

Abstract

DNA origami nanoframes with two parallel DNA sequences are used to evaluate the effect of nucleoside substituents on radiation-induced DNA damage. Double strand breaks (DSB) of DNA are counted using atomic force microscopy (AFM), and total number of lesions is evaluated using real-time polymerase chain reaction (RT-PCR). Enhanced AT or GC content does not increase the number of DNA strand breaks. Incorporation of 8-bromoadenosine results in the highest enhancement in total number of lesions; however, the highest enhancement in DSB is observed for 2′-deoxy-2′-fluorocytidine, indicating different mechanisms of radiosensitization by nucleoside analogues with the halogen substituent on base or sugar moieties, respectively. “Bystander” effects are observed, when the number of DSB in a sequence is enhanced by a substituent in the parallel DNA sequence. The present approach eliminates limitations of previously developed methods and motivates detailed studies of poorly understood conformation or bystander effects in radiation induced damage to DNA.
Understanding radiation damage to DNA is crucial for cancer radiotherapy. Irreparable damage to DNA suppresses cell proliferation, thereby exploring its mechanisms is important to the improvement of radio-therapeutic techniques.1,2 Several tools have been utilized to study radiation damage to DNA in solution, most of them relying on plasmid DNA and gel electrophoresis.3,4 These can provide quantitative data on DNA damage by ionizing radiation, although they are often influenced by the chosen analysis approach.5,6 Plasmid DNA-based systems have been useful in fundamental studies, for example, in disentangling direct and indirect effects of radiation on DNA damage3,7,8 or in studies identifying sources of synergism in chemo-radiation therapy.9−11 In certain cases, however, plasmid DNA and the necessity for stabilizing buffer solutions create a complex system that could be difficult to link to fundamental studies performed on isolated DNA building blocks.12,13 Therefore, there is also rising interest in studies of short DNA segments, incorporated into DNA strands14 or placed in a well-defined environment.15−17
DNA origami nanostructures are promising tools to study radiation damage to DNA sequences and radiosensitivity modification by sensitizers such as nucleotides functionalized with electron affinic moieties. The hundreds of synthetic oligomers that compose the origami nanostructure allow for hundreds of unique binding sites that can be extended to support a wide array of probes and active agents at fixed nanometric spaces.18 A common DNA origami-based technique used to study DNA single strand breaks utilizes DNA origami nanotriangles deposited on Si/SiO2 substrates with atomic force microscopy (AFM) as the primary characterization technique to quantify strand breaks in singulo.19 Biotinylated DNA oligomers containing sequences of interest are attached to several locations on the origami nanostructure, free-standing above the surface, which can be irradiated in vacuum either by vacuum ultraviolet (VUV) or low energy electrons (LEE). The intact strands remaining after irradiation are amplified by the addition of streptavidin for subsequent visualization by AFM.19 The technique has been tested on various oligomer sequences with and without added radiosensitizers as well as biologically relevant secondary structures like G-quadruplexes generating values for strand break cross sections to represent the effects of secondary electrons.20−22 These studies are so far limited to irradiation in vacuum. Extending these studies to systems in aqueous solution as demonstrated in the present study would be beneficial in order to determine the respective contributions of direct effects deduced from said studies and those of indirect effects caused by reactive radiolysis products like OH radicals in environments close to that experienced by DNA in cellular environments. Some exploratory studies on bare origami nanostructures were also performed, indicating another possible approach to study DNA damage in the aqueous environment using DNA origami templates. One of these made use of C60 in solution to generate 1O2 reactive species upon photoexcitation, which then react with DNA origami23 as observed through AFM. In another study, solutions containing DNA origami were irradiated by high doses of γ-rays and proton beams, demonstrating enhanced DNA origami stability compared to similarly sized plasmids, making it a suitable substrate for radiation damage studies.24
The common use of DNA origami as a substrate with free-standing sequences of interest is limited to sequences of only several nucleotides due to the tendencies of longer sequences (∼20 nucleotides) to fold.26 Here we exploit an alternative method using DNA origami nanoframes, which enables studies of longer double stranded sequences (Figure 1A). The nanoframes are based on the design of the Sugiyama group,27 where it was mostly used for state of the art mechanistic studies on various DNA sequences.28,29 The frames are ∼80 × 90 nm in dimension and are constructed from an M13mp18 single-stranded DNA scaffold and 222 strategically designed staple oligomer sequences that tie the structure together. We had to trim the blunt ends by the removal of 31 staples on the sides to prevent agglomeration during deposition on the surface as a consequence of π–π stacking in between the end bases. The frames can support a couple of DNA motifs in their inner aperture, with at least 64 base-pairs in addition to 16 nucleotides that anchor to both sides of the origami frame (Figure 1B). In this design, two duplexes/motifs can be studied at the same time, allowing for direct comparison of damage to the strands, for example, in between two different DNA sequences or in between plain DNA and its sensitized form. We irradiated the samples in solution using 16 MeV electrons, deposited the treated solutions on Si substrates, and then visualized directly the strand breaks by AFM. The AFM count is limited to double strand breaks caused both by direct hits as well as double strand breaks from multiple SSBs on both strands of the DNA motif (clustered damages). The rest of the sample was then analyzed using a real-time polymerase chain reaction (RT-PCR), which was used as a complementary analytical technique to confirm dose-dependent damage to the DNA motifs. The latter can detect the relative amount of remaining, intact DNA motifs after irradiation; however, it cannot differentiate between the different types of lesions.
Figure 1.

Scheme of the methods employed (A) and the various sequences studied, their positions in the origami nanostructure (image rendered in oxView),25 and the chemical structures of the radiosensitizers used (B).
In this work, we investigated four systems, two of which deal with the effects of the AT and GC contents of the strands (referred to here as A10 and G10 systems); the other two concern the effects of the inclusion of model radiosensitizing nucleosides (referred to here as 2FC and BrA systems) (Figure 1B). The A10 system contains a higher AT content with A10 and complementary T10 fixed within the center of the DNA duplex in the top position (Position 1). The bottom position (Position 2) supports a control strand comprised of a scrambled DNA sequence (R(A10)). In the G10 system, the A10:T10 section of the top strand is replaced by a G10:C10 sequence. For the radiosensitized systems, the 64 bp free central sequence of the 2FC strand is similar to that of the R strand but with three cytidine nucleosides replaced by 2′-deoxy-2′-fluorocytidine, while in the BrA system, three adenosine nuleosides are replaced by 8-bromoadenosine (Figure 1B). Both are representative models for two types of electrophilic radiosensitizers, with halogen atoms incorporated in the sugar unit30,31 and the base unit32,33 of the nucleosides, respectively. In both systems, the R sequence is retained in Position 2 serving as a control.
The breaks in the parallel duplex strands in the
frame aperture
can be distinguished and counted by AFM. The counts for each system
at increasing absorbed dose are shown in Figure 2e–h. The dose response of DSB counted
per DNA origami frame (NDSB, with initial DSB count 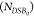 in the
unirradiated sample) can be described
by an exponential with a saturation point (eq 1) and a decay rate corresponding to the double
strand break yield (μDSB) due to
the observed stochastic nature of the process.34 The working range 0–4 kGy (see Supporting Information) before the frames become heavily damaged
is still within the quasi-linear regime estimated by fitting a linear
function whose slope is equal to μDSB (eq 2).
in the
unirradiated sample) can be described
by an exponential with a saturation point (eq 1) and a decay rate corresponding to the double
strand break yield (μDSB) due to
the observed stochastic nature of the process.34 The working range 0–4 kGy (see Supporting Information) before the frames become heavily damaged
is still within the quasi-linear regime estimated by fitting a linear
function whose slope is equal to μDSB (eq 2).
| 1 |
| 2 |
Figure 2.

Dose response of Cp values (Mean ± SD, n = 3) from RT-PCR analysis (a–d) and AFM counts of % DSB (Mean ± SD, n = 4–7) vs absorbed dose curves (e–h) of the various DNA motifs studied: A10, G10, 2FC, and BrA (blue circles) and their accompanying control (R) strand (orange squares). Linear fits were performed on each curve based on eqs 3 and 2 for data obtained from RT-PCR and AFM, respectively. Uncertainties in absorbed doses are within 10% as estimated from the measurements of dose rates on the irradiation platform.
In the case of RT-PCR,
the fraction of remaining intact strands 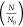 after irradiation is related to the measured Cp (crossing point or threshold
value of fluorescence detection) as described in eq 3, taking into account the primer efficiency
(E) obtained from standard curves (see Supporting Information), with
after irradiation is related to the measured Cp (crossing point or threshold
value of fluorescence detection) as described in eq 3, taking into account the primer efficiency
(E) obtained from standard curves (see Supporting Information), with  taken
as the threshold value of the control
containing N0 strands.35,36 The measured curves are shown in Figure 2a–d. During irradiation, the strands
can be nicked, and the remaining fraction of intact strands can be
related to the absorbed dose (D) by an exponential
decay function (eq 4).
The decay rate (μTL) estimates the
number of lesions per absorbed dose assuming a Poisson distribution
of nicking events. The lesions include all double strand breaks and
all damages to the modified strands that prevent PCR amplification
such as single-strand breaks and base damages. Combining both equations
relates the RT-PCR raw measurements in terms of Cp and the absorbed dose to directly obtain
the number of lesions from the slope of the linear fit of the Cp vs D curves
(eq 5).
taken
as the threshold value of the control
containing N0 strands.35,36 The measured curves are shown in Figure 2a–d. During irradiation, the strands
can be nicked, and the remaining fraction of intact strands can be
related to the absorbed dose (D) by an exponential
decay function (eq 4).
The decay rate (μTL) estimates the
number of lesions per absorbed dose assuming a Poisson distribution
of nicking events. The lesions include all double strand breaks and
all damages to the modified strands that prevent PCR amplification
such as single-strand breaks and base damages. Combining both equations
relates the RT-PCR raw measurements in terms of Cp and the absorbed dose to directly obtain
the number of lesions from the slope of the linear fit of the Cp vs D curves
(eq 5).
| 3 |
| 4 |
| 5 |
To discuss the effects of GC content, we can first look into the A10 and G10 systems. The values for PCR-evaluated lesions, AFM-evaluated DSB, and the corresponding enhancement factors when comparing the damages to the control strand in Position 2 are summarized in Table 1. Within the sensitivity of the present methods, increasing and decreasing the GC content by incorporation of contiguous G10:C10 and A10:T10 sequences do not show particular enhancement in the strand break yields. It has been observed that, with low energy electron irradiated sequences in the dry state, oligomers with contiguous guanine and cytosine strands have lesser strand break cross section compared to adenine and thymine;19 however, certain studies on sequence dependence of γ radiation damage on plasmids in solution, where indirect effects play a significant role, identify cytosine as the preferred damage site for SSB and DSB and guanine for base damage.37 It appears that both effects play a role in the damage observed for said systems within the radical scavenging conditions of the solution used in this experiment.
Table 1. Estimated Strand Break Yields of the Various DNA Motifs Studied after Irradiation with 16 MeV Electronsa.
| DNA Motif | μTL ± SD (total lesions·kGy–1) | EF (total lesions) | μDSB ± SD (DSB·kGy–1) | EF (DSB) |
|---|---|---|---|---|
| A10 | 0.73 ± 0.04 | 1.02 ± 0.07 | 0.030 ± 0.005 | 0.98 ± 0.19 |
| R(A10) | 0.71 ± 0.03 | 0.031 ± 0.006 | ||
| G10 | 0.64 ± 0.04 | 0.95 ± 0.09 | 0.043 ± 0.004 | 0.98 ± 0.11 |
| R(G10) | 0.67 ± 0.04 | 0.044 ± 0.002 | ||
| 2FC | 0.70 ± 0.06 | 0.98 ± 0.12 | 0.118 ± 0.010 | 1.27 ± 0.13 |
| R(2FC) | 0.71 ± 0.06 | 0.093 ± 0.005 | ||
| BrA | 1.19 ± 0.12 | 1.49 ± 0.16 | 0.144 ± 0.014 | 1.19 ± 0.13 |
| R(BrA) | 0.79 ± 0.04 | 0.121 ± 0.007 |
The enhancement factors are taken from the ratio of μTL or μDSB between the modified strand and the control strand (R) in each system.
For the sensitized strands, in the 2FC system in which the halogen is attached to the sugar unit of the DNA, the total lesions are the same as the control strand as revealed by RT-PCR measurements; however, the double strand break yield is enhanced. So far radiosensitization of 2′-deoxy-2′-fluorocytidine has not yet been explored in the literature, but it appears that it does have a significant effect in increasing DSB damage in irradiated DNA. As the numbers of lesions in the modified and control DNA strands are the same, the mechanism can be explained by the different evolution of clustered damages in the two systems. While clustered DNA damages will remain non-DSB in the random sequence, they will result in DSB in the 2FC enhanced sequence. This can be caused by the formation of sugar radicals known to induce irreversible DNA damages,38 possibly cutting the neighboring strands or just by the higher susceptibility of the 2FC strand to decay.
In the 8-bromoadenosine containing strand, both total lesions and DSB yields are amplified compared to the nonsensitized strand. The enhancement on DSB is lower than that on total lesions, which means that the strand breaks on the brominated base do not mostly result in DSB. The DNA sensitization by 8-bromoadenosine was explored in experiments with low-energy electrons irradiating single-stranded DNA sequences immobilized on DNA origami nanotriangles39 and explained by experiments with isolated molecules.33 An average enhancement factor of 1.9 ± 0.6 for irradiations by electrons in the 0.5–9 eV range was observed when comparing 8-bromoadenosine incorporated strands with the same sequence without the sensitizer, not far from the enhancement observed here for total lesions.
The damage profiles for 2FC and BrA indicate that the mechanisms of sensitization by these compounds are different. While BrA enhances number of single strand breaks, 2FC amplifies the number of DSB damages at the same SSB level.
It can be noted that the number of DSBs on the R strand in Position 2 is also increased by the presence of sensitized strands (R(2FC) and R(BrA) in Table 1)) when compared to the R strand in the presence of A/G enhanced strands (R(A10) and (R(G10) in Table 1). This can be explained as “bystander” effects, when release and diffusion of reactive species from the fragmentation of the sensitized strand can cause damages in the neighboring random sequence. In the present nanoframe design, the distance between the parallel strands is ∼12 nm. However, the DNA origami technique allows for change of the distance by redesigning the nanoframe and can thus elicit more detailed exploration of this effect not only with respect to different sequences of DNA strands but also the incorporation of various types of radiosensitizers such as gold nanoparticles, which we plan to explore in the future.
Lastly, we can also compare the results of the strand break yields obtained from RT-PCR and AFM to that observed in plasmid irradiated studies at similar conditions. While DSBs can be directly compared, the number of lesions do not exactly correspond to the number of single-strand breaks. As described earlier, these encompass strand breaks and base damages on the PCR amplified strand of the double stranded DNA sequence, and therefore, the given numbers are only the lowest estimates of the SSBs. The estimates for the control strands are shown in Table 2 and are within the range of strand break yields observed in the literature. It is difficult to directly compare the values due to the differences in parameters such as the radical scavenging capacities and dose rates utilized, but both absolute values and SSB to DSB ratios are within the same range. We therefore believe that the present method can be a viable substitute or complementary technique to assess radiation induced damage to DNA sequences in solution. The nanoframe approach also allows new questions to be addressed in radiation damage such as evaluating “bystander” effects with nanometer precision as well as consequences of such damage to DNA conformation.
Table 2. Strand Break Yields Obtained from This Work and Those Obtained from Gel Electrophoresis Results of High Energy Electron Irradiated Plasmid DNA in the Literature.a.
| plasmid/system | electron energy (MeV) | scavenging capacity (s–1) | dose rate (Gy s–1) | μSSB ± SD (SSB·Mbp–1·Gy–1) | μDSB ± SD (DSB·Mbp–1·Gy–1) | ref |
|---|---|---|---|---|---|---|
| 64 bp duplex on DNA origami frames | 16 | 107 | 6.7 | 10.58 ± 0.53 | 0.51 ± 0.10 | this work |
| 9.85 ± 0.7 | 0.69 ± 0.03 | |||||
| 9.64 ± 0.95 | 1.45 ± 0.08 | |||||
| 10.51 ± 0.57 | 1.89 ± 0.11 | |||||
| pBR322 | 16 | 7 × 105 | 0.2 | 33.25 ± 3.21 | 0.19 ± 0.07 | Perstin et al. 202240 |
| 47 | 12.22 ± 0.37 | 0.12 ± 0.04 | ||||
| 93 | 13.57 ± 0.57 | 0.12 ± 0.05 | ||||
| pBR322 | 6 | 106 | N.S. | 7.94 ± 0.14 | 0.97 ± 0.10 | Small et al. 201941 |
| 10 | 11.00 ± 0.66 | 1.13 ± 0.08 | ||||
| 15 | 7.71 ± 0.03 | 1.22 ± 0.01 | ||||
| pBR322 | 100 | 106 | 0.5 | 15.42 ± 0.86 | 0.35 ± 0.02 | Small et al. 202142 |
| 150 | 17.63 ± 0.57 | 0.35 ± 0.03 | ||||
| 200 | 20.19 ± 0.56 | 0.38 ± 0.02 | ||||
| 100 | 106 | ∼108 (flash) | 20.31 ± 1.20 | 0.37 ± 0.03 | ||
| 150 | 18.74 ± 0.52 | 0.37 ± 0.04 | ||||
| 200 | 21.22 ± 0.38 | 0.38 ± 0.02 |
To conclude, we demonstrated the use of DNA origami frames to study radiation damage to doubly stranded DNA sequences in aqueous solution, particularly in assessing double strand breaks and radiosensitization/enhancement factors of halogenated nucleosides by AFM. RT-PCR was used to confirm dose-dependent damage by detecting the relative amount of remaining intact DNA motifs. The introduction of contiguous sequences of A-T and G-C base pairs did not result in changes in strand break yields detectable by the present approach. Introducing fluorine on the sugar unit of the DNA does not significantly induce more lesions compared to the unsensitized DNA strand but has enhanced double strand break yields. Bromoadenine consistently shows strand break enhancement both for total lesion and for double strand breaks. The strand break yields obtained using the method are close to those obtained in plasmid irradiated with MeV electrons; hence, it can be a promising tool to study radiation damage to DNA sequences of interest.
Acknowledgments
This work was supported by the Czech Science Foundation, Project No. 21-26601X, and projects “Center for Tumor Ecology - Research of the Cancer Microenvironment Supporting Cancer Growth and Spread” (Reg. No. CZ.02.1.01/0.0/0.0/16_019/0000785) and CARAT “Carbon allotropes with rationalized nanointerfaces and nanolinks for environmental and biomedical applications” (Reg. No. CZ.02.1.01/0.0/0.0/16_026/0008382) supported by the Operational Programme Research, Development and Education.
Supporting Information Available
The Supporting Information is available free of charge at https://pubs.acs.org/doi/10.1021/acs.jpclett.2c00584.
The authors declare no competing financial interest.
Supplementary Material
References
- Huang R.-X.; Zhou P.-K. DNA Damage Response Signaling Pathways and Targets for Radiotherapy Sensitization in Cancer. Signal Transduct. Target. Ther. 2020, 5, 60. 10.1038/s41392-020-0150-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seiwert T. Y.; Salama J. K.; Vokes E. E. The Concurrent Chemoradiation Paradigm—General Principles. Nat. Clin. Pract. Oncol. 2007, 4, 86–100. 10.1038/ncponc0714. [DOI] [PubMed] [Google Scholar]
- Alizadeh E.; Orlando T. M.; Sanche L. Biomolecular Damage Induced by Ionizing Radiation: The Direct and Indirect Effects of Low-Energy Electrons on DNA. Annu. Rev. Phys. Chem. 2015, 66, 379–398. 10.1146/annurev-physchem-040513-103605. [DOI] [PubMed] [Google Scholar]
- Huwaidi A.; Kumari B.; Robert G.; Guérin B.; Sanche L.; Wagner J. R. Profiling DNA Damage Induced by the Irradiation of DNA with Gold Nanoparticles. J. Phys. Chem. Lett. 2021, 12, 9947–9954. 10.1021/acs.jpclett.1c02598. [DOI] [PubMed] [Google Scholar]
- Śmiałek M. A.; Moore S. A.; Mason N. J.; Shuker D. E. G. Quantification of Radiation-Induced Single-Strand Breaks in Plasmid DNA using a TUNEL/ELISA-Based Assay. Radiat. Res. 2009, 172, 529–536. 10.1667/RR1684.1. [DOI] [PubMed] [Google Scholar]
- Pachnerová Brabcová K.; Sihver L.; Ukraintsev E.; Štěpán V.; Davídková M. How Detection of Plasmid DNA Fragmentation Affects Radiation Strand Break Yields. Radiat. Prot. Dosim. 2019, 183, 89–92. 10.1093/rpd/ncy222. [DOI] [PubMed] [Google Scholar]
- Gao Y.; Zheng Y.; Sanche L. Low-Energy Electron Damage to Condensed-Phase DNA and Its Constituents. Int. J. Mol. Sci. 2021, 22, 7879. 10.3390/ijms22157879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong Y.; Gao Y.; Liu W.; Gao T.; Zheng Y.; Sanche L. Clustered DNA Damage Induced by 2–20 eV Electrons and Transient Anions: General Mechanism and Correlation to Cell Death. J. Phys. Chem. Lett. 2019, 10, 2985–2990. 10.1021/acs.jpclett.9b01063. [DOI] [PubMed] [Google Scholar]
- Schlatholter; Lacombe S.; Eustache P.; Porcel E.; Salado D.; Stefancikova L.; Tillement O.; Lux F.; Mowat P.; van Goethem M.-J.; Remita H.; Biegun A.; et al. Improving proton therapy by metal-containing nanoparticles: nanoscale insights. Int. J. Nanomedicine 2016, 11, 1549–1556. 10.2147/IJN.S99410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rezaee M.; Sanche L.; Hunting D. J. Cisplatin Enhances the Formation of DNA Single- and Double-Strand Breaks by Hydrated Electrons and Hydroxyl Radicals. Radiat. Res. 2013, 179, 323–331. 10.1667/RR3185.1. [DOI] [PubMed] [Google Scholar]
- Reimitz D.; Davídková M.; Mestek O.; Pinkas J.; Kočišek J. Radiomodifying effects of RAPTA C and CDDP on DNA strand break induction. Radiat. Phys. Chem. 2017, 141, 229–234. 10.1016/j.radphyschem.2017.07.015. [DOI] [Google Scholar]
- Hahn M. B.; Meyer S.; Schröter M.-A.; Seitz H.; Kunte H.-J.; Solomun T.; Sturm H. Direct Electron Irradiation of DNA in a Fully Aqueous Environment. Damage Determination in Combination with Monte Carlo Simulations. Phys. Chem. Chem. Phys. 2017, 19, 1798–1805. 10.1039/C6CP07707B. [DOI] [PubMed] [Google Scholar]
- Ribar A.; Huber S. E.; Śmiałek M. A.; Tanzer K.; Neustetter M.; Schürmann R.; Bald I.; Denifl S. Hydroperoxyl Radical and Formic Acid Formation from Common DNA Stabilizers upon Low Energy Electron Attachment. Phys. Chem. Chem. Phys. 2018, 20, 5578–5585. 10.1039/C7CP07697E. [DOI] [PubMed] [Google Scholar]
- Balasubramanian B.; Pogozelski W. K.; Tullius T. D. DNA Strand Breaking by the Hydroxyl Radical is Governed by the Accessible Surface Areas of the Hydrogen Atoms of the DNA Backbone. Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 9738–9743. 10.1073/pnas.95.17.9738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solomun T.; Hultschig C.; Illenberger E. Microarray Technology for the Study of DNA Damage by Low-Energy Electrons. Eur. Phys. J. D 2005, 35, 437–441. 10.1140/epjd/e2005-00204-8. [DOI] [Google Scholar]
- Schürmann R.; Vogel S.; Ebel K.; Bald I. The Physico-Chemical Basis of DNA Radiosensitization: Implications for Cancer Radiation Therapy. Chem. Eur. J. 2018, 24, 10271–10279. 10.1002/chem.201800804. [DOI] [PubMed] [Google Scholar]
- Li Z.; Cloutier P.; Sanche L.; Wagner J. R. Low-Energy Electron-Induced DNA Damage: Effect of Base Sequence in Oligonucleotide Trimers. J. Am. Chem. Soc. 2010, 132, 5422–5427. 10.1021/ja9099505. [DOI] [PubMed] [Google Scholar]
- Dey S.; Fan C.; Gothelf K. V.; Li J.; Lin C.; Liu L.; Liu N.; Nijenhuis M. A. D.; Saccà B.; Simmel F. C.; et al. DNA Origami. Nat. Rev. Methods Primers 2021, 1, 13. 10.1038/s43586-020-00009-8. [DOI] [Google Scholar]
- Vogel S.; Rackwitz J.; Schürman R.; Prinz J.; Milosavljević A. R.; Réfrégiers M.; Giuliani A.; Bald I. Using DNA Origami Nanostructures To Determine Absolute Cross Sections for UV Photon-Induced DNA Strand Breakage. J. Phys. Chem. Lett. 2015, 6, 4589–4593. 10.1021/acs.jpclett.5b02238. [DOI] [PubMed] [Google Scholar]
- Rackwitz J.; Bald I. Low-Energy Electron-Induced Strand Breaks in Telomere-Derived DNA Sequences—Influence of DNA Sequence and Topology. Chem. Eur. J. 2018, 24, 4680–4688. 10.1002/chem.201705889. [DOI] [PubMed] [Google Scholar]
- Keller A.; Bald I.; Rotaru A.; Cauët E.; Gothelf K. V.; Besenbacher F. Probing Electron-Induced Bond Cleavage at the Single-Molecule Level Using DNA Origami Templates. ACS Nano 2012, 6, 4392–4399. 10.1021/nn3010747. [DOI] [PubMed] [Google Scholar]
- Keller A.; Rackwitz J.; Cauët E.; Liévin J.; Körzdörfer T.; Rotaru A.; Gothelf K. V.; Besenbacher F.; Bald I. Sequence Dependence of Electron-induced DNA Strand Breakage Revealed by DNA Nanoarrays. Sci. Rep. 2015, 4, 7391. 10.1038/srep07391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ray A.; Liosi K.; Ramakrishna S. N.; Spencer N. D.; Kuzuya A.; Yamakoshi Y. Single-Molecule AFM Study of DNA Damage by 1O2 Generated from Photoexcited C60. J. Phys. Chem. Lett. 2020, 11, 7819–7826. 10.1021/acs.jpclett.0c02257. [DOI] [PubMed] [Google Scholar]
- Sala L.; Zerolová A.; Rodriguez A.; Reimitz D.; Davídková M.; Ebel K.; Bald I.; Kočišek J. Folding DNA into Origami Nanostructures Enhances Resistance to Ionizing Radiation. Nanoscale 2021, 13, 11197–11203. 10.1039/D1NR02013G. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poppleton E.; Bohlin J.; Matthies M.; Sharma S.; Zhang F.; Šulc P. Design, optimization and analysis of large DNA and RNA nanostructures through interactive visualization, editing and molecular simulation. Nucleic Acids Res. 2020, 48, e72. 10.1093/nar/gkaa417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebel K.; Bald I. Length and Energy Dependence of Low-Energy Electron-Induced Strand Breaks in Poly(A) DNA. Int. J. Mol. Sci. 2020, 21, 111. 10.3390/ijms21010111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Endo M.; Katsuda Y.; Hidaka K.; Sugiyama H. Regulation of DNA Methylation Using Different Tensions of Double Strands Constructed in a Defined DNA Nanostructure. J. Am. Chem. Soc. 2010, 132, 1592–1597. 10.1021/ja907649w. [DOI] [PubMed] [Google Scholar]
- Endo M.; Katsuda Y.; Hidaka K.; Sugiyama H. A Versatile DNA Nanochip for Direct Analysis of DNA Base-Excision Repair. Angew. Chem., Int. Ed. 2010, 49, 9412–9416. 10.1002/anie.201003604. [DOI] [PubMed] [Google Scholar]
- Sannohe Y.; Endo M.; Katsuda Y.; Hidaka K.; Sugiyama H. Visualization of Dynamic Conformational Switching of the G-Quadruplex in a DNA Nanostructure. J. Am. Chem. Soc. 2010, 132, 16311–16313. 10.1021/ja1058907. [DOI] [PubMed] [Google Scholar]
- Adhikary A.; Kumar A.; Rayala R.; Hindi R. M.; Adhikary A.; Wnuk S. F.; Sevilla M. D. One-Electron Oxidation of Gemcitabine and Analogs: Mechanism of Formation of C3′ and C2’ Sugar Radicals. J. Am. Chem. Soc. 2014, 136, 15646–15653. 10.1021/ja5083156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kopyra J.; Keller A.; Bald I. On the Role of Fluoro-Substituted Nucleosides in DNA Radiosensitization for Tumor Radiation Therapy. RSC Adv. 2014, 4, 6825–6829. 10.1039/c3ra46735j. [DOI] [Google Scholar]
- Wityk P.; Wieczór M.; Makurat S.; Chomicz-Mańka L.; Czub J.; Rak J. Dominant Pathways of Adenosyl Radical-Induced DNA Damage Revealed by QM/MM Metadynamics. J. Chem. Theory Comput. 2017, 13, 6415–6423. 10.1021/acs.jctc.7b00978. [DOI] [PubMed] [Google Scholar]
- Schürmann R.; Tanzer K.; Dabkowska I.; Denifl S.; Bald I. Stability of the Parent Anion of the Potential Radiosensitizer 8-Bromoadenine Formed by Low-Energy (< 3 eV) Electron Attachment. J. Phys. Chem. B 2017, 121, 5730–5734. 10.1021/acs.jpcb.7b02130. [DOI] [PubMed] [Google Scholar]
- González L. N.; Arruda-Neto J. D. T.; Cotta M. A.; Carrer H.; Garcia F.; Silva R. A. S.; Moreau A. L. D.; Righi H.; Genofre G. C. DNA fragmentation by gamma radiation and electron beams using atomic force microscopy. J. Biol. Phys. 2012, 38, 531–542. 10.1007/s10867-012-9270-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim S.; Yoon H.; Ryu S.; Jung J.; Lee M.; Kim D. A Comparative Evaluation of Radiation-Induced DNA Damage using Real-Time PCR: Influence of Base Composition. Radiat. Res. 2006, 165, 430–437. 10.1667/RR3507.1. [DOI] [PubMed] [Google Scholar]
- Svec D.; Tichopad A.; Novosadova V.; Pfaffl M. W.; Kubista M. How Good is a PCR Efficiency Estimate: Recommendations for Precise and Robust qPCR Efficiency Assessments. Biomol. Detect. Quantif. 2015, 3, 9–16. 10.1016/j.bdq.2015.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murray V.; Hardie M. E.; Gautam S. D. Comparison of Different Methods to Determine the DNA Sequence Preference of Ionising Radiation-Induced DNA Damage. Genes 2020, 11, 8. 10.3390/genes11010008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chatgilialoglu C.Radical and Radical Ion Reactivity in Nucleic Acid Chemistry; John Wiley & Sons, Ltd.: 2009; Chapter 4, pp 99–133. [Google Scholar]
- Schürmann R.; Tsering T.; Tanzer K.; Denifl S.; Kumar S. V. K.; Bald I. Resonant Formation of Strand Breaks in Sensitized Oligonucleotides Induced by Low-Energy Electrons (0.5–9 eV). Angew. Chem., Int. Ed. 2017, 56, 10952–10955. 10.1002/anie.201705504. [DOI] [PubMed] [Google Scholar]
- Perstin A.; Poirier Y.; Sawant A.; Tambasco M. Quantifying the DNA-Damaging Effects of FLASH Irradiation with Plasmid DNA. Int. J. Radiat. Oncol. Biol. Phys. 2022, 10.1016/j.ijrobp.2022.01.049. [DOI] [PubMed] [Google Scholar]
- Small K.; Angal-Kalinin D.; Chadwick A.; Edge R.; Henthorn N.; Jones R.; Kirkby K.; Merchant M.; Morris R.; Santina E.; et al. A Comparative Study of Biological Effects of Electrons and Co-60 Gamma Rays on pBR322 Plasmid DNA. Proc. of the 10th Int. Particle Accelerator Conf. 2019, 3533–3536. 10.18429/JACoW-IPAC2019-THPMP041. [DOI] [Google Scholar]
- Small K. L.; Henthorn N. T.; Angal-Kalinin D.; Chadwick A. L.; Santina E.; Aitkenhead A.; Kirkby K. J.; Smith R. J.; Surman M.; Jones J.; et al. Evaluating Very High Energy Electron RBE from nanodosimetric pBR322 plasmid DNA damage. Sci. Rep. 2021, 11, 3341. 10.1038/s41598-021-82772-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.