

ABSTRACT
Mitophagy and energy production are two functionalities in which PINK1 plays a key role. Loss of PINK1 is one of the genetic causes of Parkinson disease (PD), suggesting both processes are important in PD pathogenesis. Nonetheless, it remains unclear whether these processes are connected or independent of one another. Sphingolipids, including ceramide, have recently emerged as an important new player in the development of PD, however, how alterations in ceramide levels are mechanistically linked to PD remained elusive. In a recently published study, we demonstrated that ceramide accumulates in mitochondria and initiates ceramide-induced mitophagy, thereby compensating for the lack of PINK1-dependent mitophagy upon PINK1 deficiency. However, ceramide accumulation negatively affects ß-oxidation, further aggravating the electron transport chain (ETC) defect caused by PINK1 deficiency and resulting in an additional requirement for mitophagy. Thus, we showed that ceramide serves as a link between the ETC and mitophagy upon PINK1 deficiency. Interruption of this vicious cycle via stimulation of ß-oxidation or reduction of ceramide levels can provide a novel therapeutic target in the treatment of PINK1-related PD.
KEYWORDS: ß-oxidation, ceramide, mitophagy, Parkinson’s disease, PINK1
PINK1 acts together with PRKN as a key regulator of mitophagy, the autophagic clearance of defective mitochondria. Interestingly, homozygous loss of PINK1 or PRKN results in Parkinson disease (PD), suggesting mitophagy to be an important mechanism affected in PD. However, both PINK1 and PRKN play a role in a plethora of cellular processes, challenging the identification of the exact role or defects of mitophagy in PD pathogenesis. PINK1 is a key element for an efficient transfer of electrons in the mitochondrial electron transfer chain (ETC) via PINK1-dependent phosphorylation of complex I, which facilitates electron transfer. However, if and how these processes of energy metabolism and mitophagy are linked mechanistically remained elusive.
Our recent study demonstrated that the energy deficit caused by PINK1 dysfunction, in conjunction with impaired PINK1-dependent mitophagy, leads to relocalization of ceramide to mitochondria to increase mitochondrial ceramide levels and, subsequently, ceramide-induced mitophagy [1] (Figure 1). Ceramide is the basic sphingolipid and plays important roles in signal transmission, apoptosis, autophagy and mitophagy. Furthermore, ceramide synthesis occurs via different pathways and, hence, several enzymes can be targeted to affect ceramide levels. We took advantage of available drugs and genetic manipulation to lower ceramide levels in pink1-mutant flies. Remarkably and in agreement with our hypothesis, reduction of ceramide levels improves the Pink1-dependent phenotypes. Alleviated ATP levels and improved ETC efficiency upon reduction of ceramide suggest that mitochondrial ceramide accumulation is negatively correlated with ETC function. In the presence of decreased ceramide levels, mitochondria form fewer accumulations or enlargements facilitating mitophagy which is in keeping with the notion that ceramide accumulates in mitochondria to induce mitophagy. This process, however, is accompanied by the unwanted side effect that it negatively affects energy production. These findings collectively imply that ceramide provides a link between the ETC and mitophagy upon loss of Pink1.
Figure 1.
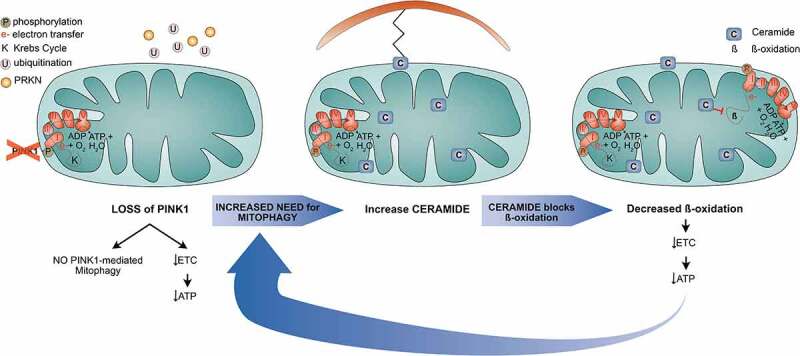
Simplified scheme of the proposed mechanism. Loss of PINK1 results in less efficient electron transfer and thus in lower energy production. This, in turn, leads to defective mitochondria that need to be cleared. However, due to lack of PINK1, PRKN cannot be recruited to the mitochondria to label mitochondrial proteins with ubiquitin for autophagic degradation. As a consequence, ceramide accumulates at the mitochondria to provoke ceramide-induced mitophagy. However, mitochondrial ceramide is negatively correlated with ß-oxidation and thus decreases ß-oxidation levels and subsequently the ETC efficiency and ATP levels further, thereby increasing the need for mitochondrial degradation, which ends in a vicious cycle.
Intriguingly, elevated mitochondrial ceramide levels induce impaired ß-oxidation corroborating a negative correlation between ß-oxidation and ceramide levels. Likewise, our data showed lower ß-oxidation levels following Pink1 deficiency. Decreased ceramide levels restore ß-oxidation levels, thereby confirming this effect on ß-oxidation to be ceramide-dependent. ß-Oxidation provides substrates originating from the breakdown of fatty acids that enter the Krebs cycle or the ETC. In a defective ETC due to loss of Pink1, an additional ceramide-dependent decrease in ß-oxidation further impairs energy production. To test if induction of ß-oxidation is sufficient to restore pink1-mutant phenotypes, we supplemented the medium with carnitine, a direct stimulator of ß-oxidation. Indeed, carnitine treatment improves Pink1-deficient phenotypes, suggesting that boosting ß-oxidation is a potential therapeutic target in the treatment of PINK1-dependent PD.
Accumulation of mitochondrial ceramide provokes ceramide-induced mitophagy and we discovered that loss of Pink1 induces the formation of LC3 puncta that colocalize with ceramide. These LC3 disappear upon reduction of ceramide, highlighting that the observed LC3 phenotypes of Pink1 deficiency are likewise ceramide dependent. In our effort to understand the underlying mechanism in relation to the impaired mitochondria, we monitored the effect of ceramide on mitophagy using the mtKeima construct. Mitophagy levels are initially not affected in pink1-mutant flies, which implies an alternative approach is activated to meet the mitophagy requirements. We provided experimental data that ceramide accumulation is responsible for the maintenance of the initial basal mitophagy levels in Pink1-deficient flies, confirming that ceramide accumulates in the mitochondria in mammalian cells in an attempt to overcome the lack of PINK1-dependent mitophagy.
The effects of loss of Pink1 in flies are well characterized and display similar defects as those observed in PINK1-mutant cells derived from patients. Importantly, previous studies showed that PINK1 function is evolutionarily conserved from flies to human. Also, our newly identified mechanism in which ceramide is redistributed to mitochondria and in which phenotypes can be rescued by lowering ceramide levels or via the stimulation of ß-oxidation, was confirmed in PINK1-deficient patient-derived fibroblasts.
To conclude, ceramide levels are increased and redistributed to mitochondria upon loss of PINK1 to provoke ceramide-induced mitophagy in an attempt to meet the increased requirement for mitochondrial clearance resulting from the PINK1-dependent impairment of the ETC. However, elevated ceramide levels negatively affect ß-oxidation, which prompts a further decline in ETC efficiency. Thus, an additionally increased demand to clear defective mitochondria is generated, finally converging into a vicious cycle. Interruption of this cycle via stimulation of ß-oxidation or decrease of ceramide levels is a potential novel therapeutic strategy in PINK1-related PD.
Funding Statement
Support was provided by European Molecular Biology Organization stipend to M. Vos (ALTF 851-2014), an Else-Kröner-Fresenius-Stiftung grant (2017_A56) and intramural funding from the University of Lübeck to M. Vos. C. Klein is supported by intramural funding from the University of Lübeck.
Disclosure statement
CK serves as medical consultant to Centogene for genetic testing in the field of movement disorders and dementia excluding Parkinson’s disease and serves on the scientific advisory board of Retromer Therapeutics.
Reference
- [1].Vos M, Dulovic-Mahlow M, Mandik F, et al. Ceramide accumulation induces mitophagy and impairs β-oxidation in PINK1 deficiency. Proc Natl Acad Sci. 2021;118:e2025347118. Available from: https://www.pnas.org/content/118/43/e2025347118 [DOI] [PMC free article] [PubMed] [Google Scholar]