

SUMMARY
Shiga toxin-producing Escherichia coli (STEC) is a known food pathogen, which main reservoir is the intestine of ruminants. The abundance of different STEC lineages in nature reflect a heterogeneity that is characterised by the differential expression of certain genotypic characteristics, which in turn are influenced by the environmental conditions to which the microorganism is exposed. Bacterial homeostasis and stress response are under the control of the alarmone guanosine tetraphosphate (ppGpp), which intrinsic levels varies across the E. coli species. In the present study, 50 STEC isolates from healthy sheep were evaluated regarding their ppGpp content, cytotoxicity and other relevant genetic and phenotypic characteristics. We found that the level of ppGpp and cytotoxicity varied considerably among the examined strains. Isolates that harboured the stx2 gene were the least cytotoxic and presented the highest levels of ppGpp. All stx2 isolates belonged to phylogroup A, while strains that carried stx1 or both stx1 and stx2 genes pertained to phylogroup B1. All but two stx2 isolates belonged to the stx2b subtype. Strains that belonged to phylogroup B1 displayed on average low levels of ppGpp and high cytotoxicity. Overall, there was a negative correlation between cytotoxicity and ppGpp.
Key words: Cytotoxicity, ppGpp, Shiga toxin, STEC
INTRODUCTION
Shiga toxin (Stx)-producing Escherichia coli (STEC) expresses and secretes a potent cytotoxin that can cause serious diseases in humans, such as bloody diarrhoea and HUS (haemolytic uraemic syndrome). Illness due to STEC has been linked to the consumption of different types of food, mostly of animal origin. Natural isolates of STEC can produce Stx1, Stx2, or both toxins; some subtypes of Stx1 and several subtypes of Stx2 exist and can come out in many different combinations [1, 2]. E. coli survival in the animal digestive tract and in foods relies on the ability to respond to environmental stresses, which in turn depends on the expression of hundreds of genes [3]. Knowledge about the molecular properties that allow pathogenic bacteria to thrive and survive in different conditions is valuable to improve their control during the food chain processing.
The nucleotides guanosine tetra and penta-phosphate, collectively known as (p)ppGpp or simply ppGpp, accumulate in response to adverse environmental conditions, such as amino acid, carbon, nitrogen and phosphate starvation [4]. ppGpp accumulation is accompanied by the stringent response, a metabolic adjustment characterised by a dramatic reduction in the synthesis of stable RNA and ribosomes and general protein inhibition [5]. In E. coli, ppGpp is synthesised by two proteins – RelA and SpoT. RelA responds to amino acid starvation when uncharged tRNAs bind to the A site of the ribosome, initiating thus the synthesis of ppGpp. SpoT is a bifunctional enzyme that displays a strong hydrolase and a weak synthetase activity, owe to the presence of two functional domains [6]. SpoT responds to several stresses (but not to amino acid shortage) by inhibiting the hydrolase activity, resulting in the stabilisation of ppGpp [7]. ppGpp is associated with diverse cellular functions, such as rRNA synthesis, mRNA elongation, amino acids, carbohydrate and lipid metabolism, DNA replication and repair and bacterial virulence [4, 8, 9]. The hallmark of the stringent response is the general inhibition of protein synthesis caused by a strong reduction in the synthesis of ribosomes. Genes that promote cell catabolism and replication are also negatively regulated by this alarmone [5]. On the other hand, some genes are positively affected by ppGpp, including those involved in amino acid biosynthesis [5] and genes related to cell survival and protection, such as rpoS that encodes the sigma factor that coordinates the general stress response [10].
The mechanism through which ppGpp exerts so many pleiotropic effects is not entirely clear. It can be partially explained by the fact that ppGpp interacts directly with RNA polymerase, shifting the affinity of the core enzyme towards alternative sigma factors [11]. ppGpp is positively associated with bacterial virulence in several species [8]. In E. coli pathotypes, such as EHEC, UPEC and EPEC ppGpp modulates the expression of virulence factors, such as the synthesis of adhesins and fimbriae important for bacterial attachment to the host cell [12–14]. However, it has been shown that ppGpp inhibits the lytic development and DNA replication of Stx bacteriophages, which suggests a negative role for ppGpp in the expression of virulence in bacteria that carry toxin-encoding phages such as STEC [15–17]. In the present study, we correlated bacterial cytotoxicity and ppGpp in a group of fifty STEC strains isolated from sheep. The levels of both ppGpp and cytotoxicity were not constant among the different isolates, and a negative correlation between ppGpp and cytotoxicity was observed.
METHODS
Bacterial strains and growth conditions
E. coli strains were isolated from rectal swabs of 23 healthy sheep in the southwest of the state of Goiás, Brazil, and plated on MacConkey agar. Colonies were biochemically identified as belonging to the species E. coli based on lactose fermentation, indole production, Voges–Proskauer assay, citrate utilisation, absence of urease and production of hydrogen sulphide. All isolates were analysed by multiplex PCR and classified into different phylogenetic groups as described by Clermont et al. [18]. The isolates were then tested for the presence of stx1, stx2 and eae genes by PCR, as described by [19]. Strains that carry one or both stx genes have been considered as belonging to the STEC pathotype. Strains EDL933 (O157:H7, stx1+, stx2+, eae+) and E2348/69 (eae+) were used as positive controls for the PCR characterisation. Strains that harboured the stx2 gene were further analysed by PCR subtyping essentially as described [1].
LB was the standard bacterial rich medium [20]. TGP is a minimal medium composed of T-salts [21] supplemented with 0·2% glucose and either 1 mM or 0·2 mM KH2PO4. Vero cells were cultivated in Dulbecco's modified Eagle medium (DMEM) supplemented with 10% Fetal Bovine Serum (FBS).
ppGpp assay
ppGpp was assayed essentially as described [22]. Briefly, bacteria were grown until the exponential phase in non-limited TGP minimal medium, at which point cells were resuspended in TGP containing a limited concentration of phosphate (0·2 mM KH2PO4 and 100 µCi/ml32P). Samples were harvested after 60 and 240 min, mixed with 2 M cold formic acid and stored overnight at −20 °C. On the next day, the cell extracts were centrifuged to precipitate debris and 5 µl of each sample were applied to PEI-cellulose thin layer chromatography (TLC) plates. The labelled nucleotides were resolved by one-dimensional TLC using 1·5 M KH2PO4 as solvent. The amounts of ppGpp on the chromatograms were estimated by measuring the radioactivity content in the ppGpp and GTP spots in a Phosphor-Imager (Molecular Dynamics). The level of ppGpp was calculated according to the formula: ppGpp = ppGpp/(GTP + ppGpp) [23]. ppGpp values represent the mean of three assays performed with three independent bacterial cultures.
Cytotoxicity assay
Cytotoxicity assay was performed as described [24]. Vero cells (1 × 105 cells/ml) were grown in 96-well plates in DMEM medium in the presence of 10% FBS for 24 h, at 37 °C, and 5% CO2. Bacteria cytotoxicity was assayed by incubating the Vero cells in the presence of a 1/10 dilution of the bacterial culture supernatant in DMEM with 2% FBS for 72 h at 37 °C and 5% CO2. The supernatant was obtained by growing the strains overnight in LB, followed by centrifugation at 1690 g for 5 min. MTT (Sigma-Aldrich) was used to determine cell viability according to the manufacturer's instructions. Strains EDL933 and DH5-α (a commensal K12 strain) were respectively used as positive and negative controls. Cytotoxicity of the isolates was normalised against the cytotoxic activity of strain EDL933. At least three independent assays were performed for each bacterial strain.
Statistical analysis
The standard error of the mean was calculated according to the formula 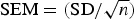 , where s.d. is the standard deviation [25]. The coefficient of variation of the mean was calculated as follows: CVmean = (SEM/mean). Data were evaluated for statistical significance using a two-tailed heteroscedastic Student's t-test.
, where s.d. is the standard deviation [25]. The coefficient of variation of the mean was calculated as follows: CVmean = (SEM/mean). Data were evaluated for statistical significance using a two-tailed heteroscedastic Student's t-test.
RESULTS
Fifty STEC strains were isolated from healthy sheep and identified as described in the Methods section. Of the 50 isolates, 27 were characterised as stx1+, 8 as stx2+ and 15 as stx1+ stx2+ (Table 1). Subtyping of the strains carrying stx2 revealed that all but two belong to the stx2b subgroup. The two remaining strains did not match any known subtype. No isolate carried the eae gene that encodes the adhesin intimin usually found in enterohaemorrhagic E. coli. The isolates were also submitted to multiplex PCR to determine their phylogenetic relationships. Most isolates belonged to phylogroup B1 (41/50) and the remainder to phylogroup A (08/50). One strain displayed an unknown phylogenetic profile. All isolates that belonged to phylogroup A carried only the gene for stx2, while phylogroup B1 was composed of bacteria carrying both stx1 and stx2 or only stx1. Bacterial virulence was evaluated by assaying cytotoxicity against Vero cells. The supernatant of overnight grown bacteria was collected and added to a monolayer of Vero cells, followed by incubation for 72 h, at which time cell survival was recorded (Table 1). On average, the stx2 isolates were the least cytotoxic (mean of 8 strains = 48·0% ± 9·8 dead cells). The stx1+2 isolates (mean of 15 strains = 73·9% ± 3·8 dead cells) and the stx1 isolates (mean of 27 strains = 86·1% ± 3·2 dead cells) displayed considerably higher cytotoxicity. These values are relative to the cytotoxic effect of the EDL933 strain, that was used as a positive control. Two-tailed Student's t-test analyses showed that the cytotoxicity means differed significantly, with P-values of 0·000025 (stx1 against stx2 isolates), 0·022 (stx1 against stx1+2) and 0·035 (stx1+2 against stx2). Figure 1A displays a graphical analysis of the cytotoxicity of each group.
Table 1.
Genetic and phenotypic profiles of 50 STEC isolates
| Strain | Phylogroup | stx1 | stx2 | eae | Cytotoxicity ± s.e.m. (CVmean)a,b | ppGpp basal level ± s.e.m. (CVmean)a,c | ppGpp induced level ± s.e.m. (CVmean)a,d |
|---|---|---|---|---|---|---|---|
| AS1 | A | − | + | − | 85 ± 22·5 (26) | 0·22 ± 0·026 (12) | 0·41 ± 0·012 (3) |
| AS2 | A | − | + | − | 96 ± 6·5 (9) | 0·30 ± 0·019 (6) | 0·41 ± 0·008 (2) |
| AS3 | A | − | +(b) | − | 26·7 ± 6·6 (35) | 0·27 ± 0·020 (7) | 0·42 ± 0·002 (1) |
| AS4 | A | − | +(b) | − | 32·3 ± 7·0 (28) | 0·30 ± 0·015 (5) | 0·33 ± 0·003 (1) |
| AS5 | A | − | +(b) | − | 38·8 ± 9·7 (35) | 0·28 ± 0·006 (2) | 0·38 ± 0·007 (2) |
| AS6 | A | − | +(b) | − | 22 ± 6·1 (27) | 0·14 ± 0·003 (2) | 0·30 ± 0·014 (5) |
| AS7 | A | − | +(b) | − | 49·9 ± 10·8 (28) | 0·18 ± 0·007 (4) | 0·31 ± 0·013 (4) |
| AS8 | A | − | +(b) | − | 33·2 ± 7·5 (29) | 0·18 ± 0·029 (17) | 0·30 ± 0·025 (8) |
| AS9 | B1 | + | +(b) | − | 90 ± 14·7 (16) | 0·12 ± 0·001 (1) | 0·29 ± 0·025 (9) |
| AS10 | B1 | + | +(b) | − | 74 ± 17·7 (24) | 0·13 ± 0·005 (4) | 0·31 ± 0·009 (3) |
| AS11 | B1 | + | +(b) | − | 65 ± 26·1 (40) | 0·13 ± 0·007 (6) | 0·32 ± 0·010 (3) |
| AS12 | B1 | + | +(b) | − | 39·9 ± 5·8 (19) | 0·13 ± 0·004 (3) | 0·34 ± 0·005 (2) |
| AS13 | B1 | + | +(b) | − | 72 ± 16·8 (23) | 0·12 ± 0·004 (4) | 0·31 ± 0·019 (6) |
| AS14 | B1 | + | +(b) | − | 96 ± 14·8 (16) | 0·10 ± 0·002 (2) | 0·25 ± 0·020 (8) |
| AS15 | B1 | + | +(b) | − | 79 ± 19·9 (25) | 0·10 ± 0·006 (7) | 0·28 ± 0·016 (6) |
| AS16 | B1 | + | +(b) | − | 84 ± 8·4 (13) | 0·10 ± 0·007 (8) | 0·20 ± 0·029 (15) |
| AS17 | B1 | + | +(b) | − | 62 ± 5·6 (9) | 0·11 ± 0·003 (3) | 0·27 ± 0·012 (5) |
| AS18 | B1 | + | +(b) | − | 69 ± 21·3 (31) | 0·09 ± 0·011 (13) | 0·17 ± 0·016 (10) |
| AS19 | B1 | + | +(b) | − | 65 ± 3·29 (5) | 0·08 ± 0·004 (5) | 0·29 ± 0·004 (1) |
| AS20 | B1 | + | +(b) | − | 71 ± 22·1 (31) | 0·08 ± 0·003 (4) | 0·29 ± 0·006 (2) |
| AS21 | B1 | + | +(b) | − | 88 ± 19·6 (22) | 0·08 ± 0·005 (8) | 0·27 ± 0·006 (2) |
| AS22 | B1 | + | +(b) | − | 63 ± 20·8 (33) | 0·09 ± 0·001 (2) | 0·27 ± 0·003 (1) |
| AS23 | B1 | + | +(b) | − | 91 ± 4·2 (5) | 0·09 ± 0·004 (5) | 0·32 ± 0·006 (2) |
| AS24 | B1 | + | − | − | 77 ± 13·3 (17) | 0·13 ± 0·016 (13) | 0·23 ± 0·014 (6) |
| AS25 | B1 | + | − | − | 91 ± 18·1 (20) | 0·09 ± 0·009 (11) | 0·26 ± 0·006 (3) |
| AS26 | B1 | + | − | − | 65 ± 17·1 (26) | 0·10 ± 0·002 (3) | 0·30 ± 0·011 (4) |
| AS27 | B1 | + | − | − | 72 ± 19·5 (27) | 0·11 ± 0·005 (5) | 0·29 ± 0·000 (0·1) |
| AS28 | B1 | + | − | − | 66 ± 25·2 (38) | 0·08 ± 0·008 (10) | 0·31 ± 0·016 (5) |
| AS29 | B1 | + | − | − | 88 ± 14·4 (16) | 0·11 ± 0·003 (3) | 0·21 ± 0·032 (15) |
| AS30 | B1 | + | − | − | 113 ± 14·4 (13) | 0·11 ± 0·007 (7) | 0·18 ± 0·002 (1) |
| AS31 | B1 | + | − | − | 95 ± 28·2 (30) | 0·10 ± 0·006 (6) | 0·22 ± 0·015 (7) |
| AS32 | B1 | + | − | − | 100 ± 13·9 (14) | 0·11 ± 0·001 (1) | 0·22 ± 0·011 (5) |
| AS33 | B1 | + | − | − | 83 ± 25·4 (31) | 0·12 ± 0·005 (4) | 0·28 ± 0·024 (9) |
| AS34 | B1 | + | − | − | 87 ± 18·0 (21) | 0·13 ± 0·021 (16) | 0·23 ± 0·013 (6) |
| AS35 | B1 | + | − | − | 79 ± 23·8 (30) | 0·18 ± 0·008 (5) | 0·28 ± 0·014 (5) |
| AS36 | B1 | + | − | − | 83 ± 19·0 (23) | 0·19 ± 0·009 (5) | 0·28 ± 0·009 (3) |
| AS37 | B1 | + | − | − | 86 ± 30·0 (35) | 0·15 ± 0·011 (7) | 0·27 ± 0·007 (3) |
| AS38 | B1 | + | − | − | 96 ± 8·5 (9) | 0·08 ± 0·008 (10) | 0·26 ± 0·007 (3) |
| AS39 | B1 | + | − | − | 90 ± 11·3 (13) | 0·11 ± 0·014 (14) | 0·28 ± 0·009 (3) |
| AS40 | B1 | + | − | − | 91 ± 22·8 (25) | 0·10 ± 0·012 (12) | 0·28 ± 0·014 (5) |
| AS41 | B1 | + | − | − | 59 ± 22·0 (37) | 0·12 ± 0·005 (5) | 0·31 ± 0·011 (4) |
| AS42 | B1 | + | − | − | 79 ± 13·4 (17) | 0·11 ± 0·003 (3) | 0·32 ± 0·014 (4) |
| AS43 | B1 | + | − | − | 50 ± 19·7 (40) | 0·11 ± 0·004 (4) | 0·22 ± 0·026 (12) |
| AS44 | B1 | + | − | − | 115 ± 7·8 (7) | 0·12 ± 0·008 (7) | 0·22 ± 0·011 (5) |
| AS45 | B1 | + | − | − | 74 ± 8·7 (12) | 0·17 ± 0·009 (5) | 0·29 ± 0·007 (2) |
| AS46 | B1 | + | − | − | 116 ± 12·2 (11) | 0·14 ± 0·004 (3) | 0·27 ± 0·008 (3) |
| AS47 | B1 | + | − | − | 93 ± 21·7 (23) | 0·15 ± 0·000 (0·3) | 0·28 ± 0·016 (4) |
| AS48 | B1 | + | − | − | 110 ± 14·2 (13) | 0·16 ± 0·002 (2) | 0·27 ± 0·022 (8) |
| AS49 | Unknown | + | − | − | 89 ± 30·3 (34) | 0·17 ± 0·004 (3) | 0·25 ± 0·010 (4) |
| AS50 | B1 | + | − | − | 77 ± 16·8 (22) | 0·20 ± 0·009 (5) | 0·36 ± 0·022 (6) |
Each values represents the mean ± s.e.m. (standard error of the mean) of three independent experiments.
Cytotoxicity against Vero cells relative to the cytotoxicity displayed by strain EDL933.
ppGpp level of exponentially growing cells.
ppGpp level of phosphate-starved cells.
Fig. 1.

Cytotoxicity and ppGpp levels in 50 STEC isolates. stx1, stx2 and stx1+2 respectively correspond to strains carrying the stx1 gene, the stx2 gene or both stx1 and stx2 genes. (a) Cytotoxicity against Vero cells; (b) ppGpp basal level and (c) ppGpp induced by phosphate starvation. In all three assays, the stx2 group of strains was significantly different from the other two groups (stx1 and stx1+2) with P < 0·05 by Students’ t-tests.
It has been reported that the alarmone ppGpp inhibits DNA replication and lytic development of Stx phages [15]. It has also been shown that the intrinsic levels of ppGpp in the species E. coli are not constant [26]. We thus asked whether the intrinsic variability in ppGpp concentration in STEC isolates would influence the virulence of the strains. If that is the case, strains that display high levels of ppGpp are expected to be less cytotoxic than low-ppGpp ones and vice-versa. To test this assumption ppGpp was assayed in all 50 STEC isolates under two different growth conditions (exponential phase and stationary phase elicited by phosphate starvation). Supplementary Figure S1 shows a representative thin-layer chromatogram of labelled nucleotides isolated from strains AS1, AS2 and AS3. The average ppGpp basal level across all strains (under exponential growth phase) was 0·14 units, ranging from 0·076 to 0·30 units (Table 1). Under phosphate-starvation, the mean ppGpp level increased to 0·28 units (ranging from 0·17 to 0·42). Mean ppGpp basal and induced levels are significantly different (P < 10−5). These results are in agreement with the finding that phosphate starvation induces the accumulation of ppGpp in E. coli [27]. Strains of the stx2 group displayed a significantly higher ppGpp basal and induced (phosphate-starved) levels than those in group stx1 (P = 0·0013 for basal levels and P = 0·0018 for induced levels) or in group stx1+stx2 (P = 0·022 for basal levels and P = 0·0036 for induced levels) (Fig. 1B and C). Overall, there was a moderate, but persistent inverse correlation between cytotoxicity and ppGpp at both basal and induced levels (Fig. 2). Taken together, these results are consistent with the idea that ppGpp plays a negative role in the virulence of STEC natural isolates.
Fig. 2.

Correlation between cytotoxicity and ppGpp level. ppGpp basal (dark grey circles) and induced levels (light grey circles) were plotted against the cytotoxicity of the 50 isolates. The correlation coefficient (r) of cytotoxicity against ppGpp basal and induced levels was 0·32 and 0·43, respectively.
DISCUSSION
Bovines and ovines are natural reservoirs of STEC strains. Both O157 and non-O157 STEC strains were implicated in human disease outbreaks [28, 29]. The main virulence factor of STEC is the production of Shiga toxins, encoded by stx1 and stx2 [2, 30–32]. The majority of STEC strains isolated in the present study carried stx1 or stx1+2 genes and belonged to phylogroups B1 or A. All stx2 strains were in phylogroup A and all stx1 and stx1+2 were in phylogroup B1. These results are in agreement with another study with STEC obtained from Brazilian sheep in which B1 was the prevalent phylogroup and that all STEC isolates belonging to phylogroup A were positive for stx2 and negative for stx1 [33]. Other studies have also shown similar distributions of stx alleles and phylogroups [34, 35].
Cytotoxicity was strongly related to the type of stx present in each isolate, strains harbouring stx1 were generally more cytotoxic than those carrying stx2. The results presented here agree with other studies that have shown that the Stx1 toxin is 10 times more potent than Stx2 against Vero cells, while the LD50 of Stx2 in mice is 100-fold lower than that of Stx1 [36, 37]. The stx1+2 isolates were less cytotoxic than the isolates carrying only stx1. An explanation for this surprising result is that different phages in the same strain can negatively interfere with each other, as reported elsewhere [38, 39]. It is worth mentioning that the two stx2 isolates that did not match a known subtype, AS1 and AS2, displayed high levels of cytotoxicity and ppGpp, contrasting with the inverse relation between these traits generally observed in the other strains.
STEC strains are lysogenic for Shiga toxin-converting lambdoid bacteriophages. The production and release of Stx is preceded by the induction of the bacteriophage, which will ultimately result in cell lysis [15, 40]. Hence, the level of Stx directly depends on the level of phage induction. Under nutrient starvation or in slow-growing bacteria, ppGpp accumulates and the production of lambda phage progeny is hampered [16, 17, 41], resulting in less Stx toxin secretion [15].
The negative effect of ppGpp on prophage induction and phage DNA replication has hitherto been tested only in a single strain, into which relA and relA spoT mutations were introduced [15, 17]. However, ppGpp levels are potentially non-uniform in E. coli under identical growth conditions, meaning that natural isolates differ in their intrinsic ppGpp levels [22, 26]. This species-wide variation has significant implications for bacteria growth and metabolism, because ppGpp controls many metabolic genes as well as gene expression networks [5]. We hypothesised that the inconstancy of ppGpp across E. coli strains would have an impact on the level of Stx production in a random assortment of STEC strains. Particularly, strains that exhibit high levels of ppGpp would produce less toxin than the ones that display low concentrations of this alarmone.
Here, we confirmed that cytotoxicity, which is directly influenced by the production of Stx, is inversely correlated to ppGpp in natural STEC isolates. The correlation is not as strong as would appear from studies with isogenic strains because variations in ppGpp level among natural isolates are never as high as the difference between a relA+ spoT+ strain and its isogenic mutants [15]. In fact, the difference between the strain with the highest and the lowest ppGpp basal level was fourfold, while under ppGpp-inducing conditions this difference was only 2·5-fold. Besides, cytotoxicity is influenced by several inputs, ppGpp being only one of them. Even if ppGpp would be the most relevant factor in Stx production, its effect in an individual strain would also depends on the strain genetic background. For instance, some phenotypes previously attributed solely to ppGpp, such as growth rate control, glycogen accumulation and others actually result from an interplay between ppGpp and RpoS, which is subject to strain variation even stronger than that observed for ppGpp [22].
ppGpp is implicated in the virulence of several bacterial species [8], usually in a positive fashion. In most cases ppGpp is required to fully induce genes associated with virulence [12–14]. Interestingly, STEC stands out as a rare case in which virulence is negatively affected by ppGpp. This divergent result is due to the fact that the main virulence factor of STEC - toxin production, is coupled to phage induction, which is inhibited by ppGpp. By inhibiting phage induction and preventing, thus, cell lysis, ppGpp fulfil its role as a promoter of cell survival under adverse conditions [42].
In conclusion, a set of 50 STEC strains isolated from healthy sheep was evaluated for cytotoxicity, ppGpp and phylogenetic association. As expected for a diverse non-clonal population, there was a high variability in the parameters tested. Most importantly, both cytotoxicity and ppGpp levels displayed a high degree of variation, which were inversely correlated to some extent. In particular, stx2 isolates correlated with a low degree of cytotoxicity and high ppGpp.
ACKNOWLEDGEMENTS
The authors are grateful to Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) for supporting this study. A.E.S. was supported by a FAPESP scholarship. B.S. is a recipient of a productivity scholarship from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Supplementary material
For supplementary material accompanying this paper visit http://dx.doi.org/10.1017/S0950268817001091.
click here to view supplementary material
REFERENCES
- 1.Scheutz F, et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. Journal of Clinical Microbiology 2012; 50: 2951–2963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Melton-Celsa AR. Shiga toxin (Stx) classification, structure, and function. Microbiology Spectrum 2014; 2: EHEC-0024-2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Chiang SM, et al. Phenotypic diversity caused by differential RpoS activity among environmental Escherichia coli isolates. Applied and Environmental Microbiology 2011; 77: 7915–7923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Cashel M, et al. Escherichia coli and Salmonella: cellular and molecular biology, chapter the stringent response. American Society for Microbiology, 1996; 2: 1458–1496. [Google Scholar]
- 5.Traxler MF, et al. The global, ppGpp-mediated stringent response to amino acid starvation in Escherichia coli. Molecular Microbiology 2008; 68: 1128–1148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Fujita C, et al. Guanosine 5′-diphosphate 3′-diphosphate (ppGpp) synthetic activities on Escherichia coli SpoT domains. Bioscience and Biotechnology Biochemistry 2002; 66: 1515–1523. [DOI] [PubMed] [Google Scholar]
- 7.Murray KD, Bremer H. Control of spoT-dependent ppGpp synthesis and degradation in Escherichia coli. Journal of Molecular Biology 1996; 259: 41–57. [DOI] [PubMed] [Google Scholar]
- 8.Dalebroux ZD, et al. ppGpp conjures bacterial virulence. Microbiology and Molecular Biology Reviews 2010; 74: 171–199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kamarthapu V, et al. ppGpp couples transcription to DNA repair in E. coli. Science 2016; 352: 993–996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gentry DR, et al. Synthesis of the stationary-phase sigma factor sigma S is positively regulated by ppGpp. Journal of Bacteriology 1993; 175: 7982–7989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jishage M, et al. Regulation of sigma factor competition by the alarmone ppGpp. Genes and Development 2002; 16: 1260–1270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Aberg A, Shingler V, Balsalobre C. (p)ppGpp regulates type 1 fimbriation of Escherichia coli by modulating the expression of the site-specific recombinase fimB. Molecular Microbiology 2006; 60: 1520–1533. [DOI] [PubMed] [Google Scholar]
- 13.Nakanishi N, et al. ppGpp with DksA controls gene expression in the locus of enterocyte effacement (LEE) pathogenicity island of enterohaemorrhagic Escherichia coli through activation of two virulence regulatory genes. Molecular Microbiology 2006; 61: 194–205. [DOI] [PubMed] [Google Scholar]
- 14.Spira B, Ferreira GM, de Almeida LG. relA enhances the adherence of enteropathogenic Escherichia coli. PLoS One 2014; 9: e91703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Nowicki D, et al. ppGpp-dependent negative control of DNA replication of Shiga toxin-converting bacteriophages in Escherichia coli. Journal of Bacteriology 2013; 195: 5007–5015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Loś JM, et al. Differential efficiency of induction of various lambdoid prophages responsible for production of Shiga toxins in response to different induction agents. Microbial Pathogenesis 2009; 47: 289–298. [DOI] [PubMed] [Google Scholar]
- 17.Nejman-Faleńczyk B, et al. Inhibition of development of Shiga toxin-converting bacteriophages by either treatment with citrate or amino acid starvation. Foodborne Pathogens and Disease 2012; 9: 13–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Clermont O, et al. The Clermont Escherichia coli phylo-typing method revisited: improvement of specificity and detection of new phylo-groups. Environmental Microbiology Reports 2013; 5: 58–65. [DOI] [PubMed] [Google Scholar]
- 19.Vidal M, et al. Single multiplex PCR assay to identify simultaneously the six categories of diarrheagenic Escherichia coli associated with enteric infections. Journal of Clinical Microbiology 2005; 43: 5362–5365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Miller JH. A Short Course in Bacterial Genetics. A Laboratory Manual and Handbook for Escherichia coli and Related Bacteria. New York: CSHL Press, 1992. [Google Scholar]
- 21.Spira B, Yagil E. The integration host factor (IHF) affects the expression of the phosphate-binding protein and of alkaline phosphatase in Escherichia coli. Current Microbiology 1999; 38: 80–85. [DOI] [PubMed] [Google Scholar]
- 22.Spira B, Hu X, Ferenci T. Strain variation in ppGpp concentration and RpoS levels in laboratory strains of Escherichia coli K-12. Microbiology 2008; 154: 2887–2895. [DOI] [PubMed] [Google Scholar]
- 23.Svitil AL, Cashel M, Zyskind JW. Guanosine tetraphosphate inhibits protein synthesis in vivo. A possible protective mechanism for starvation stress in Escherichia coli. Journal of Biological Chemistry 1993; 268: 2307–2311. [PubMed] [Google Scholar]
- 24.Rocha LB, et al. Interaction between Shiga toxin and monoclonal antibodies: binding characteristics and in vitro neutralizing abilities. Toxins (Basel) 2012; 4: 729–747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cumming G, Fidler F, Vaux DL. Error bars in experimental biology. Journal of Cell Biology 2007; 177: 7–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ferenci T, et al. The constancy of global regulation across a species: the concentrations of ppGpp and RpoS are strain-specific in Escherichia coli. BMC Microbiology 2011; 11: 62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Spira B, Yagil E. The relation between ppGpp and the PHO regulon in Escherichia coli. Molecular and General Genetics 1998; 257: 469–477. [DOI] [PubMed] [Google Scholar]
- 28.Dodson K, LeJeune J. Escherichia coli O157:h7, Campylobacter jejuni, and Salmonella prevalence in cull dairy cows marketed in northeastern Ohio. Journal of Food Protection 2005; 68: 927–931. [DOI] [PubMed] [Google Scholar]
- 29.Scallan E, et al. Foodborne illness acquired in the United States – major pathogens. Emerging Infectious Disease 2011; 17: 7–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Fuller CA, et al. Shiga toxin subtypes display dramatic differences in potency. Infection and Immunity 2011; 79: 1329–1337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Obata F, Obrig T. Role of Shiga/Vero toxins in pathogenesis. Microbiology Spectrum 2014; 2: EHEC-0005-2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Shen J, et al. Virulence characterization of non-O157 Shiga toxin-producing Escherichia coli isolates from food, humans and animals. Food Microbiology 2015; 50: 20–27. [DOI] [PubMed] [Google Scholar]
- 33.Martins FH, et al. Diversity of Shiga toxin-producing Escherichia coli in sheep flocks of Paraná state, southern Brazil. Veterinary Microbiology 2015; 175: 150–156. [DOI] [PubMed] [Google Scholar]
- 34.Blanco M, et al. Serotypes, virulence genes, and intimin types of Shiga toxin (verotoxin)-producing Escherichia coli isolates from healthy sheep in Spain. Journal of Clinical Microbiology 2003; 41: 1351–1356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Franz E, et al. Molecular hazard identification of non-O157 Shiga toxin-producing Escherichia coli (STEC). PLoS One 2015; 10: e0120353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tesh VL, et al. Comparison of the relative toxicities of Shiga-like toxins type I and type II for mice. Infection and Immunity 1993; 61: 3392–3402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Russo LM, et al. Comparisons of native Shiga toxins (Stxs) type 1 and 2 with chimeric toxins indicate that the source of the binding subunit dictates degree of toxicity. PLoS One 2014; 9: e93463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Russo LM, Melton-Celsa AR, O'Brien AD. Shiga toxin (Stx) type 1a reduces the oral toxicity of Stx type 2a. The Journal of Infectious Diseases 2016; 213: 1271–1279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Serra-Moreno R, Jofre J, Muniesa M. The CI repressors of Shiga toxin-converting prophages are involved in coinfection of Escherichia coli strains, which causes a down regulation in the production of Shiga toxin 2. Journal of Bacteriology 2008; 190: 4722–4735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Waldor MK, Friedman DI. Phage regulatory circuits and virulence gene expression. Current Opinion in Microbiology 2005; 8: 459–465. [DOI] [PubMed] [Google Scholar]
- 41.Wegrzyn G, Licznerska K, Wegrzyn A. Phage λ-new insights into regulatory circuits. Advances in Virus Research 2012; 82: 155–178. [DOI] [PubMed] [Google Scholar]
- 42.Jain V, Kumar M, Chatterji D. ppGpp: stringent response and survival. Journal of Microbiology 2006; 44: 1–10. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
For supplementary material accompanying this paper visit http://dx.doi.org/10.1017/S0950268817001091.
click here to view supplementary material