

Abstract
Background
Growing evidence proved that signal transducer and activators of transcription (STAT) proteins are cytoplasmic transcription factors known to play key roles in many cellular biological processes and may be prognostic predictors of some cancers. However, the role of each STAT family members in breast cancer (BRCA) is diverse and controversial. This study aimed to systematic mine the prognostic significance and immune infiltration of STAT family member in human BRCA.
Methods
Based on The Cancer Genome Atlas (TCGA) database, we used the Oncomine, Gene Expression Profiling Interactive Analysis (GEPIA) and The Human Protein Atlas to analyze the expression of STAT family members in normal human breast and tumor tissues. The Kaplan-Meier Plotter, GEPIA and PrognoScan were utilized to assess the prognostic value of different STATs in BRCA. Then we used the cBioPortal, STRING, GeneMANIA and Metascape to make further mutation analysis, protein-protein interaction (PPI) analysis and subsequent functional enrichment analysis. Finally, the “ESTIMATE” and “ggcorrplot” package of R 17 software were used for immune infiltration analysis.
Results
STAT2 [P<0.01, hazard ratio (HR) =1.23, 95% confidence interval (CI): 1.07–1.42] and STAT3 (P=0.018, HR =0.69, 95% CI: 0.51–0.94) could be an independent risk factor for predicting overall survival (OS). STAT4 could be used as an independent predictor of distant metastasis-free survival in BRCA based on both GSE19615 (P=0.021, HR =0.21, 95% CI: 0.06–0.79) and GSE2034 (P=0.015, HR =0.57, 95% CI: 0.37–0.90) datasets. Meanwhile, STAT5A, STAT5B and STAT6 also have been shown to independently predict the prognosis of BRCA. Additionally, the functional mechanisms of STAT4 co-expressed genes were mainly focused on immune-related pathways and its expression was associated with immune checkpoint-associated genes and immunomodulators in BRCA.
Conclusions
Our study mined the prognostic significance of STAT family members in BRCA and their correlation with immune infiltration. The results suggest that individual STATs, except STAT1, may act as a prognostic biomarker for BRCA and provide a reference for further potential immunotherapies.
Keywords: Signal transducer and activator of transcription (STAT), overall survival (OS), immune infiltration, breast cancer (BRCA)
Introduction
Breast cancer (BRCA) is currently one of the leading causes of cancer-related death in women worldwide, and the incidence rate is still increasing (1). At present, there are multiple of treatments for BRCA, including surgery, chemotherapy, radiotherapy, and endocrine therapy (2,3). Also, increasing numbers of researches have shown that the immune system plays an important role in BRCA, such as detecting the response to standard treatment or predicting the long-term survival for BRCA patients (4). Meanwhile, with the discovery of immune checkpoints, which regulate immune activation, has led to new advances for the treatment of BRCA (5). Besides, many prognostic models based on multigene testing already exist in clinical practice, such as OncotypeDX, EndoPredict, the Breast Cancer Indicator, etc. (6). However, problems of high cost and poor reproducibility continue to limit their practical clinical application. Therefore, identifying some dependable prognostic biomarkers and performing a comprehensive analysis to assess its roles in the process of tumorigenesis is still necessary for BRCA patients.
Signal transducer and activator of transcription (STAT) family members are transcriptional factors that were first discovered by Wegenka et al. in 1994 (7). Seven STAT family members have been confirmed in the human genome: STAT1 (chromosomal location: 2q32.2), STAT2 (12q13.3), STAT3 (17q21.2), STAT4 (2q32.2), STAT5A (17q21.2), STAT5B (17q21.2), and STAT6 (12q13.3) (8). STATs need to be triggered by extracellular signaling ligands such as hormones, cytokines, and growth factors, and then perform biological functions via the Janus tyrosine kinase (JAK)-STAT signaling pathway (9,10). Previous studies have reported that STATs are engaged in numerous normal physiological cellular processes, including cell proliferation, differentiation, apoptosis, and immune system regulation (11-13). However, many irregular actions and mechanisms can occur in different STAT family members, resulting in aberrant STAT regulation, which leads to different pathological events in cancer, such as malignant cell conversion and metastasis. At present, several STAT family members are even regarded as oncogenes, while others are not (10,14). Thus, the relevance of immune infiltration and prognostic value of STAT family members in BRCA still requires further study.
In this research, we aim to conduct an in-depth analysis of the STAT family using multiple databases, such as The Cancer Genome Atlas (TCGA) and Kaplan-Meier plotter, and summarize the role of STATs in the clinical prognosis and immune infiltration in BRCA, including gene expression, survival status, genetic mutation, immune infiltration, and related cellular pathways. This will provide a comprehensive insight into the role of different STAT members in cancer initiation and progression, and will help us to better understand the prospect of their potential clinical implications. We present the following article in accordance with the REMARK reporting checklist (available at https://gs.amegroups.com/article/view/10.21037/gs-22-189/rc).
Methods
Datasets source and patients selection
To facilitate our analysis, we mainly used bioinformatics techniques to assist in the prognostic analysis of STAT family members. TCGA (https://tcga-data.nci.nih.gov/) is a public database that aims to provide researchers with available datasets to improve oncology-related diagnostic methods, treatment standards, and in order to facilitate cancer prevention (15). In this study, we included 1,880 BRCA patients who had complete survival data in the TCGA database and removed patient information with missing data. Meanwhile, we downloaded and analyzed several datasets from the PrognoScan platform and directly collated into a table format for our interpretation. Finally, we downloaded the RNA-sequencing (RNA-seq) expression data of STAT family members from the Xena system (https://xenabrowser.net/datapages/) for immune profiling. The study was conducted in accordance with the Declaration of Helsinki (as revised in 2013).
Oncomine
The messenger RNA (mRNA) expression levels of the STAT family members in different types of cancers were analyzed through the Oncomine database. Oncomine is a publicly available online oncology-related database and a web-based data mining platform, which integrates many bioinformatics resources such as LocusLink, Kyoto Encyclopedia of Genes and Genomes (KEGG), and Biocarta, etc. (16-18). In this study, genes with a fold change (FC) of 2 and a P value <0.01were ranked in the top 10%, and the data types were set as mRNA.
Gene Expression Profiling Interactive Analysis (GEPIA)
GEPIA is a time-saving tool that can unlock the value of large genome data in TCGA and Genotype-Tissue Expression (GTEx). It also can provide an interactive web platform for gene expression analysis, and is a good complement to other existing gene analysis tools, such as cBioPortal and Expression Atlas (19). In this study, we used a |log2FC| cutoff of 1, q-value cutoff of 0.01, and the log scale was set as yes. We then performed comparisons between the tumor and normal breast tissues, analyzing different gene expression, associated prognostic value, and pathological stage by GEPIA.
The Human Protein Atlas (HPA)
The HPA database can map human tissues, cells, and organs based on proteomic, transcriptomics, and systems biology data. Forty-four major normal tissues and some cancer tissues were included by immunohistochemistry. In our study, the protein expression of STAT family members in normal human breast and tumor tissues were confirmed by this database (20).
Kaplan-Meier plotter
The Kaplan-Meier plotter is a website that includes numerous modules, such as gene expression information and clinical outcome data. This website was utilized to assess the prognostic value of different STATs in BRCA patients, which were constructed based on gene chips and RNA-seq data from public databases including Gene Expression Omnibus (GEO), European Genome-phenome Archive (EGA), and TCGA. The log rank test was used to compare the Kaplan-Meier survival curves. All statistical comparisons were considered significant at P<0.05 (21).
PrognoScan
Based on a large collection of publicly available cancer gene microarray datasets, the PrognoScan database use the minimal P value method to assess the relationship between gene expression and prognosis in various cancer patients (22). We extracted some STAT family-related datasets in BRCA and analyzed the expression of STATs in relation to overall survival (OS), relapse-free survival (RFS), disease-free survival (DFS), and distant metastasis-free survival (DMFS) in BRCA patients. All statistical comparisons were considered significant at P<0.05.
Survival analysis
To assess the prognostic value of individual STATs, the patients was divided into two groups based on their median gene expression (high vs. low expression) in the datasets from TCGA and PrognoScan platform. OS was utilized to assess the prognosis of BRCA patients in the datasets from both sources, whereas DFMS, DFS and RFS were only used to evaluate the prognosis of patients in the PrognoScan platform. Using the Kaplan-Meier method, survival analysis was carried out and gene expression groups were compared using the log-rank test. Cox regression analysis was used to screen the independent prognostic factors by calculating the Cox P value, adjusted hazard ratio (HR) and 95% confidence interval (CI).
cBioPortal
cBioPortal is an interactive resource that facilitates access to cancer genomic datasets (23). Based on genetic mutations-related data, the modules of STATs were provided by cBioPortal. We used this database to select the BRCA datasets, with 817 cases (TCGA, Cell 2015) (24) selected for further analysis. The Oncoprint, OS, and DFS of STAT family members were analyzed online using this method.
GeneMANIA
GeneMANIA is a user-friendly website that can predict gene function, analyze gene lists, and prioritize genes for functional assays (25). We used it to explore the relationship among different STAT family members.
Search Tool for the Retrieval of Interacting Genes (STRING)
STRING can provide important evaluation clues and integrate the protein-protein interactions (PPIs), including both direct and indirect associations (26). In this study, protein-protein associations in the STATs family from co-expression data were analyzed by STRING.
Metascape
Metascape is an efficient functional annotation analysis tool comprising interactome analysis, gene annotation, functional enrichment, and membership search (27). The Metascape database was used to explore the enrichment pathways associated with the STAT4 co-expressed genes.
Tumor Immune Estimation Resource (TIMER)
The TIMER database can be utilized to analyze the infiltration of immune-related cells in various types of tumors (28). In this study, TIMER database analysis allowed us to investigate the relationship between STAT4 expression levels and the degree of infiltration of various immune-related cells in different BRCA subtypes.
Follow-up
Follow-up was defined as the time between the diagnosis of BRCA and the occurrence of the outcome event. In this study, the primary clinical outcome endpoint was OS, which was defined as the time from the diagnosis of BRCA to death from any cause. The secondary study endpoints were DFMS, DFS and RFS. The survival status of patients lost to follow-up was recorded as “blank” in the TCGA database and excluded accordingly.
Statistical analysis
All gene expression data were standardized by log2 transformation. Normal and cancer tissues were compared using two sets of t-test. Relevance analysis between two variables was performed using Spearman or Pearson tests. All statistical analyses were processed using the “ESTIMATE” and “ggcorrplot” package of R 17 software (https://CRAN.R-project.org, version 4.0.2). A two-tailed P<0.05 was considered to be statistically significant.
Results
Divergent expression of STATs in BRCA patients
To investigate the expression levels of different STATs in BRCA patients, we analyzed the mRNA expression of STATs through the Oncomine database. The results suggested that, compared to normal tissues, the mRNA expression levels of STAT1 and STAT2 were elevated in BRCA tissues (Figure 1A), while those of STAT3, STAT4, STAT5A, and STAT5B were reduced in BRCA. We also used the GEPIA database to compare the mRNA expression levels of STATs between BRCA and normal breast tissues (Figure 1B). The results suggested that, compared to normal tissues, the expression level of STAT1 was elevated in BRCA tissues, while those of STAT5A and STAT5B were reduced. These results, which were obtained via the GEPIA database, were almost identical to those previously obtained through the Oncomine database.
Figure 1.
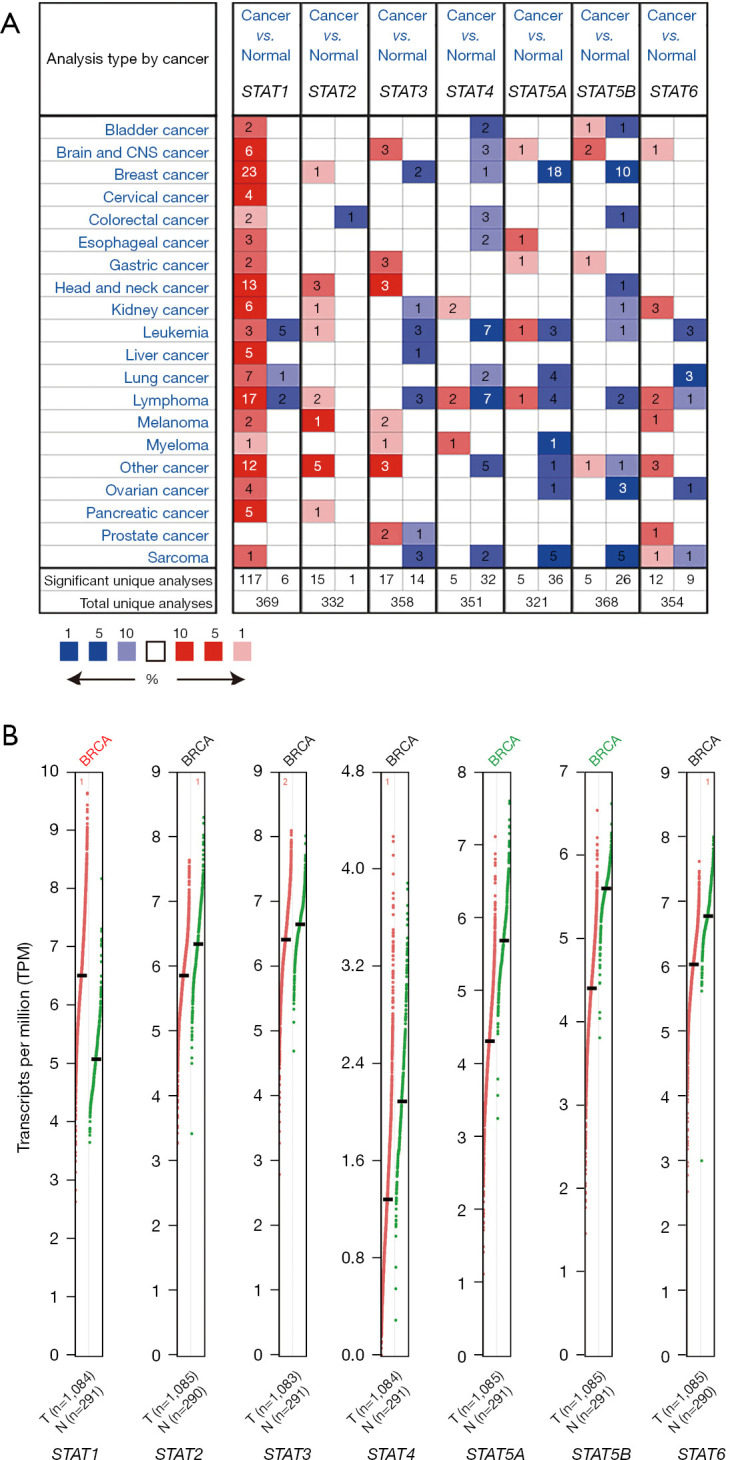
Expression of STATs in BRCA. (A) mRNA expression of STAT family members in different cancer types (Oncomine). The graph shows the number of datasets with obvious mRNA expression among the STAT family genes: upregulated (red) and downregulated (blue). The criteria were as follows: P value: 0.01, fold change: 2, gene rank: 10%, date type: mRNA, and analysis type: cancer vs. normal tissue. (B) Expression of STATs in BRCA (GEPIA). As shown in this graphic: expression levels of STAT1 were higher in BRCA tissues, while the expression levels of STAT5A and STAT5B were lower in BRCA vs. normal tissues (P<0.05). BRCA, breast cancer; CNS, central nervous system; GEPIA, Gene Expression Profiling Interactive Analysis; mRNA, messenger RNA; STAT, signal transducer and activator of transcription; N, normal tissue; T, tumor tissue.
Additionally, some typical immunohistochemical images were selected from the HPA database to analyze the protein expressions of the STAT family in human breast normal and tumor tissues. The immunohistochemical images illustrated that STAT1, STAT2, STAT3, and STAT6 had low coloring in normal breast tissues, but high staining in BRCA (Figure S1). Also, STAT5A and STAT5B had lower levels of protein expression in BRCA tissues compared with normal tissues, indicating a similar result to mRNA transcriptional levels.
We then explored the relationship between the pathological stage of BRCA patients and the expression of STATs using the GEPIA database. As shown in Figure 2, we found that the STAT3 and STAT6 groups were obviously variable, whereas the STAT1, STAT2, STAT4, STAT5A, and STAT5B groups were not obviously different. These results indicated that STAT expressions were markedly different in BRCA compared with normal tissues, and that STAT3 may play an important role in the development and progression of BRCA.
Figure 2.

Correlation between STAT expression and tumor pathological stage in breast cancer patients (GEPIA). The expression of STAT3 and STAT6 were related to the pathological stage of BRCA patients (P<0.05). TPM, transcripts per million; BRCA, breast cancer; GEPIA, Gene Expression Profiling Interactive Analysis; STAT, signal transducer and activator of transcription.
Prognostic value of STATs in BRCA patients
To analyze the prognostic value of STATs in BRCA patients, we used Kaplan-Meier plotter website to conduct a subgroup analysis. As shown in Figure 3, the mRNA expression levels of STAT4 (HR =0.78, 95% CI: 0.65–0.94, P=0.0099), STAT5A (HR =0.74, 95% CI: 0.61–0.89, P=0.0017), and STAT6 (HR =0.76, 95% CI: 0.63–0.92, P=0.0043) were significantly correlated with long OS in BRCA patients.
Figure 3.

Prognostic value of STATs in breast cancer (Kaplan-Meier plotter). High levels of mRNA expression in STAT4, STAT5A, and STAT6 were significantly correlated with long OS in breast cancer patients. HR, hazard ratio; mRNA, messenger RNA; OS, overall survival; STAT, signal transducer and activator of transcription.
Furthermore, we used the GEPIA database to validate the value of the variable expression of STATs in the progression of BRCA. The OS and DFS curves are displayed in Figure 4. As for BRCA patients, there was no obvious association between any of the STAT family members and the DFS rates. Meanwhile, we found that the patients with high transcriptional levels of STAT4 (HR =0.56, P=0.00062) were significantly related to long OS. With the exception of the STAT4 group, all other members of the STAT family did not appear to show a remarkable effect on OS. Therefore, we subsequently extracted several datasets from the PrognoScan platform and further analyzed different STATs expression in relation to OS, RFS, DFS, and DMFS in BRCA patients (Table 1), aiming to further evaluate our previous analysis. The results showed that STAT2 (P<0.01, HR =1.23, 95% CI: 1.07–1.42) and STAT3 (P=0.018, HR =0.69, 95% CI: 0.51–0.94) could be an independent risk factor for predicting OS. STAT4 could be used as an independent predictor of DMFS in BRCA based on both GSE19615 (P=0.021, HR =0.21, 95% CI: 0.06–0.79) and GSE2034 (P=0.015, HR =0.57, 95% CI: 0.37–0.90) datasets. Meanwhile, STAT5A, STAT5B and STAT6 also have been shown to independently predict the prognosis of BRCA.
Figure 4.

Prognostic value of the mRNA expression of STAT family members in BRCA (GEPIA). BRCA patients with a high transcriptional level of STAT4 (P=0.00062) were obviously related to long OS. BRCA, breast cancer; GEPIA, Gene Expression Profiling Interactive Analysis; HR, hazard ratio; mRNA, messenger RNA; OS, overall survival; STAT, signal transducer and activator of transcription; TPM, transcripts per million.
Table 1. STAT family gene expression was related to the prognosis of breast cancer in PrognoScan.
| Gene | Dataset | Endpoint | Cutpoint | Cox P value | Ln(HR) | HR (95% CI) |
|---|---|---|---|---|---|---|
| STAT1 | GSE7378 | OS | 158 | 0.734006 | 0.422818 | 1.53 (0.96–2.42) |
| E-TABM-158 | OS | 198 | 0.690165 | 0.102520 | 1.11 (0.88–1.39) | |
| GSE19615 | DFS | 76 | −1.49087 | −0.231171 | 0.79 (0.44–1.42) | |
| GSE3143 | DFS | 54 | −15.3825 | 0.034494 | 1.04 (0.48–2.22) | |
| GSE7849 | DFS | 249 | 0.543037 | 0.223837 | 1.25 (0.88–1.78) | |
| GSE3494-GPL96 | RFS | 204 | 0.380639 | 0.121462 | 1.13 (0.88–1.44) | |
| GSE4922-GPL96 | RFS | 87 | 0.570071 | 0.024605 | 1.02 (0.61–1.72) | |
| GSE2990 | RFS | 159 | 0.856193 | 0.462988 | 1.59 (0.88–2.88) | |
| GSE7390 | RFS | 125 | 0.729704 | 0.215864 | 1.24 (0.89–1.73) | |
| GSE9195 | DMFS | 87 | 0.843094 | 0.069123 | 1.07 (0.80–1.43) | |
| GSE12093 | DMFS | 136 | 1.48027 | 0.413829 | 1.51 (0.92–2.50) | |
| GSE11121 | DMFS | 200 | 0.904803 | 0.225317 | 1.25 (0.88–1.77) | |
| GSE1378 | DMFS | 286 | −1.27494 | −0.236758 | 0.79 (0.56–1.12) | |
| GSE9893 | DMFS | 117 | 0.59709 | 0.058867 | 1.06 (0.76–1.48) | |
| GSE2034 | DMFS | 125 | −1.23639 | −0.072966 | 0.93 (0.66–1.31) | |
| STAT2 | GSE3143 | OS | 158 | 0.123157 | 0.722624 | 2.06 (0.82–5.16) |
| GSE9893 | OS | 155 | 0.00447014 | 0.209301 | 1.23 (1.07–1.42) | |
| GSE1456-GPL97 | OS | 159 | 0.741445 | −0.133722 | 0.87 (0.40–1.94) | |
| E-TABM-158 | OS | 117 | 0.604802 | −0.220720 | 0.80 (0.35–1.85) | |
| GSE7390 | OS | 198 | 0.671654 | 0.116688 | 1.12 (0.66–1.93) | |
| GSE7849 | DFS | 76 | 0.89763 | −0.128759 | 0.88 (0.12–6.25) | |
| GSE7378 | DFS | 54 | 0.710306 | −0.322114 | 0.72 (0.13–3.97) | |
| GSE4922-GPL97 | DFS | 249 | 0.40129 | 0.303484 | 1.35 (0.67–2.75) | |
| GSE12276 | RFS | 204 | 0.7863 | 0.055055 | 1.06 (0.71–1.57) | |
| GSE6532-GPL570 | RFS | 87 | 0.706449 | −0.199444 | 0.82 (0.29–2.31) | |
| GSE9195 | RFS | 77 | 0.784835 | 0.233873 | 1.26 (0.24–6.77) | |
| GSE1379 | RFS | 60 | 0.688543 | 0.178464 | 1.20 (0.50–2.86) | |
| GSE1456-GPL96 | RFS | 159 | 0.90511 | −0.027443 | 0.97 (0.62–1.53) | |
| GSE1456-GPL97 | RFS | 159 | 0.391313 | 0.341762 | 1.41 (0.64–3.07) | |
| E-TABM-158 | RFS | 117 | 0.604802 | −0.220720 | 0.80 (0.35–1.85) | |
| GSE2990 | RFS | 125 | 0.904064 | 0.032345 | 1.03 (0.61–1.75) | |
| GSE7390 | RFS | 198 | 0.245853 | −0.126170 | 0.88 (0.71–1.09) | |
| GSE19615 | DMFS | 115 | 0.153716 | −0.868095 | 0.42 (0.13–1.38) | |
| GSE6532-GPL570 | DMFS | 87 | 0.695463 | −0.202646 | 0.82 (0.30–2.25) | |
| GSE9195 | DMFS | 77 | 0.255748 | −2.201030 | 0.11 (0.00–4.93) | |
| GSE12093 | DMFS | 136 | 0.956642 | −0.017426 | 0.98 (0.52–1.84) | |
| GSE11121 | DMFS | 200 | 0.444782 | 0.458926 | 1.58 (0.49–5.13) | |
| GSE2034 | DMFS | 286 | 0.0981089 | −0.424718 | 0.65 (0.40–1.08) | |
| E-TABM-158 | DMFS | 117 | 0.190977 | 1.181818 | 3.26(0.55-19.17) | |
| GSE2990 | DMFS | 125 | 0.767727 | 0.271797 | 1.31 (0.22–7.97) | |
| GSE7390 | DMFS | 198 | 0.304212 | 0.277772 | 1.32 (0.78–2.24) | |
| STAT3 | GSE3143 | OS | 158 | 0.695188 | −0.147051 | 0.86 (0.41–1.80) |
| GSE9893 | OS | 155 | 0.0177795 | −0.366907 | 0.69 (0.51–0.94) | |
| GSE1456-GPL97 | OS | 159 | 0.840228 | 0.104908 | 1.11 (0.40–3.08) | |
| E-TABM-158 | OS | 117 | 0.918875 | 0.020681 | 1.02 (0.69–1.52) | |
| GSE7390 | OS | 198 | 0.907517 | −0.030143 | 0.97 (0.58–1.61) | |
| GSE7849 | DFS | 76 | 0.0141442 | 1.593150 | 4.92 (1.38–17.56) | |
| GSE7378 | DFS | 54 | 0.456313 | −0.241100 | 0.79 (0.42–1.48) | |
| GSE4922-GPL96 | DFS | 249 | 0.971322 | 0.007425 | 1.01 (0.67–1.51) | |
| GSE12276 | RFS | 204 | 0.687901 | −0.072622 | 0.93 (0.65–1.33) | |
| GSE6532-GPL570 | RFS | 87 | 0.70237 | −0.344585 | 0.71 (0.12–4.15) | |
| GSE9195 | RFS | 77 | 0.0217172 | 1.335230 | 3.80 (1.22–11.89) | |
| GSE1379 | RFS | 60 | 0.894997 | −0.060561 | 0.94 (0.38–2.31) | |
| GSE1456-GPL97 | RFS | 159 | 0.89862 | −0.065406 | 0.94 (0.34–2.56) | |
| E-TABM-158 | RFS | 117 | 0.918875 | 0.020681 | 1.02 (0.69–1.52) | |
| GSE2990 | RFS | 62 | 0.0108244 | −1.112200 | 0.33 (0.14–0.77) | |
| GSE2990 | RFS | 125 | 0.837496 | −0.048294 | 0.95 (0.60–1.51) | |
| GSE2990 | RFS | 62 | 0.0378638 | −0.593717 | 0.55 (0.32–0.97) | |
| GSE7390 | RFS | 198 | 0.646099 | 0.082683 | 1.09 (0.76–1.55) | |
| GSE19615 | DMFS | 115 | 0.468102 | −0.499143 | 0.61 (0.16–2.34) | |
| GSE6532-GPL570 | DMFS | 87 | 0.70237 | −0.344585 | 0.71 (0.12–4.15) | |
| GSE9195 | DMFS | 77 | 0.135511 | 1.010010 | 2.75 (0.73–10.34) | |
| GSE9195 | DMFS | 77 | 0.144656 | 0.890767 | 2.44 (0.74–8.07) | |
| GSE12093 | DMFS | 136 | 0.0164555 | −0.532524 | 0.59 (0.38–0.91) | |
| GSE11121 | DMFS | 200 | 0.0203471 | −0.822380 | 0.44 (0.22–0.88) | |
| GSE2034 | DMFS | 286 | 0.0157849 | −0.254330 | 0.78 (0.63–0.95) | |
| E-TABM-158 | DMFS | 117 | 0.117872 | −0.417841 | 0.66 (0.39–1.11) | |
| GSE2990 | DMFS | 125 | 0.905535 | −0.036515 | 0.96 (0.53–1.76) | |
| GSE2990 | DMFS | 54 | 0.0858196 | −0.982659 | 0.37 (0.12–1.15) | |
| GSE7390 | DMFS | 198 | 0.0827706 | 0.387547 | 1.47 (0.95–2.28) | |
| STAT4 | GSE3143 | OS | 158 | 0.638658 | −0.158958 | 0.85 (0.44–1.66) |
| GSE9893 | OS | 155 | 0.0693526 | 0.316011 | 1.37 (0.98–1.93) | |
| GSE1456-GPL96 | OS | 159 | 0.337157 | −0.435590 | 0.65 (0.27–1.57) | |
| E-TABM-158 | OS | 117 | 0.731327 | −0.069909 | 0.93 (0.63–1.39) | |
| GSE7390 | OS | 198 | 0.899221 | −0.021423 | 0.98 (0.70–1.36) | |
| GSE7849 | DFS | 76 | 0.898074 | −0.056164 | 0.95 (0.40–2.23) | |
| GSE7378 | DFS | 54 | 0.0599192 | −1.718460 | 0.18 (0.03–1.07) | |
| GSE4922-GPL96 | DFS | 249 | 0.531574 | 0.183205 | 1.20 (0.68–2.13) | |
| GSE12276 | RFS | 204 | 0.370653 | −0.093580 | 0.91 (0.74–1.12) | |
| GSE6532-GPL570 | RFS | 87 | 0.753865 | 0.064398 | 1.07 (0.71–1.60) | |
| GSE9195 | RFS | 77 | 0.376928 | −0.541672 | 0.58 (0.17–1.93) | |
| GSE1456-GPL96 | RFS | 159 | 0.598349 | 0.245888 | 1.28 (0.51–3.19) | |
| E-TABM-158 | RFS | 117 | 0.731327 | −0.069909 | 0.93 (0.63–1.39) | |
| GSE2990 | RFS | 62 | 0.686218 | −0.069183 | 0.93 (0.67–1.31) | |
| GSE19615 | DMFS | 115 | 0.0210692 | −1.539200 | 0.21 (0.06–0.79) | |
| GSE6532-GPL570 | DMFS | 87 | 0.753865 | 0.064398 | 1.07 (0.71–1.60) | |
| GSE9195 | DMFS | 77 | 0.142219 | −1.103740 | 0.33 (0.08–1.45) | |
| GSE12093 | DMFS | 136 | 0.774322 | −0.096607 | 0.91 (0.47–1.76) | |
| GSE11121 | DMFS | 200 | 0.057605 | −0.581530 | 0.56 (0.31–1.02) | |
| GSE2034 | DMFS | 286 | 0.0147119 | −0.556861 | 0.57 (0.37–0.90) | |
| E-TABM-158 | DMFS | 117 | 0.986949 | −0.003965 | 1.00 (0.62–1.60) | |
| GSE2990 | DMFS | 125 | 0.254872 | −0.531531 | 0.59 (0.24–1.47) | |
| GSE7390 | DMFS | 198 | 0.992948 | −0.001456 | 1.00 (0.72–1.38) | |
| STAT5A | GSE3143 | OS | 158 | 0.210806 | −0.2375 | 0.79 (0.54–1.14) |
| GSE9893 | OS | 155 | 0.440632 | −0.185188 | 0.83 (0.52–1.33) | |
| GSE1456-GPL96 | OS | 159 | 0.158685 | −0.706401 | 0.49 (0.18–1.32) | |
| E-TABM-158 | OS | 117 | 0.501951 | −0.213658 | 0.81 (0.43–1.51) | |
| GSE7390 | OS | 198 | 0.71127 | −0.082341 | 0.92 (0.60–1.42) | |
| GSE7849 | DFS | 76 | 0.543889 | 0.238656 | 1.27 (0.59–2.74) | |
| GSE7378 | DFS | 54 | 0.0314761 | −2.14162 | 0.12 (0.02–0.83) | |
| GSE4922-GPL96 | DFS | 249 | 0.0172677 | −0.825435 | 0.44 (0.22–0.86) | |
| GSE12276 | RFS | 204 | 0.167323 | −0.228702 | 0.80 (0.58–1.10) | |
| GSE6532-GPL570 | RFS | 87 | 0.00227358 | −1.144590 | 0.32 (0.15–0.66) | |
| GSE9195 | RFS | 77 | 0.923972 | −0.074396 | 0.93 (0.20–4.28) | |
| GSE1378 | RFS | 60 | 0.248078 | −0.225438 | 0.80 (0.54–1.17) | |
| GSE1379 | RFS | 60 | 0.452682 | −0.228444 | 0.80 (0.44–1.44) | |
| GSE1456-GPL96 | RFS | 159 | 0.185957 | −0.683025 | 0.51 (0.18–1.39) | |
| E-TABM-158 | RFS | 117 | 0.501951 | −0.213658 | 0.81 (0.43–1.51) | |
| GSE2990 | RFS | 125 | 0.284496 | −0.312392 | 0.73 (0.41–1.30) | |
| GSE7390 | RFS | 198 | 0.763745 | 0.053695 | 1.06 (0.74–1.50) | |
| GSE19615 | DMFS | 115 | 0.00884115 | −1.729470 | 0.18 (0.05–0.65) | |
| GSE6532-GPL570 | DMFS | 87 | 0.00227358 | −1.144590 | 0.32 (0.15–0.66) | |
| GSE9195 | DMFS | 77 | 0.48064 | −0.659811 | 0.52 (0.08–3.23) | |
| GSE12093 | DMFS | 136 | 0.0598301 | −1.15754 | 0.31 (0.09–1.05) | |
| GSE11121 | DMFS | 200 | 0.0358548 | −0.681225 | 0.51 (0.27–0.96) | |
| GSE2034 | DMFS | 286 | 0.00786138 | −0.590146 | 0.55 (0.36–0.86) | |
| E-TABM-158 | DMFS | 117 | 0.406866 | −0.327397 | 0.72 (0.33–1.56) | |
| GSE2990 | DMFS | 125 | 0.319595 | −0.384966 | 0.68 (0.32–1.45) | |
| GSE7390 | DMFS | 198 | 0.996648 | −0.000910 | 1.00 (0.65–1.53) | |
| STAT5B | GSE12417-GPL150 | OS | 158 | 0.502612 | 0.243754 | 1.28 (0.63–2.60) |
| GSE12417-GPL191 | OS | 155 | 0.0739188 | 0.306504 | 1.36 (0.97–1.90) | |
| GSE12417-GPL197 | OS | 159 | 3.15E−07 | −2.43983 | 0.09 (0.03–0.22) | |
| GSE12417-GPL207 | OS | 117 | 0.314488 | 0.276225 | 1.32 (0.77–2.26) | |
| GSE12417-GPL243 | OS | 198 | 0.51792 | 0.142256 | 1.15 (0.75–1.77) | |
| GSE12417-GPL155 | DFS | 76 | 0.633366 | −0.238224 | 0.79 (0.30–2.10) | |
| GSE12417-GPL204 | DFS | 54 | 0.131027 | −0.90912 | 0.40 (0.12–1.31) | |
| GSE12417-GPL222 | DFS | 249 | 0.0659483 | −0.661075 | 0.52 (0.26–1.04) | |
| GSE12417-GPL160 | RFS | 204 | 0.182215 | −0.092228 | 0.91 (0.80–1.04) | |
| GSE12417-GPL171 | RFS | 77 | 0.899623 | −0.089730 | 0.91 (0.23–3.69) | |
| GSE12417-GPL188 | RFS | 60 | 0.033972 | 0.459418 | 1.58 (1.04–2.42) | |
| GSE12417-GPL195 | RFS | 159 | 0.0667261 | −0.967095 | 0.38 (0.14–1.07) | |
| GSE12417-GPL211 | RFS | 117 | 0.163118 | −0.805688 | 0.45 (0.14–1.39) | |
| GSE12417-GPL226 | RFS | 62 | 0.0297399 | −0.866101 | 0.42 (0.19–0.92) | |
| GSE12417-GPL231 | RFS | 125 | 0.419839 | −0.240821 | 0.79 (0.44–1.41) | |
| GSE12417-GPL242 | RFS | 198 | 0.675771 | −0.083283 | 0.92 (0.62–1.36) | |
| GSE12417-GPL149 | DMFS | 115 | 0.0255209 | −1.57939 | 0.21 (0.05–0.82) | |
| GSE12417-GPL163 | DMFS | 87 | 0.066610 | 0.964541 | 2.62 (0.94–7.35) | |
| GSE12417-GPL180 | DMFS | 77 | 0.326454 | −0.909009 | 0.40 (0.07–2.48) | |
| GSE12417-GPL181 | DMFS | 136 | 0.496678 | −0.283021 | 0.75 (0.33–1.70) | |
| GSE12417-GPL186 | DMFS | 200 | 0.00553487 | −1.30901 | 0.27 (0.11–0.68) | |
| GSE12417-GPL192 | DMFS | 286 | 0.0750032 | −0.465471 | 0.63 (0.38–1.05) | |
| GSE12417-GPL214 | DMFS | 117 | 0.372841 | −0.613899 | 0.54 (0.14–2.09) | |
| GSE12417-GPL228 | DMFS | 125 | 0.458577 | −0.291181 | 0.75 (0.35–1.61) | |
| GSE12417-GPL229 | DMFS | 54 | 0.694294 | −0.129606 | 0.88 (0.46–1.68) | |
| GSE12417-GPL236 | DMFS | 125 | 0.76281 | −0.215466 | 0.81 (0.20–3.27) | |
| GSE12417-GPL239 | DMFS | 198 | 0.289715 | 0.252804 | 1.29 (0.81–2.06) | |
| STAT6 | GSE3143 | OS | 158 | 0.993998 | −0.002656 | 1.00 (0.50–1.99) |
| GSE9893 | OS | 155 | 0.00582855 | −0.217493 | 0.80 (0.69–0.94) | |
| GSE1456-GPL96 | OS | 159 | 0.0581333 | −0.846861 | 0.43 (0.18–1.03) | |
| E-TABM-158 | OS | 117 | 0.997088 | 0.0010673 | 1.00 (0.56–1.78) | |
| GSE7390 | OS | 198 | 0.189067 | −0.177569 | 0.84 (0.64–1.09) | |
| GSE7849 | DFS | 76 | 0.232317 | 1.06851 | 2.91 (0.50–6.81) | |
| GSE7378 | DFS | 54 | 0.286081 | 0.703717 | 2.02 (0.55–7.36) | |
| GSE4922-GPL96 | DFS | 249 | 0.0879759 | −0.313232 | 0.73 (0.51–1.05) | |
| GSE12276 | RFS | 204 | 0.0735396 | −0.165275 | 0.85 (0.71–1.02) | |
| GSE6532-GPL570 | RFS | 87 | 0.0561677 | −0.971252 | 0.38 (0.14–1.03) | |
| GSE9195 | RFS | 77 | 0.573397 | −0.384807 | 0.68 (0.18–2.60) | |
| GSE1378 | RFS | 60 | 0.361895 | −0.272733 | 0.76 (0.42–1.37) | |
| GSE1456-GPL96 | RFS | 159 | 0.948818 | −0.013512 | 0.99 (0.65–1.49) | |
| E-TABM-158 | RFS | 117 | 0.959771 | 0.022162 | 1.02 (0.43–2.42) | |
| GSE2990 | RFS | 125 | 0.742506 | −0.136701 | 0.87 (0.39–1.97) | |
| GSE7390 | RFS | 198 | 0.488795 | −0.142638 | 0.87 (0.58–1.30) | |
| GSE19615 | DMFS | 115 | 0.309889 | −0.678707 | 0.51 (0.14–1.88) | |
| GSE6532-GPL570 | DMFS | 87 | 0.0561677 | −0.971252 | 0.38 (0.14–1.03) | |
| GSE9195 | DMFS | 77 | 0.706615 | −0.232473 | 0.79 (0.24–2.66) | |
| GSE12093 | DMFS | 136 | 0.205107 | −0.73496 | 0.48 (0.15–1.49) | |
| GSE11121 | DMFS | 200 | 0.000547358 | −0.603841 | 0.55 (0.39–0.77) | |
| GSE2034 | DMFS | 286 | 0.0381629 | −0.47926 | 0.62 (0.39–0.97) | |
| E-TABM-158 | DMFS | 117 | 0.300091 | 0.550172 | 1.73 (0.61–4.91) | |
| GSE2990 | DMFS | 125 | 0.936266 | −0.044824 | 0.96 (0.32–2.87) |
OS, overall survival; DFS, disease-free survival; RFS, relapse-free survival; DMFS, disease metastasis-free survival; HR, hazard ratio; CI, confidence interval. STAT, signal transducer and activator of transcription.
Genetic mutations, clinical relevance, and protein-protein network of STATs in BRCA
We utilized the cBioPortal online tool to explore the genetic mutations of STATs in BRCA. As shown in Figure 5A, two or more mutations were observed in different BRCA isoforms, with deep depletion alterations being more prevalent in mixed ductal and lobular breast carcinoma samples. As shown in Figure 5B, STATs were mutated in 72 samples of 816 patients in BRCA, accounting for 9%. In addition, STAT1, STAT2, STAT3, STAT4, STAT5A, STAT5B, and STAT6 were altered in the queried BRCA samples in the order of 1.7%, 0.7%, 3.0%, 2.2%, 2.9%, 3.0%, and 1.5%, respectively. Furthermore, the results of this database also showed a statistically significant relationship between the existence of mutations in STATs and OS (Figure 5C, P=0.0350) in BRCA patients, but not with DFS (Figure 5D, P=0.835).
Figure 5.

Mutational profile and the protein-protein interaction network of STAT family in breast cancer. (A,B) The STAT gene mutation and expression analyses in BRCA (cBioPortal). “+” means the data was included in the analysis. (C,D) STATs were altered in 72 samples of 816 patients with BRCA, accounting for 9%. Alterations in STATs were correlated with short OS in breast cancer patients, but not with DFS. (E,F) A protein-protein interaction network was constructed using the STRING and GeneMANIA databases, and shows the STATs and STATs-interacting proteins. BRCA, breast cancer; DFS, disease-free survival; STAT, signal transducer and activator of transcription; OS, overall survival.
Moreover, we further explored the potential interaction of these differentially expressed STATs using the STRING database. Several lines and points were acquired in the network (Figure 5E). The variably-expressed STATs were related to cell growth, signal transduction, and transcriptional activation. As shown in Figure 5F, the GeneMANIA analysis also indicated that the functions of the STAT family member proteins and their relevant molecules (CLNK, SH2D2A, DAPP1, SH2D1B, STAP1, SHB, PIK3R3, SH2D1A, SH3BP2, and SH2B2) were mainly related to the JAK-STAT cascade, protein binding and bridging, SH3/SH2 adaptor activity, and cellular response to growth hormone stimulus.
Co-expressed genes of STAT4 and their associated enrichment pathways
Given the remarkable prognostic correlation between STAT4 and BRCA, we searched the GEPIA database for the top 100 related genes that are co-expressed with STAT4, and performed an online data enrichment analysis of Gene Ontology (GO) and KEGG for these co-expressed genes using the Metascape database (Figure 6). The results indicated that the co-expressed genes of STAT4 were mainly enriched in the adaptive immune response, T cell activation, T cell receptor signaling pathway, regulation of cytokine production, negative regulation of immune system processes, and other biological processes. In addition, the main molecular functions, which were regulated by the co-expressed genes of STAT4, were T cell receptor binding, major histocompatibility complex (MHC) protein binding, SH2 domain binding, and non-membrane spanning protein tyrosine kinase activity. Also, the KEGG enrichment pathway, which was influenced by co-expressed genes of STAT4, was mainly focused on T cell receptor signaling pathway, T helper (Th)1, and Th2 cell differentiation, cell adhesion molecules (CAMs), natural killer cell-mediated cytotoxicity, as well as the nuclear factor-kappa B (NF-κB) and chemokine signaling pathways.
Figure 6.

Co-expressed genes of STAT4 and their associated enrichment pathways analyzed using the Metascape database demonstrated that they were mainly enriched in numerous immune-related biological processes, such as the adaptive immune response, T cell activation, T cell receptor signaling pathway, and negative regulation of immune system processes. STAT, signal transducer and activator of transcription.
Immune infiltration relevance of STAT4 in BRCA patients
Based on the fact that STAT4 was significantly enriched in the adaptive immune response and T cell receptor signaling pathways, we investigated the relationship between STAT4 gene expression and the degree of immune cell infiltration in BRCA. According to previous studies (29,30), we mainly selected six tumor-infiltrating immune cell types, including B cells, CD4+ T cells, CD8+ T cells, neutrophils, macrophages, and dendritic cells, to analyze the data from TCGA database using R 17 software (version 4.0.2). The results suggested that STAT4 expression was remarkably positively related to the level of infiltration of B cells (P=1.31e−26, r=0.31, 95% CI: 0.26–0.37), CD4+ T cells (P=2.98e−166, r=0.71, 95% CI: 0.67–0.74), CD8+ T cells (P=1.89e−131, r=0.65, 95% CI: 0.61–0.69), neutrophils (P=1.67e−171, r=0.71, 95% CI: 0.68–0.74), macrophages (P=7.14e−18, r=0.26, 95% CI: 0.20–0.31), and dendritic cells (P=3.7e−178, r=0.72, 95% CI: 0.69–0.75) (Figure 7A).
Figure 7.

Relationship between STAT4 and immune cell expression and analysis of immune infiltration scores. (A) The positive relationship between STAT4 gene expression and the degree of immune cell infiltration in breast cancer. (B) Based on three algorithms, the correlation between immune score and STAT4 expression was calculated, and the results showed that STAT4 expression was significantly correlated with immune infiltration. BRCA, breast cancer; CI, confidence interval; STAT, signal transducer and activator of transcription; TCGA, The Cancer Genome Atlas.
To further depict the underlying immune pattern of STAT4, we calculated ESTIMATE, immune, and stromal for all BRCA samples in TCGA database using the ESTIMATE package, in order to assess their correlation with STAT4 expression. As shown in Figure 7B, we observed positive correlations between STAT4 and the stromal, immune, and ESTIMATE scores in the BRCA dataset.
As shown in Figure S2, we further analyzed the correlation between STAT4 expression and immune cell infiltration in various BRCA subtypes. The results indicated that STAT4 exhibited a distinctive connection with the level of infiltration of B cells (r=0.277, P=2.14e−04), CD4+ T cells (r=0.305, P=4.30e−05), neutrophils (r=0.495, P=3.96e−12), and dendritic cells (r=0.256, P=6.66e−04) in the basal-like BRCA group. Additionally, the infiltration of CD8+ T cells (r=0.263, P=2.59e−02), CD4+ T cells (r=0.448, P=7.85e−05), neutrophils (r=0.558, P=3.55e−07), and dendritic cells (r=0.363, P=1.71e−03) were also related to the high expression level of STAT4 in the HER2 BRCA group. In the luminal A BRCA patient group, STAT4 expression was positively correlated with the infiltration of CD8+ T cells (r=0.291, P=1.59e−11), CD4+ T cells (r=0.328, P=1.95e−14), neutrophils (r=0.592, P=3.21e−50), and dendritic cells (r=0.448, P=6.73e−27). In the luminal B group, it was associated with the infiltration of CD4+ T cells (r=0.249, P=5.05e−04), neutrophils (r=0.589, P=2.58e−19), and dendritic cells (r=0.418, P=1.54e−09).
Correlations between STAT4 expression and immune checkpoints and immunomodulators
Immune checkpoint is a regulatory molecule that plays a suppressive role in the immune system. It can control the time and intensity of the immune response to maintain self-tolerance, prevent autoimmune reactions, and minimize tissue damage (31). In this study, we initially selected eight immune checkpoint-associated genes (CD274, CTLA4, HAVCR2, LAG3, PDCD1, PDCD1LG2, TIGIT, and SIGLEC15) to analyze their expression difference between BRCA and normal tissues (Figure 8A). We then further extracted STAT4 and the expression information of two kinds of immune checkpoint pathway genes from the TCGA database. As shown in Figure 8B, we observed that STAT4 had a relationship with immune checkpoint pathway genes. Furthermore, as shown in Figure 9, our finding also revealed that STAT4 was positively correlated with the majority of immunomodulators in BRCA.
Figure 8.
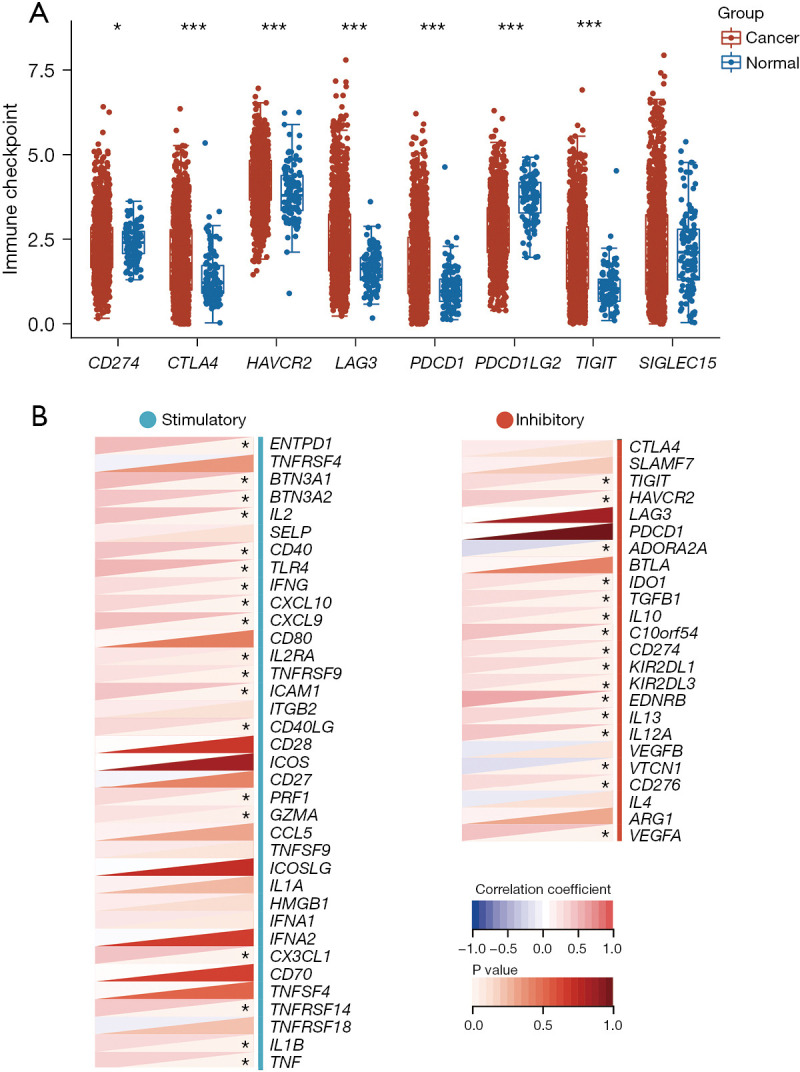
Expression of immune checkpoint genes in breast cancer and their relationship with STAT4. (A) Expression of eight immune checkpoint genes in breast tumor tissues and normal tissues, where the horizontal axis represents different genes and the vertical axis represents the distribution of expression of that gene. The different colors represent different groups, and the asterisks represent the levels of significance: *, P<0.05; ***, P<0.001. (B) Relationship between STAT4 and two kinds of immune checkpoint pathway genes (inhibitory, stimulatory). STAT, signal transducer and activator of transcription.
Figure 9.

Correlation between STAT4 and 150 immunomodulators (immunostimulators, MHC, chemokines, immunoinhibitors, and receptors). The asterisks represent levels of significance: *, P<0.05. MHC, major histocompatibility complex; STAT, signal transducer and activator of transcription.
Discussion
In this study, we mainly carried out an in-depth analysis of the mRNA expression of STATs in BRCA, and explored the relationship between STAT family genes and BRCA using multiple online databases. The result suggested that compared to normal tissues, the expression levels of STAT1 and STAT2 were higher in BRCA tissues, while those of STAT3, STAT4, STAT5A, and STAT5B were lower. We also found that STAT3 was significantly associated with the pathological stage of BRCA, which indicates that STAT3 might play an important role in the occurrence and development of BRCA (32,33). More importantly, through mutual verification by several database tools, we observed that BRCA patients with a high transcriptional level of STAT4 were apparently related with long OS. This indicates that STAT4 could have the potential to become a prognostic marker for BRCA.
To explain the molecular mechanism of STAT4 on the onset and progression of BRCA, we further selected the top 100 related genes that were co-expressed with STAT4 and analyzed their associated enrichment pathways. The results showed these co-expressed genes were mainly enriched in the adaptive immune response, T cell activation, T cell receptor signaling pathway, regulation of cytokine production, and negative regulation of immune system processes. Furthermore, the association between the immune system and STAT4 expression was investigated in depth using several databases, and revealed that STAT4 was associated with the degree of infiltration of multiple immune cells, the expression of immune checkpoint-related genes, and a majority of immunomodulators in BRCA. Therefore, mining the prognostic significance and immune infiltration of STAT family members in BRCA might facilitate the discovery of new biomarkers and further our understanding of important immunotherapies.
As an important component of signal transduction, STAT family proteins are associated with the development of a variety of cancers. Previous study has confirmed that activated STAT1 acts as a tumor suppressor in cancer cells (34). Also, the anti-carcinogenic effect of STAT1 primarily depends on the phosphorylation of tyrosine synthase (35). However, the role of STAT1 is puzzling due to the fact that Greenwood et al. observed that STAT1 positive triple-negative tumors are more aggressive (36). This is probably attributable to the fact that STAT1 can upregulate the action of various pro-inflammatory cytokines, enzymes, and chemokines, such as tumor necrosis factor α (TNF-α), cyclooxygenase 2 (COX2), and inducible nitric oxide synthase (iNOS) (37-39). In addition, the mRNA expression of STAT1 was obviously higher in BRCA tissues, but no correlation of STAT1 on the OS rate of BRCA patients was observed in our study.
Unlike STAT1, STAT2 is a cardinal and specific effector of type I interferon (IFN-I) signaling, which plays an important role in tumor immune surveillance (40). STAT2 can also regulate the expression of genes involved in the differentiation and recruitment of immune effector cells to the tumor sites (41). Therefore, it is also crucial for STAT2 to be studied. In recent years, inflammation has attracted attention with regards to tumorigenesis and malignant progression. There is some evidence that STAT family proteins, particularly STAT3, play a key role in the selective induction and maintenance of an oncogenic inflammatory microenvironment, both at the onset of malignant transformation and during cancer progression (42). Furthermore, STAT3 has been shown to be particularly active in BRCA, where it promotes cancer progression by stimulating cell proliferation, increasing angiogenesis, influencing escape to the immune system, and providing resistance to apoptosis (33,43). In our research, STAT3 expression was related to the pathological stage of BRCA, which could perhaps be explained by the mechanism described above.
STAT5A and STAT5B, two other important members of the STAT family, are two isoforms of STAT5. Similar to STAT3, structural activation of STAT5 is a major cause of tumorigenesis (44). However, emerging data has proposed that activated STAT5 could act as a predictor of better prognosis in BRCA patients (45). According to the database exploration in our study, the enhanced mRNA expression of STAT5A portended better OS in all BRCAs. This appears to be associated with the fact that STAT5A can inhibit BRCA cells via E-cadherin-mediated junctions (46). As for STAT6, we determined that the elevated mRNA expression of STAT6 was correlated with better OS using the Kaplan-Meier plotter. Also, the findings of Wang et al. (47) are generally consistent our results. Nevertheless, further investigation of larger populations is necessary.
Immunotherapy is becoming increasingly popular in cancer prevention and treatment research, and immune cells in the tumor microenvironment are regarded as an important determinant of clinical outcomes and the reaction to immunotherapy (48). In our study, we found that STAT4 was highly expressed in BRCA compared to normal breast tissue, and its higher expression was associated with better prognosis. Through bioinformatics analysis, we also found that STAT4 co-expressed genes were focused on immune-related pathways, and the STAT4 expression level was associated with the infiltration level of multiple immune cells in BRCA. Previous studies have also shown that STAT4 is engaged in a series of immune response processes, including differentiation and activation of immune cells, interferon-gamma (IFN-γ) production, interleukin (IL)-12 and -23, and IFN signaling in immune cells, and so on (49,50). In recent years, Anderson et al. found that STAT4 deficiency decreases the anti-tumor immune response and promotes the accumulation of immunosuppressive myeloid cell populations to promote tumor metastasis in head and neck squamous cell carcinoma (51). In contrast, knockdown or inhibition of STAT4 can suppress cancer cell migration, invasion, and metastasis in gastric and colorectal cancers (50,52). However, in terms of BRCA, STAT4 was still poorly studied in terms of immunity. Although there are multiple treatments for BRCA (2), it is important that more effective treatments are continually being explored. This study may offer some detailed information about immunization to support the design of new immunotherapies.
There were some limitations in this study that should be noted. Firstly, all of the data analyzed were obtained from online databases, and thus, we our findings need to clarify through more cellular experiments and long-term clinical studies. In addition, it is also necessary to further explore the potential mechanisms of action and clinically relevant applications of different STATs in BRCA.
Conclusions
In summary, based on the validation of a large amount of data, our study mined the prognostic significance and immune infiltration relevance of STAT family members. The results suggest that individual STATs, except STAT1, may act as a prognostic biomarker for BRCA and provide a reference for further potential immunotherapies.
Acknowledgments
We thank The Cancer Genome Atlas (TCGA) Database for sharing a large amount of data and the convenience provided by several online database tools.
Funding: This study was funded by the National Natural Science Fund of China (No. 82003183).
Ethical Statement: The authors are accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. The study was conducted in accordance with the Declaration of Helsinki (as revised in 2013).
Open Access Statement: This is an Open Access article distributed in accordance with the Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License (CC BY-NC-ND 4.0), which permits the non-commercial replication and distribution of the article with the strict proviso that no changes or edits are made and the original work is properly cited (including links to both the formal publication through the relevant DOI and the license). See: https://creativecommons.org/licenses/by-nc-nd/4.0/.
Reporting Checklist: The authors have completed the REMARK reporting checklist. Available at https://gs.amegroups.com/article/view/10.21037/gs-22-189/rc
Conflicts of Interest: All authors have completed the ICMJE uniform disclosure form (available at https://gs.amegroups.com/article/view/10.21037/gs-22-189/coif). The authors have no conflicts of interest to declare.
(English Language Editor: A. Kassem)
References
- 1.Britt KL, Cuzick J, Phillips KA. Key steps for effective breast cancer prevention. Nat Rev Cancer 2020;20:417-36. 10.1038/s41568-020-0266-x [DOI] [PubMed] [Google Scholar]
- 2.Tong CWS, Wu M, Cho WCS, et al. Recent Advances in the Treatment of Breast Cancer. Front Oncol 2018;8:227. 10.3389/fonc.2018.00227 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Veronesi U, Boyle P, Goldhirsch A, et al. Breast cancer. Lancet 2005;365:1727-41. 10.1016/S0140-6736(05)66546-4 [DOI] [PubMed] [Google Scholar]
- 4.Savas P, Salgado R, Denkert C, et al. Clinical relevance of host immunity in breast cancer: from TILs to the clinic. Nat Rev Clin Oncol 2016;13:228-41. 10.1038/nrclinonc.2015.215 [DOI] [PubMed] [Google Scholar]
- 5.Esteva FJ, Hubbard-Lucey VM, Tang J, et al. Immunotherapy and targeted therapy combinations in metastatic breast cancer. Lancet Oncol 2019;20:e175-86. 10.1016/S1470-2045(19)30026-9 [DOI] [PubMed] [Google Scholar]
- 6.Chen H, Pu S, Yu S, et al. A nomogram based on CENPP expression for survival prediction in breast cancer. Gland Surg 2021;10:1874-88. 10.21037/gs-21-30 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wegenka UM, Lütticken C, Buschmann J, et al. The interleukin-6-activated acute-phase response factor is antigenically and functionally related to members of the signal transducer and activator of transcription (STAT) family. Mol Cell Biol 1994;14:3186-96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Berg T. Signal transducers and activators of transcription as targets for small organic molecules. Chembiochem 2008;9:2039-44. 10.1002/cbic.200800274 [DOI] [PubMed] [Google Scholar]
- 9.Darnell JE, Jr. STATs and gene regulation. Science 1997;277:1630-5. 10.1126/science.277.5332.1630 [DOI] [PubMed] [Google Scholar]
- 10.Shao F, Pang X, Baeg GH. Targeting the JAK/STAT Signaling Pathway for Breast Cancer. Curr Med Chem 2021;28:5137-51. 10.2174/0929867328666201207202012 [DOI] [PubMed] [Google Scholar]
- 11.Miklossy G, Hilliard TS, Turkson J. Therapeutic modulators of STAT signalling for human diseases. Nat Rev Drug Discov 2013;12:611-29. 10.1038/nrd4088 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Pencik J, Pham HT, Schmoellerl J, et al. JAK-STAT signaling in cancer: From cytokines to non-coding genome. Cytokine 2016;87:26-36. 10.1016/j.cyto.2016.06.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Villarino AV, Kanno Y, O'Shea JJ. Mechanisms and consequences of Jak-STAT signaling in the immune system. Nat Immunol 2017;18:374-84. 10.1038/ni.3691 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sasidharan Nair V, Toor SM, Ali BR, et al. Dual inhibition of STAT1 and STAT3 activation downregulates expression of PD-L1 in human breast cancer cells. Expert Opin Ther Targets 2018;22:547-57. 10.1080/14728222.2018.1471137 [DOI] [PubMed] [Google Scholar]
- 15.Tomczak K, Czerwińska P, Wiznerowicz M. The Cancer Genome Atlas (TCGA): an immeasurable source of knowledge. Contemp Oncol (Pozn) 2015;19:A68-77. 10.5114/wo.2014.47136 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Rhodes DR, Yu J, Shanker K, et al. ONCOMINE: a cancer microarray database and integrated data-mining platform. Neoplasia 2004;6:1-6. 10.1016/S1476-5586(04)80047-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pruitt KD, Maglott DR. RefSeq and LocusLink: NCBI gene-centered resources. Nucleic Acids Res 2001;29:137-40. 10.1093/nar/29.1.137 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kanehisa M, Goto S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res 2000;28:27-30. 10.1093/nar/28.1.27 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tang Z, Li C, Kang B, et al. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res 2017;45:W98-W102. 10.1093/nar/gkx247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Chen W, Dai X, Chen Y, et al. Significance of STAT3 in Immune Infiltration and Drug Response in Cancer. Biomolecules 2020;10:834. 10.3390/biom10060834 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Voutsadakis IA. 8p11.23 Amplification in Breast Cancer: Molecular Characteristics, Prognosis and Targeted Therapy. J Clin Med 2020;9:3079. 10.3390/jcm9103079 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mizuno H, Kitada K, Nakai K, et al. PrognoScan: a new database for meta-analysis of the prognostic value of genes. BMC Med Genomics 2009;2:18. 10.1186/1755-8794-2-18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cerami E, Gao J, Dogrusoz U, et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov 2012;2:401-4. 10.1158/2159-8290.CD-12-0095 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ciriello G, Gatza ML, Beck AH, et al. Comprehensive Molecular Portraits of Invasive Lobular Breast Cancer. Cell 2015;163:506-19. 10.1016/j.cell.2015.09.033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Franz M, Rodriguez H, Lopes C, et al. GeneMANIA update 2018. Nucleic Acids Res 2018;46:W60-4. 10.1093/nar/gky311 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Szklarczyk D, Franceschini A, Wyder S, et al. STRING v10: protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res 2015;43:D447-52. 10.1093/nar/gku1003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zhou Y, Zhou B, Pache L, et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat Commun 2019;10:1523. 10.1038/s41467-019-09234-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Li T, Fan J, Wang B, et al. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res 2017;77:e108-10. 10.1158/0008-5472.CAN-17-0307 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Li Q, Pan Y, Cao Z, et al. Comprehensive Analysis of Prognostic Value and Immune Infiltration of Chromobox Family Members in Colorectal Cancer. Front Oncol 2020;10:582667. 10.3389/fonc.2020.582667 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Sorrelle N, Ganguly D, Dominguez ATA, et al. Improved Multiplex Immunohistochemistry for Immune Microenvironment Evaluation of Mouse Formalin-Fixed, Paraffin-Embedded Tissues. J Immunol 2019;202:292-9. 10.4049/jimmunol.1800878 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Li B, Chan HL, Chen P. Immune Checkpoint Inhibitors: Basics and Challenges. Curr Med Chem 2019;26:3009-25. 10.2174/0929867324666170804143706 [DOI] [PubMed] [Google Scholar]
- 32.Segatto I, Baldassarre G, Belletti B. STAT3 in Breast Cancer Onset and Progression: A Matter of Time and Context. Int J Mol Sci 2018;19:2818. 10.3390/ijms19092818 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Eroglu M, Kokenek-Unal TD, Akin-Bali DF, et al. STAT3 expression is correlated with pathological stage in luminal subtypes of breast carcinoma. Bratisl Lek Listy 2020;121:51-61. 10.4149/BLL_2020_008 [DOI] [PubMed] [Google Scholar]
- 34.Koromilas AE, Sexl V. The tumor suppressor function of STAT1 in breast cancer. JAKSTAT 2013;2:e23353. 10.4161/jkst.23353 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Meissl K, Macho-Maschler S, Müller M, et al. The good and the bad faces of STAT1 in solid tumours. Cytokine 2017;89:12-20. 10.1016/j.cyto.2015.11.011 [DOI] [PubMed] [Google Scholar]
- 36.Greenwood C, Metodieva G, Al-Janabi K, et al. Stat1 and CD74 overexpression is co-dependent and linked to increased invasion and lymph node metastasis in triple-negative breast cancer. J Proteomics 2012;75:3031-40. 10.1016/j.jprot.2011.11.033 [DOI] [PubMed] [Google Scholar]
- 37.Samardzic T, Jankovic V, Stosic-Grujicic S, et al. STAT1 is required for iNOS activation, but not IL-6 production in murine fibroblasts. Cytokine 2001;13:179-82. 10.1006/cyto.2000.0785 [DOI] [PubMed] [Google Scholar]
- 38.Akram M, Kim KA, Kim ES, et al. Selective inhibition of JAK2/STAT1 signaling and iNOS expression mediates the anti-inflammatory effects of coniferyl aldehyde. Chem Biol Interact 2016;256:102-10. 10.1016/j.cbi.2016.06.029 [DOI] [PubMed] [Google Scholar]
- 39.Kaur T, Mukherjea D, Sheehan K, et al. Short interfering RNA against STAT1 attenuates cisplatin-induced ototoxicity in the rat by suppressing inflammation. Cell Death Dis 2011;2:e180. 10.1038/cddis.2011.63 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Arimoto KI, Löchte S, Stoner SA, et al. STAT2 is an essential adaptor in USP18-mediated suppression of type I interferon signaling. Nat Struct Mol Biol 2017;24:279-89. 10.1038/nsmb.3378 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Yue C, Xu J, Tan Estioko MD, et al. Host STAT2/type I interferon axis controls tumor growth. Int J Cancer 2015;136:117-26. 10.1002/ijc.29004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yu H, Pardoll D, Jove R. STATs in cancer inflammation and immunity: a leading role for STAT3. Nat Rev Cancer 2009;9:798-809. 10.1038/nrc2734 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wang T, Niu G, Kortylewski M, et al. Regulation of the innate and adaptive immune responses by Stat-3 signaling in tumor cells. Nat Med 2004;10:48-54. 10.1038/nm976 [DOI] [PubMed] [Google Scholar]
- 44.Bowman T, Garcia R, Turkson J, et al. STATs in oncogenesis. Oncogene 2000;19:2474-88. 10.1038/sj.onc.1203527 [DOI] [PubMed] [Google Scholar]
- 45.Nevalainen MT, Xie J, Torhorst J, et al. Signal transducer and activator of transcription-5 activation and breast cancer prognosis. J Clin Oncol 2004;22:2053-60. 10.1200/JCO.2004.11.046 [DOI] [PubMed] [Google Scholar]
- 46.Sultan AS, Xie J, LeBaron MJ, et al. Stat5 promotes homotypic adhesion and inhibits invasive characteristics of human breast cancer cells. Oncogene 2005;24:746-60. 10.1038/sj.onc.1208203 [DOI] [PubMed] [Google Scholar]
- 47.Wang S, Yu L, Shi W, et al. Prognostic roles of signal transducers and activators of transcription family in human breast cancer. Biosci Rep 2018;38:BSR20171175. 10.1042/BSR20171175 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sun J, Zhang Z, Bao S, et al. Identification of tumor immune infiltration-associated lncRNAs for improving prognosis and immunotherapy response of patients with non-small cell lung cancer. J Immunother Cancer 2020;8:e000110. 10.1136/jitc-2019-000110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Korman BD, Kastner DL, Gregersen PK, et al. STAT4: genetics, mechanisms, and implications for autoimmunity. Curr Allergy Asthma Rep 2008;8:398-403. 10.1007/s11882-008-0077-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Cheng JM, Yao MR, Zhu Q, et al. Silencing of stat4 gene inhibits cell proliferation and invasion of colorectal cancer cells. J Biol Regul Homeost Agents 2015;29:85-92. [PubMed] [Google Scholar]
- 51.Anderson K, Ryan N, Volpedo G, et al. Immune Suppression Mediated by STAT4 Deficiency Promotes Lymphatic Metastasis in HNSCC. Front Immunol 2019;10:3095. 10.3389/fimmu.2019.03095 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zhou X, Xia Y, Su J, et al. Down-regulation of miR-141 induced by helicobacter pylori promotes the invasion of gastric cancer by targeting STAT4. Cell Physiol Biochem 2014;33:1003-12. 10.1159/000358671 [DOI] [PubMed] [Google Scholar]