

Abstract
Background
Water pollution due to uncontrolled release of chemical pollutants is an important global problem. Its effect on medically important insects, especially mosquitoes, is a critical issue in the epidemiology of mosquito-borne diseases.
Methods
In order to understand the effect of water pollutants on the demography of Anopheles stephensi, colonies were reared in clean, moderately and highly polluted water for three consecutive generations at 27 °C, 75% RH, and a photoperiod of 12:12 h (L:D). The demographic data of the 4th generation of An. stephensi were collected and analysed using the age-stage, two-sex life table.
Results
The intrinsic rate of increase (r), finite rate of increase (λ), mean fecundity (F) and net reproductive rate (R0) of An. stephensi in clean water were 0.2568 d−1, 1.2927 d−1, 251.72 eggs, and 109.08 offspring, respectively. These values were significantly higher than those obtained in moderately polluted water (r = 0.2302 d−1, λ = 1.2589 d−1, 196.04 eggs, and R0 = 65.35 offspring) and highly polluted water (r = 0.2282 d−1, λ = 1.2564 d−1, 182.45 eggs, and R0 = 62.03 offspring). Female adult longevity in moderately polluted (9.38 days) and highly polluted water (9.88 days) were significantly shorter than those reared in clean water (12.43 days), while no significant difference in the male adult longevity was observed among treatments.
Conclusions
The results of this study showed that An. stephensi can partially adapt to water pollution and this may be sufficient to extend the range of mosquito-borne diseases.
Keywords: Global water pollutants, Anopheles stephensi, Malaria vector, Life table, Adaptation
Background
Mosquitoes are important vectors of many diseases, including malaria, dengue fever, chikungunya, yellow fever, Zika, encephalitis, and filariasis [1–3]. Anopheles mosquitoes are particularly important as they transmit malaria [4]. The World Health Organization (WHO) estimated there were 241 million malaria cases globally and 627,000 deaths in 2020 which were only slightly lower than those in 2010 and 2017 [5–7]. In Iran, Anopheles stephensi is an important vector of malaria [8, 9]. This species is widely distributed in Asia including India, Pakistan, Afghanistan, Iran, Iraq, Bahrain, Oman, Saudi Arabia, Bangladesh, Myanmar, and South China [10]. Recently the distribution of this species expanded to Sri Lanka, Djibouti, Ethiopia, Sudan and Somalia [11]. In Iran, this species has a wide distribution in Khuzestan, Fars, Kerman, Hormozgan, Sistan va Baluchestan, and southern Kermanshah provinces [8, 12, 13]. It has been the main malaria vector in Iran for many decades and still causes sporadic malaria cases in the South and Southeast areas of Iran [8].
Malaria transmission by Anopheles mosquitoes is influenced by their population density, which is affected by a number of abiotic factors of aquatic habitats [14]. The physicochemical factors of aquatic habitats affect the survival, development, and fecundity of Anopheles and, consequently can play a key role in malaria outbreaks [15–17]. Usually, Anopheles mosquitoes breed in clean water bodies; however, some Anopheles larvae are able to complete their life cycle in polluted waters [18, 19]. Human domestic, industrial and agricultural activities have increased pollution in aquatic environments through wastewater, solid waste, application of detergents, pesticides and fertilizers [20, 21]. Global warming and environmental changes may increase aquatic pollution in the future [22–24]. There is the potential for adaptation of mosquitoes to these pollutants [25]. Therefore, the assessment of the impact of water pollutants on the population fitness of Anopheles is important [26, 27].
The ecological features of vector populations such as stage-specific survival, development, and fecundity are key components of disease epidemiology [28]. Life tables are used to evaluate the effect of water pollution on the growth, development, survival and fecundity of mosquitoes [29]. This offers the most comprehensive understanding of the population characteristics [30, 31], and female age-specific life tables have been widely used in numerous studies. These have been applied to studies of the survivorship and reproductive strategies of colonized culicine and anopheline mosquitoes such as Aedes aegypti [32], Culex quinquefasciatus [33], Culex tritaeniorhynchus [34], An. stephensi and Anopheles culicifacies [30]. Because the female age-specific life tables ignore the male population and cannot describe the stage differentiation, they cannot be used to predict population growth. Huang and Chi [35], Huang et al. [36] and Chi et al. [37] discussed in detail the problems of the female age-specific life tables. In contrast to the female age-specific life table, the age-stage, two-sex life table can describe the stage differentiation and include the male population [29, 38, 39].
The goal of the present study was to compare the demographic characteristics of An. stephensi using the age-stage, two-sex life table approach in clean, moderately, and highly polluted water to assess the adaptive capacity of An. stephensi in breeding sites containing different degrees of pollutions, and provide detailed information on life table data of An. stephensi in polluted water.
Methods
Anopheles stephensi strain
The beech laboratory strain of An. stephensi was colonized in 1947 [40]. This subsection describes the methods used to maintain the colony in the insectary at the Medical Entomology and Vector Control Department, Mazandaran University of Medical Sciences, where it has been maintained for at least five generations. Eggs of An. stephensi (no older than 24 h) were put in white enamel trays (38 cm in length, 25 cm in width, and 10 cm in height) containing two litres of tap water allowing no more than 200 larvae in each tray [41]. Trays were kept at 27 ± 2 °C, 75 ± 5% RH and a photoperiod 12:12 h (L: D). About 24 h after eggs hatching, 0.5 g of TetraMin® fish food was daily added to the tray as larval food [41, 42]. When the 4th instars were observed, trays were checked daily for the presence of pupae. Pupae were collected and transferred into screened plastic cups containing tap water. Emerged adults, with a 1:1 ratio of male and female (25 pairs), were transferred to a screened rearing cage (40 × 40 × 40 cm) where they could readily mate. A restrained guinea pig was supplied to females for blood-feeding for 120 min. Initially, attempts were made to sedate the guinea pigs with different drugs, however this caused unusual malformations in the legs of the emerging adult mosquitoes (becoming hook like). Unfortunately, a high proportion failed to fully hatch from the pupal case and drowned. Mosquito blood feeding was carried out according to the ethical approval No. IR.MAZUMS.REC.1398.1398.
A vial containing cotton wicks soaked in a 10% sugar solution was placed in each cage as the source of energy for adults. To minimize the growth of mold or bacteria, sugar solution and cotton wicks were replaced every three days. Mosquitoes were offered blood meals every 3 days, until all females had died. A white enamel bowl (7 cm diameter × 15 cm height) filled with 150 ml tap water was placed in the cage to collect mosquito eggs. The bowl was checked daily and newly laid eggs were transferred to trays filled with tap water.
Pollutants and preparation of polluted water samples
In this study, total organic carbon (TOC), nitrate, and phosphate which are the common contaminants of aquatic environments were considered as the pollutants. Water samples of three different qualities were prepared: clean water treatment (i.e., control treatment), moderately polluted water, and highly polluted water. Deionized water (DI) and tap water were used for the preparation of all stock solutions and polluted water. Clean water was prepared with tap water and deionized water (1:1 in ratio). All tap waters used for preparation of treatment waters were set aside for 24 h in the laboratory to remove chlorine. The chemical characteristics of moderately and highly polluted water samples containing different concentrations of nitrate, phosphate and total organic carbon (TOC) which were prepared in the laboratory are listed in Table 1. The moderately polluted water mimicked the concentration of pollutants in mosquito larval habitats in Mazandaran Province [16]. Highly polluted water had the maximum permissible amount of each component in drinking water according to the World Health Organization standard [43]. Water samples were prepared by adding humic acid, sodium nitrate (NaNO3) and potassium dihydrogen phosphate (KH2PO4) to clean water as the sources for TOC, nitrate and phosphate, respectively [44].
Table 1.
Quality of clean, moderately polluted and highly polluted water: concentration of phosphate, nitrate, and total organic carbon (TOC), pH and electrical conductivity (EC)
| Parameter | Clean water | Moderately polluted water | Highly polluted water |
|---|---|---|---|
| Phosphate (mg. L−1) | < 0.01 | 2 | 6 |
| Nitrate (mg. L−1) | < 0.01 | 5 | 50 |
| TOC (mg. L−1) | < 0.01 | 5 | 10 |
| pH | 7.33 | 7.17 | 7.24 |
| EC (µS/cm) | 380.4 | 465.6 | 544.2 |
Total organic carbon (TOC) as a measure of the amount of organic compounds in all water samples was measured according to the Standard Method 5310B using TOC-VCSH TOC Analyzer (Shimadzu, Japan). If a water sample contains gross solids or insoluble matter, it is filtered to obtain homogenous solution. After this preparatory step, sample is injected into the analyzer according to the manufacturer’s instructions. Organic carbon is oxidized (during a combustion process in the instrument) to carbon dioxide, CO2. The resulting CO2 is purged from the solution and carried to an infrared analyzer specifically tuned to quantify the CO2 contents of the sample. The instrument’s microprocessor converts the detector signal to organic carbon concentration in mg/L based on a stored calibration curve [45]. The concentration of nitrate and phosphate was determined by spectrophotometry (Hach Company DR 2000), according to the Standard Methods [46]. Electrical conductivity (EC) and pH were measured using a table pH/EC meter (Eutech Instrument). The concentration of candidate chemicals along with pH and EC in the different water samples were controlled every three days throughout the experiments.
Life table study
Age-stage, two-sex life tables were used to analyse the influence of water conditions on the development and survival of An. stephensi. The mosquitoes were raised in the three different water treatments for four generations before recording differences in the population parameters. The three water treatments were: clean, moderately and highly polluted water.
To conduct the study, newly emerged male (n = 25) and female (n = 25) adults were released into screened rearing cages (40 × 40 × 40 cm) and were offered 10% sucrose solution in vials with cotton wicks. Mosquitoes were blood-fed after mating on a restrained guinea pig. White enamel bowls (7 cm diameter × 15 cm height) filled with 150 ml of the respective water were used to collect mosquito eggs. To determine the life table, newly hatched first instar larvae (L1) were used. For this purpose, egg masses (about 200 eggs) laid within 24 h of each treatment were transferred to white enamel bowls containing each of the water treatments. After hatching, 150 newly hatched first instar larvae (L1) were individually transferred into 8-cell containers (one larva per cell; dimension of each cell: 10 × 3 × 3 cm) containing 35 ml of each water type per cell. Larvae were fed daily with a diet of TetraMin® fish food (0.0025 gr per larva). The numbers of live mosquitoes at each developmental stage [47] were observed daily. Subsequently, newly emerged males and females were reared in group (as many as emerged in circa 1:1 ratio) in cages (40 × 40 × 40 cm) and data files were prepared as group-reared life table and analysed [48]. A white paper sheet was placed on the bottom of the cage to facilitate the identification and removal of dead mosquitoes. Dead adults were sexed and removed daily. Both sugar solution and blood meal were routinely offered to the mosquitoes as detailed previously. A white enamel bowl filled with water of the respective treatment was provided for egg collection. The bowl was replaced daily. Egg masses were kept until hatched and the number of hatched eggs was recorded as daily fecundity. The procedure was repeated until all adults in the cage died. During the life table study, mosquitoes were kept at 27 ± 2 °C, 75 ± 5% RH and a photoperiod 12:12 h (L: D).
Life table data analyses
All life table raw data on the development, survival and daily fecundity of An. stephensi were analysed according to the age-stage, two-sex life table theory [38, 39] using the TWOSEX-MSChart program [49]. The age-stage-specific survival rate (sxj: the probability that a newborne egg will survive to age x and stage j); the age-specific survival rate (lx: probability that a newborne egg will survive to age x); female fecundity (F: eggs/female); the age-stage-specific fecundity (fxj: the number of hatched eggs produced by female adult at age x); the age-specific fecundity (mx: the number of eggs per individual at age x); and age-specific maternity (lxmx: the product of lx and mx) were calculated. All population parameters including the intrinsic rate of increase (r), the finite rate of increase (λ), the net reproductive rate (R0) and the mean generation time (T) were calculated.
The age-stage-specific survival rate (sxj) was calculated as:
The age-specific survival rate (lx) was calculated for both females and males using the following formula:
where δ is the last stage of the cohort.
The age-specific fecundity (mx) at age x was calculated as:
The net reproductive rate (R0) is the sum of all lxmx (age-specific maternity) which considers the survival rate and is calculated by the following equation:
The intrinsic rate of increase (r) is calculated using the Lotka-Euler equation with age indexed from zero [50] using the following formula:
The finite rate of increase (λ) was calculated as λ = er. The mean generation time (T) is the time length that a population requires to increase to R0-fold of its size as time approaches infinity and the population settles down to a stable age-stage distribution and is calculated as:
Age-stage specific life expectancy (exj) defined as the number of days that an individual of age x and stage j is expected to live, is calculated using the following equation by assuming ;
where is the probability that an individual of age x and stage j will survive to age i and stage y.
The reproductive value (vxj) represents the contribution of individuals of age x and stage j to the future population [51] and was estimated using the following formula [29, 52]:
Since anopheline mosquitoes require 8–12 days (depending on the malaria parasite species) to become infective [53, 54], the number of female adults that survived more than 10 days (Nf,AL≥10) and the proportion of these female adults in the life table cohort (Nf,AL≥10/N) were calculated.
To estimate standard errors of the life-table parameters, group-reared data was converted to individual-reared data [48]. Then the bootstrap technique was used with 100,000 resampling [55, 56] and differences between treatments were assessed using the paired bootstrap [57, 58].
The parameter of adult pre-oviposition period (APOP) was calculated based on the period between the emergence of an adult female until initiation of the first oviposition; and total pre-oviposition period (TPOP) was calculated based on the time interval from egg to the beginning of oviposition of the resulting adult. Oviposition days (Od) defined as the number of days in which females laid eggs [58, 59].
Population projection
The life table data obtained were used to project the population size of An. stephensi [36]. Ten newly laid eggs of An. stephensi were used as the initial population to project the total population size for 40 days using the computer program TIMING-MSChart (Chi 2021). The 100,000 bootstrap results of the finite rate (λ) were sorted to find the life table of 2.5th and 97.5th percentiles (i.e., the 2500th and 97,500th sorted bootstrap samples) of the finite rate. These life tables were then used to predict the variability of population growth [36].
Results
Development, survival and reproduction
The durations of all stages of An. stephensi reared in three water treatments are shown in Table 2. The effect of water quality on developmental duration of the egg, larva, pupa, and adult stage of An. stephensi were variable. The longevity of adult females raised in clean water (average of 12.43 ± 0.81 days) was significantly longer than those whose immatures were raised in moderately (9.38 ± 0.65 days) or highly (9.88 ± 0.60 days) polluted water. There were significant differences in pupal duration among treatments. The shortest pupal duration was observed in the clean water treatment. However, there was no significant difference in total pre-adult duration among treatments. The immature survival rate in clean water (0.79% ± 0.03) was significantly higher than that in highly polluted water (0.64% ± 0.04) (Table 2).
Table 2.
Developmental duration, longevity, adult pre-oviposition period (APOP), total pre-oviposition period (TPOP), oviposition days (Od), total longevity of all eggs, and mean fecundity (± SE) of Anopheles stephensi reared in different water treatments
| Stage | Clean water | Moderately polluted | Highly polluted | |||
|---|---|---|---|---|---|---|
| n | Mean ± SE | n | Mean ± SE | n | Mean ± SE | |
| Egg (days) | 150 | 1.11 ± 0.03 b | 150 | 1.14 ± 0.03 ab | 150 | 1.19 ± 0.03 a |
| 1st Instar (days) | 149 | 1.88 ± 0.03 a | 150 | 1.63 ± 0.04 b | 150 | 1.80 ± 0.03 a |
| 2nd Instar (days) | 147 | 2.20 ± 0.06 a | 150 | 2.12 ± 0.03 a | 149 | 2.09 ± 0.04 a |
| 3rd Instar (days) | 127 | 1.94 ± 0.06 a | 143 | 1.94 ± 0.05 a | 120 | 2.00 ± 0.06 a |
| 4th Instar (days) | 121 | 2.28 ± 0.01 b | 121 | 1.90 ± 0.04 ab | 108 | 2.24 ± 0.04 a |
| Pupa (days) | 118 | 1.21 ± 0.04 c | 104 | 2.02 ± 0.01 a | 96 | 1.35 ± 0.05 b |
| Pre-adult (days) | 118 | 10.52 ± 0.05 a | 104 | 10.42 ± 0.05 a | 96 | 10.50 ± 0.05 a |
| Immature survival rate (%) | 150 | 0.79 ± 0.03 a | 150 | 0.69 ± 0.04 ab | 150 | 0.64 ± 0.04 b |
| Male adult longevity (days) | 53 | 3.45 ± 0.14 a | 54 | 3.56 ± 0.19 a | 45 | 3.89 ± 0.20 a |
| Male total longevity (days) | 53 | 13.98 ± 0.18 a | 54 | 13.83 ± 0.23 a | 45 | 14.33 ± 0.26 a |
| Female adult longevity (days) | 65 | 12.43 ± 0.81 a | 50 | 9.38 ± 0.65 b | 51 | 9.88 ± 0.60 b |
| Female total longevity (days) | 65 | 22.94 ± 0.86 a | 50 | 19.96 ± 0.71 b | 51 | 20.43 ± 0.66 b |
| Total longevity (days) | 150 | 16.64 ± 0.61 a | 150 | 14.57 ± 0.43 b | 150 | 14.33 ± 0.48 b |
| APOP (days) | 63 | 1.48 ± 0.06 a | 48 | 1.40 ± 0.07 a | 50 | 1.44 ± 0.07 a |
| TPOP (days) | 63 | 12.00 ± 0.00 a | 48 | 12.00 ± 0.00 a | 50 | 12.00 ± 0.00 a |
| Oviposition days (Od) (days) | 65 | 7.84 ± 0.57 a | 48 | 5.90 ± 0.46 b | 50 | 6.06 ± 0.44 b |
| Female fecundity (F) (eggs/female) | 65 | 251.72 ± 23.38 a | 50 | 196.04 ± 24.03 ab | 51 | 182.45 ± 18.95 b |
Means in the same row followed by different lower-case letters are significantly different based on the confidence intervals of the differences between different treatments using the paired bootstrap test (P < 0.05)
Means in the same column followed by different upper-case letters denote significant difference between sexes in the same treatment
When An. stephensi was reared in clean water, the mean fecundity was 251.72 ± 23.38 eggs/female, which is the highest among all treatments and significantly higher than that of highly polluted water (182.45 ± 18.95 eggs/female, P = 0.021), but not significantly different from that in the moderately polluted water (196.04 ± 24.03 eggs/female, P = 0.095). In the clean water treatment, the oviposition days (Od: the number of oviposition days) of An. stephensi was 7.84 ± 0.57 days, which was significantly longer than those in moderately and highly polluted treatments (Table 2).
There were no significant differences in adult pre-oviposition period (APOP) and total pre-oviposition period (TPOP) among treatments. The average adults’ female longevity was 12.43 ± 0.81 days in the clean water treatment, which was significantly longer than those in moderately (9.38 ± 0.65 days) and highly polluted water (9.88 ± 0.60 days). The total longevity of female individuals in clean water treatment was also significantly longer than those in moderately and highly polluted treatments. The age-stage-specific survival rates (sxj) (Fig. 1) showed that untreated An. stephensi survived longer than those in the moderately and highly polluted water. The shorter sxj curves of male An. stephensi in comparison with those of females showed longer survival of females. There were no significant differences in male adult longevity and total male longevity among treatments.
Fig. 1.

Age-stage survival rate curves (sxj) of Anopheles stephensi in clean, moderately and highly polluted water treatments
Age-specific survival (lx), age-specific fecundity of the total population (mx), and age-specific maternity (lxmx) of An. stephensi are demonstrated in Fig. 2. The mx curve revealed the periodic reproduction of An. stephensi. The mx curve for An. stephensi reared in clean water was extended from age 12 d to 38 d, which was significantly longer than the other two treatments. The longer reproductive period in the clean water treatment was consistent with the higher number of oviposition days (Od = 7.84 ± 0.57) (Table 2). Although higher peaks in the mx curve was observed in the moderately polluted water, the net maternity (lxmx) was lower than that of the clean water treatment due to the lower survival rate (lx).
Fig. 2.

Age-specific survival rate (lx), age-specific fecundity (mx), and age-specific maternity (lxmx) of Anopheles stephensi in clean, moderately and highly polluted water treatments
Population parameters
The intrinsic rate of increase (r), finite rate of increase (λ), the net reproductive rate (R0) and mean generation time (T) are given in Table 3. The value of r, λ and R0 for An. stephensi in clean water was 0.2568 ± 0.0066 d−1, 1.2927 ± 0.0085 d−1 and 109.08 ± 14.33 offspring/individual, respectively. These values were higher than those in moderately and highly polluted water. No significant differences were observed between moderately and highly polluted water. There was no significant difference in the mean generation time (T) among treatments.
Table 3.
Means (± SE) of population parameters of Anopheles stephensi reared in clean, moderately and highly polluted water: r, intrinsic rate of increase; λ, finite rate of increase; R0, net reproductive rate; T, mean generation time; Nf,AL≥10, number of female adults with adult longevity ≥ 10 days; Nf,AL≥10/N, proportion of female adults in cohort with adult longevity ≥ 10 days
| Parameters | Clean water | Moderately polluted | Highly polluted |
|---|---|---|---|
| r (d−1) | 0.2568 ± 0.0066 a | 0.2302 ± 0.0081 b | 0.2282 ± 0.0079 b |
| λ (d−1) | 1.2927 ± 0.0085 a | 1.2589 ± 0.0102 b | 1.2564 ± 0.0098 b |
| R0 (offspring/individual) | 109.08 ± 14.33 a | 65.35 ± 10.91 b | 62.03 ± 9.47 b |
| T (d) | 18.27 ± 0.24 a | 18.16 ± 0.28 a | 18.08 ± 0.23 a |
| Nf,AL≥10 | 45.00 ± 5.62 a | 22.00 ± 4.14 b | 27.00 ± 4.68 b |
| Nf,AL≥10/N | 0.30 ± 0.0375 a | 0.1467 ± 0.0288 b | 0.18 ± 0.0312 b |
Means in the same row followed by different letters are significantly different based on the confidence intervals of the differences between treatments using the paired bootstrap test (P < 0.05)
Out of 150 eggs, 45 female adults survived longer than 10 days (Nf,AL≥10 = 45.00 ± 5.62 females) in clean water, which was significantly more than that in moderately polluted water (Nf,AL≥10 = 22.00 ± 4.14 females) and that in highly polluted water (Nf,AL≥10 = 27.00 ± 4.68 females). The proportion of these female adults in clean water was 0.3 (i.e., Nf,AL≥10/N = 0.3 ± 0.0375), which was significantly greater than that in the moderately polluted water (Nf,AL≥10/N = 0.1467 ± 0.0288), and highly polluted water (Nf,AL≥10/N = 0.18 ± 0.0312), respectively.
The life expectancy (exj) of each age-stage group of An. stephensi is illustrated in Fig. 3. It represents the time that an individual of age x and stage j is expected to live [60]. The mean life expectancy of a newly laid egg (e01) was 16.64, 14.57 and 14.33 d in clean, moderately and highly polluted water treatment, respectively (Table 2). When the first female adult emerged, life expectancy was 12.94, 9.96 and 10.43 d in the clean, moderately and highly polluted water (Fig. 3), respectively; while the life expectancy curve for the male adult began with 3.93, 3.83 and 4.33 d in the clean, moderately and highly polluted water, respectively. The lower curves of male adults showed the male adults had a shorter longevity than females. The reproductive value (vxj) is the contribution of individuals of age x and stage j to the future population (Fig. 4). At age 0, the reproductive value v01 is exactly the finite rate of increase (λ), i.e., 1.2927 ± 0.0085, 1.2589 ± 0.0102 and 1.2564 ± 0.0098 d−1, in the clean, moderately and highly polluted water, respectively. The first peak of reproductive value for An. stephensi reared in clean, moderately and highly polluted water was 38.88, 37.75, and 36.22 d−1 at age 10 d when the female adult emerged.
Fig. 3.

Age-stage-specific life expectancy (exj) of Anopheles stephensi in clean, moderately and highly polluted water treatments
Fig. 4.

Age-stage-specific reproductive value (vxj) of Anopheles stephensi in clean, moderately and highly polluted water treatments
Population projection
Population growth projections with stage structures are shown in Fig. 5. The stage overlaps during population growth are evident. The total population size at the end of 40 days and its uncertainty are shown in Fig. 6. In the clean water treatment, the population size reached 63,246 individuals after 40 days, it was much higher than those in the moderately polluted water (24,401 individuals) and highly polluted water treatments (21,732 individuals), respectively (Fig. 6).
Fig. 5.

Population projection of Anopheles stephensi in clean, moderately and highly polluted water treatments
Fig. 6.
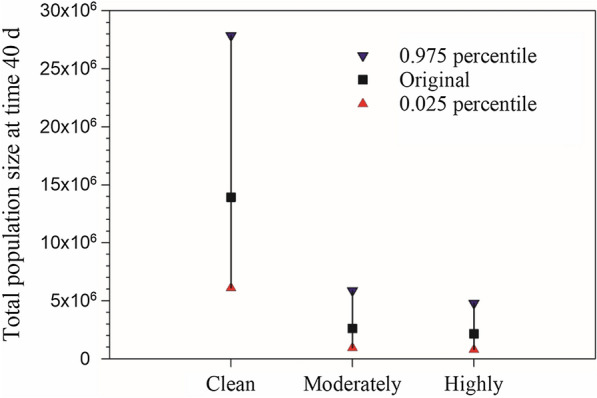
Population growth projection and the 0.025 and 0.975 confidence intervals of Anopheles stephensi in clean, moderately and highly polluted water treatments
By using the bootstrap technique, the uncertainty of population parameter of An. stephensi was estimated by using 100,000 resampling. The uncertainty of finite rate was shown in Fig. 7. The 100,000 finite rates randomly fluctuated around the mean (Fig. 7A1–A3). When these values were sorted in ascending order, a smooth curve was obtained (Fig. 7B1–B3). Frequency distribution of the 100,000 bootstrap finite rates of the An. stephensi in the different water habitats showed a normal distribution (Fig. 7C1–C3). The 0.025 and 0.975 percentiles of the finite rate of increase were obtained from the sorted data (Fig. 7B1–B3, C1–C3).
Fig. 7.

Top figures A1–A3 The unsorted finite rates of increase of 100,000 bootstrap results of the Anopheles stephensi in clean, moderately and highly polluted water treatments. The finite rates of 100,000 fluctuated randomly around the mean. Middle figures B1–B3 The 100,000 finite rates of increase were sorted in ascending order. Bottom figures C1–C3 The histogram of 100,000 finite rates. Finite rates of 0.025 and 0.975 percentiles can be observed in (B1–B3) and (C1–C3)
Discussion
In this study, the effect of water pollution on the population fitness of An. stephensi is presented. The mosquito larvae of malaria vectors can adapt to aquatic environments with different physicochemical characteristics [25, 61–63]. Anopheles species, such as An. stephensi, An. culicifacies, Anopheles gambiae and Anopheles coluzzii can adapt to a wide range of water bodies including waste or polluted waters [14, 25, 64, 65]. Knowledge on the population fitness of major malaria vectors breeding in different aquatic environments can inform disease and vector management strategies. Life table data offers comprehensive information about a population, and provide a useful tool for population ecology and pest management [36]. Based on the age-stage, two-sex life table, it was demonstrated that An. stephensi could successfully survive and reproduce when their eggs and larvae were reared in moderately or highly polluted water. Using Kaplan–Meier estimator with log-rank test, Oliver and Brooke showed similar adaptation of Anopheles arabiensis in metal polluted water [27]. Because the Kaplan–Meier method is only a descriptive method for the survival rate while ignoring the fecundity, it offers limited information about the population fitness.
In the present study, there were no significant differences in the development time of the total pre-adult duration of An. stephensi among clean, moderately polluted, and highly polluted water. However, the mean longevity of female adults emerging from clean water (12.43 d) was significantly longer than those from the moderately (9.38 d) and highly polluted water (9.88 d). It is necessary to mention here that female adult longevity influences vector potential, because the malaria parasite requires time to complete their development and sporogonic life cycle inside the mosquito body [28, 53]. This usually takes 8–12 days (intrinsic incubation period – the time required for an individual female to become infectious) depending on the malaria parasite species and the environmental factors [53, 54]. This means that at least some of the individuals in the cohorts may live longer than the intrinsic incubation period required for the sporogonic life cycle.
Despite significant differences in female longevity, there was no difference in adult pre-oviposition (APOP) and total pre-oviposition period (TPOP), i.e. the time needed to begin oviposition did not change due to water pollution. Earlier studies showed that An. gambiae and Aedes aegypti could adapt to water pollutants including cadmium chloride, copper nitrate and lead nitrate with costs of biological fitness such as reduction of egg viability, immature survivorship and reduced reproductive capacity [27, 66].
The life table parameters (r, λ, and R0) of An. stephensi were reduced in polluted water compared with those in clean water. It was revealed that An. stephensi has the ability to adapt to increasing levels of nitrate, phosphate and TOC, despite the unfavourable effect on the mosquito’s life table parameters.
However, the adaptation of mosquitoes to polluted water poses a serious challenge to the spread of diseases, such as malaria [66, 67], but the biological costs to mosquitoes should not be overlooked. For a correct assessment of the cost of biological fitness for adaptation, it is necessary to consider multiple population characteristics, as with the age-stage, two-sex life table, which includes the development duration, stage differentiation, survival and daily fecundity [38, 39]. After four generations of selection, the costs for mosquitoes to adapt to polluted water was largely seen as reduced longevity of the adult females, which consequently resulted in a lower number of oviposition events and lower lifetime fecundity. Limited studies have been performed on the underlying molecular response of Anopheles to water pollutants, e.g., tolerance to heavy metals can be caused by differential expression of metallothioneins and mucins [68, 69]. It was suggested that mosquitoes can regulate their biological program through proteome changes to counter polluted aquatic habitats [68]. Some species of Anopheles, such as An. gambiae, could genetically adapt to chemically altered aquatic habitats [66]. Also, previous studies showed that the life history parameters of mosquitoes are affected by ecological features, therefore they may undergo changes (molecular/structural) in order to adapt to the new conditions [28, 70]. Reduced fecundity can be the cost to tolerate pollutants in aquatic habitats [28, 66], as in this study. This adaptation is important for An. stephensi, which prefers to live near its human hosts and, therefore, increasingly encounters chemical pollutants in potential breeding sites, especially in urban areas.
The decreased life table parameters of Anopheles in polluted water did cause reduction in the rate of population growth. However, the long-term effect of mosquitoes' adaptation to water pollution needs to be monitored. First of all, pollution can affect the biodiversity of the aquatic organisms so that the species composition changes from natural species to compatible species [71]. Therefore, if natural enemies of the mosquito do not tolerate pollutants, they will lose the capacity of natural reduction of the mosquito population. Secondly, adaptation of Anopheles mosquitoes to polluted water can predispose them to insecticide resistance selection [72–74]. Therefore, chemical control programmes may fail to reduce the mosquito population. Some pollutants, such as nitrates and phosphates, can create a large proliferation of algae and bacteria, which is generally detrimental to aquatic organisms. This does not seem to be a serious problem for the Anopheles because their larvae and pupae breath directly from air through their siphons [75, 76], so an increase in the growth of algae and bacteria may increase mosquito larval food sources [77–79]. Hence although there is a measurable difference in life table parameters of mosquitoes in polluted water under laboratory conditions, these may be mitigated in field conditions.
The importance of this issue is understood by considering the fact that malaria incidence rate and its management can be assessed by the mathematical model such as "basic reproductive rate" (R0) [80] which has been used for the malaria risk assessment. R0 is calculated using the formula [80]. This is very important to note that changes in the life table parameters affect the density (m) and probability of living (p) of Anopheles mosquitoes, the two most important components of the disease reproduction rate (R0). According to the results of this study, the reduced life table parameters will result in the reduction of the parameters of m and p in the above equation; therefore, it seems that the disease transmission and risk of malaria would reduce. However, care must be taken to extend the negative impact of life table parameters under laboratory conditions on the R0 to the real life situation, and natural field condition studies with the same or other pollutants are recommended to conclude the impact of the water pollutants on the risk of malaria spread in a given area.
It should be noted that the present study faced with some limitations including: for individual rearing of mosquitoes, eggs would have to be singly put in small rearing trays, during this procedure, damage to the eggs might have been inevitable, the end result of which have been observed by rather small hatching rate. Therefore, the study was performed on individual L1 stage. Secondly, individual rearing of male and female adults was not possible in cages because most females did not show a desire to feed blood or if they did, they laid a small amount of eggs; therefore, this part of the study was performed as a group.
Conclusion
Findings presented in this paper demonstrate the impact of environmental pollutants including nitrate, phosphate and TOC on life table parameters such as r and R0, among others, of An. stephensi in the laboratory. Larval stages of An. stephensi prefer unpolluted water [25, 53–55], however, in the face of increasing environmental pollution on one hand and climate change leading to alteration of weather patterns (droughts, floods) on the other, they evolve to adapt to the situation enduring some fitness costs. Therefore, this adaptation may impact the distribution range of An. stephensi, as well as changes in the vectorial capacity of the mosquito, factors that may have epidemiological and vector control implications.
Acknowledgements
We would like to thank the deputy of research and technology of Mazandaran University of Medical Sciences for its support of this research.
Abbreviations
- r
The intrinsic rate of increase
- λ
Finite rate of increase
- R0
The net reproductive rate
- T
Mean generation time
- exj
Life expectancy
- sxj
Age-stage-specific survival rate
- fxj
Age-stage-specific fecundity
- lx
Age-specific survival rate
- mx
Age-specific fecundity
- lxmx
Age-specific maternity
- APOP
Adult pre-oviposition period
- TPOP
Total pre-oviposition period
Author contributions
MF and AA designed the study. MA, SHN, NH and FMH reared and collated the Anopheles data. HC, MF, and MSO undertook the analyses. MF, AA, MZ and JH led the writing of the manuscript. HC, MSO, MF, AA, JH and MZ performed the interpretation of analyses and joined the writing after the first draft. All authors contributed critically to the drafts and gave final approval for publication. All the authors read and approved the final manuscript.
Funding
Research reported in this publication was supported by Elite Researcher Grant Committee under award number 988000 from the National Institutes for Medical Research Development (NIMAD), Tehran, Iran. The Mazandaran University of Medical Sciences, Iran also contributed.
Availability of data and materials
The datasets analysed during the current study are available from the corresponding author (aenayati1372@gmail.com) on request.
Declarations
Ethics approval and consent to participate
This research was approved by the Mazandaran University of Medical Sciences ethics committee under number the IR.MAZUMS.REC.1398.1398.
Consent for publication
All the authors read the manuscript and consented for its publication in Malaria Journal.
Competing interests
The authors state that there is no conflict of interest.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Eldridge BF, Edman JD. Medical entomology: a textbook on public health and veterinary problems caused by arthropods. Berlin: Springer; 2004. p. 660. [Google Scholar]
- 2.Service M . Medical entomology for students. 5. Cambridge: Cambridge University Press; 2012. [Google Scholar]
- 3.Nikookar SH, Fazeli-Dinan M, Enayati A, Zaim M. Zika; a continuous global threat to public health. Environ Res. 2020;188:109868. doi: 10.1016/j.envres.2020.109868. [DOI] [PubMed] [Google Scholar]
- 4.Lehane MJ. The biology of blood-sucking in insects. Cambrige: Cambridge University Press; 2005. [Google Scholar]
- 5.WHO . World malaria report. 20 years of global progress and challenges. Geneva: World Health Organization; 2020. [Google Scholar]
- 6.WHO . World malaria report. Geneva: World Health Organization; 2019. [Google Scholar]
- 7.WHO . World malaria report. Geneva: World Health Organization; 2021. [Google Scholar]
- 8.Sanei-Dehkordi A, Soleimani-Ahmadi M, Jaberhashemi SA, Zare M. Species composition, seasonal abundance and distribution of potential anopheline vectors in a malaria endemic area of Iran: field assessment for malaria elimination. Malar J. 2019;18:157. doi: 10.1186/s12936-019-2795-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Madani A, Soleimani-Ahmadi M, Davoodi SH, Sanei-Dehkordi A, Jaberhashemi SA, Zare M, et al. Household knowledge and practices concerning malaria and indoor residual spraying in an endemic area earmarked for malaria elimination in Iran. Parasit Vectors. 2017;10:600. doi: 10.1186/s13071-017-2548-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Surendran SN, Sivabalakrishnan K, Gajapathy K, Arthiyan S, Jayadas TT, Karvannan K, et al. Genotype and biotype of invasive Anopheles stephensi in Mannar Island of Sri Lanka. Parasit Vectors. 2018;11:3. doi: 10.1186/s13071-017-2601-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Surendran SN, Sivabalakrishnan K, Sivasingham A, Jayadas TT, Karvannan K, Santhirasegaram S, et al. Anthropogenic factors driving recent range expansion of the malaria vector Anopheles stephensi. Public Health Front. 2019;7:53. doi: 10.3389/fpubh.2019.00053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hanafi-Bojd A, Vatandoost H, Oshaghi M, Haghdoost A, Shahi M, Sedaghat M, et al. Entomological and epidemiological attributes for malaria transmission and implementation of vector control in southern Iran. Acta Trop. 2012;121:85–92. doi: 10.1016/j.actatropica.2011.04.017. [DOI] [PubMed] [Google Scholar]
- 13.Vatandoost H, Oshaghi M, Abaie M, Shahi M, Yaaghoobi F, Baghaii M, et al. Bionomics of Anopheles stephensi Liston in the malarious area of Hormozgan province, southern Iran, 2002. Acta Trop. 2006;97:196–203. doi: 10.1016/j.actatropica.2005.11.002. [DOI] [PubMed] [Google Scholar]
- 14.Awolola T, Oduola A, Obansa J, Chukwurar N, Unyimadu J. Anopheles gambiae ss breeding in polluted water bodies in urban Lagos, southwestern Nigeria. J Vector Borne Dis. 2007;44:241–244. [PubMed] [Google Scholar]
- 15.Ndenga B, Githeko A, Omukunda E, Munyekenye G, Atieli H, Wamai P, et al. Population dynamics of malaria vectors in western Kenya highlands. J Med Entomol. 2014;43:200–206. doi: 10.1093/jmedent/43.2.200. [DOI] [PubMed] [Google Scholar]
- 16.Nikookar SH, Fazeli-Dinan M, Azari-Hamidian S, Mousavinasab SN, Aarabi M, Ziapour SP, et al. Correlation between mosquito larval density and their habitat physicochemical characteristics in Mazandaran Province, northern Iran. PLoS Negl Trop Dis. 2017;11:e0005835. doi: 10.1371/journal.pntd.0005835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nikookar S, Fazeli-Dinan M, Azari-Hamidian S, Mousavinasab S, Arabi M, Ziapour S, et al. Species composition and abundance of mosquito larvae in relation with their habitat characteristics in Mazandaran Province, northern Iran. Bull Entomol Res. 2017;107:598–610. doi: 10.1017/S0007485317000074. [DOI] [PubMed] [Google Scholar]
- 18.Sattler MA, Mtasiwa D, Kiama M, Premji Z, Tanner M, Killeen GF, et al. Habitat characterization and spatial distribution of Anopheles sp. mosquito larvae in Dar es Salaam (Tanzania) during an extended dry period. Malar J. 2005;4:4. doi: 10.1186/1475-2875-4-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kamdem C, Fossog BT, Simard F, Etouna J, Ndo C, Kengne P, et al. Anthropogenic habitat disturbance and ecological divergence between incipient species of the malaria mosquito Anopheles gambiae. PLoS ONE. 2012;7:e39453. doi: 10.1371/journal.pone.0039453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Chen M, Zeng G, Zhang J, Xu P, Chen A, Lu L. Global landscape of total organic carbon, nitrogen and phosphorus in lake water. Sci Rep. 2015;5:15043. doi: 10.1038/srep15043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.WHO . Nitrate and nitrite in drinking-water: background document for development of WHO Guidelines for Drinking-water Quality. Geneva: World Health Organization; 2003. [Google Scholar]
- 22.Shellenberger M, Nordhaus T. The death of environmentalism: global warming politics in a post-environmental world. Geopolit Hist Int Relat. 2009;1:121–163. [Google Scholar]
- 23.Manciocco A, Calamandrei G, Alleva E. Global warming and environmental contaminants in aquatic organisms: the need of the etho-toxicology approach. Chemosphere. 2014;100:1–7. doi: 10.1016/j.chemosphere.2013.12.072. [DOI] [PubMed] [Google Scholar]
- 24.Matus K, Yang T, Paltsev S, Reilly J, Nam KM. Toward integrated assessment of environmental change: air pollution health effects in the USA. Clim Change. 2008;88:59–92. doi: 10.1007/s10584-006-9185-4. [DOI] [Google Scholar]
- 25.Gunathilaka N, Fernando T, Hapugoda M, Wickremasinghe R, Wijeyerathne P, Abeyewickreme W. Anopheles culicifacies breeding in polluted water bodies in Trincomalee district of Sri Lanka. Malar J. 2013;12:285. doi: 10.1186/1475-2875-12-285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kudom AA. Larval ecology of Anopheles coluzzii in cape coast, Ghana: water quality, nature of habitat and implication for larval control. Malar J. 2015;14:447. doi: 10.1186/s12936-015-0989-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Oliver SV, Brooke BD. The effect of metal pollution on the life history and insecticide resistance phenotype of the major malaria vector Anopheles arabiensis (Diptera: Culicidae) PLoS ONE. 2018;13:e0192551. doi: 10.1371/journal.pone.0192551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yurttas H, Alten B. Geographic differentiation of life table attributes among Anopheles sacharovi (Diptera: Culicidae) populations in Turkey. J Vector Ecol. 2006;31:275–284. doi: 10.3376/1081-1710(2006)31[275:GDOLTA]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 29.Tuan SJ, Lee CC, Chi H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag Sci. 2014;70:805–813. doi: 10.1002/ps.3618. [DOI] [PubMed] [Google Scholar]
- 30.Reisen WK, Mahmood F. Horizontal life table characteristics of the malaria vectors Anopheles culicifacies and Anopheles stephensi (Diptera: Culicidae) J Med Entomol. 1980;17:211–217. doi: 10.1093/jmedent/17.3.211. [DOI] [Google Scholar]
- 31.Chi H. Timing-MSChart: a computer program for the population projection based on age-stage, two-sex life table. Taichung, Taiwan: National Chung Hsing University; http://140.120.197.173/Ecology/Download/Timing-MSChart.rar. 2020.
- 32.Crovello TJ, Hacker CS. Evolutionary strategies in life table characteristics among feral and urban strains of Aedes aegypti (L.) Evolution. 1972;26:185–196. doi: 10.1111/j.1558-5646.1972.tb00186.x. [DOI] [PubMed] [Google Scholar]
- 33.Suman D, Tikar S, Mendki M, Sukumaran D, Agrawal O, Parashar B, et al. Variations in life tables of geographically isolated strains of the mosquito Culex quinquefasciatus. Med Vet Entomol. 2011;25:276–288. doi: 10.1111/j.1365-2915.2010.00939.x. [DOI] [PubMed] [Google Scholar]
- 34.Reisen WK, Siddiqui TF, Aslam Y, Malik GM. Geographic variation among the life table characteristics of Culex tritaeniorhynchus from Asia. Ann Entomol Soc Am. 1979;72:700–709. doi: 10.1093/aesa/72.5.700. [DOI] [Google Scholar]
- 35.Huang YB, Chi H. Age-stage, two-sex life tables of Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations. Insect Sci. 2012;19:263–273. doi: 10.1111/j.1744-7917.2011.01424.x. [DOI] [Google Scholar]
- 36.Huang H-W, Chi H, Smith CL. Linking demography and consumption of Henosepilachna vigintioctopunctata (Coleoptera: Coccinellidae) fed on Solanum photeinocarpum (Solanales: Solanaceae): with a new method to project the uncertainty of population growth and consumption. J Econ Entomol. 2018;111:1–9. doi: 10.1093/jee/tox330. [DOI] [PubMed] [Google Scholar]
- 37.Chi H, You M, Atlihan R, Smith CL, Kavousi A, Özgökçe MS, et al. Age-stage, two-sex life table: an introduction to theory, data analysis, and application. Entomol Gen. 2020;40:103–124. doi: 10.1127/entomologia/2020/0936. [DOI] [Google Scholar]
- 38.Chi H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ Entomol. 1988;17:26–34. doi: 10.1093/ee/17.1.26. [DOI] [Google Scholar]
- 39.Chi H, Liu H. Two new methods for the study of insect population ecology. Bull Inst Zool Acad Sin. 1985;24:225–240. [Google Scholar]
- 40.Gorouhi MA, Oshaghi MA, Vatandoost H, Enayati AA, Raeisi A, Abai MR, et al. Biochemical basis of cyfluthrin and DDT resistance in Anopheles stephensi (Diptera: Culicidae) in malarious area of Iran. J Arthropod Borne Dis. 2018;12:310. [PMC free article] [PubMed] [Google Scholar]
- 41.Schmeid S, Seidahmed H, Soliban G, Stevens G, Thailayil A, Williams R. Methods in Anopheles research. 4. Manassas: BEI Resources; 2014. [Google Scholar]
- 42.Christiansen-Jucht C, Parham PE, Saddler A, Koella JC, Basáñez MG. Temperature during larval development and adult maintenance influences the survival of Anopheles gambiae s.s. Parasit Vectors. 2014;7:1–10. doi: 10.1186/s13071-014-0489-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Frisbie SH, Mitchell EJ, Dustin H, Maynard DM, Sarkar B. World Health Organization discontinues its drinking-water guideline for manganese. Environ Health Perspect. 2012;120:775–778. doi: 10.1289/ehp.1104693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Zazouli M, Nasseri S, Mahvi A, Mesdaghinia A, Younecian M, Gholami M. Determination of hydrophobic and hydrophilic fractions of natural organic matter in raw water of Jalalieh and Tehranspars water treatment plants (Tehran) J Appl Sci. 2007;7:2651–2655. doi: 10.3923/jas.2007.2651.2655. [DOI] [Google Scholar]
- 45.Standard Methods Committee of the American Public Health Association, American Water Works Association, and Water Environment Federation . 5310 Total organic carbon (TOC) In: Lipps WC, Baxter TE, Braun-Howland E, editors. Standard methods for the examination of water and wastewater. Washington DC: APHA Press; 2018. p. 1545. [Google Scholar]
- 46.Standard Methods Committee of the American Public Health Association, American Water Works Association, and Water Environment Federation. Lipps WC, Baxter TE, Braun-Howland E. Standard methods for the examination of water and wastewater. Washington DC: APHA Press; 2018. p. 1545. [Google Scholar]
- 47.Timmerman S. Molting and metamorphosis in mosquito larvae: a morphometric analysis. J Swiss Entomol Soc. 1998;71:373–387. [Google Scholar]
- 48.Chang C, Huang CY, Dai SM, Atlihan R, Chi H. Genetically engineered ricin suppresses Bactrocera dorsalis (Diptera: Tephritidae) based on demographic analysis of group-reared life table. J Econ Entomol. 2016;109:987–992. doi: 10.1093/jee/tow091. [DOI] [PubMed] [Google Scholar]
- 49.Chi H. TWOSEX-MSChart: a computer program for the age-stage, two-sex life table analysis. 2015;197. http://140.120.197.173/Ecology/prod02.htm
- 50.Goodman D. Optimal life histories, optimal notation, and the value of reproductive value. Am Nat. 1982;119:803–823. doi: 10.1086/283956. [DOI] [Google Scholar]
- 51.Fisher R. The genetical theory of natural selection. Oxford: Clarendon Press; 1930. p. 272. [Google Scholar]
- 52.Huang YB, Chi H. The age-stage, two-sex life table with an offspring sex ratio dependent on female age. J Agri Fore. 2011;60:337–345. [Google Scholar]
- 53.Grieco JP, Achee NL, Briceno I, King R, Andre R, Roberts D, et al. Comparison of life table attributes from newly established colonies of Anopheles albimanus and Anopheles vestitipennis in northern Belize. J Vector Ecol. 2003;28:200–207. [PubMed] [Google Scholar]
- 54.Sandosham AA, Thomas V. Malariology: with special reference to Malaya. Singapore: NUS Press; 1983. p. 408. [Google Scholar]
- 55.Efron B, Tibshirani RJ. An introduction to the bootstrap. Monographs on statistics and applied probability. Boca Raton: Chapman and Hall/CRC; 1993. p. 436. [Google Scholar]
- 56.Huang YB, Chi H. Assessing the application of the jackknife and bootstrap techniques to the estimation of the variability of the net reproductive rate and gross reproductive rate: a case study in Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae) J Agric Forest. 2012;61:37–45. [Google Scholar]
- 57.Smucker MD, Allan J, Carterette B. A comparison of statistical significance tests for information retrieval evaluation. Proceedings of the sixteenth ACM conference on Conference on information and knowledge management; 2007.
- 58.Xie W, Zhi J, Ye J, Zhou Y, Li C, Liang Y, et al. Age-stage, two-sex life table analysis of Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) reared on maize and kidney bean. Chem Biol Technol Agric. 2021;8:1–8. doi: 10.1186/s40538-021-00241-8. [DOI] [Google Scholar]
- 59.Ning S, Zhang W, Sun Y, Feng J. Development of insect life tables: comparison of two demographic methods of Delia antiqua (Diptera: Anthomyiidae) on different hosts. Sci Rep. 2017;7:4821. doi: 10.1038/s41598-017-05041-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Amir-Maafi M, Chi H. Demography of Habrobracon hebetor (Hymenoptera: Braconidae) on two pyralid hosts (Lepidoptera: Pyralidae) Ann Entomol Soc Am. 2006;99:84–90. doi: 10.1603/0013-8746(2006)099[0084:DOHHHB]2.0.CO;2. [DOI] [Google Scholar]
- 61.Edillo FE, Tripét F, Touré YT, Lanzaro GC, Dolo G, Taylor CE. Water quality and immatures of the M and S forms of Anopheles gambiae ss and An. arabiensis in a Malian village. Malar J. 2006;5:35. doi: 10.1186/1475-2875-5-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ndenga BA, Simbauni JA, Mbugi JP, Githeko AK, Fillinger U. Productivity of malaria vectors from different habitat types in the western Kenya highlands. PLoS ONE. 2011;6:e19473. doi: 10.1371/journal.pone.0019473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Sinka ME, Bangs MJ, Manguin S, Coetzee M, Mbogo CM, Hemingway J, et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: occurrence data, distribution maps and bionomic précis. Parasit Vectors. 2010;3:117. doi: 10.1186/1756-3305-3-117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ossè R, Bangana S, Aïkpon R, Kintonou J, Sagbohan H. Adaptation of Anopheles coluzzii larvae to polluted breeding sites in Cotonou. A strengthening in urban malaria transmission in Benin. Vector Biol J. 2019;4:1. [Google Scholar]
- 65.Thakare A, Ghosh C, Alalamath T, Kumar N, Narang H, Whadgar S, et al. The genome trilogy of Anopheles stephensi, an urban malaria vector, reveals structure of a locus associated with adaptation to environmental heterogeneity. Sci Rep. 2022;12:3610. doi: 10.1038/s41598-022-07462-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Mireji P, Keating J, Hassanali A, Mbogo C, Muturi M, Githure J, et al. Biological cost of tolerance to heavy metals in the mosquito Anopheles gambiae. Med Vet Entomol. 2010;24:101–107. doi: 10.1111/j.1365-2915.2010.00863.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Elkington JS. Insect population ecology: an African perspective. Nairobi: ICIPE Science Press; 1993. [Google Scholar]
- 68.Rono MK, Muturi CN, Ochieng R, Mwakubabanya R, Wachira FN, Mwangangi J, et al. Cadmium tolerance pathway in Anopheles gambiae senso stricto. Acta Trop. 2019;198:105033. doi: 10.1016/j.actatropica.2019.05.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Mireji PO, Keating J, Kenya E, Mbogo C, Nyambaka H, Osir E, et al. Differential induction of proteins in Anopheles gambiae sensu stricto (Diptera: culicidae) larvae in response to heavy metal selection. Int J Trop Insect Sci. 2006;26:214–226. doi: 10.1017/S1742758406658955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Yurttas H, Alten B, Aytekin A. Variability in natural populations of Anopheles sacharovi (Diptera: Culicidae) from southeast Anatolia, revealed by morphometric and allozymic analyses. J Vector Ecol. 2005;30:206–212. [PubMed] [Google Scholar]
- 71.Wang R, Hetai S, Shi Y, Wang Z, Xiao Y. Effects of pollution on aquatic ecology and water quality bio-assessment. Hydrobiologia. 2014;729:3–15. doi: 10.1007/s10750-013-1680-9. [DOI] [Google Scholar]
- 72.Azrag RS, Mohammed BH. Anopheles arabiensis in Sudan: a noticeable tolerance to urban polluted larval habitats associated with resistance to Temephos. Malar J. 2018;17:204. doi: 10.1186/s12936-018-2350-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Jeanrenaud AC, Brooke BD, Oliver SV. The effects of larval organic fertiliser exposure on the larval development, adult longevity and insecticide tolerance of zoophilic members of the Anopheles gambiae complex (Diptera: Culicidae) PLoS ONE. 2019;14:e0215552. doi: 10.1371/journal.pone.0215552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Jeanrenaud AC, Brooke BD, Oliver SV. Second generation effects of larval metal pollutant exposure on reproduction, longevity and insecticide tolerance in the major malaria vector Anopheles arabiensis (Diptera: Culicidae) Parasit Vectors. 2020;13:4. doi: 10.1186/s13071-020-3886-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Ha YR, Yeom E, Ryu J, Lee SJ. Three-dimensional structures of the tracheal systems of Anopheles sinensis and Aedes togoi pupae. Sci Rep. 2017;7:44490. doi: 10.1038/srep44490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Fried S, Mackie B, Nothwehr E. Nitrate and phosphate levels positively affect the growth of algae species found in Perry Pond. Tillers. 2003;4:21–24. [Google Scholar]
- 77.Walker ED, Olds EJ, Merritt RW. Gut content analysis of mosquito larvae (Diptera: Culicidae) using DAPI stain and epifluorescence microscopy. J Med Entomol. 1988;25:551–554. doi: 10.1093/jmedent/25.6.551. [DOI] [PubMed] [Google Scholar]
- 78.Merritt R, Dadd R, Walker E. Feeding behavior, natural food, and nutritional relationships of larval mosquitoes. Annu Rev Entomol. 1992;37:349–374. doi: 10.1146/annurev.en.37.010192.002025. [DOI] [PubMed] [Google Scholar]
- 79.Kaufman MG, Wanja E, Maknojia S, Bayoh MN, Vulule JM, Walker ED. Importance of algal biomass to growth and development of Anopheles gambiae larvae. J Med Entomol. 2006;43:669–676. doi: 10.1093/jmedent/43.4.669. [DOI] [PubMed] [Google Scholar]
- 80.Schofield S, Tepper M, Tuck JJ. Malaria risk assessment and preventive recommendations: a new approach for the Canadian military. Mil Med. 2007;172:1250–1253. doi: 10.7205/MILMED.172.12.1250. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets analysed during the current study are available from the corresponding author (aenayati1372@gmail.com) on request.