G-B8 | 185 ± 541 | 243 ± 221 |
| ASAP-21 | 5207 ± 6902 | N.D. |
| ASAP-168 | 111 ± 171 | 312 ± 371 |
1No significant difference (P value > 0.05) compared to B5 as tested by Mann–Whitney U Test
2Significant difference (P value = 0.0167) between ASAP-21 and B5 as tested by Mann–Whitney U Test
N.D. for not determined
To evaluate if the modified pfexo transgenic parasites and clinical isolates with PfEXO-E415G mutation have cross-resistance, parasite drug sensitivity was also assessed against a panel of antimalarial drugs (Table 3). No significant difference in IC50 values between B5 and B5-rexo-E415G-B8 parasites were observed for most of antimalarial drugs except for atovaquone (ATG) and proguanil (PG), where higher IC50 values were detected in comparison with the B5 parental line. The ASAP-21 parasite had significantly lower IC50 values for quinine (QN), chloroquine (CQ), atovaquone (ATQ) and doxycycline (DOX) but higher IC50 value for pyronaridine (PND) compared to the B5 parental line, while the ASAP-168 parasite had increased drug sensitivity against dihydroartemisinin (DHA), mefloquine (MQ), QN, and ATQ. The reduced IC50 values of QN and ATQ were observed in both ASAP-21 and ASAP-168 parasites compared to the B5 parental line.
Table 3.
In vitro susceptibility of P. falciparum, parental (B5), pfexo-modified (B5-rexo-E415G-B8) and adapted clinical parasites to dihydroartemisinin (DHA), artesunate (AS), mefloquine (MQ), quinine (QN), chloroquine (CQ), atovaquone (ATQ), lumefantrine (LUM), doxycycline (DOX), tafenoquine (TQ), cycloguanil (CYC), primaquine (PQ), proguanil (PG), and pyronaridine (PND). (mean ± SD, nM)
| Drug | IC50 (nM) | ||||
|---|---|---|---|---|---|
| B5 | B5-r exo-E415G-B8 | ASAP-21 | ASAP-168 | P-value | |
| DHA | 9 ± 4 | 14 ± 6 | 5 ± 0.2 | 2 ± 1 | 0.1213 |
| AS | 7 ± 5 | 4 ± 1 | 3 ± 0.4 | 3 ± 1 | – |
| MQ | 161 ± 58 | 82 ± 37 | 61 ± 23 | 50 ± 20 | 0.0483 |
| QN | 546 ± 180 | 564 ± 165 | 71 ± 3 | 138 ± 32 | 0.0172,3 |
| CQ | 562 ± 218 | 416 ± 219 | 64 ± 12 | 256 ± 59 | 0.0172 |
| ATQ | 48 ± 38 | 114 ± 1 | 5 ± 0.6 | 4 ± 1 | 0.0171,2,3 |
| LUM | 10 ± 8 | 4 ± 1 | 2 ± 0.2 | 3 ± 1 | – |
| DOX | 13,523 ± 2337 | 14,649 ± 1071 | 8527 ± 349 | 10,920 ± 1061 | 0.0242 |
| TQ | 263 ± 143 | 315 ± 45 | 189 ± 13 | 274 ± 554 | – |
| CYC | 2063 ± 2703 | 705 ± 15 | 7183 ± 388 | N.D. | – |
| PQ | 3094 ± 2015 | 6387 ± 218 | 5720 ± 1160 | 2458 ± 61 | – |
| PG | 12,522 ± 7247 | 23,816 ± 3786 | 2114 ± 243 | 3643 ± 473 | 0.0331 |
| PND | 28 ± 16 | 25 ± 4 | 50 ± 2 | 12 ± 6 | 0.0332 |
P-value calculated by Mann–Whitney U Test
1,2,3Significant differences between data from B5-rexo-E415G-B8, ASAP-21, and ASAP-168 compared to B5 line, respectively
N.D. for not determined
To gain a better understanding of the ART and PPQ resistance phenotypes, the survival assay (RSA0–3 h and PSA0–3 h) and PPQ-bimodal dose response curve were performed (Fig. 2). For RSA0–3 h (Fig. 2A), P. falciparum W2 was used as a control of ART-sensitive parasites, while IPC-4884 and IPC-5202 parasites were controls for ART-resistant parasites. The ASAP-21 and ASAP-168 parasites, containing the PfK13-C580Y mutation, exhibited % RSA survival rate of greater than 1, while B5 and B5-rexo-E415G-B8 parasites, carrying wild-type PfK13, exhibited % RSA survival rate of less than 1, clearly validating the correlation between PfK13-C580Y mutation and ART resistance. The % PSA survival rate of the modified pfexo transgenic lines and clinical isolates were next examined (Fig. 2B). PPQ-sensitive parasites P. falciparum W2, IPC-4884 and IPC-5202 were used as controls. The transgenic B5-rexo-E415G-B8 parasites harbouring merely PfEXO-E415G mutation had a % PSA survival rate of less than 10 (a cut-off for PPQ resistance), similar to that of the parental B5 parasites. This suggested that the presence of PfEXO-E415G mutation alone could not confer the PPQ resistance phenotype. The ASAP-21 and ASAP-168 parasites, containing the PfEXO-E415G mutation in combination with either PfCRT mutation or multiple copies of pfpm2, exhibited a % PSA survival rate higher than 10, indicative of PPQ-resistance.
Fig. 2.

Phenotypic analysis of pfexo transgenic lines and clinical isolates. (A-B) In vitro RSA0-3h and PSA0-3h survival rates, respectively, for standard laboratory-adapted clones (W2 for an ART-sensitive control, IPC-4884 and IPC-5202 for ART-resistance control), B5 (parental line, B5-rexo-E415G-B8 (modified pfexo transgenic line) and culture-adapted clinical isolates (ASAP-21 and ASAP-168). The dashed line represents the 1% survival rate cut-off that differentiates ART-resistance (≥ 1%) from ART-sensitive (< 1%) parasites in RSAs or the 10% survival rate cut-off that distinguishes PPQ-resistance (≥ 10%) from PPQ-sensitive (< 10%) parasites in PSAs. Two independent biological replicates were performed. Zero values of % survival rate were plotted as 0.001 % in logarithmic scale. (C-F) PPQ-bimodal dose-response curves of B5, B8, ASAP-21, and ASAP-168, respectively. Increasing the starting concentration and number of data points (24 points) for HRP2 ELISA dose-response curve provided a bimodal distribution of parasite response to PPQ exposure for PPQ-resistant parasites. ASAP-21 and ASAP-168 shows a second peak in PPQ concentration 0.01- 100 µM. The concentration range used to calculate the area under the curve (AUC) is indicated. Data are shown as mean values from three biological replicates with S.D
In addition to PSA0–3 h, PPQ-resistant parasites have been reported to exhibit a bimodal dose-response curve with a second peak between 0.01 and 100 µM and that the AUC correlates with the degree of PPQ resistance [16]. B5 and B5-rexo-E415G-B8 parasites did not exhibit the bimodal dose-response curve, confirming the PPQ-sensitive phenotype, whereas both ASAP-21 and ASAP-168 parasites clearly showed the bimodal dose-response curve with the AUC of 1029 and 596, respectively (Fig. 3C–F). The AUC of these parasites was in good agreement with both IC50 and IC90 values in that ASAP-21 parasites are more resistant to PPQ than ASAP-168 parasites.
Fig. 3.
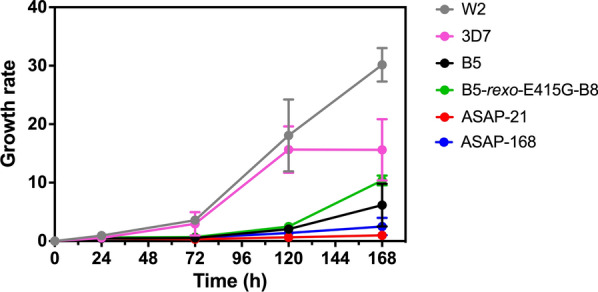
Analysis of growth rates. P. falciparum parasites with PfEXO wild-type (W2, 3D7 and B5), PfEXO-E415G transgenic line (B5-rexo-E415G-B8), and clinical isolates with the combination of PfEXO-E415G and multiple pfpm2 copies (ASAP-21 and ASAP-168) were synchronized (initiated at 0.1% parasitaemia ring stage, 2% haematocrit) and followed for 168 hours. Nucleic acids were stained with SYBR Green I and parasitaemia assessed by flow cytometry. Raw values were corrected, and data represent mean ± S.D. from 3 independent experiments
Growth assay
To assess whether the introduction of E415G mutation into PfEXO affects the parasite fitness, growth assays of B5-rexo-E415G-B8 parasites was carried out in comparison with W2, 3D7, B5 and the other two culture adapted clinical isolates (ASAP-21 and ASAP-168) (Fig. 3). Parasite lines originating from Cambodia (i.e., B5, B5-rexo-E415G-B8, ASAP-21 and ASAP-168) grew much more slowly than W2 (an Indochina clone exhibiting CQ resistance) and 3D7 (African origin) lines. Parasite B5-rexo-E415G-B8, harbouring pfexo-E415G mutation, seemed to grow slightly faster than its parental line B5. In the presence of PfK13-C580Y, PfEXO-E415G, novel PfCRT-F145I mutations and multiple copy number of pfpm2, ASAP-21 parasite had a severely retarded growth rate. ASAP-168, having the similar genetic background to ASAP-21 but without the PfCRT-F145I mutation, grew marginally better than ASAP-21 parasites. This observation implies an important contribution of the strain background toward parasite fitness, especially when PfK13-C580Y, PfCRT-F145I mutations and multiple copy number of pfpm2 were present.
Discussion
Validated molecular markers, such as pfk13 for ART resistance, have been widely employed as identification and prediction tools for the emergence of drug-resistant P. falciparum [50]. While a molecular marker of PPQ resistant P. falciparum malaria has been reported [13, 15, 16, 19, 20, 29], there is still a pressing need for validating potential new or additive molecular markers to determine effects on PPQ as a partner drug in ACT. This study found that the E415G mutation in the gene pfexo cannot alone reduce PPQ susceptibility neither improve parasite survival rates that are characteristic of PPQ resistant isolates circulating in the Greater Mekong Subregion [17].
Exonucleases are essential to genome stability, catalysing the removal of a single nucleotide monophosphate (dNMP) from the end of one strand of DNA and acting as a proof-reader during DNA replication [51]. Exonucleases are highly conserved and can be classified into families based on both sequence and functional homology, including the 5′–3′ exo C-terminal domain (CTD) superfamily, the RNAaseH domain superfamily i.e., DnaQ-like family and other 3′–5′ exonucleases. The pfexo gene from P. falciparum (PF3D7_1362500) encodes a protein with a mass of 86.7 kDa with a 3′–5′ exonuclease domain at the C-terminus. In this study, the P. falciparum B5 line used for genome editing had one mutation (K614N) in exonuclease, located in the 3′–5′ exonuclease domain. Since no in vivo relevance of this mutation has yet been proved in this study, it is suggested that the prevalence of this mutation should be determined. Nucleases may be partially or fully redundant, depending on the pathway, and such redundancy might complement functional losses. Zhang et al. [52] used bioinformatics to predict eight putative RNA exosome-associated proteins in the P. falciparum genome, including exoribonuclease functional domain-containing proteins Dis3 and Rrp6, the latter of which was found in PfEXO of this study.
CRISPR/Cas9 approaches have transformed the speed and scale with which Plasmodium genome editing can be achieved, and the approach has been employed to introduce point mutations into several P. falciparum genes [53]. In this study, instead of using P. falciparum 3D7 or Dd2 lines, the Cambodia adapted-B5 line [24] was employed with the hypothesis that the B5 line has a close genetic background to the currently circulating P. falciparum in Cambodia. It has been previously shown that different phenotypes were observed when using different parasite backgrounds. Targeted gene disruption of either pfpm2 or pfpm3 in the 3D7 genetic background caused a only slight decrease in PPQ susceptibility [54] and pfpm2 and pfpm3 overexpression in 3D7 did not alter the sensitivity of P. falciparum to PPQ [55]. However, when P. falciparum Dd2 parasites with copy number variation in pfpm2 were generated [29], pfpm2 amplification contributed to PPQ resistance, with a bimodal dose-response observed. In addition to a qPCR assay to detect pfpm2 copy number variation, the breakpoint assay, as described by Imwong et al. [56], was carried out. It was found that in parasites with a pfpm2/3 copy number above the cut-off of 1.52, 88% were confirmed to have pfpm2/3 amplification by the breakpoint assay, while for those < 1.14 (the cut-off for a single copy number) and for an intermediate value between 1.14 and 1.52, the proportion of pfpm2/3 amplified parasites using the breakpoint SNP were 4% and 38%, respectively. In this study, the ASAP-168 parasite was found to have pfpm2 copy number variation of 1.11, lower than the cut-off for a single copy number, but the breakpoint assay confirmed the gene duplication of pfpm2. As for pfpm3 copy number variation, the detection by qPCR was not 100% concordant with and pfpm2/3 breakpoint assay for B5 and B5-rexo-E415G-B8 lines as qPCR assay showed the pfpm3 multiple copy for Cambodia isolates, which was different from the previous report by Ansbro et al. [45]. It is not lost on the authors that different primers were used to identify the copy number of pfpm3 and further investigation could be done. Therefore, it is important to also apply the breakpoint assay to isolates with copy number values < 1.52 for pfpm2, to capture all isolates with pfpm2/3 amplification.
IC90 values, PPQ survival rates, and bimodal dose-response curves were used for assessing PPQ resistance in vitro [16, 17, 48]. All three assays confirmed that the engineered parasites harbouring the E415G-PfEXO mutation (B5-rexo-E415G-B8) did not show a PPQ resistant phenotype. On the contrary, parasites with the combination of PfEXO-E415G mutation with either novel PfCRT mutations or pfpm2 multiple copies demonstrated reduced IC90 susceptibility, high PPQ survival rates, and a second peak of bimodal curve (Fig. 3). If the PfEXO-E415G mutation does not alter the PPQ susceptibility, this finding also recapitulates the work of Silva et al. [29] that parasites with the pfpm2 multiple copy alone (i.e. ASAP-168) show reduced PPQ susceptibility and involved in PPQ resistance. Nonetheless, when the novel PfCRT mutation was added on top of the pfpm2 multiple copies, the level of PPQ resistance increased tremendously as judged by the IC50 and IC90 values as well as AUC between 0.01 and 100 µM of PPQ in the bimodal dose-response curve. Bopp et al. [16] showed that when exposed up to 10 µM PPQ for 12 h, PPQ resistance parasites could survive and complete their lifecycle. It was also previously evident that the introduction of novel PfCRT mutations resulted in a fitness cost for the mutations [19, 20]. Similar trends were observed in this study; parasites holding either novel PfCRT mutations or pfpm2 multiple copies grew much slower than those without mutations. It is likely that introducing PfEXO-E415G mutations may account for better fitness compared to parasites with the same genetic background. Even though the scope of this work could not address the function of PfEXO, it is evidently showed that the E415G mutation in PfEXO does not alter PPQ susceptivity, but rather affects parasite fitness.
Conclusions
In summary, this study suggests that the E415G mutation in PfEXO could still be an important marker if used in conjunction with other markers. The insertion of the PfEXO-E415G mutation did not lead to an increased PPQ-IC90 or improve %PPQ survival, suggesting that this mutation is not associated with PPQ resistance. Additionally, this specific mutation resulted in parasites that grew better than those with the same background, highlighting the importance of genetic mutations toward parasite fitness and resistance.
Supplementary Information
Additional file 1: Table S1. Oligonucleotideprimer sequences used in this study. Figure S1. DNA sequence alignment of exo gene from P. falciparum 3D7,B5 and B5-rexo-E415G-B8 parasites. Figure S2. Gene and amino acid composition of PfEXO. Figure S3. Schematic of P. falciparum plasmepsin 2/3 (pfpm2/3)gene duplication.
Acknowledgements
The following reagents were obtained through BEI Resources, NIAID, NIH: Plasmodium falicaprum, Strain IPC-4884 (MRA-1238) and IPC-5202 (MRA-1240), contributed by Didier Ménard. We would like to thank Professor Michael J Blackman (The Francis Crick Institute, UK) for providing the pDC2-Cas9-hDHFR-yFCU plasmid as well as insightful comments and suggestions, Dr. Ellen Knuepfer (The Royal Veterinary College, UK) for PkpSKIP_Pk47 plasmid, and Dr. Michele D. Spring (USAMD-AFRIMS) for valuable comments and suggestions. The authors would like to thank the clinical and laboratory staff from Molecular and Immunology section at AFRIMS-BPD who made the study possible.
Disclaimer
Material has been reviewed by the Walter Reed Army Institute of Research. There is no objection to its presentation/publication. The opinions or assertions contained herein are the private views of the author, and are not to be construed as official, or as reflecting true views of the Department of the Army or the Department of Defense. The investigators have adhered to the policies for protection of human subjects as prescribed in AR 70-25.
Author contributions
NB conceived, designed, and executed the study. BAV, MW, JSG, and NCW conceived, and supported the study. NB constructed all plasmids used in this study. NB and KK performed transfection, cloning and selection processes. KK, CT and CP performed cell culture, drug susceptibility assay, survival assay. CP performed cell growth assay and flow cytometry analysis. NB wrote the first draft. All authors read and approved the final manuscript.
Funding
This work was supported by Defense Malaria Assistance Program (DMAP). The funding source had no role in the analysis or interpretation of data, preparation of the manuscript or the decision to publish.
Availability of data and materials
All data generated or analysed during this study are included in this published article and its Additional files.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.WHO . World malaria report. Geneva: World Health Organization; 2019. [Google Scholar]
- 2.Noedl H, Se Y, Schaecher K, Smith BL, Socheat D, Fukuda MM, et al. Evidence of artemisinin-resistant malaria in western Cambodia. N Engl J Med. 2008;359:2619–2620. doi: 10.1056/NEJMc0805011. [DOI] [PubMed] [Google Scholar]
- 3.Dondorp AM, Nosten F, Yi P, Das D, Phyo AP, Tarning J, et al. Artemisinin resistance in Plasmodium falciparum malaria. N Engl J Med. 2009;361:455–467. doi: 10.1056/NEJMoa0808859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Amaratunga C, Sreng S, Suon S, Phelps ES, Stepniewska K, Lim P, et al. Artemisinin-resistant Plasmodium falciparum in Pursat province, western Cambodia: a parasite clearance rate study. Lancet Infect Dis. 2012;12:851–858. doi: 10.1016/S1473-3099(12)70181-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.WHO . Guidelines fro the treatment of malaria. 3. Geneva: World Health Organization; 2015. [PubMed] [Google Scholar]
- 6.Saunders DL, Vanachayangkul P, Lon C, Program USAMMR. National Center for Parasitology Entomology and Malaria Control et al. Dihydroartemisinin–piperaquine failure in Cambodia. N Engl J Med. 2014;371:484–485. doi: 10.1056/NEJMc1403007. [DOI] [PubMed] [Google Scholar]
- 7.Leang R, Taylor WR, Bouth DM, Song L, Tarning J, Char MC, et al. Evidence of Plasmodium falciparum malaria multidrug resistance to artemisinin and piperaquine in Western Cambodia: dihydroartemisinin–piperaquine open-label multicenter clinical assessment. Antimicrob Agents Chemother. 2015;59:4719–4726. doi: 10.1128/AAC.00835-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Spring MD, Lin JT, Manning JE, Vanachayangkul P, Somethy S, Bun R, et al. Dihydroartemisinin–piperaquine failure associated with a triple mutant including kelch13 C580Y in Cambodia: an observational cohort study. Lancet Infect Dis. 2015;15:683–691. doi: 10.1016/S1473-3099(15)70049-6. [DOI] [PubMed] [Google Scholar]
- 9.Amaratunga C, Lim P, Suon S, Sreng S, Mao S, Sopha C, et al. Dihydroartemisinin–piperaquine resistance in Plasmodium falciparum malaria in Cambodia: a multisite prospective cohort study. Lancet Infect Dis. 2016;16:357–365. doi: 10.1016/S1473-3099(15)00487-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Phuc BQ, Rasmussen C, Duong TT, Dong LT, Loi MA, Menard D, et al. Treatment failure of dihydroartemisinin/piperaquine for Plasmodium falciparum malaria, Vietnam. Emerg Infect Dis. 2017;23:715–717. doi: 10.3201/eid2304.161872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.van der Pluijm RW, Imwong M, Chau NH, Hoa NT, Thuy-Nhien NT, Thanh NV, et al. Determinants of dihydroartemisinin–piperaquine treatment failure in Plasmodium falciparum malaria in Cambodia, Thailand, and Vietnam: a prospective clinical, pharmacological, and genetic study. Lancet Infect Dis. 2019;19:952–961. doi: 10.1016/S1473-3099(19)30391-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Witkowski B, Amaratunga C, Khim N, Sreng S, Chim P, Kim S, et al. Novel phenotypic assays for the detection of artemisinin-resistant Plasmodium falciparum malaria in Cambodia: in-vitro and ex-vivo drug-response studies. Lancet Infect Dis. 2013;13:1043–1049. doi: 10.1016/S1473-3099(13)70252-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Amato R, Lim P, Miotto O, Amaratunga C, Dek D, Pearson RD, et al. Genetic markers associated with dihydroartemisinin–piperaquine failure in Plasmodium falciparum malaria in Cambodia: a genotype–phenotype association study. Lancet Infect Dis. 2017;17:164–173. doi: 10.1016/S1473-3099(16)30409-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Parobek CM, Parr JB, Brazeau NF, Lon C, Chaorattanakawee S, Gosi P, et al. Partner-drug resistance and population substructuring of artemisinin-resistant Plasmodium falciparum in Cambodia. Genome Biol Evol. 2017;9:1673–1686. doi: 10.1093/gbe/evx126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Witkowski B, Duru V, Khim N, Ross LS, Saintpierre B, Beghain J, et al. A surrogate marker of piperaquine-resistant Plasmodium falciparum malaria: a phenotype–genotype association study. Lancet Infect Dis. 2017;17:174–183. doi: 10.1016/S1473-3099(16)30415-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bopp S, Magistrado P, Wong W, Schaffner SF, Mukherjee A, Lim P, et al. Plasmepsin II–III copy number accounts for bimodal piperaquine resistance among Cambodian Plasmodium falciparum. Nat Commun. 2018;9:1769. doi: 10.1038/s41467-018-04104-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Duru V, Khim N, Leang R, Kim S, Domergue A, Kloeung N, et al. Plasmodium falciparum dihydroartemisinin–piperaquine failures in Cambodia are associated with mutant K13 parasites presenting high survival rates in novel piperaquine in vitro assays: retrospective and prospective investigations. BMC Med. 2015;13:305. doi: 10.1186/s12916-015-0539-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Agrawal S, Moser KA, Morton L, Cummings MP, Parihar A, Dwivedi A, et al. Association of a novel mutation in the Plasmodium falciparum chloroquine resistance transporter with decreased piperaquine sensitivity. J Infect Dis. 2017;216:468–476. doi: 10.1093/infdis/jix334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ross LS, Dhingra SK, Mok S, Yeo T, Wicht KJ, Kumpornsin K, et al. Emerging Southeast Asian PfCRT mutations confer Plasmodium falciparum resistance to the first-line antimalarial piperaquine. Nat Commun. 2018;9:3314. doi: 10.1038/s41467-018-05652-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Dhingra SK, Small-Saunders JL, Menard D, Fidock DA. Plasmodium falciparum resistance to piperaquine driven by PfCRT. Lancet Infect Dis. 2019;19:1168–1169. doi: 10.1016/S1473-3099(19)30543-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Thanh NV, Thuy-Nhien N, Tuyen NT, Tong NT, Nha-Ca NT, Dong LT, et al. Rapid decline in the susceptibility of Plasmodium falciparum to dihydroartemisinin–piperaquine in the south of Vietnam. Malar J. 2017;16:27. doi: 10.1186/s12936-017-1680-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Diakite SAS, Traore K, Sanogo I, Clark TG, Campino S, Sangare M, et al. A comprehensive analysis of drug resistance molecular markers and Plasmodium falciparum genetic diversity in two malaria endemic sites in Mali. Malar J. 2019;18:361. doi: 10.1186/s12936-019-2986-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Robert MG, Tsombeng FF, Gendrot M, Diawara S, Madamet M, Kounta MB, et al. Baseline ex vivo and molecular responses of Plasmodium faciparum isolates to piperaquine before implementation of dihydroartemisinin–piperaquine in Senegal. Antimicrob Agents Chemother. 2019;63:e02445-18. doi: 10.1128/AAC.02445-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Boonyalai N, Vesely BA, Thamnurak C, Praditpol C, Fagnark W, Kirativanich K, et al. Piperaquine resistant Cambodian Plasmodium falciparum clinical isolates: in vitro genotypic and phenotypic characterization. Malar J. 2020;19:269. doi: 10.1186/s12936-020-03339-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Quang HH, Chavchich M, Trinh NTM, Edgel KA, Edstein MD, Martin NJ. Multidrug-resistant Plasmodium falciparum parasites in the Central Highlands of Vietnam jeopardize malaria control and elimination strategies. Antimicrob Agents Chemother. 2021;65:e01639-20. doi: 10.1128/AAC.01639-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Si Y, Zeng W, Li N, Wang C, Siddiqui F, Zhang J, et al. In vitro susceptibility of Plasmodium falciparum isolates from the China-Myanmar border area to piperaquine and association with candidate markers. Antimicrob Agents Chemother. 2021;65:e02305-20. doi: 10.1128/AAC.02305-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Phong NC, Chavchich M, Quang HH, San NN, Birrell GW, Chuang I, et al. Susceptibility of Plasmodium falciparum to artemisinins and Plasmodium vivax to chloroquine in Phuoc Chien Commune, Ninh Thuan Province, south-central Vietnam. Malar J. 2019;18:10. doi: 10.1186/s12936-019-2640-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hussien M, Abdel Hamid MM, Elamin EA, Hassan AO, Elaagip AH, Salama AHA, et al. Antimalarial drug resistance molecular makers of Plasmodium falciparum isolates from Sudan during 2015–2017. PLoS ONE. 2020;15:e0235401. doi: 10.1371/journal.pone.0235401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Silva M, Calcada C, Teixeira M, Veiga MI, Ferreira PE. Multigenic architecture of piperaquine resistance trait in Plasmodium falciparum. Lancet Infect Dis. 2020;20:26–27. doi: 10.1016/S1473-3099(19)30689-9. [DOI] [PubMed] [Google Scholar]
- 30.Madeira F, Park YM, Lee J, Buso N, Gur T, Madhusoodanan N, et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019;47:W636–W641. doi: 10.1093/nar/gkz268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Almagro Armenteros JJ, Tsirigos KD, Sonderby CK, Petersen TN, Winther O, Brunak S, et al. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat Biotechnol. 2019;37:420–423. doi: 10.1038/s41587-019-0036-z. [DOI] [PubMed] [Google Scholar]
- 32.Blum M, Chang HY, Chuguransky S, Grego T, Kandasaamy S, Mitchell A, et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 2021;49:D344–D354. doi: 10.1093/nar/gkaa977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Trager W, Jensen JB. Human malaria parasites in continuous culture. Science. 1976;193:673–675. doi: 10.1126/science.781840. [DOI] [PubMed] [Google Scholar]
- 34.Saul A, Myler P, Elliott T, Kidson C. Purification of mature schizonts of Plasmodium falciparum on colloidal silica gradients. Bull World Health Organ. 1982;60:755–759. [PMC free article] [PubMed] [Google Scholar]
- 35.Harris PK, Yeoh S, Dluzewski AR, O’Donnell RA, Withers-Martinez C, Hackett F, et al. Molecular identification of a malaria merozoite surface sheddase. PLoS Pathog. 2005;1:241–251. doi: 10.1371/journal.ppat.0010029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lambros C, Vanderberg JP. Synchronization of Plasmodium falciparum erythrocytic stages in culture. J Parasitol. 1979;65:418–420. doi: 10.2307/3280287. [DOI] [PubMed] [Google Scholar]
- 37.Knuepfer E, Napiorkowska M, van Ooij C, Holder AA. Generating conditional gene knockouts in Plasmodium—a toolkit to produce stable DiCre recombinase-expressing parasite lines using CRISPR/Cas9. Sci Rep. 2017;7:3881. doi: 10.1038/s41598-017-03984-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Knuepfer E, Wright KE, Kumar Prajapati S, Rawlinson TA, Mohring F, Koch M, et al. Divergent roles for the RH5 complex components, CyRPA and RIPR in human-infective malaria parasites. PLoS Pathog. 2019;15:e1007809. doi: 10.1371/journal.ppat.1007809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Collins CR, Das S, Wong EH, Andenmatten N, Stallmach R, Hackett F, et al. Robust inducible Cre recombinase activity in the human malaria parasite Plasmodium falciparum enables efficient gene deletion within a single asexual erythrocytic growth cycle. Mol Microbiol. 2013;88:687–701. doi: 10.1111/mmi.12206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Janse CJ, Franke-Fayard B, Mair GR, Ramesar J, Thiel C, Engelmann S, et al. High efficiency transfection of Plasmodium berghei facilitates novel selection procedures. Mol Biochem Parasitol. 2006;145:60–70. doi: 10.1016/j.molbiopara.2005.09.007. [DOI] [PubMed] [Google Scholar]
- 41.Thomas JA, Collins CR, Das S, Hackett F, Graindorge A, Bell D, et al. Development and application of a simple plaque assay for the human malaria parasite, Plasmodium falciparum. PLoS ONE. 2016;11:e0157873. doi: 10.1371/journal.pone.0157873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ariey F, Witkowski B, Amaratunga C, Beghain J, Langlois AC, Khim N, et al. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature. 2014;505:50–55. doi: 10.1038/nature12876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Talundzic E, Okoth SA, Congpuong K, Plucinski MM, Morton L, Goldman IF, et al. Selection and spread of artemisinin-resistant alleles in Thailand prior to the global artemisinin resistance containment campaign. PLoS Pathog. 2015;11:e1004789. doi: 10.1371/journal.ppat.1004789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Pelleau S, Moss EL, Dhingra SK, Volney B, Casteras J, Gabryszewski SJ, et al. Adaptive evolution of malaria parasites in French Guiana: reversal of chloroquine resistance by acquisition of a mutation in pfcrt. Proc Natl Acad Sci USA. 2015;112:11672–11677. doi: 10.1073/pnas.1507142112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Ansbro MR, Jacob CG, Amato R, Kekre M, Amaratunga C, Sreng S, et al. Development of copy number assays for detection and surveillance of piperaquine resistance associated plasmepsin 2/3 copy number variation in Plasmodium falciparum. Malar J. 2020;19:181. doi: 10.1186/s12936-020-03249-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Chaorattanakawee S, Tyner SD, Lon C, Yingyuen K, Ruttvisutinunt W, Sundrakes S, et al. Direct comparison of the histidine-rich protein-2 enzyme-linked immunosorbent assay (HRP-2 ELISA) and malaria SYBR green I fluorescence (MSF) drug sensitivity tests in Plasmodium falciparum reference clones and fresh ex vivo field isolates from Cambodia. Malar J. 2013;12:239. doi: 10.1186/1475-2875-12-239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Chaorattanakawee S, Saunders DL, Sea D, Chanarat N, Yingyuen K, Sundrakes S, et al. Ex vivo drug susceptibility testing and molecular profiling of clinical Plasmodium falciparum isolates from Cambodia from 2008 to 2013 suggest emerging piperaquine resistance. Antimicrob Agents Chemother. 2015;59:4631–4643. doi: 10.1128/AAC.00366-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Chaorattanakawee S, Lon C, Jongsakul K, Gawee J, Sok S, Sundrakes S, et al. Ex vivo piperaquine resistance developed rapidly in Plasmodium falciparum isolates in northern Cambodia compared to Thailand. Malar J. 2016;15:519. doi: 10.1186/s12936-016-1569-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wojnarski M, Lon C, Vanachayangkul P, Gosi P, Sok S, Rachmat A, et al. Atovaquone-proguanil in combination with artesunate to treat multidrug-resistant P. falciparum malaria in Cambodia: an open-label randomized trial. Open Forum Infect Dis. 2019;6:ofz314. doi: 10.1093/ofid/ofz314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Plowe CV, Roper C, Barnwell JW, Happi CT, Joshi HH, Mbacham W, et al. World Antimalarial Resistance Network (WARN) III: molecular markers for drug resistant malaria. Malar J. 2007;6:121. doi: 10.1186/1475-2875-6-121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mason PA, Cox LS. The role of DNA exonucleases in protecting genome stability and their impact on ageing. Age (Dordr) 2012;34:1317–1340. doi: 10.1007/s11357-011-9306-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zhang Q, Siegel TN, Martins RM, Wang F, Cao J, Gao Q, et al. Exonuclease-mediated degradation of nascent RNA silences genes linked to severe malaria. Nature. 2014;513:431–435. doi: 10.1038/nature13468. [DOI] [PubMed] [Google Scholar]
- 53.Lee MCS, Lindner SE, Lopez-Rubio JJ, Llinas M. Cutting back malaria: CRISPR/Cas9 genome editing of Plasmodium. Brief Funct Genom. 2019;18:281–289. doi: 10.1093/bfgp/elz012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Mukherjee A, Gagnon D, Wirth DF, Richard D. Inactivation of plasmepsins 2 and 3 sensitizes Plasmodium falciparum to the antimalarial drug piperaquine. Antimicrob Agents Chemother. 2018;62:e02309-17. doi: 10.1128/AAC.02309-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Loesbanluechai D, Kotanan N, de Cozar C, Kochakarn T, Ansbro MR, Chotivanich K, et al. Overexpression of plasmepsin II and plasmepsin III does not directly cause reduction in Plasmodium falciparum sensitivity to artesunate, chloroquine and piperaquine. Int J Parasitol Drugs Drug Resist. 2018;9:16–22. doi: 10.1016/j.ijpddr.2018.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Imwong M, Suwannasin K, Srisutham S, Vongpromek R, Promnarate C, Saejeng A, et al. Evolution of multidrug resistance in Plasmodium falciparum: a longitudinal study of genetic resistance markers in the Greater Mekong Subregion. Antimicrob Agents Chemother. 2021;65:e0112121. doi: 10.1128/AAC.01121-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Table S1. Oligonucleotideprimer sequences used in this study. Figure S1. DNA sequence alignment of exo gene from P. falciparum 3D7,B5 and B5-rexo-E415G-B8 parasites. Figure S2. Gene and amino acid composition of PfEXO. Figure S3. Schematic of P. falciparum plasmepsin 2/3 (pfpm2/3)gene duplication.
Data Availability Statement
All data generated or analysed during this study are included in this published article and its Additional files.