

Abstract
The coronavirus disease 2019 (COVID‐19) pandemic has created unprecedented challenges worldwide. Severe acute respiratory syndrome coronavirus 2 (SARS‐CoV‐2) causes COVID‐19 and has a complex interaction with the immune system, including growing evidence of sex‐specific differences in the immune response. Sex‐disaggregated analyses of epidemiological data indicate that males experience more severe symptoms and suffer higher mortality from COVID‐19 than females. Many behavioural risk factors and biological factors may contribute to the different immune response. This review examines the immune response to SARS‐CoV‐2 infection in the context of sex, with emphasis on potential biological mechanisms explaining differences in clinical outcomes. Understanding sex differences in the pathophysiology of SARS‐CoV‐2 infection will help promote the development of specific strategies to manage the disease.
Keywords: COVID‐19, gender, immune system, SARS‐CoV‐2, sex, sex hormones
This review examines the immune response to SARS‐CoV‐2 infection in the context of sex, with emphasis on potential biological mechanisms explaining differences in clinical outcomes. Understanding sex differences in the pathophysiology of SARS‐CoV‐2 infection will help promote the development of specific strategies to manage the disease.
BIOLOGICAL SEX AND COVID‐19
The coronavirus disease 2019 (COVID‐19) pandemic, caused by the severe acute respiratory syndrome coronavirus 2 (SARS‐CoV‐2), produced enormous global health challenges. Recent epidemiological studies have uncovered critical differences between men and women with respect to COVID‐19 outcomes. Males and females have similar proportions of COVID‐19 cases, suggesting similar infection rates. 1 However, males exhibit higher disease severity and are at a higher risk of succumbing to the disease. 2 , 3 Older age is associated with higher risk of death in both sexes, but even after accounting for age, males are still more likely than females to die from SARS‐CoV‐2 infection. 1 , 4 , 5 , 6 , 7 , 8 , 9 Though males were less likely to be diagnosed with COVID‐19 in one study, their mortality rate was about twice that of females. 4 Mortality rates were higher amongst males across all age groups above 20 years in multiple European countries. 10 Certain groups of females with COVID‐19 are more likely to have worse clinical outcomes. For example, pregnant females with COVID‐19 are at increased risk of severe illness, hospitalization, intensive care unit (ICU) admission, preterm delivery and mortality. 11 , 12 , 13 , 14 , 15 Concerns have also been raised about the use of hormonal contraception in females with COVID‐19. Combined oral contraceptives are associated with a higher risk of venous thromboembolism (VTE), and COVID‐19 disease is associated with hypercoagulability. 16 , 17 , 18 Further studies are needed to evaluate the risk of VTE in females with COVID‐19 and using combined oral contraceptives.
Sex‐specific differences in clinical outcomes and immune response to other viruses have been documented. Higher mortality amongst males has been observed in other human coronavirus infections, including SARS‐CoV‐1 and Middle East respiratory syndrome‐related coronavirus (MERS‐CoV). 19 , 20 Females infected with the human immunodeficiency virus (HIV) mount stronger antiviral responses than males, possibly due to increased toll‐like receptor 7 (TLR7) and interferon alpha (IFNα) activation. 21 , 22 Higher hepatitis B surface antibody (anti‐HBs) titres have been reported in women compared with men following hepatitis B virus (HBV) vaccination. 23
There are many possible explanations for the difference in COVID‐19 outcomes between men and women, including certain behavioural and social factors. 24 However, biological factors such as sex‐related genes and sex hormones that influence immune system regulation may also play an important role. 25 This review will focus on our current understanding of biological mechanisms underlying sex differences in the immune response to SARS‐CoV‐2 infection.
SEX DIFFERENCES IN THE IMMUNE RESPONSE TO SARS‐COV‐2 INFECTION
Females and males have different immune responses to pathogens, which may explain the differing disease severity and mortality due to SARS‐CoV‐2 infection. 26 The immune system's interaction with SARS‐CoV‐2 consists of viral entry into the human cell, followed by recognition of the virus and activation of the host innate immune response, which subsequently leads to the activation of the adaptive immune response (Figure 1). 10 Gene expression in immune cells exhibits different patterns based on sex, being most pronounced in autosomal genes. 10 , 27 The sex chromosomes (X and Y) also carry a number of immune response‐related genes and are involved in immunoregulation as well. For example, transcriptional regulation of immunoregulatory genes can be influenced by the mosaic loss of the Y chromosome in leucocytes. 28
FIGURE 1.
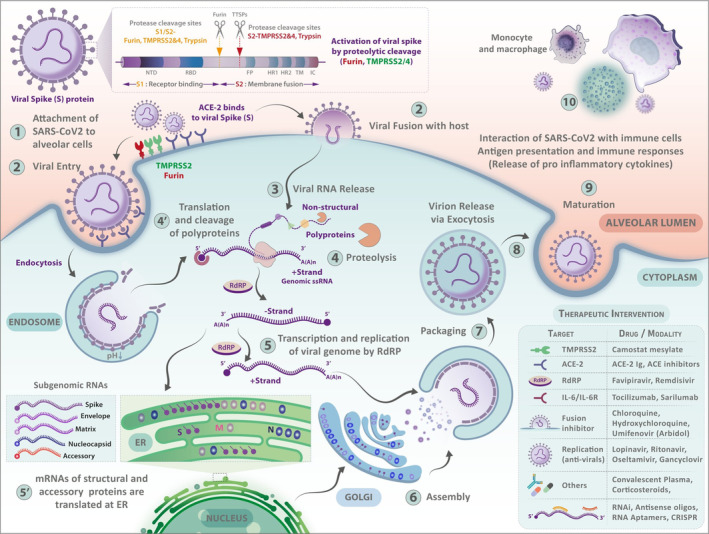
The life cycle of severe acute respiratory syndrome coronavirus 2 (SARS‐CoV‐2). (1) The cycle begins with the virus entering cells of the airway. Viral attachment is mediated by the interaction between the viral spike (S) protein and host ACE2 receptors with the serine protease TMPRSS2 co‐receptors. Furin cleaves the spike protein to make it functional for priming and activation. (2) Either by endocytosis or membrane fusion, the virus enters the host cell. (3) This event is followed by the release of viral ssRNA into the host cell. The release of the viral genome triggers the signals for activation of intracellular pattern recognition receptors such as TLR7 that usually lead to the synthesis of antiviral interferons. (4 & 4′) Subsequently, the translation and cleavage of the viral polymerase protein occur in the cytoplasm. (5 & 5′) RNA replication depends on the viral RNA‐dependent RNA polymerase (RdRp). Translation of viral structural proteins occurs via the ribosomes in the endoplasmic reticulum (ER). (6) Virion assembly occurs at ER‐Golgi junctions in concurrence with (7) the formation of mature virions inside Golgi vesicles. (8) Post‐assembly, the infective virions are released via exocytosis or through cell lysis. The inset table shows possible intervention points for SARS‐CoV‐2 at multiple stages of its life cycle. These repurposed drugs intercept the SARS‐CoV‐2 infection at crucial points, including inhibiting the viral proteins or interfering with viral entry, translation of viral proteins, assembly of new virions, viral budding, etc. Many repurposed/experimental drugs also possess off‐target side‐effects contributing to drug‐induced cytotoxicity, local tissue damage, and systemic immunosuppression
Viral entry
Coronaviruses are pleomorphic, enveloped, positive‐sense, single‐stranded RNA viruses. 29 The viral membrane includes the envelope (E) protein, transmembrane (M) glycoprotein, and spike (S) glycoprotein. 30 Due to the surface spikes, coronaviruses appear like crowns. The viral life cycle within the host involves attachment, penetration, biosynthesis, maturation, and release. After the virus binds to receptors of the host cell (attachment), it enters through receptor‐mediated endocytosis or membrane fusion (penetration). The initial cells infected are usually those along the upper respiratory tract. The corona spikes bind to the angiotensin‐converting enzyme 2 (ACE2) receptors on the surface of upper respiratory tract cells, which can facilitate entry into the cells. The normal function of the ACE2 receptor, which includes decreasing inflammation, becomes affected when the virus binds. 31 , 32 This decrease in anti‐inflammatory response may exacerbate tissue damage. 33 , 34 Located on the cell surface and intracellularly, furin is a subtilisin‐like proprotein convertase and calcium‐dependent serine endoprotease that cleaves the spike protein to make it functional for priming and activation. 35 , 36 The spike protein is primed by proteases: transmembrane serine protease 2 (TMPRSS2) and a disintegrin and metalloprotease 17 (ADAM17). Subsequently, the virus uses those cells to produce more progenies for invasion of surrounding cells. 2 , 37
A computational study demonstrated that the Omicron variant had a greater affinity for ACE2 compared with the Delta variant because of numerous mutations in the SARS‐CoV‐2 receptor‐binding domain. 38 Preliminary research also showed that Omicron preferred infecting the upper airways rather than the lungs. 39 These characteristics may contribute to the higher transmissibility and lower disease severity of Omicron. 40 , 41 , 42 , 43
Males have been found to have elevated expression of ACE2 in some studies, although the relationship between sex and ACE2 levels is highly complex. 44 , 45 , 46 One hypothesis is that men have a higher prevalence of hypertension and heart failure, which may explain the higher ACE2 compared with women. 47 It has also been shown that soluble ACE2 (sACE2) levels are similar between males and females up to age 12 years. However, by age 15, sACE2 levels in males surpass those of females. 48 Consistent with that idea, one study demonstrated that a lower dose of ACE inhibitors is needed in females compared with males for optimal therapeutic effect. 49 More research will hopefully further elucidate the relationship between sex‐related differential expression of ACE2 and vulnerability to SARS‐CoV‐2 infection. 10
Innate immune response
Females usually mount stronger innate and adaptive immune responses against pathogens than their male counterparts, which could explain their lower COVID‐19 disease severity and mortality. 26 Early immune responses to the virus involve innate viral detection, interferon (IFN) production and inflammasome activation. 50 Although cross‐reactive adaptive immune cells have been detected in SARS‐CoV‐2 unexposed individuals, the disease outcome is largely thought to be shaped by initial innate immune responses. 51 , 52
Pattern recognition receptors (PRRs) such as toll‐like receptor 7 (TLR7) that sense single‐stranded RNA may play a major role in the initial innate immune response against RNA viruses such as SARS‐CoV‐2. 53 Women may be able to clear early SARS‐CoV‐2 infection better than men because TLR7 expression in innate immune cells can be upregulated by the female sex steroid oestrogen and, furthermore, TLR7 might escape X chromosome inactivation in some cells. 54 , 55 Escaping X‐inactivation allows the TLR7 gene to be expressed more highly in females, who have two copies of the X chromosome.
After detection by PRRs, two major arms of innate antiviral responses are activated to curb the spread of the virus. Specifically, viral replication and dissemination are restricted by type I and type III IFNs, and antiviral immune cells are recruited by cytokines and chemokines. In COVID‐19, the cytokine/chemokine response is stronger than the IFN response, possibly because SARS‐CoV‐2 can more effectively evade IFN responses. 56 , 57
Sex differences in early SARS‐CoV‐2 infection may be linked to differences in the IFN response. 58 TLR7 is a crucial sensor in plasmacytoid dendritic cells (pDCs) for the production of type I IFN. IFNα production from pDCs is greater in women than in men, with oestrogen modulating this effect. 59 , 60 , 61 , 62 , 63 Females with COVID‐19 have higher plasma concentrations of IFNα. 64 IFN regulatory factor 5 (IRF5), a critical transcription factor in IFN signalling, is also more highly expressed in pDCs of females, contributing to stronger type I IFN responses. 10 , 53 Besides these differences in IFN induction, in a group of severely ill COVID‐19 patients consisting mostly of older males, type I IFN signalling was found to be inhibited by autoantibodies. 65
Innate immune cells produce proinflammatory cytokines in response to viral infections. Elevated inflammatory cytokine levels have been associated with severe COVID‐19 pathology, and clinical studies in patients indicate sex differences. Interleukin 6 (IL‐6) production after viral infection is lower in females and is associated with better prognosis. 66 Though stronger cytokine responses are typically seen in females with viral infections, COVID‐19 is an exception. Instead, higher levels of innate proinflammatory cytokines (e.g., IL‐8 and IL‐18) are seen in males. 64 Elevated plasma proinflammatory cytokines and chemokines, especially IL‐6 and inflammasome‐associated IL‐1β and IL‐18, are seen in people with severe COVID‐19, culminating in cytokine storm. 67 , 68 An extraordinary amount of IL‐1 and tumour necrosis factor (TNF) release can lead to acute inflammation and death. 66 , 69 Males have been found to have higher serum levels of IL‐8, IL‐18 and C‐C motif chemokine ligand 5 (CCL5) than females, and a significant correlation has been found between elevated serum levels of IL‐8 and reduction in antiviral lymphocytes. There is also an association between systemic inflammation and lung involvement, such as infiltration by monocytes and neutrophils, and elevated neutrophil count is linked to worse clinical outcomes. 70 Research in female mice has shown not only lower SARS‐CoV‐1 viral titres, but also lower infiltration by monocytes and macrophages and production of cytokines, resulting in milder lung injury and lower mortality compared with males. 71
Adaptive immune response
Antibody‐based protection has been shown to be a key in our fight against SARS‐CoV‐2. Females usually have greater humoral responses to viral infection and vaccination, although they also show greater autoreactivity. 72 , 73 , 74 , 75 , 76 , 77 , 78 , 79 Multiple mechanisms, which may be mediated by oestrogen, help account for differences in antibody production between the sexes: germinal centre formation, selection against autoreactive B cells, somatic hypermutation enhancement, and epigenetic accessibility of B cell‐specific loci. 10
Similar to that of humoral responses, the role of sex‐based differences in T‐cell responses in COVID‐19 has also been emerging. Regulatory T‐cell (Treg) development, 80 , 81 , 82 , 83 lymphocyte subset distribution, 84 and quality of T‐cell responses are all affected by sex. 85 , 86 Higher oestradiol levels increase Tregs, 87 and women have a higher ratio of CD4/CD8 T cells. 88 Researchers did not identify a connection between T‐cell responses and human outcomes in the SARS outbreak. 89 However, CD4+ T cells may be involved based on murine experiments. 90 Low lymphocyte levels predict COVID‐19 disease progression and are associated with severe COVID‐19 disease. 91 , 92 , 93 Males with COVID‐19 have lower lymphocyte count and higher neutrophil‐to‐lymphocyte ratios and serum C‐reactive protein (CRP) concentrations compared with females. 94 Elderly females with early SARS‐CoV‐2 infection have more robust T‐cell activation than their male counterparts. Poorer outcomes are seen in males with COVID‐19, along with weak activation of T cells during early disease, but this is not seen in females. 64 More research is necessary to characterize sex differences in the role of T cells in acute infection and lung injury 95 as well as vaccine targets. 10
The epigenetic status of immune cells also differs based on sex. The epigenetic landscape of immune cells in males changes immensely from age 62 to 64, leading to faster immunosenescence. 24 This includes elevated expression of innate proinflammatory genes and decreased expression of adaptive immune system genes. Males exhibit a more notable decline in certain naïve T cells with ageing, 24 and only males exhibit a notable decrease in B cells after age 65. 77 On the other hand, females exhibit large epigenetic changes in immune cells about 5–6 years later than males. 57 Overexpression of immune genes on the X chromosome in T cells has been associated with T‐cell‐specific X‐chromosome epigenetic modifications and incomplete X‐inactivation. 96 , 97
In addition, females may be more efficient at clearing SARS‐CoV‐2, as studies demonstrated that the virus is detected for a longer period in males. 98 , 99 To assess the efficacy of immune responses, the viral clearance rate should be further evaluated. 10
Long‐term effects
Female sex is associated with long or post‐COVID syndrome, which involves symptoms that persist after recovering from COVID‐19. 100 , 101 , 102 , 103 In one study, fatigue was observed to be the most common sequela of post‐COVID syndrome and associated with higher IL‐6 levels and female sex. Dyspnoea was more likely in men and not associated with higher IL‐6 levels. 101 Sex‐specific autoimmune responses may influence the course of recovery from COVID‐19. 104 , 105 A study found that, after asymptomatic infection, the overall autoantibody (AAB) response was greater in women. However, after ‘at least mildly symptomatic infection,’ the ‘breadth and extent of AAB reactivity’ was greater in men. 105
CRUCIAL ROLE OF SEX HORMONES IN MODULATING THE IMMUNE RESPONSE TO SARS‐COV‐2 INFECTION
Androgens
The progression of COVID‐19 is likely associated with sex hormones (Table 1). Females have more oestrogen and progesterone, whilst males have more androgens, including testosterone and dihydrotestosterone (Figure 2). 106 Androgens may regulate furin. Studies suggest that the androgen receptor influences furin and other proprotein convertases in prostate cancer cells. 35 , 107 , 108 TMPRSS2, a gene regulated by testosterone, may be more highly expressed in men, 109 , 110 which may, in turn, account for the greater COVID‐19 severity in males. 111 , 112 The genetic expression of TMPRSS2 is regulated and can be increased by the androgen receptor. 113 As a result, SARS‐CoV‐2 entry into host cells can be affected by the expression of androgen receptors and TMPRSS2. Higher androgen receptor expression may increase the risk of more severe COVID‐19 disease. 114 This is supported by clinical evidence of an association between androgenic alopecia and severe COVID‐19. 115 Smoking has also been associated with higher expression of ACE2 and TMPRSS2, possibly through upregulating the androgen pathway. 116 , 117 , 118
TABLE 1.
Relationship between sex hormones and immune response to SARS‐CoV‐2 infection
| Sex hormones | Potential effect on the immune response to SARS‐CoV‐2 infection | References |
|---|---|---|
| Androgens |
|
35, 107, 108, 113, 119, 120, 137, 138, 139, 140, 143, 144 |
| Oestrogens |
|
36, 54, 55, 61, 62, 63, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160 |
| Progesterone |
|
169 |
Abbreviations: ACE, angiotensin‐converting enzyme. ADAM, a disintegrin and metalloprotease. CCR, C‐C motif chemokine receptor. CCL, C‐C motif chemokine ligand, CXCR, C‐X‐C motif chemokine receptor. COVID‐19, coronavirus disease 2019. DPP, dipeptidyl peptidase. IFN, interferon. IL, interleukin. NF‐κB, nuclear factor kappa B. NLRP, nucleotide‐binding oligomerization domain‐like receptor, pyrin domain containing. pDCs, plasmacytoid dendritic cells. RAGE, receptor for advanced glycation end products. SARS‐CoV‐2, severe acute respiratory syndrome coronavirus 2. Th, T‐helper cell. TLR, toll‐like receptor. TMPRSS, transmembrane serine protease. TNF, tumour necrosis factor. Treg, regulatory T cell.
FIGURE 2.
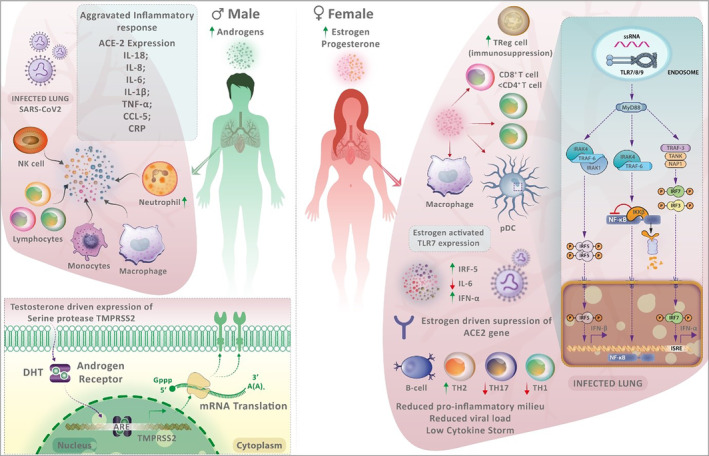
The sex differences in the immune response to severe acute respiratory syndrome coronavirus 2 (SARS‐CoV‐2) infection. Males have more testosterone and dihydrotestosterone, whilst females have higher oestrogen and progesterone levels. Men experience aggravated immune responses to SARS‐CoV‐2 infection due to various factors. The expression of transmembrane serine protease 2 (TMPRSS2), which facilitates viral entry, is regulated by androgen receptors and higher in males. Males with coronavirus disease 2019 (COVID‐19) have higher neutrophil‐to‐lymphocyte ratios, lower lymphocyte count and greater serum C‐reactive protein levels. Older males have decreased naïve T and B cells. Males experience hyperinflammation and cytokine storms, which translate into increased COVID‐19 severity. In females, SARS‐CoV‐2 infection is better controlled due to the efficient sensing of the viral genome by endosomally expressed TLR7 in immune cells. TLR7 expression is enhanced by higher levels of oestrogen in females. Such intracellular detection of the viral genome in immune cells amplifies the production of type I interferon (IFN), which confers antiviral immunity. In plasmacytoid dendritic cells of females, IRF5 expression is higher, which may explain the greater production of type I IFN in females. Additionally, oestradiol promotes regulatory T cells, and women also have increased CD4+:CD8+ T‐cell ratios, which may have an impact on COVID‐19 progression. Females exhibit more robust adaptive immunity compared with males, with pronounced effects on reducing dysregulated inflammation.
Androgen‐related treatments may provide further insight into the role of androgens in COVID‐19 progression. One study showed a lower risk of COVID‐19 infection amongst prostate cancer patients who were on androgen deprivation therapy (ADT) compared with those who were not. 119 It is thought that virus binding to the cell can be inhibited by ADT. Fewer activated androgen receptors to upregulate TMPRSS2 would be expected in patients on ADT, decreasing the risk of SARS‐CoV‐2 infection. 120 However, multiple recent studies observed that ADT did not improve COVID‐19 outcomes, including infection risk, ICU admission, hospitalization, and mortality. 121 , 122 , 123 , 124 , 125 , 126 Another study showed that prostate cancer patients with COVID‐19 had higher rates of hospitalization and mortality compared with non‐prostate genitourinary cancer patients with COVID‐19. 127
TMPRSS2 expression and SARS‐CoV‐2 entry in human lung cells have been reduced by antiandrogens. 128 , 129 , 130 Clinical trial results on antiandrogens have been mixed. A randomized controlled trial (RCT) treated COVID‐19 patients undergoing nitazoxanide and azithromycin therapy with dutasteride and found decreased viral shedding, inflammatory markers and time‐to‐remission compared with placebo. 131 Another RCT found that proxalutamide decreased the 30‐day hospitalization rate and risk ratio amongst men with COVID‐19. 132 In contrast, a third RCT determined that COVID‐19 patients treated with enzalutamide had longer hospitalization. 126
Semen samples from males with COVID‐19 have been shown to contain SARS‐CoV‐2 genetic material. 133 ACE2 and TMPRSS2 are also expressed in the testes. 134 However, the expression of TMPRSS2 in lung tissue was not significantly different between sexes, according to one study. 135 Currently, it remains unclear whether pulmonary expression of TMPRSS2 may be influenced by androgens during SARS‐CoV‐2 infection. 136 Whether SARS‐CoV‐2 viral load can be influenced by the level of TMPRSS2 expression is also uncertain.
Additionally, androgens have numerous immunosuppressive effects. They can suppress thymic function and T‐cell development, have direct or indirect effects (via antigen‐presenting cells) on T cells to decrease proinflammatory cytokine release (e.g., IFNγ and TNF) or increase anti‐inflammatory cytokine release (e.g., IL‐4 and IL‐10), reduce T helper 1 (Th1) and T helper 17 (Th17) cell differentiation, induce Treg differentiation and regulate B‐cell development and humoral immune responses. 137 , 138 , 139 , 140 Testosterone‐dependent Gr1+ cells in male autoimmune BWF1 mice inhibited T follicular helper cell development, which, in turn, controlled the formation of germinal centres and differentiation of plasma cells. 141 Furthermore, higher testosterone levels are associated with lower neutralizing antibody responses after influenza vaccination. 142
Paradoxically, these immunosuppressive effects of testosterone might be beneficial to overcome the heightened inflammatory environment that predisposes to severe COVID‐19. Recent research has revealed that males with COVID‐19 have lower testosterone levels and higher luteinizing hormone (LH) levels than healthy subjects. 143 Moreover, there is a negative association between testosterone/LH ratio and serum CRP levels. 143 Another study found a negative association between total testosterone levels and biochemical markers of COVID‐19 severity. 144 In a prospective cohort study of COVID‐19 patients, lower testosterone concentrations were associated with higher concentrations of IL‐6, CRP, IL‐1 receptor antagonist, hepatocyte growth factor, and IFNγ‐inducible protein 10. 145 In young men, there is a negative association between androgen levels and inflammatory markers. 146 Therefore, testosterone deficiency appears to be associated with higher levels of proinflammatory cytokines, whilst testosterone can decrease these cytokines. 147
The available evidence on the role of androgens in COVID‐19 leaves many questions unanswered. It appears that both high and low androgen levels can be associated with poor COVID‐19 outcomes. 35 Additional research is needed to investigate the relationship.
Oestrogens
A different line of reasoning argues that oestrogen promotes beneficial immune system activation, which may protect against severe COVID‐19. 148 Oestrogen receptors are expressed on cells of the immune system and known to impact immune‐related gene expression. In female mouse models, oestrogen decreased the severity of influenza virus infection, and mortality after SARS‐CoV‐1 infection increased when oestrogen was suppressed. 71 , 149 Amongst hospitalized patients, a negative association between oestradiol levels and COVID‐19 severity was found. 150 In addition to androgens, there is evidence that oestrogens also regulate furin. In one study, human breast cancer cells were transfected with the cDNAs of PC1 and furin, which are both proprotein convertases. Lower oestradiol concentration was associated with slower growth in the transfected cells relative to the wild‐type cells. In mice, tamoxifen therapy led to slower regression of tumours from transfected cells compared with those from wild‐type cells. These results suggest that overexpressing proprotein convertases can lead to higher dependence on oestrogen and resistance to antioestrogens. 151 Another study revealed that oestradiol and oestrogen receptor agonists regulated the expression of furin in peripheral blood leucocytes. 152 Oestrogen regulates three other molecules involved in SARS‐CoV‐2 entry into host cells: ACE2, TMPRSS2, and ADAM17. 153 , 154 , 155 Oestrogen can also suppress dipeptidyl peptidase 4 (DPP4), helping prevent another potential means of entry for SARS‐CoV‐2. 154 Moreover, oestradiol can activate adenosine receptors (part of the DPP4/adenosine pathway), which may have anti‐inflammatory effects. 154
Several other pathways that contribute to heightened inflammation in COVID‐19 are regulated by oestrogen. Oestrogen inhibits nucleotide‐binding oligomerization domain‐like receptor, pyrin domain containing 3 (NLRP3) inflammasome activation and may regulate the receptor for advanced glycation end products (RAGE) pathway to reduce lung injury. 154 Oestrogen can also decrease C‐C motif chemokine receptor 2 (CCR2), CCL2 and C‐X‐C motif chemokine receptor 3 (CXCR3), inhibit recruitment of cells of the innate immune system (e.g., monocytes and neutrophils) and regulate eosinophils. 156 , 157 , 158 In addition, oestrogen activates anti‐inflammatory cytokines (IL‐4, IL‐10), inhibits the nuclear factor kappa B (NF‐κB) pathway and decreases the release of inflammatory cytokines (IL‐1β, IL‐6, IL‐17, and TNF), thereby reducing inflammation in innate immune responses. 154 , 159 , 160 The hormone can inhibit Th1 cells, promote T helper 2 (Th2) and Treg cells, affect B‐cell development, and stimulate plasma cells to produce antibodies. 158 Oestrogen also participates in the regulation of pDCs, which are involved in the antiviral immune response by producing IFNα. 61 In response to viruses, females produce much more IFNα than males, leading to a stronger priming of the adaptive immune response. 62 , 63 Oestrogen can increase nitric oxide for vasodilation of vessels and decrease platelet aggregation to prevent thrombosis. Through these various mechanisms, oestrogen may have therapeutic potential for managing COVID‐19. 161
Given the important role of oestrogen in the immune system, there has been increased interest in and concerns about interventions that modulate the effects of oestrogen. 162 , 163 Researchers are currently examining the effect of selective oestrogen receptor modulators (SERMs) on COVID‐19. 164 , 165 , 166 An ongoing RCT is investigating the efficacy of an oestradiol and progesterone therapy for reducing COVID‐19 disease severity in hospitalized adults. 167
Progesterone
Progesterone is another hormone that may influence COVID‐19 disease progression. In hypoxemic men hospitalized with COVID‐19, short‐term subcutaneous progesterone improved clinical outcomes. 168 The anti‐inflammatory effects of progesterone may decrease the risk of hyperinflammation and cytokine storm. 169 Progesterone has been shown to increase Treg differentiation, enhance IFNα pathways, promote lung repair by inducing amphiregulin, reduce T helper cell 17 (Th17) responses and disrupt endocytic pathways used by viruses to enter host cells. 169 Taken together, the current evidence suggests that the effects of sex hormones on the immune system are highly complex and context dependent. Further investigation into the underlying pathways targeted by these sex steroids and their clinical correlation will allow us to fine‐tune our treatment approaches across the sexes.
SEX DIFFERENCES IN THE IMMUNE RESPONSE TO VACCINES
Vaccines are amongst the most critical components in our arsenal to control the spread of a rapidly spreading pandemic. They induce immunological memory using components of the pathogen, such that when exposed to the actual pathogen, immunological memory can be reactivated to rapidly mount a stronger immune response to eliminate the pathogen. Vaccine efficacy is affected by various factors including age and sex. Females generally show stronger humoral and cell‐mediated immune responses to vaccines. 53 , 170 Data from a randomized, prospective, single‐blind investigation on humoral immune responses have shown that healthy women (age 18–64) produce a stronger protective antibody response to a trivalent inactivated influenza vaccine than their male counterparts. 171 Antibody responses of females to a half dose of influenza vaccine were comparable to antibody responses of males to the full dose. Moreover, a positive association was found between levels of antibodies to the monovalent 2009 H1N1 vaccine and circulating oestradiol concentrations in females. 172 These findings explain why sex differences should be taken into account as a biological variable for adjusting sex‐personalized vaccine dosage and considering vaccine efficacy. Because adverse events are also more common in women, 173 the increased immune response to vaccines should be balanced with vaccine safety. 170
Similar sex‐related factors that influence the immune response to pathogens can potentially affect the immune response to vaccines. Hormonal differences, including oestrogen and androgen levels, may lead to discrepancies in the vaccine response. 174 In addition, X chromosomes have been proposed as one possible factor for the stronger and sometimes longer lasting immunological responses in women, since females have two, whilst males have only one. Higher expression of immune‐related molecules in females could be due to the X chromosome having abundant genes that produce immunological components (e.g., TLR7, cytokines, and chemokines) as well as regulatory molecules such as micro‐RNAs. 55 , 175 , 176 , 177 Ageing is another factor in immune response modulation that should not be overlooked. 178 As discussed earlier, males exhibit faster immunosenescence than females. 24
Females and males also experience different pathogen‐specific and non‐specific effects (NSEs) of vaccines. 179 NSEs are effects beyond those that protect against the targeted diseases. 180 They can be beneficial for certain vaccines and harmful for others and generally affect females more than males. 181 , 182 , 183 , 184 Many factors are involved in NSEs of vaccines, including innate and adaptive immune responses, heterologous immunity, and the effects of other vaccines. Potential reasons for sex differences in NSEs include the effect of X‐linked (and Y‐linked) genes, sex hormones, micro‐RNAs, and sex differences in the microbiota. 179
A CDC report showed that of 13.8 million doses of COVID‐19 vaccine administered in the United States from mid‐December 2020 to mid‐January 2021, women received 61% of the doses but accounted for 79% of the reported adverse events. 185 A systematic review and meta‐analysis revealed that COVID‐19 vaccination had higher efficacy in preventing COVID‐19 disease in men compared with women. 180 , 186 Further research is necessary to determine the role of sex differences in the immune response to the COVID‐19 vaccine.
CONCLUDING REMARKS
The purpose of this review was to explore the biological mechanisms driving sex differences in the immune response to SARS‐CoV‐2 infection and to help elucidate the complex relationship between sex and immune regulation in the context of COVID‐19. As discussed in this review, there are sex‐specific differences at various stages of SARS‐CoV‐2 infection, including viral entry, the innate immune response and the adaptive immune response. Sex hormones, including androgens, oestrogens and progesterone, may have a major role in regulating the immune system to combat the virus. Studies have also identified potential differences between females and males in the vaccine immune response. These sex‐specific differences at the cellular and molecular levels may contribute to the dissimilar clinical outcomes between females and males. Because sex is a complex, non‐binary biological trait that involves the sex chromosomes, sex hormones, and reproductive organs and because sex‐related genes and sex hormones affect the immune system—and thus disease progression and outcome—in many known and potentially unknown ways, sex should be considered in basic, translational and clinical COVID‐19 research as a biological factor that may influence therapeutic response. Further enquiry into the role of biological sex in COVID‐19 could help enhance the specificity of preventive and therapeutic strategies and improve health and well‐being.
AUTHOR CONTRIBUTIONS
J.Q.H, M.R.S, B.B and S.Z‐ Conceptualization and design, Writing Original draft. J.Q.H, M.R.S, B.B, T.S, R.E, P.C, V.S, L.B, G.H and S.Z, Writing – Review & Editing. All authors read and approved the final version of the manuscript.
FUNDING INFORMATION
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
CONFLICT OF INTEREST
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
PERMISSION TO REPRODUCE
Not applicable.
ACKNOWLEDGEMENTS
This research was supported by the Seed Funding program from the Office of the Provost at the University of Massachusetts Dartmouth, USA.
Ho JQ, Sepand MR, Bigdelou B, Shekarian T, Esfandyarpour R, Chauhan P, et al. The immune response to COVID‐19: Does sex matter? Immunology. 2022. 10.1111/imm.13487
Funding information University of Massachusetts Dartmouth
DATA AVAILABILITY STATEMENT
Not applicable.
REFERENCES
- 1. Peckham H, de Gruijter NM, Raine C, Radziszewska A, Ciurtin C, Wedderburn LR, et al. Male sex identified by global COVID‐19 meta‐analysis as a risk factor for death and ITU admission. Nat Commun. 2020;11(1):6317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Lipsky MS, Hung M. Men and COVID‐19: a pathophysiologic review. Am J Mens Health. 2020;14(5):1557988320954021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Gomez JMD, Du‐Fay‐de‐Lavallaz JM, Fugar S, Sarau A, Simmons JA, Clark B, et al. Sex differences in COVID‐19 hospitalization and mortality. J Womens Health (Larchmt). 2021;30(5):646–53. [DOI] [PubMed] [Google Scholar]
- 4. Dudley JP, Lee NT. Disparities in age‐specific morbidity and mortality from SARS‐CoV‐2 in China and the Republic of Korea. Clin Infect Dis. 2020;71(15):863–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Li LQ, Huang T, Wang YQ, Wang ZP, Liang Y, Huang TB, et al. COVID‐19 patients' clinical characteristics, discharge rate, and fatality rate of meta‐analysis. J Med Virol. 2020;92(6):577–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Grasselli G, Zangrillo A, Zanella A, Antonelli M, Cabrini L, Castelli A, et al. Baseline characteristics and outcomes of 1591 patients infected with SARS‐CoV‐2 admitted to ICUs of the Lombardy region, Italy. JAMA. 2020;323(16):1574–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Palmieri L, Vanacore N, Donfrancesco C, Lo Noce C, Canevelli M, Punzo O, et al. Clinical characteristics of hospitalized individuals dying with COVID‐19 by age Group in Italy. J Gerontol A Biol Sci Med Sci. 2020;75(9):1796–800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Biswas M, Rahaman S, Biswas TK, Haque Z, Ibrahim B. Association of sex, age, and comorbidities with mortality in COVID‐19 patients: a systematic review and meta‐analysis. Intervirology. 2020;64:1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Griffith DM, Sharma G, Holliday CS, Enyia OK, Valliere M, Semlow AR, et al. Men and COVID‐19: a biopsychosocial approach to understanding sex differences in mortality and recommendations for practice and policy interventions. Prev Chronic Dis. 2020;17:E63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Scully EP, Haverfield J, Ursin RL, Tannenbaum C, Klein SL. Considering how biological sex impacts immune responses and COVID‐19 outcomes. Nat Rev Immunol. 2020;20(7):442–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Villar J, Ariff S, Gunier RB, Thiruvengadam R, Rauch S, Kholin A, et al. Maternal and neonatal morbidity and mortality among pregnant women with and without COVID‐19 infection: the INTERCOVID multinational cohort study. JAMA Pediatr. 2021;175(8):817–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Allotey J, Stallings E, Bonet M, Yap M, Chatterjee S, Kew T, et al. Clinical manifestations, risk factors, and maternal and perinatal outcomes of coronavirus disease 2019 in pregnancy: living systematic review and meta‐analysis. BMJ. 2020;370:m3320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Zambrano LD, Ellington S, Strid P, Galang RR, Oduyebo T, Tong VT, et al. Update: characteristics of symptomatic women of reproductive age with laboratory‐confirmed SARS‐CoV‐2 infection by pregnancy status ‐ United States, January 22‐October 3, 2020. MMWR Morb Mortal Wkly Rep. 2020;69(44):1641–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. DeBolt CA, Bianco A, Limaye MA, Silverstein J, Penfield CA, Roman AS, et al. Pregnant women with severe or critical coronavirus disease 2019 have increased composite morbidity compared with nonpregnant matched controls. Am J Obstet Gynecol. 2021;224(5):510 e1–e12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Lokken EM, Huebner EM, Taylor GG, Hendrickson S, Vanderhoeven J, Kachikis A, et al. Disease severity, pregnancy outcomes, and maternal deaths among pregnant patients with severe acute respiratory syndrome coronavirus 2 infection in Washington state. Am J Obstet Gynecol. 2021;225(1):77 e1–e14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Alyousefi NA. An oral combined contraceptive user with elevated D‐dimer post COVID‐19: a case report. BMC Womens Health. 2021;21(1):320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Spratt DI, Buchsbaum RJ. COVID‐19 and hypercoagulability: potential impact on management with oral contraceptives, estrogen therapy and pregnancy. Endocrinology. 2020;161(12):bqaa121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Panda SR, Meena M. Contraceptive advice during COVID‐19 pandemic and the overlapping threat of venous thromboembolism. Eur J Obstet Gynecol Reprod Biol. 2021;260:232–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Karlberg J, Chong DS, Lai WY. Do men have a higher case fatality rate of severe acute respiratory syndrome than women do? Am J Epidemiol. 2004;159(3):229–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Alghamdi IG, Hussain II, Almalki SS, Alghamdi MS, Alghamdi MM, El‐Sheemy MA. The pattern of Middle East respiratory syndrome coronavirus in Saudi Arabia: a descriptive epidemiological analysis of data from the Saudi Ministry of Health. Int J Gen Med. 2014;7:417–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Ziegler S, Altfeld M. Sex differences in HIV‐1‐mediated immunopathology. Curr Opin HIV AIDS. 2016;11(2):209–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Guery JC. Sex differences in primary HIV infection: revisiting the role of TLR7‐driven type 1 IFN production by plasmacytoid dendritic cells in women. Front Immunol. 2021;12:729233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Ruggieri A, Gagliardi MC, Anticoli S. Sex‐dependent outcome of hepatitis B and C viruses infections: synergy of sex hormones and immune responses? Front Immunol. 2018;9:2302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Takahashi T, Iwasaki A. Sex differences in immune responses. Science. 2021;371(6527):347–8. [DOI] [PubMed] [Google Scholar]
- 25. Sharma G, Volgman AS, Michos ED. Sex differences in mortality from COVID‐19 pandemic: are men vulnerable and women protected? JACC Case Rep. 2020;2(9):1407–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Pradhan A, Olsson PE. Sex differences in severity and mortality from COVID‐19: are males more vulnerable? Biol Sex Differ. 2020;11(1):53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Schmiedel BJ, Singh D, Madrigal A, Valdovino‐Gonzalez AG, White BM, Zapardiel‐Gonzalo J, et al. Impact of genetic polymorphisms on human immune cell gene expression. Cell. 2018;175(6):1701–15 e16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Dumanski JP, Halvardson J, Davies H, Rychlicka‐Buniowska E, Mattisson J, Moghadam BT, et al. Immune cells lacking Y chromosome show dysregulation of autosomal gene expression. Cell Mol Life Sci. 2021;78(8):4019–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Perlman S, Netland J. Coronaviruses post‐SARS: update on replication and pathogenesis. Nat Rev Microbiol. 2009;7(6):439–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Neuman BW, Adair BD, Yoshioka C, Quispe JD, Orca G, Kuhn P, et al. Supramolecular architecture of severe acute respiratory syndrome coronavirus revealed by electron cryomicroscopy. J Virol. 2006;80(16):7918–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Rodrigues Prestes TR, Rocha NP, Miranda AS, Teixeira AL, Simoes ESAC. The anti‐inflammatory potential of ACE2/angiotensin‐(1‐7)/mas receptor Axis: evidence from basic and clinical research. Curr Drug Targets. 2017;18(11):1301–13. [DOI] [PubMed] [Google Scholar]
- 32. Abassi Z, Higazi AAR, Kinaneh S, Armaly Z, Skorecki K, Heyman SN. ACE2, COVID‐19 infection, inflammation, and coagulopathy: missing pieces in the puzzle. Front Physiol. 2020;11:574753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Cascella M, Rajnik M, Aleem A, Dulebohn SC, Di Napoli R. Features, evaluation, and treatment of coronavirus (COVID‐19). Treasure Island (FL): StatPearls; 2021. [PubMed] [Google Scholar]
- 34. Iwasaki M, Saito J, Zhao H, Sakamoto A, Hirota K, Ma D. Inflammation triggered by SARS‐CoV‐2 and ACE2 augment drives multiple organ failure of severe COVID‐19: molecular mechanisms and implications. Inflammation. 2021;44(1):13–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Younis JS, Skorecki K, Abassi Z. The double edge sword of testosterone's role in the COVID‐19 pandemic. Front Endocrinol (Lausanne). 2021;12:607179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Mateus D, Sebastiao AI, Carrascal MA, Carmo AD, Matos AM, Cruz MT. Crosstalk between estrogen, dendritic cells, and SARS‐CoV‐2 infection. Rev Med Virol. 2021;e2290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Hoffmann M, Kleine‐Weber H, Schroeder S, Kruger N, Herrler T, Erichsen S, et al. SARS‐CoV‐2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell. 2020;181(2):271–80 e8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Kumar S, Thambiraja TS, Karuppanan K, Subramaniam G. Omicron and Delta variant of SARS‐CoV‐2: a comparative computational study of spike protein. J Med Virol. 2022;94(4):1641–9. [DOI] [PubMed] [Google Scholar]
- 39. Diamond M, Halfmann P, Maemura T, Iwatsuki‐Horimoto K, Iida S, Kiso M, et al. The SARS‐CoV‐2 B.1.1.529 omicron virus causes attenuated infection and disease in mice and hamsters. Res Sq. 2021. [Google Scholar]
- 40. Araf Y, Akter F, Tang YD, Fatemi R, Parvez MSA, Zheng C, et al. Omicron variant of SARS‐CoV‐2: genomics, transmissibility, and responses to current COVID‐19 vaccines. J Med Virol. 2022;94:1825–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Abdullah F, Myers J, Basu D, Tintinger G, Ueckermann V, Mathebula M, et al. Decreased severity of disease during the first global omicron variant covid‐19 outbreak in a large hospital in Tshwane, South Africa. Int J Infect Dis. 2021;116:38–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Wolter N, Jassat W, Walaza S, Welch R, Moultrie H, Groome M, et al. Early assessment of the clinical severity of the SARS‐CoV‐2 omicron variant in South Africa: a data linkage study. Lancet. 2022;399(10323):437–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Iuliano AD, Brunkard JM, Boehmer TK, Peterson E, Adjei S, Binder AM, et al. Trends in disease severity and health care utilization during the early omicron variant period compared with previous SARS‐CoV‐2 high transmission periods ‐ United States, December 2020‐January 2022. MMWR Morb Mortal Wkly Rep. 2022;71(4):146–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Sama IE, Ravera A, Santema BT, van Goor H, Ter Maaten JM, Cleland JGF, et al. Circulating plasma concentrations of angiotensin‐converting enzyme 2 in men and women with heart failure and effects of renin‐angiotensin‐aldosterone inhibitors. Eur Heart J. 2020;41(19):1810–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Zhao Y, Zhao Z, Wang Y, Zhou Y, Ma Y, Zuo W. Single‐cell RNA expression profiling of ACE2, the receptor of SARS‐CoV‐2. Am J Respir Crit Care Med. 2020;202(5):756–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Viveiros A, Rasmuson J, Vu J, Mulvagh SL, Yip CYY, Norris CM, et al. Sex differences in COVID‐19: candidate pathways, genetics of ACE2, and sex hormones. Am J Physiol Heart Circ Physiol. 2021;320(1):H296–304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Salah HM, Mehta JL. Hypothesis: sex‐related differences in ACE2 activity may contribute to higher mortality in men versus women with COVID‐19. J Cardiovasc Pharmacol Ther. 2021;26(2):114–8. [DOI] [PubMed] [Google Scholar]
- 48. Sward P, Edsfeldt A, Reepalu A, Jehpsson L, Rosengren BE, Karlsson MK. Age and sex differences in soluble ACE2 may give insights for COVID‐19. Crit Care. 2020;24(1):221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Santema BT, Ouwerkerk W, Tromp J, Sama IE, Ravera A, Regitz‐Zagrosek V, et al. Identifying optimal doses of heart failure medications in men compared with women: a prospective, observational, cohort study. Lancet. 2019;394(10205):1254–63. [DOI] [PubMed] [Google Scholar]
- 50. Iwasaki A. A virological view of innate immune recognition. Annu Rev Microbiol. 2012;66:177–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Loyal L, Braun J, Henze L, Kruse B, Dingeldey M, Reimer U, et al. Cross‐reactive CD4(+) T cells enhance SARS‐CoV‐2 immune responses upon infection and vaccination. Science. 2021;374(6564):eabh1823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Song G, He WT, Callaghan S, Anzanello F, Huang D, Ricketts J, et al. Cross‐reactive serum and memory B‐cell responses to spike protein in SARS‐CoV‐2 and endemic coronavirus infection. Nat Commun. 2021;12(1):2938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Klein SL, Flanagan KL. Sex differences in immune responses. Nat Rev Immunol. 2016;16(10):626–38. [DOI] [PubMed] [Google Scholar]
- 54. Ziegler SM, Altfeld M. Human immunodeficiency virus 1 and type I interferons‐where sex makes a difference. Front Immunol. 2017;8:1224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Souyris M, Cenac C, Azar P, Daviaud D, Canivet A, Grunenwald S, et al. TLR7 escapes X chromosome inactivation in immune cells. Sci Immunol. 2018;3(19):eaap8855. [DOI] [PubMed] [Google Scholar]
- 56. Blanco‐Melo D, Nilsson‐Payant BE, Liu WC, Uhl S, Hoagland D, Moller R, et al. Imbalanced host response to SARS‐CoV‐2 drives development of COVID‐19. Cell. 2020;181(5):1036–45 e9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Nish S, Medzhitov R. Host defense pathways: role of redundancy and compensation in infectious disease phenotypes. Immunity. 2011;34(5):629–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Sallard E, Lescure FX, Yazdanpanah Y, Mentre F, Peiffer‐Smadja N. Type 1 interferons as a potential treatment against COVID‐19. Antivir Res. 2020;178:104791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Griesbeck M, Ziegler S, Laffont S, Smith N, Chauveau L, Tomezsko P, et al. Sex differences in plasmacytoid dendritic cell levels of IRF5 drive higher IFN‐alpha production in women. J Immunol. 2015;195(11):5327–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Laffont S, Rouquie N, Azar P, Seillet C, Plumas J, Aspord C, et al. X‐chromosome complement and estrogen receptor signaling independently contribute to the enhanced TLR7‐mediated IFN‐alpha production of plasmacytoid dendritic cells from women. J Immunol. 2014;193(11):5444–52. [DOI] [PubMed] [Google Scholar]
- 61. Seillet C, Laffont S, Tremollieres F, Rouquie N, Ribot C, Arnal JF, et al. The TLR‐mediated response of plasmacytoid dendritic cells is positively regulated by estradiol in vivo through cell‐intrinsic estrogen receptor alpha signaling. Blood. 2012;119(2):454–64. [DOI] [PubMed] [Google Scholar]
- 62. Berghofer B, Frommer T, Haley G, Fink L, Bein G, Hackstein H. TLR7 ligands induce higher IFN‐alpha production in females. J Immunol. 2006;177(4):2088–96. [DOI] [PubMed] [Google Scholar]
- 63. Meier A, Chang JJ, Chan ES, Pollard RB, Sidhu HK, Kulkarni S, et al. Sex differences in the toll‐like receptor‐mediated response of plasmacytoid dendritic cells to HIV‐1. Nat Med. 2009;15(8):955–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Takahashi T, Ellingson MK, Wong P, Israelow B, Lucas C, Klein J, et al. Sex differences in immune responses that underlie COVID‐19 disease outcomes. Nature. 2020;588(7837):315–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Bastard P, Rosen LB, Zhang Q, Michailidis E, Hoffmann HH, Zhang Y, et al. Autoantibodies against type I IFNs in patients with life‐threatening COVID‐19. Science. 2020;370(6515):eabd4585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Conti P, Younes A. Coronavirus COV‐19/SARS‐CoV‐2 affects women less than men: clinical response to viral infection. J Biol Regul Homeost Agents. 2020;34(2):339–43. [DOI] [PubMed] [Google Scholar]
- 67. Tan M, Liu Y, Zhou R, Deng X, Li F, Liang K, et al. Immunopathological characteristics of coronavirus disease 2019 cases in Guangzhou, China. Immunology. 2020;160(3):261–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Lucas C, Wong P, Klein J, Castro TBR, Silva J, Sundaram M, et al. Longitudinal analyses reveal immunological misfiring in severe COVID‐19. Nature. 2020;584(7821):463–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Seki M, Kohno S, Newstead MW, Zeng X, Bhan U, Lukacs NW, et al. Critical role of IL‐1 receptor‐associated kinase‐M in regulating chemokine‐dependent deleterious inflammation in murine influenza pneumonia. J Immunol. 2010;184(3):1410–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Wang J, Li Q, Yin Y, Zhang Y, Cao Y, Lin X, et al. Excessive neutrophils and neutrophil extracellular traps in COVID‐19. Front Immunol. 2020;11:2063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Channappanavar R, Fett C, Mack M, Ten Eyck PP, Meyerholz DK, Perlman S. Sex‐based differences in susceptibility to severe acute respiratory syndrome coronavirus infection. J Immunol. 2017;198(10):4046–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Bynoe MS, Grimaldi CM, Diamond B. Estrogen up‐regulates Bcl‐2 and blocks tolerance induction of naive B cells. Proc Natl Acad Sci U S A. 2000;97(6):2703–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Grimaldi CM, Cleary J, Dagtas AS, Moussai D, Diamond B. Estrogen alters thresholds for B cell apoptosis and activation. J Clin Invest. 2002;109(12):1625–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Grimaldi CM, Jeganathan V, Diamond B. Hormonal regulation of B cell development: 17 beta‐estradiol impairs negative selection of high‐affinity DNA‐reactive B cells at more than one developmental checkpoint. J Immunol. 2006;176(5):2703–10. [DOI] [PubMed] [Google Scholar]
- 75. Hill L, Jeganathan V, Chinnasamy P, Grimaldi C, Diamond B. Differential roles of estrogen receptors alpha and beta in control of B‐cell maturation and selection. Mol Med. 2011;17(3–4):211–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Pauklin S, Sernandez IV, Bachmann G, Ramiro AR, Petersen‐Mahrt SK. Estrogen directly activates AID transcription and function. J Exp Med. 2009;206(1):99–111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Marquez EJ, Chung CH, Marches R, Rossi RJ, Nehar‐Belaid D, Eroglu A, et al. Sexual‐dimorphism in human immune system aging. Nat Commun. 2020;11(1):751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Fink AL, Klein SL. The evolution of greater humoral immunity in females than males: implications for vaccine efficacy. Curr Opin Physio. 2018;6:16–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Sakiani S, Olsen NJ, Kovacs WJ. Gonadal steroids and humoral immunity. Nat Rev Endocrinol. 2013;9(1):56–62. [DOI] [PubMed] [Google Scholar]
- 80. Dragin N, Bismuth J, Cizeron‐Clairac G, Biferi MG, Berthault C, Serraf A, et al. Estrogen‐mediated downregulation of AIRE influences sexual dimorphism in autoimmune diseases. J Clin Invest. 2016;126(4):1525–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Polanczyk MJ, Carson BD, Subramanian S, Afentoulis M, Vandenbark AA, Ziegler SF, et al. Cutting edge: estrogen drives expansion of the CD4+CD25+ regulatory T cell compartment. J Immunol. 2004;173(4):2227–30. [DOI] [PubMed] [Google Scholar]
- 82. Tai P, Wang J, Jin H, Song X, Yan J, Kang Y, et al. Induction of regulatory T cells by physiological level estrogen. J Cell Physiol. 2008;214(2):456–64. [DOI] [PubMed] [Google Scholar]
- 83. Zhu ML, Bakhru P, Conley B, Nelson JS, Free M, Martin A, et al. Sex bias in CNS autoimmune disease mediated by androgen control of autoimmune regulator. Nat Commun. 2016;7:11350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Page ST, Plymate SR, Bremner WJ, Matsumoto AM, Hess DL, Lin DW, et al. Effect of medical castration on CD4+ CD25+ T cells, CD8+ T cell IFN‐gamma expression, and NK cells: a physiological role for testosterone and/or its metabolites. Am J Physiol Endocrinol Metab. 2006;290(5):E856–63. [DOI] [PubMed] [Google Scholar]
- 85. Hewagama A, Patel D, Yarlagadda S, Strickland FM, Richardson BC. Stronger inflammatory/cytotoxic T‐cell response in women identified by microarray analysis. Genes Immun. 2009;10(5):509–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Yee Mon KJ, Goldsmith E, Watson NB, Wang J, Smith NL, Rudd BD. Differential sensitivity to IL‐12 drives sex‐specific differences in the CD8+ T cell response to infection. Immunohorizons. 2019;3(4):121–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Arruvito L, Sanz M, Banham AH, Fainboim L. Expansion of CD4+CD25+and FOXP3+ regulatory T cells during the follicular phase of the menstrual cycle: implications for human reproduction. J Immunol. 2007;178(4):2572–8. [DOI] [PubMed] [Google Scholar]
- 88. Amadori A, Zamarchi R, De Silvestro G, Forza G, Cavatton G, Danieli GA, et al. Genetic control of the CD4/CD8 T‐cell ratio in humans. Nat Med. 1995;1(12):1279–83. [DOI] [PubMed] [Google Scholar]
- 89. Janice Oh HL, Ken‐En Gan S, Bertoletti A, Tan YJ. Understanding the T cell immune response in SARS coronavirus infection. Emerg Microbes Infect. 2012;1(9):e23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Chen J, Lau YF, Lamirande EW, Paddock CD, Bartlett JH, Zaki SR, et al. Cellular immune responses to severe acute respiratory syndrome coronavirus (SARS‐CoV) infection in senescent BALB/c mice: CD4+ T cells are important in control of SARS‐CoV infection. J Virol. 2010;84(3):1289–301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Yang X, Yu Y, Xu J, Shu H, Xia J, Liu H, et al. Clinical course and outcomes of critically ill patients with SARS‐CoV‐2 pneumonia in Wuhan, China: a single‐centered, retrospective, observational study. Lancet Respir Med. 2020;8(5):475–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Ruan Q, Yang K, Wang W, Jiang L, Song J. Clinical predictors of mortality due to COVID‐19 based on an analysis of data of 150 patients from Wuhan, China. Intensive Care Med. 2020;46(5):846–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Zhang X, Tan Y, Ling Y, Lu G, Liu F, Yi Z, et al. Viral and host factors related to the clinical outcome of COVID‐19. Nature. 2020;583(7816):437–40. [DOI] [PubMed] [Google Scholar]
- 94. Meng Y, Wu P, Lu W, Liu K, Ma K, Huang L, et al. Sex‐specific clinical characteristics and prognosis of coronavirus disease‐19 infection in Wuhan, China: a retrospective study of 168 severe patients. PLoS Pathog. 2020;16(4):e1008520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Halter S, Aimade L, Barbie M, Brisson H, Rouby JJ, Langeron O, et al. T regulatory cells activation and distribution are modified in critically ill patients with acute respiratory distress syndrome: a prospective single‐Centre observational study. Anaesth Crit Care Pain Med. 2020;39(1):35–44. [DOI] [PubMed] [Google Scholar]
- 96. Qu K, Zaba LC, Giresi PG, Li R, Longmire M, Kim YH, et al. Individuality and variation of personal regulomes in primary human T cells. Cell Syst. 2015;1(1):51–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Wang J, Syrett CM, Kramer MC, Basu A, Atchison ML, Anguera MC. Unusual maintenance of X chromosome inactivation predisposes female lymphocytes for increased expression from the inactive X. Proc Natl Acad Sci U S A. 2016;113(14):E2029–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Xu K, Chen Y, Yuan J, Yi P, Ding C, Wu W, et al. Factors associated with prolonged viral RNA shedding in patients with coronavirus disease 2019 (COVID‐19). Clin Infect Dis. 2020;71(15):799–806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Zheng S, Fan J, Yu F, Feng B, Lou B, Zou Q, et al. Viral load dynamics and disease severity in patients infected with SARS‐CoV‐2 in Zhejiang province, China, January‐march 2020: retrospective cohort study. BMJ. 2020;369:m1443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Bai F, Tomasoni D, Falcinella C, Barbanotti D, Castoldi R, Mule G, et al. Female gender is associated with long COVID syndrome: a prospective cohort study. Clin Microbiol Infect. 2021;28(4):611.e9‐611.e16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Ganesh R, Grach SL, Ghosh AK, Bierle DM, Salonen BR, Collins NM, et al. The female‐predominant persistent immune dysregulation of the post‐COVID syndrome. Mayo Clin Proc. 2022;97:454–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102. Nalbandian A, Sehgal K, Gupta A, Madhavan MV, McGroder C, Stevens JS, et al. Post‐acute COVID‐19 syndrome. Nat Med. 2021;27(4):601–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103. Fernandez‐de‐Las‐Penas C, Martin‐Guerrero JD, Pellicer‐Valero OJ, Navarro‐Pardo E, Gomez‐Mayordomo V, Cuadrado ML, et al. Female sex is a risk factor associated with long‐term post‐COVID related‐symptoms but not with COVID‐19 symptoms: the LONG‐COVID‐EXP‐CM multicenter study. J Clin Med. 2022;11(2):413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Salamanna F, Veronesi F, Martini L, Landini MP, Fini M. Post‐COVID‐19 syndrome: the persistent symptoms at the post‐viral stage of the disease. A systematic review of the current data. Front Med (Lausanne). 2021;8:653516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Liu Y, Ebinger JE, Mostafa R, Budde P, Gajewski J, Walker B, et al. Paradoxical sex‐specific patterns of autoantibody response to SARS‐CoV‐2 infection. J Transl Med. 2021;19(1):524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Klein SL. The effects of hormones on sex differences in infection: from genes to behavior. Neurosci Biobehav Rev. 2000;24(6):627–38. [DOI] [PubMed] [Google Scholar]
- 107. Uchida K, Chaudhary LR, Sugimura Y, Adkisson HD, Hruska KA. Proprotein convertases regulate activity of prostate epithelial cell differentiation markers and are modulated in human prostate cancer cells. J Cell Biochem. 2003;88(2):394–9. [DOI] [PubMed] [Google Scholar]
- 108. Couture F, D'Anjou F, Desjardins R, Boudreau F, Day R. Role of proprotein convertases in prostate cancer progression. Neoplasia. 2012;14(11):1032–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Okwan‐Duodu D, Lim EC, You S, Engman DM. TMPRSS2 activity may mediate sex differences in COVID‐19 severity. Signal Transduct Target Ther. 2021;6(1):100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110. Tomlins SA, Rhodes DR, Perner S, Dhanasekaran SM, Mehra R, Sun XW, et al. Recurrent fusion of TMPRSS2 and ETS transcription factor genes in prostate cancer. Science. 2005;310(5748):644–8. [DOI] [PubMed] [Google Scholar]
- 111. McCoy J, Wambier CG, Vano‐Galvan S, Shapiro J, Sinclair R, Ramos PM, et al. Racial variations in COVID‐19 deaths may be due to androgen receptor genetic variants associated with prostate cancer and androgenetic alopecia. Are anti‐androgens a potential treatment for COVID‐19? J Cosmet Dermatol. 2020;19(7):1542–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Penna C, Mercurio V, Tocchetti CG, Pagliaro P. Sex‐related differences in COVID‐19 lethality. Br J Pharmacol. 2020;177(19):4375–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113. Lin B, Ferguson C, White JT, Wang S, Vessella R, True LD, et al. Prostate‐localized and androgen‐regulated expression of the membrane‐bound serine protease TMPRSS2. Cancer Res. 1999;59(17):4180–4. [PubMed] [Google Scholar]
- 114. Mohamed MS, Moulin TC, Schioth HB. Sex differences in COVID‐19: the role of androgens in disease severity and progression. Endocrine. 2021;71(1):3–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. Goren A, Vano‐Galvan S, Wambier CG, McCoy J, Gomez‐Zubiaur A, Moreno‐Arrones OM, et al. A preliminary observation: male pattern hair loss among hospitalized COVID‐19 patients in Spain ‐ a potential clue to the role of androgens in COVID‐19 severity. J Cosmet Dermatol. 2020;19(7):1545–7. [DOI] [PubMed] [Google Scholar]
- 116. Chakladar J, Shende N, Li WT, Rajasekaran M, Chang EY, Ongkeko WM. Smoking‐mediated upregulation of the androgen pathway leads to increased SARS‐CoV‐2 susceptibility. Int J Mol Sci. 2020;21(10):3627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117. Cai G, Bosse Y, Xiao F, Kheradmand F, Amos CI. Tobacco smoking increases the lung gene expression of ACE2, the receptor of SARS‐CoV‐2. Am J Respir Crit Care Med. 2020;201(12):1557–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118. Yin J, Kasper B, Petersen F, Yu X. Association of cigarette smoking, COPD, and lung cancer with expression of SARS‐CoV‐2 entry genes in human airway epithelial cells. Front Med (Lausanne). 2020;7:619453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Montopoli M, Zumerle S, Vettor R, Rugge M, Zorzi M, Catapano CV, et al. Androgen‐deprivation therapies for prostate cancer and risk of infection by SARS‐CoV‐2: a population‐based study (N = 4532). Ann Oncol. 2020;31(8):1040–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Patel VG, Zhong X, Liaw B, Tremblay D, Tsao CK, Galsky MD, et al. Does androgen deprivation therapy protect against severe complications from COVID‐19? Ann Oncol. 2020;31(10):1419–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Karimi A, Nowroozi A, Alilou S, Amini E. Effects of androgen deprivation therapy on COVID‐19 in patients with prostate cancer: a systematic review and meta‐analysis. Urol J. 2021;18(6):577–84. [DOI] [PubMed] [Google Scholar]
- 122. Jimenez‐Alcaide E, Garcia‐Fuentes C, Hernandez V, De la Pena E, Perez‐Fernandez E, Castro A, et al. Influence of androgen deprivation therapy on the severity of COVID‐19 in prostate cancer patients. Prostate. 2021;81(16):1349–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Schmidt AL, Tucker MD, Bakouny Z, Labaki C, Hsu CY, Shyr Y, et al. Association between androgen deprivation therapy and mortality among patients with prostate cancer and COVID‐19. JAMA Netw Open. 2021;4(11):e2134330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Duarte MBO, Leal F, Argenton JLP, Carvalheira JBC. Impact of androgen deprivation therapy on mortality of prostate cancer patients with COVID‐19: a propensity score‐based analysis. Infect Agent Cancer. 2021;16(1):66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125. Gedeborg R, Styrke J, Loeb S, Garmo H, Stattin P. Androgen deprivation therapy and excess mortality in men with prostate cancer during the initial phase of the COVID‐19 pandemic. PLoS One. 2021;16(10):e0255966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Welen K, Rosendal E, Gisslen M, Lenman A, Freyhult E, Fonseca‐Rodriguez O, et al. A phase 2 trial of the effect of antiandrogen therapy on COVID‐19 outcome: no evidence of benefit, supported by epidemiology and in vitro data. Eur Urol. 2021;81:e124–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Chakravarty D, Ratnani P, Sobotka S, Lundon D, Wiklund P, Nair SS, et al. Increased hospitalization and mortality from COVID‐19 in prostate cancer patients. Cancers (Basel). 2021;13(7):1630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128. Leach DA, Mohr A, Giotis ES, Cil E, Isac AM, Yates LL, et al. The antiandrogen enzalutamide downregulates TMPRSS2 and reduces cellular entry of SARS‐CoV‐2 in human lung cells. Nat Commun. 2021;12(1):4068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. Deng Q, Rasool RU, Russell RM, Natesan R, Asangani IA. Targeting androgen regulation of TMPRSS2 and ACE2 as a therapeutic strategy to combat COVID‐19. iScience. 2021;24(3):102254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Qiao Y, Wang XM, Mannan R, Pitchiaya S, Zhang Y, Wotring JW, et al. Targeting transcriptional regulation of SARS‐CoV‐2 entry factors ACE2 and TMPRSS2. Proc Natl Acad Sci U S A. 2020;118(1):e2021450118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131. Cadegiani FA, McCoy J, Gustavo Wambier C, Goren A. Early antiandrogen therapy with Dutasteride reduces viral shedding, inflammatory responses, and time‐to‐remission in males with COVID‐19: a randomized, double‐blind, placebo‐controlled interventional trial (EAT‐DUTA AndroCoV trial ‐ biochemical). Cureus. 2021;13(2):e13047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132. McCoy J, Goren A, Cadegiani FA, Vano‐Galvan S, Kovacevic M, Situm M, et al. Proxalutamide reduces the rate of hospitalization for COVID‐19 male outpatients: a randomized double‐blinded placebo‐controlled trial. Front Med (Lausanne). 2021;8:668698. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 133. Li D, Jin M, Bao P, Zhao W, Zhang S. Clinical characteristics and results of semen tests among men with coronavirus disease 2019. JAMA Netw Open. 2020;3(5):e208292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Verma S, Saksena S, Sadri‐Ardekani H. ACE2 receptor expression in testes: implications in coronavirus disease 2019 pathogenesisdagger. Biol Reprod. 2020;103(3):449–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135. Baratchian M, McManus JM, Berk MP, Nakamura F, Mukhopadhyay S, Xu W, et al. Androgen regulation of pulmonary AR, TMPRSS2 and ACE2 with implications for sex‐discordant COVID‐19 outcomes. Sci Rep. 2021;11(1):11130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136. Stopsack KH, Mucci LA, Antonarakis ES, Nelson PS, Kantoff PW. TMPRSS2 and COVID‐19: serendipity or opportunity for intervention? Cancer Discov. 2020;10(6):779–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137. Olsen NJ, Kovacs WJ. Evidence that androgens modulate human thymic T cell output. J Investig Med. 2011;59(1):32–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Kissick HT, Sanda MG, Dunn LK, Pellegrini KL, On ST, Noel JK, et al. Androgens alter T‐cell immunity by inhibiting T‐helper 1 differentiation. Proc Natl Acad Sci U S A. 2014;111(27):9887–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Henze L, Schwinge D, Schramm C. The effects of androgens on T cells: clues to female predominance in autoimmune liver diseases? Front Immunol. 2020;11:1567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Trigunaite A, Dimo J, Jorgensen TN. Suppressive effects of androgens on the immune system. Cell Immunol. 2015;294(2):87–94. [DOI] [PubMed] [Google Scholar]
- 141. Der E, Dimo J, Trigunaite A, Jones J, Jørgensen TN. Gr1+ cells suppress T‐dependent antibody responses in (NZB× NZW) F1 male mice through inhibition of T follicular helper cells and germinal center formation. J Immunol. 2014;192(4):1570–6. [DOI] [PubMed] [Google Scholar]
- 142. Furman D, Hejblum BP, Simon N, Jojic V, Dekker CL, Thiebaut R, et al. Systems analysis of sex differences reveals an immunosuppressive role for testosterone in the response to influenza vaccination. Proc Natl Acad Sci U S A. 2014;111(2):869–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143. Ma L, Xie W, Li D, Shi L, Ye G, Mao Y, et al. Evaluation of sex‐related hormones and semen characteristics in reproductive‐aged male COVID‐19 patients. J Med Virol. 2021;93(1):456–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144. Rastrelli G, Di Stasi V, Inglese F, Beccaria M, Garuti M, Di Costanzo D, et al. Low testosterone levels predict clinical adverse outcomes in SARS‐CoV‐2 pneumonia patients. Andrology. 2021;9(1):88–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145. Dhindsa S, Zhang N, McPhaul MJ, Wu Z, Ghoshal AK, Erlich EC, et al. Association of circulating sex hormones with inflammation and disease severity in patients with COVID‐19. JAMA Netw Open. 2021;4(5):e2111398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146. Bobjer J, Katrinaki M, Tsatsanis C, Lundberg Giwercman Y, Giwercman A. Negative association between testosterone concentration and inflammatory markers in young men: a nested cross‐sectional study. PLoS One. 2013;8(4):e61466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Mohamad NV, Wong SK, Wan Hasan WN, Jolly JJ, Nur‐Farhana MF, Ima‐Nirwana S, et al. The relationship between circulating testosterone and inflammatory cytokines in men. Aging Male. 2019;22(2):129–40. [DOI] [PubMed] [Google Scholar]
- 148. Klein SL, Marriott I, Fish EN. Sex‐based differences in immune function and responses to vaccination. Trans R Soc Trop Med Hyg. 2015;109(1):9–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149. Robinson DP, Hall OJ, Nilles TL, Bream JH, Klein SL. 17beta‐estradiol protects females against influenza by recruiting neutrophils and increasing virus‐specific CD8 T cell responses in the lungs. J Virol. 2014;88(9):4711–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150. Ding T, Zhang J, Wang T, Cui P, Chen Z, Jiang J, et al. Potential influence of menstrual status and sex hormones on female severe acute respiratory syndrome coronavirus 2 infection: a cross‐sectional multicenter study in Wuhan, China. Clin Infect Dis. 2021;72(9):e240–e8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151. Cheng M, Xu N, Iwasiow B, Seidah N, Chretien M, Shiu RP. Elevated expression of proprotein convertases alters breast cancer cell growth in response to estrogen and tamoxifen. J Mol Endocrinol. 2001;26(2):95–105. [DOI] [PubMed] [Google Scholar]
- 152. Stygar D, Masironi B, Eriksson H, Sahlin L. Studies on estrogen receptor (ER) alpha and beta responses on gene regulation in peripheral blood leukocytes in vivo using selective ER agonists. J Endocrinol. 2007;194(1):101–19. [DOI] [PubMed] [Google Scholar]
- 153. Liu J, Ji H, Zheng W, Wu X, Zhu JJ, Arnold AP, et al. Sex differences in renal angiotensin converting enzyme 2 (ACE2) activity are 17beta‐oestradiol‐dependent and sex chromosome‐independent. Biol Sex Differ. 2010;1(1):6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154. Al‐Kuraishy HM, Al‐Gareeb AI, Faidah H, Al‐Maiahy TJ, Cruz‐Martins N, Batiha GE. The looming effects of estrogen in Covid‐19: a rocky rollout. Front Nutr. 2021;8:649128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155. Stelzig KE, Canepa‐Escaro F, Schiliro M, Berdnikovs S, Prakash YS, Chiarella SE. Estrogen regulates the expression of SARS‐CoV‐2 receptor ACE2 in differentiated airway epithelial cells. Am J Physiol Lung Cell Mol Physiol. 2020;318(6):L1280–L1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156. Nadkarni S, Cooper D, Brancaleone V, Bena S, Perretti M. Activation of the annexin A1 pathway underlies the protective effects exerted by estrogen in polymorphonuclear leukocytes. Arterioscler Thromb Vasc Biol. 2011;31(11):2749–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157. Nadkarni S, McArthur S. Oestrogen and immunomodulation: new mechanisms that impact on peripheral and central immunity. Curr Opin Pharmacol. 2013;13(4):576–81. [DOI] [PubMed] [Google Scholar]
- 158. Ma Q, Hao ZW, Wang YF. The effect of estrogen in coronavirus disease 2019. Am J Physiol Lung Cell Mol Physiol. 2021;321(1):L219–L27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159. Kramer PR, Kramer SF, Guan G. 17 beta‐estradiol regulates cytokine release through modulation of CD16 expression in monocytes and monocyte‐derived macrophages. Arthritis Rheum. 2004;50(6):1967–75. [DOI] [PubMed] [Google Scholar]
- 160. Breithaupt‐Faloppa AC, Correia CJ, Prado CM, Stilhano RS, Ureshino RP, Moreira LFP. 17beta‐estradiol, a potential ally to alleviate SARS‐CoV‐2 infection. Clinics (Sao Paulo). 2020;75:e1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161. Cattrini C, Bersanelli M, Latocca MM, Conte B, Vallome G, Boccardo F. Sex hormones and hormone therapy during COVID‐19 pandemic: implications for patients with cancer. Cancers (Basel). 2020;12(8):2325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162. Calderone A, Menichetti F, Santini F, Colangelo L, Lucenteforte E, Calderone V. Selective estrogen receptor modulators in COVID‐19: a possible therapeutic option? Front Pharmacol. 2020;11:1085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163. Vatansev H, Kadiyoran C, Cumhur Cure M, Cure E. COVID‐19 infection can cause chemotherapy resistance development in patients with breast cancer and tamoxifen may cause susceptibility to COVID‐19 infection. Med Hypotheses. 2020;143:110091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164. Abramenko N, Vellieux F, Tesarova P, Kejik Z, Kaplanek R, Lacina L, et al. Estrogen receptor modulators in viral infections such as SARS‐CoV‐2: therapeutic consequences. Int J Mol Sci. 2021;22(12):6551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165. Di Vincenzo A, Andrisani A, Vettor R, Rossato M. Estrogen and COVID‐19: friend or foe? Ann Oncol. 2021;32(7):933–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166. Allegretti M, Cesta MC, Zippoli M, Beccari A, Talarico C, Mantelli F, et al. Repurposing the estrogen receptor modulator raloxifene to treat SARS‐CoV‐2 infection. Cell Death Differ. 2022;29(1):156–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167. Lovre D, Bateman K, Sherman M, Fonseca VA, Lefante J, Mauvais‐Jarvis F. Acute estradiol and progesterone therapy in hospitalised adults to reduce COVID‐19 severity: a randomised control trial. BMJ Open. 2021;11(11):e053684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168. Ghandehari S, Matusov Y, Pepkowitz S, Stein D, Kaderi T, Narayanan D, et al. Progesterone in addition to standard of care vs standard of care alone in the treatment of men hospitalized with moderate to severe COVID‐19: a randomized, controlled pilot trial. Chest. 2021;160(1):74–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169. Jakovac H. Sex differences in COVID‐19 course and outcome: progesterone should not be neglected. J Appl Physiol (1985). 2020;129(5):1007–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170. Klein SL, Jedlicka A, Pekosz A. The Xs and Y of immune responses to viral vaccines. Lancet Infect Dis. 2010;10(5):338–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171. Engler RJ, Nelson MR, Klote MM, VanRaden MJ, Huang CY, Cox NJ, et al. Half‐ vs full‐dose trivalent inactivated influenza vaccine (2004–2005): age, dose, and sex effects on immune responses. Arch Intern Med. 2008;168(22):2405–14. [DOI] [PubMed] [Google Scholar]
- 172. Potluri T, Fink AL, Sylvia KE, Dhakal S, Vermillion MS, Vom Steeg L, et al. Age‐associated changes in the impact of sex steroids on influenza vaccine responses in males and females. NPJ Vaccines. 2019;4:29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173. Fischinger S, Boudreau CM, Butler AL, Streeck H, Alter G. Sex differences in vaccine‐induced humoral immunity. Semin Immunopathol. 2019;41(2):239–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174. Ruggieri A, Anticoli S, D'Ambrosio A, Giordani L, Viora M. The influence of sex and gender on immunity, infection and vaccination. Ann Ist Super Sanita. 2016;52(2):198–204. [DOI] [PubMed] [Google Scholar]
- 175. Ciarambino T, Barbagelata E, Corbi G, Ambrosino I, Politi C, Lavalle F, et al. Gender differences in vaccine therapy: where are we in COVID‐19 pandemic? Monaldi Arch Chest Dis. 2021;91(4):1669. [DOI] [PubMed] [Google Scholar]
- 176. Pinheiro I, Dejager L, Libert C. X‐chromosome‐located microRNAs in immunity: might they explain male/female differences? The X chromosome‐genomic context may affect X‐located miRNAs and downstream signaling, thereby contributing to the enhanced immune response of females. BioEssays. 2011;33(11):791–802. [DOI] [PubMed] [Google Scholar]
- 177. Schurz H, Salie M, Tromp G, Hoal EG, Kinnear CJ, Moller M. The X chromosome and sex‐specific effects in infectious disease susceptibility. Hum Genomics. 2019;13(1):2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178. Castelo‐Branco C, Soveral I. The immune system and aging: a review. Gynecol Endocrinol. 2014;30(1):16–22. [DOI] [PubMed] [Google Scholar]
- 179. Aaby P, Benn CS, Flanagan KL, Klein SL, Kollmann TR, Lynn DJ, et al. The non‐specific and sex‐differential effects of vaccines. Nat Rev Immunol. 2020;20(8):464–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180. Donzelli A, Schivalocchi A, Giudicatti G. Non‐specific effects of vaccinations in high‐income settings: how to address the issue? Hum Vaccin Immunother. 2018;14(12):2904–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181. Aaby P, Martins CL, Garly ML, Bale C, Andersen A, Rodrigues A, et al. Non‐specific effects of standard measles vaccine at 4.5 and 9 months of age on childhood mortality: randomised controlled trial. BMJ. 2010;341:c6495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182. Aaby P, Kollmann TR, Benn CS. Nonspecific effects of neonatal and infant vaccination: public‐health, immunological and conceptual challenges. Nat Immunol. 2014;15(10):895–9. [DOI] [PubMed] [Google Scholar]
- 183. Benn CS, Fisker AB, Rieckmann A, Sorup S, Aaby P. Vaccinology: time to change the paradigm? Lancet Infect Dis. 2020;20(10):e274–e83. [DOI] [PubMed] [Google Scholar]
- 184. Aaby P, Benn C, Nielsen J, Lisse IM, Rodrigues A, Ravn H. Testing the hypothesis that diphtheria‐tetanus‐pertussis vaccine has negative non‐specific and sex‐differential effects on child survival in high‐mortality countries. BMJ Open. 2012;2(3):e000707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185. Gee J, Marquez P, Su J, Calvert GM, Liu R, Myers T, et al. First month of COVID‐19 vaccine safety monitoring ‐ United States, December 14, 2020‐January 13, 2021. MMWR Morb Mortal Wkly Rep. 2021;70(8):283–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186. Bignucolo A, Scarabel L, Mezzalira S, Polesel J, Cecchin E, Toffoli G. Sex disparities in efficacy in COVID‐19 vaccines: a systematic review and meta‐analysis. Vaccines (Basel). 2021;9(8):825. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.