

Abstract
The aim of this study was to investigate the effects of pyrroloquinoline quinone (PQQ), an oxidoreductase cofactor, on the H2O2-induced premature senescence model in HEI-OC1 auditory cells and to elucidate its mechanism of action in vitro. Cells were treated with PQQ for 1 day before H2O2 (100 μM) exposure. Mitochondrial respiratory capacity was damaged in this premature senescence model but was restored in cells pretreated with PQQ (0.1 nM or 1.0 nM). A decrease in mitochondrial potential, the promotion of mitochondrial fusion and the accelerated movement of mitochondria were all observed in PQQ-pretreated cells. The protein expression of sirtuin 1 (SIRT1) and peroxisome proliferator-activated receptor gamma coactivator-1α (PGC-1α) were significantly decreased under H2O2 exposure while they were increased with PQQ pretreatment, and PGC-1α acetylation was significantly decreased. In conclusion, PQQ has a protective effect on the premature senescence model of HEI-OC1 auditory cells and is associated with the SIRT1/PGC-1α signaling pathway, mitochondrial structure, and mitochondrial respiratory capacity.
Subject terms: Senescence, Organelles
Introduction
Aging is a physiological phenomenon that occurs in all eukaryotes. Cellular senescence manifests as a stable cell arrest with active metabolism, which plays an important role in aging1,2. H2O2 (hydrogen peroxide) is one type of reactive oxygen species (ROS) which has been widely used to achieve oxidative stress-induced premature senescence within a short period of time3,4. In our previous study, short H2O2 exposure led to a significant decrease in the cell population and mitochondrial respiratory capacity and resulted in an imbalance of mitochondrial fusion/fission5. The regulation of aging also relies on ROS function in redox signaling, and the control of oxidative stress is essential6.
Pyrroloquinoline quinone (PQQ) was first reported in 1964 by Hauge et al. as a new coenzyme distinct from nicotinamide adenine dinucleotide (NADH) and flavin in glucose dehydrogenase (GDH)7, and its structure was determined by X-ray analysis in 19798,9. PQQ is contained in fruits and vegetables, such as kiwi fruit, parsley, cocoa powder, fermented soybeans (Japanese natto) and green peppers, and in human breast milk10–12. PQQ is a novel biofactor of physiological importance13, and has been reported to be involved in a variety of biological functions that are clearly beneficial to survival, such as growth and fertility in newborns14. It has also been reported to protect against glutamate-induced cell apoptosis in primary cultured hippocampal neurons15. PQQ is especially effective in neutralizing superoxide and hydroxyl radicals16,17, which cause mitochondrial dysfunction, and it also antagonizes several types of oxidative stress-induced cell damage, including reoxygenation cardiac injury18, chronic heart failure19, ethanol-induced liver damage20, and hyperoxia-induced cognitive deficits21.
PQQ is also involved in the control of redox processes in the mitochondrial respirator chain22, the attenuation of oxidative stress in mitochondria23,24, and the protection of neurons25,26. PQQ not only protects mitochondria from oxidative stress but also promotes mitochondrial biogenesis27. Dietary supplementation of PQQ improves mitochondrial amounts and lipid metabolism in rats28 and has been shown to improve respiratory quotients by increasing the mitochondrial numbers and function in mice29. PQQ prevents rotenone-induced neurotoxicity in Parkinson’s disease models by promoting mitochondrial function and regulating mitochondrial fission and fusion30. In humans, dietary supplementation with PQQ recovers the antioxidant potential, attenuates the inflammatory response, and increases urinary metabolites related to mitochondrial functions31. Considering these findings, the protective effect of PQQ in mitochondrial biogenesis is also expected for inner ear cells.
Mitochondrial biogenesis is mediated by peroxisome proliferator-activated receptor γ coactivator-1α (PGC-1α) under the control of the histone deacetylase sirtuin 1 (SIRT1)32 which has emerged as a crucial regulator of mitochondrial function in vascular smooth muscle33, liver34, kidney35–37, and heart38,39. PQQ has positive effects on biological activities and neural functions in SK-N-SH cells40, HepG2 cells41, and in C57BL/6 mice by increasing PGC-1α expression40. PQQ also stimulates mitochondrial biogenesis through phosphorylation of the cAMP response element-binding protein and an increase of PGC-1α expression in mouse hepatocytes27,42. In humans, the supplementation of PQQ has been reported to improve peak oxygen consumption and impact mitochondrial biogenesis by elevating PGC-1α protein content43. The SIRT1/PGC-1α signaling pathway modulation with the administration of PQQ may also play an important role in protecting mitochondrial function in the inner ear tissues.
This study aims to assess mitochondrial metabolic activity, mitochondrial network structure, mitochondrial motility, and the SIRT1/PGC-1α signaling pathway under PQQ treatment in the immortalized Corti-derived auditory epithelial cell line HEI-OC1 using the H2O2-induced premature senescence model.
Results
High-concentration PQQ-induced cytotoxicity
We first evaluated the cytotoxicity of PQQ in HEI-OC1 cells to confirm the safety of PQQ as a medicine. To achieve this, we examined the cell population doubling rate, metabolic enzyme activity using WST-8 formazan, and the mitochondrial membrane potential (MMP) using the JC-1 fluorescence dye, techniques that have been previously reported for evaluating PQQ and similar compounds in cell lines40,44. The population doubling rate increased significantly compared to the control when the concentration of PQQ was 1.0 nM (P < 0.05), but it then decreased at higher concentrations; the population doubling rate was greater at 1.0 nM compared to that at concentrations greater than 10 nM (Fig. 1A) (P < 0.05). The metabolic enzyme activity was increased significantly compared to the control at PQQ concentrations of 0.1 nM and 1.0 nM, whereas there was no significant difference when the concentration was greater than 10 nM (Fig. 1B). The MMP decreased significantly compared to the control when the concentration was 10 nM or greater (Fig. 1C). These results indicate that, for the HEI-OC1 auditory cells, the optimum concentration of PQQ determined during the evaluation of cell metabolism was 1.0 nM and that PQQ induces cytotoxicity when applied at higher concentrations. The optimum concentration of PQQ we found was similar to those reported previously40, thus we decided to use PQQ concentrations of 0.1 nM and 1.0 nM in the following experiments.
Fig. 1. Effects of PQQ on cell proliferation, cell metabolic activity, and the mitochondrial membrane potential in HEI-OC1 cells evaluated using multiple assays.

A Cell proliferation rate analyzed by cell counting after PQQ pretreatment for 1 day. B Cell metabolic activity measured with the WST-8 formazan assay after PQQ pretreatment for 1 day, shown in optical density (O.D.). The higher value indicates higher metabolic activity. C Mitochondrial membrane potential (MMP) measured with JC-1 fluorescence staining after PQQ pretreatment for 1 day. The ratio of red signal (relative fluorescence units (RFU) at 590 nm; mitochondrial polarized cells) to green signal (RFU at 535 nm; mitochondrial depolarized cells) is shown (n = 5 per group). Box plot shows statistical parameters as follows; central line: median; box limits: first and third quartile; whiskers: minimum and maximum. *P < 0.05, **P < 0.01, ****P < 0.0001.
PQQ pretreatment before H2O2 exposure increased cell proliferation and cell metabolism in HEI-OC1 cells but decreased the mitochondrial membrane potential
H2O2 is widely utilized as an agent to induce premature cellular senescence in cell lines45–47 including HEI-OC1 cells48. We have previously shown that premature cellular senescence was induced in HEI-OC1 cells by exposing them to H2O2 at a concentration of 100 μM for 1 h5. We evaluated the protective effect of pretreatments of 0.1 nM and 1.0 nM PQQ for 1 day on auditory cells with H2O2-induced premature senescence using population doubling rate analysis and metabolic enzyme activity analysis. The cell counts significantly increased in H2O2-exposed cells pretreated with 1.0 mM PQQ, although they remained unchanged in the other conditions (Fig. 2A). There was no significant decrease of viability in H2O2-exposed cells pretreated with PQQ (0.1 mM and 1.0 mM) (Fig. 2B). The population doubling rate decreased significantly in the H2O2-exposed group (with no PQQ pretreatment), whereas pretreatment with 0.1 nM or 1.0 nM PQQ significantly ameliorated the decrease induced by H2O2 exposure (Fig. 2C, D). These results indicate that H2O2 exposure induced premature senescence in HEI-OC1 cells and PQQ pretreatment had a protective effect against H2O2 exposure. H2O2 exposure decreased the metabolic activity of HEI-OC1 cells even when they were pretreated with PQQ, but the decrease in metabolic activity was significantly ameliorated when the cells were pretreated with 1.0 nM PQQ (Fig. 2E).
Fig. 2. Cell proliferation, cell metabolic activity, senescence markers, and mitochondrial membrane potential (MMP) in HEI-OC1 cells with premature cellular senescence induced by H2O2 exposure with and without PQQ pretreatment.

The effects of PQQ under H2O2 exposure are evaluated using multiple assays in the same manner as shown in Fig. 1. A Cell count 1 day after H2O2 exposure. B Cell viability 1 day after H2O2 exposure. C Cell proliferation rate using total cell count for 3 days after H2O2 exposed. D Cell proliferation rate using live-cell count for 3 days after H2O2 exposure. E Cell metabolic activity measured with WST-8 formazan assay. F Relative mRNA expressions of Cdkn1a (p21), Cdkn2a (p16), and Trp53 (p53) using β-actin as an internal control. RQ relative quantification. G The changes of senescence-associated β-galactosidase (SA-β-Gal) using SPiDER-βGal staining in flow cytometry analysis. H The changes of dipeptidyl peptidase DPP4 (CD26) cell surface protein marker in flow cytometry analysis. I The increased ratio in positivity rate over control of SPiDER-βGal positive cells. J The increased ratio in positivity rate over control of CD26 positive cells. K Imaging of mitochondria stained with JC-1. Green signal indicates low MMP and red signal indicates high MMP. Scale bar, 10 µm. L MMP measured with JC-1. (A–D, L: n = 6 per group, E, I, J: n = 5 per group, F: n = 3 per group). Box plot shows statistical parameters as follows; central line: median; box limits: first and third quartile; whiskers: minimum and maximum. RQ data are shown as mean ± standard deviation. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
We also evaluated the expression of cell cycle regulators (Cdkn1a (p21WAF1/CIP1), Cdkn2a (p16INK4A), and Trp53 (p53)), senescence-associated β-galactosidase (SA-β-Gal), and the dipeptidyl peptidase DPP4 (CD26) cell surface protein as cellular senescence biomarkers. The expression of the primary senescence markers p21, p16, and p53 are associated with cell cycle arrest in mediating senescence and the senescent phenotype49,50. SA-β-Gal is widely utilized as a senescence marker in aging research51–53, and has also been previously evaluated in HEI-OC1 cells2,54. CD26 is recently reported as a senescence marker55,56, and is emerging as a therapeutic target57. While H2O2 exposure increased the expression of p21 and the activity of SA-β-Gal and CD26, the expression of p21, p16, and p53 were all alleviated by 0.1 nM PQQ pretreatment (Fig. 2F–J). These results also support the theory that H2O2 exposure induces premature senescence and that PQQ pretreatment protects HEI-OC1 cells from H2O2-induced premature senescence.
The MMP was decreased by exposure to H2O2 and more significantly decreased when the HEI-OC1 cells were pretreated with PQQ (Fig. 2K, L). These results suggest that PQQ pretreatment protects HEI-OC1 cells from H2O2-induced premature senescence and that PQQ has a mitochondrial uncoupling effect. The mitochondrial reactive oxygen species (ROS) evaluated with MitoSOX Red was not decreased by exposure to H2O2 whereas PQQ pretreatment significantly increased the ROS level (Supplementary Data). These results indicate that PQQ mildly increases the ROS levels, which can help to maintain the regulation of ROS levels in HEI-OC1 cells and so protect against premature senescence.
PQQ alleviated ultrastructural damage of mitochondria induced by exposure to H2O2 in HEI-OC1 cells
We also evaluated the protective effect of PQQ pretreatment on auditory cells with H2O2-induced premature senescence using transmission electron microscopy (TEM) image analysis. The shape of the mitochondria under TEM in the control cells was normal, but the mitochondria in the H2O2-exposed cells showed ultrastructural damage, including typical matrix swelling, the loss of cristae in the majority of mitochondria, and the increased formation of autophagosomes (Fig. 3A). The mitochondrial damage was alleviated by PQQ pretreatment before H2O2 exposure; PQQ pretreatment improved the internal heterogeneity and the swollen shape of the mitochondria and increased the number of endosomes. Image analysis of the area of endosomes showed that the increase in the number of endosomes and lysosomes was significant in the 0.1 nM PQQ-pretreated group compared to the H2O2-exposed group with no PQQ pretreatment (Fig. 3B). In contrast, the number of autophagosomes was significantly increased in the H2O2-exposed group, whereas PQQ pretreatment suppressed this increase. These morphological findings indicate that PQQ alleviates ultrastructural damage to the mitochondria of HEI-OC1 cells induced by a short exposure to H2O2. In addition, PQQ pretreatment increased the number of both endosomes and lysosomes, which implies an acceleration of autophagy or the inhibition of lysosome function (Fig. 3B).
Fig. 3. Mitochondrial ultrastructure in HEI-OC1 cells exposed to H2O2 (100 µM) with and without PQQ pretreatments (0.1 nM and 1.0 nM), and control cells.

A Short exposure to H2O2 induces fine structural damage to mitochondria, which is alleviated with PQQ pretreatment. From left to right: control cells, cells exposed to H2O2, cells exposed to H2O2 after 0.1 nM PQQ pretreatment, and cells exposed to H2O2 after 1.0 nM PQQ pretreatment. AP autophagosome, DM damaged mitochondria, E endosome, ER endoplasmic reticulum, GL Golgi body, L lysosome, M mitochondria, N nucleus. Scale bar, 500 nm in top row, 200 nm bottom row. B The numerical analysis of TEM images. The endosome ratio, damaged mitochondria number, autophagosome number, and lysosome number were calculated by visual inspection (n = 5 per group). Box plot shows statistical parameters as follows; central line: median; box limits: first and third quartile; whiskers: minimum and maximum. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
The area of total mitochondria including healthy mitochondria and damaged mitochondria showed no significant difference between groups (Fig. 3B). The copy numbers of the mitochondrial DNA did not show a significant difference between groups (Supplementary Data), which indicated that the changes under the exposure to H2O2 and the treatment of PQQ mainly causes morphological and structural changes in mitochondria but not the proliferation of mitochondria in the timeframe of this study. These results indicate that H2O2 exposure or PQQ pretreatment does not alter mitochondrial proliferation but causes damages or protection in mitochondrial structure.
PQQ protected against the decline of mitochondrial respiratory capacity in the oxidative stress-induced premature senescence of HEI-OC1 cells
In order to confirm the protective effect of PQQ pretreatment in mitochondrial biogenesis and metabolic function in HEI-OC1 cells, we evaluated the oxygen consumption rate (OCR) and the extracellular acidification rate (ECAR) using a XF24 extracellular flux analyzer, which measures the fundamental parameters of the electron transport chain (ETC), basal OCR, ATP-linked respiration, maximal OCR, spare respiratory capacity and proton leak (Fig. 4A, B). The basal ATP production speed did not differ significantly between groups; however, the maximal respiration measured following the injection of a mitochondrial uncoupling agent (carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP)) decreased significantly in the H2O2-exposed group compared to the control group, while it was protected significantly in both the 0.1 nM and 1.0 nM PQQ-pretreated groups compared to the H2O2-exposed group (Fig. 4C). ECAR tended to increase only in the H2O2-exposed group, although the change was not significant (Fig. 4B, D). ATP production speed represented by OCR, ECAR, and phosphate/oxygen ratio (P/O ratio) showed a slight increase of glycolytic ATP production under H2O2 exposure which was alleviated by PQQ pretreatment, although the difference was not significant between groups (Fig. 4D). These results indicate that PQQ prevents the decline of mitochondrial respiratory capacity under H2O2 exposure in HEI-OC1 cells.
Fig. 4. Mitochondrial biogenesis in the premature cellular senescence model with and without PQQ treatment.

The effects of PQQ pretreatment in HEI-OC1 cells exposed to H2O2 were evaluated using a XF24 flux analyzer. A Oxygen consumption rate (OCR) diagram of the XF assay, normalized by the number of cells. B Extracellular acidification rate (ECAR) diagram of the XF assay. C ATP production rate quantified with OCR measured following the injection of oligomycin, maximal respiration rates measured following injection of FCCP, and cell energy phenotype profile of the XF assay. D Basal ATP production rate and ATP rate index of the XF assay (n = 5 per group). Data are shown as mean ± standard deviation. *P < 0.05.
The maximal respiration after the injection of a mitochondrial uncoupling agent indicates the mitochondrial respiratory capacity or reserve capacity, which works effectively in response to high-energy demand or oxidative stress58,59. The mechanism of improvement in respiratory capacity mainly concerns the ETC because the medium in this assay contains glucose and pyruvic acid, but not glutamate or fatty acid (Supplementary Information). To further confirm the substrates on which HEI-OC1 cells depend, the metabolism regarding fatty acid oxidation (FAO) and glutaminolysis were analyzed. The palmitic acid supplementation increased the OCR whereas respiratory capacity was decreased; indicating that fatty acid worked as an uncoupler rather than an oxidation substrate, and fatty acid was not mainly utilized in HEI-OC1 cells (Fig. 5A). Carnitine palmitoyltransferase (CPT) inhibitor, Etomoxir, decreased respiratory capacity without PQQ pretreatment, but the decrease was not significantly different compared to that with PQQ treatment, indicating that FAO was not the main pathway in the recovery of respiratory capacity (Fig. 5B). Glutamine supplementation increased the OCR and also respiratory capacity, although the protective effect of PQQ was still present under glutamine supplementation (Fig. 5C). Glutaminase (GLS) inhibitor, (bis-2-(5-phenylacetamido-1,3,4-thiadiazol-2-yl)ethyl sulfide (BPTES), disabled the protective effect by PQQ pretreatment (Fig. 5D), indicating that glutaminolysis is one of the alternative substrate pathways to protect damaged mitochondrial in HEI-OC1 cells. Other mechanisms including mitochondrial structural changes, however, may underlie in respiratory capacity recovery effect with PQQ pretreatment, because glutamine supplementation did not fully recover respiratory capacity. The expression of CPT1C (Carnitine palmitoyltransferase 1C) was increased significantly under the exposure to H2O2 compared to the control group, which indicates that the energy metabolism was promoted to FAO from main glucose metabolism under the exposure to H2O2, whereas these expressions were nearly completely recovered to control level by PQQ pretreatment (Fig. 5E). The expression of glutaminase 2 (GLS2) was also increased significantly under the exposure to H2O2 compared to the control group, but the increase of GLS2 under PQQ pretreatment did not significantly different compared to the control group (Fig. 5E). The increase in GLS2 expression did not result in an increase in OCR under glutamine treatment, suggesting that the increase in GLS2 expression may indicate stress to the cells (Fig. 5F). These results indicate that the metabolic shift from glucose substrate-dependent pathway to FAO or glutaminolysis was induced under the exposure to H2O2 regarding the mitochondrial damage, whereas the promotion was not enough to compensate respiratory capacity decrease and was rather a marker of H2O2-exposed stress, and this shift was held to the nearly normal state with PQQ pretreatment protection.
Fig. 5. Substrate-specific mitochondrial biogenesis in the premature cellular senescence model with and without PQQ treatment.

The substrate-specific effects of PQQ pretreatment in HEI-OC1 cells exposed to H2O2 were evaluated using a XF24 flux analyzer; A Oxygen consumption rate (OCR) under palmitic acid supplementation. B OCR under Cpt (Carnitine palmitoyltransferase) inhibition by Etomoxir. C OCR under glutamine supplementation. D OCR under Gls (Glutaminase) inhibition by BPTES ((bis-2-(5-phenylacetamido-1,3,4-thiadiazol-2-yl)ethyl sulfide). E Relative mRNA expressions of Cpt1a, Cpt1c, Cpt2, Gls1, and Gls2 using β-actin as an internal control. F OCR under glutamine supplementation in HEI-OC1 cells in the control group or H2O2-exposed group (n = 5 per group). Data are shown as mean ± standard deviation. *P < 0.05.
PQQ has the potential to protect against morphological and dynamic deteriorations in HEI-OC1 cells with short exposures to H2O2
We evaluated the structure and dynamic motility of the mitochondria to confirm the improvements in mitochondrial function under PQQ pretreatment. The HEI-OC1 cells were stained with TMRE fluorescence dye, which accumulates in mitochondria due to the MMP, and were observed under confocal microscopy. The early alterations in mitochondrial shape and motility represent the dynamic function of mitochondria in auditory cells5. The control group had a good balance of mitochondrial fusion and fission, while fragmented mitochondria with a balloon-like shape or hyperfused mitochondria with an enlarged shape were observed in the H2O2-exposed group (Fig. 6A). In the 0.1 nM or 1.0 nM PQQ-pretreated groups, the mitochondrial shapes had partly recovered to the normal balance (Fig. 6A). In order to quantify the recovery of the mitochondrial structures, we processed the mitochondrial images and analyzed the specific parameters of the mitochondrial network nodes (Fig. 6B)5. The mitochondrial dynamics include different types of fusion and fission mechanisms of the network nodes (tip-to-tip, tip-to-side, or side-to-side) (Fig. 6C), and the specific parameters show the condition of mitochondrial dynamics (Fig. 6D). The number of branches, the number of junctions and the average branch length all increased significantly in 0.1 nM and 1.0 nM PQQ-pretreated groups compared to the H2O2-exposed group; however, mitochondrial dynamics biased toward the fusion process was observed in the 0.1 nM or 1.0 nM PQQ-pretreated groups (Fig. 6E). These results indicate that PQQ pretreatment assists in the recovery from morphological damage caused to the mitochondria by H2O2 exposure and also aids the recovery of mitochondrial dynamics in HEI-OC1 cells.
Fig. 6. The morphological evaluation of mitochondria in the premature cellular senescence model under PQQ pretreatment.

The morphological changes with PQQ pretreatment in HEI-OC1 cells exposed to H2O2 to induce premature cellular senescence were evaluated using microscopic imaging. A Mitochondrial images stained with MitoTracker Orange CMTMRos. Scale bar, 10 µm. B The image processing flow of the skeletonized image analysis. C The different types of dynamic mitochondrial regulations including fusion and fission mechanisms involving network nodes (tip-to-tip, tip-to-side, or side-to-side). Graph representation of the mitochondrial reticulum using four node types: 1 (magenta), 2 (green), 3 (dark blue), and 4 (blue). D The descriptions of parameters in the mitochondrial network morphology. E The number of branches, junctions, and the average length of branches in each image (E: n = 5 per group; repeated experiments). Box plot shows statistical parameters as follows; central line: median; box limits: first and third quartile; whiskers: minimum and maximum. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
For the analysis of mitochondrial motility, particle tracking was derived from spatiotemporal image sequences, and then the kinetics of the moving particles were evaluated quantitatively using the TrackMate plugin in ImageJ/FIJI and @msdanalyzer on the MATLAB platform (Fig. 7A, B, E). The mean square displacement (MSD) was calculated from the tracking of the mitochondria, and the area under the curve (AUC) of the MSD plot of each group is shown in Fig. 7C, D. The mitochondrial motility in the short-exposure H2O2 group tended to decrease compared to the control group, while it increased in the PQQ-pretreated groups. The AUC of the MSD plot showed significant increases in the PQQ-pretreated groups compared to the short-exposure H2O2 group (Fig. 7D). These results also support our proposal that PQQ pretreatment protects mitochondrial dynamics under H2O2 exposure in HEI-OC1 cells.
Fig. 7. The dynamic evaluation of mitochondrial motility in the premature cellular senescence model under PQQ pretreatment.

The changes in mitochondrial dynamics with PQQ pretreatment in HEI-OC1 cells exposed to H2O2 to induce premature cellular senescence were evaluated using microscopic imaging and image processing. A Image preprocessing flow of the motility analysis. B Motility analysis of the mitochondria. C The average of the mean square displacement (MSD) of each group. D The area under curve (AUC) of the MSD of each group. E Mitochondrial motion tracking video. The mitochondrial particle tracking lasted for 3 min in 3 s of intervals under ImageJ (D: n = 5 per group; repeated experiments). Box plot shows statistical parameters as follows; central line: median; box limits: first and third quartile; whiskers: minimum and maximum. *P < 0.05.
Pretreatment with PQQ increased the expression of SIRT1 and PGC-1α in H2O2 short exposure HEI-OC1 cells, inducing deacetylation of PGC-1α
To explore the pathways by which PQQ pretreatment led to the improvement of mitochondrial function, we investigated changes in the expression of SIRT1 and PGC-1α involving the regulation of mitochondrial metabolism. The expression of SIRT1 and PGC-1α decreased significantly in the H2O2 short-exposure group, while it recovered significantly in the 1.0 nM PQQ-pretreated group (Fig. 8A–C, G). In addition, the acetylation of PGC-1α increased significantly in the H2O2 short-exposure group, while it attenuated significantly in the 1.0 nM PQQ-pretreated group (Fig. 8D, E). No significant change was observed in the mRNA expression levels of SIRT1 and PGC-1α and there was no correlation seen between the protein and mRNA expression (Fig. 8F). These results indicate that other post-translational modification factors including microRNA (miRNA) might regulate SIRT1 and PGC-1α at the transcriptional level60–63. These results indicate that H2O2 short exposure decreases the protein expression of SIRT1 and PGC-1α, while PQQ pretreatment increases the protein expression of SIRT1 and PGC-1α under H2O2 short exposure in HEI-OC1 cells, inducing the deacetylation of PGC-1α.
Fig. 8. Analysis of protein expression and protein acetylation in the premature cellular senescence model under PQQ pretreatment.

A SIRT1 and PGC-1α protein expression was analyzed by Western blotting (upper, middle) using β-actin as a loading control (lower). B Relative protein expression level of SIRT1 normalized by β-actin. n = 10 per group. C Relative protein expression level of PGC-1α normalized by β-actin. D Acetylation of PGC-1α analyzed by immunoprecipitation. E Relative acetylation level of PGC-1α normalized by PGC-1α. F Relative mRNA expressions of GAPDH, SIRT1, and PGC-1α using β-actin as an internal control. G The changes of SIRT1 expression in flow cytometry analysis. H The rate of SIRT1 positive cells (B, C, E, H: n = 5 per group, F: n = 3 per group; repeated experiments). Box plot shows statistical parameters as follows; central line: median; box limits: first and third quartile; whiskers: minimum and maximum. RQ data are shown as mean ± standard deviation. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, *****P < 0.00001.
Discussion
In this study, we demonstrated that PQQ has the potential to protect against oxidative stress-induced premature cellular senescence in auditory cells by enabling the recovery of mitochondrial function by restoring the regulation of SIRT1/PGC-1α, the protein expression of SIRT1, and the deacetylation of PGC-1α, and by facilitating the recovery of mitochondrial biogenesis including the ATP production rate and maximum respiration rate. Restored SIRT1 leads to the mediation of PGC-1α and mitochondrial biogenesis in the auditory cells with oxidative stress-induced premature senescence (Fig. 9). To our knowledge, this is the first report which indicates the effect of PQQ in protecting against the oxidative stress-induced premature cellular senescence of auditory cells.
Fig. 9. Schematic diagram of the mechanism of HEI-OC1 mitochondrial biogenesis modulation under H2O2 exposure and PQQ protection.
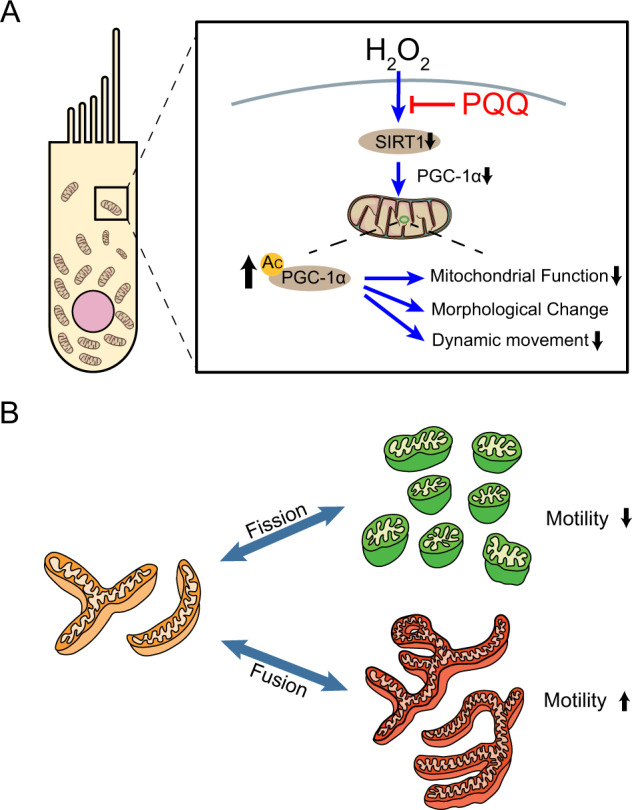
A Sirtuin 1 (SIRT1) expression and deacetylation activity are decreased in HEI-OC1 cells by H2O2 exposure. PGC-1α mediates the protective effect of SIRT1 expression and mitochondrial biogenesis and function. PQQ improves mitochondrial biogenesis and function by restoring SIRT1 expression and deacetylation activity under H2O2 exposure in HEI-OC1 cells. B Schematic diagram of mitochondrial morphological changes.
First, we confirmed the mitochondrial morphology and dynamics in auditory cells for evaluating the effect of PQQ pretreatment because these two changes represent the condition of the mitochondrial quality control system64, and previous reports indicate that PQQ improves reproduction, neonatal development, and mitochondrial function in rats by mechanisms that involve mitochondrial-related signaling pathways28. The acceleration of autophagy function was indicated in the PQQ-pretreated groups by TEM images. In addition, the increase of the fusion of autophagosome and lysosome in the degradation step of autophagy, leading to the formation of autolysosome, was also presumed by TEM image analysis. PQQ could ameliorate autophagy-dependent apoptosis via the lysosome-mitochondria axis in vascular endothelial cells65, but because these functions interact closely with each other, it is difficult to precisely separate them and evaluate their functions, so further research is needed. In addition, we focused on mitochondrial motility in auditory cells because it decreases with age in vascular smooth muscle at the same time as mitochondrial dysfunction66. In this study, we confirmed the improvement of mitochondrial morphology as well as mitochondrial motility with PQQ pretreatment in the oxidative stress-induced premature senescence of auditory cells, which supports the protective aspect of PQQ in the mitochondrial quality control system.
Mitochondrial function is dependent on mitochondrial structure and mitochondrial dynamics, including fission, fusion, and motility67. The mitochondrial transport to discrete subcellular regions may contribute to the accumulation of reactive oxygen species (ROS) in the nucleus, and the oxidative-based signaling function68. The fusion of mitochondria is dependent on the mitochondrial inner membrane potential and independent of microtubules or actin in neurons, and it directly reflects the mitochondrial condition69. The anterograde mitochondrial transport delivers healthy mitochondria to peripheral sites, while the retrograde mitochondrial transport returns damaged mitochondria to central sites, maintaining mitochondrial quality control, and the mitochondrial trafficking is crucial for neural survival70. Molecular pathways that control mitochondrial movement can be reduced to their effects on the balance of forces that act on mitochondria, driving and opposing movement71. The immobilization of mitochondria to microtubules is mediated by syntaphilin, an axonal molecule that facilitates the increase in mitochondrial volume in demyelinated axons, facilitating their survival and protecting against axonal degeneration in the central nervous system72. Mitochondrial motility and dynamics both regulate and reflect mitochondrial function in auditory cells5. The morphological and dynamic deteriorations caused by H2O2 exposure were alleviated by PQQ pretreatment in HEI-OC1 cells, which indicates that PQQ works protectively in terms of the mitochondrial function and the mitochondrial condition. The recovery of mitochondrial morphology and dynamics is the first step in the protection of mitochondrial metabolic activity in auditory cells.
The increase of expressions of FAO and glutaminolysis was observed under the exposure to H2O2, which was not enough to recover mitochondrial biogenesis, and this increase was reverted to nearly normal state by PQQ pretreatment. The induction of FAO or glutaminolysis is reported to be one of the phenotype of cellular senescence; CPT1A is overexpressed in senescent placenta-derived mesenchymal stem cells, the inhibition of CPT1A induced a change of energy metabolism and reversed senescence73, and GLS1 was identified as an essential gene for the survival of senescent cells56. The increase of expressions in FAO or glutaminolysis-related genes can draw the condition of H2O2-induced premature cellular senescence. This increase was inhibited by PQQ pretreatment, which indicates that PQQ have the protective function on mitochondria. Glutaminolysis was one of the alternative substrate pathways to protect damaged mitochondrial in HEI-OC1 cells under PQQ pretreatment, although other mechanisms including structural changes of mitochondria may underlie in respiratory capacity recovery effect with PQQ pretreatment. It has been reported that changes in mitochondrial morphology are related to the respiratory capacity of mitochondria74, indicating that mitochondrial structure is an important aspect of respiratory capacity protection. The mitochondrial respiratory capacity or reserve capacity respond to high-energy demand or oxidative stress and acts as a buffer to protect the cells58,59.
The speed of glucose consumption varies between cell lines, and the drift of the OCR in this timeframe is also observed in normal condition, indicating that the metabolism of HEI-OC1 cells is very high to decrease the glucose concentration and OCR in this timeframe. The glucose concentration in the DMEM medium during the culture of HEI-OC1 cells decreases from the initial 450 mg/dL to about 250 mg/dL within 5–6 days (Supplementary Information). This corresponds to a decrease of about 50% in 120 h and about 2% in 4 h. During the analysis of XF24, where the medium volume is very low, this decrease in glucose concentration and OCR becomes more prominent and is likely to lead to a further decrease in OCR over time during the analysis timeframe.
Importantly, the PQQ pretreatment led to the restoration of the expression of SIRT1 and PGC-1α and the deacetylation of PGC-1α in the oxidative stress-induced premature senescence of auditory cells. These results suggest that PQQ regulates mitochondrial homeostasis via SIRT1 and the deacetylation of PGC-1α in auditory cells, preventing premature senescence75. Recent studies suggest that PQQ activates SIRT1 and SIRT3 genes and increases NAD + activity in the human hepatocyte cell line HepG241, induces deacetylation of PGC-1α and enhancement of mitochondrial activity in NIH/3T3 cells after treatment with a SIRT1 selective inhibitor76, and protects skeletal muscles from denervation-induced atrophy by activating PGC-1α and improving the energy metabolic profile in C57BL6/J mice77. These effects of PQQ activating the SIRT1/PGC-1α signaling pathway in various cells support our data, including the protective effect of PQQ for the inner ear. In addition, the age-related decrease in the expression of SIRT1 and PGC-1α in cochlear tissue is observed in C57BL/6 mice, and the overexpression of SIRT1 suppressed apoptosis and promoted cell proliferation in HEI-OC1 cells78. The activation of PGC-1α promoted mitochondrial biogenesis and protected them against cisplatin (CDDP)-induced ototoxicity in HEI-OC1 cells79. Even caloric restriction, which has an anti-aging effect, induces SIRT1 expression and improves PGC-1α expression, which can regulate energy homeostasis and extend lifespan80. The mRNA expression levels of SIRT1 and PGC-1α did not correlate with the protein expression levels in this study, indicating post-translational modification factors in the protective pathway of PQQ pretreatment. We consider that the expression of SIRT1 might be also regulated by microRNA (miRNA) in auditory cells, based on the results of previous studies60–63.
In this study, we confirmed that PQQ pretreatment had a mitochondrial uncoupling effect (the MMP decrease) in the high-concentration condition, and it increased cell viability and worked protectively under H2O2 oxidative stress (Fig. 3C, D). Indeed, the decrease of MMP by PQQ has been reported in tumor cell lines at high doses of over 15 μM44. The optimum concentration analysis is crucial in evaluating the protective effect of PQQ, and from the aspect of the drug screening targeting the mitochondrial protection the mitochondrial uncoupling effect is one of the key factors in reducing damage and protecting the mitochondria in relation to ROS. We selected the optimum concentration of PQQ in this study by determining the concentration at which there was no decrease in cell numbers and no decrease in mitochondrial membrane potential. In a study evaluating the cardioprotective effect of FCCP (a mitochondrial uncoupler) in the post-ischemic functional recovery of rat hearts, the optimum concentration was also determined based on the mitochondrial oxidation without mitochondrial membrane depolarization81, and the protective pathway was ROS-dependent82. Generally, the MMP shows the condition of the mitochondria, and low membrane potential is normally considered to reflect the damaged condition; however, the MMP uncoupling effect of compounds should be interpreted carefully. The mitochondrial uncouplers including intrinsic uncoupling proteins, natural uncoupling compounds or newly developed agents which are reported to have a protective effect in mitochondria against ROS damage83. UCP2 and UCP3, homologues of the endogenous mitochondrial uncouplers expressed in the heart, also protect against mitochondrial oxidative damage induced with cardiac ischemia-reperfusion by reducing the production of ROS84. FCCP reduces brain edema, decreases neuroinflammation, and improves neurological deficits following intracerebral hemorrhage by activating AMPK85. The newly discovered agent Ppc-1 functions as a mitochondrial uncoupler, and stimulates adipocytes to release fatty acids, thus acting as an anti-obesity agent86,87. Based on these previous studies, the higher respiratory flux leads to consumption of the membrane potential component of the proton motive force by ATP synthase which leads to the generation of ATP producing a mitochondrial membrane depolarization which can be one of the key factors underlying the protective mechanisms of PQQ under H2O2 exposure. Elevated mitochondrial ROS production promotes insulin resistance and obesity88, and the anti-obesity effect is also important in considering ROS protective agents. Low concentration of a mitochondrial uncoupler induces mild ROS production and PQQ extends the lifespan by increasing endogenous ROS levels89. Quercetin, one of the antioxidant polyphenols, is also known to protect neural and cardiac tissues through its mild mitochondrial uncoupling function90–92. ROS can act as essential signaling molecules to promote metabolic health and longevity93. The mild mitochondrial uncoupling effect and the mild ROS production could be key factors in the protective pathway of PQQ. Based on these previous reports and the results in this study, we consider that the decrease in MMP dissipation at high doses of PQQ might be related to decreased cellular ATP levels and the enhanced regulation of ROS relating to apoptosis44. Previous reports have also suggested that the mitochondrial membrane permeability transition is a critical step in the induction of intrinsic apoptosis and often represents the earliest apoptotic signal94, and that the loss of mitochondrial ATP synthesis in apoptotic cells and the increase of inter-membrane creatine phosphate concentrations might be consequences of the loss of MMP caused by the translocation of proapoptotic BH3-only proteins to the mitochondria95. Because ROS can alter the expression level of many miRNAs96 and SIRT1 expression is also regulated by the miRNA pathway in oxidative stress60, ROS regulation of the uncoupling effect of PQQ can work protectively, through the miRNA post-translational modification pathway, in the oxidative stress-induced premature senescence of auditory cells.
This study demonstrated that PQQ protected mitochondrial function in HEI-OC1 auditory cells with stress-induced premature cellular senescence. The cell numbers and cell viability were protected from H2O2-induced cellular senescence by PQQ, which attenuated both H2O2-induced mitochondrial respiration decreases, as well as H2O2-induced morphological and dynamic movement changes. The mechanism underlying these findings may be associated with SIRT1, PGC-1α, and deacetylation of PGC-1α.
Methods
Cell culture and culture conditions
House Ear Institute-Organ of Corti 1 (HEI-OC1) cells derived from the auditory organ of the transgenic mouse ImmortomouseTM, which harbors a temperature-sensitive mutant of the SV40 large T antigen gene under the control of an interferon-γ-inducible promoter element. HEI-OC1 cells represent a common progenitor for sensory and supporting cells of the organ of Corti97,98. HEI-OC1 cells provided by Professor F. Kalinec (UCLA, Los Angeles, CA, USA), were cultured in high-glucose Dulbecco’s Eagle’s medium (DMEM) (21063; Life Technologies, Inc, NY., USA) containing 10% fetal bovine serum (FBS) (Life Technologies, Inc, NY., USA) with 0.06% w/v penicillin (Nacalai Tesque, Kyoto, Japan). Cells were incubated at 33 °C with 10% CO2 under permissive conditions.
PQQ treatments, cell viability assay and population doubling rate analysis
The HEI-OC1 auditory cells were incubated with PQQ (pyrroloquinoline quinone disodium salt, Mitsubishi Gas Chemical Co., Inc., Tokyo, Japan) in the culture medium for 1 day before H2O2 exposure. The HEI-OC1 auditory cells (4 × 104 cells/mL/well of 12-well plates) were washed with Dulbecco’s phosphate-buffered saline (DPBS) once, then incubated with H2O2 (100 μM; Nacalai Tesque, Kyoto, Japan) dissolved in the DMEM without FBS for 1 h. The H2O2 solution was then replaced with the normal culture medium. The control cells were prepared in the same way, but without H2O2.
In the analysis of the cell population doubling rate and the cell viability, the cells were washed with DPBS, harvested from the flasks via trypsinization (0.05% trypsin, 0.53 mM EDTA for 2 min), and diluted (1:1) in 0.4% trypan blue solution. The population doubling rate and the cell viability for HEI-OC1 cells were measured using an image-based cell counter (Countess II; Thermo Fisher Scientific Inc., USA), following the manufacturer’s suggested procedure.
The metabolic activity of the HEI-OC1 cells after PQQ pretreatment was evaluated using the WST-8 (2-[2-methoxy-4-nitrophenyl]-3-[4-nitrophenyl]-5-[2,4-isulfophenyl]-2H-tetrazolium, monosodium salt) formazan assay. Cells were plated in a 96-well plate and cultured under multiple concentrations of PQQ in the medium for 1 day. Then the medium with PQQ was replaced with the normal culture medium and assays were performed using the Cell Counting Kit-8 (WST-8) (Dojindo, Kumamoto, Japan) by incubating cells with WST-8 for 1 hr as directed by the manufacturer and measuring the absorbance at 450 nm using the infinite M200 PRO-plate-reader (TECAN, USA). The mitochondrial membrane potential (MMP) is essential for maintaining mitochondrial biogenesis, and its disruption leads to cell apoptosis. To verify PQQ function with respect to the MMP, we utilized 5, 5’, 6, 6’-tetrachloro-1, 3,3’-tetraethyl benzimidazolyl carbocyanine iodide (JC-1) mitochondrial membrane fluorescent dye in analyzing the disruption of the MMP. JC-1 is taken up by the mitochondria and forms either JC-1 dimer aggregates with emission of red fluorescence (590 ± 17.5 nm) under high-membrane potentials, or JC-1 monomers with emission of green fluorescence (530 ± 15 nm) for low membrane potentials. The HEI-OC1 cells were exposed to multiple concentrations of PQQ which was then replaced with the normal medium with JC-1 (200 nM, Biotium, USA) for 30 min before the MMP ratio assay. The fluorescence emission ratio was analyzed with the infinite M200 PRO-plate-reader (TECAN, USA), under 10% CO2 incubation at 33 °C. The analysis of the red to the green ratio of JC-1 emissions was performed and calculated at multiple reads per well (circle, 4 × 4).
Measurement of cellular senescence biomarkers
The cellular senescence biomarkers of the HEI-OC1 cells were evaluated with SA-β-Gal assay using SPiDER-βGal (SG02, DOJINDO, Japan) and CD26 (dipeptidyl peptidase-4 (DPP4)) (PE anti-mouse CD26, #137804, BioLegend, USA). In the analysis with SPiDER-βGal, the cells were washed with DPBS, harvested from the flasks via trypsinization (0.05% trypsin, 0.53 mM EDTA for 2 min), fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) (pH 7.4) (10010023; Thermo Fisher Scientific Inc., USA) for 3 min, washed with Hanks’ balanced salt solution (HBSS) with Ca and Mg without Phenol Red (09735-75; Nacalai Tesque, Kyoto, Japan) three times, incubated in 0.5 μM SPiDER-βGal in McIlvaine buffer (pH 6.0) diluted five times with distilled water at 37 °C in room air for 30 min, washed with HBSS two times and diluted in HBSS for flow cytometry analysis. In the analysis of CD26, the cells were washed with DPBS, harvested from the flasks via trypsinization (0.05% trypsin, 0.53 mM EDTA for 2 min), washed with PBS two times, stained with 1 μg/100 μL CD26 antibody in PBS with 0.1% bovine serum albumin (BSA) and 0.1% NaN3, washed with PBS three times and diluted in HBSS for flow cytometry analysis.
Flow cytometry analysis
In the staining of SIRT1, the cells were washed with DPBS, harvested from the flasks via trypsinization (0.05% trypsin, 0.53 mM EDTA for 2 min), fixed with 2% paraformaldehyde in phosphate-buffered saline (PBS) (pH 7.4) (10010023; Thermo Fisher Scientific Inc., USA) at room temperature for 10 min, washed with PBS, incubated in 0.1% Triton X-100 in PBS at room temperature for 5 min, washed with PBS two times, stained with Alexa Fluor 488 Phalloidin (12:1000, A12379; Thermo Fisher Scientific Inc., USA) and anti-SIRT1 antibody (1:500, NBP1-51641AF647 (1.77 mg/mL), Novus Biologicals, Inc.) in PBS with 0.5% BSA at room temperature for 30 min, washed with PBS two times and diluted in PBS for flow cytometry analysis.
In the staining of MitoSOX Red, the cells were washed with DPBS, harvested from the flasks via trypsinization (0.05% trypsin, 0.53 mM EDTA for 2 min), washed with HBSS 2 times, stained with MitoSOX Red (5 µM, Thermo Fisher Scientific Inc., USA) in HBSS at 37 °C in room air for 10 min, washed with HBSS two times and diluted in HBSS for flow cytometry analysis.
The distribution of fluorescent intensity in HEI-OC1 cells was analyzed using a multi-color flow cytometer (Gallios, Beckman Coulter, Brea, CA, USA). The staining of both SPiDER-βGal, CD26 and MitoSOX Red was filtered with a 40 μm cell strainer and were measured using FL2 detector (488 nm excitation, 575/20 nm emission). The staining of SIRT1 was measured using FL6 detector (633 nm excitation, 660/20 nm emission). The analysis was performed using Flowing Software (Turku Bioscience Centre, Finland), flowCore99, and Bioconductor100.
Measurement of mitochondrial function using the XF24 extracellular flux analyzer
For the further protective analysis, four groups were determined: (1) control group, no PQQ treatment; (2) H2O2 group, treated with 100 μM H2O2 for 1 h; (3) 0.1 nM PQQ group, pretreatment with 0.1 nM PQQ for 1 day followed by 100 μM H2O2 for 1 h; 4) 1.0 nM PQQ group, pretreatment with 1.0 nM PQQ for 1 day followed by 100 μM H2O2 for 1 h. Cells were cultured in XF24 plate, pretreated with PQQ for 1 day followed by 100 μM H2O2 for 1 h. A Seahorse XF24 Analyzer (Agilent Technologies, Santa Clara, CA) was used to measure mitochondrial function in HEI-OC1 cells. The cells in each of the four groups as listed above were rinsed twice with assay medium (XF assay medium (DMEM without NaHCO3, without GlutaMAX, without l-glutamine; 102353-100, Seahorse Bioscience, Billerica, MA, USA, Supplementary Data), supplemented with 25 mM (450 mg/dL) d-glucose (Otsuka Seiyaku, Tokushima, Japan) and 1 mM sodium pyruvate (Nacalai Tesque, Kyoto, Japan)) and then equilibrated for 1 hr at 33 °C in an incubator (0.03% CO2) prior to the assay. The mitochondrial basal respiration was measured before any pharmacological perturbations. The cells were then subjected to (in sequence, in final concentrations): (1) 2.5 μM Oligomycin A to inhibit ATP synthase and oxidative phosphorylation (OXPHOS); (2) 1.25 μM carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP) to induce maximal respiration; (3) 0.625 μM Rotenone; (4) 0.625 μM Antimycin A to end this reaction under assay medium. The XF24 analyzer determines the oxygen and proton concentrations in real time which reflects the rate of oxygen consumption and proton production speeds. Parameters expressed as the oxygen consumption rate (OCR) in pMoles/min or the extracellular acidification rate (ECAR) in mpH/min were key indicators of mitochondrial respiration and glycolysis. The ATP production rates were calculated according to the manufacturer’s instructions and software101,102. In the substrate oxidation stress test, the cells were prepared as above prior to the assay, and the cells were subjected to (in sequence, in final concentrations): (1) 0, 4 mM l-glutamine (Seahorse Bioscience, Billerica, MA, USA), 40 μM Etomoxir (11969, Cayman Chemical, Ann Arbor, USA) or 3 μM BPTES ((bis-2-(5-phenylacetamido-1,3,4-thiadiazol-2-yl)ethyl sulfide); 19284, Cayman Chemical, Ann Arbor, USA) (2) 5 μM Oligomycin A; (3) 2.5 μM FCCP. In the fatty acid oxidation (FAO) assay, the culture medium was washed twice and replaced with substrate-limited DMEM (102353-100, Seahorse Bioscience, Billerica, MA, USA) supplemented with 25 mM d-glucose, 1 mM sodium pyruvate, 0.5 mM l-carnitine hydrochloride (07353-51, Nacalai Tesque, Kyoto, Japan) and 0.5 mM NaOH to adjust to pH 7.4, and the cells were equilibrated for 1 h at 33 °C in an incubator (0.03% CO2) prior to the assay. In the FAO assay, the cells were subjected to (in sequence, in final concentrations): (1) conjugate of palmitatic acid (PA) (25919-62, Nacalai Tesque, Kyoto, Japan) and fatty acid-free bovine serum albumin (BSA) (08587-26, Nacalai Tesque, Kyoto, Japan) (final concentration PA: 200 µM, BSA 34 µM) or BSA (34 µM) only (2) 5 μM Oligomycin A; (3) 2.5 μM FCCP. The number of cells was counted after each XF24 analysis as follows; the cells in the XF24 plate was washed with PBS 1 time, fixed with 4% paraformaldehyde (PFA) (Nacalai Tesque, Kyoto, Japan) for 1 min, stained with Hoechst 33342 (final 5 µg/mL, Thermo Fisher Scientific Inc., USA) for 30 min, washed with PBS one time, captured with a fluorescence microscope (BZ-X710, KEYENCE, Osaka, Japan) using the DAPI filter cube, and counted automatically using ImageJ103,104 with Fiji105.
Analysis of mitochondrial dynamics and morphology
Cells were dyed with tetramethylrhodamine, ethyl ester (TMRE) (200 nM, Biotium, USA) mitochondrial fluorescence and captured with confocal microscopy focusing on mitochondrial-rich regions beside the cell nucleus. Z stacks across the depth of the cell provide 3D information about mitochondrial morphology while the time-lapse images allow the study of changes in mitochondrial dynamics over time. Images were captured at 3 s of intervals for a duration of 3 min. Supplementary Video 1 depicts a representation of a projection of Z stacks of mitochondria in a single cell with a focus on the mitochondrial-rich regions. All image processing and analysis were performed using NIS-Elements (Nikon, Japan), ImageJ103,104 with Fiji105, TrackMate106, and @msdanalyzer107 in MATLAB R2019b. The mean square displacement (MSD) and the area under curve (AUC) measurements were used for the analysis of the parameters of mitochondrial motion. The mitochondrial skeleton was vectorized to measure branches, junctions, and average branch lengths as described in our previous study5.
Western blot
The cells were washed three times with ice-cold PBS and then lysed in RIPA lysis buffer containing protease inhibitor cocktails (in final concentrations; 10 mM NaF, 1 mM Sodium orthovanadate, 1 mM PMSF, 1× cOmplete Mini (Roche, Mannheim, Germany), 0.3 µg/mL Trichostatin A, 10 mM Nicotinamide, 5 mM Beta-glycerophosphoric acid, 1 mM dithiothreitol (DTT)) for 30 min on ice. Insoluble fractions of cell lysates (2–3 mg) were removed by centrifugation at 13,000 ×g at 4 °C for 5 min and denatured by boiling in EzApply (AE-1430; ATTO Co, Japan). The protein concentrations were determined using a Pierce BCA Protein Assay Kit (Thermo Fisher Scientific, Inc., Waltham, MA, USA). The samples containing 60 μg of proteins were separated by SDS-PAGE using EzRunC + (AE-1412; ATTO Co, Japan) and transferred onto PVDF membranes using the BioRad TransBlot Turbo according to the manufacturer’s instructions (High MW 10 min for SIRT1, Mixed MW 7 min for PGC-1α and Ac-Lysine). Following blocking with 5% nonfat milk (Amersham ECL blocking agent, GE Healthcare) at room temperature for 1 hr, membranes were incubated with primary antibodies at 4 °C overnight and then incubated with horseradish peroxidase (HRP) conjugated secondary antibodies (anti-mouse 1:200,000, ab6789, Abcam; anti-rabbit 1:200,000, NA934VS, Sigma Aldrich) for 1 h at room temperature. The detection was carried out using the chemiluminescent imaging system (ImageQuant LAS 4010, UK) after incubation with SuperSignalTM West Femto Maximum Sensitivity Substrate (Thermo, USA) according to the manufacturer’s instructions. The primary antibodies used were SIRT1 (1:500, (B-7) sc-74465, Santa Cruz), PGC-1α (1:200, (D-5) sc-518025, Santa Cruz), and β-actin (1:10,000, PM053, MBL).
All blots derive from the same experiment and were processed in parallel.
Immunoprecipitation and PGC-1α acetylation
Immunoprecipitation was performed according to the manufacturer’s directions (Thermo Fisher Scientific, Inc., Waltham, MA, USA): 50 μL Dynabeads Protein G was added to the 500 μg of sample solution and shaken on a horizontal shaker for 10 min to eliminate nonspecific binding proteins before separating on the magnet until the supernatant was clear and could be removed. 4 μg primary antibody against PGC-1α and 50 μL of protein G agarose were then added into the supernatant and shaken on a horizontal shaker for 10 min at room temperature. After washing three times, the beads were eluted with 50 mM Glycine (pH 2.8) for 2 min and then separated with SAS-PAGE as described in the western blot protocol above. The membranes were incubated with acetyl-lysine HRP (1:200, (7F8) sc-81623 HRP, Santa Cruz) to stain at 4 °C overnight after blocking the transferred membrane. The membranes were then detected with the same procedure described above before being reproved with anti-PGC-1α HRP (1:200, (D-5) sc-518025 HRP, Santa Cruz) antibody and a secondary antibody (anti-mouse 1:1000 Mouse TrueBlot ULTRA, 18-8817-30, Rockland).
Quantitative RT-PCR analysis
Total RNA was isolated with NucleoSpin RNA (Macherey-Nagel, Düren, Germany) according to the manufacturer’s instructions. The concentration of extracted mRNA was measured using NanoDrop Lite (Thermo Fisher Scientific, USA) and complementary DNA (cDNA) was generated using ReverTraAce qPCR RT Master Mix with gDNA Remover (TOYOBO, Japan) according to the manufacturer’s instructions. The quality of mRNA was assessed by visualization of the 28S and 18S ribosomal RNA bands using DynaMarker, RNA High for Easy Electrophoresis (BioDynamics Laboratory Inc., Japan). The primers of genes including SIRT1 (forward GCTGACGACTTCGACGACG, reverse TCGGTCAACAGGAGGTTGTCT, final concentration 250 nM), PGC-1α (forward AAGTGGTGTAGCGACCAATCG, reverse AATGAGGGCAATCCGTCTTCA, final concentration 50 nM), Cdkn2a (p16) (forward CGCAGGTTCTTGGTCACTGT, reverse TGTTCACGAAAGCCAGAGCG, final concentration 500 nM), Cdkn1a (p21) (forward CCTGGTGATGTCCGACCTG, reverse CCATGAGCGCATCGCAATC, final concentration 500 nM), Trp53 (p53) (forward GCGTAAACGCTTCGAGATGTT, reverse TTTTTATGGCGGGAAGTAGACTG, final concentration 500 nM), Cpt1a (forward GTCCCTCCAGCTGGCTTATC, reverse CATGCGGCCAGTGGTGTCTA, final concentration 1000 nM), Cpt1c (forward TCTTCACTGAGTTCCGATGGG, reverse ACGCCAGAGATGCCTTTTCC, final concentration 125 nM), Cpt2 (forward CCCTGCATACCAGCGGATAA, reverse CATACGCAATGCCAAAGCCA, final concentration 500 nM), Gls1 (forward TTCGCCCTCGGAGATCCTAC, reverse CCAAGCTAGGTAACAGACCCT, final concentration 500 nM), Gls2 (forward CGTCCGGTACTACCTCGGT, reverse TGTCCCTCTGCAATAGTGTAGAA, final concentration 250 nM), GAPDH (forward TGAGGCCGGTGCTGAGTATGTCG, reverse CCACAGTCTTCTGGGTGGCAGTG, final concentration 125 nM) and β-actin (forward CCTCTATGCCAACACAGTGC, reverse GTACTCCTGCTTGCTGATCC, final concentration 250 nM) were purchased from FASMAC (Kanagawa, Japan). The forward and reverse primers were mixed with 100 ng cDNA and quantitative RT-PCR was performed using PowerUp SYBR Green Master Mix (Applied Biosystems, Darmstadt, Germany) or THUNDERBIRD SYBR qPCR Mix (TOYOBO, Japan). The following experimental run protocol was used: denaturation and activation program (50 °C for 120 s, 95 °C for 120 s), amplification and quantification program repeated 60 times (95 °C for 15 s, 60 °C for 60 s), melting curve program (95 °C for 15 s, 60 °C for 60 s, 95 °C for 15 s). Data collection was performed using QuantStudio 7 Flex Detection System (Thermo Fisher Scientific, USA). The 2-ΔΔCt method was applied to analyze the relative changes in gene expressions. The mRNA expression level of each gene was normalized using β-actin as an internal control.
Mitochondrial DNA copy number analysis
Total DNA was isolated with NucleoSpin Tissue (Macherey-Nagel, Düren, Germany) according to the manufacturer’s instructions. The concentration of extracted DNA was measured using NanoDrop Lite (Thermo Fisher Scientific, USA). The primers of mitochondrial and nuclear genes including mitochondrial 16 S rRNA (forward CCGCAAGGGAAAGATGAAAGAC, reverse TCGTTTGGTTTCGGGGTTTC, final concentration 1000 nM), mitochondrial NADH-ubiquinone oxidoreductase chain 1 (ND1) (forward CTAGCAGAAACAAACCGGGC, reverse CCGGCTGCGTATTCTACGTT, final concentration 500 nM), mitochondrial D-loop (forward CCAAAAAACACTAAGAACTTGAAAGACA, reverse GTCATATTTTGGGAACTACTAGAATTGATC, final concentration 1000 nM), nuclear hexokinase 2 (HK2) (forward GCCAGCCTCTCCTGATTTTAGTGT, reverse GGGAACACAAAAGACCTCTTCTGG, final concentration 500 nM) and nuclear thymidine kinase 1 (TK1) (forward GACTGTATTGAGCGGCTTCAGA, reverse CATGCTCGGTGTGAGCCATA, final concentration 500 nM) were purchased from FASMAC (Kanagawa, Japan). Quantitative PCR was performed in the same way as in quantitative RT-PCR analysis. The copy number of mitochondrial DNA was calculated using the deviations of the cycle threshold (Ct) between mitochondrial and nuclear genes according to the previous studies108,109.
Measurement of glucose concentration of the medium
The glucose concentration of the medium was measured using the following glucometers: Accu-Chek Aviva Nano/Guide (Roche Diagnostics GmbH, Mannheim, Germany), OneTouch Verio Reflect (Johnson & Johnson, New Jersey, USA), Freestyle Libre (Abbott Diabetes Care, Witney, U.K.) and NIPRO StatStrip XP2 (NIPRO, Osaka, Japan).
Statistical analysis
The statistical software GraphPad Prism and R version 4.1.0 software (R Core Team; R Foundation for Statistical Computing, Vienna, Austria, 2021) were used in data processing and statistical analysis. A one-way ANOVA with Bonferroni’s test was used to make multiple comparisons between groups. A value of P < 0.05 was considered significant. Box plot shows statistical parameters as follows; central line: median; box limits: first and third quartile; whiskers: minimum and maximum.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Supplementary information
Acknowledgements
This work was supported by ROHTO Pharmaceutical Co., Ltd. (Osaka, Japan). JSPS KAKENHI Grant Numbers 25293347, 26253081, 18K16906, 18K19602, 20H00546, 20K21646 and 21K16853. We thank Kenichi Nishimaki, Hisae Aoyagi, and Miwako Shidomi for suggesting the topic of this paper and the early stages of this work. We thank Haruko Tanaka, Atsuko Tsuyuzaki, Emi Usukura, Satoru Fukuda, and Ayano Mitsui for technical assistance. We thank Laura Mezey for reviewing this English language in this manuscript.
Author contributions
T.Y. and T.K. conceived this study. T.Y., C.F., S.I., and T.K. contributed to supervising experimental design. Y.G. and T.K. conducted the experiment. Y.G. and T.K. performed collection of the data and statistical analysis. T.Y. contributed to the final approval of the manuscript. All authors contributed to the writing of the manuscript.
Data availability
Data are available from the corresponding author on a reasonable request.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Ying Gao, Teru Kamogashira.
Change history
10/6/2022
A Correction to this paper has been published: 10.1038/s41514-022-00095-w
Contributor Information
Teru Kamogashira, Email: tkamogashira-tky@umin.ac.jp.
Tatsuya Yamasoba, Email: tyamasoba-tky@umin.ac.jp.
Supplementary information
The online version contains supplementary material available at 10.1038/s41514-022-00083-0.
References
- 1.Baker DJ, et al. Clearance of p16 Ink4a-positive senescent cells delays ageing-associated disorders. Nature. 2011;479:232–236. doi: 10.1038/nature10600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Tsuchihashi NA, et al. Autophagy through 4EBP1 and AMPK regulates oxidative stress-induced premature senescence in auditory cells. Oncotarget. 2015;6:3644–3655. doi: 10.18632/oncotarget.2874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wang Z, Wei D, Xiao H. Methods of cellular senescence induction using oxidative stress. Methods Mol. Biol. 2013;1048:135–144. doi: 10.1007/978-1-62703-556-9_11. [DOI] [PubMed] [Google Scholar]
- 4.Zdanov S, Remacle J, Toussaint O. Establishment of H2O2-induced premature senescence in human fibroblasts concomitant with increased cellular production of H2O2. Annal. New York Acad. Sci. 2006;1067:210–216. doi: 10.1196/annals.1354.025. [DOI] [PubMed] [Google Scholar]
- 5.Kamogashira T, Hayashi K, Fujimoto C, Iwasaki S, Yamasoba T. Functionally and morphologically damaged mitochondria observed in auditory cells under senescence-inducing stress. npj Aging Mech. Dis. 2017;3:2. doi: 10.1038/s41514-017-0002-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Schieber M, Chandel NS. ROS function in redox signaling and oxidative stress. Current Biology. 2014;24:R453–R462. doi: 10.1016/j.cub.2014.03.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hauge JG. Glucose dehydrogenase of bacterium anitratum: an enzyme with a novel. J. Biol. Chem. 1964;239:3630–3639. [PubMed] [Google Scholar]
- 8.Salisbury SA, Forrest HS, Cruse WBT, Kennard O. A novel coenzyme from bacterial primary alcohol dehydrogenases. Nature. 1979;280:843–844. doi: 10.1038/280843a0. [DOI] [PubMed] [Google Scholar]
- 9.Westerling J, Frank J, Duine JA. The prosthetic group of methanol dehydrogenase from hyphomicrobium X: electron spin resonance evidence for a quinone structure. Biochem. Biophys. Res. Commun. 1979;87:719–724. doi: 10.1016/0006-291x(79)92018-7. [DOI] [PubMed] [Google Scholar]
- 10.Kumazawa T, Sato S, Seno H, Ishii A, Suzuki O. Levels of pyrroloquinoline quinone in various foods. Biochem. J. 1995;307:331–333. doi: 10.1042/bj3070331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mitchell AE, Jones AD, Mercer RS, Rucker RB. Characterization of pyrroloquinoline quinone amino acid derivatives by electrospray ionization mass spectrometry and detection in human milk. Anal. Biochem. 1999;269:317–325. doi: 10.1006/abio.1999.4039. [DOI] [PubMed] [Google Scholar]
- 12.Stites TE, Mitchell AE, Rucker RB. Physiological importance of quinoenzymes and the O-quinone family of cofactors. J. Nutr. 2000;130:719–727. doi: 10.1093/jn/130.4.719. [DOI] [PubMed] [Google Scholar]
- 13.Akagawa M, Nakano M, Ikemoto K. Recent progress in studies on the health benefits of pyrroloquinoline quinone. Biosci. Biotechnol. Biochem. 2016;80:13–22. doi: 10.1080/09168451.2015.1062715. [DOI] [PubMed] [Google Scholar]
- 14.Rucker R, Chowanadisai W, Nakano M. Potential physiological importance of pyrroloquinoline quinone. Alternative Med. Rev. 2009;14:268–277. [PubMed] [Google Scholar]
- 15.Zhang Q, Shen M, Ding M, Shen D, Ding F. The neuroprotective action of pyrroloquinoline quinone against glutamate-induced apoptosis in hippocampal neurons is mediated through the activation of PI3K/Akt pathway. Toxicol. Appl. Pharmacol. 2011;252:62–72. doi: 10.1016/j.taap.2011.02.006. [DOI] [PubMed] [Google Scholar]
- 16.Misra HS, et al. Pyrroloquinoline-quinone: a reactive oxygen species scavenger in bacteria. FEBS Lett. 2004;578:26–30. doi: 10.1016/j.febslet.2004.10.061. [DOI] [PubMed] [Google Scholar]
- 17.Xu T, et al. Pyrroloquinoline quinone attenuates cachexia-induced muscle atrophy via suppression of reactive oxygen species. J. Thorac. Dis. 2018;10:2752–2759. doi: 10.21037/jtd.2018.04.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zhu BQ, Zhou HZ, Teerlink JR, Karliner JS. Pyrroloquinoline quinone (PQQ) decreases myocardial infarct size and improves cardiac function in rat models of ischemia and ischemia/reperfusion. Cardiovasc. Drugs Ther. 2004;18:421–431. doi: 10.1007/s10557-004-6219-x. [DOI] [PubMed] [Google Scholar]
- 19.Xu X, et al. Pyrroloquinoline quinone can prevent chronic heart failure by regulating mitochondrial function. Cardiovasc. Diagn. Ther. 2020;10:453–469. doi: 10.21037/cdt-20-129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jiang X, et al. Hepatoprotective effect of pyrroloquinoline quinone against alcoholic liver injury through activating Nrf2-mediated antioxidant and inhibiting TLR4-mediated inflammation responses. Process Biochem. 2020;92:303–312. [Google Scholar]
- 21.Ohwada K, et al. Pyrroloquinoline quinone (PQQ) prevents cognitive deficit caused by oxidative stress in rats. J. Clin. Biochem. Nutr. 2008;42:29–34. doi: 10.3164/jcbn.2008005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Matsushita K, Toyama H, Yamada M, Adachi O. Quinoproteins: structure, function, and biotechnological applications. Appl. Microbiol. Biotechnol. 2002;58:13–22. doi: 10.1007/s00253-001-0851-1. [DOI] [PubMed] [Google Scholar]
- 23.Tao R, et al. Pyrroloquinoline quinone preserves mitochondrial function and prevents oxidative injury in adult rat cardiac myocytes. Biochem. Biophys. Res. Commun. 2007;363:257–262. doi: 10.1016/j.bbrc.2007.08.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hoque SAM, Umehara T, Kawai T, Shimada M. Adverse effect of superoxide-induced mitochondrial damage in granulosa cells on follicular development in mouse ovaries. Free Radic. Biol. Med. 2021;163:344–355. doi: 10.1016/j.freeradbiomed.2020.12.434. [DOI] [PubMed] [Google Scholar]
- 25.Zhang Y, Feustel PJ, Kimelberg HK. Neuroprotection by pyrroloquinoline quinone (PQQ) in reversible middle cerebral artery occlusion in the adult rat. Brain Res. 2006;1094:200–206. doi: 10.1016/j.brainres.2006.03.111. [DOI] [PubMed] [Google Scholar]
- 26.Zhang L, et al. The neuroprotective effect of pyrroloquinoline quinone on traumatic brain injury. J. Neurotrauma. 2012;29:851–864. doi: 10.1089/neu.2011.1882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chowanadisai W, et al. Pyrroloquinoline quinone stimulates mitochondrial biogenesis through cAMP response element-binding protein phosphorylation and increased PGC-1α expression. J. Biol. Chem. 2010;285:142–152. doi: 10.1074/jbc.M109.030130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bauerly K, et al. Altering pyrroloquinoline quinone nutritional status modulates mitochondrial, lipid, and energy metabolism in rats. PLoS ONE. 2011;6:e21779. doi: 10.1371/journal.pone.0021779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Stites T, et al. Pyrroloquinoline quinone modulates mitochondrial quantity and function in mice. J. Nutr. 2006;136:390–396. doi: 10.1093/jn/136.2.390. [DOI] [PubMed] [Google Scholar]
- 30.Lu J, et al. Mitochondrial regulation by pyrroloquinoline quinone prevents rotenone-induced neurotoxicity in Parkinson’s disease models. Neurosci. Lett. 2018;687:104–110. doi: 10.1016/j.neulet.2018.09.031. [DOI] [PubMed] [Google Scholar]
- 31.Harris CB, et al. Dietary pyrroloquinoline quinone (PQQ) alters indicators of inflammation and mitochondrial-related metabolism in human subjects. J. Nutr. Biochem. 2013;24:2076–2084. doi: 10.1016/j.jnutbio.2013.07.008. [DOI] [PubMed] [Google Scholar]
- 32.Philp A, et al. Sirtuin 1 (SIRT1) deacetylase activity is not required for mitochondrial biogenesis or peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α) deacetylation following endurance exercise. J. Biol. Chem. 2011;286:30561–30570. doi: 10.1074/jbc.M111.261685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Li P, et al. Activation of sirtuin 1/3 improves vascular hyporeactivity in severe hemorrhagic shock by alleviation of mitochondrial damage. Oncotarget. 2015;6:36998–37011. doi: 10.18632/oncotarget.6076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Powell RD, et al. Resveratrol attenuates hypoxic injury in a primary hepatocyte model of hemorrhagic shock and resuscitation. J. Trauma Acute Care Surg. 2014;76:409–417. doi: 10.1097/TA.0000000000000096. [DOI] [PubMed] [Google Scholar]
- 35.Hasegawa K, et al. Kidney-specific overexpression of Sirt1 protects against acute kidney injury by retaining peroxisome function. J. Biol. Chem. 2010;285:13045–13056. doi: 10.1074/jbc.M109.067728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Xu S, et al. SIRT1/3 activation by resveratrol attenuates acute kidney injury in a septic rat model. Oxid. Med. Cell. Longev. 2016;2016:7296092. doi: 10.1155/2016/7296092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Deng Z, et al. SIRT1 attenuates sepsis-induced acute kidney injury via Beclin1 deacetylation-mediated autophagy activation. Cell Death Dis. 2021;12:217. doi: 10.1038/s41419-021-03508-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Jian B, Yang S, Chaudry IH, Raju R. Resveratrol improves cardiac contractility following trauma-hemorrhage by modulating Sirt1. Mol. Med. 2012;18:209–214. doi: 10.2119/molmed.2011.00365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Jian B, Yang S, Chaudry IH, Raju R. Resveratrol restores sirtuin 1 (SIRT1) activity and pyruvate dehydrogenase kinase 1 (PDK1) expression after hemorrhagic injury in a rat model. Mol. Med. 2014;20:10–16. doi: 10.2119/molmed.2013.00077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yamada Y, et al. Effects of pyrroloquinoline quinone and imidazole pyrroloquinoline on biological activities and neural functions. Heliyon. 2020;6:e03240. doi: 10.1016/j.heliyon.2020.e03240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zhang J, et al. Pyrroloquinoline quinone increases the expression and activity of Sirt1 and -3 genes in HepG2 cells. Nutr. Res. 2015;35:844–849. doi: 10.1016/j.nutres.2015.06.014. [DOI] [PubMed] [Google Scholar]
- 42.Hwang P, Willoughby DS. Mechanisms behind pyrroloquinoline quinone supplementation on skeletal muscle mitochondrial biogenesis: possible synergistic effects with exercise. J. Am. College Nutr. 2018;37:738–748. doi: 10.1080/07315724.2018.1461146. [DOI] [PubMed] [Google Scholar]
- 43.Hwang PS, et al. Effects of pyrroloquinoline quinone (PQQ) supplementation on aerobic exercise performance and indices of mitochondrial biogenesis in untrained men. J. Am. Coll. Nutr. 2020;39:547–556. doi: 10.1080/07315724.2019.1705203. [DOI] [PubMed] [Google Scholar]
- 44.Min Z, et al. Pyrroloquinoline quinone induces cancer cell apoptosis via mitochondrial-dependent pathway and down-regulating cellular Bcl-2 protein expression. J. Cancer. 2014;5:609–624. doi: 10.7150/jca.9002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Chen J-H, Ozanne SE, Hales CN. Methods of cellular senescence induction using oxidative stress. Methods Mol. Biol. 2007;371:179–189. doi: 10.1007/978-1-59745-361-5_14. [DOI] [PubMed] [Google Scholar]
- 46.Duan JJJ, Duan JJJ, Zhang Z, Tong T. Irreversible cellular senescence induced by prolonged exposure to H 2O2 involves DNA-damage-and-repair genes and telomere shortening. Int. J. Biochem. Cell Biol. 2005;37:1407–1420. doi: 10.1016/j.biocel.2005.01.010. [DOI] [PubMed] [Google Scholar]
- 47.Furukawa A, Tada-Oikawa S, Kawanishi S, Oikawa S. H2O2 accelerates cellular senescence by accumulation of acetylated p53 via decrease in the function of SIRT1 by NAD depletion. Cell. Physiol. Biochem. 2007;20:45–54. doi: 10.1159/000104152. [DOI] [PubMed] [Google Scholar]
- 48.Zhang Y, et al. PIN1 protects hair cells and auditory HEI-OC1 cells against senescence by inhibiting the PI3K/Akt/mTOR pathway. Oxid. Med. Cell. Longev. 2021;2021:1–17. doi: 10.1155/2021/9980444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.González-Gualda E, Baker AG, Fruk L, Muñoz-Espín D. A guide to assessing cellular senescence in vitro and in vivo. FEBS J. 2021;288:56–80. doi: 10.1111/febs.15570. [DOI] [PubMed] [Google Scholar]
- 50.Kumari R, Jat P. Mechanisms of cellular senescence: cell cycle arrest and senescence associated secretory phenotype. Front. Cell Dev. Biol. 2021;9:645593. doi: 10.3389/fcell.2021.645593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lee BY, et al. Senescence-associated beta-galactosidase is lysosomal beta-galactosidase. Aging Cell. 2006;5:187–195. doi: 10.1111/j.1474-9726.2006.00199.x. [DOI] [PubMed] [Google Scholar]
- 52.Itahana K, Campisi J, Dimri GP. Methods to detect biomarkers of cellular senescence: the senescence-associated beta-galactosidase assay. Methods Mol. Biol. 2007;371:21–31. doi: 10.1007/978-1-59745-361-5_3. [DOI] [PubMed] [Google Scholar]
- 53.Herranz N, Gil J. Mechanisms and functions of cellular senescence. J. Clin. Investig. 2018;128:1238–1246. doi: 10.1172/JCI95148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Zhu RZ, Li BS, Gao SS, Seo JH, Choi B-M. Luteolin inhibits H2O2-induced cellular senescence via modulation of SIRT1 and p53. Korean J. Physiol. Pharmacol. 2021;25:297–305. doi: 10.4196/kjpp.2021.25.4.297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kim KM, et al. Identification of senescent cell surface targetable protein DPP4. Genes Dev. 2017;31:1529–1534. doi: 10.1101/gad.302570.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Johmura Y, et al. Senolysis by glutaminolysis inhibition ameliorates various age-associated disorders. Science. 2021;371:265–270. doi: 10.1126/science.abb5916. [DOI] [PubMed] [Google Scholar]
- 57.Cheng XW, et al. Dose rectification of an imbalance between DPP4 and GLP-1 ameliorates chronic stress-related vascular aging and atherosclerosis? Clin. Exp. Pharmacol. Physiol. 2018;45:467–470. doi: 10.1111/1440-1681.12903. [DOI] [PubMed] [Google Scholar]
- 58.Dranka BP, Hill BG, Darley-Usmar VM. Mitochondrial reserve capacity in endothelial cells: the impact of nitric oxide and reactive oxygen species. Free Radic. Biol. Med. 2010;48:905–914. doi: 10.1016/j.freeradbiomed.2010.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Sansbury BE, Jones SP, Riggs DW, Darley-Usmar VM, Hill BG. Bioenergetic function in cardiovascular cells: the importance of the reserve capacity and its biological regulation. in. Chemico-Biological Interactions. 2011;191:288–295. doi: 10.1016/j.cbi.2010.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Chen Z, Shentu T-P, Wen L, Johnson DA, Shyy JY-J. Regulation of SIRT1 by oxidative stress-responsive miRNAs and a systematic approach to identify its role in the endothelium. Antioxid. Redox Signal. 2013;19:1522–1538. doi: 10.1089/ars.2012.4803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Tsai K-L, et al. The role of microRNAs in modulating sirtuin 1 expression. J. Clin. Gerontol. Geriatr. 2011;2:71–75. [Google Scholar]
- 62.Schwanhäusser B, et al. Global quantification of mammalian gene expression control. Nature. 2011;473:337–342. doi: 10.1038/nature10098. [DOI] [PubMed] [Google Scholar]
- 63.Baker JR, et al. Oxidative stress dependent microRNA-34a activation via PI3Kα reduces the expression of sirtuin-1 and sirtuin-6 in epithelial cells. Sci. Rep. 2016;6:35871. doi: 10.1038/srep35871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Braun RJ, Westermann B. Mitochondrial dynamics in yeast cell death and aging. in. Biochemical Soc. Transact. 2011;39:1520–1526. doi: 10.1042/BST0391520. [DOI] [PubMed] [Google Scholar]
- 65.Jiang C, et al. Pyrroloquinoline quinine ameliorates doxorubicin-induced autophagy-dependent apoptosis via lysosomal-mitochondrial axis in vascular endothelial cells. Toxicology. 2019;425:152238. doi: 10.1016/j.tox.2019.152238. [DOI] [PubMed] [Google Scholar]
- 66.Chalmers S, Saunter CD, Girkin JM, McCarron JG. Age decreases mitochondrial motility and increases mitochondrial size in vascular smooth muscle. J. Physiol. 2016;594:4283–4295. doi: 10.1113/JP271942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Woods LC, Berbusse GW, Naylor K. Microtubules are essential for mitochondrial dynamics-fission, fusion, and motility-in Dictyostelium discoideum. Front. Cell Dev. Biol. 2016;4:19. doi: 10.3389/fcell.2016.00019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Al-Mehdi AB, et al. Perinuclear mitochondrial clustering creates an oxidant-rich nuclear domain required for hypoxia-induced transcription. Sci. Signal. 2012;5:ra47. doi: 10.1126/scisignal.2002712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Mattenberger Y, James DI, Martinou JC. Fusion of mitochondria in mammalian cells is dependent on the mitochondrial inner membrane potential and independent of microtubules or actin. FEBS Lett. 2003;538:53–59. doi: 10.1016/s0014-5793(03)00124-8. [DOI] [PubMed] [Google Scholar]
- 70.Schwarz TL. Mitochondrial trafficking in neurons. Cold Spring Harb. Perspect. Biol. 2013;5:a011304. doi: 10.1101/cshperspect.a011304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Barnhart EL. Mechanics of mitochondrial motility in neurons. Curr. Opin. Cell Biol. 2016;38:90–99. doi: 10.1016/j.ceb.2016.02.022. [DOI] [PubMed] [Google Scholar]
- 72.Ohno N, et al. Mitochondrial immobilization mediated by syntaphilin facilitates survival of demyelinated axons. Proc. Natl Acad. Sci. USA. 2014;111:9953–9958. doi: 10.1073/pnas.1401155111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Seok J, et al. Alteration of fatty acid oxidation by increased CPT1A on replicative senescence of placenta-derived mesenchymal stem cells. Stem Cell Res. Ther. 2020;11:1. doi: 10.1186/s13287-019-1471-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Schulman JJ, et al. Bok regulates mitochondrial fusion and morphology. Cell Death Differ. 2019;26:2682–2694. doi: 10.1038/s41418-019-0327-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Xu C, et al. SIRT1 is downregulated by autophagy in senescence and ageing. Nat. Cell Biol. 2020;22:1170–1179. doi: 10.1038/s41556-020-00579-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Saihara K, Kamikubo R, Ikemoto K, Uchida K, Akagawa M. Pyrroloquinoline quinone, a redox-active o-quinone, stimulates mitochondrial biogenesis by activating the SIRT1/PGC-1α signaling pathway. Biochemistry. 2017;56:6615–6625. doi: 10.1021/acs.biochem.7b01185. [DOI] [PubMed] [Google Scholar]
- 77.Kuo YT, Shih PH, Kao SH, Yeh GC, Lee HM. Pyrroloquinoline quinone resists denervation-induced skeletal muscle atrophy by activating PGC-1α and integrating mitochondrial electron transport chain complexes. PLoS ONE. 2015;10:e0143600. doi: 10.1371/journal.pone.0143600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Xue T, et al. MiR-29b overexpression induces cochlear hair cell apoptosis through the regulation of SIRT1/PGC-1 signaling: implications for age-related hearing loss. Int. J. Mol. Med. 2016;38:1387–1394. doi: 10.3892/ijmm.2016.2735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Zhang W, et al. PGC-1α overexpression promotes mitochondrial biogenesis to protect auditory cells against cisplatin-induced cytotoxicity. J. Bio-X Res. 2019;2:81–86. [Google Scholar]
- 80.Rodgers JT, et al. Nutrient control of glucose homeostasis through a complex of PGC-1α and SIRT1. Nature. 2005;434:113–118. doi: 10.1038/nature03354. [DOI] [PubMed] [Google Scholar]
- 81.Brennan JP, Berry RG, Baghai M, Duchen MR, Shattock MJ. FCCP is cardioprotective at concentrations that cause mitochondrial oxidation without detectable depolarisation. Cardiovasc. Res. 2006;72:322–330. doi: 10.1016/j.cardiores.2006.08.006. [DOI] [PubMed] [Google Scholar]
- 82.Brennan JP, et al. Mitochondrial uncoupling, with low concentration FCCP, induces ROS-dependent cardioprotection independent of KATP channel activation. Cardiovasc. Res. 2006;72:313–321. doi: 10.1016/j.cardiores.2006.07.019. [DOI] [PubMed] [Google Scholar]
- 83.Demine S, Renard P, Arnould T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 2019;8:795. doi: 10.3390/cells8080795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Cadenas S. Mitochondrial uncoupling, ROS generation and cardioprotection. Biochimica et Biophysica Acta - Bioenergetics. 2018;1859:940–950. doi: 10.1016/j.bbabio.2018.05.019. [DOI] [PubMed] [Google Scholar]
- 85.Pan X, et al. Mitochondrial uncouplers confer protection by activating amp-activated protein kinase to inhibit neuroinflammation following intracerebral hemorrhage. Biol. Pharm. Bull. 2020;43:1210–1219. doi: 10.1248/bpb.b20-00108. [DOI] [PubMed] [Google Scholar]
- 86.Suzuki T, et al. Weight loss by Ppc-1, a novel small molecule mitochondrial uncoupler derived from slime mold. PLoS ONE. 2015;10:e0117088. doi: 10.1371/journal.pone.0117088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Kikuchi H, et al. Synthesis of prenylated quinolinecarboxylic acid derivatives and their anti-obesity activities. Bioorganic Med. Chem. 2015;23:66–72. doi: 10.1016/j.bmc.2014.11.024. [DOI] [PubMed] [Google Scholar]
- 88.Manna P, Jain SK. Obesity, oxidative stress, adipose tissue dysfunction, and the associated health risks: causes and therapeutic strategies. Metabolic Syndrome Related Disord. 2015;13:423–444. doi: 10.1089/met.2015.0095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Sasakura H, et al. Lifespan extension by peroxidase and dual oxidase-mediated ROS signaling through pyrroloquinoline quinone in C. elegans. J. Cell Sci. 2017;130:2631–2643. doi: 10.1242/jcs.202119. [DOI] [PubMed] [Google Scholar]
- 90.Cho I, Song H-O, Cho JH. Flavonoids mitigate neurodegeneration in aged Caenorhabditis elegans by mitochondrial uncoupling. Food Sci. Nutr. 2020;8:6633–6642. doi: 10.1002/fsn3.1956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.de Oliveira MR, et al. Quercetin and the mitochondria: a mechanistic view. Biotechnol. Adv. 2016;34:532–549. doi: 10.1016/j.biotechadv.2015.12.014. [DOI] [PubMed] [Google Scholar]
- 92.Zholobenko AV, Mouithys-Mickalad A, Dostal Z, Serteyn D, Modriansky M. On the causes and consequences of the uncoupler-like effects of quercetin and dehydrosilybin in H9c2 cells. PLoS ONE. 2017;12:e0185691. doi: 10.1371/journal.pone.0185691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Ristow M, Schmeisser S. Extending life span by increasing oxidative stress. Free Radical Biology and Medicine. 2011;51:327–336. doi: 10.1016/j.freeradbiomed.2011.05.010. [DOI] [PubMed] [Google Scholar]
- 94.Ly JD, Grubb DR, Lawen A. The mitochondrial membrane potential (δψm) in apoptosis; an update. Apoptosis. 2003;8:115–128. doi: 10.1023/a:1022945107762. [DOI] [PubMed] [Google Scholar]
- 95.Wilfling F, et al. BH3-only proteins are tail-anchored in the outer mitochondrial membrane and can initiate the activation of Bax. Cell Death Differ. 2012;19:1328–1336. doi: 10.1038/cdd.2012.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Kura B, Szeiffova Bacova B, Kalocayova B, Sykora M, Slezak J. Oxidative stress-responsive microRNAs in heart injury. Int. J. Mol. Sci. 2020;21:358. doi: 10.3390/ijms21010358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Kalinec GM, Webster P, Lim DJ, Kalinec F. A cochlear cell line as an in vitro system for drug ototoxicity screening. Audiol. Neuro-Otol. 2003;8:177–189. doi: 10.1159/000071059. [DOI] [PubMed] [Google Scholar]
- 98.Kalinec G, Thein P, Park C, Kalinec F. HEI-OC1 cells as a model for investigating drug cytotoxicity. Hear. Res. 2016;335:105–117. doi: 10.1016/j.heares.2016.02.019. [DOI] [PubMed] [Google Scholar]
- 99.Hahne F, et al. flowCore: a Bioconductor package for high throughput flow cytometry. BMC Bioinformatics. 2009;10:106. doi: 10.1186/1471-2105-10-106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Gentleman RC, et al. Bioconductor: open software development for computational biology and bioinformatics. Genome Biol. 2004;5:R80. doi: 10.1186/gb-2004-5-10-r80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Mookerjee SA, Gerencser AA, Nicholls DG, Brand MD. Quantifying intracellular rates of glycolytic and oxidative ATP production and consumption using extracellular flux measurements. J. Biol. Chem. 2017;292:7189–7207. doi: 10.1074/jbc.M116.774471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Espinosa JA, Pohan G, Arkin MR, Markossian S. Real-time assessment of mitochondrial toxicity in HepG2 cells using the Seahorse extracellular flux. Analyzer. Curr. Protoc. 2021;1:e75. doi: 10.1002/cpz1.75. [DOI] [PubMed] [Google Scholar]
- 103.Schneider CA, Rasband WS, Eliceiri KW. NIH image to ImageJ: 25 years of image analysis. Nat. Methods. 2012;9:671–675. doi: 10.1038/nmeth.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Rueden CT, et al. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017;18:529. doi: 10.1186/s12859-017-1934-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Schindelin J, et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods. 2012;9:676–682. doi: 10.1038/nmeth.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Tinevez JY, et al. TrackMate: an open and extensible platform for single-particle tracking. Methods. 2017;115:80–90. doi: 10.1016/j.ymeth.2016.09.016. [DOI] [PubMed] [Google Scholar]
- 107.Tarantino N, et al. Tnf and il-1 exhibit distinct ubiquitin requirements for inducing NEMO-IKK supramolecular structures. J. Cell Biol. 2014;204:231–245. doi: 10.1083/jcb.201307172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Quiros PM, Goyal A, Jha P, Auwerx J. Analysis of mtDNA/nDNA ratio in mice. Curr. Protoc. Mouse Biol. 2017;7:47–54. doi: 10.1002/cpmo.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Myers MB, Mittelstaedt RA, Heflich RH. Using ΦX174 DNA as an exogenous reference for measuring mitochondrial DNA copy number. Biotechniques. 2009;47:867–869. doi: 10.2144/000113222. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data are available from the corresponding author on a reasonable request.