

Abstract
Aim
Comprehensive, global information on species' occurrences is an essential biodiversity variable and central to a range of applications in ecology, evolution, biogeography and conservation. Expert range maps often represent a species' only available distributional information and play an increasing role in conservation assessments and macroecology. We provide global range maps for the native ranges of all extant mammal species harmonised to the taxonomy of the Mammal Diversity Database (MDD) mobilised from two sources, the Handbook of the Mammals of the World (HMW) and the Illustrated Checklist of the Mammals of the World (CMW).
Location
Global.
Taxon
All extant mammal species.
Methods
Range maps were digitally interpreted, georeferenced, error‐checked and subsequently taxonomically aligned between the HMW (6253 species), the CMW (6431 species) and the MDD taxonomies (6362 species).
Results
Range maps can be evaluated and visualised in an online map browser at Map of Life (mol.org) and accessed for individual or batch download for non‐commercial use.
Main conclusion
Expert maps of species' global distributions are limited in their spatial detail and temporal specificity, but form a useful basis for broad‐scale characterizations and model‐based integration with other data. We provide georeferenced range maps for the native ranges of all extant mammal species as shapefiles, with species‐level metadata and source information packaged together in geodatabase format. Across the three taxonomic sources our maps entail, there are 1784 taxonomic name differences compared to the maps currently available on the IUCN Red List website. The expert maps provided here are harmonised to the MDD taxonomic authority and linked to a community of online tools that will enable transparent future updates and version control.
Keywords: biodiversity, biogeography, conservation planning, GIS, Mammalia, mapping, species distributions
1. INTRODUCTION
Expert range maps, hand‐drawn polygons that demarcate the geographic distributions of species, have a long history in biogeography and ecology. Initially, they were established to visually illustrate the geographic limits of single or multiple species for educational and taxonomic purposes. Over time, they have also taken on an increasing role in conservation assessment and macroecology. The aim of range maps is typically to define the edges of a species' range, with limited scope for identifying ‘holes’ within that delimited area, which tends to be the focus of other methods for mapping distributions, such as species distribution models or atlases (Jetz et al., 2012).
Range maps represent aggregated expert knowledge and field experience that may implicitly incorporate both coarse and fine‐grain information (Figure 1). For example, speciation history and physiographic barriers may provide information on the broad regions that a species occupies, and information on habitat preferences, published or unpublished occurrence records and local knowledge, may be used to delimit the range within that area. The extent to which range maps demarcate populations into separate polygons can also differ. An array of data sources and types may therefore be used to generate any given range map, and the combinations will vary across species and even between different regions of a given species' range. Range maps are particularly important where observation‐ or specimen‐based occurrence records are sparse, such as in the tropics, where they may represent the only distributional knowledge of a species.
FIGURE 1.
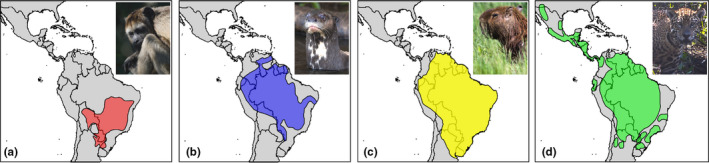
Range maps georeferenced from the Handbook of the Mammals of the World (HMW) for four South American species, (a) Alouatta caraya, (b) Pteronura brasiliensis, (c) Hydrochoerus hydrochaeris and (d) Panthera onca. Different maps species show different levels of precision (i.e., how roughly it approximates the edge of the species' range and how finely they demarcate different populations) and so will differ in their accuracy at different grain sizes
Over recent decades, expert range maps have gained further recognition as quantitative spatial objects that can be aggregated to map biogeographic patterns, including taxonomic richness, range size and population genetics (Fritz et al., 2016; Jetz & Rahbek, 2002; Kreft & Jetz, 2010; Lyons et al., 2019; Olalla‐Tárraga et al., 2011), and they can be linked with trait or phylogenetic data for further inference (Faurby et al., 2018; Jarzyna et al., 2021; Mazel et al., 2017). Efforts such as Map of Life (MOL) have begun to combine expert range maps with other spatial biodiversity data for a more comprehensive understanding of species distributions (Jetz et al., 2012). They have also been used to identify areas of importance for threatened species and conservation priorities (Schipper et al., 2008), as well as for conservation assessments (Brooks et al., 2019; Pacifici et al., 2020). They are recognised as a central, maximum extent‐delimiting data type supporting Species Distribution Essential Biodiversity Variables (SD EBVs), which provide information about the occurrence of species over a specific space–time grid at full taxon extent (Jetz et al., 2019).
However, expert range maps are limited by the somewhat ambiguous temporal scope of the distributions that they are describing (typically characterising recent distributions as records increase towards the present). Below spatial resolutions of ~100 km, they also have a false presence rate (Hurlbert & Jetz, 2007; Hurlbert & White, 2005) that covaries with ecological attributes (Ficetola et al., 2014; Jetz et al., 2008) and with technical aspects of the range map creation. For example, the distribution of a wide‐ranging species will usually have a lower level of spatial detail captured within the range map than that of a narrow endemic, and geographically variable expert knowledge will result in range maps being more precise in some areas than others. The level of precision and uncertainty and the decisions made during the creation of a map are difficult to quantify and document. They can also quickly become out‐of‐date, often due to taxonomic revisions (e.g., species splits or merges) or range boundary changes (e.g., range shifts, or the discovery or extinction of subpopulations). Although separate polygons for introduced range components are sometimes included, range maps typically exclude introductions outside of the species' native ranges. Nevertheless, range maps continue to serve an important role, especially in combination with ancillary data such as species habitat preferences and land cover information to further refine species ranges and as one of several inputs into a new generation of model‐based species distribution predictions (Merow et al., 2017; Pacifici et al., 2018; Powers & Jetz, 2019; Visconti et al., 2011).
Here, we provide fully georeferenced digital range maps for the native ranges of all extant species of mammals, globally harmonised to three taxonomies: the authoritative mammal taxonomy provided in the Mammal Diversity Database (MDD v1.2; Burgin et al., 2018), the Handbook of the Mammals of the World (HMW), which was published across nine volumes (Mittermeier et al., 2013; Wilson et al., 2016, 2017; Wilson & Mittermeier, 2009, 2011, 2014, 2015, 2018, 2019), and the Illustrated Checklist of the Mammals of the World (CMW; Burgin et al., 2020a, 2020b). Each of the three datasets are therefore represented by range maps for all species in each taxonomy, tracking any lumps or splits that occurred between them, as well as any name changes, updates to the ranges and newly described species. The maps are based primarily on the HMW and CMW, and, for the MDD, supplemented with spatial information from other primary source materials, where taxonomic revisions have occurred since the publication of the HMW and CMW compendia. The final range maps are not exact replicas of those published in any one source, as changes are introduced during the interpretation and georeferencing stages, as well as other processing steps, such as clipping to coastlines for terrestrial species.
The three taxonomic authorities represent different temporal snapshots of mammal taxonomy. The nine volumes of the HMW were published over the course of a decade. The CMW aimed to unify the HMW taxonomy into a single two‐volume publication while providing updates to taxonomy and geographic ranges. The MDD was first published in 2018 as a joint effort of the American Society of Mammalogists and the National Science Foundation‐supported VertLife Terrestrial project (vertlife.org) to centralise all published taxonomic changes to mammals as a freely available online database (mammaldiversity.org). The most recent version of the MDD builds upon the CMW and HMW compendia, but provides further updates to taxonomy, synonyms and primary reference information using a wide range of primary literature. The biggest change from the CMW to the MDD is the decision to revert the taxonomy of Bovidae, which follows Groves and Grubb (2011) in the former, to the older taxonomic arrangement presented by Grubb (Grubb, 2005a, 2005b) for the MDD v1.2 (see https://zenodo.org/record/4139818). The rest of the Perissodactyla and non‐cetacean Artiodactyla also follow this older arrangement with some modifications (e.g., Giraffa; Petzold & Hassanin, 2020). The older arrangement is viewed as more stable and justifiable in the mammalogical community (Gutiérrez & Garbino, 2018; Holbrook, 2013), and results in a reduction of 124 extant species in the MDD taxonomy that had been split in the CMW.
The datasets presented in this publication follow the FAIR principles for data sharing (Wilkinson et al., 2016). They are freely accessible from a recognised formal data repository with no restrictions on reuse for non‐commercial purposes, follow data and metadata standards that make them interoperable, and are stored in a well‐known format for spatial data (shapefiles) making them fully reusable. The work represents a collaboration with the Mammal Diversity Database (https://www.mammaldiversity.org; Burgin et al., 2018; versions on Zenodo: https://doi.org/10.5281/zenodo.4139722), an authoritative effort to address and curate the global taxonomy of mammals conducted under the auspices of the American Society of Mammalogists.
With regard to the suite of range maps developed by experts as part of Red List assessments of the International Union for the Conservation of Nature (IUCN; https://www.iucnredlist.org/resources/spatial‐data‐download), the datasets presented in our data paper provide additional range information for many mammal species. The maps available on the IUCN Red List website represent fewer extant species (5829 species, including 20 species labelled as ‘Possibly Extinct'; IUCN, 2021, downloaded 2021‐06‐03) than presented here (HMW = 6253 species, CMW = 6431 species, MDD = 6362 species; Table 1). This translates to 916, 998, and 946 species names (1227 species names across all three sources combined) in the HMW, CMW, and MDD, respectively, which are absent from the IUCN Red List website, as well as 557 species names within the IUCN taxonomy that are absent from the HMW, CMW, and MDD (total of 1784 differences in species names across sources). The range maps described here therefore provide much complementary material to those available through IUCN, and vice‐versa.
TABLE 1.
Total number of extant, non‐domesticated mammal species recognised in each taxonomic source: The Handbook of the Mammals of the World (HMW), the Checklist of the Mammals of the World (CMW), and version 1.2 of the Mammal Diversity Database (MDD). All species listed as domesticated and extinct have been removed from this summary
| HMW | CMW | MDD v1.2 | |
|---|---|---|---|
| Species | 6253 | 6431 | 6362 |
| Genera | 1289 | 1310 | 1306 |
| Orders | 27 | 27 | 27 |
| Species lacking georeferenced maps | 5 | 3 | 2 |
We provide the georeferenced range maps as shapefiles with species‐level metadata and source information for non‐commercial use. Maps are available for bulk download at https://doi.org/10.48600/mol‐7r3j‐8066 (HMW), https://doi.org/10.48600/mol‐zzrs‐q778 (CMW) and https://doi.org/10.48600/mol‐48vz‐p413 (MDD), or all three sources can be accessed from https://mol.org/datasets/?dt=range&sg=Mammals. Maps for individual species can be viewed and downloaded, along with other species‐level spatial data, on Map of Life's species pages (https://mol.org/species).
2. MATERIALS AND METHODS
Range maps were generated by digitally interpreting the maps in the source materials (starting with the HMW), georeferencing them using coastlines, rivers and other geographical features and converting the georeferenced images to vector shapefiles, before quality control and assembling all the species from each source taxonomy into a unified product. To harmonise the HMW and CMW maps, we retained any maps from HMW in which there were no taxonomic or range updates and repeated the digitisation steps with the CMW as source material for any differences. For the MDD dataset, we generated maps for any differences using the HMW or CMW maps as a base along with information from alternative sources, such as published literature.
For the range maps with HMW as the main source, we imaged the maps from each volume, which were then georeferenced using the World Geodetic System projection (WGS84; EPSG: 4326) against the coastlines and political boundaries provided by the Database for Global Administrative Areas v3.6 (GADM; Global Administrative Areas, 2018). Additional spatial data such as rivers and waterbodies were used to determine more exact locations for ground control point placement. For each image, we used a minimum of 10 ground control points that were approximately evenly spaced to ensure constant warping during transformation.
The georeferenced images were converted to GeoTIFFs with cell size equivalent to the resolution of the image using a second‐degree polynomial‐type transformation with a nearest‐neighbour resampling method. These rasters were converted to shapefiles by manually tracing range edges using either the Add Polygon Feature tool in QGIS (QGIS.org, 2020) or the Create Features panel in ArcMap and ArcGIS Pro. In cases where a species range consisted of geographically distinct areas, multipolygons were created. Due to their complexity, coastlines cannot be accurately captured via the tracing method; therefore, coastlines were only roughly outlined manually. Terrestrial species' geometries were subsequently clipped to the coastline of the GADM, while marine species' geometries were clipped to a global map of oceans (Global Administrative Areas, 2018). Species with both terrestrial and marine distributions, such as seals, were not clipped. For each completed species, we then carried out a series of quality controls (described within Technical Validation below).
Once validated, the total number of features in a volume was checked prior to merging. To ensure no geometries were lost during downstream processes, the combined shapefiles were disaggregated so that all geometries were separate before getting clipped, as described above. This clipped file was then merged back together, dissolved by scientific name, and exported as the combined geopackage consisting of range maps for all species. The number of individual features were checked again at the end of the process to confirm that no geometries were lost. Once a volume was completed, we carried out a second quality control to ensure all species in the volume were included, species names were spelled correctly, and that the geometry of each species was valid. Geometry issues were first fixed automatically using the Geospatial Data Abstraction Library (GDAL; GDAL/OGR contributors, 2020) and then using the Check Validity tool in QGIS (QGIS.org, 2020). If necessary, remaining self‐intersections and sliver polygons were fixed by hand. In these cases, satellite images were used to verify any potential errors and help inform the decision of how to correct them.
2.1. Harmonisation between data sources
To create the CMW dataset, range maps for each species in the CMW were compared to those in the HMW volumes. The majority of species did not exhibit range map changes from the HMW to the CMW, but 515 species required some degree of further edits (see the Supplementary Material). These edits fell into one of five categories (Table 2): (1) species that needed to be digitally interpreted from the maps published in the CMW, because they were newly described since the HMW (new species); (2) species that needed to be digitally interpreted directly from the CMW maps because their range changed substantially from the range in the HMW (range changes); (3) species from the HMW that had been taxonomically split into multiple species in the CMW (splits); (4) species in the HMW that had been taxonomically merged into a single species in the CMW (lumps) and (5) species in the HMW that had changes to the scientific name in the CMW taxonomy but no change in geographic range (name changes).
TABLE 2.
Summary of taxonomic and geographical differences of extant, non‐domesticated mammal species recognised in each taxonomic source: The Handbook of the Mammals of the World (HMW), the Illustrated Checklist of the Mammals of the World (CMW), and version 1.2 of the Mammal Diversity Database (MDD). All species listed as domesticated and extinct have been removed from this summary. Note that multiple categories can apply to a single taxonomic revision
| Differences in CMW from HMW | Differences in MDD v1.2 from CMW | |
|---|---|---|
| Range changes | 44 | 2 |
| Name changes | 146 | 66 |
| Splits | 94 species into 220 species | 38 species into 97 species |
| Lumps | 68 species into 32 species | 210 species into 59 species |
| New species | 95 | 20 |
Source interpretation and georeferencing of new species was performed using the same methods as described above for the HMW. For splits, if the split occurred along clear geographical boundaries such as island populations, then the polygons of the original shapefiles were manually separated into child species. However, if the sum of child species was not equal to the range map of the parent species, then each child species was digitally interpreted from the CMW map. The same rule also applied for lumping multiple child species in HMW into a single CMW species. For name changes, the HMW‐based shapefiles were simply renamed. The entire CMW dataset was combined into a single geopackage as described above.
Once conversion of shapefiles from the HMW to the CMW was complete, we then harmonised the CMW shapefiles to the MDD taxonomy to create the MDD dataset. The CMW maps were altered in the same way as described above, except using primary taxonomic descriptions to guide the modifications. For newly described species, range maps were derived directly from the source articles. If sources only presented the geocoordinates of a type specimen (type locality), then a range map was generated by buffering the point with a radius of 50 km. If range maps or type localities were not present in source materials, then range maps were hand‐drawn following maps from the CMW as well as maps drawn by experts in support of the Red List assessment effort by IUCN (IUCN, 2020; necessary for 63 species or ~1% of the total), followed by error‐checking. In all cases, the source materials used to generate each map are recorded in the species‐level citation provided in the metadata of the shapefile for each species (see ‘Usage notes’ section below). Details of all differences between the HMW and CMW, and the CMW and MDD are presented in the supplementary material.
Two species in the CMW, HMW and MDD have no range maps as knowledge of their range is too limited (Nycticeius aenobarbus and Phoniscus aerosus). Two species have no range map in the HMW, but they have range maps within the CMW and MDD (Biswamoyopterus laoensis and Ctenomys dorsalis). One species has a range map in the MDD, but not in the CMW or HMW (Makalata obscura). Three species that feature in the HMW were considered extinct in the CMW and so were excluded from this dataset and the MDD (Juscelinomys candango, Pipistrellus sturdeei and Tonatia saurophila). In all three datasets, two species considered extinct in the wild are retained as their maps are published (Elaphurus davidianus and Oryx dammah).
All georeferencing and shapefile creation were carried out in QGIS 3.10.9‐A Coruña (QGIS.org, 2020), ArcMap 10.7.1, and ArcGIS Pro 2.5.0. Quality checks and creation of geopackages was carried out in Python 3.8 in Jupyter Notebook 6.0.3 using the fiona (Gillies, 2011), geopandas (Gillies, 2007) and shapely (Jordahl et al., 2020) packages.
3. TECHNICAL VALIDATION
The HMW maps were quality controlled by published volume. 10%–30% of images per volume were randomly sampled for manual inspection by a team‐member other than the individual that had carried out the source interpretation and georeferencing. Images were checked for: (1) appropriate transformations and that the World Geodetic System (WGS84) was the reference coordinate system; (2) that each geometry accurately reflected the information in the raster and that no geometries were lost; (3) all geometries were valid; (4) no features were duplicated and (5) that metadata was complete and without translation errors in the scientific name. If over 15% of sampled species were reported to have errors, then the entire volume of species was checked again, otherwise the dataset was cleared for further processing. As there were fewer species to process for the CMW and MDD maps, all species maps were checked visually against the original source.
We also visually confirmed the aggregated products by generating global maps of species richness across all species of each taxonomic source (Figure 2). The richness maps were created by intersecting the range maps of each species with a global equal‐area grid with cell widths of 0.5° and cell height variable by latitude in the Behrmann cylindrical equal‐area projection in R 3.6.1 (R Core Team, 2019) using the sf package (Pebesma, 2018).
FIGURE 2.
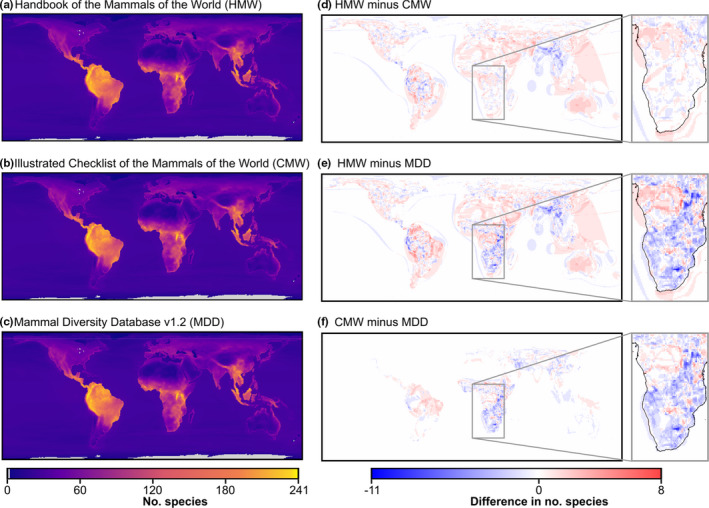
Global richness maps of terrestrial mammals generated by all species with digitisable range maps in (a) the Handbook of the Mammals of the World (HMW), (b) the Illustrated Checklist of the Mammals of the World (CMW) and (c) the CMW harmonised to the latest taxonomy from the Mammal Diversity Database (MDD), as well as the difference in species richness between (d) the HMW minus the CMW, (e) the HMW minus the MDD and (f) the CMW minus the MDD. Detailed regions (sub‐Saharan Africa) for each map of difference are presented to the right, with main differences attributed to the inclusion (HMW, CMW) or exclusion (MDD) of ungulate species from Groves and Grubb (2011). Richness maps were generated by intersecting each species' range map with a global equal‐area grid with cell widths of 0.5° and cell height variable by latitude in the Behrmann cylindrical equal‐area projection
Even though the presented range maps are provided as polygons and thus not directly tied to a specific spatial resolution, it is vital to recognise the spatial grain limitations of expert range map data. At spatial grains smaller than 200 × 200 km, the presence of errors can be substantial; at grains below ~50–100 km edge length, this issue can render the presence information from these maps uninterpretable. Without further validation or inclusion of other data, expert range maps should not be analysed in single or aggregated form at grain sizes <200 × 200 km (Hurlbert & Jetz, 2007; Jetz et al., 2008).
3.1. Comparisons with IUCN range maps
We constructed a richness map for range maps provided by experts to IUCN (6.2, accessed on 25/04/2021; IUCN, 2021) using the same intersections method as above and calculated the difference between the resulting richness map with that of the MDD. IUCN range maps include 30 additional polygon categories for some species based on combinations of information on current status, introductions, reintroductions, seasonality, vagrancy and certainty where possible. Although 90% of species are represented by only one category (‘Extant [resident]’), for the remaining 597 species, users will need to pay close attention to subsetting only those categories relevant to their studies. For example, IUCN range maps include eight extinct (or extinct in the wild) species, as well as historical ranges and areas that have not been explored (‘Not Mapped’) for other species, which would potentially need to be excluded from analyses focussing on current distributions. Therefore, for an accurate comparison with our range maps, which include only native, extant ranges, we removed all polygons labelled as ‘Probably Extant & Introduced (resident)’, ‘Extant & Introduced (resident)’, ‘Extinct & Introduced’, ‘Extinct’, ‘Introduced’ and ‘Not Mapped’.
There are considerable differences between the range maps provided in this data paper and IUCN (Figure 3), with differences of up to 158 species for a given 0.5° cell. First, there are differences between the range maps themselves due to differences in the information used to generate the maps, as well as considerations on how to treat absences within the wider distribution (i.e., how fine to resolve the boundaries and separate populations into individual polygons). The most appropriate range map will therefore depend not only on the accuracy of the information used to generate the maps but also on the user's objectives and study scale. Second, many of the differences can be attributed to differing treatments of coastlines and freshwater bodies. For example, IUCN range maps do not exclude Lake Victoria.
FIGURE 3.
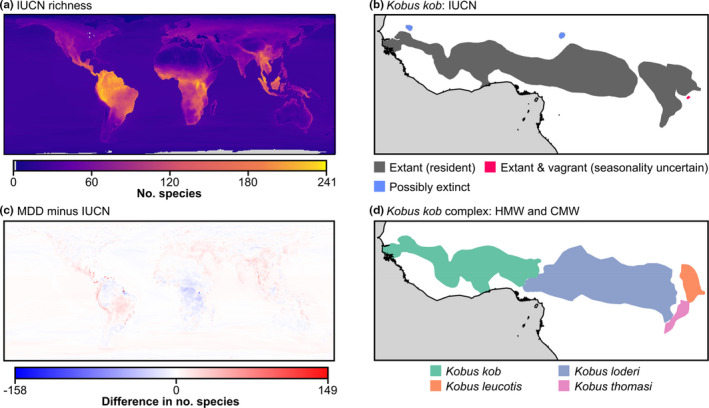
Comparisons with expert maps contributed to IUCN assessment. (a and c) Richness patterns and absolute differences with the Mammal Diversity Database (MDD) calculated as MDD minus IUCN (for details see Figure 2). (b and d) Map differences for the antelope Kobus kob in West and Central Africa. In the IUCN source (b), it is treated as a single species and includes information on the type of distribution and seasonality, but it is separated into four species in the Handbook of the Mammals of the World (HMW) and the Illustrated Checklist of the Mammals of the World (CMW) (d)
Finally, there are considerable differences in the taxonomic treatments (Table 3; Figure 3). Higher richness will be observed where a parent species has been split into multiple child species, and a given grid cell overlays the boundary between the ranges of the new species. Our data and associated taxonomic sources include more species than IUCN, with 7.2%, 10.3% and 9.1% more extant species in the HMW, CMW and MDD, respectively. Overall, this results in 1784 taxonomic differences from IUCN across the three sources. Taxonomic differences are spread widely across mammal groups. For example, there are differences in 16 out of 27 orders and 73 out of 167 families between the species represented in the MDD and IUCN range maps (Table 3). Of these, 289 are name differences (i.e., synonyms) between the MDD and IUCN, 121 species are present only within IUCN and not MDD, and 615 species are present in MDD and not IUCN, totalling over a thousand taxonomic differences in all (see the Supplementary material for details of all differences per taxonomy).
TABLE 3.
Differences between range maps of extant, non‐domesticated mammal species from the Mammal Diversity Database (MDD) version 1.2 and range maps from the International Union for the Conservation of Nature (IUCN)
| Order | Number of families with differences | Name difference | Species in MDD and not in IUCN | Species in IUCN and not in MDD |
|---|---|---|---|---|
| Afrosoricida | 2 | 5 | 0 | 0 |
| Artiodactyla | 8 | 10 | 16 | 4 |
| Carnivora | 10 | 22 | 15 | 7 |
| Chiroptera | 12 | 81 | 137 | 31 |
| Cingulata | 1 | 1 | 2 | 1 |
| Dasyuromorphia | 1 | 1 | 8 | 2 |
| Didelphimorphia | 1 | 1 | 36 | 8 |
| Diprotodontia | 5 | 14 | 11 | 1 |
| Eulipotyphla | 3 | 27 | 65 | 7 |
| Lagomorpha | 2 | 1 | 14 | 1 |
| Macroscelidea | 1 | 2 | 1 | 0 |
| Microbiotheria | 1 | 0 | 2 | 0 |
| Peramelemorphia | 1 | 0 | 3 | 0 |
| Pilosa | 1 | 0 | 6 | 0 |
| Primates | 7 | 17 | 6 | 10 |
| Rodentia | 17 | 107 | 293 | 49 |
We therefore do not profess one set of range maps to be superior to another, but instead urge users to consider the most appropriate range map(s) for their specific use‐case, based on the taxonomic treatments they wish to use, whether they wish to make modifications with regards to usage permissions, the purposes of the study in relation to the qualities each range map possesses, as well as the region under study. In particular, it is important to recognise the limitations that expert range maps generally have on the spatial grains at which they should be applied (Hurlbert & Jetz, 2007).
4. DATA RECORDS
All three datasets are hosted primarily in Map of Life (MOL, mol.org), a scalable platform for the integration and use of global species occurrence and environmental data. The aim of MOL is to support effective and global biodiversity monitoring, research, education, and decision‐making through the provision of tools and information products addressing global species distributions and their dynamics. MOL is deployed on the Google Cloud Platform, which provides the ability to work with managed, scalable services. The data is hosted within a PostgreSQL database with custom API built for both species information and mapping, which are driven by the need for various unique services that users require.
Individual mammal species maps are publicly available for visualisation and download (e.g., https://mol.org/species/map/Chaetodipus_nelsoni) under the ‘Detailed Map’ tab for any given species, along with other sources of spatial information. Each species is represented by a multipolygon that includes dataset‐ and species‐level metadata, as well as the citation information. For convenience, in order to aid the download of large numbers of species, multiple species' shapefiles have also been packaged by taxonomic ranks (order and class). Each dataset is provided with a DOI, which links to the page where they can be accessed and downloaded. The MDD dataset will form the base for version‐controlled updates to the range maps provided by Map of Life and a community of experts (see section ‘Improvements and a framework for community‐driven updates’ below).
5. IMPROVEMENTS AND A FRAMEWORK FOR COMMUNITY‐DRIVEN UPDATES
Inevitably, there will be some inaccuracies in the range maps of some species. For example, precision is necessarily limited for wide‐ranging species to ensure clarity when printed, and errors can be introduced in the printing and georeferencing procedures that get carried through to the final shapefiles. These errors will be more serious in wide‐ranging species, where a small shift in printed pixels translates to larger geographic distances. Both precision and accuracy may be spatially variable as there may be little to no knowledge in parts of a species' range but much in another. Finally, range maps may be drawn using out‐of‐date knowledge, and the ranges of species may change over time. A larger goal of this data publication, therefore, is to provide a clearly referenced and open basis for future updates, and a transparent, open implementation to track changes in distributions, knowledge, and taxonomy (Jetz et al., 2012).
The data provided has no restriction on the non‐commercial use and revision of maps. This might include the updating of maps to an alternative or new taxonomy, or spatial refinements, for example, to allow higher precision coastline and river information. The presented data will undoubtedly require further updates and improvements. We expect a range of such improvements to be provided through Map of Life, where new datasets will also support users with version‐control. In order to support this larger vision for transparently and openly improved expert maps, a new tool, soon to be released, will provide an interactive environment for experts to evaluate and improve range maps (https://mol.org/expert‐range‐mapper‐demo/welcome). The new toolset will allow range maps to be updated in a transparent and traceable manner, with all decisions directly attributable. We expect that this will allow users to clearly track version history of any particular range map, download the most appropriate map for their use case and provide additional information currently unavailable from traditional range maps. For example, areas where agreement between experts are low can potentially inform spatial uncertainty in downstream analyses.
The maps provided here only address native ranges and thus do not inform about the sometimes substantial and ongoing alien range expansions (Capinha et al., 2015; Seebens et al., 2017). Work is underway to include alien distribution data on Map of Life, incorporating information from the Global Register of Introduced and Invasive Species database (GRIIS; Pagad et al., 2018) and the global Distribution of Alien Mammals database (DAMA; Biancolini et al., 2021). While a different data type, this will enable timelier accounting for ongoing range dynamics.
Clearly, a variety of different steps are needed to generate, keep up‐to‐date, and maintain a geospatial resource addressing the distribution of extant mammals and their changes over time. By drawing on an authoritative effort from mammal experts around the world and offering a fully open data resource to the community, we hope to contribute a useful advance to this larger quest.
6. USAGE NOTES
There are two forms of citation that we recommend, depending on the usage. For citing at the dataset‐level, we recommend a citation of both the dataset and the source (HMW, CMW or MDD), and this information can be found on the download page as well as a text file included with the download. Alternatively, at the species‐level, each species' shapefile can be cited independently using the shapefile metadata, which includes the provenance of all information used to generate that species' range map (Table 4).
TABLE 4.
Details of the metadata fields in the attributes of each species' shapefile
| Metadata field | Description |
|---|---|
| sciname | The scientific name of the species |
| authors | The authors involved in generating the shapefile. As multiple people have been responsible over many years at multiple stages of georeferencing, processing and quality control, exact attribution of effort at the species‐level is not always possible. The author list reflects current and past members of MOL who have been involved in the process in no particular order |
| year | The year the shapefile was published |
| rec_source | The original source(s) of the species map if different from the dataset_source, such as a chapter within the HMW or the journal article for more recent taxonomic changes |
| dataset_source | The main compendia or taxonomic source, i.e., the HMW volume, CMW or MDD |
The recommended full citations are included under the column ‘citation’. Where the HMW is the taxonomic source, they have been constructed as: <sciname>: <authors> (<year>) from map published in <rec_source> in <vol_source>.
Where the CMW is the taxonomic source then there is no species‐level rec_source and the citation has been constructed as: <sciname>: <authors> (<year>) from map published in <dataset_source>.
Where the MDD is the taxonomic source, if there were no taxonomic revisions necessary from the CMW, then the CMW is also the species‐level rec_source: <sciname>: <authors> (<year>) from map published in <rec_source> harmonised to <dataset_source>.
If taxonomic revisions were necessary and the maps were hand‐drawn based on available information such as a journal article(s), then the rec_source reflects all the sources of the information used: <sciname>: <authors> (<year>) using information from <rec_source> harmonised to <dataset_source>.
CONFLICT OF INTEREST
The authors declare no competing interests.
BIOSKETCH
Charlie Marsh is interested in spatial ecology, focussing on the tropics, and in particular how ecological processes scale with observation and how we can incorporate this scaling into a better understanding of the effects of human disturbance. He is based at Yale University Department of Ecology & Evolutionary Biology (https://eeb.yale.edu/), the Yale Center for Biodiversity and Global Change (https://bgc.yale.edu/), and Map of Life (https://mol.org/).
Author contributions: Conception and Supervision: WJ; writing of initial draft: CJM, YVS, CJB, WAD, RCA, IDTM, JGV, AR, NU & WJ; source interpretation and georeferencing: YVS, WAD, CJB, RCA, IDTM, JGV, VLD, MD, JAH, AL, JKM, KM, MT & LV; taxonomic harmonisation: CJB, VB, JAH & NU; quality control: CJM, YVS, WAD, CJB, RCA, IDTM & JGV; development of protocols and coordination: YVS, CJM, WAD, MD & WJ; infrastructure: YVS, MD, RG, AR, AS & WJ; generation of original source maps: TLaJ, RAM, ABR, WS, DEW, AMA, LFA, JA‐C, DA, AMB, GB, JKB, JB, PEB, SFB, MAC, PC, EAC, JAC, TC, GC, ECS, VCT, TRBD, TD, CD, CRD, MDBE, EFD, CMF, GF, WLF, TF, JAF, ELG, GSTG, PG, NG, TG, JSG, JG, SMG, SG‐D, KH, MSH, Mha, MHo, KMH, SHe, AH, SHi, KEH, RLH, BAH, TH, RH, CIU, SJ, JJa, MJ, PJ, RJ, JJu, RK, CWK, TK, JLK, BK, TEL, TLeJ, YLRL, RLMN, BKL, AL, RL‐A, AL‐B, CDM, FGM, MAM, HM, SM, EM, AM, FBM, GM, TN, RWN, AO, NO‐G, UFJP, BDP, AP, MP, CP, JP, HLQ, MR, EPR, SJR, DIR, DR, JS‐B, SS‐B, CJS, CS, LKS, MSh, GS, PS, SS, RS, MSu, ABT, MT, PT, TVD, NT, DGT, ST, STT, RV, VVC, GV, JWa, RW, DW, EAW, GW, JWo & DZ.
Supporting information
Supplementary Material 1
Supplementary Material 2
Supplementary Material 3
ACKNOWLEDGEMENTS
We are grateful to the publisher of the key sources used to support this research, Lynx Edicions, Barcelona, and their support for this work. We are also indebted to the many experts who contributed to the development of these publications and, in turn, the colleagues they leaned on. We thank the Institute for the Preservation of Cultural Heritage at Yale University for support. The Map of Life team acknowledges support from Yale University, the E.O. Wilson Biodiversity Foundation, and funding through the NSF VertLife project (NSF DEB‐1441737). NSU was supported by the Arizona State University President's Special Initiative Fund and NIH 1R21AI164268‐01. No permits were required for this research.
Marsh, C. J. , Sica, Y. V. , Burgin, C. J. , Dorman, W. A. , Anderson, R. C. , et al. (2022). Expert range maps of global mammal distributions harmonised to three taxonomic authorities. Journal of Biogeography, 49, 979–992. 10.1111/jbi.14330
Handling Editor: Alexander Zizka
Contributor Information
Yanina V. Sica, Email: yanina.sica@yale.edu.
Nathan S. Upham, Email: nathan.upham@asu.edu.
Walter Jetz, Email: walter.jetz@yale.edu.
DATA AVAILABILITY STATEMENT
All range maps for the three taxonomic sources are openly available for non‐commercial use through https://mol.org/datasets or at species‐level at https://mol.org/species, or for bulk download at https://doi.org/10.48600/mol‐7r3j‐8066 (HMW), https://doi.org/10.48600/mol‐zzrs‐q778 (CMW) and https://doi.org/10.48600/mol‐48vz‐p413 (MDD).
REFERENCES
- Biancolini, D. , Vascellari, V. , Melone, B. , Blackburn, T. M. , Cassey, P. , Scrivens, S. L. , & Rondinini, C. (2021). DAMA: The global distribution of alien mammals database. Ecology, e03474. 10.1002/ecy.3474 [DOI] [PubMed] [Google Scholar]
- Brooks, T. M. , Pimm, S. L. , Akçakaya, H. R. , Buchanan, G. M. , Butchart, S. H. M. , Foden, W. , Hilton‐Taylor, C. , Hoffmann, M. , Jenkins, C. N. , Joppa, L. , Li, B. V. , Menon, V. , Ocampo‐Peñuela, N. , & Rondinini, C. (2019). Measuring terrestrial area of habitat (AOH) and its utility for the IUCN red list. Trends in Ecology & Evolution, 34(11), 977–986. 10.1016/j.tree.2019.06.009 [DOI] [PubMed] [Google Scholar]
- Burgin, C. J. , Colella, J. P. , Kahn, P. L. , & Upham, N. S. (2018). How many species of mammals are there? Journal of Mammalogy, 99(1), 1–14. 10.1093/jmammal/gyx147 [DOI] [Google Scholar]
- Burgin, C. J. , Wilson, D. E. , Mittermeier, R. A. , Rylands, A. B. , Lacher, T. E., Jr. , & Sechrest, W. (2020a). Illustrated checklist of the mammals of the world: Vol. 2: Eulipotyphla to Carnivora. Lynx Edicions. [Google Scholar]
- Burgin, C. J. , Wilson, D. E. , Mittermeier, R. A. , Rylands, A. B. , Lacher, T. E., Jr. , & Sechrest, W. (2020b). Illustrated checklist of the mammals of the world: Vol. 1: Monotremata to Rodentia. Lynx Edicions. [Google Scholar]
- Capinha, C. , Essl, F. , Seebens, H. , Moser, D. , & Pereira, H. M. (2015). The dispersal of alien species redefines biogeography in the Anthropocene. Science, 348(6240), 1248–1251. 10.1126/science.aaa8913 [DOI] [PubMed] [Google Scholar]
- Faurby, S. , Davis, M. , Pedersen, R. Ø. , Schowanek, S. D. , Antonelli, A. , & Svenning, J.‐C. (2018). PHYLACINE 1.2: The phylogenetic atlas of mammal macroecology. Ecology, 99(11), 2626. 10.1002/ecy.2443 [DOI] [PubMed] [Google Scholar]
- Ficetola, G. F. , Rondinini, C. , Bonardi, A. , Katariya, V. , Padoa‐Schioppa, E. , & Angulo, A. (2014). An evaluation of the robustness of global amphibian range maps. Journal of Biogeography, 41(2), 211–221. 10.1111/jbi.12206 [DOI] [Google Scholar]
- Fritz, S. A. , Eronen, J. T. , Schnitzler, J. , Hof, C. , Janis, C. M. , Mulch, A. , Böhning‐Gaese, K. , & Graham, C. H. (2016). Twenty‐million‐year relationship between mammalian diversity and primary productivity. Proceedings of the National Academy of Sciences, 113(39), 10908–10913. 10.1073/pnas.1602145113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- GDAL/OGR contributors . (2020). GDAL/OGR geospatial data abstraction software library. Open Source Geospatial Foundation. https://gdal.org [Google Scholar]
- Gillies, S. (2007). Shapely: Manipulation and analysis of geometric objects. toblerity.org. https://github.com/Toblerity/Shapely
- Gillies, S. (2011). Fiona is OGR's neat, nimble, no‐nonsense API. Toblerity. https://github.com/Toblerity/Fiona
- Global Administrative Areas . (2018). GADM database of Global Administrative Areas [Map].
- Groves, C. , & Grubb, P. (2011). Ungulate taxonomy. John Hopkins University Press. [Google Scholar]
- Grubb, P. (2005a). Order Artiodactyla. In Wilson D. E., & Reeder D. M. (Eds.). Mammal species of the world: A taxonomic and geographic reference (pp. 637–722). John Hopkins University Press. [Google Scholar]
- Grubb, P. (2005b). Order Perissodactyla. In Wilson D. E., & Reeder D. M. (Eds.). Mammal species of the world: A taxonomic and geographic reference (pp. 629–636). John Hopkins University Press. [Google Scholar]
- Gutiérrez, E. E. , & Garbino, G. S. T. (2018). Species delimitation based on diagnosis and monophyly, and its importance for advancing mammalian taxonomy. Zoological Research, 39(5), 301–308. 10.24272/j.issn.2095-8137.2018.037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holbrook, L. T. (2013). Taxonomy interrupted. Journal of Mammalian Evolution, 20(2), 153–154. 10.1007/s10914-012-9206-1 [DOI] [Google Scholar]
- Hurlbert, A. H. , & Jetz, W. (2007). Species richness, hotspots, and the scale dependence of range maps in ecology and conservation. Proceedings of the National Academy of Sciences, 104(33), 13384–13389. 10.1073/pnas.0704469104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hurlbert, A. H. , & White, E. P. (2005). Disparity between range map‐ and survey‐based analyses of species richness: Patterns, processes and implications. Ecology Letters, 8(3), 319–327. 10.1111/j.1461-0248.2005.00726.x [DOI] [Google Scholar]
- IUCN . (2020). The IUCN red list of threatened species. IUCN. [Google Scholar]
- IUCN . (2021). The IUCN red list of threatened species. Version 2021‐1. https://www.iucnredlist.org. Downloaded on 2021‐06‐03.
- Jarzyna, M. A. , Quintero, I. , & Jetz, W. (2021). Global functional and phylogenetic structure of avian assemblages across elevation and latitude. Ecology Letters, 4(2), 196–207. 10.1111/ele.13631 [DOI] [PubMed] [Google Scholar]
- Jetz, W. , McGeoch, M. A. , Guralnick, R. , Ferrier, S. , Beck, J. , Costello, M. J. , Fernandez, M. , Geller, G. N. , Keil, P. , Merow, C. , Meyer, C. , Muller‐Karger, F. E. , Pereira, H. M. , Regan, E. C. , Schmeller, D. S. , & Turak, E. (2019). Essential biodiversity variables for mapping and monitoring species populations. Nature Ecology & Evolution, 3(4), 539–551. 10.1038/s41559-019-0826-1 [DOI] [PubMed] [Google Scholar]
- Jetz, W. , McPherson, J. M. , & Guralnick, R. P. (2012). Integrating biodiversity distribution knowledge: Toward a global map of life—ScienceDirect. Trends in Ecology and Evolution, 27(3), 151–159. 10.1016/j.tree.2011.09.007 [DOI] [PubMed] [Google Scholar]
- Jetz, W. , & Rahbek, C. (2002). Geographic range size and determinants of avian species richness. Science, 297(5586), 1548–1551. 10.1126/science.1072779 [DOI] [PubMed] [Google Scholar]
- Jetz, W. , Sekercioglu, C. H. , & Watson, J. E. M. (2008). Ecological correlates and conservation implications of overestimating species geographic ranges. Conservation Biology, 22(1), 110–119. 10.1111/j.1523-1739.2007.00847.x [DOI] [PubMed] [Google Scholar]
- Jordahl, K. , Bossche, J. V. D. , Fleischmann, M. , Wasserman, J. , McBride, J. , Gerard, J. , Tratner, J. , Perry, M. , Badaracco, A. G. , Farmer, C. , Hjelle, G. A. , Snow, A. D. , Cochran, M. , Gillies, S. , Culbertson, L. , Bartos, M. , Eubank, N. , Maxalbert, Bilogur, A. , & Leblanc, F. (2020). Geopandas/geopandas: V0.8.1 (v0.8.1) [computer software]. Zenodo. 10.5281/ZENODO.3946761 [DOI]
- Kreft, H. , & Jetz, W. (2010). A framework for delineating biogeographical regions based on species distributions. Journal of Biogeography, 37(11), 2029–2053. 10.1111/j.1365-2699.2010.02375.x [DOI] [Google Scholar]
- Lyons, S. K. , Smith, F. A. , & Ernest, S. K. M. (2019). Macroecological patterns of mammals across taxonomic, spatial, and temporal scales. Journal of Mammalogy, 100(3), 1087–1104. 10.1093/jmammal/gyy171 [DOI] [Google Scholar]
- Mazel, F. , Wüest, R. O. , Gueguen, M. , Renaud, J. , Ficetola, G. F. , Lavergne, S. , & Thuiller, W. (2017). The geography of ecological niche evolution in mammals. Current Biology, 27(9), 1369–1374. 10.1016/j.cub.2017.03.046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merow, C. , Wilson, A. M. , & Jetz, W. (2017). Integrating occurrence data and expert maps for improved species range predictions: Expert maps & point process models. Global Ecology and Biogeography, 26(2), 243–258. 10.1111/geb.12539 [DOI] [Google Scholar]
- Mittermeier, R. A. , Rylands, A. B. , & Wilson, D. E. (2013). Handbook of the mammals of the world: Vol. 3: Primates. Lynx Edicions. [Google Scholar]
- Olalla‐Tárraga, M. Á. , McInnes, L. , Bini, L. M. , Diniz‐Filho, J. A. F. , Fritz, S. A. , Hawkins, B. A. , Hortal, J. , Orme, C. D. L. , Rahbek, C. , Rodríguez, M. Á. , & Purvis, A. (2011). Climatic niche conservatism and the evolutionary dynamics in species range boundaries: Global congruence across mammals and amphibians. Journal of Biogeography, 38(12), 2237–2247. 10.1111/j.1365-2699.2011.02570.x [DOI] [Google Scholar]
- Pacifici, M. , Rondinini, C. , Rhodes, J. R. , Burbidge, A. A. , Cristiano, A. , Watson, J. E. M. , Woinarski, J. C. Z. , & Di Marco, M. (2020). Global correlates of range contractions and expansions in terrestrial mammals. Nature Communications, 11(1), 2840. 10.1038/s41467-020-16684-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacifici, M. , Visconti, P. , & Rondinini, C. (2018). A framework for the identification of hotspots of climate change risk for mammals. Global Change Biology, 24(4), 1626–1636. 10.1111/gcb.13942 [DOI] [PubMed] [Google Scholar]
- Pagad, S. , Genovesi, P. , Carnevali, L. , Schigel, D. , & McGeoch, M. A. (2018). Introducing the global register of introduced and invasive species. Scientific Data, 5(1), 170202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pebesma, E. J. (2018). Simple features for R: Standardized support for spatial vector data. The R Journal, 10(1), 439–446. 10.32614/RJ-2018-009 [DOI] [Google Scholar]
- Petzold, A. , & Hassanin, A. (2020). A comparative approach for species delimitation based on multiple methods of multi‐locus DNA sequence analysis: A case study of the genus Giraffa (Mammalia, Cetartiodactyla). PLoS One, 15(2), e0217956. 10.1371/journal.pone.0217956 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Powers, R. P. , & Jetz, W. (2019). Global habitat loss and extinction risk of terrestrial vertebrates under future land‐use‐change scenarios. Nature Climate Change, 9(4), 323–329. 10.1038/s41558-019-0406-z [DOI] [Google Scholar]
- QGIS.org . (2020). QGIS geographic information system (3.10.9‐A Coruña) [Computer software]. QGIS Association. [Google Scholar]
- R Core Team . (2019). R: A language and environment for statistical computing. R Foundation for Statistical Computing. https://www.R‐project.org/ [Google Scholar]
- Schipper, J. , Chanson, J. S. , Chiozza, F. , Cox, N. A. , Hoffmann, M. , Katariya, V. , Lamoreux, J. , Rodrigues, A. S. L. , Stuart, S. N. , Temple, H. J. , Baillie, J. , Boitani, L. , Lacher, T. E. , Mittermeier, R. A. , Smith, A. T. , Absolon, D. , Aguiar, J. M. , Amori, G. , Bakkour, N. , … Young, B. E. (2008). The status of the world's land and marine mammals: Diversity, threat, and knowledge. Science, 322(5899), 225–230. 10.1126/science.1165115 [DOI] [PubMed] [Google Scholar]
- Seebens, H. , Blackburn, T. M. , Dyer, E. E. , Genovesi, P. , Hulme, P. E. , Jeschke, J. M. , Pagad, S. , Pyšek, P. , Winter, M. , Arianoutsou, M. , Bacher, S. , Blasius, B. , Brundu, G. , Capinha, C. , Celesti‐Grapow, L. , Dawson, W. , Dullinger, S. , Fuentes, N. , Jäger, H. , … Essl, F. (2017). No saturation in the accumulation of alien species worldwide. Nature Communications, 8(1), 14435. 10.1038/ncomms14435 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Visconti, P. , Pressey, R. L. , Giorgini, D. , Maiorano, L. , Bakkenes, M. , Boitani, L. , Alkemade, R. , Falcucci, A. , Chiozza, F. , & Rondinini, C. (2011). Future hotspots of terrestrial mammal loss. Philosophical Transactions of the Royal Society B: Biological Sciences, 366(1578), 2693–2702. 10.1098/rstb.2011.0105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilkinson, M. D. , Dumontier, M. , Aalbersberg, I. J. , Appleton, G. , Axton, M. , Baak, A. , Blomberg, N. , Boiten, J.‐W. , da Silva Santos, L. B. , Bourne, P. E. , Bouwman, J. , Brookes, A. J. , Clark, T. , Crosas, M. , Dillo, I. , Dumon, O. , Edmunds, S. , Evelo, C. T. , Finkers, R. , … Mons, B. (2016). The FAIR guiding principles for scientific data management and stewardship. Scientific Data, 3(1), 160018. 10.1038/sdata.2016.18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson, D. E. , Lacher, T. E., Jr. , & Mittermeier, R. A. (2016). Handbook of the mammals of the world. Vol. 6: Lagomorphs and Rodents. Lynx Edicions. [Google Scholar]
- Wilson, D. E. , Lacher, T. E., Jr. , & Mittermeier, R. A. (2017). Handbook of the mammals of the world: Vol. 7: Rodents II. Lynx Edicions. [Google Scholar]
- Wilson, D. E. , & Mittermeier, R. A. (2009). Handbook of the mammals of the world. Vol. 1: Carnivores. Lynx Edicions. [Google Scholar]
- Wilson, D. E. , & Mittermeier, R. A. (2011). Handbook of the mammals of the world. Vol. 2: Hoofed mammals. Lynx Edicions. [Google Scholar]
- Wilson, D. E. , & Mittermeier, R. A. (2014). Handbook of the mammals of the world. Vol. 4: Sea mammals. Lynx Edicions. [Google Scholar]
- Wilson, D. E. , & Mittermeier, R. A. (2015). Handbook of the mammals of the world. Vol. 5: Monotremes and marsupials. Lynx Edicions. [Google Scholar]
- Wilson, D. E. , & Mittermeier, R. A. (2018). Handbook of the mammals of the world. Vol. 8: Insectivores, sloths and colugos. Lynx Edicions. [Google Scholar]
- Wilson, D. E. , & Mittermeier, R. A. (2019). Handbook of the mammals of the world. Vol. 9: Bats. Lynx Edicions. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Material 1
Supplementary Material 2
Supplementary Material 3
Data Availability Statement
All range maps for the three taxonomic sources are openly available for non‐commercial use through https://mol.org/datasets or at species‐level at https://mol.org/species, or for bulk download at https://doi.org/10.48600/mol‐7r3j‐8066 (HMW), https://doi.org/10.48600/mol‐zzrs‐q778 (CMW) and https://doi.org/10.48600/mol‐48vz‐p413 (MDD).