

Abstract
Agricultural intensification is one of the major threats to global biodiversity and ecosystem services. Sustainable management of agricultural lands can reduce these impacts, but few efforts have been made in the context of paddy rice fields, especially in simplified landscapes composed of large monocultures separated by fragments of natural lands, such as in Taiwan or elsewhere in Asia. In this study, during a pest control intervention, we examined the effects of management practices on insect communities under conventional and organic farming systems in the paddy fields of northern Taiwan in 2016. Our results showed that organic practices did not increase the species richness or abundance of the four insect groups (total, predators/parasitoids, pests, and other insects). In addition, the composition of the insect communities did not differ between organic and conventional farming systems. Both the abundance and richness of predator/parasitoid insects were similar between conventional and organic farming systems. The pest abundance in the organic systems could be suppressed by other unmeasured predators and integrated management practices, which showed similar levels to the conventional systems with pesticide applications. The results of this study suggest that farming systems may not influence insect biodiversity in simplified landscapes during pest-control intervention.
Keywords: Biodiversity, Conventional production, Organic production, Sustainability, Pesticide
BACKGROUND
Agricultural intensification is one of the major threats to global biodiversity (Firbank et al. 2008). High levels of human appropriation of terrestrial net primary production involves the adoption of improved crop varieties and the application of pesticides and chemical fertilizers, which cause environmental degradation and other ecological impacts (e.g., Jonsson et al. 2012; Inclán et al. 2015). For example, decreased ecological complexity leads to a decline in the farmland biodiversity of Europe (Kleijn et al. 2011). In addition, the issue of biodiversity loss has spurred growing concerns about declining ecosystem services, such as the impacts of reduced diversity and the abundance of arthropod natural enemies on pest suppression (Caprio et al. 2015) and possible disruptions of plant-pollinator interactions (Batary et al. 2013). The management of agricultural lands can differentially influence nutrient cycling (Horth and Campbell 2018).
Pest management strategies are effective in preventing economic damage to organic crops (Zehnder et al. 2007). Based on the restricted use of pesticides and chemical fertilizers, organic farming has been proposed to mitigate biodiversity loss and ecosystem disservices on agricultural land (Tuck et al. 2014) and could enhance agricultural sustainability (Liu et al. 2016). For example, organic farming has been proposed to be an agri-environment scheme (AES) based on a key policy of the European Union aiming to counteract the negative effects of intensified agricultural management (Batary et al. 2015). However, different taxa show inconsistent responses to organic farming (Hole et al. 2005; Rahmann 2011), possibly due to differences in mobility or other biological traits. In addition, surrounding landscapes can influence communities, e.g., insect predators (Ali et al. 2020). Case studies have shown that organic farming does not always enhance the diversity of predatory arthropods or suppress the diversity of insect pests (Winqvist et al. 2011; Froidevaux et al. 2017; Lundin et al. 2017). This can occur when other management practices (e.g., use of resistant crop varieties and manual removal of pests) have been reported to compensate for the loss of efficient pest suppression under farming systems that do not apply pesticides in fields (Alam et al. 2016; Brzozowski and Mazourek 2018). Other studies have indicated that while organic farms host richer communities of sessile or less mobile organisms (plants and caterpillars) than conventional farms, the diversity of vagile taxa (e.g., birds) is not different between the two farming systems (Puig-Montserrat et al. 2017).
Rice (Oryza sativa L.) is one of the most important food crops in regions with warm weather and abundant moisture levels, mainly within Asia. The effects of organic farming compared to conventional farming on biodiversity levels are seldom investigated in rice fields (Katayama et al. 2019), and previous studies have mainly focused on the impacts on local generalist predator communities (e.g., carabid beetles, spiders, bats, and frogs) (Ohwaki 2015; Baba and Tanaka 2016; Toffoli and Rughetti 2017). In addition to focusing on generalist predators, previous studies have explored few extensions to other predatory, phytophagous, or neutral/other assemblages (e.g., insect communities in organic and nonorganic rice ecosystems of Indonesia) (Ovawanda et al. 2016). General conclusions and recommendations can be made only after considering a wider variety of taxa, agricultural systems, and bioclimatic regions.
The objective of this study was to examine the overall effects of farming systems (each with a different set of integrated management practices) on insect communities in rice fields of simplified landscapes to provide suggestions for the sustainable control of key rice pests. In Taiwan and elsewhere in Asia, intensive farming is characterized by small-area fields usually belonging to different owners with independent farming practices. Under the prevailing agricultural features, field-based management is a feasible strategy, but its efficacy remains an open question in simplified landscapes. This is especially true during pest control interventions when pesticides can diffuse among fields. In our simplified landscape, shortly after pesticide applications, we compared the species richness, abundance, and community structure of insect communities in conventionally and organically managed rice fields. We also investigated the potential for pest control when considering the predation of natural enemies and management practices (e.g., using a resistant rice variety in our organic farming system). With this research design, we asked (1) if organic farming increases species richness and abundance and (2) whether higher natural pest predator abundances and either similar or lower pest abundances occur in organic fields compared to conventional fields. The difference in insect richness or abundance can result from their local dynamics and habitat preference under the influence of different farming systems.
MATERIALS AND METHODS
Study area
The study area is located in northern Taiwan (Jiaoxi Township, Yilan County; 24°47'45.9"N, 121°45'27.0"E) and is primarily composed of agricultural lands (Fig. 1). Organic farmers in the area established production-selling organizations in 1987 and gradually enlarged their organic paddy fields. During our study, the total area of organic fields was one-third the area of conventional fields. The organic and conventional fields were arranged in a crisscross pattern, with some organic fields concentrated in the northern and southern parts of the study area. The fields with different farming systems were generally mixed together in the simplified landscape. Conventional fields were sometimes right next to organic fields. In addition to the paddy fields, some abandoned fields, houses, and other crop fields were present in the study area. In this area, there was one growing season, from late winter to mid-summer.
Fig. 1.

Rice fields of different farming systems sampled in northern Taiwan.
Organic and conventional farming systems have different sets of management practices (e.g., pesticides, fertilizers, and crop varieties). Within this study area, the paddy fields grow the rice varieties Taichung Sen 10 or Taiken 11 with organic or conventional farming methods, respectively. The variety Taichung Sen 10 has enhanced resistance to arthropod pests (e.g., planthoppers and leafhoppers) compared to the other variety (Cheng and Chang 1979). The organic farmers do not apply chemical fertilizers or pesticides to their fields, following the government’s organic standard in Taiwan. These farmers applied green manures as organic fertilizers during the fallow period, and they removed harmful grasses and insects manually during the rice-growing seasons. They also apply bitter tea meal to kill the golden apple snail (Pomacea canaliculata) before transplanting the rice seedlings. In conventional farms, the chemical fertilizers used include N, P2O5, and K2O. Conventional farmers apply Phorate 10% CG (IRAC 1B) to control the leaf miners, thrips, and leafhoppers once in May. To control diseases of rice blast and sheath blight, a fungicide (6% probenazole) is applied to conventional farms once in May. The farmers use hand-held pesticide sprayers to control these diseases and pests.
Insect sampling
From late May to early July 2016 until or during the rice tillering stage (one to two weeks after pesticide applications), we collected 20 insect samples under each of the organic and conventional farming systems (Fig. 1). By sweep netting with an insect net (handle length: 95 cm; net diameter: 40 cm), each of the 40 samples had a sampling ridge around one paddy field (see the next paragraph for details; Fig. S1). The sweeping approach achieves robust sampling by forcefully sweeping the net through the rice canopy (e.g., Rashid et al. 2006; Ali et al. 2020). The captured insects were brought back to the laboratory for identification. Most of the sampled insects were identified at the species level, while the others were identified at the genus or family level. In addition, these taxa were categorized as predators/parasitoids, rice pests, or other (Table S1).
For each sample, we sampled the insects once by walking 100 m along the ridge and used a sampling width of 1 m in the field at both sides of the ridge (Fig. S1). There were 60 swings of the net along the 100 m. This sampling effort is comparable to other studies using sweep nets in rice fields of Asia (e.g., Sulaiman et al. 2013; Hashim et al. 2017; Katayama et al. 2019). Both sides of the sampling ridge of each sample belonged to either organic or conventional fields. There were two or three fields sampled along each sampling ridge, and different samples did not share the same fields. All of the fields were enclosed in 2 × 2 km. The study field area ranged from 500 and 2000 m2. Each field was separated from the neighboring fields by a ridge. There were no houses, gardens or bodies of water adjacent to the study fields. These organic fields were generally surrounded by sampled and/or unsampled conventional fields in the simplified landscape.
Data analysis
The generalized additive mixed model (GAMM), which models nonlinear relationships between covariates and outcomes, was used to identify the effects of the farming system on species richness and on the abundance of the total insect community, four most abundant taxa (see Tables 1 and S1), predators/parasitoids, pests, and other insects. The species richness of each group was calculated as the number of taxa present in each group. In the GAMM, the farming system is a binary variable (either a conventional or organic system) in the model. Considering the interacting communities linked by species dispersal (Ovaskainen and Abrego 2020), spatial autocorrelation (i.e., based on the interactions among sampling sites based on their geographic distance) was implemented for model adjustment. In addition, the faction of nonnatural land use (e.g., farm, road, and house) within each circular area (with a diameter of 1 km around one sampling field) and field area each had a nonlinear effect based on a smoothing function in the GAMM. A model with a Poisson distribution was set up for each insect group (response variables). We set the significance level α for the GAMMs at α = 0.05, and the GAMMs were performed by the package ‘mgcv’ (Wood 2011) within the R statistical framework (version 3.6.1).
Table 1.
Species richness and abundance (mean ± SD) under conventional and organic farming systems and their GAMM (generalized additive mixed model) analyses on the effects of farming systems on the farms and the nonnatural land surrounding each field (n = 20 fields for each farming system)
| Value | GAMM | |||||
| Conventional | Organic | Farming (t; p-value) | Nonnatural land (F; p-value; EDF) | Field area (F; p-value; EDF) | ||
| Species richness of total insects | 5.5 ± 2.7 | 5.6 ± 2.2 | -0.19; 0.85 | 0.23; 0.64; 1.0 | 0.37; 0.55; 1.0 | |
| Species richness of predators/parasitoids | 1.7 ± 0.8 | 1.6 ± 1.0 | -0.17; 0.86 | 0.01; 0.92; 1.0 | 0.05; 0.83; 1.0 | |
| Species richness of pests | 1.4 ± 1.1 | 1.1 ± 0.9 | -1.90; 0.07 | 0.17; 0.68; 1.0 | 0.90; 0.35; 1.0 | |
| Species richness of others | 2.4 ± 1.5 | 2.8 ± 1.4 | 0.40; 0.69 | 0.67; 0.42; 1.0 | 0.07; 0.80; 1.0 | |
| Total abundance | 16.9 ± 10.2 | 14.4 ± 8.9 | -0.68; 0.50 | 5.38; 0.09; 2.0 | 30.03; 0.00; 7.9 | |
| Abundance of predators/parasitoids | 5.7 ± 2.6 | 5.2 ± 6.4 | -1.39; 0.17 | 3.81; 0.06; 1.0 | 0.09; 0.76; 1.0 | |
| Abundance of pests | 4.8 ± 4.8 | 2.9 ± 3.2 | -2.32; 0.03 | 9.28; 0.04; 2.4 | 11.96; 0.28; 6.3 | |
| Abundance of others | 6.3 ± 5.0 | 6.3 ± 5.6 | -0.58; 0.57 | 4.39; 0.33; 2.3 | 19.54; 0.02; 7.5 | |
| Abundance of AS | 3.0 ± 3.3 | 3.0 ± 3.7 | -0.09; 0.93 | 7.58; 0.05; 2.2 | 0.25; 0.62; 1.0 | |
| Abundance of OS | 2.6 ± 2.1 | 2.4 ± 2.8 | -0.54; 0.59 | 0.56; 0.46; 1.0 | 3.92; 0.06; 1.0 | |
| Abundance of HO | 2.2 ± 3.3 | 0.6 ± 1.1 | -3.80; 0.00 | 2.58; 0.12; 1.0 | 0.02; 0.89; 1.0 | |
| Abundance of CP | 2.5 ± 3.5 | 1.0 ± 1.9 | -0.92; 0.37 | 11.37; 0.00; 1.0 | 1.34; 0.25; 1.0 | |
EDF stands for the estimated degrees of freedom.
We used nonmetric multidimensional scaling (NMDS), based on a Bray-Curtis dissimilarity matrix computed on a log (x+1)-transformation of taxon abundance, to ordinate the insect community samples. Stress values of less than 0.2 indicated credible information in the NMDS plots. ANOSIM (analysis of similarities) was used to determine whether there were significant differences between farming systems. NMDS and ANOSIM were performed by the package ‘vegan’ within the R statistical framework (Oksanen et al. 2016).
RESULTS
In total, 626 insects were captured, and 32 taxa of insects were found in the paddy fields, of which 13 taxa were detected in both organic and conventional fields (see Table S1). Organic and conventional fields contained 24 and 21 taxa, respectively. Among all taxa, Atractomorpha sinensis (Orthoptera; other group) and Orthetrum sabina (Odonata; predator group) were the most abundant species (up to 20.4 and 16.3%, respectively), in both conventional and organic farming fields. In addition, Harmonia octomaculata (Coleoptera) and Cletus punctiger (Hemiptera) were the most abundant predators (13.1%) and pests (15.1%), respectively, in the conventional farming fields. Except for H. octomaculata, the farming system did not have significant effects on the abundance of the most common species (2.2 vs. 0.6 individuals). In addition, nonnatural land had significant effects on the abundance of C. punctiger and A. sinensis. However, the field area did not influence their abundance (Table 1).
Except for pest abundance, there was similar species richness and abundance for total insects, predators, pests, and others under different farming systems (Table 1). We found insignificant differences between the abundance (16.9 vs. 14.4 individuals) and species richness (5.5 vs. 5.6 taxa) levels of total insects for conventional and organic farming systems. Nonnatural land or field area had significant effects on the abundance of pests or total insects and others, respectively. These two environmental factors did not influence the species richness of all insect groups. Similarly, the abundances of predators/parasitoids (5.7 vs. 5.2 individuals) were not significantly different under conventional or organic farming systems. The same results were found for the species richness of predators/parasitoids (1.7 vs. 1.6 taxa). The abundance (4.8 vs. 2.9 individuals) or species richness (1.4 vs. 1.1 taxa) of pests showed significant or insignificant differences between the suppression effectiveness of conventional and organic farming systems, respectively. Conventional and organic farming systems had similar influences on both the abundance (6.3 vs. 6.3 individuals) and species richness (2.4 vs. 2.8 taxa) of other insects.
The two-dimensional NMDS ordination of insect communities under conventional and organic farming systems shows that the different farming systems did not appear to be well separated for these insect communities (Fig. 2). The result of ANOSIM (R = 0.03; p-value = 0.18) can help clarify the interpretation of the NMDS ordination, which shows insignificant differences between insect communities under conventional and organic farming systems.
Fig. 2.
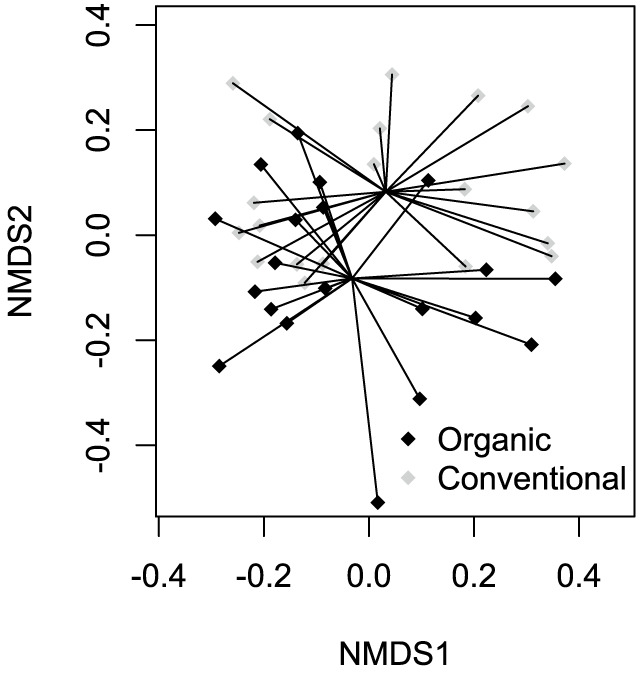
Two-dimensional NMDS ordination of 40 insect communities sampled under different farming systems in northern Taiwan (stress = 0.18).
DISCUSSION
The overall effects of organic and conventional farming systems (each with a different set of integrated management practices) on insect communities were similar in our simplified landscape during the pest-control intervention. Our results show that organic farming did not increase the species richness or abundance but could have higher nonpesticidal pest suppression than conventional farming (one to two weeks after pesticide applications). Pest suppression could result from their local decline and/or low habitat preference under the action of farming systems. The abundance and species richness of overall and other/neutral assemblages did not differ between farming systems. In addition, we did not observe any significant differences in community structure between organic and conventional farming. Predator/parasitoid abundance and richness were similar for organic and conventional farming, and pest abundance could be suppressed by other unmeasured predators and integrated management practices (e.g., resistant rice varieties and the manual removal of pests) and showed similar levels in organic fields without pesticide applications. A meta-analysis showed that biodiversity levels for organic farming are roughly one-third greater than those for conventional farming (Tuck et al. 2014).
The sampling efforts and species richness and abundance of insects in this study were comparable to other studies based on the sweep-net method in Asian paddy fields (e.g., Sulaiman et al. 2013; Hashim et al. 2017; Katayama et al. 2019). Collecting a smaller number of individuals would influence species richness because of the higher absence probability of rare species, which can result in an underestimated number of species. However, rare species contribute little to abundance and community structure, and our sampling protocol caused systemic, consistent bias among all samples. This situation will not influence our statistical results by comparing the effects of the farming systems. In our study, the simplified landscape complexity in Taiwan could explain why biodiversity and nonpesticidal pest suppression were usually independent of the farming system. This is especially true during pest-control interventions when pesticide spreading might homogenize the effects of farming systems on insect communities because the organic and conversational fields are mixed together in the simplified landscape (see below).
We did not observe any significant differences in abundance, species richness, or community composition of total insects between farming systems. The intensive use of pesticides in conventional paddy fields has raised concerns about their impact on nontarget organisms (Ohwaki 2015; Maltchik et al. 2017; Martinez-Eixarch et al. 2017). There are some cases showing lower richness and abundance in organic farms (e.g., no hypothesized changes in macroinvertebrates due to water-level management to control weeds and soil tillage) (Dalzochio et al. 2016). At least one possible explanation could account for our findings: organic fields may be influenced by leaching or aerial pesticide applications in neighboring conventional fields (Reichenberger et al. 2007). This would delimit the differences between community structures because the arthropod communities of organic fields would also be exposed to agrochemical threats. Because pesticides degrade over time, pesticide persistence in conventional rice crops influences the structural similarity of insect communities. In addition, the regional species pool is simplified based on the intensive-farming landscape, and these overall species might tend to be generalists, which could inhabit both organic and conventional fields. Another study found that species richness was influenced by the landscape context irrespective of management type (Schmidt et al. 2005). Lower species richness is typically found in structurally simple landscapes (Sunderland and Samu 2000). However, one case study showed clear differences in community composition despite a similar species pool being shared by different types of vineyards (Masoni et al. 2017).
Although we did not find a higher abundance of predators/parasitoids under organic farming systems, pest suppression under organic farming systems can be partly attributed to the two integrated management practices (i.e., resistant rice varieties and the manual removal of harmful insects). In addition, the potentially enhanced abundance of other unmeasured predators, e.g., spiders, bats, and frogs (Ohwaki 2015; Baba and Tanaka 2016; Toffoli and Rughetti 2017), could also result in higher predation rates in organic fields, which would partly explain the lower abundances of pests under organic than conventional farming systems. Other studies showed that organic farming resulted in higher abundances of predators/parasitoids and changes in their community composition (Inclán et al. 2015; Birkhofer et al. 2016; Katayama 2016), which partly or wholly suppressed pest populations under organic farming, replacing the pesticides used in conventional fields (Crowder et al. 2010).
Indeed, organic farming seems to enhance the species richness and abundance of many common taxa, but its effects are often species-specific and trait-or context-dependent (Winqvist et al. 2012). Spiders are one of the major predators in paddy fields (Deb 2009) and have been shown to occur more often in organic farming systems than in conventional farming systems (Birkhofer et al. 2016), which could be attributed to their lower dispersion abilities and higher dependency on local factors. In both organic and conventional systems, we did not observe some major pests in the fields, e.g., the brown rice planthopper Nilaparvata lugens and the green rice leafhopper Nephotettix cincticeps in Taiwan (Cheng and Chiu 1999), which may be attributed to the enhanced resistance of the rice variety (Taichung Sen 10) to arthropod pests (e.g., planthoppers and leafhoppers) in organic fields. Our major objective was to study farming effects and sampling was conducted only after pesticide applications, so any depressive effects on insect communities in conventional fields and/or a seasonal occurrence of these major taxa would also result in their absence in our samples.
CONCLUSIONS
Rice is an important food crop, and the intensive use of agrichemicals raises concerns about their threats to communities of nontarget organisms (Wandscheer et al. 2017). This study investigated the influence of farming systems on predatory, phytophagous, and neutral/other assemblages seldom addressed in rice field studies. There is no question as to whether organic farming can decrease the negative effects and enhance biodiversity compared to conventional farming (Bengtsson et al. 2005; Tuck et al. 2014). This may also be true in simplified landscapes, but pesticide spreading among fields can obscure the influences of different farming systems during pest control interventions. For example, we did not observe the hypothesized responses of the insect communities in organic and conventional rice fields, but the enhancement of nonpesticidal pest suppression (through their local decline and/or low habitat preference) could occur in organic fields under a set of integrated management practices. In Taiwan and elsewhere in Asin, small-area fields with independent owners and farming practices are integrated into the simplified landscape. This prevailing feature has limited the range of management strategies available. However, our results suggest the rethinking of landscape-based management implemented in areas with intensive farming. In addition, under the current socioeconomic conditions, considerable regional efforts are necessary as investments for the hypothesized farming efficacy.
Supplementary materials
Conceptual diagram of hand-net sweep sampling in this study.
Relative abundances of insects (%) under organic and conventional farming (n = 20 fields for each farming system).
Acknowledgments
This study was supported by grants from the Council of Agriculture [Grant No. 105AgriSci-12.4.2-Forest-G1 (3)], the Ministry of Science and Technology, Taiwan (Grant No. 105-2621-M-005-005), and the “Innovation and Development Center of Sustainable Agriculture” project administered by the “Featured Areas Research Center program” within the framework of Taiwanese Ministry of Education’s “Higher Education Sprout Project”.
Footnotes
Authors’ contributions: Conceptualization, CNK, MCC, and HJL; methodology and software, CNK and MCC; data collection and curation, CNK, LMJ, and YJL; writing—original draft preparation, CNK and MCC; writing—review and editing, All authors; supervision, CNK and HJL; funding acquisition, CNK and HJL.
Availability of data and materials: The datasets used and analyzed are available from the corresponding author on reasonable request.
Competing interests: All authors declare that they have no conflict of interest.
Consent for publication: Not applicable.
Ethics approval consent to participate: Not applicable.
References
- Alam MZ, Crump AR, Haque MM, Islam MS, Hossain E, Hasan SB, Hasan SB, Hossain MS. 2016. Effects of integrated pest management on pest damage and yield components in a rice agro-ecosystem in the barisal region of bangladesh. Front Environ Sci 4:22. doi:10.3389/fenvs.2016.00022.
- Ali MP, Kabir MMM, Haque SS, Afrin S, Ahmed N, Pittendrigh B, Qin X. 2020. Surrounding landscape influences the abundance of insect predators in rice field. BMC Zool 5:8. doi:10.1186/s40850-020-00059-1.
- Baba YG, Tanaka K. 2016. Environmentally friendly farming and multi-scale environmental factors influence generalist predator community in rice paddy ecosystems of Japan. NIAES Series 6:169–177.
- Batary P, Dicks LV, Kleijn D, Sutherland WJ. 2015. The role of agri-environment schemes in conservation and environmental management. Conserv Biol 29:1006–1016. doi:10.1111/cobi.12536. . [DOI] [PMC free article] [PubMed]
- Batary P, Sutcliffe L, Dormann CF, Tscharntke T. 2013. Organic farming favours insect-pollinated over non-insect pollinated forbs in meadows and wheat fields. PLoS ONE 8:e54818. doi:10.1371/journal.pone.0054818. . [DOI] [PMC free article] [PubMed]
- Bengtsson J, Ahnstrom J, Weibull AC. 2005. The effects of organic agriculture on biodiversity and abundance: a meta-analysis. J Appl Ecol 42:261–269. doi:10.1111/j.1365-2664.2005.01005.x.
- Birkhofer K, Arvidsson F, Ehlers D, Mader VL, Bengtsson J, Smith HG. 2016. Organic farming affects the biological control of hemipteran pests and yields in spring barley independent of landscape complexity. Landsc Ecol 31:567–579. doi:10.1007/s10980-015-0263-8.
- Brzozowski L, Mazourek M. 2018. A sustainable agricultural future relies on the transition to organic agroecological pest management. Sustainability 10:2023. doi:10.3390/su10062023.
- Caprio E, Nervo B, Isaia M, Allegro G, Rolando A. 2015. Organic versus conventional systems in viticulture: Comparative effects on spiders and carabids in vineyards and adjacent forests. Agr Syst 136:61–69. doi:10.1016/j.agsy.2015.02.009.
- Cheng CH, Chang WL. 1979. Studies on varietal resistance to brown planthopper in Taiwan. Brown planthopper: Treat to rice production in Asia. Los Banos, Philippines: IRRI. pp. 251–257.
- Cheng CH, Chiu YI. 1999. Review of changes involving rice pests and their control measures in Taiwan since 1945. Plant Prot Bull 41:9–34.
- Crowder DW, Northfield TD, Strand MR, Snyder WE. 2010. Organic agriculture promotes evenness and natural pest control. Nature 466:109–112. doi:10.1038/nature09183. . [DOI] [PubMed]
- Dalzochio MS, Baldin R, Stenert C, Maltchik L. 2016. Can organic and conventional agricultural systems affect wetland macroinvertebrate taxa in rice fields? Basic Appl Ecol 17:220–229. doi:10.1016/j.baae.2015.10.009.
- Deb D. 2009. Biodiversity and complexity of rice farm ecosystems:an empirical assessment. Open Ecol J 2:112–129. doi:10.2174/1874 213000902010112.
- Firbank LG, Petit S, Smart S, Blain A, Fuller RJ. 2008. Assessing the impacts of agricultural intensification on biodiversity: a British perspective. Philos T R Soc B 363:777–787. doi:10.1098/rstb.2007.2183. . [DOI] [PMC free article] [PubMed]
- Froidevaux JSP, Louboutin B, Jones G. 2017. Does organic farming enhance biodiversity in Mediterranean vineyards? A case study with bats and arachnids. Agr Ecosyst Environ 249:112–122. doi:10.1016/j.agee.2017.08.012. . [DOI] [PMC free article] [PubMed]
- Hashim NA, Aziz M, Basari N, Saad K, Jasmi A, Hamid SA. 2017. Diversity and guild structure of insects during rice flowering stage at a selected rice field in Penang, Malaysia. Malays Appl Biol 46:161–169.
- Hole DG, Perkins AJ, Wilson JD, Alexander IH, Grice PV, Evans AD. 2005. Does organic farming benefit biodiversity? Biol Conserv 122:113–130. doi:10.1016/j.biocon.2004.07.018.
- Horth L, Campbell LA. 2018. Supplementing small farms with native mason bees increases strawberry size and growth rate. J Appl Ecol 55:591–599. doi:10.1111/1365-2664.12988.
- Inclán DJ, Cerretti P, Gabriel D, Benton TG, Sait SM, Kunin WE, Gillespie MAK, Marini L. 2015. Organic farming enhances parasitoid diversity at the local and landscape scales. J Appl Ecol 52:1102–1109. doi:10.1111/1365-2664.12457.
- Jonsson M, Buckley HL, Case BS, Wratten SD, Hale RJ, Didham RK. 2012. Agricultural intensification drives landscape-context effects on host-parasitoid interactions in agroecosystems. J Appl Ecol 49:706–714. doi:10.1111/j.1365-2664.2012.02130.x.
- Katayama N. 2016. Bird diversity and abundance in organic and conventional apple orchards in northern Japan. Sci Rep 6:34210. doi:10.1038/srep34210. . [DOI] [PMC free article] [PubMed]
- Katayama N, Osada Y, Mashiko M, Baba YG, Tanaka K, Kusumoto Y, Okubo S, Ikeda H, Natuhara Y. 2019. Organic farming and associated management practices benefit multiple wildlife taxa: a large-scale field study in rice paddy landscapes. J Appl Ecol 56:1970–1981. doi:10.1111/1365-2664.13446.
- Kleijn D, Rundlof M, Scheper J, Smith HG, Tscharntke T. 2011. Does conservation on farmland contribute to halting the biodiversity decline? Trends Ecol Evol 26:474–481. doi:10.1016/j.tree.2011.05.009. . [DOI] [PubMed]
- Liu HT, Meng J, Bo WJ, Cheng D, Li Y, Guo LY et al. 2016. Biodiversity management of organic farming enhances agricultural sustainability. Sci Rep 6:23816. doi:10.1038/srep23816. . [DOI] [PMC free article] [PubMed]
- Lundin O, Svensson GP, Larsson MC, Birgersson G, Hederstrom V, Lankinen A, Anderbrant O, Rundlöf M. 2017. The role of pollinators, pests and different yield components for organic and conventional white clover seed yields. Field Crop Res 210:1–8. doi:10.1016/j.fcr.2017.05.014.
- Maltchik L, Stenert C, Batzer DP. 2017. Can rice field management practices contribute to the conservation of species from natural wetlands? Lessons from Brazil. Basic Appl Ecol 18:50–56. doi:10.1016/j.baae.2016.10.002.
- Martinez-Eixarch M, Curco A, Ibanez C. 2017. Effects of agri-environmental and organic rice farming on yield and macrophyte community in Mediterranean paddy fields. Paddy Water Environ 15:457–468. doi:10.1007/s10333-016-0563-x.
- Masoni A, Frizzi F, Bruhl C, Zocchi N, Palchetti E, Chelazzi G, Santini G. 2017. Management matters: a comparison of ant assemblages in organic and conventional vineyards. Agr Ecosyst Environ 246:175–183. doi:10.1016/j.agee.2017.05.036.
- Ohwaki A. 2015. Ground arthropod communities in paddy fields during the dry period: comparison between different farming methods. J Asia-Pac Entomol 18:413–419. doi:10.1016/j.aspen.2015.05.001.
- Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O'Hara RB et al. 2016. vegan: Community Ecology Package. R package version 2.3-5. Available at https://CRAN.R-project.org/package= vegan.
- Ovaskainen O, Abrego N. 2020. Joint Species Distribution Modelling: With Applications in R. Cambridge: Cambridge University Press.
- Ovawanda EA, Witjaksono W, Trisyono YA. 2016. Insect biodiversity in organic and non-organic rice ecosystem in the District of Bantul. J Perlin Tan Indo 20:15–21. doi:10.22146/jpti.16619.
- Puig-Montserrat X, Stefanescu C, Torre I, Palet J, Fabregas E, Dantart J, Arrizabalaga A, Flaquer C. 2017. Effects of organic and conventional crop management on vineyard biodiversity. Agr Ecosyst Environ 243:19–26. doi:10.1016/j.agee.2017.04.005.
- Rahmann G. 2011. Biodiversity and organic farming: What do we know? Landbauforschung 61:189–208.
- Rashid T, Johnson DT, Bernhardt JL. 2006. Sampling rice stink bug (Hemiptera: Pentatomidae) in and around rice fields. Environ Entomol 35:102–111. doi:10.1603/0046-225x-35.1.102.
- Reichenberger S, Bach M, Skitschak A, Frede HG. 2007. Mitigation strategies to reduce pesticide inputs into ground-and surface water and their effectiveness; A review. Sci Total Environ 384:1–35. doi:10.1016/j.scitotenv.2007.04.046. . [DOI] [PubMed]
- Schmidt MH, Roschewitz I, Thies C, Tscharntke T. 2005. Differential effects of landscape and management on diversity and density of ground-dwelling farmland spiders. J Appl Ecol 42:281–287. doi:10.1111/j.1365-2664.2005.01014.x.
- Sulaiman N, Isahak A, Sahid I, Maimon A. 2013. Diversity of pest and non-pest insects in an organic paddy field cultivated under the System of Rice Intensification (SRI): A case study in Lubok China, Melaka, Malaysia. J Food Agric Environ 11:2861–2865.
- Sunderland K, Samu F. 2000. Effects of agricultural diversification on the abundance, distribution, and pest control potential of spiders: a review. Entomol Exp Appl 95:1–13. doi:10.1046/j.1570-7458. 2000.00635.x.
- Toffoli R, Rughetti M. 2017. Bat activity in rice paddies: Organic and conventional farms compared to unmanaged habitat. Agr Ecosyst Environ 249:123–129. doi:10.1016/j.agee.2017.08.022.
- Tuck SL, Winqvist C, Mota F, Ahnstrom J, Turnbull LA, Bengtsson J. 2014. Land-use intensity and the effects of organic farming on biodiversity: a hierarchical meta-analysis. J Appl Ecol 51:746–755. doi:10.1111/1365-2664.12219. . [DOI] [PMC free article] [PubMed]
- Wandscheer ACD, Marchesan E, da Silva MF, Aramburu BB, de David R, Trivisiol VS, da Silva AL. 2017. Impact of fungicide and insecticide use on non-target aquatic organisms in rice paddy fields. Ciência Rural 47:01, e20151475. doi:10.1590/0103-8478cr20151475.
- Winqvist C, Ahnstrom J, Bengtsson J. 2012. Effects of organic farming on biodiversity and ecosystem services: taking landscape complexity into account. Ann Ny Acad Sci 1249:191–203. doi:10.1111/j.1749-6632.2011.06413.x. . [DOI] [PubMed]
- Winqvist C, Bengtsson J, Aavik T, Berendse F, Clement LW, Eggers S et al. 2011. Mixed effects of organic farming and landscape complexity on farmland biodiversity and biological control potential across Europe. J Appl Ecol 48:570–579. doi:10.1111/j.1365-2664.2010.01950.x.
- Wood SN. 2011. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J R Stat Soc Ser B-Stat Methodol 73:3–36. doi:10.1111/j.1467-9868.2010.00749.x.
- Zehnder G, Gurr G, Kühne S, R Wade M, D Wratten S, Wyss E. 2007. Arthropod pest management in organic crops. Annu Rev Entomol 52:57–80. doi:10.1146/annurev.ento.52.110405.091337. . [DOI] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Conceptual diagram of hand-net sweep sampling in this study.
Relative abundances of insects (%) under organic and conventional farming (n = 20 fields for each farming system).