

Abstract
Knowledge of the growth and maturation of human antral follicles is based mainly on concepts and deductions from clinical observations and animal models. To date, new experimental approaches and in vitro data contributed to a deep comprehension of gonadotropin receptors’ functioning and may provide new insights into the mechanisms regulating still unclear physiological events. Among these, the production of androgen in the absence of proper LH levels, the programming of follicular atresia and dominance are some of the most intriguing. Starting from evolutionary issues at the basis of the gonadotropin receptor signal specificity, we draw a new hypothesis explaining the molecular mechanisms of the antral follicular growth, based on the modulation of endocrine signals by receptor-receptor interactions. The “heteromer hypothesis” explains how opposite death and life signals are delivered by gonadotropin receptors and other membrane partners, mediating steroidogenesis, apoptotic events, and the maturation of the dominant follicle.
Keywords: Antral follicle, FSH, LH, GPER, Heteromers, Atresia
Evolutionary insights in gonadotropins physiology
Human reproductive functions are supported by gonadotropins, which are glycoprotein hormones inducing the synthesis of sex steroids and gametogenesis during the fertile age. Two of these molecules, the follicle-stimulating (FSH) and the luteinizing hormone (LH), are produced by the pituitary, upon stimulation by gonadotropin-releasing hormone (GnRH) and bind specific membrane receptors (FSHR; LHCGR) [1, 2]. These molecules belong to the class A, or “rhodopsin-like,” superfamily of G protein-coupled receptors (GPCRs), expressed in the gonads of both males and females. A third gonadotropin, the choriogonadotropin (CG), induces progesterone synthesis required to support fetal growth in females. While CG exists only in primates, FSH and LH are present in vertebrates and in a large part of invertebrates, together with the structurally similar thyroid-stimulating hormone (TSH), suggesting that they have been overall conserved across evolution [3]. All these hormones have a dimeric structure and share a common α subunit, while they have similar, but unique β-subunits specifically binding the receptor. Considering the overall sequence identity of the β-subunits, which is about 40% among FSH, LH, and TSH, and 80% between hCG and LH, it was suggested that these hormones evolved from a common ancestral gene and underwent duplications over time [4, 5]. These differences would be due to gene mutations gradually accumulated during evolution, and particular interest was given to genes coding the unique LH and the several hCG β-subunits of primates (LHB and CGBs), which are eight in humans, including two pseudogenes [3]. Studies comparing these gene sequences suggested that CGBs evolved by repeated duplications of an ancestral LHB gene. Interestingly, CGB genes have a nucleotide sequence extension, likely due to the loss of the original stop codon [6], resulting in additional 24 amino acids in the hCGβ protein. This carboxyl-terminal peptide carries additional glycosylations, resulting in specific hormone activities and biochemical properties [7–11], such as a longer half-life than LH [12]. Reflecting a certain grade of similarity, glycoprotein hormone receptors share 45% of sequence identity in the transmembrane domain and 70% in the extracellular portion that binds the ligand [4].
From an evolutionary point of view, the specificity of ligand-hormone binding played a fundamental role in determining the separation of endocrine functions [13]. While genes encoding glycoprotein hormone subunits and receptors are present in all metazoan species, ligand-receptor signaling complexes do not imply the separation of specific endocrine signals in all of them [14]. It means that physiological events, such as growth, osmoregulation, and reproduction, are not under the control of distinct hypothalamic-pituitary endocrine axes in the early metazoan [13]. The complexity of neuroendocrine control specificity increases in tetrapods and, especially, in mammals, where reproductive, thyroid, and other functions rely on highly refined and separated pathways, diversified through high specificity of glycoprotein hormone receptor signaling modules. One of the highest levels of complexity of the gonadotropin receptor system is achieved in humans and closest primates, where reproductive functions are regulated by three gonadotropins, which manage specific physiological events acting with relatively high specificity through two different receptors [15]. This issue suggests that human reproductive functions are characterized by an extremely fine regulation of the gametogenic program, which, however, relies on few key amino acid residues of gonadotropin receptors. An example is provided by mutations of a couple of amino acids in the LHCGR extracellular domain, falling in close proximity to interaction sites with the hormone, leading to loss of LH- and hCG-specific signals [16]. In fact, point mutations of LHCGR and FSHR hotspots for ligand binding or signal transduction were described as causes of reproductive pathological conditions, such as ovarian failure, ovarian hyperstimulation syndrome, or miscarriage [16–20], due to cross-interaction between gonadotropins and other receptors. Similar conclusions may be achieved in light of cases of hCG hypersensitivity in familial gestational hyperthyroidism, where high levels of the chorionic hormone may lead to interference with thyroid function due to cross-interaction between hCG and specific TSHR variants [21]. Given these considerations, we may conclude that a limited number of key amino acid residues work as a specificity barrier to avoid promiscuous activation of receptors and redundant endocrine signals [4], while the control of human reproductive functions requires specificity of ligand-receptor binding.
The human “three-gonadotropin” system would optimize the specialization of endocrine signals, especially in the ovary, where they modulate unique, not completely replaceable, and sometimes opposite functions. Despite molecular issues conferring high specificity to the hormone receptor binding [22–24], it is worth noting that gonadotropin receptors, as well as TSHR, induce the activation of largely similar intracellular signaling modules in target cells [3, 25–29]. These pathways consist in a qualitatively similar early signaling signature, overall mediated by the same molecules. For instance, depending on Gαs protein receptor coupling, gonadotropin binding results in intracellular cyclic adenosine monophosphate (cAMP) increase and protein kinase A (PKA) activation. Coupling to Gαi protein results in rapid phosphorylation of extracellularly regulated kinases 1 and 2 (ERK1/2), which may be sustained over time by receptor interactors β-arrestins. Gq protein coupling is linked to phospholipase C (PLC) activation and intracellular calcium ion (Ca2+) increase, while several other interactors would induce protein kinase B (AKT) phosphorylation, mammalian target of rapamycin (mTOR) activation, receptor recycling, and degradation. Other effector molecules common to gonadotropin receptors are known, increasing the complexity of the intracellular signaling networks activated by different ligand-receptor complexes [3, 26–29]. Interestingly, besides these known concepts from the classical gonadotropin receptor signaling, recent advances revealed a new mode of actions of class A GPCRs, including biased and intracellular activation, transactivation, dimerization, and biphasic behavior, providing a more complex view of the endocrine regulation [30]. These modes of action occur differently in the two sexes and are the new frontier in the comprehension of gonadotropin receptor functioning and of unknown aspects of human gonadal physiology.
Human gametogenesis is regulated by a complex network of signals, which converge into strictly stage-specific steroidogenic, apoptotic, and mitogenic signals. Although gonadotropins regulate gonadal functions and reproduction of both sexes, there are differences in the regulation of hormone-dependent signals between men and women (Fig. 1). In the male reproductive system, FSH acts on Sertoli cells activating mitogenic signals delivered mainly through cAMP/PKA, ERK1/2, and AKT, resulting in trophic effects and physical support of gamete maturation [31]. However, FSH seems not to play necessarily a major role in spermatogenesis since the administration of gonadotropin is beneficial only for a limited number of infertile patients, mainly idiopathic [32]. Rather, the synthesis of testosterone by Leydig cells is a requisite to support spermatogenesis [33, 34], a function that, in men, is committed to LH via activation mainly of cAMP/PKA-, ERK1/2-, and protein kinase C (PKC)-pathways [35–37]. Therefore, differently from FSH, proper LH action is fundamental to maintain spermatogenesis. In women, both gonadotropins are required to support oocyte selection and growth and to induce ovulation. The functional unit of the ovary is the follicle, which supports oocyte development through a complex crosstalk between different cell components, such as granulosa and theca cells, as well as a fine endocrine regulation operated by hormones and growth factors [38–40]. The follicular response to FSH begins during the recruitment of a pool of secondary follicles, becoming antral, which is accompanied by increasing levels of FSHR mRNA transcripts in granulosa cells [41] (Fig. 2). During this stage, FSH is essential to mediate cAMP/PKA-dependent steroidogenic signals required to produce estrogens, which are proliferative and anti-apoptotic factors acting in follicular cells [45, 46]. Estrogens exert negative feedback in the pituitary, inhibiting the secretion of FSH, which is replaced by LH. At this stage, the decline of FSH levels, occurring in the presence of estrogens, is assumed to be responsible for atresia, a degenerative process involving non-dominant follicles based on apoptotic events [47]. Follicular dominance is characterized by the shift from FSH to LH dependence, supported by the expression of increasing LHCGR transcripts. During this stage, and differently from what occurs in the testis, FSHR and LHCGR are co-expressed in granulosa cells [41] and may interact displaying a unique and physiologically relevant behavior supporting the mid-antral follicle maturation [48, 49]. The periovulatory stage of the menstrual cycle is characterized by marked LH responsivity of follicular cells, resulting in relatively marked ERK1/2 and AKT activation, over cAMP production, progesterone synthesis, and metabolic state changes of granulosa cells, becoming lutein after ovulation [50].
Fig. 1.

Differences between male and female gonadal late gametogenesis. In the testis, FSHR and LHCGR are expressed in Sertoli and Leydig cells, respectively. While Sertoli cells have mainly trophic functions, Leydig cells synthesize androgens fundamental to support spermatogenesis. In the ovary, at the antral stage, theca cells produce androgens under LH stimulation, while FSHR and LHCGR are co-expressed in granulosa cells, which produce estrogens and support oocyte maturation
Fig. 2.
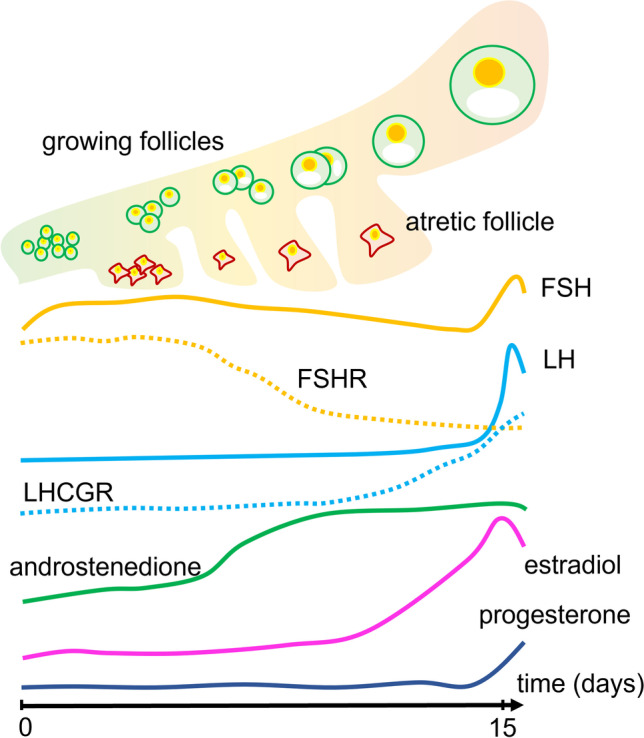
Hormone and receptor levels during the antral follicle stage. Follicle growth and atresia are accompanied by variations of gonadotropin levels (orange and light-blue lines) and receptors (dotted lines). Changes in sex steroid hormone levels are represented by green (androstenedione), pink (estradiol), and blue (progesterone) lines. Timeline is to be referred to the first day of the menstrual cycle (day 0) until ovulation (day 15) (adapted and merged from [42–44])
Receptor-receptor interactions impact early antral follicle functions
Although hormone-induced signaling could be simplistically described as the result of one activated receptor that independently triggers intracellular signals, the issue is more complex and other membrane molecules may cooperate in the modulation of endocrine signals [51]. As several GPCRs [52, 53], gonadotropin receptors are known to act as dimers or oligomers, consisting of two or more interacting molecular units [1, 2, 54, 55]. Under this conformation, one receptor bound to the hormone may transactivate the partner receptor, which would be the effector that triggers signal transduction [30, 56]. Moreover, one of the two partner receptors may act as an allosteric modulator of the other, altering its pharmacological and signal activation profiles [57–59]. For gonadotropin receptors, these concepts were demonstrated by in vitro studies, which found that they may form structural complexes possibly ameliorating the deleterious effect of heterozygous mutations [60]. Interestingly, oligomers may consist of assemblies of different receptors, forming heteromers [61]. Although the existence of class A receptor heteromers in vivo is debated [61], an increasing number of studies revealed that FSHR and LHCGR may form heteromeric complexes in vitro [48, 62–64]. The existence of functional units of assembled FSHR/LHCGR poses a new hypothesis to explain how antral follicle development is regulated.
Classically, it is known that FSH induces the expression of LHCGR in the ovary, via upregulation of LHCGR promoter activity [65]. These signals would lead to increasing LHCGR protein expression levels along with antral follicle maturation, possibly detected in early small antral follicles [66], although the presence of LHCGR was suggested also in secondary follicles [66]. These data would require further confirmation, given the known, sub-optimal anti-LHCGR/FSHR antibody specificity [67]. In any case, they suggest that the early antral stage is characterized by very low LHCGR expression levels [41, 66, 68, 69], pointing out the marginal role of LH in activating the synthesis of androgens during the window of FSH dependence. However, androgens are the substrate to produce estrogens [70], which induce major trophic effects required to support antral follicle growth [45, 71–75]. These hormones would be produced by LHCGR-expressing theca cells. However, these cells are very poorly represented at the early antral stage, when the theca layer and granulosa cells are not fully differentiated into two mature and distinct androgen- and estrogen-producing functional units, respectively [76]. Although speculative, this hypothesis suggests that, at the secondary to antral follicle transition, androgenic functions could be supported by pre-thecal cells expressing both granulosa and theca cell transcripts, such as FSHR, LHCGR, CYP19A1, and CYP17A1 [77–79]. On the one hand, these data must be considered cautiously since they were obtained from non-human models. On the other hand, they suggest that functional LHCGR should be expressed at the early antral stage and be able to mediate the androgenic signal, even in the absence of sufficient LH molecules. A theory based on the “spare” LH receptors concept was largely used to explain how androgen synthesis might occur in the ovary, even in the absence of proper LH levels [80]. This hypothesis explains that maximal activation of the target effect, i.e. the synthesis of androgens, occurs at relatively low levels of receptor occupancy [81]. However, there is no evidence for androgenic support by “spare” LHCGRs in the human ovary [49], given that in vivo data are missing, and that LH has a weak steroidogenic potential [82, 83]. Rather, intermolecular cooperation and physical interactions between gonadotropin receptors were found in vitro and, at least in part, in vivo [62, 84–87]. Some of these experiments demonstrated that ligand-binding deficient receptors may be transactivated by signaling-deficient receptors bound to the hormone [88, 89]. These data are suggestive of the existence of LH-like stimuli delivered through LHCGR, upon heteromerization with FSHR bound to FSH (Fig. 3). This hypothesis is strengthened by the fact that, in the early antral stage, FSHR expression is 100-fold higher than LHCGR [41], favoring LHCGR to form heteromers with largely available FSHRs. Moreover, it might explain why androgen production is particularly elevated during controlled ovarian stimulation for assisted reproduction, compared to a natural cycle, in the presence of supraphysiological (exogenous) FSH levels stimulating immature early antral follicles [90]. Finally, our hypothesis is in line with data from patients with inactivating LHB mutations; they do not impair follicle progression from secondary to early antral transition, neither androgen nor estrogen production in the early antral stage [91]. In those cases, functional LHCGRs may form heteromers with FSHRs, likely supporting the secondary to early antral follicle functions. Although this view provides new insights on how gonadotropin receptors may support the antral follicular growth, there are limitations to be considered; among all, no reports describe FSHR/LHCGR heteromers in vivo, due to missing methods reliably detecting oligomers in human living tissues. Most of the data about gonadotropin receptor heteromers were from in vitro studies, where transfected cell models overexpressing both FSHR and LHCGR were used, thus providing an environment favoring interactions between membrane proteins. However, further support may come from mice, even if it they are multiovulatory species and some features of their ovarian physiology could not find a parallel in humans. In female mice, gonadotropin production was depleted by hypophysectomy, obtaining folliculogenesis blockade. Interestingly, follicle maturation, growth, and ovulation were restored upon administration of exogenous FSH [92–96], suggesting that these activities were supported completely by the follitropin, which acts through Fshr interacting with Lhr in the gonads. This was not observed in Lhr knockout mice, where exogenous FSH and hCG failed in recovering antral folliculogenesis and ovulation [92]. Taken together, these data converge on the requirement of functional LHCGR, rather than LH production, to support androgenic signals in the early antral follicle.
Fig. 3.
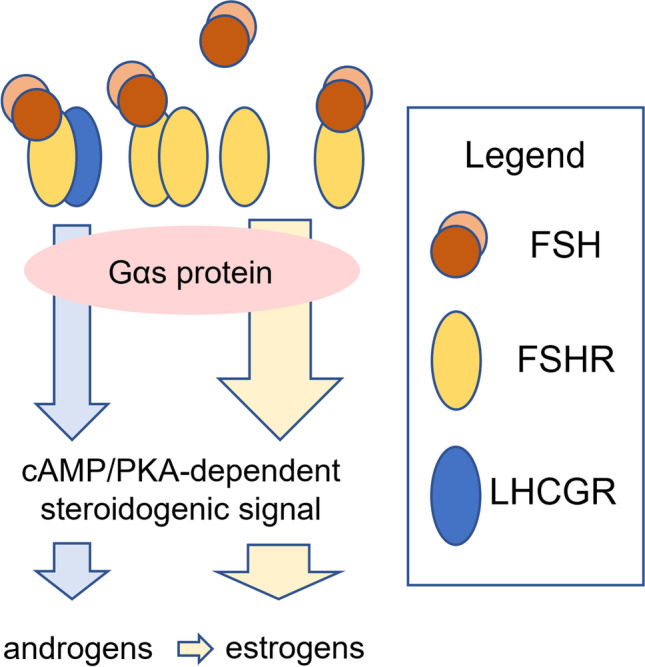
Androgen production during the early antral stage. Poorly expressed LHCGRs heteromerize with FSHRs, cooperating to induce LH-like stimuli through FSH binding to its receptor. Blue arrows indicate LH-like stimuli, while yellow arrows are FSH-dependent steroidogenic signals
FSHR-specific signals regulate follicular atresia in the early antral stage
FSH plays a central role during the early/mid ovarian antral stage, when FSHRs are maximally expressed in granulosa cells [41] and possibly in the oocyte [68]. FSHR could be expressed even in preantral follicles, although relatively weakly, where the hormone could impact early follicle growth via activation of signals upregulating cell proliferation and differentiation [97]. However, the physiology of the ovarian antral stage is not completely clear, and still, there are missing points, including the role of gonadotropins and their receptors. For instance, while it is well known that FSH supports antral follicle growth-stimulating estrogen synthesis in granulosa cells, recent studies suggest that FSH and its receptor could be responsible for the dominant follicle selection in humans [98, 99]. This function would be exerted via activation of pro-apoptotic signals not fully known in multi-follicular species like rodents [100]. In humans, the selection of the dominant follicle occurs at the days 4–5 of the menstrual cycle, traditionally supposed to be due to declining FSH levels in response to increased estrogen production. From this point of view, FSH would exert exclusive, and essential life signals directly induced in granulosa cells and, likely, the oocytes, supporting the growth of antral follicles [101]. This concept is strengthened by the evidence provided by clinical treatments for couple infertility, which may consist in the controlled ovarian stimulation with exogenous FSH, resulting in multi-follicular growth [102]. However, most of these considerations are affected by limitations; they do not consider that endocrine signals are delivered thanks to a couple of actors, i.e., the hormone and its receptor, thus ignoring the relevance of prolonged, high FSHR expression levels during the antral stage. Moreover, ovarian follicles are exposed to proliferative factors, such as growth factors and sex steroids, which hardly allow identifying the proliferative role of FSH in vivo.
In the ovary, the highest number of FSHR mRNA transcripts is detected in small antral follicles [41, 97], concomitantly with high FSH levels, and shortly preceding atresia of non-dominant follicles. Therefore, selection of the dominant follicle may be temporarily associated with more than one physiological event: on the one hand, the classical knowledge of ovarian physiology provides that atresia is due to declining FSH levels. On the other hand, follicle selection is a process that might be triggered few hours before the exacerbation of the atretic process, and in the presence of relatively high FSHR levels. The latter view reflects two events likely occurring in vivo and demonstrated in vitro: (I) the action of FSH in the ovary is linked to downregulation of FSHR mRNA transcripts [103–106] and (II) pro-apoptotic signals may be delivered through overexpressed FSHRs [98, 99]. Taken together, the model explaining the human dominant follicle selection may be updated by revisiting the role of FSHR, which would have a biphasic behavior, in light of recent in vitro data. At the preantral stage, relatively low FSHR expression levels [97] might be fundamental to recruit a pool of follicles and to lead them into the antral stage. In this case, FSHR would boost life signals, possibly through the activation of kinases, such as ERK1/2, which are related to proliferative signals [107–109]. Although clinical data supporting this hypothesis are not available, the rationale is provided by in vitro studies. They demonstrated that ERK1/2 activators, such as β-arrestins and Gαi proteins [110], are preferentially recruited when FSHR is expressed at low levels [98, 111], triggering follicular maturation.
Another event related to FSH action is steroidogenesis, which is upregulated via cAMP/PKA activation, leading to the transcription of genes coding steroidogenic enzymes [112]. Estrogens are proliferative factors required to support antral follicular growth and are synthesized upon the action of FSH through FSHR. In fact, the receptor is maximally expressed during the early antral stage [113], allowing to effectively deliver the cAMP/PKA-dependent steroidogenic signal, and differently to what likely occurs at the preantral stage, which is a physiological window dependent on ERK1/2 and other kinases. The functional differences between these two stages could rely on the biphasic behavior of FSHR, which is preferentially coupled to the Gαs protein when it is overexpressed [98]. This is fundamental for FSHR to trigger adequate cAMP increase, which likely occurs even in the absence of the ligand, due to the high constitutive activity of overexpressed receptors [99]. Interestingly, while these intracellular events are fundamental to provide enough estrogens to support follicular growth, they are strictly linked to the activation of death signals. FSH induces the activation of the p38 mitogen-activated kinase (p38 MAPK) via a cAMP/PKA-dependent mechanism [114]. In pre-ovulatory granulosa cells, p38 MAPK, together with other enzymes, such as the Jun N-terminal kinase (JNK), is associated with cytoskeletal rearrangements and apoptosis [115]. These signals are extremely stage-specific and connect steroidogenesis with pro-apoptotic stimuli. Thus, it is not surprising that intracellular cAMP increase was associated with pro-apoptotic protein p53 activation [116] and that not consistent, steroidogenic human cell lines, permanently overexpressing FSHR exist so far [49, 99, 117]. Most importantly, these issues provide the rationale for FSHR as a determinant of follicular atresia, which occurs as a “dark side” of the steroidogenic process. Interestingly, while follicular cells are collapsing, the production of an adequate amount of sex steroid hormones would be ensured by the compartmentalization of steroidogenic processes into organelles [118]. This issue points out the relevance of cytoskeletal rearrangements occurring in granulosa cells [115], supporting apoptosis and steroidogenic activity at the same time.
New insights on the dominant follicle rescuing
The expression of FSHRs is a transitory event in ovarian cells. The number of these receptors declines from the early/mid-antral stage on, when LHCGR transcripts increase [41], together with serum LH levels, achieving the maximal expression in the luteal phase [41, 119]. This evidence is suggestive of cAMP-dependent, pro-apoptotic signals linked to FSHR overexpression, detailed in the previous section, which should be inhibited immediately after the programming of atretic follicles to avoid exaggerated cell death. At the molecular level, this mechanism could rely on the replacement of FSHR by LHCGR, whose expression is upregulated by several factors, such as FSH [65]. In fact, upon LH binding, LHCGR upregulates ovarian cell proliferation and survival via activation of ERK1/2- and AKT-dependent signals [46, 82, 83, 99, 120, 121]. Especially, the ERK1/2 anti-apoptotic role was described even in the dominant follicle [122] and it is not surprising that ovarian granulosa cell death is linked to reduced ERK1/2 activity [123, 124]. Therefore, ERK1/2- and cAMP/PKA-pathways may exert stage-specific and opposite roles in granulosa cells, although they may be connected by intracellular crosstalk [26, 27].
While the above considerations could explain how life signals support follicle growth, the molecular mechanism underlying follicular dominance is still widely unclear. To date, it is explained with the “negative feedback model,” in which estrogens would suppress the proliferative role of FSH [47]. This model was provided by observations in vivo, indicating that the blockade of estrogenic effects by antibodies or clomiphene citrate would increase FSH levels and the number of growing antral follicles [125–127]. Under physiological conditions, FSH levels decline at the early/mid-antral stage and the dominant follicle would be able to survive at low hormone concentration, due to the highest number of expressed FSHRs [47]. However, these views are likely affected by confounding factors intrinsic to the in vivo model since it is conflicting with the known proliferative effects modulated by estrogens [46, 128, 129], as well as with the requirement of these sex steroids to support vital functions of the growing dominant follicle [73, 130, 131]. Moreover, they are in contrast with the pro-apoptotic functions associated with FSHR overexpression and could be revised considering recent in vitro data, which bring new insights on the programming of follicular dominance as a molecular mechanism that, once again, could rely on receptor-receptor interactions occurring in ovarian cells. These data are based on the finding that a class A sex steroid GPCR (GPER) [132] functions as a transmembrane partner of FSHR [98]. It is a membrane receptor responsible for the “non-genomic” effects of estrogens [133] and associated with proliferative events in the ovary [134–136]. FSHR/GPER may form heteromers on the granulosa cell surface, with the preferential assembly of complexes containing a higher number of FSHR than GPER molecules [98]. In response to FSH binding, these functional units inhibit cAMP signaling and shift the FSHR signal to AKT activation, upregulating cell survival and reducing apoptosis [98]. This mechanism is underpinned by a GPER-associated inhibitory protein complex [137], which blocks the activation of the Gαs protein signaling linked to FSHR [98]. Interestingly, since the membrane estrogen receptor had no effect on LHCGR-mediated cAMP, the blockade of Gαs protein signaling is specific to FSHR [98]. While these data reflect structural differences and physiological roles between FSHR and LHCGR [46, 83], they are suggestive of survival signals occurring in the dominant follicle. From this point of view, the follicular fate could rely on the capability of GPER to block FSHR-mediated apoptosis during the early/mid-antral stage before the fall of follitropin receptor transcripts (Fig. 4). Later, FSHR is replaced by LHCGR expression after the dominant follicle selection [41], when anti-apoptotic and proliferative, rather than steroidogenic signals, are activated by LH.
Fig. 4.

“Receptor heteromer” hypothesis of the antral follicle selection. The very early antral stage, as likely secondary follicles, is characterized by a relatively low number of FSHRs, which are preferentially coupled to intracellular interactors mediating FSH-induced proliferative signals crucial for the proliferation of granulosa cells. The programming of follicular dominance relies on the overexpression of FSHR in follicles becoming atretic, deputed to the synthesis of sex steroids upon FSH stimulation. Estrogens are required to support vital functions of the growing dominant follicle. The latter is rescued from atresia by FSHR/GPER heteromers, shifting FSH-induced stimuli from death to AKT-dependent survival signals. While estrogen synthesis continues during the atretic process, via compartmentalization of steroidogenic signals, LHCGRs replace FSHRs in the dominant follicle
The ability of FSHR/GPER heteromers to reprogram cAMP/death signals into proliferative stimuli, sustaining oocyte survival, may have pathophysiological implications. First, a high FSHR:GPER mRNA expression ratio negatively impacts follicular maturation in vivo, leading to a poor ovarian response to controlled ovarian stimulation cycles [41]. The treatment consists of the clinical induction of multi-follicular maturation by exogenous administration of FSH [138] in women considered as “normo-” or “poor-responders,” depending on the number of oocytes obtained [139, 140]. Since low oocyte yield is correlated to a higher FSHR:GPER ratio, we may hypothesize that a low number of GPER molecules negatively impact cell survival, due to ineffective counterbalancing of FSHR-induced death signals [41]. Moreover, the proliferative potential retained by these heteromers may upregulate the growth of hormone-dependent cancer cells expressing the two receptors [136, 141] and represent a potential target for a specific drug or disruptive agents [142].
The later stage is dominated by the presence of LHCGR, which accompanies the maturation of the dominant follicle until ovulation upon the action of increasing LH levels [41]. This hormone has a relatively short half-life (about 20 min) [3], likely optimizing proliferative and anti-apoptotic pulses operated through ERK1/2 and AKT phosphorylation, rather than through cAMP [82, 83, 120]. Interestingly, the activation of these signals could require LHCGR internalization into intracellular compartments, as a key feature of the receptor mode of action upon LH binding [143]. It is worth mentioning that, in the clinical context of controlled ovarian stimulation for assisted reproduction, hCG may be added to FSH and support the multi-follicular growth [138]. Since hCG is a natural ligand of LHCGR, it is commonly assumed that the choriogonadotropin may provide “LH-like” activity. To this purpose, hCG is successfully used in clinics. However, considering the differences existing between the two hormones [3], it is illogical to assume that hCG has the same activity as LH, even if, in granulosa cells, hCG activates an intracellular signaling pattern partially overlapping that of LH. While LH predominantly induces cell proliferation and survival and displays weak potency to induce steroidogenesis, hCG has relatively high efficacy in activating cAMP and exerts its main action in supporting progesterone production during pregnancy [82, 83, 120]. These characteristics reflect distinct biochemical and pharmacological properties of the two hormones, which evolved in primates to optimize specific physiological roles [62]. Therefore, the use of hCG during the antral stage should boost the steroidogenic signal, resulting indeed in a barely higher number of oocytes collected, due to enhanced anti-apoptotic signals related to the estrogenic action [46, 98], likely at the cost of lower quality than those obtained under an FSH + LH protocol [144]. While these considerations may not be easily confirmed by clinical studies, due to the requirement of relatively high sample size, these data underlie the irreplaceable role of LH during the mid/late antral stage.
Conclusions
We provided a new picture illustrating ovarian follicle recruitment, growth, and selection, where FSHR would play a key role throughout the antral stage. It would be responsible of androgen synthesis during the early antral stage, supporting LH-like stimuli delivered upon cooperation with LHCGR, as a requisite for estradiol production. These sex steroids are fundamental to support follicle growth during the antral stage and are induced by FSH. This hormone acts through overexpressed FSHR, which potently triggers cAMP/PKA activation, causing atresia. The early/mid-antral stage is characterized by the triggering of selective processes leading to the maturation of a single, dominant follicle, which is rescued by heteromerization of FSHR with the membrane estrogen receptor GPER, which shifts FSH-induced cAMP production towards AKT-dependent survival signals. In the later stage, FSHR is replaced by LHCGR, driving the dominant follicle through final maturation and ovulation, by ERK1/2- and AKT-dependent proliferative and anti-apoptotic signals triggered by an irreplaceable action of LH.
Acknowledgements
The authors are grateful to MUR for supporting the Department of Biomedical, Metabolic, and Neural Sciences (University of Modena and Reggio Emilia, Italy) in the context of the Departments of Excellence Programme. KZ and AGG were supported by a Trialect Corporation’s fellowship.
Declarations
Conflict of interest
The authors declare no competing interests.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Simoni M, Gromoll J, Nieschlag E. The follicle-stimulating hormone receptor: biochemistry, molecular biology, physiology, and pathophysiology. Endocr Rev. 1997;18:739–773. doi: 10.1210/edrv.18.6.0320. [DOI] [PubMed] [Google Scholar]
- 2.Ascoli M, Fanelli F, Segaloff DL. The lutropin/choriogonadotropin receptor, a 2002 perspective. Endocr Rev. 2002;23:141–174. doi: 10.1210/edrv.23.2.0462. [DOI] [PubMed] [Google Scholar]
- 3.Casarini L, Santi D, Brigante G, Simoni M. Two hormones for one receptor: evolution, biochemistry, actions, and pathophysiology of LH and hCG. Endocr Rev. 2018;39:549–592. doi: 10.1210/er.2018-00065. [DOI] [PubMed] [Google Scholar]
- 4.Costagliola S, Urizar E, Mendive F, Vassart G. Specificity and promiscuity of gonadotropin receptors. Reproduction. 2005;130:275–281. doi: 10.1530/rep.1.00662. [DOI] [PubMed] [Google Scholar]
- 5.Talmadge K, Vamvakopoulos NC, Fiddes JC. Evolution of the genes for the β subunits of human chorionic gonadotropin and luteinizing hormone. Nature. 1984;307:37–40. doi: 10.1038/307037a0. [DOI] [PubMed] [Google Scholar]
- 6.Fiddes JC, Goodman HM. The cDNA for the β-subunit of human chorionic gonadotropin suggests evolution of a gene by readthrough into the 3′-untranslated region. Nature. 1980;286:684–687. doi: 10.1038/286684a0. [DOI] [PubMed] [Google Scholar]
- 7.Furuhashi M, Suganuma N. Effect of additional N-glycosylation signal in the N-terminal region on intracellular function of the human gonadotropin alpha-subunit. Endocr J. 2003;50:245–253. doi: 10.1507/endocrj.50.245. [DOI] [PubMed] [Google Scholar]
- 8.Garcia-Campayo V, Sugahara T, Boime I. Unmasking a new recognition signal for O-linked glycosylation in the chorionic gonadotropin beta subunit. Mol Cell Endocrinol. 2002;194:63–70. doi: 10.1016/S0303-7207(02)00189-2. [DOI] [PubMed] [Google Scholar]
- 9.Shao K, Balasubramanian SV, Pope CM, Bahl OP. Effect of individual N-glycosyl chains in the beta-subunit on the conformation of human choriogonadotropin. Mol Cell Endocrinol. 1998;146:39–48. doi: 10.1016/S0303-7207(98)00194-4. [DOI] [PubMed] [Google Scholar]
- 10.Koistinen H, Koel M, Peters M, Rinken A, Lundin K, Tuuri T, et al. Hyperglycosylated hCG activates LH/hCG-receptor with lower activity than hCG. Mol Cell Endocrinol. 2019;479:103–109. doi: 10.1016/j.mce.2018.09.006. [DOI] [PubMed] [Google Scholar]
- 11.Van Loenen HJ, Van Gelderen-Boele S, Flinterman JF, Merz WE, Rommerts FFG. The relative importance of the oligosaccharide units in human chorionic gonadotropin (CG) for LH/CG receptor activation in rat Leydig cells and mouse Leydig tumor cells. J Endocrinol. 1995;147:367–375. doi: 10.1677/joe.0.1470367. [DOI] [PubMed] [Google Scholar]
- 12.Matzuk MM, Hsueh AJW, Lapolt P, Tsafriri A, Keene JL, Boime I, et al. The biological role of the carboxyl-terminal extension of human chorionic gonadotroin/3-subunit. Endocrinology. 1990;126:376–383. doi: 10.1210/endo-126-1-376. [DOI] [PubMed] [Google Scholar]
- 13.Sower SA, Freamat M, Kavanaugh SI. The origins of the vertebrate hypothalamic-pituitary-gonadal (HPG) and hypothalamic-pituitary-thyroid (HPT) endocrine systems: new insights from lampreys. Gen Comp Endocrinol. 2009;161:20–29. doi: 10.1016/j.ygcen.2008.11.023. [DOI] [PubMed] [Google Scholar]
- 14.Buechi HB, Bridgham JT. Evolution of specificity in cartilaginous fish glycoprotein hormones and receptors. Gen Comp Endocrinol. 2017;246:309–320. doi: 10.1016/j.ygcen.2017.01.007. [DOI] [PubMed] [Google Scholar]
- 15.Casarini L, Santi D, Marino M. Impact of gene polymorphisms of gonadotropins and their receptors on human reproductive success. Reproduction. 2015;150:R175–R184. doi: 10.1530/REP-15-0251. [DOI] [PubMed] [Google Scholar]
- 16.Lazzaretti C, Secco V, Paradiso E, Sperduti S, Rutz C, Kreuchwig A, et al. Identification of key receptor residues discriminating human chorionic gonadotropin (hCG)- and luteinizing hormone (LH)-specific signaling. Int J Mol Sci. 2020;22:1–14. doi: 10.3390/ijms22010151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bentov Y, Kenigsberg S, Casper RF. A novel luteinizing hormone/chorionic gonadotropin receptor mutation associated with amenorrhea, low oocyte yield, and recurrent pregnancy loss. Fertil Steril. 2012;97:1165–1168. doi: 10.1016/j.fertnstert.2012.02.002. [DOI] [PubMed] [Google Scholar]
- 18.Montanelli L, Van Durme JJJ, Smits G, Bonomi M, Rodien P, Devor EJ, et al. Modulation of ligand selectivity associated with activation of the transmembrane region of the human follitropin receptor. Mol Endocrinol. 2004;18:2061–2073. doi: 10.1210/me.2004-0036. [DOI] [PubMed] [Google Scholar]
- 19.Smits G, Olatunbosun O, Delbaere A, Pierson R, Vassart G, Costagliola S. Ovarian hyperstimulation syndrome due to a mutation in the follicle-stimulating hormone receptor. N Engl J Med. 2003;349:760–766. doi: 10.1056/NEJMoa030064. [DOI] [PubMed] [Google Scholar]
- 20.Vasseur C, Rodien P, Beau I, Desroches A, Gérard C, de Poncheville L, et al. A chorionic gonadotropin-sensitive mutation in the follicle-stimulating hormone receptor as a cause of familial gestational spontaneous ovarian hyperstimulation syndrome. N Engl J Med. 2003;349:753–759. doi: 10.1056/NEJMoa030065. [DOI] [PubMed] [Google Scholar]
- 21.Thachil J, Agarwal S. Understanding the COVID-19 coagulopathy spectrum. Anaesthesia. 2020;75:1432–1436. doi: 10.1111/anae.15141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ulloa-Aguirre A, Zariñán T, Jardón-Valadez E, Gutiérrez-Sagal R, Dias JA. Structure-function relationships of the follicle-stimulating hormone receptor. Front Endocrinol (Lausanne). 2018;9:707. doi: 10.3389/fendo.2018.00707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Banerjee AA, Mahale SD. Role of the Extracellular and intracellular loops of follicle-stimulating hormone receptor in its function. Front Endocrinol (Lausanne). 2015;6:110. doi: 10.3389/fendo.2015.00110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Puett D, Li Y, DeMars G, Angelova K, Fanelli F. A functional transmembrane complex: the luteinizing hormone receptor with bound ligand and G protein. Mol Cell Endocrinol. 2007;260–262:126–136. doi: 10.1016/j.mce.2006.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kleinau G, Neumann S, Grüters A, Krude H, Biebermann H. Novel insights on thyroid-stimulating hormone receptor signal transduction. Endocr Rev. 2013;34:691–724. doi: 10.1210/er.2012-1072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Casarini L, Crépieux P. Molecular mechanisms of action of FSH. Front Endocrinol (Lausanne). 2019;10:305. doi: 10.3389/fendo.2019.00305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Casarini L, Simoni M. Recent advances in understanding gonadotropin signaling. Fac Rev. 2021;10:41. doi: 10.12703/r/10-41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ulloa-Aguirre A, Reiter E, Crepieux P. FSH receptor signaling: complexity of interactions and signal diversity. Endocrinology. 2018;159:3020–3035. doi: 10.1210/en.2018-00452. [DOI] [PubMed] [Google Scholar]
- 29.Sayers N, Hanyaloglu AC. Intracellular follicle-stimulating hormone receptor trafficking and signaling. Front Endocrinol (Lausanne). 2018;9:653. doi: 10.3389/fendo.2018.00653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wang W, Qiao Y, Li Z. New insights into modes of GPCR activation. Trends Pharmacol Sci. 2018;39:367–386. doi: 10.1016/j.tips.2018.01.001. [DOI] [PubMed] [Google Scholar]
- 31.Casarini L, Crépieux P, Reiter E, Lazzaretti C, Paradiso E, Rochira V, et al. FSH for the treatment of male infertility. Int J Mol Sci. 2020;21:2270. doi: 10.3390/ijms21072270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Santi D, Crépieux P, Reiter E, Spaggiari G, Brigante G, Casarini L, et al. Follicle-stimulating hormone (FSH) action on spermatogenesis: a focus on physiological and therapeutic roles. J Clin Med. 2020;9:1014. doi: 10.3390/jcm9041014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Weiss J, Axelrod L, Whitcomb RW, Harris PE, Crowley WF, Jameson JL. Hypogonadism caused by a single amino acid substitution in the beta subunit of luteinizing hormone. N Engl J Med. 1992;326:179–183. doi: 10.1056/NEJM199201163260306. [DOI] [PubMed] [Google Scholar]
- 34.Latronico AC, Anasti J, Arnhold IJP, Rapaport R, Mendonca BB, Bloise W, et al. Brief report: testicular and ovarian resistance to luteinizing hormone caused by inactivating mutations of the luteinizing hormone-receptor gene. N Engl J Med. 1996;334:507–512. doi: 10.1056/NEJM199602223340805. [DOI] [PubMed] [Google Scholar]
- 35.Martinat N, Crépieux P, Reiter E, Guillou F. Extracellular signal-regulated kinases (ERK) 1, 2 are required for luteinizing hormone (LH)-induced steroidogenesis in primary Leydig cells and control steroidogenic acute regulatory (StAR) expression. Reprod Nutr Dev. 2005;45:101–108. doi: 10.1051/rnd:2005007. [DOI] [PubMed] [Google Scholar]
- 36.Manna PR, Jo Y, Stocco DM. Regulation of Leydig cell steroidogenesis by extracellular signal-regulated kinase 1/2: role of protein kinase A and protein kinase C signaling. J Endocrinol. 2007;193:53–63. doi: 10.1677/JOE-06-0201. [DOI] [PubMed] [Google Scholar]
- 37.Ulloa-Aguirre A, Lira-Albarrán S. Clinical applications of gonadotropins in the male. Prog Mol Biol Transl Sci. 2016;143:121–174. doi: 10.1016/bs.pmbts.2016.08.003. [DOI] [PubMed] [Google Scholar]
- 38.Reis FM, Cobellis L, Luisi S, Driul L, Florio P, Faletti A, et al. Paracrine/autocrine control of female reproduction. Gynecol Endocrinol. 2000;14:464–475. doi: 10.3109/09513590009167720. [DOI] [PubMed] [Google Scholar]
- 39.Poulsen LC, Bøtkjær JA, Østrup O, Petersen KB, Yding Andersen C, Grøndahl ML, et al. Two waves of transcriptomic changes in periovulatory human granulosa cells. Hum Reprod. 2020;35:1230–1245. doi: 10.1093/humrep/deaa043. [DOI] [PubMed] [Google Scholar]
- 40.Chen YJ, Hsiao PW, Lee MT, Mason JI, Ke FC, Hwang JJ. Interplay of PI3K and cAMP/PKA signaling, and rapamycin-hypersensitivity in TGFbeta1 enhancement of FSH-stimulated steroidogenesis in rat ovarian granulosa cells. J Endocrinol. 2007;192:405–419. doi: 10.1677/JOE-06-0076. [DOI] [PubMed] [Google Scholar]
- 41.Jeppesen JV, Kristensen SG, Nielsen ME, Humaidan P, Dal Canto M, Fadini R, et al. LH-receptor gene expression in human granulosa and cumulus cells from antral and preovulatory follicles. J Clin Endocrinol Metab. 2012;97:E1524–31. doi: 10.1210/jc.2012-1427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Stricker R, Eberhart R, Chevailler MC, Quinn FA, Bischof P, Stricker R. Establishment of detailed reference values for luteinizing hormone, follicle stimulating hormone, estradiol, and progesterone during different phases of the menstrual cycle on the Abbott ARCHITECT® analyzer. Clin Chem Lab Med. 2006;44:883–887. doi: 10.1515/CCLM.2006.160. [DOI] [PubMed] [Google Scholar]
- 43.Broekmans FJ. Individualization of FSH doses in assisted reproduction: facts and fiction. Front Endocrinol (Lausanne). 2019;10:181. [DOI] [PMC free article] [PubMed]
- 44.Khan DR, Fournier É, Dufort I, Richard FJ, Singh J, Sirard MA. Meta-analysis of gene expression profiles in granulosa cells during folliculogenesis. Reproduction. 2016;151:R103–10. doi: 10.1530/REP-15-0594. [DOI] [PubMed] [Google Scholar]
- 45.Palter SF, Tavares AB, Hourvitz A, Veldhuis JD, Adashi EY. Are estrogens of import to primate/human ovarian folliculogenesis? Endocr Rev. 2001;22:389–424. doi: 10.1210/edrv.22.3.0433. [DOI] [PubMed] [Google Scholar]
- 46.Casarini L, Riccetti L, De Pascali F, Gilioli L, Marino M, Vecchi E, et al. Estrogen modulates specific life and death signals induced by LH and hCG in human primary granulosa cells in vitro. Int J Mol Sci. 2017;18:926. doi: 10.3390/ijms18050926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zeleznik AJ. The physiology of follicle selection. Reprod Biol Endocrinol. 2004;2:31. doi: 10.1186/1477-7827-2-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Jonas KC, Hanyaloglu AC. Impact of G protein-coupled receptor heteromers in endocrine systems. Mol Cell Endocrinol. 2017;449:21–27. doi: 10.1016/j.mce.2017.01.030. [DOI] [PubMed] [Google Scholar]
- 49.Casarini L, Santi D, Simoni M, Potì F. “Spare” luteinizing hormone receptors: facts and fiction. Trends Endocrinol Metab. 2018;29:208–217. doi: 10.1016/j.tem.2018.01.007. [DOI] [PubMed] [Google Scholar]
- 50.Riccetti L, Sperduti S, Lazzaretti C, Casarini L, Simoni M. The cAMP/PKA pathway: steroidogenesis of the antral follicular stage. Minerva Ginecol. 2018;70:516–524. doi: 10.23736/S0026-4784.18.04282-X. [DOI] [PubMed] [Google Scholar]
- 51.Light A, Hammes SR. Membrane receptor cross talk in steroidogenesis: recent insights and clinical implications. Steroids. 2013;78:633–638. doi: 10.1016/j.steroids.2012.12.016. [DOI] [PubMed] [Google Scholar]
- 52.Fuxe K, Marcellino D, Borroto-Escuela DO, Frankowska M, Ferraro L, Guidolin D, et al. The changing world of G protein-coupled receptors: from monomers to dimers and receptor mosaics with allosteric receptor-receptor interactions. J Recept Signal Transduct Res. 2010;30:272–283. doi: 10.3109/10799893.2010.506191. [DOI] [PubMed] [Google Scholar]
- 53.Ferré S, Ciruela F, Casadó V, Pardo L. Oligomerization of G protein-coupled receptors: still doubted? Prog Mol Biol Transl Sci. 2020;169:297–321. doi: 10.1016/bs.pmbts.2019.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Jiang X, Fischer D, Chen X, McKenna SD, Liu H, Sriraman V, et al. Evidence for follicle-stimulating hormone receptor as a functional trimer. J Biol Chem. 2014;289:14273–14282. doi: 10.1074/jbc.M114.549592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Gomes I, Jordan BA, Gupta A, Rios C, Trapaidze N, Devi LA. G protein coupled receptor dimerization: implications in modulating receptor function. J Mol Med (Berl) 2001;79:226–242. doi: 10.1007/s001090100219. [DOI] [PubMed] [Google Scholar]
- 56.Jonas KC, Rivero-Müller A, Huhtaniemi IT, Hanyaloglu AC. G protein-coupled receptor transactivation: from molecules to mice. Methods Cell Biol. 2013;117:433–450. doi: 10.1016/B978-0-12-408143-7.00023-2. [DOI] [PubMed] [Google Scholar]
- 57.Agwuegbo UC, Jonas KC. Molecular and functional insights into gonadotropin hormone receptor dimerization and oligomerization. Minerva Ginecol. 2018;70:539–548. doi: 10.23736/S0026-4784.18.04287-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Milligan G, Smith NJ. Allosteric modulation of heterodimeric G-protein-coupled receptors. Trends Pharmacol Sci. 2007;28:615–620. doi: 10.1016/j.tips.2007.11.001. [DOI] [PubMed] [Google Scholar]
- 59.Han Y, Moreira IS, Urizar E, Weinstein H, Javitch JA. Allosteric communication between protomers of dopamine class A GPCR dimers modulates activation. Nat Chem Biol. 2009;5:688–695. doi: 10.1038/nchembio.199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Thomas RM, Nechamen CA, Mazurkiewicz JE, Muda M, Palmer S, Dias JA. Follice-stimulating hormone receptor forms oligomers and shows evidence of carboxyl-terminal proteolytic processing. Endocrinology. 2007;148:1987–1995. doi: 10.1210/en.2006-1672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Franco R, Martínez-Pinilla E, Lanciego JL, Navarro G. Basic pharmacological and structural evidence for class a g-protein-coupled receptor heteromerization. Front Pharmacol. 2016;7:76. [DOI] [PMC free article] [PubMed]
- 62.Casarini L, Riccetti L, Paradiso E, Benevelli R, Lazzaretti C, Sperduti S, et al. Two human menopausal gonadotrophin (hMG) preparations display different early signaling in vitro. Mol Hum Reprod. 2020;26:894–905. doi: 10.1093/molehr/gaaa070. [DOI] [PubMed] [Google Scholar]
- 63.Jonas KC, Chen S, Virta M, Mora J, Franks S, Huhtaniemi I, Hanyaloglu AC. Temporal reprogramming of calcium signalling via crosstalk of gonadotrophin receptors that associate as functionally asymmetric heteromers. Sci Rep. 2018;8:2239. doi: 10.1038/s41598-018-20722-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Feng X, Zhang M, Guan R, Segaloff DL. Heterodimerization between the lutropin and follitropin receptors is associated with an attenuation of hormone-dependent signaling. Endocrinology. 2013;154:3925–3930. doi: 10.1210/en.2013-1407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Law NC, Weck J, Kyriss B, Nilson JH, Hunzicker-Dunn M. Lhcgr expression in granulosa cells: roles for PKA-phosphorylated β-catenin, TCF3, and FOXO1. Mol Endocrinol. 2013;27:1295–1310. doi: 10.1210/me.2013-1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Yung Y, Aviel-Ronen S, Maman E, Rubinstein N, Avivi C, Orvieto R, et al. Localization of luteinizing hormone receptor protein in the human ovary. Mol Hum Reprod. 2014;20:844–849. doi: 10.1093/molehr/gau041. [DOI] [PubMed] [Google Scholar]
- 67.Chrusciel M, Ponikwicka-Tyszko D, Wolczynski S, Huhtaniemi I, et al. Extragonadal FSHR expression and function-is it real? Front Endocrinol (Lausanne). 2019;10:32. [DOI] [PMC free article] [PubMed]
- 68.Méduri G, Charnaux N, Driancourt M-A, Combettes L, Granet P, Vannier B, et al. Follicle-stimulating hormone receptors in oocytes? J Clin Endocrinol Metab. 2002;87:2266–2276. doi: 10.1210/jcem.87.5.8502. [DOI] [PubMed] [Google Scholar]
- 69.Pavlik R, Wypior G, Hecht S, Papadopoulos P, Kupka M, Thaler C, et al. Induction of G protein-coupled estrogen receptor (GPER) and nuclear steroid hormone receptors by gonadotropins in human granulosa cells. Histochem Cell Biol. 2011;136:289–299. doi: 10.1007/s00418-011-0846-7. [DOI] [PubMed] [Google Scholar]
- 70.Miller WL, Auchus RJ. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr Rev. 2011;32:81–151. doi: 10.1210/er.2010-0013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Drummond AE, Findlay JK. The role of estrogen in folliculogenesis. Mol Cell Endocrinol. 1999;151:57–64. doi: 10.1016/S0303-7207(99)00038-6. [DOI] [PubMed] [Google Scholar]
- 72.Guo Y, Guo KJ, Huang L, Tong XG, Li X. Effect of estrogen deprivation on follicle/oocyte maturation and embryo development in mice. Chin Med J. 2004;117:498–502. [PubMed] [Google Scholar]
- 73.Britt KL, Drummond AE, Cox VA, Dyson M, Wreford NG, Jones MEE, et al. An age-related ovarian phenotype in mice with targeted disruption of the Cyp 19 (aromatase) gene. Endocrinology. 2000;141:2614–2623. doi: 10.1210/endo.141.7.7578. [DOI] [PubMed] [Google Scholar]
- 74.Peluso JJ, Delidow BC, Lynch J, White BA. Follicle-stimulating hormone and insulin regulation of 17 beta-estradiol secretion and granulosa cell proliferation within immature rat ovaries maintained in perifusion culture. Endocrinology. 1991;128:191–196. doi: 10.1210/endo-128-1-191. [DOI] [PubMed] [Google Scholar]
- 75.Richards JS, Uilenbroek JT, Jonassen JA. Follicular growth in the rat: a reevaluation of the roles of FSH and LH. Adv Exp Med Biol. 1979;112:11–26. doi: 10.1007/978-1-4684-3474-3_2. [DOI] [PubMed] [Google Scholar]
- 76.Orisaka M, Tajima K, Tsang BK, Kotsuji F. Oocyte-granulosa-theca cell interactions during preantral follicular development. J Ovarian Res. 2009;2:9. doi: 10.1186/1757-2215-2-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Aoyama M, Shiraishi A, Matsubara S, Horie K, Osugi T, Kawada T, et al. Identification of a new theca/interstitial cell-specific gene and its biological role in growth of mouse ovarian follicles at the gonadotropin-independent stage. Front Endocrinol (Lausanne) 2019;10:553. doi: 10.3389/fendo.2019.00553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Logan KA, Juengel JL, McNatty KP. Onset of steroidogenic enzyme gene expression during ovarian follicular development in sheep. Biol Reprod. 2002;66:906–916. doi: 10.1095/biolreprod66.4.906. [DOI] [PubMed] [Google Scholar]
- 79.Jing R, Gu L, Li J, Gong Y. A transcriptomic comparison of theca and granulosa cells in chicken and cattle follicles reveals ESR2 as a potential regulator of CYP19A1 expression in the theca cells of chicken follicles. Comp Biochem Physiol - Part D Genomics Proteomics. 2018;27:40–53. doi: 10.1016/j.cbd.2018.04.002. [DOI] [PubMed] [Google Scholar]
- 80.Kim I, Greenwald GS. Evidence for rapid loss of spare hCG receptors in the corpora lutea of the hypophysectomized rat. Mol Cell Endocrinol. 1985;40:123–128. doi: 10.1016/0303-7207(85)90166-2. [DOI] [PubMed] [Google Scholar]
- 81.van Rossum J, Ariens EJ. Receptor-reserve and threshold-phenomena. II. Theories on drug-action and a quantitative approach to spare receptors and threshold values. Arch Int Pharmacodyn Thérapie. 1962;136:385–413. [PubMed] [Google Scholar]
- 82.Casarini L, Riccetti L, De Pascali F, Nicoli A, Tagliavini S, Trenti T, et al. Follicle-stimulating hormone potentiates the steroidogenic activity of chorionic gonadotropin and the anti-apoptotic activity of luteinizing hormone in human granulosa-lutein cells in vitro. Mol Cell Endocrinol. 2016;422:103–114. doi: 10.1016/j.mce.2015.12.008. [DOI] [PubMed] [Google Scholar]
- 83.Casarini L, Lispi M, Longobardi S, Milosa F, La Marca A, Tagliasacchi D, et al. LH and hCG action on the same receptor results in quantitatively and qualitatively different intracellular signalling. PLoS One. 2012;7:e46682. [DOI] [PMC free article] [PubMed]
- 84.Jonas KC, Fanelli F, Huhtaniemi IT, Hanyaloglu AC. Single molecule analysis of functionally asymmetric G protein-coupled receptor (GPCR) oligomers reveals diverse spatial and structural assemblies. J Biol Chem. 2015;290:3875–3892. doi: 10.1074/jbc.M114.622498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Lee CW, Ji I, Ji TH. Use of defined-function mutants to access receptor-receptor interactions. Methods. 2002;27:318–323. doi: 10.1016/S1046-2023(02)00089-0. [DOI] [PubMed] [Google Scholar]
- 86.Mazurkiewicz JE, Herrick-Davis K, Barroso M, Ulloa-Aguirre A, Lindau-Shepard B, Thomas RM, et al. Single-molecule analyses of fully functional fluorescent protein-tagged follitropin receptor reveal homodimerization and specific heterodimerization with lutropin receptor. Biol Reprod. 2015;92:100. [DOI] [PMC free article] [PubMed]
- 87.Rivero-Müller A, Chou YY, Ji I, Lajic S, Hanyaloglu AC, Jonas K, et al. Rescue of defective G protein-coupled receptor function in vivo by intermolecular cooperation. Proc Natl Acad Sci U S A. 2010;107:2319–2324. doi: 10.1073/pnas.0906695106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Ji I, Lee C, Song Y, Michael Conn P, Ji TH. Cis- and Trans-activation of hormone receptors: the LH receptor. Mol Endocrinol. 2002;16:1299–1308. doi: 10.1210/mend.16.6.0852. [DOI] [PubMed] [Google Scholar]
- 89.Ji I, Lee C, Jeoung M, Koo Y, Sievert GA, Ji TH. Trans-activation of mutant follicle-stimulating hormone receptors selectively generates only one of two hormone signals. Mol Endocrinol. 2004;18:968–978. doi: 10.1210/me.2003-0443. [DOI] [PubMed] [Google Scholar]
- 90.Fanchin R, Schonäuer LM, Righini C, Frydman N, Frydman R, Taieb J. Serum anti-Müllerian hormone dynamics during controlled ovarian hyperstimulation. Hum Reprod. 2003;18:328–332. doi: 10.1093/humrep/deg043. [DOI] [PubMed] [Google Scholar]
- 91.Arnhold IJ, Lofrano-Porto A, Latronico AC. Inactivating mutations of luteinizing hormone beta-subunit or luteinizing hormone receptor cause oligo-amenorrhea and infertility in women. Horm Res. 2009;71:75–82. doi: 10.1159/000183895. [DOI] [PubMed] [Google Scholar]
- 92.Pakarainen T, Zhang FP, Nurmi L, Poutanen M, Huhtaniemi I. Knockout of luteinizing hormone receptor abolishes the effects of follicle-stimulating hormone on preovulatory maturation and ovulation of mouse graafian follicles. Mol Endocrinol. 2005;19:2591–2602. doi: 10.1210/me.2005-0075. [DOI] [PubMed] [Google Scholar]
- 93.Wang XN, Greenwald GS. Human chorionic gonadotropin or human recombinant follicle-stimulating hormone (FSH)-induced ovulation and subsequent fertilization and early embryo development in hypophysectomized FSH-primed mice. Endocrinology. 1993;132:2009–2016. doi: 10.1210/endo.132.5.8477652. [DOI] [PubMed] [Google Scholar]
- 94.Tapanainent JS, Lapolt PS, Perlas E, Hsueh AJW. Induction of ovarian follicle luteinization by recombinant follicle-stimulating hormone. Endocrinology. 1993;133:2875–2880. doi: 10.1210/endo.133.6.8243314. [DOI] [PubMed] [Google Scholar]
- 95.Lapolt PS, Nishimori K, Fares FA, Perlas E, Boime I, Hsueh AJW. Enhanced stimulation of follicle maturation and ovulatory potential by long acting follicle-stimulating hormone agonists with extended carboxyl-terminal peptides. Endocrinology. 1992;131:2514–2520. doi: 10.1210/endo.131.6.1446593. [DOI] [PubMed] [Google Scholar]
- 96.Galway AB, Lapolt PS, Tsafriri A, Dargan CM, Boime I, Hsueh AJW. Recombinant follicle-stimulating hormone induces ovulation and tissue plasminogen activator expression in hypophysectomized rats. Endocrinology. 1990;127:3023–3028. doi: 10.1210/endo-127-6-3023. [DOI] [PubMed] [Google Scholar]
- 97.Kristensen SG, Ebbesen P, Andersen CY. Transcriptional profiling of five isolated size-matched stages of human preantral follicles. Mol Cell Endocrinol. 2015;401:189–201. doi: 10.1016/j.mce.2014.12.012. [DOI] [PubMed] [Google Scholar]
- 98.Casarini L, Lazzaretti C, Paradiso E, Limoncella S, Riccetti L, Sperduti S, et al. Membrane estrogen receptor (GPER) and follicle-stimulating hormone receptor (FSHR) heteromeric complexes promote human ovarian follicle survival. iScience. 2020;23:101812. [DOI] [PMC free article] [PubMed]
- 99.Casarini L, Reiter E, Simoni M. β-arrestins regulate gonadotropin receptor-mediated cell proliferation and apoptosis by controlling different FSHR or LHCGR intracellular signaling in the hGL5 cell line. Mol Cell Endocrinol. 2016;437:11–21. doi: 10.1016/j.mce.2016.08.005. [DOI] [PubMed] [Google Scholar]
- 100.Chaffin CL, Vandevoort CA. Follicle growth, ovulation, and luteal formation in primates and rodents: a comparative perspective. Exp Biol Med (Maywood) 2013;238:539–548. doi: 10.1177/1535370213489437. [DOI] [PubMed] [Google Scholar]
- 101.Chun SY, Eisenhauer KM, Minami S, Billig H, Perlas E, Hsueh AJW. Hormonal regulation of apoptosis in early antral follicles: follicle-stimulating hormone as a major survival factor. Endocrinology. 1996;137:1447–1456. doi: 10.1210/endo.137.4.8625923. [DOI] [PubMed] [Google Scholar]
- 102.Hillier SG. Current concepts of the roles of follicle stimulating hormone and luteinizing hormone in folliculogenesis. Hum Reprod. 1994;9:188–191. doi: 10.1093/oxfordjournals.humrep.a138480. [DOI] [PubMed] [Google Scholar]
- 103.Minegishi T, Tano M, Nakamura K, Nakamura M, Igarashi S, Ito I, et al. Regulation of follicle-stimulating hormone receptor. Horm Res. 1996;46(Suppl 1):37–44. doi: 10.1159/000185180. [DOI] [PubMed] [Google Scholar]
- 104.Minegishi T, Tano M, Nakamura K, Karino S, Miyamoto K, Ibuki Y. Regulation of follicle-stimulating hormone receptor messenger ribonucleic acid levels in cultured rat granulosa cells. Mol Cell Endocrinol. 1995;108:67–73. doi: 10.1016/0303-7207(94)03457-5. [DOI] [PubMed] [Google Scholar]
- 105.Nordhoff V, Sonntag B, Von Tils D, Götte M, Schüring AN, Gromoll J, et al. Effects of the FSH receptor gene polymorphism p.N680S on cAMP and steroid production in cultured primary human granulosa cells. Reprod BioMed Online. 2011;23:196–203. doi: 10.1016/j.rbmo.2011.04.009. [DOI] [PubMed] [Google Scholar]
- 106.Zhang YM, Roy SK. Downregulation of follicle-stimulating hormone (FSH)-receptor messenger RNA levels in the hamster ovary: effect of the endogenous and exogenous FSH. Biol Reprod. 2004;70:1580–1588. doi: 10.1095/biolreprod.103.026898. [DOI] [PubMed] [Google Scholar]
- 107.Donaubauer EM, Law NC, Hunzicker-Dunn ME. Follicle-stimulating hormone (FSH)-dependent regulation of extracellular regulated kinase (ERK) phosphorylation by the mitogen-activated protein (MAP) kinase phosphatase MKP3. J Biol Chem. 2016;291:19701–19712. doi: 10.1074/jbc.M116.733972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Donaubauer EM, Hunzicker-Dunn ME. Extracellular signal-regulated kinase (ERK)-dependent phosphorylation of Y-box-binding protein 1 (YB-1) enhances gene expression in granulosa cells in response to follicle-stimulating hormone (FSH) J Biol Chem. 2016;291:12145–12160. doi: 10.1074/jbc.M115.705368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Kayampilly PP, Menon KMJ. Inhibition of extracellular signal-regulated protein kinase-2 phosphorylation by dihydrotestosterone reduces follicle-stimulating hormone-mediated cyclin D2 messenger ribonucleic acid expression in rat granulosa cells. Endocrinology. 2004;145:1786–1793. doi: 10.1210/en.2003-1029. [DOI] [PubMed] [Google Scholar]
- 110.Smith JS, Pack TF, Inoue A, Lee C, Zheng K, Choi I, et al. Noncanonical scaffolding of Gαi and β-arrestin by G protein-coupled receptors. Science. 2021;371:eaay1833. [DOI] [PMC free article] [PubMed]
- 111.Tranchant T, Durand G, Gauthier C, Crépieux P, Ulloa-Aguirre A, Royère D, et al. Preferential β-arrestin signalling at low receptor density revealed by functional characterization of the human FSH receptor A189 V mutation. Mol Cell Endocrinol. 2011;331:109–118. doi: 10.1016/j.mce.2010.08.016. [DOI] [PubMed] [Google Scholar]
- 112.Amsterdam A, Tajima K, Frajese V, Seger R. Analysis of signal transduction stimulated by gonadotropins in granulosa cells. Mol Cell Endocrinol. 2003;202:77–80. doi: 10.1016/S0303-7207(03)00066-2. [DOI] [PubMed] [Google Scholar]
- 113.Channing C, Schaerf F, Anderson L, Tsafriri A. Ovarian follicular and luteal physiology - PubMed [Internet]. Internatioal Rev. Physiol. 1980 [cited 2022 Jan 16]. p. 117–201. Available from: https://pubmed.ncbi.nlm.nih.gov/6248477/. [PubMed]
- 114.Maizels ET, Cottom J, Jones JCR, Hunzicker-dunn M. Follicle stimulating hormone (FSH) activates the p38 mitogen-activated protein kinase pathway, inducing small heat shock protein phosphorylation and cell rounding in immature rat ovarian granulosa cells. Endocrinology. 1998;139:3353–3356. doi: 10.1210/endo.139.7.6188. [DOI] [PubMed] [Google Scholar]
- 115.Uma J, Muraly P, Verma-Kumar S, Medhamurthy R. Determination of onset of apoptosis in granulosa cells of the preovulatory follicles in the bonnet monkey (Macaca radiata): correlation with mitogen-activated protein kinase activities. Biol Reprod. 2003;69:1379–1387. doi: 10.1095/biolreprod.103.017897. [DOI] [PubMed] [Google Scholar]
- 116.Amsterdam A, Gold RS, Hosokawa K, Yoshida Y, Sasson R, Jung Y, et al. Crosstalk among multiple signaling pathways controlling ovarian cell death. Trends Endocrinol Metab. 1999;10:255–262. doi: 10.1016/S1043-2760(99)00164-2. [DOI] [PubMed] [Google Scholar]
- 117.Revankar CM, Vines CM, Cimino DF, Prossnitz ER. Arrestins block G protein-coupled receptor-mediated apoptosis. J Biol Chem. 2004;279:24578–24584. doi: 10.1074/jbc.M402121200. [DOI] [PubMed] [Google Scholar]
- 118.Amsterdam A, Dantes A, Selvaraj N, Aharoni D. Apoptosis in steroidogenic cells: structure-function analysis. Steroids. 1997;62:207–211. doi: 10.1016/S0039-128X(96)00182-1. [DOI] [PubMed] [Google Scholar]
- 119.Sperduti S, Lazzaretti C, Paradiso E, Anzivino C, Villani MT, De Feo G, et al. Quantification of hormone membrane receptor FSHR, GPER and LHCGR transcripts in human primary granulosa lutein cells by real-time quantitative PCR and digital droplet PCR. Gene Reports. 2021;23:101194. doi: 10.1016/j.genrep.2021.101194. [DOI] [Google Scholar]
- 120.Riccetti L, Yvinec R, Klett D, Gallay N, Combarnous Y, Reiter E, et al. Human luteinizing hormone and chorionic gonadotropin display biased agonism at the LH and LH/CG receptors. Sci Rep. 2017;7:940. doi: 10.1038/s41598-017-01078-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Johnson AL, Bridgham JT, Swenson JA. Activation of the Akt/protein kinase B signaling pathway is associated with granulosa cell survival. Biol Reprod. 2001;64:1566–1574. doi: 10.1095/biolreprod64.5.1566. [DOI] [PubMed] [Google Scholar]
- 122.Peter AT, Dhanasekaran N. Apoptosis of granulosa cells: a review on the role of MAPK-signalling modules. Reprod Domest Anim. 2003;38:209–213. doi: 10.1046/j.1439-0531.2003.00438.x. [DOI] [PubMed] [Google Scholar]
- 123.Gebauer G, Peter AT, Onesime D, Dhanasekaran N. Apoptosis of ovarian granulosa cells: Correlation with the reduced activity of ERK-signaling module. J Cell Biochem. 1999;75:547–554. doi: 10.1002/(SICI)1097-4644(19991215)75:4<547::AID-JCB1>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- 124.Shiota M, Sugai N, Tamura M, Yamaguchi R, Fukushima N, Miyano T, et al. Correlation of mitogen-activated protein kinase activities with cell survival and apoptosis in porcine granulosa cells. Zool Sci. 2003;20:193–201. doi: 10.2108/zsj.20.193. [DOI] [PubMed] [Google Scholar]
- 125.Boostanfar R, Jain JK, Mishell DR, Paulson RJ. A prospective randomized trial comparing clomiphene citrate with tamoxifen citrate for ovulation induction. Fertil Steril. 2001;75:1024–1026. doi: 10.1016/S0015-0282(01)01749-6. [DOI] [PubMed] [Google Scholar]
- 126.Kettel LM, Roseff SJ, Berga SL, Mortola JF, Yen SSC. Hypothalamic-pituitary-ovarian response to clomiphene citrate in women with polycystic ovary syndrome. Fertil Steril. 1993;59:532–538. doi: 10.1016/S0015-0282(16)55795-1. [DOI] [PubMed] [Google Scholar]
- 127.Zeleznik AJ, Hutchison JS, Schuler HM. Passive immunization with anti-oestradiol antibodies during the luteal phase of the menstrual cycle potentiates the perimenstrual rise in serum gonadotrophin concentrations and stimulates follicular growth in the cynomolgus monkey (Macaca fascicularis) J Reprod Fertil. 1987;80:403–410. doi: 10.1530/jrf.0.0800403. [DOI] [PubMed] [Google Scholar]
- 128.Robker RL, Richards JS. Hormone-induced proliferation and differentiation of granulosa cells: a coordinated balance of the cell cycle regulators cyclin D2 and p27Kip1. Mol Endocrinol. 1998;12:924–940. doi: 10.1210/mend.12.7.0138. [DOI] [PubMed] [Google Scholar]
- 129.Liu W, Xin Q, Wang X, Wang S, Wang H, Zhang W, et al. Estrogen receptors in granulosa cells govern meiotic resumption of pre-ovulatory oocytes in mammals. Cell Death Dis. 2017;8:e2662. [DOI] [PMC free article] [PubMed]
- 130.Findlay JK, Kerr JB, Britt K, Liew SH, Simpson ER, Rosairo D, et al. Ovarian physiology: follicle development, oocyte and hormone relationships. Anim Reprod. 2018;6:16–19. [Google Scholar]
- 131.Richards JAS, Russell DL, Robker RL, Dajee M, Alliston TN. Molecular mechanisms of ovulation and luteinization. Mol Cell Endocrinol. 1998;145:47–54. doi: 10.1016/S0303-7207(98)00168-3. [DOI] [PubMed] [Google Scholar]
- 132.Barton M, Filardo EJ, Lolait SJ, Thomas P, Maggiolini M, Prossnitz ER. Twenty years of the G protein-coupled estrogen receptor GPER: historical and personal perspectives. J Steroid Biochem Mol Biol. 2018;176:4–15. doi: 10.1016/j.jsbmb.2017.03.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Carmeci C, Thompson DA, Ring HZ, Francke U, Weigel RJ. Identification of a gene (GPR30) with homology to the G-protein-coupled receptor superfamily associated with estrogen receptor expression in breast cancer. Genomics. 1997;45:607–617. doi: 10.1006/geno.1997.4972. [DOI] [PubMed] [Google Scholar]
- 134.Heublein S, Mayr D, Friese K, Jarrin-Franco MC, Lenhard M, Mayerhofer A, et al. The G-protein-coupled estrogen receptor (GPER/GPR30) in ovarian granulosa cell tumors. Int J Mol Sci. 2014;15:15161–15172. doi: 10.3390/ijms150915161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Han N, Heublein S, Jeschke U, Kuhn C, Hester A, Czogalla B, et al. The G-protein-coupled estrogen receptor (GPER) regulates trimethylation of histone H3 at lysine 4 and represses migration and proliferation of ovarian cancer cells in vitro. Cells. 2021;10:1–23. doi: 10.3390/cells10030619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Czogalla B, Partenheimer A, Jeschke U, von Schönfeldt V, Mayr D, Mahner S, et al. β-arrestin 2 is a prognostic factor for survival of ovarian cancer patients upregulating cell proliferation. Front Endocrinol (Lausanne). 2020;11:554733. doi: 10.3389/fendo.2020.554733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Broselid S, Berg KA, Chavera TA, Kahn R, Clarke WP, Olde B, et al. G protein-coupled receptor 30 (GPR30) forms a plasma membrane complex with membrane-associated guanylate kinases (MAGUKs) and protein kinase A-anchoring protein 5 (AKAP5) that constitutively inhibits cAMP production. J Biol Chem. 2014;289:22117–22127. doi: 10.1074/jbc.M114.566893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Prodromidou A, Anagnostou E, Mavrogianni D, Liokari E, Dimitroulia E, Drakakis P, et al. Past, present, and future of gonadotropin use in controlled ovarian stimulation during assisted reproductive techniques. Cureus. 2021;13:e15663. [DOI] [PMC free article] [PubMed]
- 139.Ferraretti AP, La Marca A, Fauser BCJM, Tarlatzis B, Nargund G, Gianaroli L. ESHRE consensus on the definition of “poor response” to ovarian stimulation for in vitro fertilization: the Bologna criteria. Hum Reprod. 2011;26:1616–1624. doi: 10.1093/humrep/der092. [DOI] [PubMed] [Google Scholar]
- 140.Polyzos NP, Sunkara SK. Sub-optimal responders following controlled ovarian stimulation: an overlooked group? Hum Reprod. 2015;30:2005–2008. doi: 10.1093/humrep/dev149. [DOI] [PubMed] [Google Scholar]
- 141.Heublein S, Mayr D, Vrekoussis T, Friese K, Hofmann SS, Jeschke U, et al. The G-protein coupled estrogen receptor (GPER/GPR30) is a gonadotropin receptor dependent positive prognosticator in ovarian carcinoma patients. PLoS One. 2013;8:e71791. [DOI] [PMC free article] [PubMed]
- 142.Roy N, Mascolo E, Lazzaretti C, Paradiso E, D'Alessandro S, Zaręba K, et al. Endocrine disruption of the follicle-stimulating hormone receptor signaling during the human antral follicle growth. Front Endocrinol (Lausanne). 2021;12:791763. doi: 10.3389/fendo.2021.791763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Lyga S, Volpe S, Werthmann RC, Götz K, Sungkaworn T, Lohse MJ, et al. Persistent cAMP signaling by internalized LH receptors in ovarian follicles. Endocrinology. 2016;157:1613–1621. doi: 10.1210/en.2015-1945. [DOI] [PubMed] [Google Scholar]
- 144.Santi D, Casarini L, Alviggi C, Simoni M. Efficacy of follicle-stimulating hormone (FSH) alone, FSH + luteinizing hormone, human menopausal gonadotropin or FSH + human chorionic gonadotropin on assisted reproductive technology outcomes in the “personalized” medicine era: a meta-analysis. Front Endocrinol (Lausanne). 2017;8:114. doi: 10.3389/fendo.2017.00114. [DOI] [PMC free article] [PubMed] [Google Scholar]