

 This work is licensed under a
This work is licensed under a Abstract
Oocyte activation deficiency (OAD) remains the predominant cause of total/low fertilization rate in assisted reproductive technology. Phospholipase C zeta (PLCZ1) is the dominant sperm-specific factor responsible for triggering oocyte activation in mammals. OAD has been linked to numerous PLCZ1 abnormalities in patients experiencing failed in vitro fertilization or intracytoplasmic sperm injection cycles. While significant efforts have enhanced our understanding of the clinical relevance of PLCZ1, and the potential effects of genetic variants upon functionality, our ability to apply PLCZ1 in a diagnostic or therapeutic role remains limited. Artificial oocyte activation is the only option for patients experiencing OAD but lacks a reliable diagnostic approach. Immunofluorescence analysis has revealed that the levels and localization patterns of PLCZ1 within sperm can help us to indirectly diagnose a patient’s ability to induce oocyte activation. Screening of the gene encoding PLCZ1 protein is also critical if we are to fully determine the extent to which genetic factors might play a role in the aberrant expression and/or localization patterns observed in infertile patients. Collectively, these findings highlight the clinical potential of PLCZ1, both as a prognostic indicator of OAD and eventually as a therapeutic agent. In this review, we focus on our understanding of the association between OAD and PLCZ1 by discussing the localization and expression of this key protein in human sperm, the potential genetic causes of OAD, and the diagnostic tools that are currently available to us to identify PLCZ1 deficiency and select patients that would benefit from targeted therapy.
Male infertility
The incidence of infertility is rising as the global population continues to grow and is now considered to affect 8–12% of couples worldwide (Agarwal et al. 2021). According to the European Society for Human Reproduction and Embryology, the causes of infertility can be classified as sole-male factor (20–30%), sole-female factor (20–35%), due to both partners (25–40%), or idiopathic (10–20%) (ESHRE 2018). The Human Fertilization and Embryology Authority (HFEA) reports that the most common reasons for patients seeking in vitro fertilization (IVF) treatment are male infertility (37%), female infertility (31%), and cases involving unexplained infertility (32%) (HFEA 2018). Currently, male infertility is responsible for 25–30% of all cases of infertility and can arise as a result of testicular deficiency (such as varicocele), post-testicular impairment, or other genetic/molecular abnormalities that affect the production, quality, and function of sperm (Esteves 2016, Leslie et al. 2020, Agarwal et al. 2021). The field of assisted reproductive technology (ART) is revolutionizing the field of reproductive medicine and strives continuously to provide infertile patients with access to appropriate and efficient treatments. Indeed, intracytoplasmic sperm injection (ICSI) has now become the most commonly applied technique and accounts for two-thirds of all global ART treatments; conventional IVF only accounts for one-third of all treatments (Haddad et al. 2021). Regardless of the undoubtable success of ART, total fertilization failure (TFF), in which all of the mature oocytes retrieved from the female fail to fertilize in one cycle, occurs in 5–20% of all IVF cycles and 1–3% of all ICSI cycles (Cardona Barberán et al. 2020).
The principal cause of TFF following ICSI is oocyte activation deficiency (OAD). This is when an oocyte fails to undergo further maturation and complete fertilization following sperm fusion. Over recent years, OAD has been linked to numerous PLCZ1 abnormalities in patients experiencing failed IVF or ICSI cycles (Nikiforaki et al. 2016, Yeste et al. 2016a). Numerous studies have shown that PLCZ1 is responsible for inducing Ca2+ oscillations in the activating oocyte; these oscillations control crucial cell signalling pathways in the early embryo. Since the discovery of PLCZ1 in 2002, a significant body of research has demonstrated the fundamental role of PLCZ1 as the sperm-specific oocyte activation factor and have provided strong evidence to support a causal link between PLCZ1 deficiency and male infertility (Saunders et al. 2002, Sanders & Swann 2016, Swann & Lai 2016, Parrington et al. 2019). Previous research has demonstrated that patients with reduced levels of PLCZ1, or with a total absence of PLCZ1, are unable to initiate the Ca2+ oscillations that are required for successful oocyte activation. Furthermore, some reports have shown that levels of PLCZ1 protein and mutations in the gene encoding PLCZ1, are strongly associated with male infertility/subfertility (Kashir 2020). In addition, reports have shown that sperm from fertile patients can evoke variable Ca2+ responses; thus, such sperm may be responsible for the failure of some cycles (Swann 2018, Meng et al. 2020). Further investigations successfully demonstrated the association between the localization and expression levels of PLCZ1 and successful ICSI cycles (Yelumalai et al. 2015).
Yoon et al. were the first to inject human sperm from patients with a history of ICSI failure into mouse oocytes; this practice identified the absence of the normal Ca2+ oscillations and resulted in oocyte activation failure (Yoon et al. 2008). In addition, these authors demonstrated that the co-injection of Plcz1 mRNA into mouse oocytes, alongside PLCZ1-deficient sperm, successfully rescued OAD, thus reinforcing the fundamental role of PLCZ1 for the treatment of oocyte activation (Yoon et al. 2008). These findings highlighted the fact that investigating the specific expression of PLCZ1 protein in human sperm can facilitate our understanding of their ability to trigger oocyte activation. Ever since, research has become increasingly focused on the design of therapeutic and diagnostic applications for PLCZ1 and the translation of such technology to the clinical setting (Kashir et al. 2013, Nomikos et al. 2013a, b). In this review, we focus on our understanding of the association between OAD and PLCZ1 by discussing the localization and expression of this key protein in human sperm, the potential genetic causes of OAD, and the current diagnostic tools that are available to us to identify cases of PLCZ1 deficiency.
What is known about the localization patterns and expression levels of PLCZ1 in sperm
Previous research has shown that the specific localization patterns and expression levels of PLCZ1 in sperm are associated with OAD. Immunofluorescence analysis has revealed that PLCZ1 is predominantly localized in the equatorial segment of human sperm but is also localized in the acrosome, post-acrosome, tail, or a combination of these locations (Fig. 1) (Grasa et al. 2008, Kashir et al. 2013, Yelumalai et al. 2015, Ferrer-Vaquer et al. 2016, Yeste et al. 2016b). In human sperm, the most dominant region for PLCZ1 localization is the equatorial region; this makes biological sense because this would facilitate the release of PLCZ1 protein into the ooplasm following gamete fusion to initiate the sequence of events that are vital for embryogenesis (Nomikos et al. 2012). The localization of PLCZ1 in multiple spatial regions has raised speculation that different populations of this protein may potentially be responsible for differential functions (Grasa et al. 2008), although this theory has yet to be proven. For example, Grasa et al. reported that the equatorial localization of PLCZ1 remained consistent during sperm capacitation and the acrosome reaction and suggested that PLCZ1 localization in the acrosomal region of the sperm head may mediate the acrosome reaction rather than triggering oocyte activation (Grasa et al. 2008). It is possible that the acrosomal population of PLCZ1 may play a role in fertilization; however, due to the timing of the acrosome reaction in relation to sperm–oocyte fusion, there are significant doubts with regards to what functional role this particular population of PLCZ1 might play. Furthermore, reports have demonstrated a potential shift in PLCZ1 localization to the post-acrosomal region following capacitation (Grasa et al. 2008, Young et al. 2009). Interestingly, another study put forward the hypothesis that a truncated isoform of PLCZ1, inexplicably termed ‘NYD-SP7’ was localized within the acrosomal regions of murine and human sperm and served as a decapacitation factor (Bi et al. 2009). However, confusion remains regarding the precise nature and intention of this study since NYD-SP7 lacked the inherent domain structure of the PLCZ1 protein, thus rendering it functionally inactive. Some researchers have reported the localization of PLCZ1 to the sperm tail. It remains unknown whether these findings are legitimate or merely represent a technical flaw related to immunological detection technology. Examination of the current literature clearly shows that various different PLCZ1 localization patterns have been reported in sperm, although most researchers seem to agree that the most valid and justifiable localization pattern of PLCZ1 is in the equatorial region. However, it is important that future studies identify the specific reasons for potential PLCZ1 localization in the sperm tail. A previous study by Bedford-Guaus et al. (2011) achieved calcium responses in oocytes by simply injecting the tails of equine sperm. Thus, we cannot omit the possibility that PLCZ1 localized in the tail may exert functionality in terms of activation or in downstream processes. However, Kashir et al. claimed that the localization of PLCZ1 in the sperm tail is simply an artefact; these authors noted that antibody specificity remains a notable problem and that we should disregard the localization of PLCZ1 in the sperm tail (Kashir et al. 2020). Researchers have yet to consider the potential function of PLCZ1 populations in the sperm tail; further research is needed to address this possibility, especially in human sperm (Bedford-Guaus et al. 2011).
Figure 1.

Representative immunofluorescence images of PLCZ1 localization in sperm from a fertile male. Images were captured as (A) bright-field or stained with (B) DAPI and (C) FITC-PLCZ1. Merged images are shown in (D). PLCZ1 is expressed in the equatorial (yellow arrows), acrosomal (white arrow), and postacrosomal (asterisk) regions in the sperm. Scale bar = 5 μm. Reproduced with permission from Meng et al. (2020).
The most demanding conundrum in PLCZ1 research is the significant variability of the expression levels and localization patterns of PLCZ1 that has been described in both fertile donors and infertile patients (Grasa et al. 2008, Kashir et al. 2013). Studies have reported significant variation in both the expression level and localization pattern of PLCZ1 within the same ejaculate and when compared between different donors and patients (Yoon et al. 2008, Heytens et al. 2009, Kashir et al. 2011a, b, 2012a, Escoffier et al. 2015a, Ferrer-Vaquer et al. 2016, Javadian-Elyaderani et al. 2016, Dai et al. 2020, Rahimizadeh et al. 2020). These variations in PLCZ1 content between the ejaculates of fertile and infertile men raise significant questions as to why such disparity occurs. One explanation could be that levels of PLCZ1 are reduced during spermatid elongation; however, further investigations are needed to specifically determine the localizations patterns of PLCZ1 within the testes, particularly in humans.
Previous immunofluorescent analysis of single sperm, from both fertile and infertile men, also showed wide variation in two key parameters of interest: total levels and localization patterns of PLCZ1 (Grasa et al. 2008, Kashir et al. 2013). However, despite the fact that some fertile controls can exhibit similar levels of PLCZ1 expression to infertile patients, immunofluorescence analyses have clearly confirmed that the relative fluorescence (RF) intensity of PLCZ1 in human sperm is significantly lower in OAD patients than in fertile males (Heytens et al. 2009, Kashir et al. 2013), thus providing us with the capability to perform diagnostic assays.
Other researchers have reported the inability of patients with globozoospermia, a condition in which the sperm are round and lack the acrosome, to evoke long-term Ca2+ oscillations following the injection of their sperm into mouse oocytes. Previous studies have shown that PLCZ1 is localized in the perinuclear theca of the equatorial and post-acrosomal region of human sperm (Escoffier et al. 2015b). Patients with globozoospermia have absent or abnormal levels of PLCZ1 in their sperm and show punctuated localization patterns (Heytens et al. 2009, Kashir et al. 2012b). This is not surprising as an individual with DPY19L2-dependent globozoospermia was found to produce defective sperm featuring loss of the acrosome and surrounding materials including PLCZ1 (Escoffier et al. 2015b). Subsequent studies showed that patients diagnosed with partial or complete globozoospermia are unable to fertilize oocytes naturally and that this deficiency is related to reduced levels or the total absence of PLCZ1 (Taylor et al. 2010, Abadi et al. 2016, Tavalaee & Nasr-Esfahani 2016, Tavalaee et al. 2018, Cheung et al. 2020).
Motile sperm organelle morphology examination (MSOME) has been used to study the specific morphology of globozoospermic sperm and successfully identify a sub-population of sperm exhibiting a small acrosomal bud (Sermondade et al. 2011). Subsequent ICSI cycles, involving the injection of globozoospermic sperm featuring these small acrosomal buds, permitted globozoospermic patients to achieve live birth without the use of artificial oocyte activators (AOAs) (Sermondade et al. 2011). Given these interesting findings, Kashir et al. identified that almost half (43%) of the sperm exhibiting acrosomal bud morphology expressed PLCZ1 within their head regions (Fig. 2) (Kashir et al. 2012b), albeit with highly abnormal localization patterns. Thus, the use of MSOME to identify acrosomal buds in the sperm heads of globozoospemic patients can provide a useful tool with which to guide clinical treatment by allowing the selection of sperm containing PLCZ1 from populations of sperm that do not contain PLCZ1.
Figure 2.
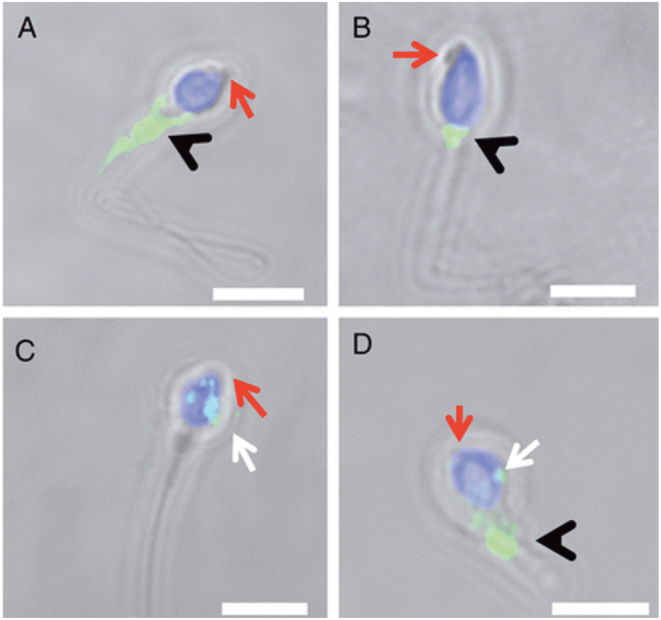
Representative confocal images of PLCZ1 immunofluorescence in motile sperm organelle morphology examination (MSOME)-selected globozoospermic sperm exhibiting an acrosomal bud (red arrow). PLCZ1 (pale green staining) was localized to the midpiece (black arrowheads, A and B), or as a punctate pattern in the sperm head (white arrows, C), or in combination (D). White scale bar represents 5 µm. Reproduced with permission from Kashir et al. (2012b).
The procedure used for semen analysis comprises basic macroscopic and microscopic diagnostic tests, including volume, concentration, motility, and viability; these tests are performed by trained clinical scientists in ART laboratories. However, tests relating to the functional aspects of the sperm, such as DNA fragmentation, mitochondrial function, and reactive oxygen species, require the use of more advanced techniques which are often not available in ART units (Agarwal et al. 2021). At present, the specific causes of male factor infertility or subfertility remain poorly understood (Agarwal et al. 2021). Furthermore, human sperm are far from being homogeneous and only a small proportion of sperm are considered as being ‘normal’ in a given semen sample. In ART laboratories, clinical scientists select, when possible, the best sperm to use for ICSI; however, the selection process is still very subjective and, in some cases, normozoopermic sperm are not available. We know that PLCZ1 is expressed at the early stage of spermatogenesis in the testis; thus, events that damage or alter spermatogenesis may contribute to variations of PLCZ1 expression in subsequent stages. With this in mind, studies have assessed the correlation between PLCZ1 expression and localization and sperm integrity. Researchers have reported different conclusions in terms of the correlation between PLCZ1 and DNA oxidation status. One group reported that higher levels of oxidative stress were correlated with lower PLCZ1 expression levels; however, other groups found no evidence of such a correlation (Park et al. 2015, Janghorban-Laricheh et al. 2016, Tavalaee et al. 2017). Some authors argued that this discrepancy may be due to the assays used. Furthermore, low semen quality, such as in patients with varicocele (a chronic genital heat stress condition), has been shown to have low levels of PLCZ1. It has been shown that PLCZ1 is heat-sensitive; therefore, this condition may disrupt the final translation of the protein (Sanusi et al. 2015). Accordingly, sperm from infertile males with teratozoospermia (abnormal head morphology) have previously been demonstrated to exhibit abnormal localization or the absence of PLCZ1 but showed a variance in PLCZ1 expression in comparison to controls (Azad et al. 2018). However, in another report, involving patients with asthenoteratozoospermia, in which motility and morphology are abnormal, showed no difference in PLCZ1 localization (Rahimizadeh et al. 2020). Other authors reported a correlation between sperm health and PLCZ1; however, the authors were not able to identify a specific correlation between morphology and PLCZ1 (Kashir et al. 2020). Additional studies, with larger cohorts, are now needed to investigate PLCZ1 with respect to abnormal and normal sperm morphology. We know that in the case of TFF, sperm are unable to induce Ca2+ oscillations. Furthermore, studies have shown that within an ejaculate, human sperm are able to cause different patterns of Ca2+ spikes in oocytes, thus reinforcing the idea that each sperm may contain a different amount of PLCZ1 or that the level of PLCZ1 activity in each sperm is variable (Ferrer-Buitrago et al. 2018). In addition, we know that PLCZ1 activity differs across species; studies have shown that a certain amount of PLCζ is required for the oocyte to achieve successful embryo development and that this amount varies across different species (Yoon et al. 2008, Nomikos et al. 2013b). This observation concurs with other studies reporting variability in the levels and localization patterns of PLCZ1 in fertile and infertile men. This raises the question as to why sperm have different levels, localization patterns, and functional activities.
Most notably, PLCZ1 levels, localization patterns, and the proportion of human sperm exhibiting PLCZ1, have been found to correlate significantly with fertilization rates following ICSI, thus indicating that PLCZ1 is a promising biomarker for diagnosing OAD and predicting the outcome of ICSI (Yelumalai et al. 2015, Nazarian et al. 2019). However, this concept has been questioned since another study concluded that PLCZ1 levels and localization patterns were not correlated with ICSI outcome (Ferrer-Vaquer et al. 2016). Similar to findings in humans, a recent study, conducted in stallions, reported that the injection of sperm with higher PLCZ1 protein levels into bovine oocytes led to a better embryonic cleavage rate (Amoroso-Sanches et al. 2019). Collectively, the existing body of evidence clearly indicates that PLCZ1 is a practical clinical biomarker that can be used to indirectly predict the oocyte activation ability of a given sperm sample. However, immunocytochemical staining alone is not sufficient to fully determine the PLCZ1 status of sperm. Screening of the gene encoding the PLCZ1 protein is also critical if we are able to fully determine the extent to which genetic factors might play a role in the aberrant expression and/or localization patterns observed in infertile patients.
PLCZ1 gene abrogation, recurrent inheritance mutations, and protein dysfunction
The human PLCZ1 gene consists of 15 exons and is located on the reverse strand of chromosome 12. This gene encodes a PLCZ1 protein that is composed of a series of key functional domains (EF, X, Y, and C2) with a linker region that connects the catalytic X and Y domains. An obvious hypothesis is that PLCZ1 deficiency is linked to genetic alterations in the PLCZ1 gene or the molecular mechanisms that govern the expression of this gene. Such alterations could readily manifest in a dysfunctional protein and exert a direct impact on oocyte activation capability. Consequently, significant efforts have been made to study PLCZ1 at the genomic level. Critically, several mutations have been detected in the human PLCZ1 gene; some of these mutations have also been demonstrated to impair the functionality of the translated protein (Table 1) (Heytens et al. 2009, Kashir et al. 2011c, Escoffier et al. 2015a, Ferrer-Vaquer et al. 2016, Torra-Massana et al. 2019, Kashir 2020). Several of these mutations have been modelled in the laboratory to confirm whether the functionality of the translated protein was impaired under normal physiological conditions.
Table 1.
Literature-based summary of PLCZ1 mutations identified in infertile males with/without follow-up functional investigation. Reproduced from Kashir (2020) with permission.
| Mutation | Domain affected | In vitro phenotype | In vivo phenotype | References |
|---|---|---|---|---|
| I120M | EF-X linker | Predicted alteration of local protein folding | OAD; low fertilization success | Torra-Massana et al. (2019) |
| C196X | X | Predicted alteration of local protein folding | OAD; reduced/absent PLCZ1 in patient sperm; abnormal PLCZ1 localization | Dai et al. (2020) |
| C196* | X | No recombinant protein produced by mammalian cells; reduced activation success following cRNA injection in mouse oocytes | OAD; reduced/absent PLCZ1 in patient sperm; low fertilization success | Mu et al. (2020), Yan et al. (2020) |
| R197H | X | Predicted alteration of local protein folding | OAD; low fertilization success | Ferrer-Vaquer et al. (2016), Torra-Massana et al. (2019) |
| L224P | X | Predicted alteration of local protein folding | OAD; low fertilization success | Torra-Massana et al. (2019) |
| H233L | X | Reduced expression in mammalian cells; reduced/absent oscillations following cRNA injections in mouse oocytes; reduced embryogenesis in mouse; predicted alteration of local protein folding | OAD; reduced/absent PLCZ1 in patient sperm; abnormal PLCZ1 localization; low fertilization success | Kashir et al. (2011b,b, 2012a), Ferrer-Vaquer et al. (2016), Torra-Massana et al. (2019) |
| L246F | X | Predicted alteration of local protein folding | OAD; reduced/absent PLCZ1 in patient sperm; abnormal PLCZ1 localization | Dai et al. (2020) |
| L277P | X | Predicted alteration of local protein fold; reduced activation success following cRNA injection in human oocytes | OAD; reduced/absent PLCZ1 in patient sperm; low fertilization success | Yan et al. (2020) |
| T324fs | X-Y linker | Truncated recombinant protein produced by mammalian cells; reduced activation success following cRNA injection in mouse oocytes | OAD; low fertilization success | Mu et al. (2020) |
| V326K fs*25 | X-Y linker | Predicted frameshift truncation of protein | OAD; low fertilization success | Torra-Massana et al. (2019) |
| S350P | Y | Predicted alteration of local protein folding | OAD; reduced/absent PLCZ1 in patient sperm; abnormal PLCZ1 localization | Dai et al. (2020) |
| N377del | Y | Predicted alteration of local protein fold; no activation success following cRNA injection in human oocytes | OAD; reduced/absent PLCZ1 in patient sperm; low fertilization success | Yan et al. (2020) |
| A384V | Y | Predicted alteration of local protein fold; n +C16o activation success following cRNA injection in human oocytes | OAD; reduced/absent PLCZ1 in patient sperm; low fertilization success | Yan et al. (2020) |
| H398P | Y | Reduced expression in mammalian cells; reduced/absent oscillations following cRNA injections in mouse oocytes; predicted alteration of local protein folding | OAD; reduced/absent PLCZ1 in patient sperm; abnormal PLCZ1 localization | Heytens et al. (2009), Kashir et al. (2011b,b, 2012a) |
| R412fs | Y | Truncated recombinant protein produced by mammalian cells; reduced activation success following cRNA injection in mouse oocytes | OAD; low fertilization success | Mu et al. (2020) |
| P420L | Y | Reduced recombinant protein produced by mammalian cells; reduced activation success following cRNA injection in mouse oocytes | OAD; low fertilization success | Mu et al. (2020), Yuan et al. (2020b) |
| K448N | Y | Predicted alteration of local protein fold; reduced activation success following cRNA injection in human oocytes | OAD; reduced/absent PLCZ1 in patient sperm; low fertilization success | Yan et al. (2020) |
| I489F | C2 | Reduced/absent oscillations following cRNA injections in mouse oocytes; reduced embryogenesis in mouse; predicted alteration of local protein fold; similar enzymatic properties but dramatically reduced substrate binding | OAD; reduced/absent PLCZ1 in patient sperm; abnormal PLCZ1 localization | Escoffier et al. (2015a), Nomikos et al. (2017) |
| S500L | C2 | Predicted alteration of local protein folding | OAD; low fertilization success | Ferrer-Vaquer et al. (2016), Torra-Massana et al. (2019) |
| R553P | C2 | Reduced/absent fertilization following cRNA injections in mouse oocytes; predicted alteration of local protein fold; mouse fertilization and embryogenesis comparable following injection of higher levels of mutant cRNA | Comparable levels of PLCZ1 in patient sperm | Yuan et al. (2020a) |
| L576P | C2 | Predicted to affect C2 domain structure or C2-catalytic domain interaction | Low fertilization success; live-birth without AOA in the third cycle | Yuan et al. (2020b) |
| M578T | After C2 | Predicted alteration of local protein fold; No activation success following cRNA injection in human oocytes | OAD; reduced/absent PLCZ1 in patient sperm; low fertilization success | Yan et al. (2020), Yuan et al. (2020b) |
AOA, artificial oocyte activation; OAD, oocyte activation deficiency; PLCZ1, phospholipase C zeta.
Heytens et al. were the first to identify a mutation in the PLCZ1 gene. These authors screened the DNA of a normozoospermic male with OAD and identified a substitution mutation (H398P) that resulted in a single amino acid change in the final protein; this mutation exerted adverse effects upon the functional capability of the protein (Heytens et al. 2009). At this time, the authors used three-dimensional (3D) structural modelling to show that the substitution mutation destabilized the protein structure and interfered with its ability to bind to substrate. Further investigation demonstrated that this mutation abolished the hydrolytic activity of the PLCZ1 protein and a general failure to elicit Ca2+ oscillations in the ooplasm (Nomikos et al. 2011). Subsequently, a second mutation (H233L) was discovered in the same patient. Additional inheritance analysis showed that the H398P and H233L mutations were located on different alleles and that one mutation (H233L) was inherited via the maternal lineage (Kashir et al. 2011c). Subsequent genotyping of single sperm from this infertile patient revealed that all of his sperm exhibited either the H398P or H233L mutation but never both, thus confirming that the two mutations were heterozygous. These findings indicated, for the first time, that PLCZ1-linked infertility can be inherited via the maternal lineage (Kashir et al. 2011c). In addition, the transfection of both mutations into human embryonic kidney cells (HEK293T) via plasmid DNA showed that the fluorescence level of recombinant mutant protein was significantly lower when a single mutation was transfected, thus indicating that a combination of these mutations may influence the stability of the protein more than a single mutation (Kashir et al. 2012a).
Subsequent research identified a homozygous mutation (I489F) in two brothers with normozoospermia who experienced TFF following ICSI and indicated that another brother (who was fertile) possessed the same mutation but in a heterozygous state. In addition, 3D modelling and liposome-binding assays showed that this mutation caused an amino acid substitution that led to a significant alteration in enzymic activity (Escoffier et al. 2015a, Nomikos et al. 2017).
Since then, two novel heterozygous mutations and a previously reported mutation have been identified in PLCZ1 (S500l, R197H, & H233L) in patients with failed fertilization; the researchers involved comments that heterozygote mutations may only reduce the functionality of the protein, as reported previously. However, no functional analyses were performed to investigate whether these mutations could specifically impair PLCZ1 functionality (Ferrer-Vaquer et al. 2016). More recently, Torra-Massana et al. reported not only the further detection of R197H, H233L, and S500L but also discovered three newly identified mutations (I120M, L224P, and V326K fs*25, a frameshift mutation) (Torra-Massana et al. 2019). Further bioinformatics analysis (involving Phyre2 software) suggested that the L224P and V326K fs*25 mutations exert detrimental effects on PLCζ protein function (Torra-Massana et al. 2019). These authors subsequently microinjected cRNA of their corresponding mutations into in vitro-matured human oocytes and found that the R197H and H233L mutations resulted in low activation rates (30 and 30.77%, respectively). In contrast, the L224P and S500L mutations had no influence on oocyte activation rates (64.29 and 60%, respectively). These findings highlighted the fact that not all genetic variations are detrimental to the functionality of PLCZ1 protein. Furthermore, the V326K fs*25 mutation was located in the XY-linker region, an important region that regulates enzyme activity; this mutation was caused by the deletion of two nucleotides (A and G), thus resulting in a frameshift and a premature stop codon downstream of the mutation. This was the first PLCZ1 mutation to be identified in the XY-linker region. Bioinformatics analysis further indicated that the V326K fs*25 mutation would inevitably result in the translation of a truncated form of PLCZ1 protein. In line with these predictions, the injection of a truncated form of PLCZ1 cRNA into human invitro-matured oocytes resulted in TFF (0/10), thus demonstrating that this particular mutation can exert a devastating effect on protein function (Torra-Massana et al. 2019).
More recently, Dai et al. identified a novel nonsense mutation in PLCZ1 (C196X, the substitution of cysteine with an undetermined amino acid) and two novel missense mutations (S350Pand L246F) (Dai et al. 2020). These mutations were identified from three infertile males who experienced low fertilization rates (0–7.7%). Further 3D modelling suggested that the C196X mutation resulted in a premature termination codon that may impair protein function. In line with this, the infertile patient who exhibited the C196X mutation showed no PLCZ1 expression in his sperm. The other two mutations, S350P and L246F, also showed hydrogenic changes in the secondary structure of PLCZ1 protein. Interestingly, PLCZ1 was found to be expressed in sperm from patients exhibiting these two mutations. Unfortunately, the authors did not ascertain whether PLCZ1 expression levels were also reduced in these mutant sperm. Nevertheless, all three patients received AOA treatment in subsequent ICSI cycles and achieved significantly improved fertilization rates (from 4.4 to 56%), resulting in two successful ongoing pregnancies.
Research by Mu et al. further identified five mutations in four different families: C196*, R197H, P420L, R412fs, and T324fs). Interestingly, one mutation was recurrent in two families (C196*) and another (T324fs) was located within the XY linker. So far, only one other mutation has been identified in this critical part of the gene (Mu et al. 2020). Mu et al. compared the level of expression of each of these mutations in vitro and noted that all five mutations led to a significantly reduced level, or the total absence, of the final protein. Furthermore, reduced pronuclei formation was observed when the corresponding cRNAs were injected into mouse oocytes, thus suggesting that these mutations affected the functionality of PLCZ1 (Mu et al. 2020). All four families achieved successful fertilization after AOA treatment. Another study reported one known mutation and five novel mutations (C196*, M578T, A384V, N377del, L277P, and K448N); two families presented with the C196* mutation (Yan et al. 2020). Injection of cRNAs corresponding to the L277P and K448N mutations led to reduced pronuclear formation (27 and 11%, respectively) upon oocyte injection in comparison to the WT (86%). However, injection of cRNAs corresponding to M578T, A384V, and N377del failed to induce pronuclear formation. It is important to note that two of these mutations were homozygous, while the other three were heterozygous, thus re-affirming the autosomal recessive inheritance of PLCZ1. In this study, 14 patients were recruited by virtue of their low fertilization rate or due to TFF and the detection of sperm defects by the mouse activation assay (MOAT) (Yan et al. 2020). Only five patients were chosen for subsequent AOA treatment because of their mutations and the lack of protein PLCZ1 expression, as determined by Western blotting. Four patients opted for AOA treatment. Unfortunately, this study failed to report the expression levels and localization patterns of PLCZ1 for each patient (Yan et al. 2020). Also, genetic screening has assisted in the diagnosis and treatment of infertility detected in five cases, further investigations are now needed to elucidate the cause of infertility in the other nine patients (Yuan et al. 2020a). Furthermore, studies have shown that patients with heterozygous mutations presented with fertilization failure or were infertile. However, other reports have shown that the fathers of these patients were fertile; therefore, such inheritance may not necessarily affect fertility (Torra-Massana et al. 2019).
More recently, another mutation was discovered in a patient with normal semen parameters (R553P) (Yuan et al. 2020a). Western blotting analysis revealed the presence of PLCZ1 protein in the patient’s sperm; however, TFF was observed after ICSI in 50 oocytes. Further analysis revealed that the mutation was likely to disrupt protein folding and thus impact on protein functionality. The authors reported that the female patient had achieved a successful pregnancy with a previous partner, thus reinforcing the link between PLCZ1 deficiency and fertilization failure. The latest mutation that we would like to report in this review by Yuan et al. has extended our knowledge relating to failed fertilization and autosomal recessive inheritance patterns, particularly with regards to their discovery of three novel mutations in two different males (Yuan et al. 2020b). Indeed, in this study, DNA sequencing of one patient experiencing TFF showed that the patient had inherited the M578T mutation from his father and P420L from his mother (Yuan et al. 2020b). This study provided further evidence that PLCZ1 mutations can be inherited from the maternal lineage and that more attention is needed to focus on the inheritance of subfertility. The other patient with low fertilization capacity possessed the L576P mutation; unfortunately, no other genetic data were acquired for this particular patient. Interestingly, none of these mutations exerted impact on the transcriptional expression of PLCZ1 (Yuan et al. 2020b). The authors assumed that the homozygous L576P mutation may result in a partial loss of PLCZ1 function as the patient was subsequently able to conceive without AOA treatment. However, the authors postulated that the heterozygous mutations (M578T and P420L) may lead to TFF due to their presence within the catalytic domain of the protein. Collectively, these data show that many mutations have been identified in the human PLCZ1 gene. Some of these are known to exert a more deleterious effect on protein function than others. The relative effects of these mutations appear to depend on their specific location within the PLCZ1 gene and their specific mode of inheritance. However, although genetic screening strategies have clearly identified patients who possess PLCZ1 mutations and that some of these mutations can impair the functionality of the PLCZ1 protein, it is highly evident that such mutations only manifest in a very small population of patients who exhibit OAD. Consequently, genetic mutations do not appear to be the main driver of PLCZ1 deficiency in the population of patients who experience fertilization failure. This hypothesis has been confirmed by our own clinical studies, carried out in collaboration with Oxford Fertility and Ninewells Hospital in Dundee (unpublished data). Despite screening over 220 patients experiencing ICSI failure, only a very small number of these patients were shown to possess PLCZ1 mutations. Of the patients screened by our diagnostic assay thus far, 37% had low levels of PLCZ1 or abnormal localization patterns, only 3% had mutations, 5% had sperm that were completely devoid of PLCZ1; however, the remainder (55%) had no PLCZ1 issues, at least using our current assays (unpublished data). However, a substantial proportion of the patients screened by our ongoing research programme were shown to exhibit clear deficiency in terms of PLCZ1 levels. These findings appear to suggest that PLCZ1 deficiency is more likely to be associated with undetermined mechanisms related to gene expression or repression, rather than mutation per se. At present, we do not know exactly why this is the case, although it is tempting to suspect that there are several mechanisms that can affect gene expression at the transcriptional and/or translational levels. For example, only a few studies have investigated the bidirectional promoter of PLCZ1; only one variant has been identified in the CAPZA3 promoter but only in the vicinity of PLCZ1 (Javadian-Elyaderani et al. 2016); the authors suspected that this variant may not affect the transcription of PLCZ1. Further studies are needed to fully elucidate the precise role of the bidirectional promoter with regards to the levels of gene transcription.
Indeed, multiple molecules are involved in gene expression at the transcriptional and translational levels, including RNA polymerase, enhancers, silencers, and RNA molecules, including coding and non-coding RNAs (Khalil et al. 2009). It would be therefore very interesting if future studies investigate whether abnormalities in these factors can impair PLCZ1 expression, which eventually contributes to the dysfunction of the PLCZ1 protein and male infertility.
PLCZ1 assays and the urgency to develop specific guidelines to justify the use of AOAs
Despite the beneficial effects of therapeutic approaches involving AOAs in patients that experience ICSI failure, there is an understandable and ongoing debate relating to the safety profile of these synthetic agents. Indeed, it is well-known that AOAs induce a single, prolonged Ca2+ transient instead of the normal oscillatory patterns. Currently, the HFEA recognizes that the use of AOAs may lead to aneuploidy during embryogenesis and has thus recommended that this treatment should be limited to infertile patients with fertilization failure and only where significant male factor has been identified (HFEA 2017). Indeed, AOA may not be the best treatment for couples with OAD (Vanden Meerschaut et al. 2012, Van Blerkom et al. 2015). Consequently, the use of AOAs should not be considered as a routine procedure in ART. Indeed, the HFEA currently considers AOAs as a treatment add-on and has placed AOA treatment on their ‘Amber’ list, thus implying that there is insufficient evidence relating to the efficiency and safety of AOAs (HFEA 2017). Only a small proportion of infertility clinics perform AOA; those that do only consider treatment according to a history of fertilization failure after ICSI or the suspicion of OAD (Tesarik et al. 2002, Montag et al. 2012, Nicopoullos et al. 2015). Van Blerkom et al. expressed significant concern over the fact that AOA treatment might become more frequently used for ICSI cycles to ensure appropriate pronuclear formation and to eliminate the potential for fertilization failure (Van Blerkom et al. 2015). AOA is recommended only in cases of previously failed fertilization, for couples where there is a possible defect of the sperm or the oocyte and where all other options have been exhausted. It is important that clinicians take into consideration previous cycles and patient histories before justifying the use of AOA. Furthermore, it is important that patients are aware of the procedure, and what it involves, as there are still too many unknowns associated with AOA, especially in cases where there is no specific justification. In the UK, it is a requirement to justify such treatment; however, this might not be the case in other countries where more flexibility is permitted. Van Blerkom and colleagues raised the concern that AOA could be used negligibly in ICSI procedures in non-justified cases but to eliminate the possibility of fertilization failure, thus improving success rate. AOA should only be advised to specific cases and should not be used routinely. There is a paucity of data to support the universal recommendation of AOA in clinical practice; the HFEA states that randomized, controlled trials and follow-up studies are required to specifically address the clinical use of AOAs. Given these concerns, it is imperative to establish standard guidelines for the clinical use of AOAs.
Several methods have been used to investigate patients who have experienced recurrent ICSI failure or to evaluate the ability of sperm to activate oocytes, including the heterologous injection of human sperm into mouse oocytes (MOAT) (Heindryckx et al. 2005), the measurement of calcium changes in human oocytes upon the injection of patient sperm (the human oocyte calcium assay) (Ferrer-Buitrago et al. 2018), or the assessment of centrosomal function in sperm (Terada et al. 2009). Electrical charges upon the sperm membrane have also been used to select sperm that contain PLCZ1; this test was considered to facilitate successful fertilization (Chan et al. 2006, Khakpour et al. 2019). However, only a limited number of clinics have introduced these approaches and a major concern is the inability of fertility units to recreate these tests as they involve specialist equipment or resources that are acquired from animal models (Heindryckx et al. 2005, 2008, Vanden Meerschaut et al. 2013). Furthermore, we know that the potency of PLCZ1 differs between the mouse and human. Therefore, it is important that essays take this disparity into consideration when developing assays that can be used in real clinical settings.
Over the years, PLCZ1 expression has been determined by measuring the levels of PLCZ1 mRNA and RNA (Aghajanpour et al. 2011, Kashir et al. 2013, Abadi et al. 2016, Khakpour et al. 2019) or by quantifying the levels of PLCZ1 protein in sperm by immunoblotting (Yan et al. 2020, Yuan et al. 2020b), cell sorting (Khakpour et al. 2019), and immunofluorescence analysis (Kashir et al. 2013). In addition, other forms of dysfunction can also lead to OAD, such as DNA fragmentation in the sperm head, inadequate oocyte maturation, low oocyte numbers, or even technical problems encountered during ART. Sperm analysis remains pivotal in the evaluation of male factor infertility in clinic, and additional laboratory tests have also been proposed to patients seeking fertility treatment, including DNA integrity; thus, it is reasonable to see that researchers have attempted to correlate PLCZ1 levels, DNA fragmentation, and oxidation. However, the findings of such studies are very limited and further investigations are required before drawing firm conclusions (Taylor et al. 2010, Park et al. 2015, Tavalaee et al. 2017). Other studies have reported that the localization and total levels of PLCZ1 are not correlated with male age in humans although a recent study has reported otherwise in several mouse strains (Yeste et al. 2016b, Kashir et al. 2021). As discussed previously, it is clear that PLCZ1 is an important physiological factor that triggers oocyte activation. However, over recent years, it has become debatable whether PLCZ1 is the sole factor required for fertilization (Hachem et al. 2017, Nozawa et al. 2018, Swann 2020). Indeed, two separate studies have shown that Plcz1-KO mice were not sterile. Interestingly, subsequent ICSI in both studies using mutant sperm into mouse oocytes neither trigger Ca2+ oscillations nor oocyte activation; however, when Plcz1 KO mice were mated with WT female mice, both obtained few, but viable, pups (Hachem et al. 2017, Nozawa et al. 2018). A recent review hypothesized that a ‘sperm cryptic-activating factor (SCAF)’ is responsible for initiating Ca2+ oscillations only in the absence of PLCZ1 (Jones 2018). However, what exactly SCAF is and how it activates an oocyte remain unknown. Despite conflicting debates, the overall evidence suggests that PLCZ1 is the dominant physiological trigger of normal Ca2+ oscillations, especially in monospermic fertilization.
It is clear that PLCZ1 has the potential to provide useful evidence for the justification of AOA in the clinical setting as required by the HFEA and in addition to basic semen analyses. Quantitative analysis of PLCZ1 immunofluorescence can provide an informative biomarker to predict the oocyte activation ability of sperm from a given patient (Kashir et al. 2013). Over the years, some researchers investigating PLCZ1 localization and expression in sperm have been challenged by widespread scepticism due to the polyclonal nature of the antibodies they were using and the apparent variability in the data generated by such assays. However, routine laboratory assays have successfully been developed and validated by peptide-blocking experiments, thus proving the efficacy of the protocols applied (Grasa et al. 2008, Heytens et al. 2009, Kashir et al. 2011a, 2013, Yelumalai et al. 2015, Yeste et al. 2016b). In a recent study, Kashir et al. proposed that inadequate specificity between the PLCZ1 antibody and its antigen hindered the visualization efficacy during PLCZ1 immunoanalysis and advocated the use of an antigen unmasking technique when carrying out such assays (Kashir et al. 2017). However, detailed experiments in our laboratory found no difference between our routine in-house assay with regards to the immunofluorescent detection of PLCZ1 when carried out with and without the antigen masking technique (Meng et al. 2022).
Immunofluorescent assays of the expression levels and localization patterns of PLCZ1 have indicated significant variation among both fertile and infertile patients. Of most concern was the fact that some infertile patients clearly exhibited lower levels of PLCZ1 than fertile controls (Meng et al. 2020). This indicates that the establishment of a simple ‘threshold or cut-off’ value for PLCZ1 levels for diagnostic purposes will be non-viable. Interestingly, we developed a more robust bioassay that permits the classification of ‘PLCZ1 deficiency’ by incorporating the combination of mean PLCZ1 level and the specific proportion of sperm in a given patient sample exhibiting PLCZ1 (Meng et al. 2020). In our study, we demonstrated that the combination of RF PLCZ1 levels and the percentage of sperm containing PLCZ1 can facilitate the selection of suitable patients for AOA treatment. In our study, we found that infertile men presenting with less than 71% of their sperm containing PLCZ1 and with a mean PLCZ1 level of 15.57 arbitrary are most likely to benefit from AOA treatment (Fig. 3). In this study, we also described the importance of using this specific threshold to diagnose patients with ‘normal’ or ‘abnormal’ sperm parameters to determine their eligibility for AOA treatment. As discussed extensively in this review, both fertile and infertile men are known to exhibit variable PLCZ1 levels; therefore, is it important that we consider both levels and the proportion of sperm containing the PLCZ1 protein before making a clinical decision. Furthermore, we found that 80% of patients exhibiting a significant ‘PLCZ1 deficiency’ (a low expression level and a low proportion of sperm exhibiting PLCZ1) who opted for AOA treatment achieved significant improvements in their fertilization rate (approximately 40% higher) and pregnancy/live birth rates (both 40% per initiated cycle), thus demonstrating that this more robust and reproducible PLCZ1 assay was more efficient with regards to diagnosing PLCZ1 deficiency (Meng et al. 2020). However, additional studies are now needed to help patients with a low sperm count. For example, this essay is currently limited to patients who produce a sufficient number of sperm for quantification. By using the algorithm, our laboratory-based in-house PLCZ1 assay can evaluate PLCZ1 expression in infertile males, therefore identifying whether the state of infertility is male-related (Fig. 4). This allows the development of an individually targeted strategy for the management of AOA, which can save effort, time, and cost for both IVF clinics and patients. Collectively, these findings highlight the clinical potential of PLCZ1, both as a prognostic indicator of OAD and eventually as a therapeutic protocol. While we pursue further research, it is important that we educate patients, via online forums, webinars, or awareness events, so that they fully understand the relationship between OAD and PLCZ1 and our current ability to diagnose PLCZ1 deficiency but are also aware of our current diagnostic and therapeutic limitations and the lack of evidence-based data relating to the widespread application of AOAs.
Figure 3.
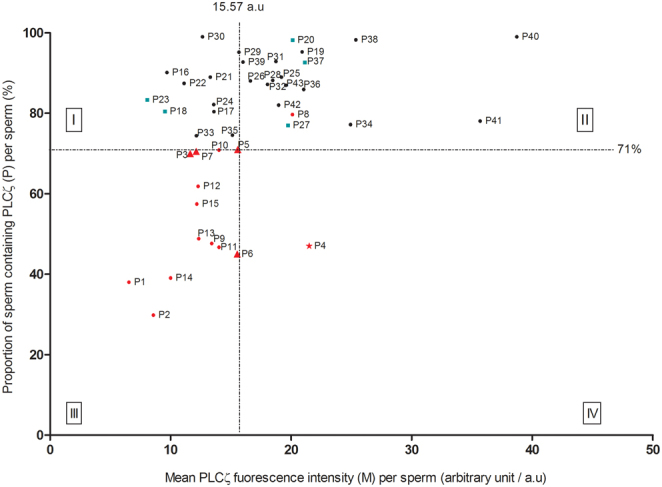
Artificial oocyte activation outcomes associated with two PLCZ1 parameters: mean PLCZ1 level in sperm (x-axis) and the proportion of sperm containing PLCZ1 (y-axis). ‘‘I, II, III, IV’ represent the four categories assigned by the two cutoffs. Patients with PLCZ1 deficiency are labelled in red (the red triangle: history with live birth with artificial oocyte activation (AOA); red dots: patients did not receive AOA; red star: patients received AOA but only chemical pregnancy). Patients who have had live births or ongoing pregnancy without AOA were labelled as green cubes, and patients with unknown outcomes of subsequent in vitro fertilization/intracytoplasmic sperm injection treatment were shown as black dots. For further details, see Meng et al. (2020). Reproduced with permission.
Figure 4.

The suggested clinical algorithm for assessing the eligibility of a patient for artificial oocyte activation. AOA, artificial oocyte activation; ICSI, intracytoplasmic sperm injection. Reproduced with permission from Meng et al. (2020).
Conclusion
Following the discovery of PLCZ1, a substantial body of evidence has confirmed the indispensable role of PLCZ1 in oocyte activation and clearly demonstrated that the expression and localization of PLCZ1 in human sperm are associated with male infertility. Infertile males show lower PLCZ1 levels in their sperm than fertile males. In addition, PLCZ1 was found to be predominantly expressed in the equatorial region of the sperm but sometimes in combination with other regions. Immunofluorescence staining is currently used to assess PLCZ1 expression and localizations in most studies, and the implementation of a robust and specific bioassay is a key requirement for future research and the evaluation of PLCZ1 levels and localization patterns to help with the justification of artificial oocyte activators in the clinic. Moreover, genetic testing of PLCZ1 can lead to the discovery of novel mutations that affect the expression/potency of the protein and therefore, oocyte activation ability. For this reason, we now need to recruit a larger cohort of fertile and infertile patients for genetic screenings so that we can gain a more comprehensive understanding of gene variations in terms of fertility status.
Declaration of interest
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of this review.
Funding
The PLCZ1 research in our laboratory has been funded by project grants from the Royal Society (UK) and the Oxford University Medical Research Fund awarded to K Coward, the Saint Glee International Foundation (awarded to X Meng), and a European Commission, FP7-People Programme, Marie Curie-IEF (Grant Number: 626061; awarded to M Yeste), and the Nuffield Department of Women’s & Reproductive Health. Some of our PLCZ1 research has been funded by student scholarships awarded by the Governments of Malaysia and Brunei.
Author contribution statement
C J and X M drafted the manuscript. K C revised the manuscript. All authors approved the final version for publication.
Acknowledgements
The authors would like to thank Prof. Marc Yeste, Dr Junaid Kashir, Dr Walaa Ramadan, Dr Suseela Yelumalai, Dr Siti Nornadhirah Amdani, and Dr Sally Galal for their contributions to the PLCζ research carried out in our laboratory. The authors would also like to thank Prof. Chris Barratt and Dr Sarah Martins da Silva (Ninewells Hospital, University of Dundee), and Dr Sheryl Homa (Andrology Solutions, London) for assisting with patient recruitment.
References
- Abadi MK-D, Tavalaee M, Shahverdi A, Nasr-Esfahani MH.2016Evaluation of PLCζ and PAWP expression in Globozoospermic individuals. Cell Journal 18 438. ( 10.22074/cellj.2016.4572) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agarwal A, Baskaran S, Parekh N, Cho C-L, Henkel R, Vij S, Arafa M, Selvam MKP, Shah R.2021Male infertility. Lancet 23319–333. ( 10.1016/S0140-6736(2032667-2) [DOI] [PubMed] [Google Scholar]
- Aghajanpour S, Ghaedi K, Salamian A, Deemeh MR, Tavalaee M, Moshtaghian J, Parrington J, Nasr-Esfahani MH.2011Quantitative expression of phospholipase C zeta, as an index to assess fertilization potential of a semen sample. Human Reproduction 262950–2956. ( 10.1093/humrep/der285) [DOI] [PubMed] [Google Scholar]
- Amoroso-Sanches F, Gonzalez-Castro R, Stokes J, Carnevale E.2019180 Stallion sperm phospholipase C zeta affects cleavage rates after intracytoplasmic injection in bovine oocytes. Reproduction, Fertility and Development 31214–215. ( 10.1071/RDv31n1Ab180) [DOI] [Google Scholar]
- Azad N, Nazarian H, Ghaffari Novin M, Masteri Farahani R, Piryaei A, Heidari MH.2018Phospholipase C zeta parameters in sperm from polymorphic teratozoospermic men. Annals of Anatomy 21563–70. ( 10.1016/j.aanat.2017.09.007) [DOI] [PubMed] [Google Scholar]
- Bedford-Guaus SJ, Mcpartlin LA, Xie J, Westmiller SL, Buffone MG, Roberson MS.2011Molecular cloning and characterization of phospholipase C zeta in equine sperm and testis reveals species-specific differences in expression of catalytically active protein. Biology of Reproduction 8578–88. ( 10.1095/biolreprod.110.089466) [DOI] [PubMed] [Google Scholar]
- Bi Y, Xu WM, Wong HY, Zhu H, Zhou ZM, Chan HC, Sha JH.2009NYD-SP27, a novel intrinsic decapacitation factor in sperm. Asian Journal of Andrology 11 229–239. ( 10.1038/aja.2009.6) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cardona Barberán A, Boel A, Vanden Meerschaut F, Stoop D, Heindryckx B.2020Diagnosis and treatment of male infertility-related fertilization failure. Journal of Clinical Medicine 9 3899. ( 10.3390/jcm9123899) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan PJ, Jacobson JD, Corselli JU, Patton WC.2006A simple zeta method for sperm selection based on membrane charge. Fertility and Sterility 85481–486. ( 10.1016/j.fertnstert.2005.07.1302) [DOI] [PubMed] [Google Scholar]
- Cheung S, Xie P, Parrella A, Keating D, Rosenwaks Z, Palermo GD.2020Identification and treatment of men with phospholipase Cζ-defective spermatozoa. Fertility and Sterility 114535–544. ( 10.1016/j.fertnstert.2020.04.044) [DOI] [PubMed] [Google Scholar]
- Dai J, Dai C, Guo J, Zheng W, Zhang T, Li Y, Lu C, Gong F, Lu G, Lin G.2020Novel homozygous variations in PLCZ1 lead to poor or failed fertilization characterized by abnormal localization patterns of PLCζ in sperm. Clinical Genetics 97347–351. ( 10.1111/cge.13636) [DOI] [PubMed] [Google Scholar]
- Escoffier J, Lee HC, Yassine S, Zouari R, Martinez G, Karaouzène T, Coutton C, Kherraf ZE, Halouani L, Triki Cet al. 2015aHomozygous mutation of PLCZ1 leads to defective human oocyte activation and infertility that is not rescued by the WW-binding protein PAWP. Human Molecular Genetics 25878–891. ( 10.1093/hmg/ddv617) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Escoffier J, Yassine S, Lee HC, Martinez G, Delaroche J, Coutton C, Karaouzene T, Zouari R, Metzler-Guillemain C, Pernet-Gallay Ket al. 2015bSubcellular localization of phospholipase Cζ in human sperm and its absence in DPY19L2-deficient sperm are consistent with its role in oocyte activation. Molecular Human Reproduction 21157–168. ( 10.1093/molehr/gau098) [DOI] [PMC free article] [PubMed] [Google Scholar]
- ESHRE2018ART Fact Sheet [Online]. European Society of Human Reproduction and Embryology. (available at: https://www.eshre.eu/Accreditation-and-Certification/Nurses-Midwives-Certification/Information-for-exam-applicants/Reading-list) [Google Scholar]
- Esteves SC.2016Novel concepts in male factor infertility: clinical and laboratory perspectives. Journal of Assisted Reproduction and Genetics 331319–1335. ( 10.1007/s10815-016-0763-8) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrer-Buitrago M, Dhaenens L, Lu Y, Bonte D, Vanden Meerschaut F, De Sutter P, Leybaert L, Heindryckx B.2018Human oocyte calcium analysis predicts the response to assisted oocyte activation in patients experiencing fertilization failure after ICSI. Human Reproduction 33416–425. ( 10.1093/humrep/dex376) [DOI] [PubMed] [Google Scholar]
- Ferrer-Vaquer A, Barragan M, Freour T, Vernaeve V, Vassena R.2016PLCζ sequence, protein levels, and distribution in human sperm do not correlate with semen characteristics and fertilization rates after ICSI. Journal of Assisted Reproduction and Genetics 33747–756. ( 10.1007/s10815-016-0718-0) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grasa P, Coward K, Young C, Parrington J.2008The pattern of localization of the putative oocyte activation factor, phospholipase Cζ, in uncapacitated, capacitated, and ionophore-treated human spermatozoa. Human Reproduction 232513–2522. ( 10.1093/humrep/den280) [DOI] [PubMed] [Google Scholar]
- Hachem A, Godwin J, Ruas M, Lee HC, Buitrago MF, Ardestani G, Bassett A, Fox S, Navarrete F, De Sutter P.2017PLCζ is the physiological trigger of the Ca 2+ oscillations that induce embryogenesis in mammals but offspring can be conceived in its absence. Development 1442914–2924. ( 10.1242/dev.150227) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haddad M, Stewart J, Xie P, Cheung S, Trout A, Keating D, Parrella A, Lawrence S, Rosenwaks Z, Palermo GD.2021Thoughts on the popularity of ICSI. Journal of Assisted Reproduction and Genetics 38101–123. ( 10.1007/s10815-020-01987-0) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heindryckx B, Van Der Elst J, De Sutter P, Dhont M.2005Treatment option for sperm-or oocyte-related fertilization failure: assisted oocyte activation following diagnostic heterologous ICSI. Human Reproduction 202237–2241. ( 10.1093/humrep/dei029) [DOI] [PubMed] [Google Scholar]
- Heindryckx B, De Gheselle S, Gerris J, Dhont M, De Sutter P.2008Efficiency of assisted oocyte activation as a solution for failed intracytoplasmic sperm injection. Reproductive Biomedicine Online 17662–668. ( 10.1016/s1472-6483(1060313-6) [DOI] [PubMed] [Google Scholar]
- Heytens E, Parrington J, Coward K, Young C, Lambrecht S, Yoon SY, Fissore RA, Hamer R, Deane CM, Ruas Met al. 2009Reduced amounts and abnormal forms of phospholipase C zeta (PLCzeta) in spermatozoa from infertile men. Human Reproduction 242417–2428. ( 10.1093/humrep/dep207) [DOI] [PubMed] [Google Scholar]
- HFEA 2017Human fertilisation and embryology authority. (available at: https://www.hfea.gov.uk/treatments/explore-all-treatments/treatment-add-ons/) [Google Scholar]
- HFEA 2018Fertility Treatment 2014–2016: Trends and Figures [Online]. Human Fertilisation and Embryology Authority. (available at: https://www.hfea.gov.uk/media/2563/hfea-fertility-trends-and-figures-2017-v2.pdf) [Google Scholar]
- Janghorban-Laricheh E, Ghazavi-Khorasgani N, Tavalaee M, Zohrabi D, Abbasi H, Nasr-Esfahani MH.2016An association between sperm PLCζ levels and varicocele? Journal of Assisted Reproduction and Genetics 331649–1655. ( 10.1007/s10815-016-0802-5) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Javadian-Elyaderani S, Ghaedi K, Tavalaee M, Rabiee F, Deemeh MR, Nasr-Esfahani MH.2016Diagnosis of genetic defects through parallel assessment of PLCζ and CAPZA3 in infertile men with history of failed oocyte activation. Iranian Journal of Basic Medical Sciences 19 281–289. [PMC free article] [PubMed] [Google Scholar]
- Jones KT.2018Mammalian sperm contain two factors for calcium release and egg activation: phospholipase C zeta and a cryptic activating factor. Molecular Human Reproduction 24465–468. ( 10.1093/molehr/gay038) [DOI] [PubMed] [Google Scholar]
- Kashir J.2020Increasing associations between defects in phospholipase C zeta and conditions of male infertility: not just ICSI failure? Journal of Assisted Reproduction and Genetics 371273–1293. ( 10.1007/s10815-020-01748-z) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kashir J, Heynen A, Jones C, Durrans C, Craig J, Gadea J, Turner K, Parrington J, Coward K.2011aEffects of cryopreservation and density-gradient washing on phospholipase C zeta concentrations in human spermatozoa. Reproductive Biomedicine Online 23263–267. ( 10.1016/j.rbmo.2011.04.006) [DOI] [PubMed] [Google Scholar]
- Kashir J, Jones C, Lee HC, Rietdorf K, Nikiforaki D, Durrans C, Ruas M, Tee ST, Heindryckx B, Galione Aet al. 2011bLoss of activity mutations in phospholipase C zeta (PLCζ) abolishes calcium oscillatory ability of human recombinant protein in mouse oocytes. Human Reproduction 263372–3387. ( 10.1093/humrep/der336) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kashir J, Konstantinidis M, Jones C, Lemmon B, Chang Lee H, Hamer R, Heindryckx B, Deane CM, De Sutter P, Fissore RA.2011cA maternally inherited autosomal point mutation in human phospholipase C zeta (PLCζ) leads to male infertility. Human Reproduction 27222–231. ( 10.1093/humrep/der384) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kashir J, Konstantinidis M, Jones C, Heindryckx B, De Sutter P, Parrington J, Wells D, Coward K.2012aCharacterization of two heterozygous mutations of the oocyte activation factor phospholipase C zeta (PLCζ) from an infertile man by use of minisequencing of individual sperm and expression in somatic cells. Fertility and Sterility 98423–431. ( 10.1016/j.fertnstert.2012.05.002) [DOI] [PubMed] [Google Scholar]
- Kashir J, Sermondade N, Sifer C, Oo SL, Jones C, Mounce G, Turner K, Child T, Mcveigh E, Coward K.2012bMotile sperm organelle morphology evaluation-selected globozoospermic human sperm with an acrosomal bud exhibits novel patterns and higher levels of phospholipase C zeta. Human Reproduction 273150–3160. ( 10.1093/humrep/des312) [DOI] [PubMed] [Google Scholar]
- Kashir J, Jones C, Mounce G, Ramadan WM, Lemmon B, Heindryckx B, De Sutter P, Parrington J, Turner K, Child Tet al. 2013Variance in total levels of phospholipase C zeta (PLC-ζ) in human sperm may limit the applicability of quantitative immunofluorescent analysis as a diagnostic indicator of oocyte activation capability. Fertility and Sterility 99107, .e3–117.e3. ( 10.1016/j.fertnstert.2012.09.001) [DOI] [PubMed] [Google Scholar]
- Kashir J, Buntwal L, Nomikos M, Calver BL, Stamatiadis P, Ashley P, Vassilakopoulou V, Sanders D, Knaggs P, Livaniou Eet al. 2017Antigen unmasking enhances visualization efficacy of the oocyte activation factor, phospholipase C zeta, in mammalian sperm. Molecular Human Reproduction 2354–67. ( 10.1093/molehr/gaw073) [DOI] [PubMed] [Google Scholar]
- Kashir J, Mistry BV, Busaleh L, Abu-Dawas R, Nomikos M, Ajlan A, Abu-Dawud R, Alyacoub N, Alhassan S, Lai FAet al. 2020Phospholipase C zeta profiles are indicative of optimal sperm parameters and fertilisation success in patients undergoing fertility treatment. Andrology 81143–1159. ( 10.1111/andr.12796) [DOI] [PubMed] [Google Scholar]
- Kashir J, Mistry BV, Gumssani MA, Rajab M, Abu-Dawas R, Almohanna F, Nomikos M, Jones C, Abu-Dawud R, Al-Yacoub Net al. 2021Advancing male age differentially alters levels and localization patterns of PLCzeta in sperm and testes from different mouse strains. Asian Journal of Andrology 23 178–187. ( 10.4103/aja.aja_67_20) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khakpour S, Sadeghi E, Tavalaee M, Bahadorani M, Nasr-Esfahani MH.2019Zeta method: a noninvasive method based on membrane charge for selecting spermatozoa expressing high level of phospholipaseCζ. Andrologia 51 e13249. ( 10.1111/and.13249) [DOI] [PubMed] [Google Scholar]
- Khalil AM, Guttman M, Huarte M, Garber M, Raj A, Rivea Morales D, Thomas K, Presser A, Bernstein BE, van Oudenaarden Aet al. 2009Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. PNAS 10611667–11672. ( 10.1073/pnas.0904715106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leslie SW, Siref LE, Khan MA.2020Male infertility. [Updated 12 Aug 2021]. In StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing. [Google Scholar]
- Meng X, Melo P, Jones C, Ross C, Mounce G, Turner K, Child T, Coward K.2020Use of phospholipase C zeta analysis to identify candidates for artificial oocyte activation: a case series of clinical pregnancies and a proposed algorithm for patient management. Fertility and Sterility 114163–174. ( 10.1016/j.fertnstert.2020.02.113) [DOI] [PubMed] [Google Scholar]
- Meng X Jones C Melo P Ross C Mounce G Child T & Coward K. 2022Antigen unmasking does not improve the visualization of phospholipase C zeta in human spermatozoa. Asian Journal of Andrology 241–8. ( 10.4103/aja202168) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montag M, Köster M, Van Der Ven K, Bohlen U, Van Der Ven H.2012The benefit of artificial oocyte activation is dependent on the fertilization rate in a previous treatment cycle. Reproductive Biomedicine Online 24521–526. ( 10.1016/j.rbmo.2012.02.002) [DOI] [PubMed] [Google Scholar]
- Mu J, Zhang Z, Wu L, Fu J, Chen B, Yan Z, Li B, Zhou Z, Wang W, Zhao Let al. 2020The identification of novel mutations in PLCZ1 responsible for human fertilization failure and a therapeutic intervention by artificial oocyte activation. Molecular Human Reproduction 2680–87. ( 10.1093/molehr/gaaa003) [DOI] [PubMed] [Google Scholar]
- Nazarian H, Azad N, Nazari L, Piryaei A, Heidari MH, Masteri-Farahani R, Karimi M, Ghaffari-Novin M.2019Effect of artificial oocyte activation on intra-cytoplasmic sperm injection outcomes in patients with lower percentage of sperm containing phospholipase Cζ: A randomized clinical trial. Journal of Reproduction and Infertility 203–9. ( 10.1016/j.fertnstert.2015.07.993) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicopoullos JD, Whitney E, Wells V, Batha S, Faris R, Abdalla H.2015Case reports to suggest an algorithm for management of total fertilisation failure prior to use of donor gametes. Journal of Assisted Reproduction and Genetics 321679–1684. ( 10.1007/s10815-015-0564-5) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nikiforaki D, Meerschaut FV, De Roo C, Lu Y, Ferrer-Buitrago M, De Sutter P, Heindryckx B.2016Effect of two assisted oocyte activation protocols used to overcome fertilization failure on the activation potential and calcium releasing pattern. Fertility and Sterility 105798–806.e2. ( 10.1016/j.fertnstert.2015.11.007) [DOI] [PubMed] [Google Scholar]
- Nomikos M, Elgmati K, Theodoridou M, Calver BL, Cumbes B, Nounesis G, Swann K, Lai FA.2011Male infertility-linked point mutation disrupts the Ca2+ oscillation-inducing and PIP2 hydrolysis activity of sperm PLCζ. Biochemical Journal 434211–217. ( 10.1042/BJ20101772) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nomikos M, Swann K, Lai FA.2012Starting a new life: Sperm PLC-zeta mobilizes the Ca2+ signal that induces egg activation and embryo development: an essential phospholipase C with implications for male infertility. BioEssays 34126–134. ( 10.1002/bies.201100127) [DOI] [PubMed] [Google Scholar]
- Nomikos M, Kashir J, Swann K, Lai FA.2013aSperm PLCζ: from structure to Ca2+ oscillations, egg activation and therapeutic potential. FEBS Letters 5873609–3616. ( 10.1016/j.febslet.2013.10.008) [DOI] [PubMed] [Google Scholar]
- Nomikos M, Yu Y, Elgmati K, Theodoridou M, Campbell K, Vassilakopoulou V, Zikos C, Livaniou E, Amso N, Nounesis Get al. 2013bPhospholipase Cζ rescues failed oocyte activation in a prototype of male factor infertility. Fertility and Sterility 9976–85. ( 10.1016/j.fertnstert.2012.08.035) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nomikos M, Stamatiadis P, Sanders JR, Beck K, Calver BL, Buntwal L, Lofty M, Sideratou Z, Swann K, Lai FA.2017Male infertility-linked point mutation reveals a vital binding role for the C2 domain of sperm PLCζ. Biochemical Journal 4741003–1016. ( 10.1042/BCJ20161057) [DOI] [PubMed] [Google Scholar]
- Nozawa K, Satouh Y, Fujimoto T, Oji A, Ikawa M.2018Sperm-borne phospholipase C zeta-1 ensures monospermic fertilization in mice. Scientific Reports 8 1315. ( 10.1038/s41598-018-19497-6) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park JH, Kim SK, Kim J, Kim JH, Chang JH, Jee BC, Kim SH.2015Relationship between phospholipase C zeta immunoreactivity and DNA fragmentation and oxidation in human sperm. Obstetrics & Gynecology Science 58232–238. ( 10.5468/ogs.2015.58.3.232) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parrington J, Arnoult C, Fissore RA.2019The eggstraordinary story of how life begins. Molecular Reproduction and Development 864–19. ( 10.1002/mrd.23083) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahimizadeh P, Topraggaleh TR, Nasr-Esfahani MH, Ziarati N, Mirshahvaladi S, Esmaeili V, Seifi S, Eftekhari-Yazdi P, Shahverdi A.2020The alteration of PLCζ protein expression in unexplained infertile and asthenoteratozoospermic patients: a potential effect on sperm fertilization ability. Molecular Reproduction and Development 87115–123. ( 10.1002/mrd.23293) [DOI] [PubMed] [Google Scholar]
- Sanders JR, Swann K.2016Molecular triggers of egg activation at fertilization in mammals. Reproduction 152R41–R50. ( 10.1530/REP-16-0123) [DOI] [PubMed] [Google Scholar]
- Sanusi R, Yu Y, Nomikos M, Lai FA, Swann K.2015Rescue of failed oocyte activation after ICSI in a mouse model of male factor infertility by recombinant phospholipase Cζ. Molecular Human Reproduction 21783–791. ( 10.1093/molehr/gav042) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saunders CM, Larman MG, Parrington J, Cox LJ, Royse J, Blayney LM, Swann K, Lai FA.2002PLCζ: a sperm-specific trigger of Ca2+ oscillations in eggs and embryo development. Development 1293533–3544. ( 10.1242/dev.129.15.3533) [DOI] [PubMed] [Google Scholar]
- Sermondade N, Hafhouf E, Dupont C, Bechoua S, Palacios C, Eustache F, Poncelet C, Benzacken B, Levy R, Sifer C.2011Successful childbirth after intracytoplasmic morphologically selected sperm injection without assisted oocyte activation in a patient with globozoospermia. Human Reproduction 262944–2949. ( 10.1093/humrep/der258) [DOI] [PubMed] [Google Scholar]
- Swann K.2018The role of Ca2+ in oocyte activation during in vitro fertilization: insights into potential therapies for rescuing failed fertilization. Biochimica et Biophysica Acta: Molecular Cell Research 18651830–1837. ( 10.1016/j.bbamcr.2018.05.003) [DOI] [PubMed] [Google Scholar]
- Swann K.2020The soluble sperm factor that activates the egg: PLCzeta and beyond. Reproduction 160V9–V11. ( 10.1530/REP-20-0079) [DOI] [PubMed] [Google Scholar]
- Swann K, Lai FA.2016The sperm phospholipase C-ζ and Ca2+ signalling at fertilization in mammals. Biochemical Society Transactions 44267–272. ( 10.1042/BST20150221) [DOI] [PubMed] [Google Scholar]
- Tavalaee M, Nasr-Esfahani MH.2016Expression profile of PLC ζ, PAWP, and TR-KIT in association with fertilization potential, embryo development, and pregnancy outcomes in globozoospermic candidates for intra-cytoplasmic sperm injection and artificial oocyte activation. Andrology 4850–856. ( 10.1111/andr.12179) [DOI] [PubMed] [Google Scholar]
- Tavalaee M, Kiani-Esfahani A, Nasr-Esfahani MH.2017Relationship between potential sperm factors involved in oocyte activation and sperm DNA fragmentation with intra-cytoplasmic sperm injection clinical outcomes. Cell Journal 18 588–596. ( 10.22074/cellj.2016.4725) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tavalaee M, Nomikos M, Lai FA, Nasr-Esfahani MH.2018Expression of sperm PLCζ and clinical outcomes of ICSI-AOA in men affected by globozoospermia due to DPY19L2 deletion. Reproductive Biomedicine Online 36348–355. ( 10.1016/j.rbmo.2017.12.013) [DOI] [PubMed] [Google Scholar]
- Taylor SL, Yoon SY, Morshedi MS, Lacey DR, Jellerette T, Fissore RA, Oehninger S.2010Complete globozoospermia associated with PLCζ deficiency treated with calcium ionophore and ICSI results in pregnancy. Reproductive Biomedicine Online 20559–564. ( 10.1016/j.rbmo.2009.12.024) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terada Y, Hasegawa H, Takahashi A, Ugajin T, Yaegashi N, Okamura K.2009Successful pregnancy after oocyte activation by a calcium ionophore for a patient with recurrent intracytoplasmic sperm injection failure, with an assessment of oocyte activation and sperm centrosomal function using bovine eggs. Fertility and Sterility 91935.e11–935.e14. ( 10.1016/j.fertnstert.2008.09.043) [DOI] [PubMed] [Google Scholar]
- Tesarik J, Rienzi L, Ubaldi F, Mendoza C, Greco E.2002Use of a modified intracytoplasmic sperm injection technique to overcome sperm-borne and oocyte-borne oocyte activation failures. Fertility and Sterility 78619–624. ( 10.1016/s0015-0282(0203291-0) [DOI] [PubMed] [Google Scholar]
- Torra-Massana M, Cornet-Bartolomé D, Barragán M, Durban M, Ferrer-Vaquer A, Zambelli F, Rodriguez A, Oliva R, Vassena R.2019Novel phospholipase C zeta 1 mutations associated with fertilization failures after ICSI. Human Reproduction 341494–1504. ( 10.1093/humrep/dez094) [DOI] [PubMed] [Google Scholar]
- Van Blerkom J, Cohen J, Johnson M.2015A plea for caution and more research in the ‘experimental’ use of ionophores in ICSI. Reproductive Biomedicine Online 30323–324. ( 10.1016/j.rbmo.2015.02.002) [DOI] [PubMed] [Google Scholar]
- Vanden Meerschaut F, Nikiforaki D, De Gheselle S, Dullaerts V, Van Den Abbeel E, Gerris J, Heindryckx B, De Sutter P.2012Assisted oocyte activation is not beneficial for all patients with a suspected oocyte-related activation deficiency. Human Reproduction 271977–1984. ( 10.1093/humrep/des097) [DOI] [PubMed] [Google Scholar]
- Vanden Meerschaut F, Leybaert L, Nikiforaki D, Qian C, Heindryckx B, De Sutter P.2013Diagnostic and prognostic value of calcium oscillatory pattern analysis for patients with ICSI fertilization failure. Human Reproduction 2887–98. ( 10.1093/humrep/des368) [DOI] [PubMed] [Google Scholar]
- Yan Z, Fan Y, Wang F, Yan Z, Li M, Ouyang J, Wu L, Yin M, Zhao J, Kuang Yet al. 2020Novel mutations in PLCZ1 cause male infertility due to fertilization failure or poor fertilization. Human Reproduction 35472–481. ( 10.1093/humrep/dez282) [DOI] [PubMed] [Google Scholar]
- Yelumalai S, Yeste M, Jones C, Amdani SN, Kashir J, Mounce G, Da Silva SJM, Barratt CL, Mcveigh E, Coward K.2015Total levels, localization patterns, and proportions of sperm exhibiting phospholipase C zeta are significantly correlated with fertilization rates after intracytoplasmic sperm injection. Fertility and Sterility 104561–8.e4. ( 10.1016/j.fertnstert.2015.05.018) [DOI] [PubMed] [Google Scholar]
- Yeste M, Jones C, Amdani SN, Patel S, Coward K.2016aOocyte activation deficiency: a role for an oocyte contribution? Human Reproduction Update 2223–47. ( 10.1093/humupd/dmv040) [DOI] [PubMed] [Google Scholar]
- Yeste M, Jones C, Amdani SN, Yelumalai S, Mounce G, Da Silva SJM, Child T, Coward K.2016bDoes advancing male age influence the expression levels and localisation patterns of phospholipase C zeta (PLCζ) in human sperm? Scientific Reports 627543. ( 10.1038/srep27543) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoon SY, Jellerette T, Salicioni AM, Lee HC, Yoo MS, Coward K, Parrington J, Grow D, Cibelli JB, Visconti PEet al. 2008Human sperm devoid of PLC, zeta 1 fail to induce Ca 2+ release and are unable to initiate the first step of embryo development. Journal of Clinical Investigation 1183671–3681. ( 10.1172/JCI36942) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young C, Grasa P, Coward K, Davis LC, Parrington J.2009Phospholipase C zeta undergoes dynamic changes in its pattern of localization in sperm during capacitation and the acrosome reaction. Fertility and Sterility 91 (Supplement) 2230–2242. ( 10.1016/j.fertnstert.2008.05.021) [DOI] [PubMed] [Google Scholar]
- Yuan P, Yang C, Ren Y, Yan J, Nie Y, Yan L, Qiao J.2020aA novel homozygous mutation of phospholipase C zeta leading to defective human oocyte activation and fertilization failure. Human Reproduction 35977–985. ( 10.1093/humrep/dez293) [DOI] [PubMed] [Google Scholar]
- Yuan P, Zheng L, Liang H, Lin Q, Ou S, Zhu Y, Lai L, Zhang Q, He Z, Wang W.2020bNovel mutations in the PLCZ1 gene associated with human low or failed fertilization. Molecular Genetics and Genomic Medicine 8 e1470. ( 10.1002/mgg3.1470) [DOI] [PMC free article] [PubMed] [Google Scholar]