

SUMMARY
A health emergency situation occurred in Bulgaria in 2007 when positive rabies cases were notified in Sofia district in the central-western part of the country, suggesting a southward spread of the disease for the first time in the last 10 years. Phylogenetic analysis on 49 isolates sampled between 2009 and 2011 showed, for the first time, evidence of the existence of NEE and D clustered lineages in Bulgaria. Their geographical distribution clearly reveals the permeability of natural barriers, as already suggested by the disease spread that occurred across the Balkan mountain range in 2007. The monitoring and passive surveillance programmes conducted since the first 2009 oral vaccination campaign, the spatio-temporal evolution of the disease in the country since 2007, and the need for further investigation of the role of jackals in virus dispersion are discussed.
Key words: Bulgaria, epidemiology, oral vaccination, phylogeny, rabies virus
Bulgaria is a South-eastern European country covering some 110 994 km2 bordered by Romania to the North (608 km), Serbia and the Republic of Macedonia to the West (318 and 148 km), Greece and Turkey to the South (494 and 240 km) and the Black Sea to the East (354 km). Bulgaria's history of rabies began with the occurrence of rabies in dogs – especially stray dogs – followed by a second epidemic phase with the development of rabies in wildlife via the red fox (Vulpes vulpes), which acted as both reservoir and vector. As already demonstrated throughout the history of rabies in Europe, Bulgaria also provides a good example of spillover in dogs and foxes as reservoirs. Rabies was officially notified in Bulgaria for the first time in 1896. Up to the last 50 years of the 20th century, rabies occurred in the classical form of ‘street rabies', with dogs being the main reservoir of infection [1]. National measures on rabies control, including the compulsory vaccination of dogs since 1949, resulted in a substantial decrease in the number of outbreaks. Wildlife-mediated rabies was documented for the first time in Bulgaria in 1974 in the northern part of the country. From the 1990s, an average of 15 rabies cases per year was detected until 2006 [2], through passive surveillance defined as investigations carried out on suspect animals exhibiting signs suggestive of the disease or acting abnormally along with a number of animals found dead, including road kills [3].
Following the integration of Bulgaria in the European Union in 2007 and since the implementation of the first oral rabies vaccination (ORV) campaigns in 2009, an additional sampling plan was established, known as ORV monitoring (or active surveillance) [4]. Foxes killed by hunters were sampled in vaccinated areas to evaluate the efficacy of ORV campaigns in terms of bait consumption (bait uptake), herd immunity and rabies incidence. The brains of sampled animals from both surveillance and monitoring programmes were submitted for rabies diagnosis to the National Reference Laboratory for Rabies (NDRVMI) in Sofia. The diagnostic technique used was the gold-standard fluorescent antibody test followed by the mouse inoculation test as a confirmatory test and also for producing and storing the strains. The methods used were those described in the WHO manual [5]. While the number of reported rabies cases increased from nine in 2006 to 59 in 2009, there has been a marked fall in the number of reported cases since 2009, leading to a single case in both 2011 and 2012 (Fig. 1). The decrease in the number of detected cases since 2009 could highlight the potential success of the ORV programmes initiated in 2009 [6]. However, this decrease is also connected to a reduction in the number of samples analysed under the passive surveillance programme since 2009 (Fig. 1). While rabies control programmes from 2006 to 2008 focused on passive surveillance only (554 tested samples), the implementation of ORV campaigns from 2009 integrated sampling of animals from ORV monitoring in the rabies diagnosis scheme (2061 samples tested under monitoring and 272 under passive surveillance over the 2009–2012 period), significantly increasing the total number of samples submitted for rabies diagnosis. The monitoring consisted of the sampling of healthy foxes in vaccinated areas and was designed to complement (not replace) sampling of suspect wild and domestic animals for rabies surveillance [3]. While ORV is in progress, it is necessary to increase and maintain the pressure through passive surveillance sampling in the whole country in order to allow a more accurate evaluation and comparison of rabies incidence between years and to be able to detect the last rabies foci. This need remains pertinent due to the current rabies situation in bordering countries. Rabies in Romania and Serbia has been considered endemic for many years [1] and the epidemiological situation of fox rabies in South-eastern Europe has recently been updated, revealing a resurgence of rabies in the Republic of Macedonia since July 2011 [7] and in Northern Greece since October 2012 (K. E. Tasioudi, personal communication). These new data in two countries bordering Bulgaria confirm the need to maintain a high level of surveillance throughout the country.
Fig. 1.

[colour online]. Evolution of sampling in passive surveillance and oral rabies virus monitoring, and number of positive cases detected from 2006 to 2012.
Spatial evolution of rabies in Bulgaria has shown for a long time the presence of rabies restricted to the North of the country [2]. The reason for the definitely predominant spread of rabies in North Bulgaria in 2007 may be linked to the geographical features of the country. The northern part of Bulgaria is separated from the southern part by a natural geographical barrier, the Balkan mountains (Stara Planina mountain range). This mountain range initially acted as a temporary natural barrier for the spread of rabies from North to South. The rabies extension that occurred in 2007 to the South of the country highlights the permeability of natural barriers as already described in Western Europe in the 1980s and 1990s, and more recently in Italy [8]. The opportunistic red fox, which is well-suited to a wide range of landscapes, can potentially spread rabies in every type of environment. Recent Italian ORV campaigns conducted in 2010 have shown evidence of the existence of rabies virus (RABV) in high-altitude mountainous areas [8]. Vaccine baits were initially distributed below an altitude of 1000 m in winter and below 1500 m in spring to avoid repeated freeze–thaw cycles. Unexpected fox rabies hotspots were detected above these threshold altitudes, probably representing local residual rabies foci which may have contributed to maintaining the infectious cycle. Based on the results, it was finally decided to implement ORV at altitudes up to 2300 m [8]. As a result, high-altitude mountains can no longer be considered an impermeable barrier to the spread of rabies. Regarding domestic animals, a more random extension of the disease may be feasible with the use of modern transportation by humans and domestic animals, allowing long distances to be covered in a short time. This supports the need to proceed, as soon as a rabies case is detected, with the compulsory vaccination of all dogs and cats in the area in addition to strict legislation on stray dog control, to avoid any unexpected expansion of the disease.
Between 2006 and October 2012, 165 confirmed rabies cases were reported in Bulgaria. Of these positive cases, 120 (73%) were wild animals, including 110 (92%) foxes and ten (0·8%) jackals (Canis aureus). Domestic animals represented 45 (27%) cases including 18 (40%) dogs and 22 (49%) cats. The red fox, which represented 67% of all positive cases, appears to remain the main reservoir of rabies in Bulgaria. Because the history of rabies in Europe has been experienced with many sporadic spillover episodes between species, the co-existence of red foxes and jackals in Bulgaria, two close members of the Canidae family, raises the question of the involvement of jackals in rabies transmission within the country. While in Turkey jackal cases have rarely been detected [9], in Israel, red foxes and also, to a lesser extent, golden jackals have been identified as primary vectors, maintaining endemic wildlife rabies in that country [10]. Although there is no doubt about the current red fox's involvement in maintaining the disease in Bulgaria, the potential impact of the jackal's biology on rabies dynamics requires closer investigation. An assessment of the effectiveness of bait attractiveness and immunogenicity in jackals would also be useful as, to our knowledge, it has never been assessed with the type of baits used during the Bulgarian ORV programmes.
This first ORV programme implemented in 2009 covered the North of the country and was extended to four administrative districts in the centre – Sofia town, and Sofia, Pernik and Kyustendil districts (corresponding to a total of 18 districts and 61882 km2 or 55 037 km2 excluding settlements) due to the detection of 16 cases in the southern part of the country in 2007 (Fig. 2). In 2010 and 2011, following the detection of rabies cases outside the 2009 vaccination area, the ORV area was extended to include a new administrative district in the South-east of the country (Bourgas district with the municipalities of Pomorie and Ruen). The new vaccination area covered 62 985 km2 (55 995 km2 excluding settlements) and included 19 districts. While two ORV campaigns were conducted in spring and autumn 2009, only one was carried out in spring 2010 and spring 2011. The vaccine bait density used for all campaigns was 20 baits/km2, representing a total of 1 100 740 oral vaccines per campaign in 2009 and 1 119 900 baits in 2010 and in 2011. Following the detection of a rabies case in November 2011 in the Republic of Macedonia, just 13 km from the Bulgarian border, the vaccination area was again extended to include a vaccination belt west of Blagoevgrad district (border of the Republic of Macedonia). In 2012, the oral vaccination of foxes was consequently undertaken over an area of 66 550 km2, including 20 administrative districts. A total of 2 662 000 baits were dropped for the spring and autumn campaigns (national programme for control and eradication of rabies in Bulgaria, 2012). Over the 2009–2012 period, six vaccination campaigns were finally carried out, with a total of 7 103 280 doses distributed. The vaccine baits (Lysvulpenpruced; Bioveta, Czech Republic), contained a modified live-virus (SAD Bern) vaccine strain. The aerial distribution of the vaccine baits involved fixed-wing aircraft fitted with a fully automated computer-aided system with GPS which recorded both flight data and the location of dropped baits. The aircraft flew at an altitude of 150–200 m above the ground at an average speed of 150–200 km/h. Vaccine baits were dropped along parallel flight lines 500 m apart. Bait uptake was assessed by analysing the fluorescence of tetracycline under ultraviolet light in a tooth section from red fox lower jaws [11]. Blood samples collected from foxes shot in the vaccinated areas were titrated using an indirect ELISA kit (Platelia Rabies II kit; Bio-Rad, France) as described previously [12].
Fig. 2.
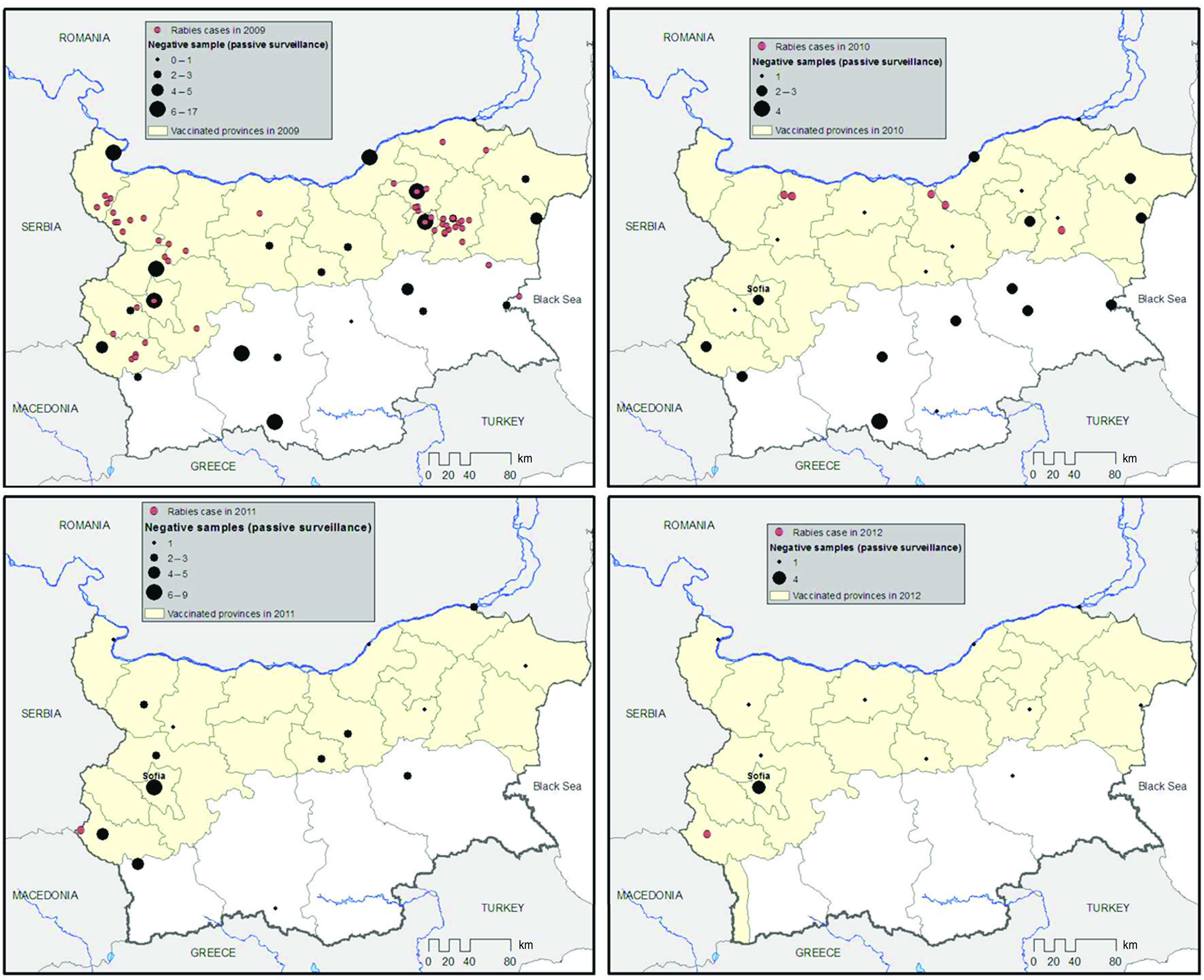
[colour online]. Location of positive and negative cases (passive surveillance) in Bulgaria from 2009 to 2012 and location of oral vaccination areas.
Analysis of the annual seroprevalence of foxes between 2009 and 2011 from a total of 544 blood samples showed stable results (2009: 18·3% positives for 191 samples tested; 2010: 17·0% positives for 112 samples tested; 2011: 18·7% positives for 241 samples tested). Of 923 tooth samples submitted for the detection of tetracycline, 195 were found positive, showing evidence of bait consumption. The bait uptake rate was estimated at 11% in 2009 (n = 225 tested) and 12% in 2010 (n = 242 tested), rising to 31% in 2011 (n = 456 tested). These data differ significantly from year to year (χ2 = 52, P χ2 < 0·001). It is considered that about 70% of the fox population need to be immunized to be able to control rabies [4]. The seroprevalence (19% in 2011) and the proportion of tetracycline-positive samples (31% in 2011) in this study indicated a lower vaccination coverage than expected. This could be due to the recency of ORV in Bulgaria plus the fact that only one ORV campaign was performed in both 2010 and 2011, during spring, when two-thirds of the fox population are cubs which are more difficult to assess by the baiting strategy. Two vaccination campaigns conducted annually during spring and autumn for several years have led to a lasting elimination of the disease in most of Western Europe [4]. Furthermore, the commercial ELISA test used has already shown a lack of sensitivity that could explain the unexpectedly low proportion of seroprevalence in this study [12].
Forty-nine Bulgarian isolates selected over the 2009–2011 period, covering the variability of regions and species in which RABV was identified (fox, n = 36; jackal, n = 2; cat, n = 6; dog, n = 2; cow, n = 1; goat, n = 1; sheep, n = 1) were included in a phylogenetic analysis. For each tested sample, 25 μl of 10% (v/w) brain homogenate suspension was deposited on an FTA® card (Whatman), inactivating the viruses and protecting RNA from degradation, as described previously by Picard-Meyer et al. [13]. RNA extraction, cDNA synthesis and PCR amplification of the nucleoprotein (N) gene with primers JW12 and JW6 were performed as described previously [14]. The glycoprotein (G) gene (740 bp fragment) was amplified with primers GH3 (positions 3891–3908) and GH4 (positions 4621–4602) [15]. One-step hnRT–PCR amplification was performed as described by Picard-Meyer et al. [16]. The phylogenetic analysis was performed after alignment of the consensus sequences. The bootstrap probabilities of each node were computed using the Neighbour-Joining method and bootstrap values over 70% were considered as statistically significant. Duplicate sequences of 100% identity were removed from the dataset. Ten partial N-gene sequences and ten partial G-gene sequences were finally included in the subsequent analysis. Analysis of the sequences revealed that all the strains corresponded to RABV field isolates and clearly differed from the sequence of the SAD Bern RABV strain used during ORV. Detailed analysis of the G and N genes provided an identical phylogenetic tree topology, therefore only the N-gene analysis is referred to in this study. The nucleotide difference in sequences revealed the existence of at least three distinct RABVs. The supplementary online material presents more detailed information on the phylogenetic analysis: the complete list of rabies viruses included in the analysis (Table S1) and the resulting Neighbour-Joining phylogenetic tree (Fig. S1). Half of the isolates clustered with the Eastern Europe (EE) group [15] (GenBank accession numbers: JX679399, JX679402, JX679407, JX679408) while the second distinctive lineage clustered with the North-eastern Europe (NEE) group [15]. (GenBank accession numbers: JX679401, JX679403–JX679406). Finally, a single fox isolate sampled in 2009 joined the centre of the European part of Russia (D) group with 98% nucleotide identity within the group, thus constituting the third lineage of the Bulgarian samples studied (GenBank accession number: JX679399). The geographical distribution of the different phylogroups (EE group, n = 25; NEE group, n = 23; D group, n = 1) is represented in Figure 3. The first study attempting to characterize a panel of Bulgarian RABVs was investigated by Johnson et al. [17]. The phylogenetic analysis of 10 isolates collected from 2002 to 2003, one isolate collected in 1994 and one isolate in 1997, suggested that the majority of samples belonged to a single specific Bulgarian group. The possible existence of a sublineage between samples from eastern and western districts was suggested. In 2011, McElhinney et al. [18] showed that the Bulgarian isolates fitted the EE group that initially clustered isolates from Austria, Poland, Czech Republic, Hungary, Romania, Croatia, Bosnia-Herzegovina, Montenegro, Serbia and Macedonia [15, 18, 19] and also fitted a newly identified Serbian fox (SF) group. Our results corroborate the presence of the EE group in the West of the country but not the presence of the SF group. There is no clear explanation regarding the absence of the SF group. It could reflect changes in the spatial dynamics of the disease or could imply that more samples should have been analysed. This study identified, for the first time in Bulgaria, RABVs belonging to the NEE group (known to be located in Western Russia, Finland, Estonia, Latvia, Lithuania, Poland, Romania and Slovakia [15, 19, 20]) and to the D group (initially identified in Romania, Hungary, West Turkey and West Russia [19, 20]). The single group D isolate of the study was detected in a fox sample from Pleven city in North Bulgaria in 2009. As both NEE and D groups had previously been identified in Romania [19], their detection in the north-western part of Bulgaria shows evidence of the movement of infected hosts between Romania and Bulgaria, probably resulting from a westward Ukrainian or Moldavian incursion. Although the Danube River, which defines the boundary between Romania and Bulgaria (with the exception of a 130-km stretch of land), could act as a natural barrier, the movement of RABV between the two countries has been clearly identified. The virus could be spread by rabid animals crossing the 130-km stretch of land forming a common boundary or crossing the river using a bridge, dam or other human infrastructures, as was observed when rabies entered Serbia from Romania and Hungary in 1977 [1]. Mountainous areas have also been shown to physically impede the movement of vectors. The Balkans range runs from West to East, dividing Bulgaria in two across the middle of the country. The phylogenetic analysis provides a picture of the lineage distribution, the country being clearly divided into North and South. Our results show that the RABV variant distribution does not exactly correlate to the landscape, as the EE variant found in the West of Bulgaria is found both in the North-west and South-west, inside and outside the Balkan mountain region. A higher barrier permeability and higher host species density implies greater connectivity and a better spread of the disease. This study clearly reveals an example of permeability in a mountainous landscape as previously described in Italy [8]. The spread of the EE group from the North-west to the South-west of Bulgaria could have been stimulated by a high fox population density. Further investigations into the population dynamics of red foxes could help clarify the spread of rabies.
Fig. 3.

[colour online]. Location of the three different Bulgarian lineage groups based on the phylogenetic analysis of the 49 rabies virus isolates sampled between 2009 and 2011.
Supplementary Material
Supplementary information supplied by authors.
ACKNOWLEDGEMENTS
We thank Tsvetanka Gencheva from the Bulgarian National Diagnostic Research Veterinary Medical Institute and Melanie Biarnais from the French Agency for Food, Environmental and Occupational Health & Safety for their excellent technical assistance. This research was jointly funded by the European Commission and the Bulgarian Food Safety Agency.
Supplementary material
For supplementary material accompanying this paper visit https://doi.org/10.1017/S0950268813001556.
click here to view supplementary material
DECLARATION OF INTEREST
None.
REFERENCES
- 1.Lontai I. Rabies in Hungary, Romania, Moldova and Bulgaria. In: King AA, Fooks AR, Aubert M, Wandeler AI, eds. Historical Perspective of Rabies in Europe and the Mediterranean Basin. Paris: OIE, 2004, pp. 119–127. [Google Scholar]
- 2.European Commission. Report on the task force meeting of the rabies subgroup. ITask force meeting of the rabies subgroup. Plovdiv, Bulgaria: European Commission, 2011, pp. 1–9. [Google Scholar]
- 3.Cliquet F, et al. Development of harmonised schemes for monitoring and reporting of rabies in animals in the European Union. EFSA Scientific Report, 2010, pp. 1–60.
- 4.European Commission. The oral vaccination of foxes against rabies. Strasbourg: European commission – Health and Consumer Protection, 23 October 2002. [Google Scholar]
- 5.Meslin FX, Kaplan MM, Koprowski H. Laboratory Techniques in Rabies, 4th edn. Geneva: World Health Organization, 1996, pp. 476. [Google Scholar]
- 6.Ilieva D. Epizootic characteristics and prevalence of rabies in Bulgaria. Journal of Agricultural Science 2011; 10: 59–64. [Google Scholar]
- 7.Kirandjiski T, et al. First reported cases of rabies in the republic of Macedonia Veterinary Record 2012; 170: 1–2. [DOI] [PubMed] [Google Scholar]
- 8.Mulatti P, et al. Emergency oral rabies vaccination of foxes in Italy in 2009–2010: identification of residual rabies foci at higher altitudes in the Alps. Epidemiology and Infection 2012; 140: 591–598. [DOI] [PubMed] [Google Scholar]
- 9.Johnson N, et al. Rabies epidemiology and control in Turkey: past and present. Epidemiology and Infection 2010; 138: 305–312. [DOI] [PubMed] [Google Scholar]
- 10.Yakobson BA, et al. Rabies vaccination programme for red foxes (Vulpes vulpes) and golden jackals (Canis aureus) in Israel (1999–2004). In: Dodet B, Schudel A, Pastoret PP, Lombard M, eds. First International Conference on Rabies in Europe. Basel: Karger, 2006, pp. 133–140. [PubMed] [Google Scholar]
- 11.Robardet E, et al. First European interlaboratory comparison of tetracycline and age determination with red fox teeth following oral rabies vaccination programs. Journal of Wildlife Diseases 2012; 48: 858–868. [DOI] [PubMed] [Google Scholar]
- 12.Knoop EV, et al. Evaluation of a commercial rabies ELISA as a replacement for serum neutralization assays as part of the pet travel scheme and oral vaccination campaigns of foxes. Berliner und Münchener Tierärztliche Wochenschrift 2010; 123: 278–285. [PubMed] [Google Scholar]
- 13.Picard-Meyer E, Barrat J, Cliquet F. Use of filter paper (FTA) technology for sampling, recovery and molecular characterisation of rabies viruses. Journal of Virological Methods 2007; 140: 174–182. [DOI] [PubMed] [Google Scholar]
- 14.Heaton PR, et al. Heminested PCR assay for detection of six genotypes of rabies and rabies-related viruses. Journal of Clinical Microbiology 1997; 35: 2762–2766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bourhy H, et al. Ecology and evolution of rabies virus in Europe. Journal of General Virology 1999; 80: 2545–2557. [DOI] [PubMed] [Google Scholar]
- 16.Picard-Meyer E, et al. Development of a hemi-nested RT-PCR method for the specific determination of European bat lyssavirus 1. Comparison with other rabies diagnostic methods. Vaccine 2004; 22: 1921–1929. [DOI] [PubMed] [Google Scholar]
- 17.Johnson N, et al. Evidence for trans-border movement of rabies by wildlife reservoirs between countries in the Balkan Peninsular. Veterinary Microbiology 2007; 120: 71–76. [DOI] [PubMed] [Google Scholar]
- 18.McElhinney LM, et al. Molecular diversity and evolutionary history of rabies virus strains circulating in the Balkans. Journal of General Virology 2011; 92: 2171–2180. [DOI] [PubMed] [Google Scholar]
- 19.Turcitu MA, et al. Molecular epidemiology of rabies in Romania provides evidence for a high degree of heterogeneity and virus diversity. Virus Research 2010; 150: 28–33. [DOI] [PubMed] [Google Scholar]
- 20.McElhinney LM, et al. Molecular epidemiology of rabies viruses in Europe. In: Dodet B, Schudel A, Pastoret PP, Lombard M, eds. First International Conference on Rabies in Europe. Basel: Karger, 2006, pp. 17–28. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary information supplied by authors.
For supplementary material accompanying this paper visit https://doi.org/10.1017/S0950268813001556.
click here to view supplementary material