

SUMMARY
We report the molecular characterization of methicillin-resistant Staphylococcus aureus (MRSA) with resistance to tigecycline and to daptomycin isolated from intensive-care-unit patients in Brazil over a 6-month period. Thirty-six isolates (25 from infection sites, 11 from nasal sites) recovered from 23 patients who presented with MRSA infection during this period were characterized by pulsed-field gel electrophoresis, multilocus sequence typing, staphylococcal cassette chromosome mec (SCCmec) typing, and antimicrobial susceptibility profiling. Ten isolates from six patients and two isolates from different patients were resistant to tigecycline and daptomycin, respectively. Eight pulsotypes were identified and one, type A, accounted for 21 isolates from 12 patients; type A isolates were SCCmecII as were a further nine isolates of other pulsotypes. All but four of the total isolates were sequence type (ST) 5 or ST105 and classified as clonal complex (CC) 5; the historically prevalent lineage in Brazil, ST239-SCCmecIII, was identified in only three patients. Tigecycline-resistant strains were all ST105-SCCmecII and two patients were nasally colonized by strains of the same pulsotype found in infection sites. Two ST5-SCCmecII were daptomycin resistant after 48 h incubation. The origin and mechanism of these resistant strains remains unknown and further studies are warranted to determine whether such clones are becoming endemic in Brazilian hospitals and to assess their impact on infection control practice.
Key words: Antibiotic resistance, bacterial typing, Staphylococcus aureus
Methicillin-resistant Staphylococcus aureus (MRSA) is a major nosocomial pathogen throughout the world but in Brazil it accounts for more than half of all S. aureus isolates in most hospitals [1].
Several clones of MRSA have spread globally. The prevalent clone in Brazil and wider Latin America [Brazilian epidemic clone (BEC), sequence type 239-staphylococcal cassette chromosome mecIII (ST239-SCCmecIII)] has been identified in 19 Brazilian cities, and in 14 different states between 1995 and 1997 [2], although several other international lineages have also been identified in Brazil [3].
A wide range of resistance to several antimicrobial agents is a hallmark of MRSA but most isolates remain susceptible to vancomycin, teicoplanin and linezolid and these are the primary drugs of choice for the treatment of MRSA infections in Brazil [4]. Two new compounds, tigecycline and daptomycin, were approved for use in Brazil in 2005 and 2008, respectively, for the treatment of complicated skin and skin structure infections and intra-abdominal infections caused by MRSA. Daptomycin is a cyclic lipopeptide targeting the bacterial cell membrane and several diverse mechanisms have been implicated in the development of resistance [5]. Tigecycline, by contrast, belongs to the class of glycylcyclines which have a high affinity for the bacterial ribosome and resistance is apparently linked to overexpression of an efflux pump protein [6].
Two recent surveys of antimicrobial resistance of a large number of bacterial isolates from Latin America and worldwide have confirmed an extremely low frequency or complete absence of tigecycline resistance in S. aureus including MRSA [7, 8]; daptomycin resistance is similarly reported to be uncommon [9]. Tigecycline is not recommended for susceptibility testing of S. aureus by the Clinical and Laboratory Standards Institute (CLSI) [10], although it is listed by the European Committee on Antimicrobial Susceptibility testing (EUCAST) [11]. Since most clinical laboratories in Brazil follow the CLSI recommendations, susceptibility testing is normally restricted to Gram-negative isolates. The finding reported here, of a cluster of MRSA isolates with reduced susceptibility to tigecycline and daptomycin was therefore considered to be significant – warranting further investigation of the antimicrobial resistance and molecular characteristics of the isolates.
The Rizoleta Tolentino Neves Hospital in Belo Horizonte city, capital of Minas Gerais state and the sixth most populous city in Brazil, has 333 beds with a 30-bed adult intensive care unit (ICU) and a five-bed infantile ICU. Twenty-three patients were infected by MRSA between June and December 2009 yielding 25 isolates from infection sites (blood, soft tissues, bone, etc.) and 11 nasal colonizing isolates. All isolates were identified by the Vitek® 2 system (bioMérieux, France) and confirmed as MRSA by mecA PCR and SCCmec element subtyping [12]. Isolates were also screened for Panton–Valentine leukocidin (PVL) by PCR as described previously [13].
The minimum inhibitory concentration (MIC) of oxacillin, tigecycline, teicoplanin, quinupristin-dalfopristin, linezolid, daptomycin and vancomycin for all isolates was determined by Etest® (bioMérieux). In addition, the MIC of vancomycin (Sigma-Aldrich Co., USA), daptomycin (Hospira Inc., USA) and tigecycline (Pfizer Inc., USA) were also evaluated by the broth microdilution method and susceptibility breakpoints for vancomycin and daptomycin were interpreted according to CLSI [10]. Tigecycline interpretive criteria were those established for staphylococci by EUCAST [11], i.e. MIC ⩽0·5 mg/l indicating susceptibility.
All isolates were screened for heterogenous vancomycin-intermediate S. aureus (hVISA) phenotype by one-point population analysis. Overnight cultures in brain heart infusion (BHI) broth were adjusted to 0·5 McFarland turbidity and 100-μl aliquots were spread onto BHI agar plates containing 4 mg/l vancomycin. The plates were read after 24 h and 48 h incubation at 37°C. Isolates were interpreted as being VISA if more than 1000 colonies grew within 48 h and as hVISA if fewer than 1000 colonies grew. Isolates recovered from the screening medium were cultured in BHI broth at 37°C overnight and cells were diluted in saline to the 0·5 McFarland standard; 100 μl of this suspension and serial tenfold dilutions were spread on BHI agar plates with various concentrations of vancomycin (0–8 mg/l) and colonies were counted after 48 h at 37°C.
Isolates were subjected to multilocus sequence typing (MLST) as previously described [14], and the resulting sequences of the seven housekeeping genes were compared to those deposited in the MLST database (http://saureus.mlst.net/site). Pulsed-field gel electrophoresis (PFGE) of SmaI DNA digests was performed and interpreted according to Tenover et al. [15] and analysed with Bionumerics v. 6.5 software (Applied Maths, Belgium) using the unweighted pair-group method with arithmetic mean based on Dice coefficients. Optimization and tolerance values were set to 0·5% and 1·25%, respectively. Isolates were considered to be the same pulsotype if profiles showed greater than 80% similarity with others.
All isolates were resistant to oxacillin and harboured the mecA gene. There was universal susceptibility to linezolid, teicoplanin and vancomycin. Sixteen isolates were classified as intermediately susceptible to quinupristin-dalfopristin (from both infection and carriage sites), and 10 and two isolates, respectively, were resistant to tigecycline and daptomycin (Fig. 1). The latter daptomycin-resistant isolates (MIC 2 mg/l) were isolated from infection sites of different patients and it is of note that they were not detected on subculture of microdilution assays before 48 h incubation although CLSI recommends reading of plates at 24 h. Six patients harboured tigecycline-resistant strains, four with single isolates from infected sites (blood, joint, bone), and two with three and four isolates from both nasal colonization and infections.
Fig. 1.
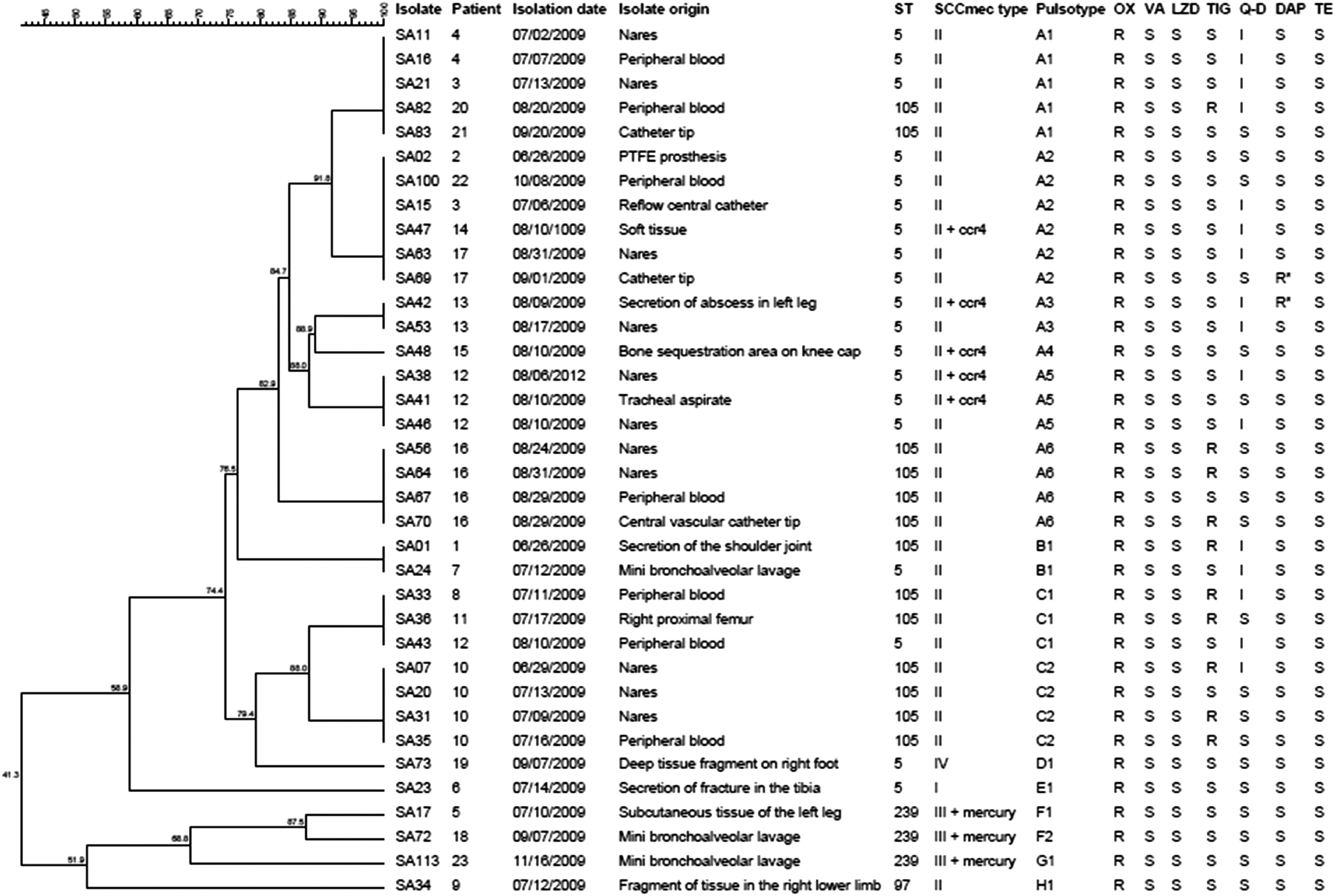
Dendrogram of the PFGE patterns, clinical source, sequence type and SCCmec element, and antimicrobial resistance of MRSA study isolates. ST, Sequence type; OX, oxacillin; VA, vancomycin; LZD, linezolid; TIG, tigecycline; Q-D, quinupristin-dalfopristin; DAP, daptomycin; TE, teicoplanin; R, resistant; I, intermediate level of resistance; S, susceptible; R*, resistant after 48 h; + ccr4, presence of an extra-SCCmec recombinase 4; + mercury, presence of SCC mercury.
Two isolates showed growth on the hVISA screening assay but were not confirmed as the hVISA phenotype on subsequent testing on a range of vancomycin concentrations by comparison with the positive control strain Mu3 of MRSA.
PFGE analysis differentiated the 36 isolates into eight pulsotypes, A–H (Fig. 1). Pulsotype A accounted for 21 (53·8%) isolates from 12 patients and this type was further discriminated into six subtypes by cluster analysis within the 80% similarity index. All pulsotype A isolates were SCCmecII, as were a further nine isolates of other pulsotypes, and were allocated to two closely related sequence types ST5 and ST105 of clonal complex (CC) 5 which also accounted for isolates of pulsotypes B and C. Pulsotypes D and E were also grouped as CC5 lineages, but were of assorted SCCmec types. Tigecycline-resistant strains were all of ST105-SCCmecII and distributed among three pulsotypes from six different patients, two of whom were colonized by strains of the same pulsotype as the infecting strain. Strain genotypes corresponding to international strain lineages were 12 isolates of ST5-SCCmecII (New York/Japan clone), single isolates of ST5-SCCmecI (Cordobes/Chilean clone) and ST5-SCCmecIV (paediatric clone) and three ST239-SCCmecIII isolates (BEC). None of the isolates harboured the genes encoding for PVL.
Although this study involves only one Brazilian hospital, the results reflect the decline in the prevalence of the BEC lineage and its replacement by other international clones, notably CC5 [3]. However, unlike other studies, we have identified a significant aggravating factor of clonal expansion of CC5 strains resistant to tigecycline but markedly less so to daptomycin. Indeed, reports of susceptibility studies of large series of MRSA isolates both within Latin America and worldwide, have identified none or very few isolates to be resistant to tigecycline [7–9]. Therefore the finding of almost one-third of isolates from ICU patients with a MIC of >0·5 mg/l for this drug over just 6 months is strongly suggestive of a nosocomial outbreak within the unit, which to our knowledge has not been previously reported.
The two cases of decreased susceptibility to daptomycin merit comment. Several mechanisms have been cited for daptomycin resistance but Camargo et al. [16] observed that a laboratory-generated strain of S. aureus with reduced susceptibility to daptomycin in vitro, had an altered membrane and cell wall and grew more slowly than the parental strain. Here, daptomycin resistance was detected only after 48 h of incubation which is consistent with a slower growth of such strains.
In the present study, molecular typing showed that at least two patients (nos. 10 and 16, Fig. 1) were first colonized in the nares with tigecycline-resistant strains prior to their recovery in samples from clinical infection. This indicates that colonization with these strains persists and emphasizes the importance and value of surveillance cultures from patients on entry into the ICU in order that efficient control management can be applied by the Infection Control Committee in the nosocomial environment. Further, this could be interpreted as suggesting de novo acquisition of the organisms from other patients or the hospital environment, but the fact that most were CC5 lineages would imply very widespread contamination of the ward environment and there is no evidence to support this.
This study has two significant limitations. First, although there is no evidence that this lineage spread to other units of the hospital, no isolates except for those presented here were available for comparison. Second, it was not possible to investigate whether tigecycline resistance arose as a consequence of treatment with other agents owing to the lack of clinical data. However, given the fact that most isolates were susceptible to the standard agents used to treat MRSA, e.g. linezolid and teicoplanin, it is unlikely that the observed resistance to tigecycline was due to prior antibiotic treatment.
In conclusion, cases of infection by MRSA strains resistant to the newest drugs available and their dissemination are of major concern. Although the mechanisms of resistance remain to be elucidated, this study has enabled the identification of predominant lineages of MRSA associated with infection and colonization and gives an insight into the emergence of clinically significant resistance. Further studies into the prevalent MRSA clones in the wider hospital and region are therefore necessary to determine the potential disease burden and impact on infection control practices and antimicrobial usage to limit their spread.
ACKNOWLEDGEMENTS
We thank Hoberdan Pereira and Hyllo Baeta Marcello Júnior from Geraldo Lustosa Laboratório Médico for sending the MRSA isolates and Simoni Camila Bogni, Waldomiro Thiago Corsi and Rani Hale Dabul for technical assistance. This work was supported by Novartis S.A. and Fundação de Amparo à pesquisa do Estado de São Paulo (2010/02619-0 and A.N.G.D. 2010/11890-9). This study was approved by the Ethics Committee at FCFRP-USP (CEP/FCFRP no. 46/2011).
DECLARATION OF INTEREST
None.
REFERENCES
- 1.Carvalho KS, Mamizuka EM, Gontijo Filho PP. Methicillin/oxacillin-resistant Staphylococcus aureus as a hospital and public health threat in Brazil. Brazilian Journal of Infectious Diseases 2010; 14: 71–76. [DOI] [PubMed] [Google Scholar]
- 2.Oliveira GA, et al. Characterization of the Brazilian endemic clone of methicilin-resistant Staphylococcus aureus (MRSA) from hospitals throughout Brazil. Brazilian Journal of Infectious Diseases 2001; 5: 163–170. [DOI] [PubMed] [Google Scholar]
- 3.Schuenck RP, et al. Staphylococcus aureus isolates belonging to different multilocus sequence types present specific virulence gene profiles. FEMS Immunology & Medical Microbiology 2012; 65: 501–504. [DOI] [PubMed] [Google Scholar]
- 4.Brazilian Health Surveillance Agency (ANVISA). (http://www.anvisa.gov.br/servicosaude/controle/rede_rm/cursos/rm_controle/opas_web/modulo3/gramp_staphylo4.htm). Accessed 15 January 2013.
- 5.Bayer AS, Schneider T, Sahl HG. Mechanisms of daptomycin resistance in Staphylococcus aureus: role of the cell membrane and cell wall. Annals of the New York Academy of Sciences 2013; 1277: 139–158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.McAleese F, et al. A novel MATE family efflux pump contributes to the reduced susceptibility of laboratory-derived Staphylococcus aureus mutants to tigecycline. Antimicrobial Agents and Chemotherapy 2005; 49: 1865–1871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sader HS, Flamm RK, Jones RN. Tigecycline activity tested against antimicrobial resistant surveillance subsets of clinical bacteria collected worldwide (2011). Diagnostic Microbiology and Infectious Disease (in press). [DOI] [PubMed] [Google Scholar]
- 8.Garza-González E, Dowzicky MJ. Changes in Staphylococcus aureus susceptibility across Latin America between 2004 and 2010. Brazilian Journal of Infectious Diseases 2013; 17: 13–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zhanel GG, et al. Antimicrobial susceptibility of 22 746 pathogens from Canadian hospitals: results of the CANWARD 2007–11 study. Journal of Antimicrobial Chemotherapy 2013; 68 (Suppl. 1): i7–i22. [DOI] [PubMed] [Google Scholar]
- 10.Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing; 20th Informational Supplement. CLSI document M100-S20. Wayne, Pennsylvania: Clinical and Laboratory Standards Institute, 2010. [Google Scholar]
- 11.European Committee on Antimicrobial Susceptibility Testing. (http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/Breakpoint_table_v_2.0_120221.pdf). Accessed 15 January 2013.
- 12.Kondo Y, et al. Combination of multiplex PCRs for staphylococcal cassette chromosome mec type assignment: rapid identification system for mec, ccr, and major differences in junkyard regions. Antimicrobial Agents and Chemotherapy 2007; 51: 264–274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lina G, et al. Involvement of Panton-Valentine leukocidin-producing Staphylococcus aureus in primary skin infections and pneumonia. Clinical Infectious Diseases 1999; 29: 1128–1132. [DOI] [PubMed] [Google Scholar]
- 14.Enright MC, et al. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. Journal of Clinical Microbiology 2000; 38: 1008–1015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Tenover FC, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. Journal of Clinical Microbiology 1995; 33: 2233–2239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Camargo IL, et al. Serial daptomycin selection generates daptomycin-nonsusceptible Staphylococcus aureus strains with a heterogeneous vancomycin-intermediate phenotype. Antimicrobial Agents and Chemotherapy 2008; 52: 4289–4299. [DOI] [PMC free article] [PubMed] [Google Scholar]