

Abstract
The pandemic burden caused by the SARS-CoV-2 coronavirus constitutes a global public health emergency. Increasing understanding about predisposing factors to infection and severity is now a priority. Genetic, metabolic, and environmental factors can play a crucial role in the course and clinical outcome of COVID-19. We aimed to investigate the putative relationship between genetic factors associated to obesity, metabolism and lifestyle, and the presence and severity of SARS-CoV-2 infection. A total of 249 volunteers (178 women and 71 men, with mean and ± SD age of 49 ± 11 years) characterized for dietary, lifestyle habits and anthropometry, were studied for presence and severity of COVID-19 infection, and genotyped for 26 genetic variants related to obesity, lipid profile, inflammation, and biorhythm patterns. A statistically significant association was found concerning a protective effect of APOE rs7412 against SARS-CoV-2 infection (p = 0.039; OR 0.216; CI 0.084, 0.557) after correction for multiple comparisons. This protective effect was also ascribed to the APOɛ2 allele (p = 0.001; OR 0.207; CI 0.0796, 0.538). The genetic variant rs7412 resulting in ApoE2, genetic determinant of lipid and lipoprotein levels, could play a significant role protecting against SARS-CoV-2 infection.
Subject terms: Preventive medicine, Risk factors
Introduction
COVID-19 pandemic that we are currently experiencing caused by SARS-CoV-2 has affected nearly 350 million people and caused almost 5.6 million deaths globally as of January 20221. Impaired immune response or exacerbated pro-inflammatory response are associated to the main complications of people affected of COVID-19. In addition, emerging evidence demonstrates close link between metabolism and the viral disease process2.
The clinical course of COVID-19 is highly variable. Elderly or patients with some comorbidities such as diabetes mellitus, hypertension, renal dysfunction, cardiovascular diseases (CVDs) and cancer are the most vulnerable groups to suffer severe or fatal cases due to this infection3–5.
Effective control of metabolic parameters (control of glucose, lipid levels, blood pressure and chronic inflammation) might represent a successful approach to dealing with the acute effects of infection by decreasing virus entry into cells and reducing local inflammatory response. Studies have also shown that the nutritional status (determined by Nutritional Risk Tools, NRT) of old patients with COVID-19 has a significant impact on their clinical outcomes6.
Recently, the importance of genotype-environment interactions in shaping the different metabolic phenotypes has been stressed, where diet, exercise, circadian rhythms and gut microbiome appear as the most important environmental factors7,8. The metabolic variability among individuals in the response to diet should be taken into consideration to individualize healthy eating advice, which will promote a reduction in the risk of non-communicable diseases (NCDs). Genetic factors are unmodifiable, but their effects can be modulated through lifestyle and environmental factors6.
COVID-19’s aetiology (its cause or set of causes), beyond SARS-CoV-2 infection, is currently the subject of a large research effort worldwide, given that the disease, at least in its more severe manifestations, occurs through a host huge inflammatory reaction. In this way, several studies have suggested associations between the lipid profile and the infection status or severity of this disease9–11. These studies disclose a severe deregulation of lipoproteins leading to increased triglycerides (TG), together with an alteration of lipoprotein particle levels and an increase in VLDL subclasses with intermediate size9. In turn, other studies showed that COVID-19 adult patients presented lower levels of high-density lipoprotein (HDL-C), which was correlated with a higher risk of developing severe events. In addition, increased TG/HDL-C ratio was associated with disease severity and mortality in COVID-19 patients10,11. Therefore, the current state of research reflects a promising scenario for the discovery of useful metabolism-markers for the early identification of patients at high risk of infection and serious diagnosis.
Likewise, an involvement of circadian rhythms regarding the regulation of the immune system as well as in the severity of infections has been observed12,13. Although the knowledge concerning the mechanisms of interaction at a molecular level of the biological clock and viral infections is still scarce, it is known that viruses are likely to alter the biological processes of infected cells in order to promote their replication and to spread to various tissues. Possible deregulations of circadian rhythms could have a substantial impact on the severity of the symptoms14.
In this perspective, Precision Nutrition is a complementary therapeutic strategy in the management of metabolic and immune alterations associated to COVID-19 poor prognosis. Personalized nutrition combines different aspects to obtain a holistic view of the individual, such as nutrigenetics, detailed phenotyping, and a big set of data obtained from omics technologies (metabolomics, metagenomics, transcriptomics, lipidomics…). Importantly, nutritional interventions based on omics knowledge represents an effective tool to fight against infections and chronic conditions. Hence, Precision Nutrition should also be considered as a complementary tool within the COVID-19 pandemic15.
Although COVID-19 is not primarily a metabolic disease, proper control of some metabolic parameters such as glycemia, blood pressure and lipid levels represent a key therapeutic approach to prevent and ameliorate the acute effects of this viral infection. Within the framework of Precision Nutrition, herein we have analyzed 26 genetic variants in metabolic genes related to obesity, inflammation, chronobiology and nutrient sensing to evaluate their putative associations to the susceptibility and evolution of symptomatology to SARS- CoV2 infection. We identify minor allele rs7412 polymorphism in APOE gene is associated to diminished susceptibility to SARSCoV-2 infection.
Results
Descriptive analysis of COVID-19 disease in the GENYAL platform
A study to identify single nucleotide genetic polymorphisms involved in the SARS-CoV-2 infection was conducted in the GENYAL population. A total of 371 volunteers from this cohort answered the COVID-19 questionnaire designed for this study. COVID-19 questionnaire plus genotyping data were finally restricted to a total of 249 subjects with enough reliable data (178 women and 71 men, with mean and ± SD age of 49 ± 11 years) presented in Table 1. Of the total sample, 210 subjects were considered young adults (< 60 years old, mean age of 46 ± 10 years) while 39 subjects were senior adults (≥ 60 years old; 65 ± 3 years). Table 1 shows the descriptive data of the study sample categorized by gender, age, and infection status (“infected”: 50; “not infected”: 141 subjects; and volunteers with suspected SARS-CoV-2 infection without diagnosis, labeled "not clear": 58). Considering the potential impact of comorbidities on SARS-CoV-2 susceptibility and outcome of SARS-CoV-2 infection, possible associations between comorbidities and SARS-CoV-2 infection status were tested and the results of the analysis are presented in Table 1. No associations were found between SARS-CoV-2 infection status and the studied comorbidities, with the exception of immunosuppression. The data according to the severity of the symptoms (for those infected) are shown in Table S1. No significant associations were found between the studied SNPs involved in lipid metabolism, obesity, inflammation and biorhythms, and COVID-19 severity. Likewise, no positive association was found between SARS-CoV-2 infection and the lifestyle parameters analyzed.
Table 1.
Descriptive phenotypic and clinical data of the study sample categorized by gender, age and SARS-CoV-2 infection.
| Variables | Total % (n = 249) | Male (n = 71) | Female (n = 178) | p | Young Adults (Age ≤ 60) (n = 210) | Older Adults (Age ≥ 60) (n = 39) | p | Virus Infection (n = 50) | Not Clear (n = 58) | No Infection (n = 141) | p |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Age | 49.13 ± 11.23 | 47 ± 12 | 50 ± 11 | 0.030 | - | - | - | 45.0 ± 9.0 | 50.91 ± 11.1 | 49.85 ± 11.7 | 0.003 |
| BMI (kg/m2) | 27.44 ± 5.09 | 29.0 ± 4.7 | 26.8 ± 5.1 | 0.002 | 27.37 ± 5.1 | 27.86 ± 5.1 | 0.582 | 28.26 ± 5.84 | 26.55 ± 4.34 | 27.52 ± 5.07 | 0.350 |
| WC (cm) | 91.47 ± 15.2 | 100.4 ± 15.2 | 88.1 ± 13.8 | < 0,001 | 90.95 ± 15.1 | 94.18 ± 15.4 | 0.232 | 94.79 ± 16.92 | 90.36 ± 13.47 | 90.83 ± 15.24 | 0.379 |
| Obesity | |||||||||||
| No | 68.02 | 67.61 | 70.79 | 0.648 | 70.00 | 69.23 | 1 | 62.00 | 68,97 | 73,05 | 0,350 |
| Yes | 31.98 | 32.39 | 29.21 | 30.00 | 30.77 | 38.00 | 31,03 | 26,95 | |||
| Hypertension | |||||||||||
| No | 86.44 | 81.69 | 92.70 | 0.020 | 90.48 | 84.61 | 0.262 | 88.89 | 82,76 | 92,91 | 0,098 |
| Yes | 13.56 | 18.31 | 7.30 | 9.52 | 15.38 | 11.11 | 17,24 | 7,09 | |||
| Diabetes | |||||||||||
| No | 96.79 | 97.18 | 96.63 | 1 | 97.14 | 94.87 | 0.615 | 98.00 | 94.83 | 97.16 | 0.615 |
| Yes | 3.21 | 2.82 | 3.37 | 2.86 | 5.13 | 2.00 | 5.17 | 2.84 | |||
| Heart disease | |||||||||||
| No | 93.57 | 94.37 | 93.26 | 1 | 94.29 | 89.74 | 0.288 | 92.00 | 91.38 | 95.04 | 0.496 |
| Yes | 6.43 | 5.63 | 6.74 | 5.71 | 10.26 | 8.00 | 8.62 | 4.96 | |||
| Chronic lung disease | |||||||||||
| No | 92.77 | 94.37 | 92.13 | 0.787 | 92.38 | 94.87 | 0.747 | 92.00 | 89.66 | 94.33 | 0.442 |
| Yes | 7.23 | 5.63 | 7.87 | 7.62 | 5.13 | 8.00 | 10.34 | 5.67 | |||
| Chronic kidney disease | |||||||||||
| No | 98.80 | 98.59 | 98.88 | 1 | 98.57 | 100 | 1 | 98.00 | 100.00 | 98.58 | 0.754 |
| Yes | 1.20 | 1.41 | 1.12 | 1.43 | 0.00 | 2.00 | 0.00 | 1.42 | |||
| Immunosuppression | |||||||||||
| No | 94.78 | 98.59 | 93.26 | 0.117 | 94.29 | 97.44 | 0.698 | 92.00 | 89.66 | 97.87 | 0.026 |
| Yes | 5.22 | 1.41 | 6.74 | 5.71 | 2.56 | 8.00 | 10.34 | 2.13 | |||
| Liver pathology | |||||||||||
| No | 97.99 | 98.59 | 97.75 | 1 | 98.57 | 94.87 | 0.176 | 98.00 | 98.28 | 97.87 | 1 |
| Yes | 2.01 | 1.41 | 2.25 | 1.43 | 5.13 | 2.00 | 1.72 | 2.13 | |||
| Cancer | |||||||||||
| No | 98.80 | 98.59 | 98.88 | 1 | 98.57 | 100.00 | 1 | 96.00 | 100.00 | 99.29 | 0.145 |
| Yes | 1.20 | 1.41 | 1.12 | 1.43 | 0.00 | 4.00 | 0.00 | 0.71 | |||
| Pregnancy | |||||||||||
| No | 98.31 | 98.10 | 100.00 | 1 | 98.00 | 100.00 | 97.87 | 0.655 | |||
| Yes | 1.69 | 1.90 | 0.00 | 2.00 | 0.00 | 2.13 | |||||
| Tobacco consumption (cigarettes/day) | |||||||||||
| 0 | 89.39 | 90.00 | 89.14 | 1 | 89.37 | 89.47 | 0.254 | 97.96 | 87.72 | 87.05 | 0.172 |
| 1–5 | 4.08 | 4.29 | 4.00 | 3.38 | 7.89 | 0.00 | 7.02 | 4.32 | |||
| > 5 | 6.53 | 5.71 | 6.86 | 7.25 | 2.63 | 2.04 | 5.26 | 8.63 | |||
| Physical exercise | |||||||||||
| Inactive | 39.36 | 30.99 | 42.7 | 0.114 | 39.52 | 38.46 | 1 | 46.00 | 43.10 | 35.46 | 0.328 |
| Active | 60.64 | 69.01 | 57.3 | 60.48 | 61.54 | 54.00 | 56.90 | 64.54 | |||
| Risk classification | |||||||||||
| Low | 44.58 | 43.66 | 44.94 | 0.768 | 52.86 | 0.00 | 0,001 | 45.39 | 36.21 | 52.00 | 0.219 |
| Medium | 37.75 | 40.85 | 36.52 | 34.76 | 53.85 | 40.43 | 37.93 | 30.00 | |||
| High | 17.67 | 15.49 | 18.54 | 12.38 | 46.15 | 14.18 | 25.86 | 18.00 | |||
Continuous variables: Mean ± SD. Categorical variables: %. BMI Body mass index. WC waist circumference. Physical exercise: Inactive, 0 times of physical activity performance per week; Active, one or more times of physical activity performance per week. Risk classification: Low, No associated risk factors; Medium, 1 associated risk factor; High, More than 1 associated risk factor. Significance level after correction for multiple comparisons p ≤ 0.05.
Associations with genetic variants related to metabolism, obesity, inflammation, and chronobiology on SARS-CoV-2 infection
The objective of this study was to identify putative associations between SARS-CoV-2 infection and genetic factors involved in metabolic or inflammation processes. In this way, 26 SNPs corresponding to genes related to lipid metabolism, obesity, inflammation, and biorhythms were tested for their association with SARS-CoV-2 infection and severity. The frequencies obtained were similar to the European frequencies and the Hardy Weinberg equilibrium was calculated as shown in Table S2. Ordinal logistic regression models were derived for each SNP, adjusted by sex, age, risk factors, risk of virus exposition, hypertension, symptoms severity, and smoking status. An additive model was adopted for the SNPs, where the effect of a homozygotic minor allele genotype was twice the effect of the heterozygotic allele genotype. As predicted endpoint, the three-levels infection status were used (“not infected” vs “not clear” vs “infected”). Figure 1 displays the odds-ratios and p values (before and after Bonferroni correction for multiple tests) obtained for each of the SNPs in these models.
Figure 1.
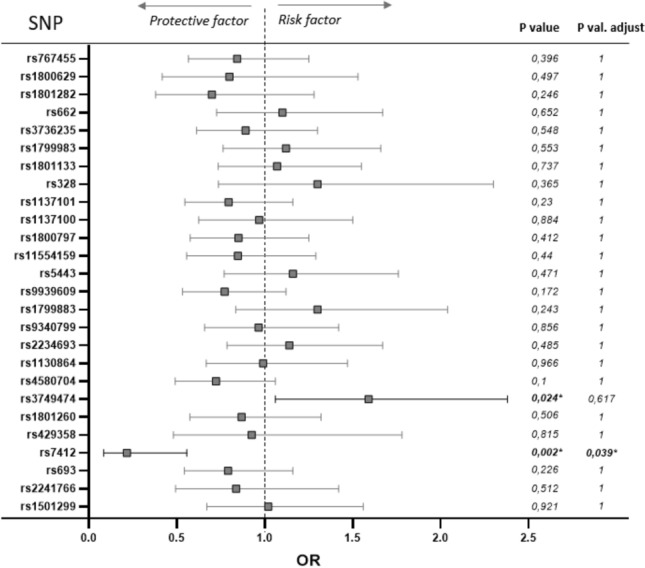
Odds ratio (OR) representing the risk of COVID-19 infection associated to genetic variability of 26 single nucleotide polymorphisms (SNPs) analyzed in 249 volunteers. Confidence interval (CI): 95%. Associations adjusted by sex, age, risk factors, risk of virus exposition, hypertension, symptoms severity and smoking status. The p-values were calculated for an additive model and corrected by multiple SNP comparisons by the Bonferroni test.
APOE rs7412 and APOE2 involvement in SARS-CoV-2 infection
A significant association between the genetic variant APOE rs7412 and SARS-CoV-2 infection status was found after adjusting for multiple testing corrections. For the additive model, a higher protective effect for each T allele was found with respect to SARS-CoV-2 infection (OR: 0.22; CI: 0.084, 0.557; p = 0.039), which is present in the minor ɛ2 allele of APOE. Also, for the dominant model (CC vs CT + TT) the higher protective effect for the T allele (OR: 0.213; CI: 0.081, 0.561; p = 0.046) was found. These results are presented in Table 2. Bootstrap statistics are displayed in Table S3, which shows a small optimism correction and good calibration, signatures of good external predictive power of the model.
Table 2.
Association of SARS-CoV-2 Infection and APOE rs7412 genotype according to genetic models.
| Model | No infection (n = 141) | Not clear (n = 58) | Infection (n = 50) | OR (CI) | p value | p value adjusta | |
|---|---|---|---|---|---|---|---|
| Additive | CC | 118 (83.7) | 53 (91.4) | 47 (94) | 0.216 (0.084, 0.557) | 0.002 | 0.039 |
| CT | 22 (15.6) | 5 (8.6) | 3 (6) | ||||
| TT | 1 (0.7) | 0 (0) | 0 (0) | ||||
| Dominant | CC | 118 (83.7) | 53 (91.4) | 47 (94) | 0.213 (0.081, 0.561) | 0.002 | 0.046 |
| CT + TT | 23 (16.3) | 5 (8.6) | 3 (6) | ||||
| Codominant | CC | 118 (83.7) | 53 (91.4) | 47 (94) | 0.227 (0.086, 0.602) | 0.011 | 0.289 |
| CT | 22 (15.6) | 5 (8.6) | 3 (6) | ||||
| TT | 1 (0.7) | 0 (0) | 0 (0) | ||||
aAdjusted by sex, age, risk factors, risk of virus exposure, severity, tobacco consumption and arterial hypertension; Corrected by multiple SNP comparisons (26) by Bonferroni test.
Figure 2 describes the distribution of rs7412 genotypes vs infection status. rs7412 has a T minor allele and a C major allele; in our sample, the CC, CT, and TT genotypes were present in a total of 218, 30 and 1 individuals, respectively. As observed in Fig. 2, for the additive model, when moving from the “not infected” to the “not clear” to the “infected” group, there is a proportional enrichment of the CC genotype in the genetic composition of the corresponding individuals, while the TT genotype is only present in the “not infected” group; the heterozygote also shows a decrease in its contribution upon higher levels of infection, but in a lower proportion.
Figure 2.

Percentage of the population classified according to SARS-CoV-2 infection status and the APOE rs7412 genotype in the additive model. p value = 0.039 adjusted by Bonferroni multiple corrections.
APOE gene has three main alleles, ɛ2, ɛ3 and ɛ4, coding for three corresponding isoforms of apolipoprotein E: E2, E3 and e4, respectively. Table S4 displays the distribution of genotypes in our samples. The main isoform E3 and the minor isoform E4 of ApoE protein have strong affinity to Low Density Lipoprotein Receptor (LDLR) and have been associated to a higher risk of Alzheimer’s disease. On the contrary, E2 isoform has been described as neuroprotective, and presents low affinity for the LDL receptor. In terms of sequences, these isoforms are defined by the amino acids occurring at positions 112 and 158, and determined by rs429358 together with rs7412: E2 isoform: Cys112 (rs429358-T), Cys158 (rs7412-T); E3 isoform: Cys112 (rs429358-T), Arg158 (rs7412-C) and E4 isoform has Arg112 (rs429358-C), Arg158 (rs7412-C). Thus, to determine a putative association between APOE isoform and SARS-CoV-2infection, we next evaluated the combination of the two SNPs rs7412 and rs429358 in the risk of infection by mean of an ordinal logistic model with the same covariables as before, by using a model that included two variables representing the number of ε2 and ε4 alleles, respectively, in the genotype. Table 3 describes the distribution of the three alleles vs infection classes, together with the statistics of the resulting model where a protective effect was found only for ɛ2 allele (OR: 0.207; CI: 0.0796, 0.538; p = 0.001). No significant association was found for ɛ4 (OR: 0.784; 0.402, 1.53; p = 0.476). On the basis of these results, ɛ2 allele seems to be a favorable marker against SARS-CoV-2 infection.
Table 3.
Association of SARS-CoV-2 Infection and APOE alleles according to additive allelic models.
| Allele | No. copies | No infection (n = 141) | Not clear (n = 58) | Infection (n = 50) | OR (CI) | p value | p value adjusta |
|---|---|---|---|---|---|---|---|
| ε2 | 0 | 118(83.69) | 53(91.38) | 47(94) | 0.207(0.0796,0.538) | 0.001 | 0.031 |
| 1 | 22(15.6) | 5(8.62) | 3(6) | ||||
| 2 | 1(0.71) | 0(0) | 0(0) | ||||
| ε4 | 0 | 120(85.11) | 53(91.38) | 40(80) | 0.784(0.402,1.53) | 0.67 | 1 |
| 1 | 18(12.77) | 5(8.62) | 9(18) | ||||
| 2 | 3(2.13) | 0(0) | 1(2) | ||||
| ε3 | 0 | 5(3.55) | 0(0) | 1(2) | Not in modelb | ||
| 1 | 38(26.95) | 10(17.24) | 12(24) | ||||
| 2 | 98(69.5) | 48(82.76) | 37(74) |
Adjusted by sex, age, risk factors, virus exposure, severity, tobacco consumption, and arterial hypertension.
aCorrected by multiple comparisons by Bonferroni test.
bThis allele is not included as it would result in perfect collinearity.
T allele of CLOCK rs3749474 might be involved in a potential increased risk of SARS-CoV-2 infection
In addition to the association observed with APOE genetic variant, the analysis of the genotypes performed showed a positive association between the risk of SARS-CoV-2 infection and CLOCK gene rs3749474 polymorphism (p = 0.024), although statistical significance was lost after Bonferroni correction (Fig. 1). This genetic variant has been associated with modulation of metabolic traits, such as energy intake16, obesity17, and biorhythms18,19, and it has also been related to variations in the levels of several cytokines16. The best fit was obtained with the additive model, in which each copy of the minor allele T was associated with an increased risk of COVID-19 disease (OR = 1.59, CI: 1.06–2.38) (Table 4). The genotype frequencies obtained from this polymorphism were similar to those described for the European population when analyzing the study population as a whole. However, when stratifying for positive infection, it was observed that among the participants infected by SARS-CoV-2, the TT genotype was present in 22% of the participants, meanwhile only 7.8% of this genotype was present in the group of uninfected subjects (Fig. 3).
Table 4.
Association of SARS-CoV-2 Infection and CLOCK rs3749474 genotype according to genetic models.
| No infection (n = 141) | Not clear (n = 58) | Infection (n = 50) | OR (CI) | p value | p value adjusta | |
|---|---|---|---|---|---|---|
| Additive | ||||||
| CC | 69 (48.9) | 26 (44.8) | 17 (34) | 1.59 (1.06, 2.38) | 0.024 | 0.617 |
| CT | 61 (43.3) | 27 (46.6) | 22 (44) | |||
| TT | 11 (7.8) | 5 (8.6) | 11 (22) | |||
| Dominant | ||||||
| CC | 69 (48.9) | 26 (44.8) | 17 (34) | 1.59 (0.932, 2.7) | 0.089 | 1 |
| CT + TT | 72 (51.1) | 32 (55.2) | 33 (66) | |||
| Codominant | ||||||
| CC | 69 (48.9) | 26 (44.8) | 17 (34) | 1.28 (0.724, 2.26) | 0.041 | 1 |
| CT | 61 (43.3) | 27 (46.6) | 22 (44) | |||
| TT | 11 (7.8) | 5 (8.6) | 11 (22) | |||
aAdjusted by sex, age, risk factors, risk of virus exposure, severity, tobacco consumption and arterial hypertension; Corrected by multiple SNP comparisons (26) by Bonferroni test.
Figure 3.

Percentage of the population classified according to SARS-CoV-2 infection status and the CLOCK rs3749474 genotype in additive model.
In view of these results, an interaction model between APOE rs7412 and CLOCK rs3749474 variants was performed. We noted that the combination of the two polymorphisms together significantly enhanced the predictive effect that APOE variant on the risk of SARS-CoV-2 infection compared to the individual analysis (p = 0.02), suggesting a cooperative role of both genetic variants in relation to the infectivity of the virus.
This remarkable finding could be an initial indicator of the potential predictive value of polymorphic variants in the CLOCK gene on SARS-CoV2 infection, as well as the correlation that the two SNPs, APOE and CLOCK, have together regarding susceptibility to COVID-19 infection.
Finally, we investigated the possible modulating effects of diet or medication on SARS-CoV-2 infection. Healthy eating index (IAS), energy intake, total lipids and carbohydrates, simple sugars and saturated fatty acids consumption were tested, as well as consumption of anti-inflammatory, hypotensive or lipid lowering drugs for medication. No significant correlation with SARS-CoV-2 infection or symptoms severity was found in any case.
Discussion
The present work has focused on the study of genetic factors involved in SARS-CoV-2 infection and its severity in order to broaden the knowledge of the genetic influence on the probability of infection. To this end, single nucleotide genetic variants in genes related to lipid metabolism, inflammation and biorhythms have been analyzed.
A statistically significant association between the genetic variant APOE rs7412 and the SARS-CoV-2 infection status was detected for the additive and dominant models, that remained significant after Bonferroni multiple-test correction. Both models suggest that the minor T allele (dominant model) or each copy of the T allele (additive model), would elicits a protective effect against SARS-CoV-2 infection. Nonetheless, given the cross-sectional nature of this study, it is not possible to fully demonstrate from here a causal involvement of the ε2 allele in the observed decrease in infection. However, the significant association reported in this work opens the door to future controlled longitudinal trials capable of confirming this hypothesis, of great interest given the need to understand the risk factors of COVID-19 disease.
This protective effect may be related to multiple processes where apolipoprotein E is implicated including cardiovascular function, immune system and inflammation20,21. Previous studies have evidenced the association of APOE alleles with susceptibility to infection by viruses, with elevated levels of lipoprotein(a) (Lp(a)), with local extracellular vesicle biogenesis in the brain, with the levels of the Willebrand factor and the modulation of angiotensin converting enzyme 2 (ACE2)20,22–27. These processes may be related to a prothrombotic condition which, together with the inflammatory character of SARS-CoV-2 infection, may lead to a higher infection capacity and severity. In addition, different isoforms of apolipoprotein E can give rise to multiple phenotypes, with numerous factors intervening in their biogenesis and expression regulation. For this reason, the multifaceted associations to APOE gene require multiple approaches to be taken into account20.
The knowledge provided in the current literature regarding COVID-19, refers mainly to APOE alleles (ɛ2, ɛ3, and ɛ4) in the population due to the multiallelic nature of apolipoprotein E gene. Nonetheless, the results of the present study revealed a protective effect of the minor T allele of the SNP rs7412 which is found only in the minor ɛ2 allele of APOE. Consequently, these results are complementary to those observed in other studies, where the ɛ4 allele increases the risk of SARS-CoV-2 infection and severity of COVID-19 the symptoms27–30.
In addition, genome wide significant associations studies (GWAS) related to rs7412 (minor T allele) and higher odds of surviving and longevity have been found31,32. On one hand, studies have shown that the rs7412 genetic variant had the strongest association with LDL-TG and RLP-C as well as a significant association with increased TG and HDL-C levels, and with decreased LDL-TG, LDL-C, total cholesterol, non-HDL-C, and Lp(a) levels33. In addition, several studies have reported a significant association between the APOE2-determining allele of rs7412 and Lp(a) concentrations34,35. These investigations similarly revealed a decrease in Lp(a) in the population carrying the allele associated with rs741234,35. In the same time, elevated levels of this lipoprotein have been proposed to be associated with the risk of SARS-CoV-2 infection and with a very high risk of thrombosis23. Therefore, these mechanisms could help to explain the potential influence of this genetic variant on SARS-CoV-2 infection. For this reason, this study may help to provide new avenues for studying ApoE purposes, evaluating not only the effect exerted by ɛ4 but also the protective effect of rs7421 and ɛ2 allele against COVID-19.
Subsequently, the effect of the APOE gene alleles and the risk of SARS-CoV-2 infection was assessed. As alleles ɛ2, ɛ3 and ɛ4 in APOE gene correspond to genotypes rs7412-T/rs429358-T; rs7412-C/rs429358-T; and rs7412-C/rs429358-C, respectively, we evaluated the combination of the two SNPs rs7412 and rs429358 in the risk of infection. In the resulting model, a protective effect was found only for ɛ2 allele after adjusting for the Bonferroni correction. These results, consequently, are in accordance with our hypothesis on the protective effect exerted by the ɛ2 allele. In the same time, published literature reports the beneficial character of apolipoprotein E2 according to cardiovascular diseases, neuroprotection, and other age-related disease traits and survival implicated in the promotion of longevity27,36,37. This way, a lower affinity for the LDL receptor, reduced hypertension and/or increased neuroprotective capacity are proposed mechanisms which may contribute to the protective mechanism against SARS-CoV-2 infection38,39. Therefore, the multifaceted nature of APOE pleomorphic gene, specifically the ɛ2 allele, may have a protective effect against COVID-19 disease.
Besides that, as COVID-19 pandemic evolves, several discouraging long-term effects are being observed. These include neurological manifestations such as smell/taste disorders and non-specific and possibly systemic neuropathological symptoms40. This seems to be due to the neurotropic character of the virus, where potential entry of the virus into the vascular endothelial cells of the brain is possible40,41. In this scenario, a possible neuroprotective effect could be also hypothesized by the rs7412 or the ɛ2 allele, given its beneficial character in relation to neurodegenerative mechanisms41.
Diet and medication do not seem to interfere with infection status and rs7412. However, more studies should be conducted in this direction. This study has been performed in a mainly non-pathological population and a larger sample size may be necessary for future analysis.
The immuno-inflammatory activation as a consequence of a viral infection is a complex system which is affected by multiple factors, including the host circadian clock14,42. Disruptions in circadian rhythms have been reported to affect both viral replication and dissemination within the host, as well as innate and adaptive immune responses, which can modulate the infective capacity of the virus and its clinical manifestations13,14,42,43. Regarding the infection by SARS-CoV-2, in addition to the previously mentioned, it has been reported that the circadian clock machinery is closely involved in the transcriptional expression of ACE2, the main responsible receptor for the viral entry of SARS-CoV-2 into the cells44. In view of the previous evidence, several authors accentuated the urgent need to study the potential contribution of circadian rhythms on COVID-19 disease to clarify whether this relationship can play a role in the spread of this pandemic45–47. In this sense, our study shows a positive association between the minor T allele of the rs3749474 variant of the CLOCK gene (a core component of the circadian clock) and the risk of infection by SARS-CoV-2 (p = 0.024), although this significance was lost after adjusting for Bonferroni correction, probably due to the low sample size. Despite the fact that the genotype frequencies of this polymorphism were similar to those described in the European population, infected subjects presented an increased incidence of TT + CT genotypes compared to non-infected subjects, pointing to a higher infection rate in the T allele carriers of this polymorphism. This experimental finding describes for the first time a potential direct relationship between a gene involved in the circadian clock and the modulation of SARS-CoV-2 infection, thus opening the door to future studies in this field.
The rs3749474 polymorphism of the CLOCK gene has also been associated with altered metabolic features. Subjects who carry the T allele of this variant show a higher energy intake16 BMI and risk of obesity17,48. After numerous clinical reports, it is a statement that obese patients, and especially those with a higher BMI, are severely affected by COVID-19 and present a worse outcome than patients with lower BMI49–51. Again, this supports the link described in our study between the rs3749474 polymorphism and SARS-CoV-2 infection.
It is noteworthy that of all the polymorphisms studied in our population, APOE and CLOCK polymorphisms have shown significant associations with SARS-CoV-2 infection, and their statistical significance is also enhanced when they are analyzed together in the same model. This suggests a cooperative role of both genes related to the infectivity of this virus. Other authors have previously described cases where the CLOCK gene and APOE genotypes can interact together modulating the progression of inflammatory-based diseases52,53. This reinforces the cooperative role proposed for these polymorphisms on the susceptibility to infection by SARS-CoV-2, as well as the progression and severity of other inflammatory-based diseases in which these genes may play a major role.
The relationship between the severity of the symptoms in the group of individuals infected by SARS-CoV-2 and the analyzed genetic polymorphisms was also assessed. However, no significance was noticed for any of the SNPs tested. These results could suggest that APOE and CLOCK could be involved in modulating the risk of SARS-CoV-2 infection as reported, but they may not have a major influence on the progress of the disease once established. However, our study has some limitations. In the first place, the sample size of the infected population might not be large enough to appreciate a significant influence of APOE and CLOCK in the severity of symptoms. Therefore, it would be advisable to test this association in a large number of infected subjects to ensure if these factors could affect the severity of symptoms caused by COVID-19 disease. Eventually, because of the complicated epidemiological situation Spain was suffering while the study was being performed, the limited availability of diagnostic tests, and the retrospective nature of the study, it was not possible to verify firmly the participants’ SARS-CoV-2 infection. Nonetheless, a validation and verification mechanism of past infection would have been interesting and will be considered in new investigations. Future studies with larger sample size are warranted to contrast the results obtained in this research. Additionally, analyzing these associations in an ethnically diverse population could be relevant as ethnicity has an impact on these variants.
To conclude, although more studies need to be conducted to corroborate the current results, we observed than the minority allele (T) of the genetic variant APOE rs7412 and the APOɛ2 allele, may confer a certain protection against SARS-CoV-2 infection. This observation will allow the performance of fully longitudinal studies to confirm this novel hypothesis from a causative point of view. In addition, the data here obtained and publicly shared will allow the estimation of appropriate sample sizes and power for these future trials.
Materials and methods
Subjects and study protocol
Subjects that participated in this study were recruited through the Platform for Clinical Trials in Nutrition and Health (GENYAL), at IMDEA Food Institute (Madrid, Spain). Given the novelty of the COVID pandemic, no information about the variability of the data to collect was available, precluding the use of sample size calculations, so we used the largest sample available. Volunteers included in the GENYAL database were contacted to participate in this study. A total of 371 volunteers of European ancestry were recruited with the objective to link data related to genetic variants, anthropometric measurements, dietary patterns and lifestyle with SARS-CoV-2 infection and severity of COVID-19 disease. Inclusion criteria were the following: previous participation in GENYAL Platform studies, adequate level to understand the research propose, agreement to voluntarily participate in the study and provide informed consent. Only those who refused to give their consent to participate in this study were excluded. CONSORT flow diagram is shown in Fig. S1. Although it could have been better to start with a larger sample, we decided to carry on with the sample available at our hands, on the interest on finding novel associations and risk factors for this novel COVID19 pandemic, and using our resources and experience in the nutrigenomics area.
This study was performed according to the guidelines present in the Declaration of Helsinki and all procedures involving human subjects were approved by the Research Ethics Committee of the IMDEA Food Foundation (CEI 27-666). Written informed consent to enroll in the study was obtained from all the participants. The trial registration number from ClinicalTrials.gov is NCT04067921.
Anthropometry and lifestyle parameters
Anthropometric measurements such as height, weight and waist circumference were measured following standard validated techniques54,55. Body weight was assessed using the body composition monitor BF511 (Omron Healthcare UK Ltd., Kyoto, Japan). Height was assessed using a stadiometer (Leicester-Biological Medical Technology SL, Barcelona). Waist circumferences were measured using a Seca 201 non-elastic tape (Quirumed, Valencia, Spain). The dietary patterns of each participant were assessed using a validated 72-h dietary food record and a food frequency questionnaire56. Energy intake, healthy eating index, macro and micronutrients from the dietary records were analyzed using the DIAL software (2.16 version Alce Ingeniería, Madrid, Spain). Regarding lifestyle data, physical activity parameters (Inactive: 0 times of physical activity performance per week/Active: one or more times of physical activity performance per week) and smoking habits (cigarettes/day) were collected from every participants57.
DNA extraction and genotyping
Single nucleotide polymorphisms related to lipid metabolism, obesity, circadian rhythm, immune system, and estrogens were previously screened and selected according to previously published procedures58,59. Blood samples were obtained from study participants and stored at − 80 °C until DNA extraction. Genomic DNA from each participant was isolated using the QIAamp DNA Blood Mini Kit (Qiagen Sciences, Inc, Germantown, MD, USA). DNA concentration and quality were assured in a nanodrop ND-2000 spectrophotometer (ThermoScientific, Waltham, MA, USA). Gene genotyping was performed using TaqMan OpenArray plates with the QuantStudio™ 12 K Flex Real-Time PCR System (Life Technologies Inc., Carlsbad, CA, USA) according to the manufacturer’s instructions60. The results obtained were analyzed with the TaqMan Genotyper software61. Duplicates of the samples were made, obtaining more than 99% of technically valid genotypes.
COVID-19 data collection
The participants answered a detailed survey of 24 questions related to COVID-19 disease. Questions were stratified into 4 different sections designed to determine: 1—the degree of exposure to SARS-CoV-2, 2—the diagnosis of the disease, 3—symptomatology and severity of the symptoms and 4—associated risk factors. The responses to each of these sections were analyzed and categorized in different clusters according to the following criteria:
Degree of exposure to SARS-CoV-2 Low (Constant home confinement + No contact with people infected by COVID-19), Medium (Rest of participants), High (Coexistence with a person infected by COVID-19/No adherence to COVID-19 security guidelines/Participants with possible exposure to COVID-19 at work).
Diagnosis of the disease Not infected (Negative PCR/Negative serological test/No perception of symptoms or disease), Not clear (Possible infection but no diagnostic test was performed), Infected by SARS-CoV-2 (Positive PCR / Positive serological test/Typical COVID-19 symptomatology / Diagnosis by medical presumption).
Symptomatology and severity (Table 5) Null (No symptoms/Symptoms described by participant as chronic and not associated with COVID-19), Mild-moderate (Mild and moderate symptoms 1–3 weeks/Mild symptoms > 3 weeks/Severe symptoms ≤ 1 week), Severe (Hospitalization period/Pneumonia caused by COVID19/Moderate or severe symptoms > 3 weeks/Severe symptoms 1–3 weeks).
Associated risk factors Low (0 risk groups), Medium (1 risk group), High (> 1 risk group). Risk groups associated with COVID-19 described: hypertension, diabetes, heart disease, chronic lung disease, chronic kidney disease, immunosuppression, liver pathology, cancer and pregnancy.
Table 5.
COVID-19 symptomatology used to define severity of disease.
| Null | Mild | Moderate | Severe |
|---|---|---|---|
| No symptoms | Mild/moderate symptoms of ≤ 1 week duration | Mild/moderate symptoms for 1–3 weeks | Hospitalization |
| Severe symptoms for ≤ 1 week | Pneumonia | ||
| Mild symptoms for 1–3 weeks | Mild symptoms for > 3 weeks | Moderate/severe symptoms for > 3 weeks | |
| Severe symptoms lasting 1–3 weeks |
Symptoms Cough, disnea, expectoration (lower respiratory tract secretions), rhinorrhoea, sore throat, chills, fever (mild: ≥ 37.5–< 38 °C; moderate: ≥ 38–< 39 °C ; severe: ≥ 39 °C), diarrhea (mild: < 4 depositions/day; moderate: 4–7 depositions/day; severe: > 7 depositions/day), nausea, vomiting, headache, dizziness, fatigue and myalgia, anosmia, dysgeusia, musculoskeletal pain, loss of appetite, sadness or distress.
Statistical analysis
The R software, version 3.6.1, was used for all the statistical analyses62. A significance level of 0.05 was used in bilateral tests. Missing data was multiple imputed using the mice package, based on conditional chained equations, and using the default imputation methods, namely predictive mean matching for numeric variables, logistic regression for binary variables, polytomous regression for categorical data, and proportional odds logistic regression for ordinal variables63. Pooled estimates of parameters and 95% confidence intervals, as well as of p values were obtained afterwards from the multiple imputations64. The rms package was used to derive ordinal logistic models in the proportional-odds version. These models were fitted to predict infection status for the whole dataset, and symptoms severity in the infected stratum of the sample. A wide set of 26 SNPs related to metabolism and nutrition was tested for significance in the prediction of these endpoints, and sex, age, risk, exposition to infection, hypertension, symptoms severity and smoking status were used as adjustment variables. Multiple-test correction of p-values was controlled with the Bonferroni method. The model was validated through bootstrap, using 2000 resamples. An additive model was adopted for the SNPs, where the effect of a homozygotic minor allele genotype is twice the effect of the heterozygotic allele genotype. The use of a codominant or a dominant model did not improve the fit in general, so they were not used. The significance of medication and diet variables was tested by including them in the model in the form of an additive term or an interaction with the rs7412 SNP.
Supplementary Information
Acknowledgements
The authors want to express their gratitude to the GENYAL Platform of Clinical Trials in Nutrition and Health of IMDEA Food Institute for their support, as well as the Counceling of Education of the Community of Madrid for their support in the constitution of GENYAL, and all the volunteers participating in this study. This research was funded by the Plan Nacional I + D + i PID2019 ‐110183RB ‐C21, FACINGLCOVID-CM project. Funding REACT EU Program (Comunidad de Madrid and The European Regional Development Fund. ERDF. European Union). Regional Government of Community of Madrid (Y2020/BIO-6350). Genomic Laboratory GENYALLab (REDLAB, the laboratories network of the Community of Madrid, under the registration number 440). GENYAL LAB participates in ALIBIRD2020-CM project (S2018/BAA-4343): "Therapeutic formulas of precision nutrition for cancer", supported by program call of R & D Activities among Research Groups of the Community of Madrid (Technologies 2018) and co-financed with European Union Structural Funds.
Author contributions
Conceptualization, A.R.M., G.R., and A.M.; methodology, software, validation, formal analysis, investigation, resources, data curation, I.E.S., G.C., CMFD, writing—original draft preparation, I.E.S., G.C., C.M.F.D., M.G.C.; writing—review and editing I.E.S., G.C., C.M.F.D., M.G.C.; visuali-zation, supervision, A.R.M., G.R., A.M.; project administration, I.E.S.; funding acquisition, G.R., A.R.M. All authors have read and agreed to the published version of the manuscript.
Data and code availability
The datasets presented in this article are not readily available because are part of the GENYAL Platform for clinical trials in nutrition and health (https://www.food.imdea.org/services/ Platform-Clinical-Trials-Nutrition-and-Health) database. This is a database that is currently registered as a collection under the Spanish rules which by policy of the Center will be public afterwards once the data of the entire expected population is gathered. Requests to access the datasets should be directed to ana.ramirez@imdea.org.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Isabel Espinosa-Salinas, Gonzalo Colmenarejo and Cristina M. Fernández-Díaz.
Contributor Information
Isabel Espinosa-Salinas, Email: mariaisabel.espinosa@imdea.org.
Ana Ramírez de Molina, Email: ana.ramirez@imdea.org.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-022-10923-4.
References
- 1.WHO Coronavirus Disease (COVID-19) Dashboard. https://covid19.who.int.
- 2.Zhou F, et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet. 2020;395:1054–1062. doi: 10.1016/S0140-6736(20)30566-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Guan W-J, et al. Clinical characteristics of coronavirus disease 2019 in China. N. Engl. J. Med. 2020;382:1708–1720. doi: 10.1056/NEJMoa2002032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wu Z, McGoogan JM. Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in China: Summary of a report of 72 314 cases from the Chinese Center for Disease Control and Prevention. JAMA. 2020;323:1239–1242. doi: 10.1001/jama.2020.2648. [DOI] [PubMed] [Google Scholar]
- 5.Yang J, et al. Prevalence of comorbidities and its effects in patients infected with SARS-CoV-2: A systematic review and meta-analysis. Int. J. Infect. Dis. 2020;94:91–95. doi: 10.1016/j.ijid.2020.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Calder PC. Nutrition, immunity and COVID-19. BMJ. 2020 doi: 10.1136/bmjnph-2020-000085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cheng S, et al. Metabolite profiling identifies pathways associated with metabolic risk in humans. Circulation. 2012;125:2222–2231. doi: 10.1161/CIRCULATIONAHA.111.067827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Holmes E, et al. Human metabolic phenotype diversity and its association with diet and blood pressure. Nature. 2008;453:396–400. doi: 10.1038/nature06882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bruzzone C, et al. SARS-CoV-2 infection dysregulates the metabolomic and lipidomic profiles of serum. iScience. 2020;23:101645. doi: 10.1016/j.isci.2020.101645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wang G, et al. Low high-density lipoprotein level is correlated with the severity of COVID-19 patients: An observational study. Lipids Health Dis. 2020;19:204. doi: 10.1186/s12944-020-01382-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhang B, et al. Triglyceride to high-density lipoprotein cholesterol ratio is an important determinant of cardiovascular risk and poor prognosis in coronavirus disease-19: A retrospective case series study. Diabetes Metab. Syndr. Obes. 2020;13:3925–3936. doi: 10.2147/DMSO.S268992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Borrmann H, McKeating JA, Zhuang X. The Circadian clock and viral infections. J. Biol. Rhythms. 2020 doi: 10.1177/0748730420967768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhuang X, Rambhatla SB, Lai AG, McKeating JA. Interplay between circadian clock and viral infection. J. Mol. Med. 2017;95:1283–1289. doi: 10.1007/s00109-017-1592-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Mazzoccoli G, Vinciguerra M, Carbone A, Relógio A. The Circadian clock, the immune system, and viral infections: The intricate relationship between biological time and host-virus interaction. Pathogens. 2020;9:83. doi: 10.3390/pathogens9020083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ramos-Lopez O, et al. Exploring host genetic polymorphisms involved in SARS-CoV infection outcomes: Implications for personalized medicine in COVID-19. Int. J. Genom. 2020;2020:1–8. doi: 10.1155/2020/6901217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Garaulet M, et al. Genetic variants in human CLOCK associate with total energy intake and cytokine sleep factors in overweight subjects (GOLDN population) Eur. J. Hum. Genet. 2010;18:364–369. doi: 10.1038/ejhg.2009.176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Garaulet M, et al. CLOCK gene is implicated in weight reduction in obese patients participating in a dietary programme based on the Mediterranean diet. Int. J. Obes. (Lond.) 2010;34:516–523. doi: 10.1038/ijo.2009.255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Espinosa-Salinas I, et al. Polymorphic appetite effects on waist circumference depend on rs3749474 CLOCK gene variant. Nutrients. 2020;12:1846. doi: 10.3390/nu12061846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Camblor Murube M, et al. Polymorphism of CLOCK gene rs3749474 as a modulator of the circadian evening carbohydrate intake impact on nutritional status in an adult sample. Nutrients. 2020;12:1142. doi: 10.3390/nu12041142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Inal J. Biological factors linking ApoE ε4 variant and severe COVID-19. Curr. Atheroscler. Rep. 2020;22:1–2. doi: 10.1007/s11883-020-00896-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kasparian K, Graykowski D, Cudaback E. Commentary: APOE e4 genotype predicts severe COVID-19 in the UK Biobank community cohort. Front. Immunol. 2020;11:1939. doi: 10.3389/fimmu.2020.01939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Burt TD, et al. Apolipoprotein (apo) E4 enhances HIV-1 cell entry in vitro, and the APOE epsilon4/epsilon4 genotype accelerates HIV disease progression. Proc. Natl. Acad. Sci. USA. 2008;105:8718–8723. doi: 10.1073/pnas.0803526105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Moriarty PM, et al. Lipoprotein(a) and its potential association with thrombosis and inflammation in COVID-19: A testable hypothesis. Curr. Atheroscler. Rep. 2020;22:1–8. doi: 10.1007/s11883-020-00867-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Peng KY, et al. Apolipoprotein E4 genotype compromises brain exosome production. Brain. 2019;142:163–175. doi: 10.1093/brain/awy289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rieker C, et al. Apolipoprotein E4 expression causes gain of toxic function in isogenic human induced pluripotent stem cell-derived endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2019;39:e195–e207. doi: 10.1161/ATVBAHA.118.312261. [DOI] [PubMed] [Google Scholar]
- 26.Wozniak MA, et al. Apolipoprotein E-epsilon 4 protects against severe liver disease caused by hepatitis C virus. Hepatology. 2002;36:456–463. doi: 10.1053/jhep.2002.34745. [DOI] [PubMed] [Google Scholar]
- 27.Finch CE, Kulminski AM. The ApoE locus and COVID-19: Are we going where we have been? J. Gerontol. A. 2020 doi: 10.1093/gerona/glaa200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kuo C-L, et al. APOE e4 genotype predicts severe COVID-19 in the UK Biobank community cohort. J. Gerontol. A. 2020 doi: 10.1093/gerona/glaa131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kuo C-L, et al. ApoE e4e4 genotype and mortality with COVID-19 in UK Biobank. J. Gerontol. A. 2020 doi: 10.1093/gerona/glaa169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Goldstein MR, Poland GA, Graeber CW. Does apolipoprotein E genotype predict COVID-19 severity? QJM. 2020 doi: 10.1093/qjmed/hcaa142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Deelen J, et al. A meta-analysis of genome-wide association studies identifies multiple longevity genes. Nat. Commun. 2019;10:3669. doi: 10.1038/s41467-019-11558-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Shinohara M, et al. APOE2 is associated with longevity independent of Alzheimer’s disease. eLife. 2020;9:e62199. doi: 10.7554/eLife.62199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Saeed A, et al. Remnant-like particle cholesterol, low-density lipoprotein triglycerides, and incident cardiovascular disease. J. Am. Coll. Cardiol. 2018;72:156–169. doi: 10.1016/j.jacc.2018.04.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Mack S, et al. A genome-wide association meta-analysis on lipoprotein (a) concentrations adjusted for apolipoprotein (a) isoforms. J. Lipid Res. 2017;58:1834–1844. doi: 10.1194/jlr.M076232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.König M, et al. Cohort profile: Role of lipoproteins in cardiovascular disease: The LipidCardio study. BMJ Open. 2019;9:534. doi: 10.1136/bmjopen-2019-030097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Abondio P, et al. The genetic variability of APOE in different human populations and its implications for longevity. Genes. 2019;10:222. doi: 10.3390/genes10030222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Sebastiani P, et al. A serum protein signature of APOE genotypes in centenarians. Aging Cell. 2019;18:e13023. doi: 10.1111/acel.13023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kulminski AM, et al. Protective role of the apolipoprotein E2 allele in age-related disease traits and survival: Evidence from the Long Life Family Study. Biogerontology. 2016;17:893–905. doi: 10.1007/s10522-016-9659-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kuo C-L, Pilling LC, Atkins JL, Kuchel GA, Melzer D. ApoE e2 and aging-related outcomes in 379,000 UK Biobank participants. medRxiv. 2020 doi: 10.1101/2020.02.12.20022459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Almqvist J, et al. Neurological manifestations of coronavirus infections: A systematic review. Ann. Clin. Transl. Neurol. 2020 doi: 10.1002/acn3.51166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Monteiro-Junior RS. COVID-19: Thinking about further mental and neurological disorders. Med. Hypotheses. 2020;143:109894. doi: 10.1016/j.mehy.2020.109894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Murakami M, Tognini P. The Circadian clock as an essential molecular link between host physiology and microorganisms. Front. Cell Infect. Microbiol. 2019;9:469. doi: 10.3389/fcimb.2019.00469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Edgar RS, et al. Cell autonomous regulation of herpes and influenza virus infection by the circadian clock. Proc. Natl. Acad. Sci. USA. 2016;113:10085–10090. doi: 10.1073/pnas.1601895113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Herichova I, Zsoldosova K, Vesela A, Zeman M. Effect of angiotensin II infusion on rhythmic clock gene expression and local renin-angiotensin system in the aorta of Wistar rats. Endocr. Regul. 2014;48:144–151. doi: 10.4149/endo_2014_03_144. [DOI] [PubMed] [Google Scholar]
- 45.Meira e cruz M, Miyazawa M, Gozal D. Putative contributions of circadian clock and sleep in the context of SARS-CoV-2 infection. Eur. Respir J. 2020;55:2001023. doi: 10.1183/13993003.01023-2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lim RK, Wambier CG, Goren A. Are night shift workers at an increased risk for COVID-19? Med. Hypotheses. 2020;144:110147. doi: 10.1016/j.mehy.2020.110147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ray S, Reddy AB. COVID-19 management in light of the circadian clock. Nat. Rev. Mol. Cell Biol. 2020;21:494–495. doi: 10.1038/s41580-020-0275-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Loria-Kohen V, et al. Polymorphism in the CLOCK gene may influence the effect of fat intake reduction on weight loss. Nutrition. 2016;32:453–460. doi: 10.1016/j.nut.2015.10.013. [DOI] [PubMed] [Google Scholar]
- 49.Stefan N, Birkenfeld AL, Schulze MB, Ludwig DS. Obesity and impaired metabolic health in patients with COVID-19. Nat. Rev. Endocrinol. 2020;16:341–342. doi: 10.1038/s41574-020-0364-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Yang J, Hu J, Zhu C. Obesity aggravates COVID-19: A systematic review and meta-analysis. J. Med. Virol. 2020 doi: 10.1002/jmv.26237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Caci G, et al. COVID-19 and obesity: Dangerous liaisons. J. Clin. Med. 2020;9:2511. doi: 10.3390/jcm9082511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Bessi V, et al. Influence of ApoE genotype and clock T3111C interaction with cardiovascular risk factors on the progression to Alzheimer’s disease in subjective cognitive decline and mild cognitive impairment patients. J. Pers. Med. 2020;10:45. doi: 10.3390/jpm10020045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Chen H, Huang C, You C, Wang Z, Si-qing H. Polymorphism of CLOCK gene rs 4580704 C > G is associated with susceptibility of Alzheimer’s disease in a Chinese population. Arch. Med. Res. 2013;44:203–207. doi: 10.1016/j.arcmed.2013.01.002. [DOI] [PubMed] [Google Scholar]
- 54.Durnin, J. V. & Fidanza, F. Evaluation of nutritional status. Bibl Nutr Dieta 20–30 (1985). [PubMed]
- 55.Norton, K., Olds, T. & Mazza, J. C. Técnicas de Medición en Antropometría. In Antropométrica: Un libro de referencia sobre mediciones corporales humanas para la Educación en Deportes y Salud 23–30 (Biosystem, Servicio Educativo, 2000).
- 56.Aguirre-Jaime A, et al. Validation of a food intake frequency questionnaire adapted for the study and monitoring of the adult population of the Canary Islands, Spain. Rev. Esp. Salud Publ. 2008;82:509–518. doi: 10.1590/S1135-57272008000500006. [DOI] [PubMed] [Google Scholar]
- 57.Espinosa-Salinas I, et al. GCKR rs780094 polymorphism as a genetic variant involved in physical exercise. Genes. 2019;10:570. doi: 10.3390/genes10080570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Zhang L, Ptáček LJ, Fu Y-H. Diversity of human clock genotypes and consequences. Prog. Mol. Biol. Transl. Sci. 2013;119:51–81. doi: 10.1016/B978-0-12-396971-2.00003-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Espinosa-Salinas, M. I. Identificación de SNPs Implicados en la Diferente Respuesta a Componentes de la Dieta y Asociación con Enfermedades Relacionadas con la Alimentación: Estudios Nutrigenéticos. (Universidad Autónoma de Madrid, 2017).
- 60.QuantStudio qPCR Product Portfolio. https://www.thermofisher.com/us/en/home/life-science/pcr/real-time-pcr/real-time-pcr-instruments/quantstudio-qpcr-product-portfolio.html (n.d.).
- 61.Applied Biosystem. TaqMan® Genotyper Software. Getting Started Guide. (Life Technologies Corporation, 2010).
- 62.R: What is R? https://www.r-project.org/about.html (n.d.).
- 63.Azur MJ, Stuart EA, Frangakis C, Leaf PJ. Multiple imputation by chained equations: What is it and how does it work? Int. J. Methods Psychiatr. Res. 2011;20:40–49. doi: 10.1002/mpr.329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Schomaker M, Heumann C. Bootstrap inference when using multiple imputation. Stat. Med. 2018;37:2252–2266. doi: 10.1002/sim.7654. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets presented in this article are not readily available because are part of the GENYAL Platform for clinical trials in nutrition and health (https://www.food.imdea.org/services/ Platform-Clinical-Trials-Nutrition-and-Health) database. This is a database that is currently registered as a collection under the Spanish rules which by policy of the Center will be public afterwards once the data of the entire expected population is gathered. Requests to access the datasets should be directed to ana.ramirez@imdea.org.