

Abstract
Background
Venom production has evolved independently many times in the animal kingdom, although it is rare among mammals. Venomous shrews produce toxins in their salivary glands and use their venoms to hunt and store prey. Thus far, the toxicity and composition of shrew venoms have been studied only in two shrew species: the northern short-tailed shrew, Blarina brevicauda, and the Eurasian water shrew, Neomys fodiens. Venom of N. fodiens has potent paralytic activity which enables hunting and storing prey in a comatose state. Here, we assayed the hemolytic effects of extracts from salivary glands of N. fodiens and the common shrew, Sorex araneus, in erythrocytes of Pelophylax sp. frogs. We identified toxins in shrew venom by high-performance liquid chromatography coupled to tandem mass spectrometry.
Results
Our results prove, confirming a suggestion made four centuries ago, that S. araneus is venomous. We also provide the first experimental evidence that shrew venoms produce potent hemolysis in frog erythrocytes. We found significant concentration-dependent effects of venoms of N. fodiens and S. araneus on hemolysis of red blood cells evaluated as hemoglobin release. Treatment of erythrocytes with N. fodiens venom at concentrations of 1.0 and 0.5 mg/ml and with S. araneus venom at concentration of 1.0 mg/ml caused an increased release of hemoglobin. Our findings confirm that hemolytic effects of N. fodiens venom are stronger than those produced by S. araneus venom. We identified four toxins in the venom of N. fodiens: proenkephalin, phospholipase A2 (PLA2), a disintegrin and metalloproteinase domain-containing protein (ADAM) and lysozyme C, as well as a non-toxic hyaluronidase. In the venom of S. araneus we found five toxins: proenkephalin, kallikrein 1-related peptidase, beta-defensin, ADAM and lysozyme C. PLA2 and ADAMs are likely to produce hemolysis in frog erythrocytes.
Conclusions
Our results clearly show that shrew venoms possess hemolytic action that may allow them to hunt larger prey. Since a member of the numerous genus Sorex is venomous, it is likely that venom production among shrews and other eulipotyphlans may be more widespread than it has previously been assumed.
Supplementary Information
The online version contains supplementary material available at 10.1186/s40851-022-00191-5.
Keywords: cytotoxicity, eulipotyphlans, hemolysis, mammalian venom, prey hunting, shrews, venom evolution
Background
The use of venom for prey acquisition, predator deterrence and competition is widespread in the animal kingdom [1, 2]. However, in mammals venom production is rare and restricted to the members of four extant orders, the monotremes, eulipotyphlans, chiropterans and primates [3–6]. Most venomous mammals belong to the order Eulipotyphla [7], which consists of moles, shrews, solenodons and hedgehogs [8], but venomous representatives have been found only among shrews and solenodons thus far [7, 9–15].
Venomous eulipotyphlans use their venoms to capture and immobilize prey for long-term storage [4, 6, 16]. Such food hoarding can save energy and time spent on foraging and capturing prey, and minimize the risk of predation [7, 16]. Thus, prey hoarding brings considerable benefits to eulipotyphlans, especially the Soricinae shrews which, due to extremely high metabolic rate, have to consume large amounts of food to meet their energetic demands [17, 18].
Studies on the toxicity and composition of eulipotyphlan venoms are scarce. Among shrews, only venoms of the northern short-tailed shrew, Blarina brevicauda, and the Eurasian water shrew, Neomys fodiens (Fig. 1a), have been analyzed thus far [9, 12, 13, 15, 19]. The venom of B. brevicauda has proteolytic and hypotensive activity [12], while venom of N. fodiens is known to produce potent paralytic effects [9, 13]. Pucek [9] was the first to observe the paralysis of the limbs and posterior part of the body in experimental animals after treatment with the crude extract from salivary glands of N. fodiens. Recently, Kowalski et al. [13] confirmed strong neurotoxic and lower cardioinhibitory activity of the venom of N. fodiens.
Fig. 1.

Venomous shrew species (a) The Eurasian water shrew, Neomys fodiens. (b) The common shrew, Sorex araneus. Photos: M. Mounier (a) and S. von Merten (b)
Animal venoms usually are of complex nature in both composition and toxicity [20–22]. For instance, venoms of many taxa such as snakes, scorpions, marine snails and jellyfish produce potent cytotoxic effects [23–30] which enable them to rapidly subdue prey and avoid retaliatory injuries, especially if the attacked prey is large or potentially dangerous [7]. Proteases, phospholipases, and cytolytic or hemolytic agents are among the primary molecules responsible for the venom cytotoxicity [26]. Snake venom metalloproteinases (SVMPs) are known to provide hemotoxic activity of snake venoms, particularly in species from the Viperidae family and Crotalinae subfamily [31, 32]. Because eulipotyphlans often prey upon larger prey, some of which may be even larger than themselves [11, 16, 33–35], it is likely that their venoms also display cytotoxicity, though it has not yet been verified.
Our recent study confirmed that N. fodiens venom contains a phospholipase A2 (PLA2) [13]. PLA2 is known for varied toxic activities, including hemolysis [24, 25, 27, 28]. Thus, the presence of PLA2 in the venom of N. fodiens might be indicative of hemolytic effects of its venom. With this in mind, in the present study we aimed to determine the hemolytic activity of salivary gland extracts of N. fodiens and, for comparison, the common shrew, Sorex araneus (Fig. 1b) hitherto considered non-venomous. To achieve our goal we performed physiological bioassays on the erythrocytes of Pelophylax sp. frogs. We hypothesized that N. fodiens venom would produce potent and concentration-dependent hemolysis in frog erythrocytes, while saliva of S. araneus [13] would not present any hemolytic effects. Because only few venom proteins have been found in shrew venoms so far [12–15, 19], we also aimed to identify new toxins in the venom of N. fodiens and non-toxic molecules in the saliva of S. araneus. To do so, we performed proteomic analyses to determine protein content in both, the whole extract from venom glands of both shrew species and in fractions separated by high-performance liquid chromatography.
Results
Shrew venoms produce hemolysis in frog erythrocytes
According to our predictions, Triton® X-100 (positive control) added to the red blood cells of frogs caused hemolysis when compared to the treatment with Ringer’s solution (RS; negative control; non-parametric Mann–Whitney U-test: U = 9.0, p < 0.005; Fig. 2).
Fig. 2.
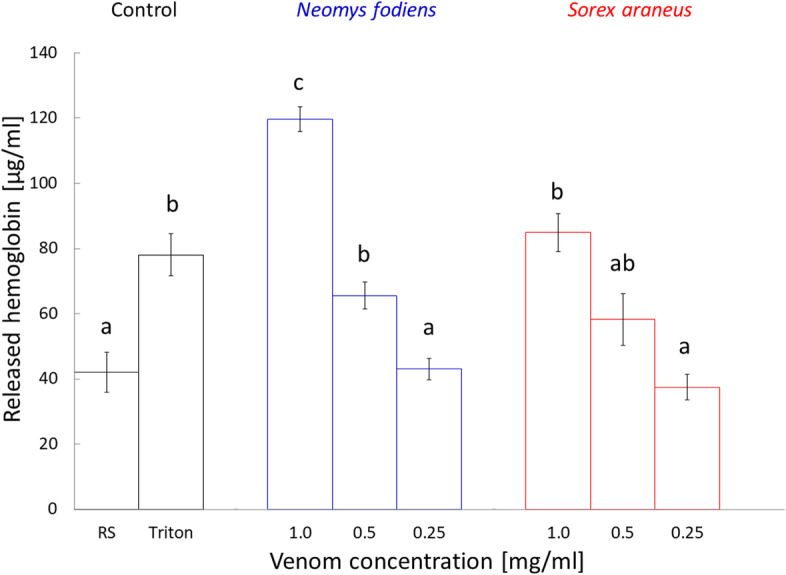
Hemolysis produced by shrew venoms expressed as concentration of released hemoglobin. Bars represent mean ± SEM. Different letters represent significant differences between treatments, controls and species
The venom of N. fodiens added to the erythrocytes resulted in a concentration-dependent hemolytic response (non-parametric Kruskal–Wallis test: χ2 = 47.5, df = 2, p < 0.0001; Fig. 2), expressed as μg of released hemoglobin per ml. Treatment of erythrocytes with the venom at concentration of 1.0 mg/ml produced a significant increased release of hemoglobin as compared with control treatments (Fig. 2): RS (parametric Student’s t-test: t = -10.7, df = 28, p < 0.0001) and Triton (U = 18, p < 0.0001). At concentration of 0.5 mg/ml, it caused a higher hemoglobin release as compared with RS (t = -3.2, df = 28, p < 0.01), but similar to that caused by Triton (U = 151, p = 0.17). Venom of N. fodiens at concentration of 0.25 mg/ml did not cause significant changes in hemoglobin concentration as compared with RS (U = 86.5, p = 0.72), but the concentration of released hemoglobin was lower in comparison to the treatment with Triton (U = 209, p < 0.0001).
Venom of N. fodiens at concentration of 1.0 mg/ml added to erythrocytes produced a higher increase in hemoglobin concentration as compared with the treatments with venom concentrations of 0.5 and 0.25 mg/ml (Fig. 2; t = 9.6, df = 40, p < 0.0001, and U = 441, p < 0.0001, respectively). Also, addition of venom at concentration of 0.5 mg/ml caused a higher hemoglobin release in comparison to the treatment with venom concentration of 0.25 mg/ml (U = 370.5, p < 0.001).
Addition of the venom of S. araneus to the red blood cells also resulted in a concentration-dependent hemolytic response (χ2 = 31.7, df = 2, p < 0.0001; Fig. 2). Treatment with venom at concentration of 1.0 mg/ml caused a significant increased release of hemoglobin when compared with RS (U = 2, p < 0.001), but it was similar to the treatment with Triton (U = 27, p = 0.33). At concentration of 0.5 mg/ml, it did not produce changes in hemoglobin concentration as compared with RS (U = 17, p = 0.13) and Triton (U = 59, p = 0.07). Similarly, addition of S. araneus venom at concentration of 0.25 mg/ml to erythrocytes did not cause any changes in the concentration of released hemoglobin when compared with RS (U = 47, p = 0.90), but the hemoglobin concentration was lower when compared to the treatment with Triton (U = 110, p < 0.0001).
Treatment of the red blood cells with S. araneus venom at concentration of 1.0 mg/ml revealed a higher increase in the concentration of released hemoglobin in comparison to the treatments with venom at concentrations of 0.5 and 0.25 mg/ml (Fig. 2; U = 41, p < 0.05, and U = 70, p < 0.001, respectively). There were no differences in concentrations of released hemoglobin when treatments with venom at concentrations of 0.5 and 0.25 mg/ml were compared (U = 52, p = 0.11).
Venom of N. fodiens has stronger hemolytic activity than venom of S. araneus
We found significant differences in hemolytic activity of venoms of N. fodiens and S. araneus (χ2 = 63.4, df = 5, p < 0.0001). N. fodiens venom at concentration of 1.0 mg/ml added to erythrocytes caused a higher increase in hemoglobin release than treatment with S. araneus venom at the same concentration (Fig. 2; U = 141, p < 0.0001). There were no differences in concentrations of released hemoglobin when treatments with N. fodiens and S. araneus venoms at concentration of 0.5 mg/ml were compared (U = 88, p = 0.47). The same was true for comparison of addition of N. fodiens and S. araneus venoms at concentrations of 0.25 mg/ml (U = 121, p = 0.52).
Venom of N. fodiens at concentration of 1.0 mg/ml caused a higher increase in hemoglobin release than S. araneus venom at concentrations of 0.5 and 0.25 mg/ml (U = 147, p < 0.0001, and U = 210, p < 0.0001, respectively). Also, addition of N. fodiens venom at concentration of 0.5 mg/ml caused a higher hemoglobin release in comparison to the treatment with S. araneus venom concentration of 0.25 mg/ml (U = 189, p = 0.0001).
Venom of S. araneus at concentration of 1.0 mg/ml caused a higher increase in hemoglobin release than N. fodiens venom at concentrations of 0.5 and 0.25 mg/ml (U = 112, p = 0.04, and U = 142, p < 0.0001, respectively). There were no differences in concentrations of released hemoglobin when treatments with S. araneus venom at concentration of 0.5 mg/ml and N. fodiens venom at concentration of 0.25 mg/ml were compared (U = 106, p = 0.09).
Toxins identified in shrew venoms
We analyzed protein content in both the whole extract from venom glands of N. fodiens and S. araneus and in fractions separated by high-performance liquid chromatography (Fig. 3a,b) using MS/MS based proteomic techniques. We identified four toxins: proenkephalin-A, phospholipase A2 (PLA2), a disintegrin and metalloproteinase domain-containing protein (ADAM) and lysozyme C (Table 1; Fig. 3c) in the extract from venom glands of N. fodiens. We also found a hyaluronidase, a toxin spreading factor, in N. fodiens venom. We found five toxins, namely proenkephalin-A, kallikrein 1-related peptidase, beta-defensin 7, ADAM and lysozyme C (Table 1; Fig. 3c) in the extract from venom glands of S. araneus.
Fig. 3.

Chromatographic separation of the extract from venom glands of Neomys fodiens (a) and Sorex araneus (b). Numbers indicate the fractions selected to identify toxins. (c) Toxins identified in the selected fractions (see Additional files 1 and 2 in the Supplementary information section for the complete list of proteins identified in the extracts from venom glands of both shrew species). Abbreviations: ADAM, a disintegrin and metalloproteinase domain-containing protein; KLK1, kallikrein 1-related peptidase; PLA2, phospholipase A2. ( +) indicates the presence while (-) indicates the absence of toxin in the given fraction
Table 1.
Toxins identified in venoms of the Eurasian water shrew, Neomys fodiens, and the common shrew, Sorex araneus
| Shrew species | Accession code | Identified peptides | Protein name |
|---|---|---|---|
| Neomys fodiens | P01211 |
K.LPSLKTWETCK.E K.KYGGFMK.R K.YGGFMK.R K.RYGGFLK.R |
Proenkephalin-A |
| P14422 | FAKFLSYK | Phospholipase A2 | |
| Q9Z0F8 | SEDIKDFSR | Disintegrin and metalloproteinase domain-containing protein 17 | |
| P12067 | YWCNDGK | Lysozyme C | |
| gi|521,028,001 | KDIEFYIPK | Hyaluronidase PH-20a | |
| Sorex araneus | P01211 | YGGFMK + Oxidation (M) | Proenkephalin-A |
| Q61754 | DKSNDLMLLR | Kallikrein 1-related peptidase b24 | |
| Q91V70 | FQIPEK | Beta-defensin 7 | |
| Q10741 | LYSDGKK | Disintegrin and metalloproteinase domain-containing protein 10 | |
| P12067 | AWVAWR | Lysozyme C |
ahyaluronidase does not display toxic activity but is a toxin spreading factor commonly found in animal venoms
Non-toxic molecules identified in shrew venoms
We identified 194 proteins in N. fodiens venom gland extract involved in various aspects of cell functioning such as: cell maintenance, cell cycle regulation, cell division, cell migration, cell adhesion and cell–cell interactions, cell aging and apoptosis, intra- and extracellular transport, signal transduction, energy metabolism, lipid metabolism, immune and stress response, and blood coagulation.
We found 112 proteins in the venom gland extract of S. araneus involved in cell maintenance, cell cycle regulation and cell division, cell migration and cell–cell interactions, cell respiration and apoptosis, intra- and extracellular transport, signal transduction, energy metabolism, lipid metabolism, immune and stress response, and blood coagulation. All proteins determined in the extracts from venom glands of N. fodiens and S. araneus and in separated fractions are listed in Supplementary Tables A1 and A2, respectively (see Additional files 1 and 2).
Discussion
Shrew venoms are known to display strong paralytic and hypotensive in vitro and in vivo effects, which may enable hunting and storing prey in a comatose state [7, 9, 12, 13, 15]. Our study provides the first experimental evidence that shrew venoms produce potent hemolysis in the red blood cells of frogs, allowing shrews to hunt and kill larger prey such as amphibians. Intriguingly, for the first time, our results confirm the toxicity of saliva of S. araneus, which enables us to classify the common shrew as a venomous mammal. These findings have important implications for understanding the ecological functions and evolution of venoms in eulipotyphlans.
Venoms of many animal taxa, including snakes and lizards, fish, wasp and ants, jellyfish and anemones – but not mammals – have been proven to produce hemolytic effects [36–43]. For instance, venoms of Australian elapid snakes Austrelaps superbus and Pseudechis colletti produce high direct hemolysis of washed rabbit erythrocytes due to the hydrolysis of phosphatidylcholine in the cell membrane to fatty acids and glycerophosphorylcholine by phospholipase B [23]. Fernández et al. [39] demonstrated strong hemolytic effects induced by the venom of the Eastern coral snake, Micrurus fulvius, after intravenous injection into mice and dogs. Similarly, Lenske et al. [41] observed severe hemolysis in a dog envenomed by a red-bellied black snake, Pseudechis porphyriacus, venom. Also, venoms of the Gila monsters (genus Heloderma) H. suspectum and H. horridum are known to produce hemolysis [37]. Gilatoxins present in their saliva are devoid of PLA2 activity, but display proteolytic, hemorrhagic and hemolytic properties [37].
Cytolysin, a cytolytic toxin isolated from Scorpaena plumieri scorpionfish venom, showed a potent hemolytic activity on washed rabbit erythrocytes [38]. Venom of the Central American scorpion Didymocentrus krausi leads to direct hemolysis on human and rabbit erythrocytes [30]. Also, hemolysin isolated from Pogonomyrmex badius ant venom causes direct hemolysis on bovine and mouse washed erythrocytes [36, 44].
Our results confirmed, for both shrew species, significant concentration-dependent effects of venom on hemolysis of the red blood cells of frogs. Similar concentration-dependent hemolytic effects have recently been found in cnidarian venoms [40, 43]. It is noteworthy that most studies on hemolytic action of animal venoms have been performed on laboratory animals (including mice, rabbits and dogs) or humans. In our study, we used Pelophylax sp. frogs, which in nature are hunted by both N. fodiens and S. araneus [16, 33, 45, 46]. Therefore, we conclude that venom may help shrews to subdue and consume larger prey such as frogs.
Our results also show that hemolytic effects of N. fodiens venom are stronger than those produced by S. araneus venom. Both field observations and laboratory experiments indicate that N. fodiens is more effective in hunting and consuming frogs than S. araneus [33, 47]. This might be because N. fodiens produces stronger venom than S. araneus does, as demonstrated in this study. Thus, shrew venoms might have evolved not only to paralyze and immobilize prey to make food stores but also to kill and eat it quickly to meet their high energetic demands [7]. These findings are useful for understanding ecological functions of venom and predator–prey interactions.
Animal venoms are extremely rich and complex sources of biologically active molecules (usually proteins) with diverse toxicity [20–22] that are responsible for the pathophysiological consequences of envenoming [20]. Venom of a single animal may contain more than 1000 toxins [22, 48] making the venom a potent tool in hunting prey and repelling predators and competitors [2, 22]. Intriguingly, despite the varied diets of shrews [33, 47, 49, 50], toxin diversity in shrew venoms appears to be relatively low suggesting that the use of venom by shrews may be of recent evolutionary origin [7, 15].
Venom of B. brevicauda consists of six toxins, including blarina toxin (BLTX), soricidin, kallikrein-1 serine protease paralog (KLK1-BL2), PLA2, antileukoproteinase, and a tissue factor pathway inhibitor 2 protein, as well as four non-toxic proteins: hyaluronidase, blarinasin, and two KLK1 paralogs [12, 15, 19, 51]. The venom of N. fodiens also appears to have a relatively simple composition [13]. In the present study, we identified 199 proteins (see Additional file 1), including four toxins (Table 1), in the extract from venom glands of N. fodiens, and 117 proteins (see Additional file 2), including five toxins (Table 1), in the extract from venom glands of S. araneus. Thus, our results also confirm the simple composition of shrew venoms. Admittedly, it seems still possible to identify new molecules with toxic activity in shrew venoms, but it is not likely that their content would reach several hundred toxins as in venoms of Conus snails [48].
Lysozyme C, PLA2 and hyaluronidase identified here in N. fodiens venom have been previously reported in this species [3, 13]. Similarly, lysozyme C, KLK1 and beta-defensin have been found in the saliva of S. araneus [13]. Proenkephalin and a disintegrin and metalloproteinase domain-containing protein (ADAM) are reported here for both shrew species for the first time. Proenkephalin, which contains the known toxin peptide soricidin, has recently been characterized in the venom of B. brevicauda [15]. Therefore, identification of this molecule in venoms of N. fodiens and S. araneus suggests that soricidin may be more common among venomous shrews.
Metalloproteinases (ADAMs) have not been found in shrew venoms thus far, but they commonly occur in venoms of other taxa, including snakes, scorpions and jellyfish [24, 29, 30, 32, 52]. These molecules display hemolytic activity [32, 52]; therefore it is possible that hemolysis produced by shrew venoms in frog erythrocytes may be caused, at least partially, by ADAMs.
PLA2, here found only in the venom of N. fodiens, is widely distributed among elapid and viperid snake venoms and in scorpion venoms [27, 53, 54], and is also known for its cytotoxicity, including hemolytic activity [27, 29, 40, 55, 56]. For instance, PLA2 from Cerastes ceratses venom produces indirect hemolysis [56]. Lazcano-Pérez et al. [40] confirmed a dose-dependent hemolytic activity of cnidarian venom, which contains PLA2. Hemolytic effects of PLA2 from marine snail and wasp venoms have been also demonstrated [25, 42]. Thus, we consider PLA2 to be the most likely candidate toxin that could be responsible for the hemolytic properties of venom of N. fodiens. However, further studies with isolated venom components are required to confirm whether PLA2 or ADAM contained in shrew venoms produces hemolytic effects. The mode of action of these molecules also needs further validation.
Because lysozyme C is involved in antimicrobial defense [57, 58], it was thought to help in the maintenance of oral hygiene in venomous mammals [7]. Although there are some components with antimicrobial activity in venoms of B. brevicauda and N. fodiens [13, 15], this venom function has never been empirically tested in shrews [7]. Defensins, here identified in the venom of S. araneus, have previously been found in platypus venom [59]. These molecules exhibit a myo- and neurotoxic activity, modifying voltage-sensitive sodium channels, which usually results in a potent analgesic effect [60].
Kallikrein-like proteins, here also found in the venom of S. araneus, have earlier been detected in the venom of B. brevicauda [12]. BLTX has a tissue kallikrein-like protease activity. This toxin cleaves kininogens to kinins, such as bradykinin, a common mediator of inflammation that increases vascular permeability and lowers blood pressure [12]. Other kallikrein-1 paralogs in B. brevicauda venom have recently been discovered by Hanf and Chavez [15]. Therefore, these kinins are thought to be the primary toxic agents of shrew venom responsible for symptoms such as dyspnea, hypotension and hypokinesia, recorded previously in pharmacological studies [12].
As in our previous study [13], here we found a hyaluronidase in the venom of N. fodiens. This non-toxic protein is commonly present in many animal venoms, including venoms of snakes, spiders and ants [44, 61, 62], and acts as a toxin spreading factor [60, 61]. Due to its ability to hydrolyze connective tissue, hyaluronidase facilitates the action of other venom components [26]. Thus, it is possible that also in the venom of N. fodiens hyaluronidase facilitates the spread of venom proteins [13]. However, how the toxins are spread still requires investigation.
We provide here, for the first time, a comprehensive list of non-toxic proteins contained in venoms of both shrew species. Similarly, numerous non-toxic components have recently been identified in the venom and saliva of the Hispaniolan solenodon, Solenodon paradoxus [14]. These proteins may be responsible for the maintenance of homeostasis, synthesis of complex toxins, and their secretion into the extracellular compartment and secretion upregulation. We also found many proteins related to cell division and cell cycle regulation, which might indicate a high epithelial cell turnover in the venom glands of shrews.
The high number of non-toxic components found in our study may result from analyzing the whole extracts from salivary glands of shrews instead of crude venom, of which only limited amounts can be extracted in shrews [7]. As animals usually inject only venom into a target species during a bite, we recommend extracting only venom for future biochemical studies. On the other hand, to understand the molecular mechanisms underlying the evolution of shrew venoms it is necessary to study not only the venom toxins but also the molecular machinery related to venom production and secretion.
Proteins recruited into venoms and acting as toxins might also be present in non-venomous animals, but devoid of toxic activity [20, 21]. None of the venom proteins directly identified here show similarity to toxins reported in other venomous mammals. Therefore, in future studies we suggest using genomic, transcriptomic and proteomic data to predict venom protein identity based on sequence similarity to previously described animal toxins. Finally, to validate the toxicological role of venom proteins, it is also necessary to ascertain their mode of action.
Venom has evolved multiple times throughout the animal kingdom, but is rare among mammals [5, 63, 64]. However, it must be emphasized that most of potentially venomous mammal species have not yet been studied [7]. One of the first suggestions that shrews are venomous is from a description of the European common shrew, Sorex araneus, in Historie of Foure-footed Beasts: “It is a ravening beast, feigning itself gentle and tame, but being touched it biteth deep, and poisoneth deadly. It beareth a cruel mind, desiring to hurt anything, neither is there any creature it loveth” [65]. Even the Latin name (aranea) suggests that it would have venomous bites like a spider. However, shrew venoms are not harmful or deadly to humans. According to our personal experience, a bite of a water shrew on a finger causes at most some pain, swelling or numbness of the finger for 1–2 days.
Recent studies confirmed that B. brevicauda and N. fodiens produce toxic substances in their salivary glands [12, 13, 15], but toxic effects of the saliva of S. araneus were not confirmed [13]. Here, our study provides the first experimental evidence that saliva of the common shrew has cytotoxic activity leading to hemolysis in the red blood cells of frog. Thus, our results prove, confirming a suggestion made four centuries ago [65], that S. araneus is indeed venomous. This is also the first report confirming venom production in a member of the genus Sorex, which includes 86 shrew species [66]. This suggests that more venomous shrew species may be discovered in the future. Two more Sorex species – S. palustris and S. cinereus – are already suspected of being venomous [4, 7], but the toxicity of their saliva has not yet been studied. Further research will allow us to investigate questions about how mammalian venoms have evolved. It is likely that venom production in shrews, and other eulipotyphlans, may be more widespread than it has previously been assumed. However, much more research effort is required to determine which shrew species do or do not produce venom and how widespread venom truly is. Finally, because animal toxins have applications in medicine and pharmacy [19, 67, 68], characterization of new toxins in eulipotyphlan venoms, and in so numerous and widespread species as S. araneus, provides a promising avenue to explore bioactive venom components as therapeutic agents in the future [69, 70].
Conclusions
Our results clearly show that shrew venoms produce hemolysis in frog erythrocytes, which suggests that this may allow these shrews to hunt larger prey. Because the venom of N. fodiens causes stronger hemolytic effects than venom of S. araneus, N. fodiens may be more effective in killing and storing such large prey as frogs than S. araneus. Since a member of the numerous genus Sorex is venomous, it is likely that venom production among shrews and other eulipotyphlans may be more widespread than it has previously been assumed.
Materials and methods
Animals
Shrews
Trapping sessions were performed in the suburbs of Poznań (western Poland) from July to September 2017. In total, we captured 12 water shrews and 12 common shrews. The captured animals were transported to the laboratory and placed separately into large (39 × 21 × 28 cm; 23 l) terraria equipped with bedding (a mixture of peat, moss and sand). Each terrarium contained a shelter (upturned clay flowerpot) and a bowl of water. Food (minced beef and live mealworms, earthworms and snails) and water were provided ad libitum. Shrews were kept in an animal room under controlled conditions (temperature: 21 ± 1 °C; humidity: 65–70%; artificial photoperiod: 12L:12D). After a week, they were killed using approved methods to obtain their submandibular salivary glands, in which toxins are produced [13].
Frogs
Nine frogs of the genus of Pelophylax sp. were captured using a net near ponds and small water tanks located in the Morasko district of Poznań (western Poland). Captured animals were placed into plastic transporters and carried to the laboratory, where their body mass (mean mb [g] ± SEM: 126.2 ± 12.8) and snout to vent length (mean SVL [mm] ± SEM: 92.4 ± 3.84) were measured. Next, they were placed into large (46 × 30 × 28 cm; 39 l) aqua-terraria (up to 3 animals per terrarium) equipped with bedding (a mixture of peat and sand). The terraria were regularly irrigated to maintain high humidity. Each terrarium contained a shelter (clay flowerpot) and a water tank to allow frogs to submerge in water. Food (live mealworms and crickets) and water were provided ad libitum. Frogs were kept (until the experimental procedure began) in the animal room under controlled conditions (temperature: 20 ± 1 °C; humidity: 65–70%; artificial photoperiod: 12L:12D).
Venom collection and sample preparation
Shrews were killed by cervical vertebrae dislocation, and their submandibular salivary (venom) glands were dissected to obtain toxic saliva [13]. Pairs of glands (n = 2; from each shrew species) designed for hemolytic bioassays were transferred into 600 μl of Ringer’s solution (RS) for frogs (125 mM NaCl; 3 mM KCl; 1.8 mM CaCl2; 10 mM glucose; 5 mM HEPES). Pairs of glands (n = 10, from each shrew species) designed for chromatographic separation were transferred into 600 μl of methanol. Tissues were next homogenized, and samples were centrifuged at 10,000 × g and 4 °C for 30 min. The supernatants were collected, and the protein content was determined using a Direct Detect spectrometer (MERCK Millipore, Warsaw, Poland).
Chromatographic separation
Supernatants suspended in methanol were used for separation before peptide analysis by reverse phase high-performance liquid chromatography (RP-HPLC). Separation was performed using a Dionex Ultimate 3000 chromatographic system comprising a dual pump programmable solvent module. Supernatants were analyzed using a BioBasic-18 analytical column (5 μm, 150 × 4.6 mm; Thermo Fisher Scientific). The samples were eluted with a gradient of 5–60% acetonitrile (ACN)/0.1% TFA with a flow rate of 0.5 ml/min for 55 min. The eluent was monitored at 214 nm, and fractions were collected into 1.5-ml tubes.
Hemolytic assay
Frogs were killed using approved methods [13] and blood was sampled from the heart and put into a tube containing 5 μl of 3.2% sodium citrate, an anticoagulant. Two blood samples were additionally taken using heparinized capillary tubes to estimate the hematocrit (mean Ht [%] ± SEM: 28.3 ± 0.77). Blood (50 μl) was re-suspended in 50 μl of RS and centrifuged at 1500 × g for 5 min at 4 °C. The supernatants were then removed, 100 μl of RS was added and the procedure was repeated two more times. The pellets were finally diluted with 100 μl of RS. Next, 10- and 100-fold dilutions were prepared, and were used for the experiments. The erythrocytes were then incubated (final volume 100 μl) at 25 °C for 1 h in the presence of N. fodiens saliva (with protein concentrations of 1.0, 0.5, and 0.25 mg/ml) or S. araneus saliva (with the same protein concentrations), and centrifuged at 1500 × g for 5 min at 4 °C. Samples containing red blood cells with RS were used as a negative control, whereas samples with Triton® X-100 as a positive control. The number of replicates (n) for each venom concentration was as follows: 21 for N. fodiens, 7 to 10 for S. araneus, 9 for RS and 11 for Triton. The released hemoglobin was measured in the final supernatants using a spectrophotometer at 415 nm, and compared with a standard curve constructed with bovine hemoglobin [71]. The results were expressed as μg hemoglobin per ml.
Protein and toxin identification
Proteomic analysis was carried out at the Mass Spectrometry Laboratory, Institute of Biochemistry and Biophysics, Polish Academy of Sciences, Warsaw. Peptides from the whole extract and separated fractions (that proved to be the most active [13] and with the highest protein content; Fig. 1d,e) of N. fodiens and S. araneus venom glands were analyzed by liquid chromatography coupled to tandem mass spectrometry LC-(MS–MS/MS) using a Nano-Acquity LC system (Waters, Milford, MA) and an OrbitrapVelos mass spectrometer (Thermo Electron Corp., San Jose, CA).
Before performing the analysis, the proteins were subjected to an ion-solution digestion procedure. Proteins were (1) reduced with 50 mM TCEP for 30 min at 60 °C, (2) alkylated with 200 mM MMTA for 30 min at room temperature and (3) digested overnight with trypsin (sequencing Grade Modified Trypsin—Promega V5111). Next, the samples were applied to an RP-18 precolumn (nanoACQUITY Symmetry® C18—Waters 186,003,514) using water containing 0.1% TFA as a mobile phase and were transferred to a nano-HPLC RP-18 column (nanoACQUITY BEH C18 – Waters 186,003,545). The samples were eluted with a gradient of 0–35% acetonitrile in the presence of 0.05% formic acid with a flow rate of 250 nl/min for 180 min. The column was directly coupled to the ion source of the spectrometer working within data dependent on the MS to MS/MS switch. To ensure a lack of cross contamination from previous samples, each analysis was preceded by a blank run.
The proteins were identified by a Mascot Search (Matrix Science, London, UK) against the SwissProt and NCBInr databases. Because only a few toxins have been identified in shrew venoms so far, and only in two shrew species, we also searched for toxin/protein sequences found in other venomous animal taxa. The search parameters were as follows: type of search: MS/MS Ion Search; enzyme specificity: trypsin; fixed methylthio modification of cysteine; variable modifications: methionine oxidation; mass values: monoisotopic; protein mass: unrestricted; peptide mass tolerance: 30 ppm; fragment mass tolerance: 0.1 D; number of missed cleavage sites allowed: 1; instrument type: HCD. Peptides with Mascot scores exceeding the threshold value of p < 0.05 were considered positively identified.
Data analysis
All data are presented as the mean value ± SEM (standard error of the mean) of the indicated number of replicates (n). Before the statistical analysis, the normality of the data distribution was checked using the Shapiro–Wilk test. To determine the effects of saliva of both shrew species on hemolysis in frog erythrocytes the Kruskal–Wallis test was used. The mean differences between the treatment (number of replicates for each venom concentration: n = 21 for N. fodiens and n = 7–10 for S. araneus) and control groups (n = 9 for RS and n = 11 for Triton) were determined using the paired two-sided Student’s t-test or Wilcoxon signed-rank test. To indicate statistically significant differences between hemolytic activity of N. fodiens saliva and S. araneus saliva, the Mann–Whitney U-test was performed. Non-parametric tests (Wilcoxon signed-rank and Mann–Whitney) were used when the datasets of non-normal distributions were compared. The statistical analyses were carried out using R software [72]. Significant results were considered those with a p-value of p < 0.05.
Supplementary Information
Additional file 1: Table A1. Protein identification in the extract from venom glands of the Eurasian water shrew, Neomys fodiens, based on tandem mass spectrometry analysis. Toxins are shown in bold.
Additional file 2: Table A2. Protein identification in the extract from venom glands of the common shrew, Sorex araneus, based on tandem mass spectrometry analysis. Toxins are shown in bold.
Acknowledgements
We are very grateful to Urszula Eichert and Marianna Pejka for help in the field work and performing physiological assays.
Abbreviations
- ADAM
A disintegrin and metalloproteinase domain-containing protein
- ADAMs
Metalloproteinases
- BLTX
Blarina Toxin
- Ht
Hematocrit
- KLK1-BL2
Kallikrein-1 serine protease paralog
- LC-(MS–MS/MS)
Liquid Chromatography Coupled to Tandem Mass Spectrometry
- mb
Body mass
- PLA2
Phospholipase A2
- RP-HPLC
Reverse Phase High-performance Liquid Chromatography
- RS
Ringer’s Solution
- SVL
Snout to Vent Length
- SVMPs
Snake Venom Metalloproteinases
Authors’ contributions
All authors conceived and designed the experiments. KK and LR captured experimental animals. KK and PM performed the experiments and analyzed the data. KK wrote the first draft of the paper. All authors revised and edited the manuscript. All authors read and approved the final manuscript.
Funding
The research was financially supported by a grant from the National Science Centre, Poland (no. 2015/17/N/NZ8/01567 to K.K.) and, in part, by the budgets of the Department of Systematic Zoology and Department of Animal Physiology and Developmental Biology (Faculty of Biology, AMU, Poznań).
Availability of data and materials
All data are available in the main text and the supplementary information files. Further information and requests for data should be directed to and will be fulfilled by the corresponding author.
Declarations
Ethics approval and consent to participate
Trapping procedure, handling, housing of animals and all measurements were done in accordance with relevant guidelines and regulations. All experimental protocols were approved by licensing committee: the Regional Directorate for Environmental Protection in Poznań, Poland (decision # WPN-II.6401.249.2017.AC). The study was carried out in compliance with the ARRIVE guidelines.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Krzysztof Kowalski, Email: k.kowalski@umk.pl.
Paweł Marciniak, Email: pmarcin@amu.edu.pl.
Leszek Rychlik, Email: leszek.rychlik@amu.edu.pl.
References
- 1.Bücherl W, Buckley EE, Deulofeu V. Venomous Animals and Their Venoms. Venomous Vertebrates. London: Academic Press; 1968. [Google Scholar]
- 2.Zhang Y. Why do we study animal toxins? Zool Res. 2015;36(4):183–222. doi: 10.13918/j.issn.2095-8137.2015.4.183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Dufton MJ. Venomous mammals. Pharmacol Ther. 1992;53(2):199–215. doi: 10.1016/0163-7258(92)90009-O. [DOI] [PubMed] [Google Scholar]
- 4.Ligabue-Braun R, Verli H, Carlini CR. Venomous mammals: a review. Toxicon. 2012;59(7–8):680–695. doi: 10.1016/j.toxicon.2012.02.012. [DOI] [PubMed] [Google Scholar]
- 5.Rode-Margono JE, Nekaris KIA. Cabinet of curiosities: venom systems and their ecological function in mammals, with a focus on primates. Toxins. 2015;7(7):2639–2658. doi: 10.3390/toxins7072639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ligabue-Braun R. Venom use in mammals: evolutionary aspects. In: Gapalakrishnakone P, Malhotra A, editors. Evolution of venomous animals and their toxins. Toxinology. The Netherlands: Springer-Dordrecht; 2016. pp. 235–57. [Google Scholar]
- 7.Kowalski K, Rychlik L. Venom use in eulipotyphlans: an evolutionary and ecological approach. Toxins. 2021;13(3):231. doi: 10.3390/toxins13030231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Douady CJ, Douzery EJP. Hedgehogs, shrews, moles, and solenodons (Eulipotyphla) In: Hedges SB, Kumar S, editors. The Timetree of Life. New York: Oxford University Press; 2009. pp. 495–498. [Google Scholar]
- 9.Pucek M. The effect of the venom of the European water shrew (Neomys fodiens fodiens Pennant) on certain experimental animals. Acta Theriol. 1959;3(6):93–108. doi: 10.4098/AT.arch.59-6. [DOI] [Google Scholar]
- 10.Pucek M. Neomys anomalus Cabrera, 1907 – a venomous mammal. Bull Acad Pol Sci. 1969;17:569–573. [Google Scholar]
- 11.Tomasi TE. Function of venom in the short-tailed shrew. Blarina brevicauda J Mammal. 1978;59(4):852–854. doi: 10.2307/1380150. [DOI] [PubMed] [Google Scholar]
- 12.Kita M, Nakamura Y, Okumura Y, Ohdachi SD, Oba Y, Yoshikuni M, Kido H, Uemura D. Blarina toxin, a mammalian lethal venom from the short-tailed shrew Blarina brevicauda: isolation and characterization. PNAS. 2004;101(20):7542–7547. doi: 10.1073/pnas.0402517101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kowalski K, Marciniak P, Rosiński G, Rychlik L. Evaluation of the physiological activity of venom from the Eurasian water shrew Neomys fodiens. Front Zool. 2017;14:46. doi: 10.1186/s12983-017-0230-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Casewell NR, Petras D, Card DC, Suranse V, Mychajliw AM, Richards D, Koludarov I, Albulescu LO, Slagboom J, Hempel BF, Ngum NM, Kennerley RJ, Brocca JL, Whiteley G, Harrison RA, Bolton FMS, Debono J, Vonk FJ, Alföldi J, Johnson J, Karlsson EK, Lindblad-Toh K, Mellor IR, Süssmuth RD, Fry BG, Kuruppu S, Hodgson WC, Kool J, Castoe TA, Barnes I, Sungar K, Undheim EAB, Turvey ST. Solenodon genome reveals convergent evolution of venom in eulipotyphlan mammals. PNAS. 2019;116(51):25745–25755. doi: 10.1073/pnas.1906117116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hanf ZR, Chavez AS. A comprehensive multi-omic approach reveals a relatively simple venom in a diet generalist, the Northern short-tailed shrew. Blarina brevicauda Genome Biol Evol. 2020;12(7):1148–1166. doi: 10.1093/gbe/evaa115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kowalski K, Rychlik L. The role of venom in the hunting and hoarding of prey differing in body size by the Eurasian water shrew. Neomys fodiens J Mammal. 2018;99(2):351–362. doi: 10.1093/jmammal/gyy013. [DOI] [Google Scholar]
- 17.Churchfield S. The Natural History of Shrews. 1. London: Christopher Helm/A & C Black; 1990. [Google Scholar]
- 18.Taylor JRE. Evolution of energetic strategies in shrews. In: Wójcik JM, Wolsan M, editors. Evolution of Shrews. Białowieża: Mammal Research Institute of Polish Academy of Sciences; 1998. pp. 309–346. [Google Scholar]
- 19.Bowen CV, DeBay D, Ewart HS, Gallant P, Gormley S, Ilenchuk TT, Iqbal U, Lutes T, Martina M, Mealing G, Merkley N, Sperker S, Moreno MJ, Rice C, Syvitski RT, Stewart JM. In vivo detection of human TRPV6-rich tumors with anti-cancer peptides derived from soricidin. PLoS ONE. 2013;8(3):e58866. doi: 10.1371/journal.pone.0058866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fry BG, Roelants K, Champagne DE, Scheib H, Tyndall JD, King GF, Nevalainen TJ, Norman J, Lewis RJ, Norton RS, Renjifo C, de la Vega RCR. The toxicogenomic multiverse: convergent recruitment of proteins into animal venoms. Annu Rev Genom Hum Genet. 2009;10:483–511. doi: 10.1146/annurev.genom.9.081307.164356. [DOI] [PubMed] [Google Scholar]
- 21.Casewell NR, Wüster W, Vonk FJ, Harrison RA, Fry BG. Complex cocktails: the evolutionary novelty of venoms. Trends Ecol Evol. 2013;28(4):219–229. doi: 10.1016/j.tree.2012.10.020. [DOI] [PubMed] [Google Scholar]
- 22.Utkin YN. Animal venom studies: current benefits and future developments. World J Biol Chem. 2015;6(2):28–33. doi: 10.4331/wjbc.v6.i2.28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bernheimer AW, Weinstein SA, Linder R. Isoelectric analysis of some Australian elapid snake venoms with special reference to phospholipase B and hemolysis. Toxicon. 1986;24(8):841–849. doi: 10.1016/0041-0101(86)90109-1. [DOI] [PubMed] [Google Scholar]
- 24.Soto JG, Perez JC, Minton SA. Proteolytic, hemorrhagic and hemolytic activities of snake venoms. Toxicon. 1988;26(9):875–882. doi: 10.1016/0041-0101(88)90328-5. [DOI] [PubMed] [Google Scholar]
- 25.Zarai Z, Bacha AB, Horchani H, Bezzine S, Zouari N, Gargouri Y, Mejdoub H. A novel hepatopancreatic phospholipase A2 from Hexaplex trunculus with digestive and toxic activities. Arch Biochem Biophys. 2010;494(2):121–129. doi: 10.1016/j.abb.2009.11.020. [DOI] [PubMed] [Google Scholar]
- 26.Lee H, Jung E, Kang C, Yoon WD, Kim JS, Kim E. Scyphozoan jellyfish venom metalloproteinases and their role in the cytotoxicity. Toxicon. 2011;58(3):277–284. doi: 10.1016/j.toxicon.2011.06.007. [DOI] [PubMed] [Google Scholar]
- 27.Estrada-Gómez S, Vargas Muñoz LJ, Saldarriaga-Córdoba M, Quintana Castillo JC. Venom from Opisthacanthus elatus scorpion of Colombia, could be more hemolytic and less neurotoxic than thought. Acta Trop. 2016;153:70–78. doi: 10.1016/j.actatropica.2015.09.019. [DOI] [PubMed] [Google Scholar]
- 28.Cedro RCA, Menaldo DL, Costa TR, Zoccal KF, Sartim MA, Santos-Filho NA, Faccioli LH, Sampaio SV. Cytotoxic and inflammatory potential of a phospholipase A2 from Bothrops jararaca snake venom. J Venom Anim Toxins incl Trop Dis. 2018;24:33. doi: 10.1186/s40409-018-0170-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Choudhary I, Hwang DH, Lee H, Yoon WD, Chae J, Han CH, Yum S, Kang C, Kim E. Proteomic analysis of novel components of Nemopilema nomurai jellyfish venom: deciphering the mode of action. Toxins. 2019;11(3):153. doi: 10.3390/toxins11030153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rojas-Azofeifa D, Sasa M, Lomonte B, Diego-García E, Ortiz N, Bonilla F, Murillo R, Tytgat J, Díaz C. Biochemical characterization of the venom of Central American scorpion Didymocentrus krausi Francke, 1978 (Diplocentridae) and its toxic effects in vivo and in vitro. Comp Biochem Physiol C. 2019;217:54–67. doi: 10.1016/j.cbpc.2018.11.021. [DOI] [PubMed] [Google Scholar]
- 31.Fox JW, Serrano SMT. Timeline of key events in snake venom metalloproteinase research. J Proteom. 2009;72(2):200–209. doi: 10.1016/j.jprot.2009.01.015. [DOI] [PubMed] [Google Scholar]
- 32.Olaoba OT, Karina Dos Santos P, Selistre-de-Araujo HS, Ferreira de Souza DH. Snake Venom Metalloproteinases (SVMPs): A structure-function update. Toxicon: X. 2020;7:100052. doi: 10.1016/j.toxcx.2020.100052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Buchalczyk T, Pucek Z. Food storage of the European water shrew, Neomys fodiens (Pennant, 1771) Acta Theriol. 1963;7(19):376–379. doi: 10.4098/AT.arch.63-22. [DOI] [Google Scholar]
- 34.Robinson DE, Brodie ED., Jr Food hoarding behavior in the short-tailed shrew Blarina brevicauda. Am Midl Nat. 1982;108(2):369–375. doi: 10.2307/2425498. [DOI] [Google Scholar]
- 35.Lopez-Jurado LF, Mateo JA. Evidence of venom in the Canarian shrew (Crocidura canariensis): immobilizing effects on the Atlantic lizard (Gallotia atlantica) J Zool. 1996;239(2):394–395. doi: 10.1111/j.1469-7998.1996.tb05460.x. [DOI] [Google Scholar]
- 36.Schmidt JO, Blum MS. Pharmacological and toxicological properties of harvester ant, Pogonomyrmex badius, venom. Toxicon. 1978;16(6):645–651. doi: 10.1016/0041-0101(78)90192-7. [DOI] [PubMed] [Google Scholar]
- 37.Hendon RA, Tu AT. Biochemical characterization of the lizard toxin gilatoxin. Biochem. 1981;20(12):3517–3522. doi: 10.1021/bi00515a033. [DOI] [PubMed] [Google Scholar]
- 38.Andrich F, Carnielli JBT, Cassoli JS, Lautner RQ, Santos RAS, Pimenta AMC, de Lima ME, Figueiredo SG. A potent vasoactive cytolysin isolated from Scorpaena plumieri scorpionfish venom. Toxicon. 2010;56(4):487–496. doi: 10.1016/j.toxicon.2010.05.003. [DOI] [PubMed] [Google Scholar]
- 39.Fernández ML, Quartino PY, Arce-Bejarano R, Fernández J, Camacho LF, Gutiérrez JM, Kuemmel D, Fidelio G, Lomonte B. Intravascular hemolysis induced by phospholipases A2 from the venom of the Eastern coral snake, Micrurus fulvius: Functional profiles of hemolytic and non-hemolytic isoforms. Toxicol Lett. 2018;286:39–47. doi: 10.1016/j.toxlet.2017.11.037. [DOI] [PubMed] [Google Scholar]
- 40.Lazcano-Pérez F, Zavala-Moreno A, Rufino-González Y, Ponce-Macotela M, García-Arredondo A, Cuevas-Cruz M, Gómez-Manzo S, Marcial-Quino J, Arreguín-Lozano B, Arreguín-Espinosa R. Hemolytic, anticancer and antigiardial activity of Palythoa caribaeorum venom. J Venom Anim Toxins incl Trop Dis. 2018;24:12. doi: 10.1186/s40409-018-0149-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lenske E, Padula AM, Leister E, Boyd S. Severe haemolysis and spherocytosis in a dog envenomed by a red-bellied black snake (Pseudechis porphyriacus) and successful treatment with a bivalent whole equine IgG antivenom and blood transfusion. Toxicon. 2018;151:79–83. doi: 10.1016/j.toxicon.2018.07.005. [DOI] [PubMed] [Google Scholar]
- 42.Gong J, Yuan H, Gao Z, Hu F. Wasp venom and acute kidney injury: the mechanisms and therapeutic role of renal replacement therapy. Toxicon. 2019;163:1–7. doi: 10.1016/j.toxicon.2019.03.008. [DOI] [PubMed] [Google Scholar]
- 43.Jafari H, Honari H, Zargan J, Jahromi ST. Identification and hemolytic activity of jellyfish (Rhopilema sp., Scyphozoa: Rhizostomeae) venom from the Persian Gulf and Oman Sea. Biodiversitas. 2019;20(4):1228–32. doi: 10.13057/biodiv/d200440. [DOI] [Google Scholar]
- 44.Schmidt JO, Blum MS. A harvester ant venom: chemistry and pharmacology. Sci. 1978;200(4345):1064–1066. doi: 10.1126/science.653354. [DOI] [PubMed] [Google Scholar]
- 45.Lyapkov SM. The influence of predators on brown frogs during and after metamorphosis. In: Kuzʹmin SL, editor. Advances in Amphibian Research in the Former Soviet Union. Sofia-Moscow: Pensoft Publishers; 1996. pp. 149–159. [Google Scholar]
- 46.Ivanter EV, Korosov AV, Makarov AM. Study of the trophic relations of small insectivorous mammals. Biol Bull. 2015;42(9):845–856. doi: 10.1134/S1062359015090058. [DOI] [Google Scholar]
- 47.Churchfield S. The feeding ecology of the European water shrew. Mammal Rev. 1985;15(1):13–21. doi: 10.1111/j.1365-2907.1985.tb00381.x. [DOI] [Google Scholar]
- 48.Davis J, Jones A, Lewis RJ. Remarkable inter- and intra-species complexity of conotoxins revealed by LC/MS. Peptides. 2009;30(7):1222–1227. doi: 10.1016/j.peptides.2009.03.019. [DOI] [PubMed] [Google Scholar]
- 49.Churchfield S. Dietary separation in three species of shrew inhabiting water-cress beds. J Zool (Lond) 1984;204(2):211–228. doi: 10.1111/j.1469-7998.1984.tb02371.x. [DOI] [Google Scholar]
- 50.Churchfield S, Rychlik L. Diets and coexistence in Neomys and Sorex shrews in Białowieża forest, eastern Poland. J Zool. 2006;269(3):381–390. doi: 10.1111/j.1469-7998.2006.00115.x. [DOI] [Google Scholar]
- 51.Kita M, Okumura Y, Ohdachi SD, Oba Y, Yoshikuni M, Nakamura Y, Kido H, Uemura D. Purification and characterisation of blarinasin, a new tissue kallikrein-like protease from the short-tailed shrew Blarina brevicauda: comparative studies with blarina toxin. Biol Chem. 2005;386(2):177–182. doi: 10.1515/BC.2005.022. [DOI] [PubMed] [Google Scholar]
- 52.Burin SM, Menaldo DL, Sampaio SV, Frantz FG. An overview of the immune modulating effects of enzymatic toxins from snake venoms. Int J Biol Macromol. 2005;109:664–671. doi: 10.1016/j.ijbiomac.2017.12.101. [DOI] [PubMed] [Google Scholar]
- 53.Doley R, Zhou X, Kini RM. Snake venom phospholipase A2 enzymes. In: Mackessy SP, editor. Handbook of venoms and toxins of reptiles. New York: CRC Press; 2010. pp. 173–198. [Google Scholar]
- 54.Harris JB, Scott-Davey T. Secreted phospholipases A2 of snake venoms: effects on the peripheral neuromuscular system with comments on the role of phospholipases A2 in disorders of the CNS and their uses in industry. Toxins. 2013;5(12):2533–2571. doi: 10.3390/toxins5122533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Vogel CW, Plückthun A, Müller-Eberhard HJ, Dennis EA. Hemolytic assay for venom phospholipase A2. Anal Biochem. 1981;118(2):262–268. doi: 10.1016/0003-2697(81)90189-5. [DOI] [PubMed] [Google Scholar]
- 56.Fatah C, Samah S, Fatima LD. Antiplatelet and anticoagulant activities of two phospholipase A2s purified from Cerastes cerastes venom: structure-function relationship. J Biochem Mol Toxicol. 2018;32(12):e22219. doi: 10.1002/jbt.22219. [DOI] [PubMed] [Google Scholar]
- 57.Low DHW, Sunagar K, Undheim EAB, Ali SA, Alagon AC, Ruder T, Jackson TNW, Pineda Gonzales S, King GF, Jones A, Antunes A, Fry BG. Dracula’s children: molecular evolution of vampire bat venom. J Proteom. 2013;89:95–111. doi: 10.1016/j.jprot.2013.05.034. [DOI] [PubMed] [Google Scholar]
- 58.van Hoek ML. Antimicrobial peptides in reptiles. Pharmaceuticals. 2014;7(6):723–753. doi: 10.3390/ph7060723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Whittington CM, Papenfuss AT, Locke DP, Mardis ER, Wilson RK, Abubucker S, Mitreva M, Wong ESW, Hsu AL, Kuchel PW, Belov K, Warren WC. Novel venom gene discovery in the platypus. Genome Biol. 2010;11(9):R95. doi: 10.1186/gb-2010-11-9-r95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Fry BG, Casewell NR, Wüster W, Vidal N, Young B, Jackson TNW. The structural and functional diversification of the Toxicofera reptile venom system. Toxicon. 2012;60(4):434–448. doi: 10.1016/j.toxicon.2012.02.013. [DOI] [PubMed] [Google Scholar]
- 61.Sannaningaiah D, Subbaiah GK, Kempaiah K. Pharmacology of spider venom toxins. Toxin Rev. 2014;33(4):206–220. doi: 10.3109/15569543.2014.954134. [DOI] [Google Scholar]
- 62.Gowda R, Rajaiah R, Angaswamy N, Krishna S, Sannanayak VB. Biochemical and pharmacological characterization of Trimersurus malabaricus snake venom. J Cell Biochem. 2018;119(7):5904–5912. doi: 10.1002/jcb.26782. [DOI] [PubMed] [Google Scholar]
- 63.Folinsbee KE. Evolution of venom across extant and extinct eulipotyphlans. C R Palevol. 2013;12(7–8):531–542. doi: 10.1016/j.crpv.2013.05.004. [DOI] [Google Scholar]
- 64.Arbuckle K. Evolutionary context of venom in animals. In: Gopalakrishnakone P, Malhotra A, editors. Evolution of Venomous Animals and Their Toxins. Toxinology. The Netherlands: Springer-Dordrecht; 2017. pp. 3–31. [Google Scholar]
- 65.Topsell RE. Historie of Foure-footed Beasts. London: Jaggard; 1607. [Google Scholar]
- 66.Mammal Diversity Database. https://mammaldiversity.org. Accessed 14 Feb 2021.
- 67.Koh DCI, Armugam A, Jeyaseelan K. Snake venom components and their applications in biomedicine. Cell Mol Life Sci. 2006;63(24):3030–3041. doi: 10.1007/s00018-006-6315-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Robinson SD, Vetter I. Pharmacology and therapeutic potential of venom peptides. Biochem Pharmacol. 2020;181:114207. doi: 10.1016/j.bcp.2020.114207. [DOI] [PubMed] [Google Scholar]
- 69.Clark GC, Casewell NR, Elliott CT, Harvey AL, Jamieson AG, Strong PN, Turner AD. Friends or foes? Emerging impacts of biological toxins. Trends Biochem Sci. 2019;44(4):365–379. doi: 10.1016/j.tibs.2018.12.004. [DOI] [PubMed] [Google Scholar]
- 70.Bordon KCF, Cologna CT, Fornari-Baldo EC, Pinheiro-Júnior EL, Cerni FA, Amorim FG, Anjolette FAP, Cordeiro FA, Wiezel GA, Cardoso IA, Ferreira IG, de Oliveira IS, Boldrini-França J, Pucca MB, Baldo MA, Arantes EC. From animal poisons and venoms to medicines: Achievements, challenges and perspectives in drug discovery. Front Pharmacol. 2020;11:1132. doi: 10.3389/fphar.2020.01132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Santamaría A, Sánchez-Rodríguez J, Zugasti A, Martínez A, Galván-Arzate S, Segura-Puertas L. A venom extract from the sea anemone Bartholomea annulata produces haemolysis and lipid peroxidation in mouse erythrocytes. Toxicol. 2002;173(3):221–228. doi: 10.1016/S0300-483X(02)00035-5. [DOI] [PubMed] [Google Scholar]
- 72.R Development Core Team R . A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing; 2018. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Table A1. Protein identification in the extract from venom glands of the Eurasian water shrew, Neomys fodiens, based on tandem mass spectrometry analysis. Toxins are shown in bold.
Additional file 2: Table A2. Protein identification in the extract from venom glands of the common shrew, Sorex araneus, based on tandem mass spectrometry analysis. Toxins are shown in bold.
Data Availability Statement
All data are available in the main text and the supplementary information files. Further information and requests for data should be directed to and will be fulfilled by the corresponding author.