

Abstract
Plant pathogens evade basal defense systems and attack different organs and tissues of plants. Genetic engineering of plants with genes that confer resistance against pathogens is very effective in pathogen control. Conventional breeding for disease resistance in ornamental crops is difficult and lagging relative to that in non-ornamental crops due to an inadequate number of disease-resistant genes. Therefore, genetic engineering of these plants with defense-conferring genes is a practical approach. We used rice BSR2 encoding CYP78A15 for developing transgenic Torenia fournieri Lind. lines. The overexpression of BSR2 conferred resistance against two devastating fungal pathogens, Rhizoctonia solani and Botrytis cinerea. In addition, BSR2 overexpression resulted in enlarged flowers with enlarged floral organs. Histological observation of the petal cells suggested that the enlargement in the floral organs could be due to the elongation and expansion of the cells. Therefore, the overexpression of BSR2 confers broad-spectrum disease resistance and induces the production of enlarged flowers simultaneously. Therefore, this could be an effective strategy for developing ornamental crops that are disease-resistant and economically more valuable.
Keywords: torenia, BSR2, disease resistance, genetic engineering, Botrytis cinerea, Rhizoctonia solani, flower size, ornamental crops
1. Introduction
Phytopathogens evade the basal defense systems of host plants to obtain specific nutrients; most of them infect only specific hosts. However, they infect different organs and tissues, depending on the host plant species. In addition, some of them infect a wide range of host plants and organs, and they cause devastating diseases in many crops. The fungus Botrytis cinerea causes gray mold in more than 200 plant species, including vegetables, fruit crops, and ornamental crops; it can infect many tissues, including leaves, stems, flowers, and fruits [1]. Rhizoctonia solani, a soil-borne fungus, infects leaves and causes sheath blight in rice [2]. In addition, it causes root rot in sugar beet and damping off in tomatoes and many ornamental crops [3] by infecting the roots. Thus, R. solani infects different tissues and causes devastating diseases, depending on the host plant species. A control strategy against these pathogens would be the introduction of disease-resistant genes into the host plants using the conventional breeding approach. However, there are few genes that have been found to be effective against these pathogens. Ornamental crops are considered high-quality products; therefore, they should have higher disease resistance compared to that of other crops. Ornamental crops cover many species and varieties, which makes conventional breeding more difficult in these plants. Therefore, introducing defense-conferring genes in these crops by genetic engineering could be an efficient approach.
The overexpression (OX) of rice (Oryza sativa L.) BROAD-SPECTRUM RESISTANCE2 (BSR2) gene, encoding a cytochrome P450 (CYP78A15), confers resistance against the bacterial pathogen Pseudomonas syringae pv. tomato DC3000 (PstDC3000), and the fungal pathogens Colletotrichum higginsianum and R. solani in Arabidopsis thaliana [4]. In addition, BSR2-OX Arabidopsis exhibits enlarged reproductive organs, slower growth, and extremely low seed fertility compared to that in the wild-type (WT) plants [4]. Some members of the CYP78As, such as CYP78A5, A6, and A9 influence the morphogenesis of reproductive organs [5,6,7], which could be attributed to a novel mobile growth signal distinct from that of the classical phytohormone involved in this process [5,6]. Similar to that in Arabidopsis, BSR2-OX confers resistance to two R. solani isolates in rice; these rice exhibit larger seeds and flowers, slower growth, and extremely low seed fertility compared to that in WT plants [4]). BSR2-OX tomato displays broad-spectrum disease resistance against fungi, such as B. cinerea and R. solani, and against bacteria, such as P. syringae pv. tomato and Ralstonia pseudosolanacearum. However, the overexpression of BSR2 does not induce remarkable morphological changes in tomato [8]. We validated whether BSR2-OX can confer disease resistance and induce larger flowers in the ornamental crop, Torenia fournieri Lind., which is not only a model but also as a commercially distributed ornamental crop. T. fournieri was chosen because it has a short generation time, small size, and can be propagated using stem cuttings; therefore, it is easy to obtain plant samples in a short time. In addition, its genome size is small (2n = 18, 171 Mbp) [9], and tissue culture and transformation methods for this plant are established.
2. Results
2.1. Overexpression of BSR2 Conferred Resistance against Two Fungal Pathogens in T. fournieri
The BSR2 expression vector contains rice BSR2 cloned downstream of the constitutive 35S promoter; it was used for tomato transformation [8]. Using this vector, several transgenic T. fournieri lines were generated. The overexpression of BSR2 was confirmed using quantitative RT-PCR (qRT-PCR) of independent T0 plant lines (Figure 1). Plants propagated through cuttings were used for the subsequent experiments. For the convenience of propagation, several different OX lines were used in different experiments. First, we examined whether BSR2-OX T. fournieri was resistant to R. solani isolate (MAFF235116; AG-4 ⅢA) infection in the roots. All WT plants died; however, 80% of the plants from the two BSR2-OX lines survived until five days after inoculation (Figure 2). Therefore, BSR2 conferred resistance to R. solani in T. fournieri. Next, we evaluated whether BSR2-OX T. fournieri exhibited resistance to B. cinerea (MAFF237249) inoculation using the drop method [8] in leaves. Leaves of three BSR2-OX T. fournieri lines showed restricted lesion formation around the drop inoculation points; however, leaves of the WT plants developed extended lesions three days after inoculation (Figure 3a). The lesion sizes in the three BSR2-OX lines were significantly lesser than that in the WT plants (Figure 3b).
Figure 1.
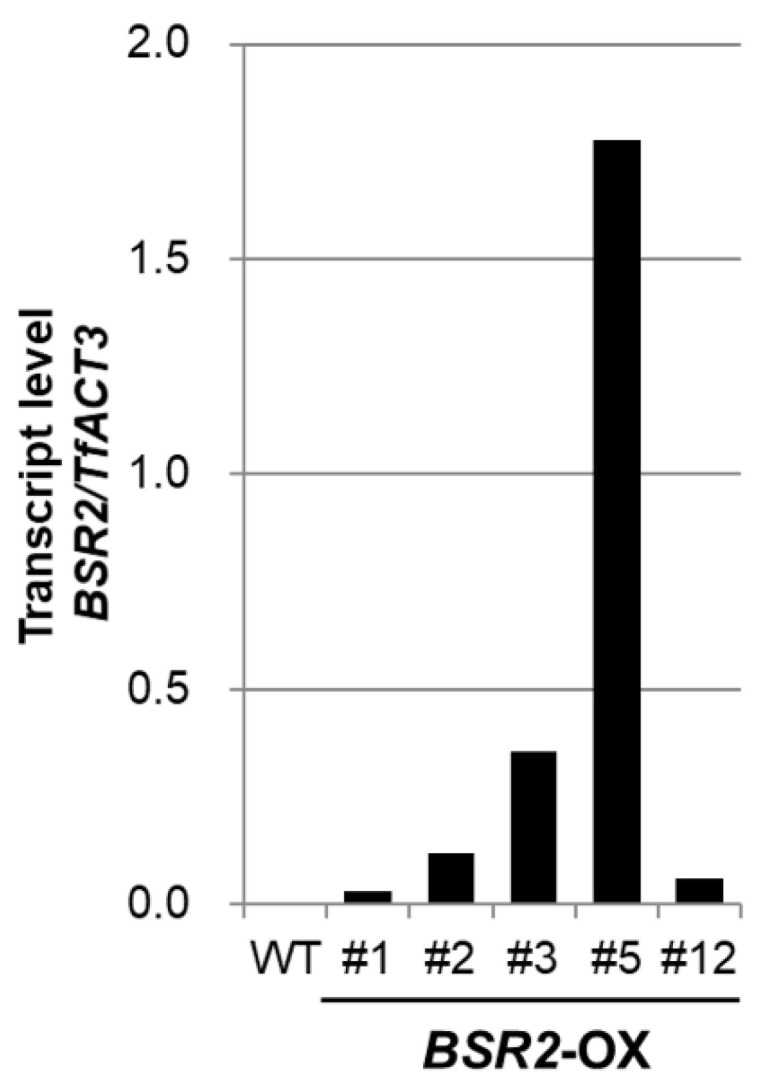
Transcript levels of BSR2 in transgenic torenia lines.
Figure 2.

Survival ratio of transgenic torenia against R. solani (MAFF235116: AG-4IIIA) in the soil inoculation assay. Two-week-old BSR2-OX lines were transplanted into the infected soil inoculated with R. solani, and the survival ratios were calculated 5 days after inoculation (n = 5).
Figure 3.

(a) Phenotypic changes and (b) lesion size of B. cinerea (MAFF237249) drop inoculation on leaves. Leaves of three BSR2-OX lines were drop inoculated with 6 × 103 spore/mL of B. cinerea (MAFF237249). Error bars represent standard deviations (n = 8). * values significantly different from WT (* p < 0.05; Dunnett’s tests).
2.2. Overexpression of BSR2 Resulted in Enlarged Flowers with Enlarged Floral Organs
Flower or petal morphology is an important breeding target, because it has the most influence on the commercial value. Enlarged flowers are observed when BSR2 is overexpressed in Arabidopsis and rice [4]. We investigated the size of each floral organ in the BSR2-OX Arabidopsis lines before investigating the same in the BSR2-OX T. fournieri lines. The lengths of the floral organs, the petal, sepal, stamen, and pistil, in the two BSR2-OX Arabidopsis lines at four days after flowering were 39.6–47.2%, 72.2–72.8%, 8.4–71.5%, and 100–131% longer than that in the WT, respectively (Figure S1).
Next, we investigated the morphological traits in BSR2-OX T. fournieri lines (6 days after flowering). All BSR2-OX T. fournieri lines had larger flowers than that in the WT. The representative two lines were further analyzed. The floral diameters of the two BSR2-OX T. fournieri lines were 26.9–30.8% larger than that in the WT (Figure 4a,b). We measured the sizes of floral organs (Figure 4c). The sepal, outer stamen, inner stamen, pistil, and ovary longitudinal lengths of the two BSR2-OX T. fournieri lines were 15.9, 23.5–26.5, 17.4–30.4, 9.1–15.2, and 28.6–35.7% longer than that in the WT lines, respectively (Figure 4d–h). Therefore, the overexpression of BSR2 enlarged floral organs in T. fournieri. In addition, the BSR2-OX T. fournieri plants were taller at the reproductive stage (Figure S2), similar to that in BSR2-OX Arabidopsis [4]. Enlarged flowers are often a result of polyploidization through tissue culture and transformation. Flow cytometry using 4′,6-diamidino-2-phenylindole (DAPI) was used to examine the nuclei samples of WT and the representative BSR2-OX T. fournieri lines; the relative fluorescence intensity of the nuclei was measured. The mean values at the main (2C) peak were almost similar between the WT and BSR2-OX lines (Figure S3a,b). In addition, a mixture of the WT and each OX line nuclei samples exhibited only one main peak (Figure S3c). Therefore, polyploidization does not occur in the tested BSR2-OX lines, indicating that the detected phenotypes are the result of BSR2 overexpression.
Figure 4.

Comparison of floral morphology between WT and BSR2-OX torenia. Comparison of (a) flowers and (b) floral diameter between WT and BSR2-OX lines 6 days after flowering. (c) Schematic representation of torenia flower. Comparison of longitudinal lengths of (d) sepals, (e) outer stamens, (f) inner stamens, (g) pistils, and (h) ovaries 6 days after flowering. Error bars represent standard deviation (n = 3). * values significantly different from WT (* p < 0.05; Dunnett’s tests).
Petals with a strong phenotype (Figure 5a) were examined to determine whether the cell size was affected. Sections of the basal parts of the petals, 7 days after flowering, were histologically observed (Figure 5b–d). The length and area of the epidermal cells (ep) and the area of parenchyma cells (pa) adjacent to the vascular cells (va) were measured. The lateral direction length, longitudinal direction length, and area of the BSR2-OX ep were 49.4, 24.3, and 34.0% longer or larger than that of the WT ep, respectively (Figure 5e–g). The area of BSR2-OX pa was 73.2% larger than that in the WT pa (Figure 5h). Therefore, the enlargement of the floral organs was attributed to the hypertrophy of the cells.
Figure 5.

Comparison between the cellular morphology of the petal in WT and BSR2-OX torenia. (a) Comparison of flowers 7 days after flowering. (b–d) Cross-section of the basal part of (c) WT and (d) BSR2-OX#5 petal. ep, epidermal cell; pa, parenchyma cell; va, vascular cell. (e) Lateral and (f) longitudinal dimensions of WT and BSR2-OX epidermal cells in the basal part of the petal. Area of (g) epidermal cells and (h) parenchyma cells adjacent to the vascular cells in the basal part of the petal in WT and BSR2-OX. Error bars represent standard deviation. n = (e) 19–25, (f) 21–25, (g) 19–25, and (h) 10–13. * values significantly different from the WT (* p < 0.05 and *** p < 0.001, t-test).
3. Discussion
In this study, we used two necrotrophic pathogens: B. cinerea, which causes gray mold in many aboveground organs including leaves, stems, flowers, and fruits; and R. solani, which causes various diseases including damping off in roots. BSR2-OX T. fournieri showed resistance to B. cinerea inoculated in leaves (aboveground organs) and to R. solani introduced in roots (underground organs), similar to that in BSR2-OX Arabidopsis and tomato [8]. Therefore, the constitutive overexpression of BSR2 could confer disease resistance in both aboveground and underground organs. The CYP78A subfamily, including BSR2, is distributed widely in land plants including moss, Arabidopsis, tomato, soybean, and wheat [10,11,12,13,14]. Therefore, the BSR2 orthologs could have a function similar to that of BSR2 in plant immunity. BSR2 encodes cytochrome P450; therefore, it would oxidize a substrate to form a “product” related to disease resistance. Arabidopsis, tomato, and T. fournieri plants overexpressing BSR2 are resistant to two necrotrophic fungi; therefore, the “product” could be common among these plants, activating immunity and countering the potent toxins and enzymes derived from both fungi.
In Arabidopsis, CYP78A5(KLUH), A6, and A9 regulate reproductive organ size [6,7,15]. CYP78A5 (KLUH)-OX Arabidopsis has enlarged flower organs because of increased cell proliferation [6]. CYP78A9-OX and CYP78A6-OX Arabidopsis have larger floral organs, fruits, and seeds [5,7,15]. In CYP78A6-OX Arabidopsis, there is an increase in the cell size and cell number in the reproductive organs [7]. BSR2 is homologous to Arabidopsis CYP78A6, A8, and A9 [4]. The size of each floral organ increased in BSR2-OX Arabidopsis and T. fournieri; therefore, to evaluate the change in cell size, we performed histological studies. The enlargement of the floral organs was due to the elongation and expansion of the cells; however, we could not deny the possibility of increased cell number.
The overexpression of BSR2, from the monocot rice, confers disease resistance against two fungal pathogens and induces enlargement (around 30% in diameter) of the flowers in the dicot T. fournieri. These findings encourage the use of BSR2 for conferring broad-spectrum disease resistance and for inducing larger flowers in other ornamental crops. Arabidopsis CYP78A9 influences the flavanol biosynthesis pathway [15]; maize CYP78A1 exhibits lauric acid 12-monooxygenase activity [16]; and tomato CYP78A5 orthologue (SlKLUH) influences lipid metabolism [17]. However, CYP78A family members, excluding BSR2, have not been reported to be involved in disease resistance. This is likely due to a lack of research about the same thus far. The future studies should focus on identifying the secondary metabolites biosynthesized by the BSR2 protein, CYP78A15. This would enable elucidating the mechanism underlying the defense against broad-spectrum pathogens and the flower morphogenesis, and it could also lead to more efficient breeding in ornamental crops.
From an agricultural point of view, gray mold caused by B. cinerea results in severe economic losses in many important ornamental crops [18]. Gray mold is also a major post-harvest disease as well as a growth stage disease of roses [19]. In this study, leaves in cut branches of BSR2-OX T. fournieri plants showed B. cinerea resistance, suggesting that BSR2-OX would be effective in the post-harvest stage; in other words, protection by BSR2-OX might be effective in the cut flowers. Generally, commercially important ornamental crops require protection from fungal diseases in both growth and post-harvest stages. Hence, the overexpression of BSR2 could be an attractive tool in the ornamental crops.
T. fournieri ‘Crown Violet’ used as a WT material in this work is derived from F1 hybrid seeds of ‘Crown Mix’; many phenotypes change in the subsequent generations due to segregation; therefore, seed proliferation of BSR2-OX T. fournieri plants was not expected from a practical point of view. Instead, vegetative propagation via cuttings would be more practical. Generally, the overexpression of BSR2 would lead to low fertility, considering the low fertility of BSR2-OX Arabidopsis and BSR2-OX rice [4], although we do not have data relating to fertility in this study. However, when low fertility is not preferable, the use of a leaf-specific and/or floral organ (e.g., petal)-specific promoter [20] would be effective in maintaining fertility. The expression of BSR2 under such mixed promoters would help confer disease resistance and induce enlarged flowers simultaneously without decreasing fertility.
4. Materials and Methods
4.1. Plant and Microbial Materials and Culture
T. fournieri Lind. ‘Crown Violet (CrV)’ was used as a WT material. Violet-flowered CrV cultivar selected from F1 hybrid seeds of ‘Crown Mix’ (Sakata Seed Co., Yokohama, Japan) was kindly provided by Dr. Ryutaro Aida (Institute of Vegetable and Floriculture Science, NARO, Tsukuba, Japan). One vigorously growing F1 hybrid plant was propagated vegetatively by herbaceous cutting by Dr. Ryutaro Aida and used as the experimental line.
The WT and transgenic T. fournieri plants were cultured and maintained on MS medium with sucrose and/or trehalose as a carbon source, as described previously [21]. Sterile moistened black peat moss (Sakata Super Mix A) was used for T. fournieri soil culture. BSR2-OX Arabidopsis lines and Col-0 (WT) were grown, as described previously [4].
The fungal pathogens R. solani (MAFF235116: AG-4ⅢA) and B. cinerea (MAFF237249) were used to test disease resistance in T. fournieri. The pathogens were cultured, as described previously [8].
4.2. T. fournieri Transformation
In order to generate transgenic T. fournieri plants overexpressing BSR2 (Os08g0547300), the recombinant binary plasmid pBIG2113SF-BSR2 [4] was used. T. fournieri transformation was performed, as described previously [22,23]; 20 μg L−1 of hygromycin-B was used for the selection. T0 plants propagated through cuttings were used for the experiments.
4.3. RNA Extraction and Quantitative Real-Time RT-PCR (qRT-PCR) Analysis
Total RNA was extracted and purified from T. fournieri leaves using Sepasol-RNA Super G reagent (Nacalai Tesque, Kyoto, Japan) and qRT-PCR was performed, as described previously [4]. The primers used for qRT-PCR were: BSR2: 5′-GGACTAAGACGAGGAGAGGGAAG-3′ and 5′-AACGTAGGGGCATTTCTACTCAA-3′; TfACT3: 5′-AAGATATGCATTGGAGTTGTGAGTG-3′ and 5′-TCCAAGCTAAGGTAGCAAAACGA-3′.
4.4. Fungal Pathogen Resistance Assay
An inoculum suspension of R. solani was prepared as described previously [8] and mixed with sterile moistened black peat moss (Sakata Super Mix A). Plants were cut about two weeks before inoculation; rooting was promoted in water. They were transferred to the infected soil and grown under long-day conditions (16 h light and 8 h dark) at 25 °C for 5 days; survival ratios were calculated. An inoculum suspension of B. cinerea (6 × 103 conidia/mL) was prepared, as previously described [8]. Vegetative branches with one node and leaves were cut from T. fournieri plants, and then, the cuttings were inserted into the floral form with water. B. cinerea inoculum (5 µL) was dropped on the leaves of the cut branches. The inoculated branches were incubated under humid, long-day conditions, at 25 °C, and the lesion sizes were measured three days after inoculation.
4.5. Flow Cytometry
Flow cytometry using a Ploidy Analyser/Flow cytometer (Partec GmbH, Munster, Germany) was performed, as described previously [24,25]. The staining solution contained 10 mM Tris, 2 mM MgCl2, 0.1% (v/v) Triton X-100, and 2 mg L−1 DAPI at pH 7.5.
Acknowledgments
We thank Hiroyasu Yamaguchi (NIVFS, Japan) for providing a flow cytometer and much advice regarding flow cytometry. We thank Takuo Fujimoto (NIVFS, Japan) for kind advice regarding data analysis of flow cytometry. We also thank Lois Ishizaki, Tomiko Senba, Chiyoko Umeda, and Yuka Yamazaki (NIAS, Japan) for their support with overall technical assistance.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/ijms23094735/s1.
Author Contributions
M.M. and N.O. designed this research. S.M., K.S., N.O., H.K. and Y.K. conducted experiments. S.M. and M.M. analyzed data and wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the grants for the Research Program on Development of Innovative Technology (NARO Bio-oriented Technology Research Advancement Institution (Number 29004A)), and by the JSPS KAKENHI Grant (Number JP20H02953).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Williamson B., Tudzynski B., Tudzynski P., van Kan J.A. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007;8:561–580. doi: 10.1111/j.1364-3703.2007.00417.x. [DOI] [PubMed] [Google Scholar]
- 2.Molla K.A., Karmakar S., Molla J., Bajaj P., Varshney R.K., Datta S.K., Datta K. Understanding sheath blight resistance in rice: The road behind and the road ahead. Plant Biotechnol. J. 2020;18:895–915. doi: 10.1111/pbi.13312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Benson D.M., Cartwright D.K. Rhizoctonia Species: Taxonomy, Molecular biology, Ecology, Pathology and Disease Control. Springer; Berlin/Heidelberg, Germany: 1996. Ornamental diseases incited by Rhizoctonia spp. pp. 303–314. [Google Scholar]
- 4.MAEDA S., Dubouzet J.G., Kondou Y., Jikumaru Y., Seo S., Oda K., Matsui M., Hirochika H., Mori M. The rice CYP78A gene BSR2 confers resistance to Rhizoctonia solani and affects seed size and growth in Arabidopsis and rice. Sci. Rep. 2019;9:587. doi: 10.1038/s41598-018-37365-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ito T., Meyerowitz E.M. Overexpression of a gene encoding a cytochrome P450, CYP78A9, induces large and seedless fruit in Arabidopsis. Plant Cell. 2000;12:1541–1550. doi: 10.1105/tpc.12.9.1541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Anastasiou E., Kenz S., Gerstung M., MacLean D., Timmer J., Fleck C., Lenhard M. Control of plant organ size by KLUH/CYP78A5-dependent intercellular signaling. Dev. Cell. 2007;13:843–856. doi: 10.1016/j.devcel.2007.10.001. [DOI] [PubMed] [Google Scholar]
- 7.Fang W., Wang Z., Cui R., Li J., Li Y. Maternal control of seed size by EOD3/CYP78A6 in Arabidopsis thaliana. Plant J. 2012;70:929–939. doi: 10.1111/j.1365-313X.2012.04907.x. [DOI] [PubMed] [Google Scholar]
- 8.Maeda S., Yokotani N., Oda K., Mori M. Enhanced resistance to fungal and bacterial diseases in tomato and Arabidopsis expressing BSR2 from rice. Plant Cell Rep. 2020;39:1493–1503. doi: 10.1007/s00299-020-02578-0. [DOI] [PubMed] [Google Scholar]
- 9.Kikuchi S., Tanaka H., Shiba T., Mii M., Tsujimoto H. Genome size, karyotype, meiosis and a novel extra chromosome in Torenia fournieri, T. baillonii and their hybrid. Chromosome Res. 2006;14:665–672. doi: 10.1007/s10577-006-1077-y. [DOI] [PubMed] [Google Scholar]
- 10.Katsumata T., Fukazawa J., Magome H., Jikumaru Y., Kamiya Y., Natsume M., Kawaide H., Yamaguchi S. Involvement of the CYP78A subfamily of cytochrome P450 monooxygenases in protonema growth and gametophore formation in the moss Physcomitrella patens. Biosci. Biotechnol. Biochem. 2011;75:331–336. doi: 10.1271/bbb.100759. [DOI] [PubMed] [Google Scholar]
- 11.Bak S., Beisson F., Bishop G., Hamberger B., Höfer R., Paquette S., Werck-Reichhart D. Cytochromes p450. Arab. Book. 2011;9:e0144. doi: 10.1199/tab.0144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chakrabarti M., Zhang N., Sauvage C., Muños S., Blanca J., Cañizares J., Diez M.J., Schneider R., Mazourek M., McClead J., et al. A cytochrome P450 regulates a domestication trait in cultivated tomato. Proc. Natl. Acad. Sci. USA. 2013;110:17125–17130. doi: 10.1073/pnas.1307313110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wang X., Li Y., Zhang H., Sun G., Zhang W., Qiu L. Evolution and association analysis of GmCYP78A10 gene with seed size/weight and pod number in soybean. Mol. Biol. Rep. 2015;42:489–496. doi: 10.1007/s11033-014-3792-3. [DOI] [PubMed] [Google Scholar]
- 14.Ma M., Zhao H., Li Z., Hu S., Song W., Liu X. TaCYP78A5 regulates seed size in wheat (Triticum aestivum) J. Exp. Bot. 2016;67:1397–1410. doi: 10.1093/jxb/erv542. [DOI] [PubMed] [Google Scholar]
- 15.Sotelo-Silveira M., Cucinotta M., Chauvin A.L., Chávez Montes R.A., Colombo L., Marsch-Martínez N., de Folter S. Cytochrome P450 CYP78A9 is involved in Arabidopsis reproductive development. Plant Physiol. 2013;162:779–799. doi: 10.1104/pp.113.218214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Imaishi H., Matsuo S., Swai E., Ohkawa H. CYP78A1 preferentially expressed in developing inflorescences of Zea mays encoded a cytochrome P450-dependent lauric acid 12-monooxygenase. Biosci. Biotechnol. Biochem. 2000;64:1696–1701. doi: 10.1271/bbb.64.1696. [DOI] [PubMed] [Google Scholar]
- 17.Li Q., Chakrabarti M., Taitano N.K., Okazaki Y., Saito K., Al-Abdallat A.M., van der Knaap E. Differential expression of SIKLUH controlling fruit and seed weight is associated with changes in lipid metabolism and photosynthesis-related genes. J. Exp. Bot. 2021;72:1225–1244. doi: 10.1093/jxb/eraa518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Dean R., Van Kan J.A., Pretorius Z.A., Hammond-Kosack K.E., Di Pietro A., Spanu P.D., Rudd J.J., Dickman M., Kahmann R., Ellis J., et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012;13:414–430. doi: 10.1111/j.1364-3703.2011.00783.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Liu X.T., Cao X.Q., Shi S.C., Zhao N., Li D.D., Fang P.H., Chen X., Qi W.C., Zhang Z. Comparative RNA-Seq analysis reveals a critical role for brassinosteroids in rose (Rosa hybrida) petal defense against Botrytis cinerea infection. BMC Genet. 2018;19:62. doi: 10.1186/s12863-018-0668-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sasaki K., Yamaguchi H., Kasajima I., Narumi T., Ohtsubo N. Generation of Novel Floral Traits Using a Combination of Floral Organ-Specific Promoters and a Chimeric Repressor in Torenia fournieri Lind. Plant Cell Physiol. 2016;57:1319–1331. doi: 10.1093/pcp/pcw081. [DOI] [PubMed] [Google Scholar]
- 21.Yamaguchi H., Sasaki K., Shikata M., Aida R., Ohtsubo N. Trehalose drastically extends the in vitro vegetative culture period and facilitates maintenance of Torenia fournieri plants. Plant Biotechnol. 2011;28:263–266. doi: 10.5511/plantbiotechnology.11.0124c. [DOI] [Google Scholar]
- 22.Aida R., Shibata M. Agrobacterium-Mediated Transformation of Torenia (Torenia fournieri) Breed. Sci. 1995;45:71–74. doi: 10.1270/jsbbs1951.45.71. [DOI] [Google Scholar]
- 23.Aida R. A protocol for transformation of Torenia. Methods Mol. Biol. 2012;847:267–274. doi: 10.1007/978-1-61779-558-9_23. [DOI] [PubMed] [Google Scholar]
- 24.Mishiba K., Ando T., Mii M., Watanabe H., Kokubun H., Hashimoto G., Marchesi E. Nuclear DNA content as an index character discriminating taxa in the genus Petunia sensu Jussieu (Solanaceae) Ann. Bot. 2000;85:665–673. doi: 10.1006/anbo.2000.1122. [DOI] [Google Scholar]
- 25.Yamaguchi H., Shimizu A., Degi K., Morishita T. Effects of dose and dose rate of gamma ray irradiation on mutation induction and nuclear DNA content in chrysanthemum. Breed. Sci. 2008;58:331–335. doi: 10.1270/jsbbs.58.331. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data are contained within the article.