

Abstract
Skeletal muscle cells are noteworthy for their syncytial nature, with each myofiber accumulating hundreds or thousands of nuclei derived from resident muscle stem cells (MuSCs). These nuclei are accrued through cell fusion, which is controlled by the two essential fusogens Myomaker and Myomerger that are transiently expressed within the myogenic lineage. While the absolute requirement of fusion for muscle development has been known for decades, the underlying need for the magnitude of multinucleation in muscle remains mysterious. Possible advantages of multinucleation include the potential it affords for transcriptional diversity within these massive cells, and as a means of increasing DNA content to support optimal cell size and function. In this article, we review recent advances that elucidate the relationship between myonuclear numbers and establishment of myofiber size, and discuss how this new information refines our understanding of the concept of myonuclear domains (MND), the cytoplasmic volumes that each resident myonucleus can support. Finally, we explore the potential consequences and costs of multinucleation and its impacts on myonuclear transcriptional reserve capacity, growth potential, myofiber size regulation, and muscle adaptability. We anticipate this report will not only serve to highlight the latest advances in the basic biology of syncytial muscle cells but also provide information to help design the next generation of therapeutic strategies to maintain muscle mass and function.
Keywords: skeletal muscle size, multinucleation, myonuclear domain, cell fusion, transcriptional output
1. Introduction
Skeletal myofibers, which grow to become amongst the largest cells of the body attaining sizes ranging between 4.9-42 cm in the human hindlimb [1], are also unique for their level of multinucleation. Between several hundred and thousands of nuclei are acquired and accumulated per myofiber primarily during development, but also upon adaptation to increased workload, or in response to chronic injury as encountered in the muscular dystrophies. The exclusive source of these myonuclei are fusogenic myoblasts, the mononuclear muscle progenitor cells derived from resident muscle stem cells (MuSCs) [2], which can fuse with myofibers to contribute nuclei [3]. Fusion of myoblasts proceeds by a unique stepwise mechanism, where the membrane remodeling activities typically driven by a single fusogen are divided between the two myogenic fusogens Myomaker (gene symbol Mymk) and Myomerger/Myomixer/Minion (gene symbol Mymx) [4–9]. Myomaker, a seven transmembrane protein, is necessary for the formation of membrane hemifusion connections, the initial step that allows for exchange of membrane lipids between fusing cells without mixing of cytosolic contents [10]. Co-expression of Myomerger, drives the reaction beyond the hemifusion step to fusion completion [9]. While the reason for such a divided fusion reaction remains to be ascertained, it is possible a binary system of this nature ensures fidelity between fusing partners, providing a regulatory checkpoint that is particularly important in a tissue that has a lifelong reliance on fusion and myonuclear accrual for optimal maintenance and function. We will not discuss in detail molecular mechanisms underlying myoblast fusion and instead refer readers to recent excellent reviews that explore membrane events associated with the fusion reaction [11–13].
Within myofibers, resident myonuclei share a common cytoplasm and under normal, homeostatic conditions occupy a peripheral, subsarcolemmal position. Resident nuclei are uniformly distributed along the length of the myofibers [14] except at the neuromuscular junctions (NMJs) and myotendinous junctions (MTJs) where they tend to cluster. Each myonucleus is thought to transcriptionally regulate a finite volume of cytoplasm in its immediate vicinity called its myonuclear domain (MND), which forms the basis of the MND theory [15–17]. Derived from Strassburger’s concept of a “sphere of influence”, at the practical level, the concept of an MND reflects the limits of biosynthetic support a single myonucleus can provide, and therefore, serves as a metric of myofiber size. Indeed, MND volume sizes, which tend to be smaller in type I (slow,oxidative) compared to type II (fast, glycolytic) myofibers [18], correlate with myofiber size in both fiber types [19].
In addition to the MND, which is analogous to DNA:cytoplasm ratios in non-muscle cells and is viewed at least partially as a quantitative metric predicated on transcriptional output, there can also be qualitative differences between the resident myonuclei within each myofiber that go beyond fiber-type differences; the strongest evidence of such transcriptional specialization within myofibers is the NMJ, where myonuclei that cluster below the endplate have distinct transcriptional profiles [20–22]. Whether broader transcriptional diversity exists within myofibers, and how such heterogeneity might be related to myonuclear numbers is incipiently understood. Overall, we are only just beginning to elucidate why skeletal myofibers need to both accrue as many nuclei as they do during development, and continue to add more nuclei in the adult through fusion of MuSC descendants; how that accrual and accumulation might be regulated during development and adaptation; and, what the relationship is between myonuclear numbers and myofiber size regulation to optimize growth, function and adaptability. It is these recent and revealing advances that will be reviewed in this article.
2. DNA content as a determinant of cell size
The conventional explanation for multinucleation is based on the long-standing evidence of a correlation between DNA content and cell size in many cell types. This relationship appears to be evolutionarily conserved as revealed by experiments from the early 1900s in sea urchin embryos where size of larval cells varied proportionally with the amount of DNA received [23, 24]. Similarly, in both S. cerevisiae and the green alga Eudorina, cell size scales linearly with DNA content [25, 26]. In mammals, megakaryocytes and trophoblast giant cells increase in size during differentiation with polyploidization [27, 28], while across different human cell-types, nuclear DNA content varies with cell size [29]. In mammalian cardiomyocytes, which can be multinucleated and/or polyploid through endoreplication, growth and volume may correlate with DNA content [30]. Growth in cell size is fundamental to normal development and function of organ systems and organisms, and this is especially true for skeletal muscle, where postnatal increases in muscle size and mass occur predominantly through longitudinal and radial enlargement of existing myofibers. Mirroring findings in rodents, average fiber (Types I and II) cross-sectional area in humans increases from 147.5 μm2 at 1 week after birth, to 7586 μm2 at 18 years of age [31]; consistent with previous quantitative analyses of myofiber size and myonuclear content [32, 33], this growth was found to be associated with a concomitant increase in myonuclear content.
It is important to note that accumulating nuclei through cell fusion is not a common mechanism to increase DNA content. Somatic cells typically increase DNA content and gene copy number through endomitosis or endoreplication, where cycles of DNA re-synthesis occur without accompanying karyokinesis or cytokinesis. In fact, in Drosophila larval body wall muscle, after initial myoblast fusion establishes myonuclear numbers in myofibers, further increases in DNA content, which scale with myofiber size, occur via endoreplication [34, 35]. However, in vertebrate skeletal myofibers resident myonuclei are incapable of DNA synthesis [36, 37]. Therefore, the only mechanism by which vertebrate myofibers can increase DNA content is by accretion of additional nuclei through cell fusion with myoblasts [3].
A causal relationship between MuSC-derived myonuclear accrual and developmental growth in myofiber size was recently tested using novel genetic mouse-models. When myonuclear accretion was blocked in neonates by deleting Myomaker expression exclusively in MuSCs at various neonatal time points, myofiber volume was reduced by 33%-54% when compared to controls [38]. Using a model in which MuSCs were deleted in prepubertal mice, Bachman et al. [39] demonstrated that blocking even relatively modest levels of myonuclear accrual (between 10%-15%) during developmental growth significantly reduced (by 15%-19%) myofiber CSA. These findings confirm that myonuclear (DNA) content is the ultimate determinant of optimal developmental growth and size in skeletal myofibers.
3. Transcriptional diversity within myofibers
An additional reason why myofibers require multinucleation could be that global transcriptional activity within these syncytial cells is allocated between resident nuclei; given the size of these cells, such a division of transcriptional labor would require the accumulation of nuclei in greater numbers, with different cohorts within the same myofiber being assigned divergent and specific transcriptional functions. Alternatively, evidence from lower organisms such as the syncytial filamentous fungus Ashbya gossypii indicates that despite occupying a common cytoplasm, sister nuclei can demonstrate variable and asynchronous behaviours [40, 41]. This raises the possibility that nuclei in syncytial cells, including resident myonuclei, may not be amenable to global regulation and instead act independently, necessitating accrual of large numbers of nuclei for sufficient order to emerge from the incident transcriptional noise.
Whether such transcriptional diversity even exists within myofibers remained unexplored until recently when three groups independently addressed the question by subjecting mouse hindlimb muscles to single-nucleus RNA-sequencing. Unlike single-cell RNA-sequencing, this approach allowed inclusion of syncytial nuclei in the RNA-sequencing analyses [42–44]. Petrany et al. [43], and Kim et al. [44] confirmed strong spatial transcriptional heterogeneity in myofibers with identification of unique transcriptional profiles in myonuclei localized to the post-synaptic NMJs and terminal MTJs. Beyond such functional compartmentalization, Petrany et al. [43] also reported existence of distinct myonuclear populations during postnatal development and in aging muscle. In homeostatic healthy adult muscle though, the transcriptional heterogeneity detected indicated modest variations in gene expression rather than sharply divergent transcriptional profiles. Additional myonuclear sub-types were characterized by Kim et al. [44], which included clusters enriched for transcripts targeting expression of nuclear-encoded mitochondrial genes or endoplasmic reticulum (ER)-associated protein processing genes. It is unclear if this reflected a level of functional compartmentalization that could independently account for the magnitude of multinucleation observed in normal skeletal myofibers. In addition, using a combination of single-nucleus RNA-sequencing and single-nucleus ATAC-sequencing, Dos Santos et al. [42] reported that a majority of the myonuclei within a given myofiber type are synchronized in terms of myosin isoform being expressed. This synchronization of gene expression was coordinated early during development, potentially discounting the likelihood that resident myonuclei within adult myofibers pursue distinct transcriptional profiles, independent of each other. It remains to be ascertained however if this coordination also entails temporal synchronization of gene expression, where expression of a given gene is always turned on and off at the same time across all resident myonuclei; results in Newlands et al. [45] suggest this might not be the case, with the observed transcriptional oscillatory activity indicating some temporal independence between myonuclei, with frequency of such active loci also reflecting developmental maturation of myofibers.
Also, there is evidence that MuSC activation and myonuclear accrual are necessary for NMJ maintenance and regeneration upon injury. When Liu et al. [46] depleted MuSCs, age-related NMJ degeneration was accelerated and occurred at a younger age, with loss of function. Denervation, which causes NMJ disruption, leads to MuSC activation and fusion, with the majority of MuSC-derived myonuclear activity occurring proximal to NMJs, and furthermore, MuSC deletion impaired NMJ regeneration [47, 48]. There is early evidence that at the MTJ, there are myonuclei derived from fibroblasts that have adopted a myogenic fate [49, 50], which raises the intriguing possibility that cells other than MuSC-derived myoblasts contribute to the myonuclear population at the MTJ. Whether the numbers of such non-MuSC-derived myonuclei are also regulated and impact broader transcriptional profiles and myofiber growth remains to be elucidated. Taken together though, excluding the functionally distinct compartments of NMJ and MTJ, current data indicate that a need for transcriptional diversity is unlikely to be primary driver of myonuclear accrual and multinucleation.
3. Myonuclear domain theory
The MND theory described earlier would appear to provide a simple and sufficient explanation for the observed reliance of developmental growth on myonuclear accrual; if the cytoplasmic volume supported by a given myonucleus is limited, the only way a myofiber can grow in size is by adding additional MNDs, which requires nuclear numbers to also increase. On closer scrutiny though, there is a great deal about the theory that remains unclear and is only now being tested and explored. Firstly, it is unknown to what extent MND limits are determined by myonuclear transcriptional output or if other contributory factors also exist. One factor is the ability, or lack thereof, to transport anabolic products over large intracellular distances within myofibers; this might be especially challenging in muscle cells, which are densely and almost entirely packed with sarcomeric myofibrillar content and lack the kind of specialized axonal transport mechanisms found in neurons. Indeed, mRNA diffusion appears to get increasingly restricted as sarcomerogenesis progresses, and becomes dependent on microtubule-based transport in myofibers [51]. Existence of such restrictions on and requirements for cellular transport in myofibers and more broadly in striated muscle cells [51, 52] might explain why other contractile and organellar gene products have also been reported to be confined to a limited area around originating myonuclei [53–55], and could also account for the relatively uniform placement of myonuclei along the length of skeletal myofibers. Geometric and statistical analyses of their spatial distribution indicate that myonuclei are optimally positioned to minimize transport distances [14, 56]. Perturbations in the normal spatial distribution of myonuclei therefore have the potential to disrupt MNDs and compromise myofiber health, with accumulating evidence indicating that misplacement of nuclei can be pathogenic [57–59] and may also contribute to age-related diminution of function [19]. Besides potential disruption in the transport of transcripts from such misplaced myonuclei, another factor to consider is the evidence that syncytial nuclei are responsive to local cytoplasmic differences on very small scales; findings in an Ashbya mutant, where syncytial nuclei get clustered, show that transcriptional activity varies with nuclear position even within such nuclear clusters, revealing a sensitivity to cytoplasmic environments on submicrometer scales [60]. Mislocalization of myonuclei from their normal positions might therefore disrupt their ability to respond to MND-specific transcriptional demands or place them in locations where they receive aberrant signals. Indeed, this might be one the reasons why myonuclei are peripherally positioned, where removed from the dense sarcomeric architecture they can efficiently receive signals that modulate their transcriptional output. Other potential factors contributing to MND maintenance include translational efficiency, and protein and RNA turnover rates [61].
Secondly, the MND theory is premised on the assumption that resident myonuclei support a constant volume of cytoplasm. If true, this would mean that MND volumes should be relatively uniform and myonuclei numbers should scale linearly with cell volume for all myofibers, irrespective of their size and myonuclear content. We already know this not to be the case given the well documented differences in MND volumes between type I and type II myofibers [18]. Furthermore, Bruusgaard et al. [14] and more recently, Hansson et al. [62], have shown in myofibers isolated from both mouse EDL and human vastus lateralis samples, that nuclear numbers in fact scale sub-linearly with volume. What this reveals is that in myofibers with higher numbers of nuclei, each myonucleus supports a larger MND when compared to a nucleus from myofibers that harbor fewer nuclei. This scaling relationship between nuclei numbers and volume was accentuated upon denervation, showing that even under experimental conditions that promote atrophy, myonuclei in myofibers with more nuclei continue to maintain larger MNDs when compared to myofibers with fewer nuclei. The reasons why MND volumes are different between larger and smaller fibers are not definitively known and this is discussed later in the article. Even with the caveats that transcriptional activity could not be assessed in these studies, and that MND volumes are unlikely to be precisely synonymous with myonuclear transcriptional output, these findings confirm that cytoplasmic volumes supported by myonuclei are not uniform, even within the same fiber-type.
It might be telling though, that the same studies also revealed that, at least in type II fibers, myonuclear numbers unfailingly scale linearly with myofiber surface (sarcolemmal) area both during development and in adult muscle [14, 62]. This linear relationship between myonuclear numbers and myofiber surface area raises the intriguing possibility that a metric involving sarcolemmal area, such as surface area to volume (SA:V) ratio, might be an important, yet commonly ignored, determinant that links myonuclear numbers with MND size and myofiber volume. Interestingly, Johnston et al. [63] have proposed that there is an optimal myofiber size, where myofibers try to achieve the largest volume possible for a given surface area, with limits being dictated primarily by diffusion constraints. This hypothesis therefore predicts a bias towards lowering SA:V ratios because doing so affords a metabolic advantage to the animal by minimizing costs of maintaining ionic homeostasis across the sarcolemma. While this was originally proposed based on work in cold-water fishes [63, 64], Jimenez et al. [65] analyzed absolute and fractional ATP costs across a broad range of fiber sizes spanning 16-species to find that there indeed may be a metabolic advantage for fibers to achieve the largest volume possible for a given surface area. Even though experimental evidence of a similar paradigm in mammals is pending, the fact that in rats there is a linear dependence of skeletal muscle oxygen consumption on Na+-K+-ATPase activity [66], which is the primary regulator of membrane potential, supports such a likelihood. Importantly, given the strong evidence that myofiber surface area scales linearly with nuclear numbers [14, 62], a defining role for SA:V ratio in establishing MND sizes only re-emphasizes that myonuclear numbers are the primary determinants of myofiber size. It would however be premature to assume that this regulation occurs purely through modulation of myonuclear transcriptional output; while the focus of this review is mostly limited to the role of nuclei and DNA content in establishment of myofiber size, protein turnover rates [67], and modulation of the non-linear interaction between metabolic rates and diffusion distances within myofibers [65, 68, 69] are just some additional potential mechanisms by which MND volumes might be regulated.
Even if diffusion capabilities are a limiting factor in MND size, the biosynthetic capacity of a myonucleus remains key, because the extent and efficiency of diffusion of an anabolic product are ultimately determined by the amount of that product synthesized. In fact, the magnitude of transcript dispersion from myonuclei has been found to correlate with transcriptional output levels rather than transcript length or encoded protein activity [51]. The idea of a finite MND volume has been commonly interpreted to mean that the intrinsic biosynthetic capacity of a myonucleus in healthy adult myofibers is fully utilized, and limiting [15, 62]. We will explore this assumption as it is directly salient to concepts related to myofiber size regulation and adaptability that have been subject to an intense and lively debate in the field [70–74]. Skeletal myofibers even after fully attaining adult size, retain a lifelong ability to respond to elevated workloads by gaining in volume and myofibrillar content. The generation of such physiological hypertrophy to optimally and sustainably meet functional demands we call “adaptive growth.” This definition encompasses both a potential reliance on the recruitment of additional myonuclei through fusion to augment volume by increasing the number of MNDs (Fig. 1A), and/or the harnessing of any available residual plasticity in pre-existing myonuclei to increase MND volumes (Fig. 1B). So defined, adaptive growth is understood to be distinct from the normal variability observed in both non-pathological MND volumes within and between fiber types [14, 18, 62] and upon pathology or aging [19, 75]. And importantly, it does not conflate an ability to enlarge MND through pharmacological or genetic interventions, with the capacity to develop sustained, physiological hypertrophy [71, 74].
Fig. 1. Relationship between myonuclear content, myonuclear domain and growth in syncytial myofibers.
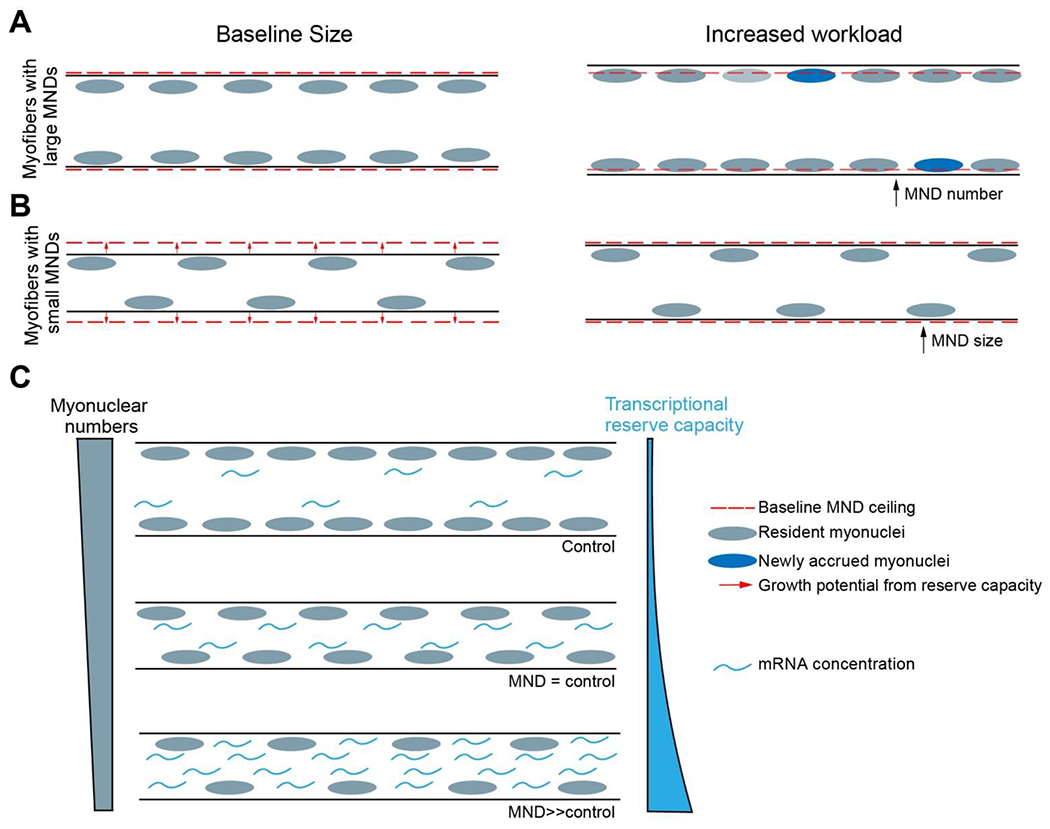
Each resident myonucleus supports a finite volume of cellular content called its myonuclear domain (MND), which reflects its biosynthetic capacity. The sum total of all MND volumes within a myofiber dictates a theoretical ceiling on the size it can potentially achieve, termed baseline MND ceiling (dashed red line). (A) In myofibers with large MNDs myofiber baseline size (left panel) approximates this ceiling with resident myonuclear biosynthetic capacity posited to be fully utilized. Adaptive growth in response to increased workload (right panel) in such myofibers requires accrual of new myonuclei (blue nuclei) and is therefore generated by increasing MND numbers. The baseline MND ceiling gets reset at a higher level (not depicted). (B) In myofibers with smaller MNDs, baseline size (left panel) is maintained below the MND ceiling, which leaves reserve potential (red arrows) to be utilized for growth during increased workload. In these fibers, myonuclear accrual is therefore not essential for adaptive growth, which is generated, at least initially, by increasing pre-existing MND sizes and not MND numbers. (C) Titration of myonuclear numbers in mice has revealed the existence of a myonuclear transcriptional reserve capacity to support MNDs much larger than typically generated in normal adult myofibers. The magnitude of reserve capacity elicited correlates with mRNA concentrations and is inversely related to myonuclear content. It progressively diminishes as myonuclear numbers approach normal, control levels. This inverse relationship could explain limits in myonuclear biosynthetic capacities and the roles of MND volumes, ceilings, and plasticity in the generation of adaptive growth in adult myofibers.
The recent development of various exercise models that elicit physiological hypertrophy in mice [76] has finally made possible the targeted interrogation of the role of MuSCs, fusion and myonuclear accrual in adaptive growth. These models should prove superior to the supraphysiologic stimulus of synergist ablation [70, 71, 77, 78]. Subjecting mice deficient in MuSCs to their progressive weighted wheel running (PoWeR) protocol, Englund et al. [79] report blunting of both muscle hypertrophy and myofiber cross-sectional area (CSA) enlargement. Abrogation of PoWeR-induced myonuclear accrual also elicited aberrant adaptive responses with impairment in muscle function, therefore closely mirroring findings previously reported by Goh et al. [80]. When mice were subjected to a high-intensity interval treadmill protocol (HIIT) where the amount of work progressively increased during each week of the eight week regimen, muscle weights, myofiber CSA, and strength were augmented, and these enhancements were associated with ~30% increase in total myonuclear numbers [80]. While myonuclei were progressively accrued over the entire duration of the regimen, growth in CSA and mass occurred principally during the latter four-week duration of the protocol. Blocking accrual via abrogation of myoblast fusogenicity elicited exercise intolerance, loss of hypertrophy and maladaptive remodelling in hind-limb muscle. Taken together, these studies provided strong evidence of an obligatory requirement of MuSC activation, myoblast fusion, and myonuclear accrual for adaptive growth to support optimal exercise-induced functional remodeling.
It has been suggested that the myonuclear accrual typically observed during such exercise regimens is more a response to exercise-induced myofiber damage and not for growth [74]. This is indeed a possibility, and practically, it may be nearly impossible to separate exercise-induced muscle injury from increased workload when utilizing exercise models such as PoWeR or HIIT. Nevertheless, when fusogenicity was abrogated four weeks into the HIIT protocol, so that myonuclear accrual was blocked specifically during the period growth occurred but after majority of the HIIT-induced increase in myonuclear numbers (68% of total) had taken place, hypertrophy continued to be blocked [80]. This absence of growth could not be dismissed as secondary to impaired running or maladaptive remodeling, because blocking fusogenicity at 4-week time point of HIIT caused neither exercise intolerance nor detectable fibrotic remodeling. Rather, the continued absence of hypertrophy revealed that simply harboring additional nuclei in the context of an exercise stimulus does not necessarily cause adaptive growth, and that continued accrual of myonuclei is likely essential for a physiological hypertrophic response to occur.
However, the argument put forth by Murach et al. [74] coupled with findings in the HIIT study [80], where substantial myonuclear accrual occurred during the initial four-weeks of the regimen without detectable growth, highlights an important point that myonuclear accrual and accumulation may not occur exclusively for purposes of growth [81]. Amongst other things, accrual might occur as a reparative response to myofiber injury, or alternatively, myonuclei might indeed be elicited for growth, which however does not manifest until their numbers reach a threshold where the effects of injury can be circumvented, if not mitigated. Future work will likely clarify this intriguing question. However, when results from the synergistic ablation surgical model of overload, where attenuation of hypertrophy upon abrogation of myonuclear accrual has been reported by four independent groups [73, 82–84] are reviewed in conjunction with several independent human studies [85–89] it is clear that the vast majority of evidence confirms a definitive link between myonuclear accrual and adaptive growth in response to increased workload.
4. MND plasticity and growth
Does this mean that increases in MND volumes in individual myofibers can never occur during functional hypertrophy? On the contrary, much of the developmental radial growth in the mouse, accounting for a 3-fold expansion of the MND, occurs in myofibers between post-natal day 21 and adulthood after robust myonuclear accrual has ceased [90, 91]. This demonstrates that systemic growth signals can drive enlargement of MND volumes in support of functional hypertrophy. It is possible that the well documented effects of various genetic and pharmacological interventions on MND and myofiber size elicit similar responses [71, 92–96], although confirmatory mechanistic understanding remains rudimentary. However, simply the fact that MND volumes are responsive to pharmacological agents and expand without myonuclear accrual does not in itself implicate a role for accrual independent expansion in adaptive growth. So, is there any evidence that MND plasticity, in the absence of myonuclear accretion, contributes to exercise-induced growth? The answer may lie in the concept of the “myonuclear domain ceiling”, an upper threshold for MND volume, which is thought to correlate with maximal myonuclear transcriptional output; any additional enlargement of the MND beyond this ceiling or threshold is thought to require accrual of additional nuclei (Fig. 1A). This theoretical ceiling is believed to be met in the biggest myofibers, which also typically harbor the largest MND volumes as described above [14, 62] (Fig. 1A), whereas in smaller myofibers with relatively smaller MND volumes, potential for enlargement remains available without the requirement of myonuclear accretion (Fig. 1B). The existence of such myofibers with disproportionately smaller MNDs has been described primarily in the context of aging-related atrophy [75, 97], but are likely also found in normal, healthy muscle [97].
The paradigm that baseline MND volumes determine the reliance on myonuclear accrual for adaptive growth, was recently tested by Snijders et al. [98], who hypothesized that exercise training in older adults would promote a decline in the proportion of smaller myofibers principally by eliciting residual plasticity rather than myonuclear content to increase MND sizes. Their cluster analysis approach indicated that after subjecting healthy older adults to 12-weeks of resistance training, resident myonuclei were able to support hypertrophic growth in the smallest myofibers without requiring an increase in myonuclear numbers, whereas in the largest myofibers (where MND size has met the ceiling and therefore, no residual plasticity exists) growth was best predicted by an increase in myonuclear content rather than domain size. Replication of findings in younger individuals and in mouse models, where sampling is less of an issue, would help exclude the possibility that results seen in smaller fibers are not just a case of atrophied fibers in older individuals regaining previously held volumes without requiring myonuclear accrual because they already harbor adequate number of nuclei to achieve that size [99, 100]. Another obvious unknown is the reason for the existence of such smaller myofibers with MNDs, which may, as shown in Fig. 1B, be maintained below the “threshold ceiling” in normal muscle. Specifically, if potential for enlargement is available, why do such myofibers not grow until that threshold is met? Reasons could include a positional effect, where the location of such fibers within the larger muscle alleviates growth demands. Alternatively, there is the intriguing possibility that the linear relationship between myonuclear numbers and surface area serves to limit developmental growth and myofiber size [101], even when transcriptional potential to achieve larger sizes exists. Despite such unknowns, the data discussed above probably best showcase that an obligatory requirement of myonuclear accrual need not preclude a contribution, even if limited, of MND plasticity to optimal adaptive growth. Such a role for residual MND plasticity in adaptive growth is consistent with the evidence that resident myonuclei have some reserve capacity to increase transcription during workload-induced hypertrophy [102]. It would be interesting to determine what fraction of this increased transcriptional output is related to rRNA synthesis and ribosomal biogenesis, which correlate with translational capacity that is known to be augmented during both developmental and adaptive growth [103–105].
Taken together, a balanced review of our current knowledge base establishes that both an absolute requirement of myonuclear accrual for adaptive growth and MND plasticity can and do coexist; indeed, baseline MND sizes may determine both the extent of MND plasticity, and reliance on myonuclear accrual for adaptive hypertrophic growth [Figures 1A and 1B]. This reveals the lingering controversy, that has posited the existence of one to negate or disprove the other, to be more about an incorrectly framed question rather than antagonistic data; the fact that MND plasticity exists does not discount the overwhelming evidence that optimal adaptive growth in response to increased workload requires fusion-mediated myonuclear accrual. At the same time, an obligatory role for myonuclear accrual in adaptive growth does not obviate the evidence that myonuclei/MNDs in some fibers possess residual transcriptional plasticity that facilitates increases in MND volumes to support such adaptive growth. Resolution of this controversy should provide an opportunity for the field to turn its attention to more salient questions regarding regulation of MND volume, myonuclear and MND plasticity, and myofiber size establishment and adaptive remodeling.
One such fascinating question that emerges from our current knowledge base is what might determine the intrinsic transcriptional capacities of myonuclei within syncytial myofibers? Clues that nuclear numbers themselves play a key role in determining the range of transcriptional output and therefore, the potential for MND plasticity have begun to emerge from studies in mice [38, 106] and zebrafish [107]. When Cramer et al. [38] lowered myonuclear numbers during neonatal development by rendering MuSCs non-fusogenic at different time-points after birth, the resultant myofibers that had fewer resident myonuclei developed to form functionally normal, albeit smaller, muscle with enlarged MNDs [38, 62]. Similarly, when myoblast fusion was impaired in zebrafish by deletion of myogenin, the resultant mutant fish were smaller with reduced muscle content, but functionally normal and viable [108]. Despite a 90% reduction in myonuclei numbers, myogenin-null myofibers developed normal sarcomere lengths, with enlargement of MNDs [107]. While a deeper analysis of MND regulation in the zebrafish model is unavailable, the evidence of a reserve capacity to generate and sustain larger MNDs during development in two independent models highlights the likelihood that the molecular paradigms revealed in mice are evolutionarily conserved.
The myonuclear titration study carried out in mice by the Millay group [38] revealed a previously unknown correlation between MND sizes and mRNA concentrations, as assessed by the relative contribution of mRNA to total RNA content [109]. Besides providing a potentially novel metric for MND size, the same results also revealed existence of a reserve capacity in myonuclei accrued during postnatal development to support higher mRNA transcript content and larger MNDs. Notably, the magnitude of this reserve capacity elicited (mRNA concentration) was found to have an inverse relationship with the number of resident myonuclei, consistent with the evidence that it does not appear to be elicited when a full complement of myonuclei is accrued (Fig. 1C). In the mouse line named Δ2w, where myonuclei numbers were reduced by approximately half, MND volumes were increased nearly 1.5-fold with a ~4-fold increase in transcript content normalized to nuclei numbers. In contrast, in the mouse line named Δ3w, where myonuclei numbers were reduced by just 25%, the increase in relative transcript content was far more modest, with no detectable increase in MNDs. Therefore, while multinucleation is clearly essential for normal development and growth, we speculate that these results indicate there is a cost paid at the level of individual nuclei where, as their numbers increase, a progressive diminution in their ability to utilize their reserve transcriptional capacities occurs. We should stress that this does not mean that myofibers with more nuclei will have smaller MNDs compared to myofibers with fewer nuclei and, as explained above, in fact the opposite is true. As demonstrated in the scaling analysis by Hansson et al. [62], even within the Δ2w model, myofibers with higher number of nuclei had larger MNDs compared to Δ2w myofibers with fewer nuclei, indicating that general scaling trends were preserved.
The cost of multinucleation should therefore be conceived not in terms of an overall reduction in transcriptional output per se, but rather the progressive limitation of the myonuclear reserve transcriptional capacities, which ultimately defines the range of transcriptional mRNA output a resident myonucleus can support. Based on this idea, one might predict that myonuclei within smaller fibers, which typically have fewer nuclei, retain some access to their transcriptional reserve capacities; is this then the reason why, as described above [97, 98], smaller myofibers seem to harbor some residual MND plasticity that is elicited during adaptive growth? Clearly, this paradigm about how myonuclear numbers act to define their own range of transcriptional output and might therefore have a role in determining the myonuclear domain “ceiling” needs to be further developed through analysis of single myofibers, and the molecular circuitry underlying this regulation also remains to be determined. One exciting possibility is that the nuclei influence one another in a manner akin to quorum-sensing between bacteria [110] that could involve production of auto inhibitory transcripts that accumulate as myonuclei numbers increase, which once a certain threshold concentration is achieved results in feedback that limits access to reserve transcriptional capacities. Alternatively, the concentration of certain essential transcription factors might be limiting within myofibers, leading to a competitive diminution of transcriptional output as myonuclear numbers increase. Whatever the mechanism, establishment of such a range of mRNA output on a per nuclear basis during development would explain the key properties of MNDs that determine size, plasticity, and adaptive growth in myofibers.
6. Conclusion
The progress reviewed thus far bodes well for the field to make significant advances in deciphering the phenomenon and mechanisms regulating multinucleation and associated growth and maintenance of skeletal myofibers. Some specific goals that now seem achievable include deciphering how myofiber surface area scales with resident myonuclear numbers at the molecular level; identifying the ancillary factors that may cooperate with myonuclear numbers and transcriptional output to determine MND volumes; molecularly defining the decision by adapting myofibers to elicit and accrue new myonuclei; and, characterizing potential transcriptional diversity within myofibers when adapting to diverse workload demands. In addition to broadening and deepening our knowledge base, future work towards these goals will have the potential to help design novel therapeutic strategies to address challenges in the areas of aging, cachexia, and other muscle wasting pathologies.
8. Acknowledgements
We thank members of the Millay and Gundersen laboratories for critical reading of the review. We apologize to those whose work we are unable to cite due to space limitations. The work in the Millay laboratory is funded by grants to D.P.M. from the Children’s Hospital Research Foundation, Pew Charitable Trusts, and National Institutes of Health (R01AR068286, R01AG059605). V.P. is a member of the Molkentin Lab.
Abbreviations
- MuSCs
muscle stem cells
- MND
myonuclear domain
- NMJ
neuromuscular junction
- MTJ
myotendinous junction
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
7. References
- [1].Arnold EM, Delp SL, Fibre operating lengths of human lower limb muscles during walking, Philos. Trans. R. Soc. Lond. B. Biol. Sci. 366 (2011) 1530–1539. doi: 10.1098/rstb.2010.0345 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Mauro A, Satellite cell of skeletal muscle fibers, J. Biophys. Biochem. Cytol. 9 (1961) 493–495. doi: 10.1083/jcb.9.2.493 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Moss FP, Leblond CP, Satellite cells as the source of nuclei in muscles of growing rats, Anat. Rec. 170 (1971) 421–435. doi: 10.1002/ar.1091700405 [DOI] [PubMed] [Google Scholar]
- [4].Margam NN, Duncan R, Myomaker and Myomerger: It Takes Two to Make One, Dev. Cell 46 (2018) 676–678. doi: 10.1016/j.devcel.2018.09.005 [DOI] [PubMed] [Google Scholar]
- [5].Millay DP, O’Rourke JR, Sutherland LB, Bezprozvannaya S, Shelton JM, Bassel-Duby R, Olson EN, Myomaker is a membrane activator of myoblast fusion and muscle formation, Nature 499 (2013) 301–305. doi: 10.1038/nature12343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Bi P, Ramirez-Martinez A, Li H, Cannavino J, McAnally JR, Shelton JM, Sánchez-Ortiz E, Bassel-Duby R, Olson EN, Control of muscle formation by the fusogenicmicropeptidemyomixer, Science 356 (2017) 323–327. doi: 10.1126/science.aam9361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Quinn ME, Goh Q, Kurosaka M, Gamage DG, Petrany MJ, Prasad V, Millay DP, Myomerger induces fusion of non-fusogenic cells and is required for skeletal muscle development, Nat. Commun. 8 (2017) 15665. doi: 10.1038/ncomms15665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Zhang Q, Vashisht AA, O’Rourke J, Corbel SY, Moran R, Romero A, Miraglia L, Zhang J, Durrant E, Schmedt C, Sampath SC, Sampath SC, The microprotein Minion controls cell fusion and muscle formation, Nat. Commun 8 (2017) 15664. doi: 10.1038/ncomms15664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Leikina E, Gamage DG, Prasad V, Goykhberg J, Crowe M, Diao J, Kozlov MM, Chernomordik LV , Millay DP, Myomaker and Myomerger Work Independently to Control Distinct Steps of Membrane Remodeling during Myoblast Fusion, Dev. Cell 46 (2018) 767–780. doi: 10.1016/j.devcel.2018.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Gamage DG, Leikina E, Quinn ME, Ratinov A, Chernomordik LV, Millay DP, Insights into the localization and function of myomaker during myoblast fusion, J. Biol. Chem. 292 (2017) 17272–17289. doi: 10.1074/jbc.M117.811372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Sampath SC, Sampath SC, Millay DP, Myoblast fusion confusion: the resolution begins, Skelet. Muscle 8 (2018) 3. doi: 10.1186/s13395-017-0149-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Brukman NG, Uygur B, Podbilewicz B, Chernomordik LV, How cells fuse, J. Cell Biol 218 (2019) 1436–1451. doi: 10.1083/jcb.201901017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Petrany MJ, Millay DP, Cell Fusion: Merging Membranes and Making Muscle, Trends Cell Biol. 29 (2019) 964–973. doi: 10.1016/j.tcb.2019.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Bruusgaard JC, Liestøl K, Ekmark M, Kollstad K, Gundersen K, Number and spatial distribution of nuclei in the muscle fibres of normal mice studied in vivo, J. Physiol. 551 (2003) 467–478. doi: 10.1113/jphysiol.2003.045328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Cheek D, Holt A, Hill D, Talbert JL, Skeletal Muscle Cell Mass and Growth: The Concept of the Deoxyribonucleic Acid Unit, Pediatr. Res. 5 (1971) 312–328. doi: 10.1203/00006450-197107000-00004 [DOI] [Google Scholar]
- [16].Cheek DB, The control of cell mass and replication. The DNA unit--a personal 20-year study, Early Hum. Dev 12 (1985)211–239. doi: 10.1016/0378-3782(85)90144-6. [DOI] [PubMed] [Google Scholar]
- [17].Allen DL, Roy RR.Edgerton VR, Myonuclear domains in muscle adaptation and disease, Muscle Nerve 22 (1999) 1350–1360. doi: . [DOI] [PubMed] [Google Scholar]
- [18].Tseng BS, Kasper CE, Edgerton VR, Cytoplasm-to-myonucleus ratios and succinate dehydrogenase activities in adult rat slow and fast muscle fibers, Cell Tissue Res. 275 (1994) 39–49. doi: 10.1007/BF00305374. [DOI] [PubMed] [Google Scholar]
- [19].Cristea A, Qaisar R, Edlund PK, Lindblad J, Bengtsson E, Larsson L, Effects of aging and gender on the spatial organization of nuclei in single human skeletal muscle cells, Aging Cell 9 (2010) 685–697. doi: 10.1111/j.1474-9726.2010.00594.x. [DOI] [PubMed] [Google Scholar]
- [20].Lin W, Burgess RW, Dominguez B, Pfaff SL, Sanes JR, Lee KF, Distinct roles of nerve and muscle in postsynaptic differentiation of the neuromuscular synapse, Nature 410 (2001) 1057–1064. doi: 10.1038/35074025. [DOI] [PubMed] [Google Scholar]
- [21].Sanes JR, Lichtman JW, Induction, assembly, maturation and maintenance of a postsynaptic apparatus, Nat. Rev. Neurosci. 2 (2001) 791–805. doi: 10.1038/35097557. [DOI] [PubMed] [Google Scholar]
- [22].Li L, Xiong WC, Mei L, Neuromuscular junction formation, aging, and disorders, Annu. Rev. Physiol. 80 (2018) 159–188. doi: 10.1146/annurev-physiol-022516-034255. [DOI] [PubMed] [Google Scholar]
- [23].Wilson EB, The cell in development and heredity, MacMillan, New York, 1926, pp. 916–923. [Google Scholar]
- [24].Amodeo AA, Skotheim JM, Cell-size control, Cold Spring Harb. Perspect. Biol. 8 (2016) doi: 10.1101/cshperspect.a019083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Mortimer RK, Radiobiological and Genetic Studies on a Polyploid Series (Haploid to Hexaploid) of Saccharomyces cerevisiae, Rad. Res. 9 (1958) 312–326. [PubMed] [Google Scholar]
- [26].Tautvydas KJ, Evidence for chromosome endoreduplication in Eudorina californica, a colonial alga, Differentiation 5 (1976) 35–42. doi: 10.1111/j.1432-0436.1976.tb00889.x. [DOI] [PubMed] [Google Scholar]
- [27].Zimmet J, Ravid K, Polyploidy: occurrence in nature, mechanisms, and significance for the megakaryocyte-platelet system, Exp. Hematol. 28 (2000) doi: 10.1016/s0301-472x(99)00124-1. [DOI] [PubMed] [Google Scholar]
- [28].Ullah Z, Lee CY, Lilly MA, DePamphilis ML, Developmental^ programmed endoreduplication in animals, Cell Cycle 8 (2009) 1501–1509. doi: 10.4161/cc.8.10.8325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Gillooly JF, Hein A, Damiani R, Nuclear DNA Content Varies with Cell Size across Human Cell Types, Cold Spring Harb. Perspect. Biol 7 (2015) a019091. doi: 10.1101/cshperspect.aOl9091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Rota M, Hosoda T, De Angelis A, Arcarese ML, Esposito G, Rizzi R, Tillmanns J, Tugal D, Musso E, Rimoldi O, Bearzi C, Urbanek K, Anversa P, Leri A, Kajstura J, The young mouse heart is composed of myocytes heterogeneous in age and function, Circ. Res. 101 (2007) 387–399. doi: 10.1161/CIRCRESAHA.107.151449. [DOI] [PubMed] [Google Scholar]
- [31].Verdijk LB, Snijders T, Drost M, Delhaas T, Kadi F, van Loon LJC, Satellite cells in human skeletal muscle; from birth to old age, Age 36 (2014) 545–547. doi: 10.1007/s11357-013-9583-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Vassilopoulos D, Lumb EM, Emery AE, Karyometric changes in human muscle with age, Eur. Neurol. 16 (1977) 31–34. doi: 10.1159/000114877. [DOI] [PubMed] [Google Scholar]
- [33].Oertel G, Morphometric analysis of normal skeletal muscles in infancy, childhood and adolescence. An autopsy study, J. Neurol. Sci. 88 (1988) 303–313. doi: 10.1016/0022-510x(88)90227-4. [DOI] [PubMed] [Google Scholar]
- [34].Demontis F, Perrimon N, Integration of Insulin receptor/Foxo signaling and dMyc activity during muscle growth regulates body size in Drosophila, Development 136 (2009) 983–993. doi: 10.1242/dev.027466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Windner SE, Manhart A, Brown A, Mogilner A, Baylies MK, Nuclear Scaling Is Coordinated among Individual Nuclei in Multinucleated Muscle Fibers, Dev. Cell 49 (2019) 48–62. doi: 10.1016/j.devcel.2019.02.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Moss FP, Leblond CP, Nature of dividing nuclei in skeletal muscle of growing rats, J. Cell Biol. 44 (1970) 459–462. doi: 10.1083/jcb.44.2.459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Stockdale FE, Holtzer H, DNA synthesis and myogenesis, Exp. Cell Res. 24 (1961) 508–520. doi: 10.1016/0014-4827(61)90450-5. [DOI] [PubMed] [Google Scholar]
- [38].Cramer AAW, Prasad V, Eftestøl E, Song T, Hansson KA, Dugdale HF, Sadayappan S, Ochala J, Gundersen K, Millay DP, Nuclear numbers in syncytial muscle fibers promote size but limit the development of larger myonuclear domains, Nat. Commun. 11 (2020) 6287. doi: 10.1038/s41467-020-20058-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Bachman JF, Klose A, Liu W, Paris ND, Blanc RS, Schmalz M, Knapp E, Chakkalakal JV, Prepubertal skeletal muscle growth requires Pax7-expressing satellite cell-derived myonuclear contribution, Development 145 (2018) doi: 10.1242/dev.167197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Gladfelter AS, Hungerbuehler AK, Philippsen P, Asynchronous nuclear division cycles in multinucleated cells, J. Cell Biol. 172 (2006) 347–362. doi: 10.1083/jcb.200507003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Nair DR, D’Ausilio CA, Occhipinti P, Borsuk ME, Gladfelter AS, A conserved G1 regulatory circuit promotes asynchronous behavior of nuclei sharing a common cytoplasm, Cell Cycle (2010) 3771–3779. doi: 10.4161/cc.9.18.12999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Dos Santos M, Backer S, Saintpierre B, Izac B, Andrieu M, Letourneur F, Relaix F, Sotiropoulos A, Maire P, Single-nucleus RNA-seq and FISH identify coordinated transcriptional activity in mammalian myofibers, Nat. Commun 11 (2020) 5102. doi: 10.1038/s41467-020-18789-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Petrany MJ, Swoboda CO, Sun C, Chetal K, Chen X, Weirauch MT , Salomonis N, Millay DP, Single-nucleus RNA-seq identifies transcriptional heterogeneity in multinucleated skeletal myofibers, Nat. Commun 11 (2020) 6374. doi: 10.1038/s41467-020-20063-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Kim M, Franke V, Brandt B, Lowenstein ED, Schöwel V, Spuler S, Akalin A, Birchmeier C, Single-nucleus transcriptomics reveals functional compartmentalization in syncytial skeletal muscle cells, Nat. Commun 11 (2020) 6375. doi: 10.1038/s41467-020-20064-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Newlands S, Levitt LK, Robinson CS, Karpf AB, Hodgson VR, Wade RP, Hardeman EC, Transcription occurs in pulses in muscle fibers, Genes Dev. 12 (1998) 2748–58. doi: 10.1101/gad.12.17.2748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Liu W, Klose A, Forman S, Paris ND, Wei-LaPierre L, Cortés-Lopéz M, Tan A, Flaherty M, Miura P, Dirksen RT, Chakkalakal JV, Loss of adult skeletal muscle stem cells drives age-related neuromuscular junction degeneration, Elife 6 (2017) e26464. doi: 10.7554/eLife.26464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Liu W, Wei-LaPierre L, Klose A, Dirksen RT, Chakkalakal JV, Inducible depletion of adult skeletal muscle stem cells impairs the regeneration of neuromuscular junctions, Elife 4 (2015) e09221. doi: 10.7554/eLife.09221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Ulintz PJ, Larouche J, Mohiuddin M, Macias JC, Kurpiers SJ, Liu W, Choi JJ, Brown LA, Markworth JF, de Silva K, Levi BD, Merajver SD, Chakkalakal JV, Jang YC, Brooks SV, Aguilar CA, Single Cell Deconstruction of Muscle Stem Cell Heterogeneity During Aging Reveals Sensitivity to the Neuromuscular Junction, bioRxiv 2020.05.28.121426; doi: 10.1101/2020.05.28.121426 [DOI] [Google Scholar]
- [49].Yaseen-Badarneh W, Kraft-Sheleg O, Zaffryar-Eilot S, Melamed S, Sun C, Millay DP, Hasson P, Fibroblast fusion to the muscle fiber regulates myotendinous junction formation, bioRxiv 2020.07.20.213199; doi: 10.1101/2020.07.20.213199 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].de Lima JE, Blavet C, Bonnin M-A, Hirsinger E, Comai G, Yvernogeau L, Bellenger L, Mella S, Nassari S, Robin C, Schweitzer R, Fournier-Thibault C, Tajbakhsh S, Relaix F, Duprez D, BMP signalling directs a fibroblast-to-myoblast conversion at the connective tissue/muscle interface to pattern limb muscles, bioRxiv 2020.07.20.211342; doi: 10.1101/2020.07.20.211342 [DOI] [Google Scholar]
- [51].Denes LT, Kelley CP, Wang ET, Microtubule-based Transport is Essential to Distribute RNA and Nascent Protein in Skeletal Muscle, bioRxiv 2021.02.26.433059; doi: 10.1101/2021.02.26.433059 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Scarborough EA, Uchida K, Vogel M, Erlitzki N, Iyer M, Phyo SA, Bogush A, Kehat I, Prosser BL, Microtubules orchestrate local translation to enable cardiac growth, Nat. Commun 12 (2021) 1547. doi: 10.1038/s41467-021-21685-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [53].Merlie JP, Sanes JR, Concentration of acetylcholine receptor mRNA in synaptic regions of adult muscle fibres, Nature 317 (1985) 66–68. doi: 10.1038/317066a0. [DOI] [PubMed] [Google Scholar]
- [54].Pavlath GK, Rich K, Webster SG, Blau HM, Localization of muscle gene products in nuclear domains, Nature 337 (1989) 570–573. doi: 10.1038/337570a0. [DOI] [PubMed] [Google Scholar]
- [55].Ralston E, Hall ZW, Restricted distribution of mRNA produced from a single nucleus in hybrid myotubes, J. Cell Biol. 119 (1992) 1063–1068. doi: 10.1083/jcb.119.5.1063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Hansson KA, Solbrå AV, Gundersen K, Bruusgaard JC, Computational Assessment of Transport Distances in Living Skeletal Muscle Fibers Studied In Situ, Biophys. J. 119 (2020) 2166–2178. doi: 10.1016/j.bpj.2020.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Metzger T, Gache V, Xu M, Cadot B, Folker ES, Richardson BE, Gomes ER, Baylies MK, MAP and kinesin-dependent nuclear positioning is required for skeletal muscle function, Nature 484 (2012) 120–124. doi: 10.1038/nature10914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Folker ES, Baylies MK, Nuclear positioning in muscle development and disease, Front. Physiol 4 (2013) 363. doi: 10.3389/fphys.2013.00363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Roman W, Gomes ER, Nuclear positioning in skeletal muscle, Semin. Cell Dev. Biol. 82 (2018) 51–56. doi: 10.1016/j.semcdb.2017.11.005. [DOI] [PubMed] [Google Scholar]
- [60].Dundon SER, Chang S-S, Kumar A, Occhipinti P, Shroff H, Roper M, Gladfelter AS, Clustered nuclei maintain autonomy and nucleocytoplasmic ratio control in a syncytium, Mol. Biol. Cell 27 (2016) 2000–2007. doi: 10.1091/mbc.E16-02-0129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [61].Liu J-X, Höglund A-S, Karlsson P, Lindblad J, Qaisar R, Aare S, Bengtsson E, Larsson L, Myonuclear domain size and myosin isoform expression in muscle fibres from mammals representing a 100,000-fold difference in body size, Exp. Physiol. 94 (2009) 117–129. doi: 10.1113/expphysiol.2008.043877. [DOI] [PubMed] [Google Scholar]
- [62].Hansson KA, Eftestøl E, Bruusgaard JC, Juvkam I, Cramer AW, Malthe-Sørenssen A, Millay DP, Gundersen K, Myonuclear content regulates cell size with similar scaling properties in mice and humans, Nat. Commun 11 (2020) 6288. doi: 10.1038/s41467-020-20057-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Johnston IA, Abercromby M, Vieira VLA, Sigursteindóttir RJ, Kristjánsson BK, Sibthorpe D, Skúlason S, Rapid evolution of muscle fibre number in post-glacial populations of Arctic charr Salvelinus alpines, J. Exp. Biol. 207 (2004) 4343–4360. doi: 10.1242/jeb.01292. [DOI] [PubMed] [Google Scholar]
- [64].Johnston IA, de la Serrana DG., Devlin RH, Muscle fibre size optimisation provides flexibility for energy budgeting in calorie-restricted coho salmon transgenic for growth hormone, J. Exp. Biol. 217 (2014) 3392–3395. doi: 10.1242/jeb.107664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Jimenez AG, Dillaman RM, Kinsey ST, Large fibre size in skeletal muscle is metabolically advantageous, Nat. Commun. 4 (2013) 2150. doi: 10.1038/ncomms3150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Asano Y, Liberman UA, Edelman IS, Thyroid thermogenesis. Relationships between Na+-dependent respiration and Na+ + K+-adenosine triphosphatase activity in rat skeletal muscle, J. Clin. Invest. 57 (1976) 368–379. doi: 10.1172/JCI108288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].van Wessel T, de Haan A, van der Laarse WJ, Jaspers RT, The muscle fiber type-fiber size paradox: hypertrophy or oxidative metabolism? Eur. J. Appl. Physiol. 110 (2010) 665–694. doi: 10.1007/s00421-010-1545-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Kinsey ST, Hardy KM, Locke BR, The long and winding road: influences of intracellular metabolite diffusion on cellular organization and metabolism in skeletal muscle, J. Exp. Biol. 210 (2007) 3505–3512. doi: 10.1242/jeb.000331. [DOI] [PubMed] [Google Scholar]
- [69].Rome LC, Lindstedt SL, The Quest for Speed: Muscles Built for High-Frequency Contractions, News Physiol. Sci. 13 (1998) 261–268. doi: 10.1152/physiologyonline. [DOI] [PubMed] [Google Scholar]
- [70].O’Connor RS, Pavlath GK, Satellite Cell addition is obligatory for skeletal muscle hypertrophy, J. Appl. Physiol. 103 (2007) 1099–1100. doi: 10.1152/japplphysiol.00101.2007. [DOI] [PubMed] [Google Scholar]
- [71].McCarthy JJ, Esser KA, Satellite cell addition is not obligatory for skeletal muscle hypertrophy, J. Appl. Physiol. 103 (2007) 1100–1102 doi: 10.1152/japplphysiol.00101.2007a. [DOI] [PubMed] [Google Scholar]
- [72].McCarthy JJ, Mula J, Miyazaki M, Erfani R, Garrison K, Farooqui AB, Srikuea R, Lawson BA, Grimes B, Keller C, Zant GV, Campbell KS, Esser KA, Dupont-Versteegden EE, Peterson CA, Effective fiber hypertrophy in satellite cell-depleted skeletal muscle, Development 138 (2011) 3657–3666. doi: 10.1242/dev.068858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].Egner IM, Bruusgaard JC, Gundersen K, Satellite cell depletion prevents fiber hypertrophy in skeletal muscle, Development 143 (2016) 2898–2906. doi: 10.1242/dev.134411. [DOI] [PubMed] [Google Scholar]
- [74].Murach KA, Englund DA, Dupont-Versteegden EE, McCarthy JJ., Peterson CA, Myonuclear Domain Flexibility Challenges Rigid Assumptions on Satellite Cell Contribution to Skeletal Muscle Fiber Hypertrophy, Front. Physiol 9 (2018) 635. doi: 10.3389/fphys.2018.00635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].Brack AS, Bildsoe H, Hughes SM, Evidence that satellite cell decrement contributes to preferential decline in nuclear number from large fibres during murine age-related muscle atrophy, J. Cell Sci. 118 (2005) 4813–4821. doi: 10.1242/jcs.02602. [DOI] [PubMed] [Google Scholar]
- [76].Murach KA, McCarthy JJ, Peterson CA, Dungan CM, Making Mice Mighty: recent advances in translational models of load-induced muscle hypertrophy, J. Appl. Physiol. 129 (2020) 516–521. doi: 10.1152/japplphysiol.00319.2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].McCarthy JJ, Dupont-Versteegden EE, Fry CS, Murach KA, Peterson CA, Methodological issues limit interpretation of negative effects of satellite cell depletion on adult muscle hypertrophy, Development 144 (2017) 1363–1365. doi: 10.1242/dev.145797. [DOI] [PubMed] [Google Scholar]
- [78].Egner IM, Bruusgaard JC, Gundersen K, An apparent lack of effect of satellite cell depletion on hypertrophy could be due to methodological limitations, Development 144 (2017) 1365–1367. doi: 10.1242/dev.148163. [DOI] [PubMed] [Google Scholar]
- [79].Englund DA, Figueiredo VC, Dungan CM, Murach KA, Peck BD, Petrosino JM, Brightwell CR, Dupont AM, Neal AC, Fry CS, Accornero F, McCarthy JJ, Peterson CA, Satellite Cell Depletion Disrupts Transcriptional Coordination and Muscle Adaptation to Exercise, Function 2 (2021) zqaa033. doi: 10.1093/function/zqaa033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [80].Goh Q, Song T, Petrany MJ, Cramer AAW, Sun C, Sadayappan S, Lee S-J, Millay DP, Myonuclear accretion is a determinant of exercise-induced remodeling in skeletal muscle, Elife 8 (2019) e44876. doi: 10.7554/eLife.44876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Masschelein E, D’Hulst G, Zvick J, Hinte L, Soro-Arnaiz I, Gorski T, von Meyenn F, Bar-Nur O, De Bock K, Exercise promotes satellite cell contribution to myofibers in a load-dependent manner, Skelet. Muscle 10 (2020) 21. doi: 10.1186/s13395-020-00237-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [82].Fry CS, Lee JD, Jackson JR, Kirby TJ, Stasko SA, Liu H, Dupont-Versteegden EE, McCarthy JJ, Peterson CA, Regulation of the muscle fiber microenvironment by activated satellite cells during hypertrophy, FASEB J. 28 (2014) 1654–1665. doi: 10.1096/fj.13-239426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Goh Q, Millay DP, Requirement of myomaker-mediated stem cell fusion for skeletal muscle hypertrophy, Elife 10 (2017) e20007. doi: 10.7554/eLife.20007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [84].Randrianarison-Huetz V, Papaefthymiou A, Herledan G, Noviello C, Faradova U, Collard L, Pincini A, Schol E, Decaux JF, Maire P, Vassilopoulos S, Sotiropoulos A, Srf controls satellite cell fusion through the maintenance of actin architecture, J. Cell Biol. 217 (2018) 685–700. doi: 10.1083/jcb.201705130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [85].Kadi F, Thornell LE, Concomitant increases in myonuclear and satellite cell content in female trapezius muscle following strength training, Histochem. Cell Biol. 113 (2000) 99–103. doi: 10.1007/s004180050012. [DOI] [PubMed] [Google Scholar]
- [86].Olsen S, Aagaard P, Kadi F, Tufekovic G, Verney J, Olesen JL, Suetta C, Kjaer M, Creatine supplementation augments the increase in satellite cell and myonuclei number in human skeletal muscle induced by strength training, J. Physiol. 573 (2006) 525–534. doi: 10.1113/jphysiol.2006.107359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [87].Petrella JK, Kim J-S, Cross JM, Kosek DJ, Bamman MM, Efficacy of myonuclear addition may explain differential myofiber growth among resistance-trained young and older men and women, Am. J. Physiol. Endocrinol. Metab 291 (2006) E937–46. doi: 10.1152/ajpendo.00190.2006. [DOI] [PubMed] [Google Scholar]
- [88].Petrella JK, Kim J-S, Mayhew DL, Cross JM, Bamman MM, Potent myofiber hypertrophy during resistance training in humans is associated with satellite cell-mediated myonuclear addition: a cluster analysis, J. Appl. Physiol. 104 (2008) 1736–1742. doi: 10.1152/japplphysiol.01215.2007. [DOI] [PubMed] [Google Scholar]
- [89].Lundberg TR, Martínez-Aranda LM, Sanz G, Hansson B, von Walden F, Tesch PA, Fernandez-Gonzalo R, Early accentuated muscle hypertrophy is strongly associated with myonuclear accretion, Am. J. Physiol. Regul. Integr. Comp. Physiol 319 (2020) R50–R58. doi: 10.1152/ajpregu.00061.2020. [DOI] [PubMed] [Google Scholar]
- [90].White RB, Biérinx A-S, Gnocchi VF, Zammit PS, Dynamics of muscle fibre growth during postnatal mouse development, BMC Dev. Biol. 10 (2010) 21. doi: 10.1186/1471-213X-10-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [91].Davis TA, Fiorotto ML, Regulation of muscle growth in neonates, Curr. Opin. Clin. Nutr. Metab. Care 12 (2009) 78–85. doi: 10.1097/MCO.0b013e32831cef9f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [92].Beermann DH, Butler WR, Hogue DE, Fishell VK, Dalrymple RH, Ricks CA, Scanes CG, Cimaterol-induced muscle hypertrophy and altered endocrine status in lambs, J. Anim. Sci. 65 (1987) 1514–1524. doi: 10.2527/jas1987.6561514x. [DOI] [PubMed] [Google Scholar]
- [93].Amthor H., Otto A, Vulin A, Rochat A, Dumonceaux J, Garcia L, Mouisel E, Hourdé C, Macharia R, Friedrichs M, Relaix F, Zammit PS, Matsakas A, Patel K, Partridge T, Muscle hypertrophy driven by myostatin blockade does not require stem/precursor-cell activity, Proc. Natl. Acad. Sci. U S A 106 (2009) 7479–7484. doi: 10.1073/pnas.0811129106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [94].Blaauw B., Canato M, Agatea L, Toniolo L, Mammucari C, Masiero E, Abraham R, Sandri M, Schiaffino S, Reggiani C, Inducible activation of Akt increases skeletal muscle mass and force without satellite cell activation, FASEB J. 23 (2009) 3896–3905. doi: 10.1096/fj.09-131870. [DOI] [PubMed] [Google Scholar]
- [95].Raffaello A., Milan G, Masiero E, Carnio S, Lee D, Lanfranchi G, Goldberg AL, Sandri M, JunB transcription factor maintains skeletal muscle mass and promotes hypertrophy, J. Cell Biol. 191 (2010) 101–113. doi: 10.1083/jcb.201001136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [96].Englund DA, Peck BD, Murach KA, Neal AC, Caldwell HA, McCarthy JJ, Peterson CA, Dupont-Versteegden EE, Resident muscle stem cells are not required for testosterone-induced skeletal muscle hypertrophy, Am. J. Physiol. Cell Physiol. 317 (2019) C719–C724. doi: 10.1152/ajpcell.00260.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [97].Karlsen A, Couppé C, Andersen JL, Mikkelsen UR, Nielsen RH, Magnusson SP, Kjaer M, Mackey AL, Matters of fiber size and myonuclear domain: Does size matter more than age? Muscle Nerve 52 (2015) 1040–1046. doi: 10.1002/mus.24669. [DOI] [PubMed] [Google Scholar]
- [98].Snijders T, Holwerda AM, van Loon LJC, Verdijk LB, Myonuclear content and domain size in small versus larger muscle fibres in response to 12 weeks of resistance exercise training in older adults, Acta Physiol. (2020) e13599. doi: 10.1111/apha.13599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [99].Bruusgaard JC, Egner IM, Larsen TK, Dupre-Aucouturier S, Desplanches D, Gundersen K, No change in myonuclear number during muscle unloading and reloading, J. Appl. Physiol. 113 (2012) 290–296. doi: 10.1152/japplphysiol.00436.2012. [DOI] [PubMed] [Google Scholar]
- [100].Gundersen K, Muscle memory and a new cellular model for muscle atrophy and hypertrophy, J. Exp. Biol 219 (2016) 235–242. doi: 10.1242/jeb.124495. [DOI] [PubMed] [Google Scholar]
- [101].Karlsen A, Bechshøft RL, Malmgaard-Clausen NM, Andersen JL, Schjerling P, Kjaer M, Mackey AL, Lack of muscle fibre hypertrophy, myonuclear addition, and satellite cell pool expansion with resistance training in 83–94-year-old men and women, Acta Physiol. 227 (2019) e13271. doi: 10.1111/apha.13271. [DOI] [PubMed] [Google Scholar]
- [102].Kirby TJ, Patel RM, McClintock TS, Dupont-Versteegden EE, Peterson CA, McCarthy JJ, Myonuclear transcription is responsive to mechanical load and DNA content but uncoupled from cell size during hypertrophy, Mol. Biol. Cell 27 (2016) 788–798. doi: 10.1091/mbc.E15-08-0585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [103].Nader GA, Hornberger TA, Esser KA, Translational control: implications for skeletal muscle hypertrophy, Clin. Orthop. Relat. Res. 403 (2002) S178–87. doi: 10.1097/00003086-200210001-00021. [DOI] [PubMed] [Google Scholar]
- [104].Figueiredo VC, McCarthy JJ, Regulation of Ribosome Biogenesis in Skeletal Muscle Hypertrophy, Physiology 34 (2019) 30–42. doi: 10.1152/physiol.00034.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [105].Figueiredo VC, Revisiting the roles of protein synthesis during skeletal muscle hypertrophy induced by exercise, Am. J. Physiol. Regul. Integr. Comp. Physiol 317 (2019) R709–R718. doi: 10.1152/ajpregu.00162.2019. [DOI] [PubMed] [Google Scholar]
- [106].Nikolaou S, Cramer AAW, Hu L, Goh Q, Millay DP, Cornwall R, Proteasome inhibition preserves longitudinal growth of denervated muscle and prevents neonatal neuromuscular contractures, JCI Insight, 4 (2019) e128454. doi: 10.1172/jci.insight.128454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [107].Ganassi M, Badodi S, Wanders K, Zammit PS, Hughes SM, Myogenin is an essential regulator of adult myofibre growth and muscle stem cell homeostasis, Elife 9 (2020) e60445. doi: 10.7554/eLife.60445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [108].Ganassi M, Badodi S, Quiroga HPO, Zammit PS, Hinits Y, Hughes SM, Myogenin promotes myocyte fusion to balance fibre number and size, Nat. Commun. 9 (2018) 4232. doi: 10.1038/s41467-018-06583-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [109].Zhurinsky J, Leonhard K, Watt S, Marguerat S, Bähler J, Nurse P, A coordinated global control over cellular transcription, Curr. Biol. 20 (2010) 2010–2015. doi: 10.1016/j.cub.2010.10.002. [DOI] [PubMed] [Google Scholar]
- [110].Miller MB, Bassler BL, Quorum sensing in bacteria, Annu. Rev. Microbiol. 55 (2001) 165–199. doi: 10.1146/annurev.micro.55.1.165. [DOI] [PubMed] [Google Scholar]