

Abstract
Diabetes and obesity are metabolic diseases that have become alarming conditions in recent decades. Their rate of increase is becoming a growing concern worldwide. Recent studies have established that the composition and dysfunction of the gut microbiota are associated with the development of diabetes. For this reason, strategies such as the use of prebiotics to improve intestinal microbial structure and function have become popular. Consumption of prebiotics for modulating the gut microbiota results in the production of microbial metabolites such as short-chain fatty acids that play essential roles in reducing blood glucose levels, mitigating insulin resistance, reducing inflammation, and promoting the secretion of glucagon-like peptide 1 in the host, and this accounts for the observed remission of metabolic diseases. Prebiotics can be either naturally extracted from non-digestible carbohydrate materials or synthetically produced. In this review, we discussed current findings on how the gut microbiota and microbial metabolites may influence host metabolism to promote health. We provided evidence from various studies that show the ability of prebiotic consumption to alter gut microbial profile, improve gut microbial metabolism and functions, and improve host physiology to alleviate diabetes and obesity. We conclude among other things that the application of systems biology coupled with bioinformatics could be essential in ascertaining the exact mechanisms behind the prebiotic–gut microbe–host interactions required for diabetes and obesity improvement.
Keywords: prebiotics, obesity, diabetes, gut microbiota, biotherapeutics, dietary fiber
1. Introduction
The condition of obesity and diabetes has risen drastically in the last decade, leading to a public health emergency. In a recent study, 463 million people were estimated to suffer from diabetes worldwide and the number is expected to increase in the coming years [1]. Diabetes is typically preceded by insulin resistance, where insulin action in peripheral tissues including the liver, skeletal muscles, and adipose tissues are impaired. This results in reduced insulin-stimulated glucose disposal, reduced lipolysis rates, and decreased insulin-induced suppression of hepatic glucose production [2]. There is increasing evidence that disruption of the gut microbiota function and composition could contribute to the pathogenesis of metabolic diseases such as diabetes [3] and obesity [4,5,6]. Consequently, it is crucial to evaluate the cross talk between the gut microbial composition in the gut, the development of metabolic disorders, and the potential therapeutic strategies to prevent these metabolic syndromes.
The mammalian gastrointestinal tract (GIT) is home to trillions of microorganisms, collectively known as the gut microbiota (GM) [7]. The GM is defined as an ecological community of commensal microorganisms that live symbiotically and pathogenically in the gut [8]. Colonization of neonatal gut may start during birth [9]. GM represents a complex ecosystem, consisting of numerous diverse sets of microorganisms such as viruses, fungi, bacteria, archaea, and phages, deeply implicated in different functions of host metabolism [10]. The most abundant phyla consists of Firmicutes (64%), Bacteroidetes (23%), Proteobacteria (8%), and Actinobacteria (3%) [11]. GM makes a crucial contribution to the production of enzymes that are not encoded by the human genome, for example, the breakdown of polysaccharides, polyphenols, and the synthesis of vitamins [12]; is pivotal for human development and physiology [13]; and plays a vital role in regulatory functions in health and disease [14].
The composition of the GM differs between person-to-person and can fluctuate significantly within an individual [15]. Variation in GM composition could be caused not only by differences in the host’s genome, but also by environmental factors, such as antibiotic use, lifestyle, hygiene, and diet administration [16,17]. Significant alterations in gut microbial composition (dysbiosis) can be unfavorable and can predispose an individual to disease. For instance, acute and chronic disorders such as obesity, inflammatory bowel disease, irritable bowel syndrome, diabetes, colon cancer, and antibiotic-associated diarrhea have all been associated with dysbiosis [12,18,19].
Food is considered as a substrate that greatly contributes to the growth of GM and has a significant influence on its composition [20]. In 1980, it was proposed that definite components of the diet could promote the proliferation of specific bacterial strains inhabiting in the GIT, which are associated with the benefit of the host’s health [21]. The dietary intervention with prebiotics can be classified as dietary fibers; however, not all fibers can be considered as prebiotics [22]. Dietary modulation of GM with prebiotics has shown great potential as an agent to ameliorate and perpetuate a balanced microbial composition to improve health and well-being [23,24,25,26].
In this review, we discussed prebiotics, their classification, and the modulatory capacity of GM for health promotion in the host. We also discussed in vivo and in vitro studies and human clinical trials to provide better insight into the benefits of prebiotics on health. Finally, we focused on the therapeutic uses of prebiotics in the treatment/prevention of obesity and type 2 diabetes mellitus (T2DM).
2. Prebiotics
Prebiotics are a class of nutritional compounds categorized together, not necessarily by structural affinity, but by the potential to promote the growth and/or activity of specific beneficial bacteria (probiotics) in the GM. The concept of prebiotics came into recognition due to Glenn Gibson and Marcel Roberfroid in 1995 [23]. A prebiotic is known as “a non-digestible food constituent that beneficially influences the host by selectively promoting the growth and/or activity of one or a restricted number of bacteria in the colon, and thus improving the host health” [27]. In 2004, prebiotics were upgraded to include four criteria: (1) resistance to hydrolysis by mammalian enzymes, gastric acidity, and gastrointestinal absorption; (2) they should only be fermented by GM; (3) induce systemic or luminal effects that are beneficial to host health; and (4) selectively stimulate the growth and activity of GM associated with health and well-being [28]. The health benefits of prebiotics are diverse and include immune modulation through increased immune-regulatory interleukins and intestinal-specific immunoglobulins; reduction of pro-inflammatory interleukins [29,30]; and production of short-chain fatty acids (SCFAs) such as acetate, propionate, and butyrate [31] (Figure 1). SCFAs are carboxylic acids with aliphatic tails of one to six carbons that are produced by anaerobic fermentation of dietary fibers in the intestine by the GM [32]. SCFAs are an important indicator of bacterial fermentation in the colon and are known to improve the gut health by maintaining intestinal barrier integrity [33], mucus production [34], protection against inflammation, and reduction in colorectal cancer and obesity [35].
Figure 1.

Mechanism of action of prebiotic supplementation. Prebiotic administration in a regular diet increases bacterial growth and functionality of specific species or genera, leading to modulation of the GM and showing a strong bifidogenic effect. The goblet cells play a key role in the production of mucus, which helps to protect the mucous membrane and form a layer in the colon that helps to reduce the inflammation caused by the bacterial interaction with intestinal epithelial cells. The modulated GM ferments prebiotics to form SCFAs (acetate, propionate, and butyrate), from which health benefits can be accrued. The production of antimicrobial agents and the reduction in the pH levels of the intestine due to prebiotic supplementation can suppress and restrict the growth of pathogenic bacteria, which can lead to positive health effects.
Among the abundant food ingredients available, some peptides and proteins, particular lipids, and non-digestible carbohydrates are components of prebiotics [36]. The chemical structures of these components are not absorbed in the upper part of the GIT or hydrolyzed by the digestive enzymes of humans. Hence, these ingredients are called colonic foods [37]. In colonic food, non-digestible carbohydrates are naturally occurring and meet all the criteria of prebiotics. These carbohydrates include non-starch polysaccharides, resistant starch, and non-digestible oligosaccharides [38]. However, not all of them are prebiotics [39]. In order to be classified as prebiotics, carbohydrates must fulfil the following criteria: (i) they are dietary fibers with a degree of polymerization (DP) between three and nine [40], and (ii) the endogenous enzymes produced in the small intestine should not hydrolyze them [41]. It should be taken into account that fermentation and fiber solubility are generally not curtailed [22].
Bacterial genera that promote health such as Lactobacillus and Bifidobacterium is proliferated by the administration of prebiotics, so that the fermented metabolites can be easily absorbed by the mammalian gut and have an influence on host physiology [42] (Figure 1). The prebiotics share several characteristics with dietary fiber, which includes partial or total resistance to digestion and fermentation by the GM. Due to its selectivity, prebiotics highlight the key condition to be demonstrated in an in-vivo experiment (including complex human or animal GM) using validated and relevant methodologies to quantify a wide variety of species that make up the GM [43].
Through characteristic and selective assimilation of prebiotics by subsequent fermentation, there is a production of SCFAs at high levels, having immunomodulation and metabolic effects on the host [44]. In this case, a reduction in the intestinal pH is also observed, creating an environment that competitively hinders the growth of pathogenic bacteria [45]. Some prebiotics prevent the adhesion of pathogenic microbiota to the GIT by mimicking an intestinal binding site [46] (Figure 1).
The application of prebiotics is well known in pharmaceuticals, and products for people with diabetes (as a natural sweetener) [47]. The large number of scientific data on prebiotics has focused on compounds associated with two major chemical groups: fructo-oligosaccharides and galacto-oligosaccharides [48]. They can be derived and/or extracted from food sources such as seeds, whole grains, legumes, chicory roots, Jerusalem artichokes, onions, garlic, and some vegetables [49], but in a recent study it was found that some aquatic plants (seaweeds and microalgae) contain prebiotics [50]. Prebiotics include a variety of forms such as fructo-oligosaccharides (FOS), galacto-oligosaccharides (GOS), human milk oligosaccharides (HMO), lactulose, lactosucrose, inulin, resistant starches (RS), arabinoxylans (AX), xylooligosaccharides (XOS), and pectin [24]. More attention has been given by researchers towards FOS as a prebiotic in improving human health [51].
3. Classification of Prebiotics
As mentioned above, there are many types of prebiotics that can be classified into different groups [52]. They differ in structure and can have a health benefit to the host through numerous different mechanisms [44]. Prebiotics also have the potential to modulate GM by selectively stimulating the growth of Bifidobacteria and Lactobacilli, by assimilation via beneficial GM and subsequent fermentation. In the fermentation process, these GM produce high levels of butyrate, isobutyrate, valerate, propionate, and acetate, which has various physiological functions in an organism [53]. The majority of prebiotics are mostly the subset of carbohydrate groups, more specifically, oligosaccharide carbohydrates. There are many relevant articles on oligosaccharide carbohydrates [54,55], but there are also few pieces of evidence showing that prebiotics are not only carbohydrates [56].
3.1. Inulin (Fructan)
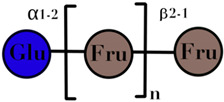
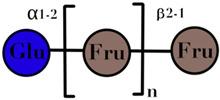
Inulin-type prebiotics are members of an immense group called “fructans”. Fructans constitute a group of compounds that confine all naturally occurring plant oligosaccharides and polysaccharides in which one or more fructosyl–fructose linkages form the majority of glycosidic bonds [57]. Hence, they are the primarily polymers of fructose units. Fructans can also be characterized by the DP, which refers to the number of repeated units in a polymer or oligomer chain [58]. The category of fructans consists of inulin and oligofructose (FOS) [59] (Table 1).
Table 1.
Summary of the structure and formula of prebiotics.
| Abbreviation | Chemical Composition | DP | Chemical Formula | References |
|---|---|---|---|---|
| Inulin | linear chain of fructose with β(2→1) linkages | 3–60 |
 n = up to 100 |
[59,60] |
| FOS | linear chain of fructose with β(2→1) linkages | <10 |
 n = 1–5 |
[59,60] |
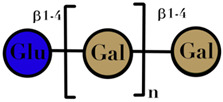
| GOS | Chain of galactosyl residues and a terminal glucose linked by β-(1–2), β-(1–3), β-(1–4), or β-(1–6) glycosidic bonds | 2–8 |
 n = 1–4 |
[60,103] |
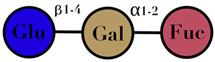
| HMO | composed of five monosaccharides: glucose, galactose, N-acetylglucosamine, fucose, and N-acetylneuraminic acid or sialic acid | <7 |
 2′fucosyllactose 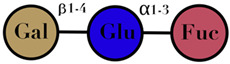 3′-fucosyllactose 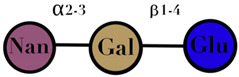 3′-sialyllactose 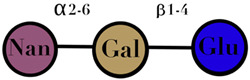 6′-sialyllactose 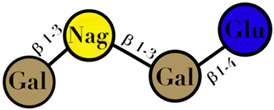 Lacto-N-tetraose 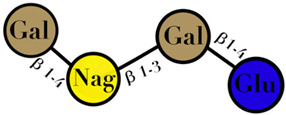 Lacto-N-neotetraose 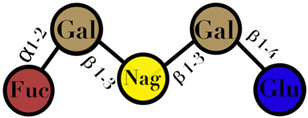 Lacto-N-fucopentaose I |
[104,105] |
| Lactulose | consisting of galactose and fructose moieties | - |
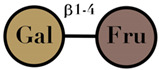
|
[106,107] |
| Lactosucrose | composed of galactose, fructose, and glucose monomers | - |
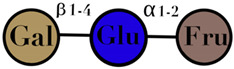
|
[108] |
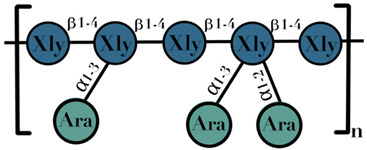
| AX | β-1,4-linked D-xylopyranoside units substituted with arabinose residues on the c(o)-2 or c(o)-3 position | 1–60 |
|
[109] |
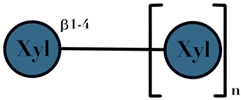
| XOS | xylose moeities linked by β-(1→4) glyosidic bonds | 2–4 |
 n = 2–4 |
[110,111,112] |
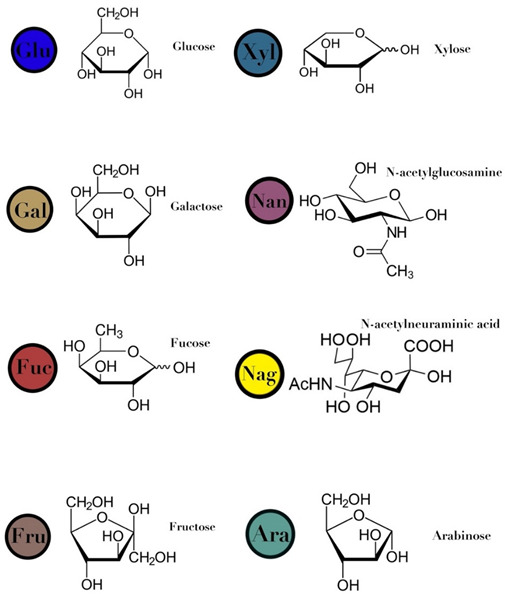
| Symbols used in Table 1: their meaning and chemical structure. |
|
|||
Inulin is a collective term that comprises all linear fructans with β (2→1) fructosyl–fructose glycosidic bonds [60]; this specific type of glycosidic bond gives inulin its distinctive physiological and structural properties. Inulin-type fructans resist enzymatic hydrolysis by small intestine digestive enzymes and human salivary enzymes because of the beta configuration bonds between fructose monomers [61]. Chemically, the linear chain of inulin is either an α-D-glucopyranosyl-[β-D-fructofuranosyl](n-1)-β-D-fructofuranoside (GpyFn) or αβ-D-fructopyranosyl-[β-D-fructofuranosyl](n-1)-β-D-fructofuranoside (FpyFn) [62].
3.2. Fructo-Oligosaccharides (Fructan)
Another type of fructans i.e., FOS, is a natural component that can be found in plants [63]. FOS are commercially prepared from chicory in a hydrolysis reaction using inulinase and may also be derived in an enzymatic synthetic reaction via the transfer of fructosyl units from sucrose molecules [64]. When presented structurally, FOS consist of a sucrose molecule linked by a chain of 3–30 fructosyl units. FOS are oligomeric linear fructans with β-(2–1) or β-(2–6) fructosyl-fructose linkages with the first monomer of the chain either being α-D- glucopyranosyl or β- D -fructopyranosyl residue [60]. The DP of inulin is up to 60 and the DP of FOS is less than 10 [65] (Table 1).
3.3. Galactooligosaccharides
GOS are the product of lactose extension and are included among non-digestible oligosaccharides. They are arranged in two subgroups: (i) with excess galactose at C3, C4 and C6; and (ii) manufactured from lactose through enzymatic trans-glycosylation [66]. The mixture of the product depends on the reaction conditions and the enzymes used. β-galactosidase of various origins, such as Aspergillus oryzae, Bacillus circulans, and Cryptococcus laurentii, is used for the industrial production of GOS [67]. The general constituents of this oligosaccharide are from tri- to penta-saccharide with β (1→6), β (1→3), and β (1→4) linkages. This category of GOS is known as trans-galacto-oligosaccharide [68]. Culture studies of Bifidobacteria and most of Lactobacilli and enterobacteria, including some streptococci-metabolized trans-oligosaccharide, with Bifidobacteria showed robust growth [69]. There are some GOS derived from the isomers of lactose, due to influential factors such as the source of the enzyme, temperature, pH, and substrate concentration. They are also considered as prebiotics [70] (Table 1).
3.4. Human Milk Oligosaccharides
HMO are complex and non-digestible carbohydrates, recently classified as prebiotic substances. They are present in high abundance in maternal breast milk (10–15 g/L) [57,71,72]. The length of the HMO chain can vary from 3 to 15 carbohydrate units and is synthesized in the mammary gland [73]. The HMO concentration in the lactating mother is higher during the early stages and gradually decreases over time [74,75,76]. Structurally, HMO are composed of five monosaccharides: glucose, galactose, N-acetylglucosamine, fucose, and N-acetylneuraminic acid or sialic acid [77,78,79]. They are synthesized from a lactose core (galactose-β (1→4) glucose) by glycosyl transferases in the lactocyte. Some HMO are branched with a fucose or sialic acid monosaccharide residue attached to the lactose core via α1–2/3/4 and α2–3/6 linkages, due to the action of fucosyltransferases and sialyltransferases, respectively [80,81]. Among its several types, less than 50 HMO have a representative abundance in human breast milk. HMO 2′-fucosyllactose has been identified as the most abundant HMO in breast milk [82]. Breast milk due to its high levels of 2′-fucosyllactose has shown advantages for the infant because of its efficiency to promote an early high Bifidobacteria-dominated GM [83]. Several experiments conducted on the supplementation of HMO documented beneficial effects on the overall health of an individual, which includes modification of the GM [78,81,84], effects on immune development [78,85,86], anti-adhesive antimicrobial effects [87], and brain development [88,89] (Table 1).
3.5. Glucose-Derived Oligosaccharides
An example of glucose-derived oligosaccharide is polydextrose (PDX), which is non-digestible and widely used in the food industry [90]. PDX is a randomly bonded glucose polymer with an average DP of 12, but ranging from 2 to 120. This molecule contains the combination of α- and β-linked 1→2, 1→3, 1→4, and 1→6 glycosidic linkages [91]. PDX has been acknowledged as a soluble fiber that has beneficial effects on gut health, satiety, and postprandial glycemia [92]. Daily intake of 4–12 g of PDX has been found to have a large improvement in physiological functions without showing any adverse effect [93].
3.6. Resistant Starches
The starch that is resistant to the upper gut digestion is termed as RS [94]. RS cannot be digested by human pancreatic amylase in the small intestine, reaching the colon, promoting health benefits by producing a high level of butyrate, suggesting it to be classified as a prebiotic [39,95]. RS consumption has been related to improving the diabetes condition by reducing postprandial glycemic and insulinemic responses, and is also associated with decreased levels of cholesterol and triglycerides [96].
3.7. Pectic Oligosaccharides
Pectic oligosaccharides (POS) originate from a polysaccharide, known as pectin, which is a structural element of intracellular regions and cell walls of the plants and is vastly present in fruits and vegetable waste materials [97]. Chemically, POS are based on the extension of rhamnose or galacturonic acid, and different types of sugars (galactose, xylose, and arabinose) or ferulic acid are linked to the side chains [98]. In humans, gastric juice and saliva are not capable of degrading pectin. Furthermore, digestive enzymes like trypsin, pepsin, and rennet cannot breakdown pectin in the small intestine [99]. It has been reported that pectin undergoes slow fermentation and exhibits prebiotic effects by producing SCFAs [100]. It has been shown that pectic oligosaccharide has the potential to show bifidogenic effects [101]. Experiments conducted on pectic oligosaccharides revealed health benefits that include antiobesity, anticancer, and antioxidant properties [102].
3.8. Lactulose
Lactulose is a synthetically produced non-digestible ketose disaccharide that consists of galactose and fructose linked by a bond resistant to lactase [113]. Lactulose is extracted from lactose (milk sugar), chemically known as 4-O-β-d-galactopyranosyl-d-fructose, and the enzyme used for the biocatalytic production is β-galactosidase [114]. It is used medically for the treatment of constipation [115]. The human small intestinal mucosa does not have the enzymes to breakdown lactulose, and hence it reaches the large bowel unchanged [116]. Lactulose is metabolized by colonic bacteria to monosaccharides and then to methane, volatile fatty acids, and hydrogen [117]. In human studies, the lactulose have a significantly modified GM by increasing Bifidobacterium, Lactobacillus, and Streptococcus, and having favorable health benefits [81,118] (Table 1).
3.9. Lactosucrose
Lactosucrose is also known as galactosylsucrose, lactosylfructoside, and galactosucrose, and is synthetically produced trisaccharide, which is composed of galactose, fructose, and glucose monomers [119]. Raffinose, an isomer of lactulose, has a potential bifidogenic effect [120]. Lactosucrose is used as a commercial food supplement in many healthy foods and beverages with the intention of altering gastrointestinal functions and improving health [121]. Lactosucrose has shown promising effects as a bifidogenic compound modulating immune functions [122,123] (Table 1).
3.10. Arabinoxylans
Arabinoxylans (AX) are predominant non-cellulosic polysaccharides of cell walls in plants. AX were first identified by Hoffman and Gortner in 1927, as viscous gum in wheat flour [124]. Their structural properties, heterogeneity, and recovery depend on their location, which is strongly influenced by the other components of the cell wall [125]. AX are called as “pentosans” as they consist of pentoses xylose and arabinose. Chemically, it is heteroxylan consisting of a backbone of β-1,4-linked D-xylopyranoside units substituted with arabinose residues on the c(o)-2 or c(o)-3 position [126]. The DP of AX is between 1 and 60 [109]. AX have the potential to show high technological importance. There are several biological studies that have been reported on the behalf of AX, including antioxidant activity [127], cholesterol-lowering agents [128], blood sugar modifiers [129], and immunity enhancers [130] (Table 1).
3.11. Xylooligosaccharides
Xylooligosaccharides (XOS) or xylan are to be considered as the second most abundant biopolymer in the plant kingdom. These are the sugar oligomers of β-1,4-linked xylose (a pentose sugar) found naturally in food sources such as honey, bamboo shoots, fruits, vegetables, and milk [131]. On the basis of substituted groups, xylan can be categorized into three classes: (i) glucuronoxylan, (ii) neutral arabinoxylan, and (iii) glucuronoarabinoxylan [132].The DP of the XOS used in commercial food products ranges from 2 to 10 [133]. The complete utilization of XOS is based on the activities of a number of enzymes, including β-xylosidase, α-glucuronides, and acetyl esterases released by different strains of GM, and produces SCFAs [134]. XOS have shown a bifidogenic effect [135], with the support of in-vivo animal studies, and offers modification in the composition and activity of the GM [136] (Table 1).
Excess consumption of prebiotics can promote severe discomfort in an individual, therefore, optimal intake is necessary [137] (Table 2).
Table 2.
Recommended intake of prebiotics.
4. Efficacy of Prebiotics on Gut Microbiota Composition: In Vivo and In Vitro Studies
The experiments conducted on the administration of prebiotics have shown selective changes in the GM composition. Different categories of prebiotics can stimulate the growth of various indigenous bacterial communities in the GM. Collective evidence from animal model trials, human studies, and in-vitro modeling systems has concluded that they affect the composition of GM, leading to proliferation in health-promoting organisms such as Bifidobacteria and Lactobacilli [146,147,148]. Prebiotics have ameliorative properties such as maintaining intestinal integrity and homeostasis, production of SCFAs, and regulation of gastrointestinal transit [115]. Indeed, it has been suggested that the use of prebiotics should have ameliorative properties on gastrointestinal diseases like irritable bowel disease, Chron’s disease, and ulcerative colitis [149]. Selective stimulation of GM growth and/or activity is potentially associated with health protection and well-being [24,150].
4.1. Inulin
Inulin is a non-digestible oligosaccharide that is fermented by the GM and has resistance to the degradation by the human digestive enzymes. It reaches the colon almost as an intact molecule and acts as a fermentable substrate for GM [62]. In vivo and in vitro studies on inulin concluded that it has selective stimulation of bacterial growth; this has been observed in numerous studies carried out either in defined pure culture fermentation or by using human feces [151,152,153]. Inulin supplementation for 19 days to a group of 10 elderly women with a dose beginning at 20 g/day from days 1 to 8 and gradually increasing to 40 g/day during days 9 to 19, showed a significant increase in Bifidobacteria that can be utilized during fermentation, and a decrease in the number of Enterococci and Enterobacteriaceae, while no statistically significant changes were observed in Bacteroides, Clostridia, or Faecalibacterium prausnitzii [154]. In another study conducted on 10 healthy volunteers with inulin supplementation of 8 g per day for 14 days, a significant increase in Bifidobacteria was shown. In this case, a number of Clostridia increased also, but the magnitude of Clostridia was one tenth of Bifidobacteria. These data supported a bifidogenic effect of inulin [155]. Importantly, inulin fermentation leads to the production of SCFAs. In an experiment conducted on rats cecum (colonic part of the GIT), it was demonstrated that inulin has significantly higher efficiency in producing SCFAs compared with other dietary fibers [156] (Table 3).
Table 3.
Prebiotic efficiency in modulating the GM.
| Prebiotics | Model | Strategy/Duration of Feeding | Dose Supplemented | Form | No. of Applications | Re-Calculated Dose * | Fecal Microbial Changes Relative to Control | Reference |
|---|---|---|---|---|---|---|---|---|
| Inulin | 17 elderly women (mean age = 76.4 years body weight not reported) |
8 days, (3 days adaptation) Feeding was continued for 8 days | 20 g/day and increased to 40 g/day |
Dissolved in drinking water | Once/day | 285.7 mg/kg/day and increased to 571.4 mg/kg/day |
significant ↑ in Bifidobacteria ↓ in Enterococci and Enterobacteriaceae |
[154] |
| 10 healthy volunteers (age = between 20 and 55 years Body weight not reported) |
14 days | 8 g/day | Dissolved in drinking water | Twice/day | 114.3 mg/kg/day | significant ↑ in Bifidobacteria ↑ in the number of Clostridia |
[155] | |
| Germ-free adult male Fischer rats (age = 10 weeks and body weight = 280 g) | 8 weeks | 1.84 g/day of the diet | Mixed with chow | During the day | 6.57 g/kg/day | significant ↑ in producing SCFAs | [156] | |
| FOS | Male Wistar rats (age = 2 months and body weight 403.2 ± 48.1 g) |
7 days | 8% of the diet | Mixed with chow | During the day | 3.4 g/kg/day | ↑ the bioavailability of nutritionally important minerals | [158] |
| FOS + GOS | 10 Male C57BL/6J mice (age 8 weeks old mice; mean body weight = 28 g) | 10 weeks | 0.3 –0.4 g/mouse/day | Dissolved in drinking water | During the day | 1.1–1.43 g/kg/day | ↑ Akkermansia abundance | [164] |
| GOS | 18 healthy human (age and body weight not indicated) | 3 weeks | 2.5 g/day 5 g/day, 10 g/day |
Administered in edible chews | Once/day | 35.7 mg/kg/day 71.4 mg/kg/day, 142 mg/kg/day |
significant ↑ in abundance of Bifidobacteria and Faecalibacterium prausnitzii, ↓ in Bacteroides |
[161] |
| Mud crab (age not reported and body weight 63.6 ± 8.8 g) |
24 h | 0.05 g/day | Dissolved in water | During the day | 786 mg/kg/day | ↑ Bacteroidetes | [163] | |
| PDX/FOS | 77 Children (age 5.8 ± 1.3; body weight not reported) | 2 weeks | 4.17 g PDX + 0.45 g FOS | Dissolved in drinking water | Once/day | PDX 200 mg/kg/day + FOS 22 mg/kg/day |
↑ in number of Bifidobacterium and Lactobacillus | [176] |
| PDX | 20 Healthy men (Age = 27.5 6 ± 4.33; body weight = 86.26 ± 13.48 kg) | 21 days | 21 g/day | Mixed in bar | Once/day | 243.4 mg/kg/day | ↑ in number of Faecalibacterium, Phascolarctobacterium, and Dialister | [175] |
| 15 Healthy volunteers (age = 18–50 body weight not reported) |
3 weeks | 8 g/day | Powder | Once/day | 243.5 mg/kg/day | ↑ Ruminococcus intestinalis, Clostridium clusters I, II and IV, significantly ↓ levels of Lactobacillus and Enterococcus group |
[177] | |
| RS | 6 Male C57BL/6J mice (18–20 month old and body weight not reported) | 8 weeks | 0.54 g/day | Mixed with chow | During the day | 18 g/kg/day | ↑ in number of Bacteroidetes, Bifidobacterium and Akkermansia species | [178] |
| Sprague-Dawley rats (age 6 weeks and body weight not reported) | 12 weeks | 27% of the diet | Mixed with chow | During the day | 18 g/kg/day | ↑ in SCFAs | [180] | |
| POS | Pigs’ fecal inoculum (age 4 years and the mean body weight 233.0 ± 10.02 kg) | 48 h | 9 g/of POS to 1 mL of inoculum | Mixed with the chow | - | - | ↑ in SCFAs | [183] |
| Lactulose | 12 healthy volunteers (age = (24 to 31 years and body weight not reported) | 4 weeks | 20 g/day | Mixed with chow | Twice/day | 285.7 mg/kg/day | ↑ in number of Bifidobacterium and Lactobacillus. | [186] |
| Lactosucrose | Red seabream Pagrus major (age and body weight not reported) |
9 months | 20 mg/kg/day | Mixed with chow | Once/day | 20 mg/kg/day | ↑ production of SCFAs | [192] |
| 8 Shepherd dogs (body weight = 22 to 32 kg; mean age = 13.5 months) | 2 weeks | 1.5 g/day | Mixed with chow | Twice/day | 55.6 mg/kg/day | ↓ in the levels of f Clostridium pefringms ↑ Bifidobacterium |
[122] | |
| 16 Broiler chickens (20–62 days and body weight not reported) |
62 days | 825 mg/day | Mixed with chow | During the day | 458 mg/kg/day | ↑ in the number of Bifidobacterium ↓ the number of Bacteriodaceae; Staphylococci; and total anaerobic bacteria, C. perfringens |
[198] | |
| 8 Cats (Mean agae + 7; body weight 3.5 kg) |
2 weeks | 50 mg of lactosucrose/day | Mixed with the chow | During the day | 14 mg/kg/day | ↑ in Lactobacilli and Bifidobacterium ↓ in Clostridium perfringens, clostridia, Spirochaetaceae, and Enterobacteriaceae |
[185] | |
| AX | 10 human children (mean age, 3 years, 7 months body weight not reported) (in vitro) |
48 h | 10 g/liter | Dissolved in drinking water | - | -- | ↑ in number of Lactobacillus | [196] |
| XOS | 12 healthy adult women (mean age for women = 33.6 years and body weight not reported) and 11 healthy men (mean age = 30.1 and body weight not reported) | 8 weeks | 1.4 g/day or 2.8 g/day | Capsule | Once/day | 20 or 40 mg/kg/day | ↑ Bacteroides fragilis, ↑ Bifidobacterium | [199] |
| 13 elderly human (body weight = 58.6 ± 10.1 kg body weight not reported) |
3 weeks | 4 g/day | Mixed with chow | Once/day | 68.3 mg/kg/day | ↑ in number of Bifidobacterium species | [200] |
↑—Increase, ↓—Decrease, * unless indicated, the average adult human weight was estimated as 70 kg and the average rat weight was estimated to be 280 g.
4.2. FOS
FOS have great potential as ingredients due to their prebiotic activity and low caloric value. Gibson and Roberfroid [23] showed the bifidogenic characteristics of FOS using 15 g per day as dietary supplementation. The GM was modulated and there was a significant decrease in the number of Bacteroides, Fusobacterium, and Clostridium. Therefore, it was concluded that FOS is better utilized by Bifidobacteria, and, on the other hand, they can cause unfavorable changes for harmful bacteria in the GIT [23].
It was verified that the addition of NeosugarR (a trade name for fructooligosaccharide) to the human diet, i.e., 15 g per day, can cause a 10-fold increase in the population of Bifidobacteria in the large intestine [138]. In addition to its bifidogenic property, the regular and adequate intake of FOS has beneficial effects in the case of disorders associated with obesity, diarrhea, osteoporosis, atherosclerotic, gastrointestinal disorders, cardiovascular, and T2DM diseases [157]. The fermentation of FOS by GM generates SCFAs and organic acids that decrease luminal pH, thereby enhancing the bioavailability of nutritionally important minerals [158]. It was also found that a diet supplemented with FOS promotes the production of butyrate, which influences lipid metabolism in humans [159] (Table 3).
4.3. GOS
GOS are a type of non-digestible fiber with prebiotic activity [133], which has also been demonstrated by a dynamic in-vitro colon model and the 13C labeling technique with GOS consumption. The results showed an increase in Bifidobacterium longum, B. bifidum, B. catenulatum, Lactobacillus gasseri, and L. salivarius, but changes in numbers of Enterobacteriaceae (a family of Gram-negative bacteria that includes some harmless symbionts) and several familiar pathogens, such as Salmonella, Yersinia pestis, Klebsiella, Escherichia coli, and Shigella, were rather negligible [160]. In another study, the prebiotic activity of GOS was analyzed by pyrosequencing of fecal samples from healthy human volunteers with GOS administration. The data obtained showed a statistically significant increase in Bifidobacteria and Faecalibacterium prausnitzii, and a decrease in Bacteroides [161]. It was also concluded that 90% of GOS resist digestion in the upper GIT and then enter the colon, which then get intact to the tract and act as fermentation substrates for the resident microbiota [162] (Table 3). An in-vitro study showed that the fermentation of GOS by GM generates SCFAs and organic acids that decrease luminal pH, thereby enhancing the bioavailability of nutritionally important minerals [163]. Interestingly, GOS administration showed anxiolytic effects in both animals [164] and humans [165] (Table 3).
4.4. HMO
One of the multifarious functions of HMO is that they act as prebiotics and stimulate the colonization of beneficial GM [166]. In vitro studies provided strong evidence that HMO promotes the growth of selective Bifidobacteria [78]. Bifidobacterium longum subsp. infantis proliferates well on 2′-FL, as the sole source of carbohydrates [81,84,167,168]. These Bifidobacterium longum subsp. infantis produce SCFAs, which create an environment that favors the growth of commensal bacteria and prevents the adhesion of pathogenic bacteria [169]. Some structures of HMO are similar to the intestinal epithelial cell surface glycan receptors, which serve as decoy receptors to prevent pathogen binding and increase pathogen removal [78]. A study on HMO supplementation suggested that breast-fed infants have a higher number of Bifidobacteria compared to the formula-fed infants [170]. In a human study, investigation into the interaction between Bifidobacteria and Eubacterium hallii demonstrated that E. hallii consume acetate, lactate, and 1,2-propanediol (which are the products formed by the fermentation of HMOs by Bifidobacteria) and eventually lead to the production of butyrate and propionate [171] (Table 3). On the other hand, the study conducted on bioengineered 2′-FL showed inhibition of the adhesion of Campylobacter jejuni, Salmonella enterica, E.coli, and Pseudomonas aeruginosa to an intestinal human cell line [172]. Research on HMO, specifically 2′-FL, has shown that it is even more potent than standard commercial prebiotics, such as FOS, and has many different functions, including immune, GM, and cognition benefits [173] (Table 3).
4.5. PDX
In vitro studies have indicated that PDX has all the characteristics to be a prebiotic [174,175]. It has been shown that daily intake can beneficially modify the composition and activity of GM. In a study in humans, PDX favored intestinal function and improved the ease of bowel movement. Furthermore, it inhibited the absorption of glucose in the small intestine and the fermentation for the production of SCFAs in the large intestine favoring the reduction of gut pH [176]. Supplementation with PDX in healthy humans with a dose of 8 g per day for 3 weeks showed a significant increase in the number of Ruminococcus intestinalis, the main producer of butyrate, and slow fermentation of PDX in the colon was observed [177]. Another study carried out in healthy adult males with 21 g of PDX supplementation per day significantly suppressed the number of phylum Firmicutes and significantly increased the number of bacteroidetes when compared to the control group [174]. These data concluded that PDX supplementation had a positive impact on the bacterial composition of GM (Table 3).
4.6. RS
A number of studies demonstrated that RS is capable of modifying the GM composition towards the heath benefit of the host. An experiment carried out in mice for 8 weeks showed that mice fed with diets containing high amylose RS2 (one of the types of RS) were colonized by higher levels of Bifidobacterium, Akkermansia, and Allobactum [178]. The nutritional intervention study revealed that RS, when supplemented in the diet, can induce a 10-fold increase of gut Bifidobacteria [179]. On the other hand, one of the byproducts of RS is SCFAs. A study carried out on rats for 12 weeks treated with two concentrations of RS (0 and 27% weight of diet) showed an increase in propionate, butyrate, and acetate [180] (Table 3).
4.7. POS
POS is a new class of prebiotics that derives SCFAs from the GM fermentation [181]. In an in vitro study, the POS from the citrus peel and sugar beet pulp were fermented by the human fecal samples, leading to an increase in the bacterial population of eight different groups. POS from sugar beet showed the highest bifidogenic effect and utmost SCFAs concentration. On the other hand, the POS from citrus peel showed an increase in the population of Lactobacillus [182]. In a recent study, it was concluded that the concentration of SCFAs was higher in the POS supplementation, when compared to FOS [183] (Table 3).
4.8. Lactulose
An investigation conducted on lactulose degradation determined that human and calf β-galactosidases do not degrade it [184]. An in-vitro study performed using fecal samples on agars, and an analysis of enzymes produced and putrefactive compounds of lactulose fermentation, concluded a selective and significant increase in Bifidobacteria, decreasing the abundance of streptococci, bacteroides, C. perfringens, and Lactobacilli [185]. Studies carried out on humans demonstrated that lactulose selectively and significantly modifies GM by increasing Lactobacillus, Bifidobacterium, and Streptococcus [186] (Table 3).
4.9. Lactosucrose
Strong evidence has been observed in the administration of lactosucrose, selectively promoting the number of Bifidobacteria in in-vitro and in-vivo studies on animals and humans [187,188,189]. Lactosucrose fermentation was evaluated using Bifidobacterium, Lactobacillus, and Streptococcus probiotic strains in the in-vitro study, and the results led to the growth of four bacterial strains: Lactobacillus casei, Lactobacillus reuteri, Lactobacillus acidophilus, and Streptococcus salivarius [187]. Animal studies have shown a significant increase in Lactobacilli and Bifidobacteria, while restraining the levels of pathogens, such as Clostridium perfringens, Staphylococci, and Bacteroidaceae, after the consumption of lactosucrose [122,185]. Lactosucrose fermentation by the GM produces SCFAs and shows a consequent reduction in the pH of fecal contents [190,191]. An in-vitro study on different fish species such as Pagrus major, Cyprinus carpio L., and Oncorhynchus mykiss showed that the lactosucrose fermentation results in the production of SCFAs and gases, concluding that the lactosucrose can also be fermented in herbivorous, omnivorous, and carnivorous fishes [192,193,194] (Table 3).
4.10. AX
AX are not digested by the enzymes produced by the GIT, thus these provide the carbon source for the GM that inhibits the large bowel [195]. Many experiments have been performed on the regular supplementation of AX, resulting in an enhancement in the proliferation of the growth of health-promoting bacteria. In-vitro studies of AX, implemented in anaerobic batch cultures inoculated with human feces, demonstrated that fermentation of wheat endosperm AX resulted in the production of acetate, propionate, and butyrate [196]. In the in-vitro digestibility test carried out on pigs, it was established that only 15% of the ingested AX is recovered in the feces, while the major fraction of AX is fermented in the cecum, which represents the high fermentability of AX [197] (Table 3).
4.11. XOS
Animal studies have furnished evidence that oral administration of XOS remarkably increases fecal weight, bone properties, fecal moisture, and number of Bifidobacteria, with a parallel increase in SCFAs production in mice [201], rats [202], and humans (elderly) [200]. A recent study on a healthy human adult demonstrated that XOS intake increases Bifidobacterium counts without affecting the number of Lactobacillus [199]. The potential of Bifidobacteria to metabolize XOS is based on the activity of their xylan-degrading enzyme systems. Human study on the prebiotic XOS and their effects on modulating the GM in vivo is limited, particularly regarding the efficiency (Table 3).
Prebiotics are also able to remodulate the composition of the GM. Compared to a different category of prebiotics, only the fructans (inulin and FOS), GOS, and lactulose had highly selective effects on human GM modification [203]. As mentioned before, fermentation products of prebiotics such as SCFAs also have modulatory effects on the gut pH [204]. The pH alteration can have an influence on the population of acid-sensitive species, such as Bacteroides, and promote butyrate formation by Firmicutes [205].
5. Prebiotics for the Treatment of Obesity and Diabetes
Globally, the population of diabetes patient is increasing, imposing a great social and economic burden on public health [206,207]. T2DM is a chronic metabolic syndrome of abnormal lipid and glucose metabolism that leads to neuropathy, retinopathy, leg ulcers, and gangrene [208]. The factors that could have an impact on T2DM development are obesity, genetics, smoking, age, hypertension, and sedentary lifestyle [207]. In recent studies, it has been proposed that the remolding of the GM composition from obesity could lead to the pathogenesis of T2DM [209,210,211,212,213].
As mentioned above, the two dominant bacteria groups in human GIT are Bacteroidetes and Firmicutes [209]. A link between obesity and GM composition has been reported in humans, showing an increase in the number of Firmicutes and a decrease in the diversity of Bacteroidetes [213] (Figure 2).
Figure 2.

An overview of the improvement in the health of obese/T2DM patients treated by modulating their GM using prebiotics supplementation in a regular diet. Administration of prebiotics has the potential to modulate GM composition in patients suffering from T2DM and obesity and can be used as a therapeutic approach to cure the adverse effects of metabolic diseases. The daily intake of prebiotics in a designed diet has a major influence on GM by decreasing gut permeability, bacterial translocation, and reducing LPS-induced inflammation. However, this diet increases SCFAs and bifidogenecity in the gut, leading to lower TC levels, lipogenesis, LDL triglycerides, and adiposity, eventually resulting in lower risk of cardiovascular diseases.
Prebiotics have gained a considerable place in the management of obesity and diabetes due to their ability to modulate GM composition, thereby affecting the status of GIT and exerting anti-diabetic effects [214,215]. As prebiotics consist of different forms, their supplementation can be considered as a dietary therapy for the prevention and treatment of T2DM [216], and also in the fight against obesity by affecting food intake and appetite and metabolic activities [10] (Figure 2).
FOS have numerous desirable characteristics such as low calories, safety for diabetics, no carcinogenicity, and bifidus-stimulating functionality [65]. Due to these properties, FOS are considered a functional food ingredient that improves health status [217]. Increasing studies demonstrated the functional properties of FOS including the reduction of blood glucose levels, cholesterol levels, and lowering of blood pressure [218,219,220] (Table 4).
Table 4.
Effect of different prebiotics on the treatment of obesity and diabetes in animal and human studies.
| Prebiotic Used | Tested Species | Dose | Re-Calculated Dose | Period | Outcomes | Reference |
|---|---|---|---|---|---|---|
| FOS | 27 women with Type-2 diabetes, age = 20–65 years; 76.0 (12.2) | 10 g/day | 131.6 mg/kg/day | 8 weeks |
|
[220] |
| FOS | 10-week-old C57BL/6J mice, body weight not reported | 0.3 g/mouse/day | 8 weeks |
|
[226] | |
| GOS | 6 rats alloxan-induced diabetic rats, 6 weeks old; Average weight = 90 g |
100 g/kg of diet | 1.11 g/kg of diet | 42 days |
|
[227] |
| Human (women with overweight age 18–65 years and body weight not reported) |
5.5 g/day of GOS | 5.5 g/kg/day | 12 weeks |
|
[228] | |
| PDX | Rats (Wistar rats age not reported and body weight 43.0 ± 4.5 g) |
5 g/100 g diet | 5 g/100 g diet | 60 days |
|
[229] |
| RS | Human (over weight and obese adults—11 men and 22 women age 18–69 years and body weight not reported) |
15 g/kg/day of HAM-RS2 v. 30 g/kg/day HAM-RS21 | 15 g/kg/day of HAM-RS2 v. 30 g/kg/day HAM-RS21 | 4 weeks |
|
[230] |
| Human (diabetic adults age = 55 ± 2.4 years body weight not reported) |
40 g/day of | 571.4 mg/kg/day | 12 weeks |
|
[231] | |
| Lactulose | Human (patients with obesity age and body weight not reported) |
8.2 g/day | - | 2 days |
|
[232] |
| AX | Rats (wild type rats with high cholesterol diet age 7 weeks body weight not reported) |
8% corn arabinoxylan | 5.8 g/kg/day | 20 days |
|
[233] |
| Human (T2DM); mean age = 55 years and body weight not reported) |
49.2 g/day | 702.9 mg/kg/day | 35 days |
|
[234] | |
| XOS | Rats (Male Wistar rats treated with streptozotocin to induce diabetes, age = 8 weeks; body weight = 180 ± 8 g) |
0.325 g/day | 1.81 mg/kg/day | 5 weeks |
|
[235] |
↑—Increase, ↓—Decrease.
Meanwhile, inulin as a prebiotic has shown mixed results on the glycemic scale [221,222]. A study carried out on 54 middle-aged (between 35 and 65 years) healthy adults (men and women) as a double-blind, randomized, placebo-controlled parallel groups with 10 g of inulin supplementation for 8 weeks did not show any significant changes in the body weight [223]. A decrease in plasma insulin level was observed after 4 weeks of treatment and remained lower up to the 8th week, along with lower plasma triglyceride concentrations. Total cholesterol (TC) was lower in the inulin-supplemented group when compared to the placebo group. The study concluded that inulin supplementation may influence the degradation of triglyceride-rich lipoprotein particles [223]. In human trials conducted on obese women treated with inulin, greater proportions of Bifidobacterium and Faecalibacterium were observed, an effect that coincided with reduced fat mass and serum lipopolysaccharide [224]. The important role of Bifidobacterium in the fight against obesity has recently been demonstrated by Bifidobacterium longum both in preclinical obesity models and in humans [225].
XOS studies indicated that they have the potential to reduce serum cholesterol and triglycerides levels, which are the main risk factors for diabetes. The administration of XOS to wild-type rats for 28 days showed a reduction in LDL levels, TC in serum, triglycerides, and body weight [236] (Table 4).
The consumption of RS improved insulin sensitivity in subjects with metabolic syndrome and appears to have a favorable effect on insulin sensitivity [231] (Table 4).
In summary, prebiotics show efficiency not only in modulating or restructuring and stabilizing the host microbiome, but also in the regulation of many mechanisms associated with the development and metabolic consequences of obesity. Furthermore, prebiotics should be enriched in popular foods, increasing the chances of consistent consumption and improving overall health. At least, dietary prebiotic supplementation represents a safe, well-tolerated, and inexpensive therapeutic approach and should be considered as a potential therapy for the treatment and prevention of T2DM and obesity (Figure 2).
6. Conclusions and Future Perspectives
Recent studies have proven that increased inflammatory state (as seen in obesity and diabetes) has a paramount influence on glucose metabolism, and eubiosis ensures appropriate immune responses. This implies that the implementation of appropriate GM modulatory strategies could be a new and promising therapy against metabolic diseases. Meanwhile, the appropriate dosage, duration of treatment, and long-term effects of the intervention of different prebiotics remain unknown. For this reason, more clinical trials are needed before prebiotics can be rationally suggested for the prevention and/or treatment of obesity and diabetes.
Although in-vivo and in-vitro studies conducted on animals and humans revealed that many prebiotics increase the growth of Bifidobacterium spp., and Lactobacillus spp. and cause a diverse change in the Bacteroidetes and Firmicutes phyla, it is still not fully understood how these carbohydrates interact with the GM with their widely diversified structures. Further research is required to reveal the mechanisms of these carbohydrates’ structures on the GM and the host. In addition, it is well established that GM ferments the prebiotics, leading to the formation of SCFAs and acidification of the colonic contents. These by-products formed by the fermentation process play an extensive role in maintaining the host’s health and ameliorate diseases. Despite advances in our understanding of prebiotics, there remain numerous knowledge gaps concerning the SCFAs molecular signaling mechanisms and their association with prebiotic chemical composition and structural conformations, along with their modulatory effects at the genetic, cellular, organelle, and systemic levels.
Meanwhile, the application of systems biology coupled with bioinformatics could be a powerful strategy to unveil mechanistic insights into the action of prebiotics on the gut microorganisms and lead to an understanding of how these compounds (and their metabolites) alter both microbial and host metabolic functions at the molecular level. These insights and population-based studies could uncover new strategies to improve dietary relevance and clinical efficacy.
Acknowledgments
We thank Virginija Bukelskienė for the correction of the first draft of the manuscript.
Abbreviations
GIT—gastrointestinal tract, GM—gut microbiota, SCFAs—short-chain fatty acids, DP—degree of polymerization, FOS—fructo-oligosaccharides, GOS—galacto-oligosaccharides, HMO—human milk oligosaccharide, PDX—polydextrose, RS—resistant starch, POS—pectin oligosaccharides, AX—arabinoxylans, XOS—Xylooligosaccharide, T2DM—Type-2 diabetes mellitus, TC—total cholesterol, LDL—low-density lipoprotein, HDL—high-density lipoprotein.
Author Contributions
Conceptualization, A.M.; investigation, A.M.; writing—original draft preparation, A.M., E.B.-M.D., D.B. and A.B.; writing—review and editing, A.M., A.B., E.B.-M.D. and D.B.; visualization, A.M.; supervision, A.B.; project administration, A.B. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Funding Statement
This project received funding from the European Regional Development Fund (project No. 01.2.2-LMT-K-718-02-0014) under grant agreement with the Research Council of Lithuania (LMT LT).
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Saeedi P., Petersohn I., Salpea P., Malanda B., Karuranga S., Unwin N., Colagiuri S., Guariguata L., Motala A.A., Ogurtsova K., et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9(th) edition. Diabetes Res. Clin. Pract. 2019;157:107843. doi: 10.1016/j.diabres.2019.107843. [DOI] [PubMed] [Google Scholar]
- 2.Morigny P., Houssier M., Mouisel E., Langin D. Adipocyte lipolysis and insulin resistance. Biochimie. 2016;125:259–266. doi: 10.1016/j.biochi.2015.10.024. [DOI] [PubMed] [Google Scholar]
- 3.Pascale A., Marchesi N., Govoni S., Coppola A., Gazzaruso C. The role of gut microbiota in obesity, diabetes mellitus, and effect of metformin: New insights into old diseases. Curr. Opin. Pharmacol. 2019;49:1–5. doi: 10.1016/j.coph.2019.03.011. [DOI] [PubMed] [Google Scholar]
- 4.Arnoriaga-Rodríguez M., Mayneris-Perxachs J., Contreras-Rodríguez O., Burokas A., Ortega-Sanchez J.-A., Blasco G., Coll C., Biarnés C., Castells-Nobau A., Puig J., et al. Obesity-associated deficits in inhibitory control are phenocopied to mice through gut microbiota changes in one-carbon and aromatic amino acids metabolic pathways. Gut. 2021;70:2283–2296. doi: 10.1136/gutjnl-2020-323371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Arnoriaga-Rodríguez M., Mayneris-Perxachs J., Burokas A., Contreras-Rodríguez O., Blasco G., Coll C., Biarnés C., Miranda-Olivos R., Latorre J., Moreno-Navarrete J.-M., et al. Obesity Impairs Short-Term and Working Memory through Gut Microbial Metabolism of Aromatic Amino Acids. Cell Metab. 2020;32:548–560.e7. doi: 10.1016/j.cmet.2020.09.002. [DOI] [PubMed] [Google Scholar]
- 6.Arnoriaga-Rodríguez M., Mayneris-Perxachs J., Burokas A., Pérez-Brocal V., Moya A., Portero-Otin M., Ricart W., Maldonado R., Fernández-Real J.-M. Gut bacterial ClpB-like gene function is associated with decreased body weight and a characteristic microbiota profile. Microbiome. 2020;8:59. doi: 10.1186/s40168-020-00837-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Milani C., Duranti S., Bottacini E., Casey E., Turroni F., Mahony J., Belzer C., Montes A.S., Mancabelli L., Lugli G.A., et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017;81:1–67. doi: 10.1128/MMBR.00036-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Adak A., Khan M.R. An insight into gut microbiota and its functionalities. Cell. Mol. Life Sci. 2019;76:473–493. doi: 10.1007/s00018-018-2943-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Avershina E., Lundgård K., Sekelja M., Dotterud C., Storrø O., Øien T., Johnsen R., Rudi K. Transition from infant- to adult-like gut microbiota. Environ. Microbiol. 2016;18:2226–2236. doi: 10.1111/1462-2920.13248. [DOI] [PubMed] [Google Scholar]
- 10.Cerdó T., García-Santos J.A., Bermúdez M.G., Campoy C. The role of probiotics and prebiotics in the prevention and treatment of obesity. Nutrients. 2019;11:635. doi: 10.3390/nu11030635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Peterson J., Garges S., Giovanni M., McInnes P., Wang L., Schloss J.A., Bonazzi V., McEwen J.E., Wetterstrand K.A., Deal C., et al. The NIH Human Microbiome Project. Genome Res. 2009;19:2317–2323. doi: 10.1101/gr.096651.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rowland I., Gibson G., Heinken A., Scott K., Swann J., Thiele I., Tuohy K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018;57:1–24. doi: 10.1007/s00394-017-1445-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dominguez-Bello M.G., Godoy-Vitorino F., Knight R., Blaser M.J. Role of the microbiome in human development. Gut. 2019;68:1108–1114. doi: 10.1136/gutjnl-2018-317503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Megur A., Baltriukienė D., Bukelskienė V., Burokas A. The microbiota–gut–brain axis and Alzheimer’s disease: Neuroinflammation is to blame? Nutrients. 2021;13:37. doi: 10.3390/nu13010037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sommer F., Bäckhed F. The gut microbiota-masters of host development and physiology. Nat. Rev. Microbiol. 2013;11:227–238. doi: 10.1038/nrmicro2974. [DOI] [PubMed] [Google Scholar]
- 16.Moschen A.R., Wieser V., Tilg H. Dietary factors: Major regulators of the Gut’s microbiota. Gut Liver. 2012;6:411–416. doi: 10.5009/gnl.2012.6.4.411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Turnbaugh P.J., Ley R.E., Hamady M., Fraser-Liggett C.M., Knight R., Gordon J.I. The Human Microbiome Project. Nature. 2007;449:804–810. doi: 10.1038/nature06244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Marchesi J.R., Adams D.H., Fava F., Hermes G.D.A., Hirschfield G.M., Hold G., Quraishi M.N., Kinross J., Smidt H., Tuohy K.M., et al. The gut microbiota and host health: A new clinical frontier. Gut. 2016;65:330–339. doi: 10.1136/gutjnl-2015-309990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mayneris-Perxachs J., Arnoriaga-Rodríguez M., Luque-Córdoba D., Priego-Capote F., Pérez-Brocal V., Moya A., Burokas A., Maldonado R., Fernández-Real J.-M. Gut microbiota steroid sexual dimorphism and its impact on gonadal steroids: Influences of obesity and menopausal status. Microbiome. 2020;8:136. doi: 10.1186/s40168-020-00913-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.De Angelis M., Garruti G., Minervini F., Bonfrate L., Portincasa P., Gobbetti M. The Food-gut Human Axis: The Effects of Diet on Gut Microbiota and Metabolome. Curr. Med. Chem. 2019;26:3567–3583. doi: 10.2174/0929867324666170428103848. [DOI] [PubMed] [Google Scholar]
- 21.Janssen A.W.F., Kersten S. The role of the gut microbiota in metabolic health. FASEB J. 2015;29:3111–3123. doi: 10.1096/fj.14-269514. [DOI] [PubMed] [Google Scholar]
- 22.Slavin J. Fiber and prebiotics: Mechanisms and health benefits. Nutrients. 2013;5:1417–1435. doi: 10.3390/nu5041417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gibson G.R., Roberfroid M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995;125:1401–1412. doi: 10.1093/jn/125.6.1401. [DOI] [PubMed] [Google Scholar]
- 24.Davani-Davari D., Negahdaripour M., Karimzadeh I., Seifan M., Mohkam M., Masoumi S.J., Berenjian A., Ghasemi Y. Prebiotics: Definition, types, sources, mechanisms, and clinical applications. Foods. 2019;8:92. doi: 10.3390/foods8030092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Vallianou N., Stratigou T., Christodoulatos G.S., Tsigalou C., Dalamaga M. Probiotics, Prebiotics, Synbiotics, Postbiotics, and Obesity: Current Evidence, Controversies, and Perspectives. Curr. Obes. Rep. 2020;9:179–192. doi: 10.1007/s13679-020-00379-w. [DOI] [PubMed] [Google Scholar]
- 26.Da Silva T.F., Casarotti S.N., de Oliveira G.L.V., Penna A.L.B. The impact of probiotics, prebiotics, and synbiotics on the biochemical, clinical, and immunological markers, as well as on the gut microbiota of obese hosts. Crit. Rev. Food Sci. Nutr. 2021;61:337–355. doi: 10.1080/10408398.2020.1733483. [DOI] [PubMed] [Google Scholar]
- 27.Salminen S., Collado M.C., Endo A., Hill C., Lebeer S., Quigley E.M.M., Sanders M.E., Shamir R., Swann J.R., Szajewska H., et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021;18:649–667. doi: 10.1038/s41575-021-00440-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Holscher H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes. 2017;8:172–184. doi: 10.1080/19490976.2017.1290756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Pluta R., Ułamek-Kozioł M., Januszewski S., Czuczwar S.J. Gut microbiota and pro/prebiotics in Alzheimer’s disease. Aging. 2020;12:5539–5550. doi: 10.18632/aging.102930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Shokryazdan P., Faseleh Jahromi M., Navidshad B., Liang J.B. Effects of prebiotics on immune system and cytokine expression. Med. Microbiol. Immunol. 2017;206:1–9. doi: 10.1007/s00430-016-0481-y. [DOI] [PubMed] [Google Scholar]
- 31.Macfarlane G.T., Macfarlane S. Fermentation in the human large intestine: Its physiologic consequences and the potential contribution of prebiotics. J. Clin. Gastroenterol. 2011;45:120–127. doi: 10.1097/MCG.0b013e31822fecfe. [DOI] [PubMed] [Google Scholar]
- 32.Venegas D.P., De La Fuente M.K., Landskron G., González M.J., Quera R., Dijkstra G., Harmsen H.J.M., Faber K.N., Hermoso M.A. Short chain fatty acids (SCFAs)mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019;10:277. doi: 10.3389/fimmu.2019.00277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Peng L., Li Z.-R., Green R.S., Holzman I.R., Lin J. Butyrate enhances the intestinal barrier by facilitating tight junction assembly via activation of AMP-activated protein kinase in Caco-2 cell monolayers. J. Nutr. 2009;139:1619–1625. doi: 10.3945/jn.109.104638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Gaudier E., Rival M., Buisine M.-P., Robineau I., Hoebler C. Butyrate enemas upregulate Muc genes expression but decrease adherent mucus thickness in mice colon. Physiol. Res. 2009;58:111–119. doi: 10.33549/physiolres.931271. [DOI] [PubMed] [Google Scholar]
- 35.O’Keefe S.J.D. Diet, microorganisms and their metabolites, and colon cancer. Nat. Rev. Gastroenterol. Hepatol. 2016;13:691–706. doi: 10.1038/nrgastro.2016.165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wan M.L.Y., Ling K.H., El-Nezami H., Wang M.F. Influence of functional food components on gut health. Crit. Rev. Food Sci. Nutr. 2019;59:1927–1936. doi: 10.1080/10408398.2018.1433629. [DOI] [PubMed] [Google Scholar]
- 37.Bengmark S. Colonic food: Pre- and probiotics. Am. J. Gastroenterol. 2000;95:9–11. doi: 10.1016/S0002-9270(99)00807-2. [DOI] [PubMed] [Google Scholar]
- 38.Zdunczyk Z. Physiological effect of low digestible oligosaccharides in diets for animals and humans. Pol. J. Food Nutr. Sci. 2004;13:115–130. [Google Scholar]
- 39.Leeman A., Sjöö M., Eliasson A.-C., Björck I. Resistant starch formation in temperature treated potato starches varying in amylose/amylopectin ratio. Carbohydr. Polym. 2006;65:306–313. doi: 10.1016/j.carbpol.2006.01.019. [DOI] [Google Scholar]
- 40.Dai F.J., Chau C.F. Classification and regulatory perspectives of dietary fiber. J. Food Drug Anal. 2017;25:37–42. doi: 10.1016/j.jfda.2016.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Howlett J.F., Betteridge V.A., Champ M., Craig S.A.S., Meheust A., Jones J.M. The definition of dietary fiber—Discussions at the Ninth Vahouny Fiber Symposium: Building scientific agreement. Food Nutr. Res. 2010;54:1–5. doi: 10.3402/fnr.v54i0.5750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Slizewska K., Chlebicz-Wójcik A. The in vitro analysis of prebiotics to be used as a component of a synbiotic preparation. Nutrients. 2020;12:1272. doi: 10.3390/nu12051272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Mohanty D.P., Mohapatra S., Misra S., Sahu P.S. Milk derived bioactive peptides and their impact on human health—A review. Saudi J. Biol. Sci. 2016;23:577–583. doi: 10.1016/j.sjbs.2015.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bindels L.B., Delzenne N.M., Cani P.D., Walter J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015;12:303–310. doi: 10.1038/nrgastro.2015.47. [DOI] [PubMed] [Google Scholar]
- 45.Markowiak P., Śliżewska K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients. 2017;9:1021. doi: 10.3390/nu9091021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Simpson H.L., Campbell B.J. Review article: Dietary fibre-microbiota interactions. Aliment. Pharmacol. Ther. 2015;42:158–179. doi: 10.1111/apt.13248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hanau S., Almugadam S.H., Sapienza E., Cacciari B., Manfrinato M.C., Trentini A., Kennedy J.F. Schematic overview of oligosaccharides, with survey on their major physiological effects and a focus on milk ones. Carbohydr. Polym. Technol. Appl. 2020;1:100013. doi: 10.1016/j.carpta.2020.100013. [DOI] [Google Scholar]
- 48.Swanson K.S., de Vos W.M., Martens E.C., Gilbert J.A., Menon R.S., Soto-Vaca A., Hautvast J., Meyer P.D., Borewicz K., Vaughan E.E., et al. Effect of fructans, prebiotics and fibres on the human gut microbiome assessed by 16S rRNA-based approaches: A review. Benef. Microbes. 2020;11:101–129. doi: 10.3920/BM2019.0082. [DOI] [PubMed] [Google Scholar]
- 49.Singla V., Chakkaravarthi S. Applications of prebiotics in food industry: A review. Food Sci. Technol. Int. 2017;23:649–667. doi: 10.1177/1082013217721769. [DOI] [PubMed] [Google Scholar]
- 50.De Jesus Raposo M.F., de Morais A.M.M.B., de Morais R.M.S.C. Emergent Sources of Prebiotics: Seaweeds and Microalgae. Mar. Drugs. 2016;14:27. doi: 10.3390/md14020027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Flesch A.G.T., Poziomyck A.K., Damin D.C. The therapeutic use of symbiotics. Braz. Arch. Dig. Surg. 2014;27:206–209. doi: 10.1590/S0102-67202014000300012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lordan C., Thapa D., Ross R.P., Cotter P.D. Potential for enriching next-generation health-promoting gut bacteria through prebiotics and other dietary components. Gut Microbes. 2020;11:1–20. doi: 10.1080/19490976.2019.1613124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Den Besten G., Van Eunen K., Groen A.K., Venema K., Reijngoud D.J., Bakker B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013;54:2325–2340. doi: 10.1194/jlr.R036012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Guarino M.P.L., Altomare A., Emerenziani S., Di Rosa C., Ribolsi M., Balestrieri P., Iovino P., Rocchi G., Cicala M. Mechanisms of action of prebiotics and their effects on gastro-intestinal disorders in adults. Nutrients. 2020;12:1037. doi: 10.3390/nu12041037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Uauy R. Foreword. Br. J. Nutr. 2009;101:S1. doi: 10.1017/S0007114509990511. [DOI] [PubMed] [Google Scholar]
- 56.Gibson G.R., Hutkins R., Sanders M.E., Prescott S.L., Reimer R.A., Salminen S.J., Scott K., Stanton C., Swanson K.S., Cani P.D., et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017;14:491–502. doi: 10.1038/nrgastro.2017.75. [DOI] [PubMed] [Google Scholar]
- 57.Cardoso B.B., Amorim C., Silvério S.C., Rodrigues L.R. Novel and emerging prebiotics: Advances and opportunities. Adv. Food Nutr. Res. 2021;95:41–95. doi: 10.1016/bs.afnr.2020.08.001. [DOI] [PubMed] [Google Scholar]
- 58.Kelly G. Inulin-type prebiotics: A review (Part 2) Altern. Med. Rev. 2009;14:36–55. [PubMed] [Google Scholar]
- 59.Yeung C.K., Glahn R.P., Welch R.M., Miller D.D. Prebiotics and iron bioavailability—Is there a connection? J. Food Sci. 2005;70:584–590. doi: 10.1111/j.1365-2621.2005.tb09984.x. [DOI] [Google Scholar]
- 60.Ibrahim O.O. Functional Oligosaccharide: Chemicals Structure, Manufacturing, Health Benefits, Applications and Regulations. J. Food Chem. Nanotechnol. 2018;4:65–76. doi: 10.17756/jfcn.2018-060. [DOI] [Google Scholar]
- 61.Roberfroid M. Prebiotics: The concept revisited. J. Nutr. 2007;137:830S–837S. doi: 10.1093/jn/137.3.830S. [DOI] [PubMed] [Google Scholar]
- 62.Cui S.W., Roberts K.T. Dietary Fiber. Fulfilling the Promise of Added-Value Formulations. Academic Press; Cambridge, MA, USA: 2009. [Google Scholar]
- 63.Chatterjee P., Ojiambo M. Food Ingredients. Volume 11. Elsevier Inc.; Amsterdam, The Netherlands: 2014. [Google Scholar]
- 64.Korczak R., Slavin J.L. Fructooligosaccharides and appetite. Curr. Opin. Clin. Nutr. Metab. Care. 2018;21:377–380. doi: 10.1097/MCO.0000000000000502. [DOI] [PubMed] [Google Scholar]
- 65.Yun J.W. Fructooligosaccharides—Occurrence, preparation, and application. Enzyme Microb. Technol. 1996;19:107–117. doi: 10.1016/0141-0229(95)00188-3. [DOI] [Google Scholar]
- 66.Crittenden R., Playne M. Production, properties and applications of food-grade oligosaccharides. Trends Food Sci. Technol. 1996;7:353–361. doi: 10.1016/S0924-2244(96)10038-8. [DOI] [Google Scholar]
- 67.Sako T., Tanaka R. Encyclopedia of Dairy Sciences. 2nd ed. Elsevier Ltd.; Amsterdam, The Netherlands: 2011. Prebiotics: Types; pp. 354–364. [DOI] [Google Scholar]
- 68.Commercially Available Oligosaccharides. [(accessed on 1 April 2022)]. Available online: https://agris.fao.org/agris-search/search.do?recordID=BE9601560.
- 69.Tanaka R., Takayama H., Morotomi M., Kuroshima T., Ueyama S., Matsumoto K., Kuroda A., Mutai M. Effects of Administration of TOS and Bifidobacterium breve 4006 on the Human Fecal Flora. Bifidobact. Microflora. 1983;2:17–24. doi: 10.12938/bifidus1982.2.1_17. [DOI] [Google Scholar]
- 70.Mahoney R.R. Galactosyl-oligosaccharide formation during lactose hydrolysis: A review. Food Chem. 1998;63:147–154. doi: 10.1016/S0308-8146(98)00020-X. [DOI] [Google Scholar]
- 71.Bao Y., Chen C., Newburg D.S. Quantification of neutral human milk oligosaccharides by graphitic carbon high-performance liquid chromatography with tandem mass spectrometry. Anal. Biochem. 2013;433:28–35. doi: 10.1016/j.ab.2012.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Thongaram T., Hoeflinger J.L., Chow J., Miller M.J. Human milk oligosaccharide consumption by probiotic and human-associated bifidobacteria and lactobacilli. J. Dairy Sci. 2017;100:7825–7833. doi: 10.3168/jds.2017-12753. [DOI] [PubMed] [Google Scholar]
- 73.Akkerman R., Faas M.M., de Vos P. Non-digestible carbohydrates in infant formula as substitution for human milk oligosaccharide functions: Effects on microbiota and gut maturation. Crit. Rev. Food Sci. Nutr. 2019;59:1486–1497. doi: 10.1080/10408398.2017.1414030. [DOI] [PubMed] [Google Scholar]
- 74.Xu G., Davis J.C., Goonatilleke E., Smilowitz J.T., German J.B., Lebrilla C.B. Absolute Quantitation of Human Milk Oligosaccharides Reveals Phenotypic Variations during Lactation. J. Nutr. 2017;147:117–124. doi: 10.3945/jn.116.238279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Chaturvedi P., Warren C.D., Altaye M., Morrow A.L., Ruiz-Palacios G., Pickering L.K., Newburg D.S. Fucosylated human milk oligosaccharides vary between individuals and over the course of lactation. Glycobiology. 2001;11:365–372. doi: 10.1093/glycob/11.5.365. [DOI] [PubMed] [Google Scholar]
- 76.Thurl S., Munzert M., Henker J., Boehm G., Müller-Werner B., Jelinek J., Stahl B. Variation of human milk oligosaccharides in relation to milk groups and lactational periods. Br. J. Nutr. 2010;104:1261–1271. doi: 10.1017/S0007114510002072. [DOI] [PubMed] [Google Scholar]
- 77.Thomson P., Medina D.A., Garrido D. Human milk oligosaccharides and infant gut bifidobacteria: Molecular strategies for their utilization. Food Microbiol. 2018;75:37–46. doi: 10.1016/j.fm.2017.09.001. [DOI] [PubMed] [Google Scholar]
- 78.Bode L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology. 2012;22:1147–1162. doi: 10.1093/glycob/cws074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Cheng Y.-J., Yeung C.-Y. Recent advance in infant nutrition: Human milk oligosaccharides. Pediatr. Neonatol. 2021;62:347–353. doi: 10.1016/j.pedneo.2020.12.013. [DOI] [PubMed] [Google Scholar]
- 80.Morozov V., Hansman G., Hanisch F.G., Schroten H., Kunz C. Human Milk Oligosaccharides as Promising Antivirals. Mol. Nutr. Food Res. 2018;62:1–46. doi: 10.1002/mnfr.201700679. [DOI] [PubMed] [Google Scholar]
- 81.Asakuma S., Hatakeyama E., Urashima T., Yoshida E., Katayama T., Yamamoto K., Kumagai H., Ashida H., Hirose J., Kitaoka M. Physiology of Consumption of Human Milk Oligosaccharides by Infant Gut-associated Bifidobacteria. J. Biol. Chem. 2011;286:34583–34592. doi: 10.1074/jbc.M111.248138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Vandenplas Y., Berger B., Carnielli V.P., Ksiazyk J., Lagström H., Luna M.S., Migacheva N., Mosselmans J.M., Picaud J.C., Possner M., et al. Human milk oligosaccharides: 2’-fucosyllactose (2’-FL) and lacto-n-neotetraose (LNnT) in infant formula. Nutrients. 2018;10:1161. doi: 10.3390/nu10091161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Corona L., Lussu A., Bosco A., Pintus R., Cesare Marincola F., Fanos V., Dessì A. Human Milk Oligosaccharides: A Comprehensive Review towards Metabolomics. Children. 2021;8:804. doi: 10.3390/children8090804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Marcobal A., Barboza M., Froehlich J.W., Block D.E., German J.B., Lebrilla C.B., Mills D.A. Consumption of human milk oligosaccharides by gut-related microbes. J. Agric. Food Chem. 2010;58:5334–5340. doi: 10.1021/jf9044205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Donovan S.M., Comstock S.S. Human Milk Oligosaccharides Influence Neonatal Mucosal and Systemic Immunity. Ann. Nutr. Metab. 2016;69((Suppl. 2)):42–51. doi: 10.1159/000452818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Comstock S.S., Li M., Wang M., Monaco M.H., Kuhlenschmidt T.B., Kuhlenschmidt M.S., Donovan S.M. Dietary Human Milk Oligosaccharides but Not Prebiotic Oligosaccharides Increase Circulating Natural Killer Cell and Mesenteric Lymph Node Memory T Cell Populations in Noninfected and Rotavirus-Infected Neonatal Piglets. J. Nutr. 2017;147:1041–1047. doi: 10.3945/jn.116.243774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Bode L. The functional biology of human milk oligosaccharides. Early Hum. Dev. 2015;91:619–622. doi: 10.1016/j.earlhumdev.2015.09.001. [DOI] [PubMed] [Google Scholar]
- 88.Jacobi S.K., Yatsunenko T., Li D., Dasgupta S., Yu R.K., Berg B.M., Chichlowski M., Odle J. Dietary Isomers of Sialyllactose Increase Ganglioside Sialic Acid Concentrations in the Corpus Callosum and Cerebellum and Modulate the Colonic Microbiota of Formula-Fed Piglets. J. Nutr. 2016;146:200–208. doi: 10.3945/jn.115.220152. [DOI] [PubMed] [Google Scholar]
- 89.Rousseaux A., Brosseau C., Le Gall S., Piloquet H., Barbarot S., Bodinier M. Human Milk Oligosaccharides: Their Effects on the Host and Their Potential as Therapeutic Agents. Front. Immunol. 2021;12:680911. doi: 10.3389/fimmu.2021.680911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Luo Y., Tu Y., Ren F., Zhang H. Characterization and functional properties of Maillard reaction products of β-lactoglobulin and polydextrose. Food Chem. 2021;377:131749. doi: 10.1016/j.foodchem.2021.131749. [DOI] [PubMed] [Google Scholar]
- 91.Lahtinen S.J., Knoblock K., Drakoularakou A., Jacob M., Stowell J., Gibson G.R., Ouwehand A.C. Effect of molecule branching and glycosidic linkage on the degradation of polydextrose by gut microbiota. Biosci. Biotechnol. Biochem. 2010;74:2016–2021. doi: 10.1271/bbb.100251. [DOI] [PubMed] [Google Scholar]
- 92.Caselato V., Ouwehand A., Flavors I., Andreollo N.A. Polydextrose: Physiological Function, and Effects on Health. Nutrients. 2016;8:553. doi: 10.3390/nu8090553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Zhong J., Luo B., Xiang M., Liu H., Zhai Z., Wang T., Craig S.A.S. Studies on the effects of polydextrose intake on physiologic funtions in Chinese people. Am. J. Clin. Nutr. 2000;72:1503–1509. doi: 10.1093/ajcn/72.6.1503. [DOI] [PubMed] [Google Scholar]
- 94.Yao N., Paez A.V., White P.J. Structure and function of starch and resistant starch from corn with different doses of mutant amylose-extender and floury-1 alleles. J. Agric. Food Chem. 2009;57:2040–2048. doi: 10.1021/jf8033682. [DOI] [PubMed] [Google Scholar]
- 95.Fuentes-Zaragoza E., Sánchez-Zapata E., Sendra E., Sayas E., Navarro C., Fernández-Lõpez J., Pérez-Alvarez J.A. Resistant starch as prebiotic: A review. Starch/Staerke. 2011;63:406–415. doi: 10.1002/star.201000099. [DOI] [Google Scholar]
- 96.Marlatt K.L., White U.A., Beyl R.A., Peterson C.M., Corby K., Marco M.L., Keenan M.J., Martin R.J., Aryana K.J., Ravussin E., et al. Role of resistant starch on diabetes risk factors in people with prediabetes: Design, conduct, and baseline results of the STARCH trial. Contemp. Clin. Trials. 2018;65:99–108. doi: 10.1016/j.cct.2017.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Tingirikari J.M.R. Microbiota-accessible pectic poly- and oligosaccharides in gut health. Food Funct. 2018;9:5059–5073. doi: 10.1039/C8FO01296B. [DOI] [PubMed] [Google Scholar]
- 98.Yoo H.D., Kim D., Paek S.H., Oh S.E. Plant cell wall polysaccharides as potential resources for the development of novel prebiotics. Biomol. Ther. 2012;20:371–379. doi: 10.4062/biomolther.2012.20.4.371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Yang J., Maldonado-Gómez M.X., Hutkins R.W., Rose D.J. Production and in vitro Fermentation of Soluble, Non-digestible, Feruloylated Oligo- and Polysaccharides from Maize and Wheat Brans. J. Agric. Food Chem. 2014;62:159–166. doi: 10.1021/jf404305y. [DOI] [PubMed] [Google Scholar]
- 100.Moon J.S., Shin S.Y., Choi H.S., Joo W., Cho S.K., Li L., Kang J.-H., Kim T.-J., Han N.S. In vitro digestion and fermentation properties of linear sugar-beet arabinan and its oligosaccharides. Carbohydr. Polym. 2015;131:50–56. doi: 10.1016/j.carbpol.2015.05.022. [DOI] [PubMed] [Google Scholar]
- 101.Wilkowska A., Nowak A., Antczak-Chrobot A., Motyl I., Czyżowska A., Paliwoda A. Structurally Different Pectic Oligosaccharides Produced from Apple Pomace and Their Biological Activity In Vitro. Foods. 2019;8:365. doi: 10.3390/foods8090365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Zhang W., Xu P., Zhang H. Pectin in cancer therapy: A review. Trends Food Sci. Technol. 2015;44:258–271. doi: 10.1016/j.tifs.2015.04.001. [DOI] [Google Scholar]
- 103.Vera C., Córdova A., Aburto C., Guerrero C., Suárez S., Illanes A. Synthesis and purification of galacto-oligosaccharides: State of the art. World J. Microbiol. Biotechnol. 2016;32:197. doi: 10.1007/s11274-016-2159-4. [DOI] [PubMed] [Google Scholar]
- 104.Zhang W., Wang T., Chen X., Pang X., Zhang S., Obaroakpo J.U., Shilong J., Lu J., Lv J. Absolute quantification of twelve oligosaccharides in human milk using a targeted mass spectrometry-based approach. Carbohydr. Polym. 2019;219:328–333. doi: 10.1016/j.carbpol.2019.04.092. [DOI] [PubMed] [Google Scholar]
- 105.German J., Freeman S., Lebrilla C., Mills D. Human milk oligosaccharides: Evolution, structures and bioselectivity as substrates for intestinal bacteria. Nestle Nutr. Work. Ser. Pediatr. Progr. 2008;62:205–218. doi: 10.1159/000146322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Ackerman D.L., Craft K.M., Townsend S.D. Infant food applications of complex carbohydrates: Structure, synthesis, and function. Carbohydr. Res. 2017;437:16–27. doi: 10.1016/j.carres.2016.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Tungland B.C., Meyer D. Nondigestible oligo-and polysaccharides (dietary fiber): Their physiology and role in human health and food. Compr. Rev. Food Sci. Food Saf. 2002;1:90–109. doi: 10.1111/j.1541-4337.2002.tb00009.x. [DOI] [PubMed] [Google Scholar]
- 108.Xu W., Liu Q., Yu S., Zhang T., Mu W. Synthesis of Lactosucrose Using a Recombinant Levansucrase from Brenneria goodwinii. Appl. Biochem. Biotechnol. 2018;186:292–305. doi: 10.1007/s12010-018-2743-1. [DOI] [PubMed] [Google Scholar]
- 109.Langenaeken N.A., De Schutter D.P., Courtin C.M. Arabinoxylan from non-malted cereals can act as mouthfeel contributor in beer. Carbohydr. Polym. 2020;239:116257. doi: 10.1016/j.carbpol.2020.116257. [DOI] [PubMed] [Google Scholar]
- 110.Gérard D., Méline T., Muzard M., Deleu M., Plantier-Royon R., Rémond C. Enzymatically-synthesized xylo-oligosaccharides laurate esters as surfactants of interest. Carbohydr. Res. 2020;495:108090. doi: 10.1016/j.carres.2020.108090. [DOI] [PubMed] [Google Scholar]
- 111.Aachary A.A., Prapulla S.G. Corncob-Induced endo-1,4-β-d-Xylanase of Aspergillus oryzae MTCC 5154: Production and Characterization of Xylobiose from Glucuronoxylan. J. Agric. Food Chem. 2008;56:3981–3988. doi: 10.1021/jf073430i. [DOI] [PubMed] [Google Scholar]
- 112.Carvalho A.F.A., de Oliva Neto P., da Silva D.F., Pastore G.M. Xylo-oligosaccharides from lignocellulosic materials: Chemical structure, health benefits and production by chemical and enzymatic hydrolysis. Food Res. Int. 2013;51:75–85. doi: 10.1016/j.foodres.2012.11.021. [DOI] [Google Scholar]
- 113.Shendurse A.M., Khedkar C.D. Lactose. In: Caballero B., Finglas P.M., Toldrá F., editors. Encyclopedia of Food and Health. Academic Press; Oxford, UK: 2016. pp. 509–516. [Google Scholar]
- 114.Panesar P.S., Bali V. Prebiotics. In: Caballero B., Finglas P.M., Toldrá F., editors. Encyclopedia of Food and Health. Academic Press; Oxford, UK: 2016. pp. 464–471. [Google Scholar]
- 115.Sivaprakasam S., Prasad P.D., Singh N. Benefits of short-chain fatty acids and their receptors in inflammation and carcinogenesis. Pharmacol. Ther. 2016;164:144–151. doi: 10.1016/j.pharmthera.2016.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Prasad S., Dhiman R.K., Duseja A., Chawla Y.K., Sharma A., Agarwal R. Lactulose improves cognitive functions and health-related quality of life in patients with cirrhosis who have minimal hepatic encephalopathy. Hepatology. 2007;45:549–559. doi: 10.1002/hep.21533. [DOI] [PubMed] [Google Scholar]
- 117.Elkington S.G. Lactulose. Gut. 1970;11:1043–1048. doi: 10.1136/gut.11.12.1043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Song M., Zeng J., Jia T., Gao H., Zhang R., Jiang J., Li G., Su T. Effects of sialylated lactulose on the mouse intestinal microbiome using Illumina high-throughput sequencing. Appl. Microbiol. Biotechnol. 2019;103:9067–9076. doi: 10.1007/s00253-019-10169-7. [DOI] [PubMed] [Google Scholar]
- 119.Mu W., Chen Q., Wang X., Zhang T., Jiang B. Current studies on physiological functions and biological production of lactosucrose. Appl. Microbiol. Biotechnol. 2013;97:7073–7080. doi: 10.1007/s00253-013-5079-3. [DOI] [PubMed] [Google Scholar]
- 120.Fernando W.M.U., Hill J.E., Zello G.A., Tyler R.T., Dahl W.J., Van Kessel A.G. Diets supplemented with chickpea or its main oligosaccharide component raffinose modify faecal microbial composition in healthy adults. Benef. Microbes. 2010;1:197–207. doi: 10.3920/BM2009.0027. [DOI] [PubMed] [Google Scholar]
- 121.Silvério S.C., Macedo E.A., Teixeira J.A., Rodrigues L.R. Perspectives on the biotechnological production and potential applications of lactosucrose: A review. J. Funct. Foods. 2015;19:74–90. doi: 10.1016/j.jff.2015.09.014. [DOI] [Google Scholar]
- 122.Terada A., Hara H., Oishi T., Matsui S., Mitsuoka T., Nakajyo S., Fujimori I., Hara K. Effect of Dietary Lactosucrose on Faecal Flora and Faecal Metabolites of Dogs. Microb. Ecol. Health Dis. 1992;5:87–92. doi: 10.3109/08910609209141294. [DOI] [Google Scholar]
- 123.Hino K., Kurose M., Sakurai T., Inoue S., Oku K., Chaen H., Fukuda S. Effect of Dietary Lactosucrose (4G-β-D-Galactosylsucrose) on the Intestinal Immune Functions in Mice. J. Appl. Glycosci. 2007;54:169–172. doi: 10.5458/jag.54.169. [DOI] [Google Scholar]
- 124.Hoffman W.F., Gortner R.A. The preparation and analysis of the various proteins of wheat flour with special reference to the globulin, albumin and proteose fractions. Cereal Chem. 2021;4:221–229. [Google Scholar]
- 125.Bastos R., Coelho E., Coimbra M.A. 8-Arabinoxylans from cereal by-products: Insights into structural features, recovery, and applications. In: Galanakis C., editor. Woodhead Publishing Series in Food Science, Technology and Nutrition. Woodhead Publishing; Sawston, UK: 2018. pp. 227–251. [Google Scholar]
- 126.Izydorczyk M.S. Handbook of Hydrocolloids. 2nd ed. Elsevier; Amsterdam, The Netherlands: 2009. Arabinoxylans; pp. 653–692. [DOI] [Google Scholar]
- 127.Paz-Samaniego R., Méndez-Encinas M., Fierro-Islas J.M., Marquez-Escalante J., Rascón-Chu A., Martinez-Lopez A.L., Carvajal-Millan E. Ferulic Acid: Antioxidant Properties, Uses and Potential Health Benefits. Nova Science Publishers; Hauppauge, NY, USA: 2014. Ferulated arabinoxylans recovered from low-value maize by-products: Gelation and antioxidant capacity; pp. 151–164. [Google Scholar]
- 128.Mendis M., Simsek S. Arabinoxylans and human health. Food Hydrocoll. 2014;42:239–243. doi: 10.1016/j.foodhyd.2013.07.022. [DOI] [Google Scholar]
- 129.Lu Z.X., Walker K.Z., Muir J.G., Mascara T., O’Dea K. Arabinoxylan fiber, a byproduct of wheat flour processing, reduces the postprandial glucose response in normoglycemic subjects. Am. J. Clin. Nutr. 2000;71:1123–1128. doi: 10.1093/ajcn/71.5.1123. [DOI] [PubMed] [Google Scholar]
- 130.Mendis M., Leclerc E., Simsek S. Arabinoxylans, gut microbiota and immunity. Carbohydr. Polym. 2016;139:159–166. doi: 10.1016/j.carbpol.2015.11.068. [DOI] [PubMed] [Google Scholar]
- 131.Althubiani A.S., Al-Ghamdi S.B., Samreen, Qais F.A., Khan M.S., Ahmad I., Malak H.A. Plant-Derived Prebiotics and Its Health Benefits. Elsevier Inc.; Amsterdam, The Netherlands: 2018. [Google Scholar]
- 132.Gibson G.R., Probert H.M., Van Loo J., Rastall R.A., Roberfroid M.B. Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics. Nutr. Res. Rev. 2004;17:259–275. doi: 10.1079/NRR200479. [DOI] [PubMed] [Google Scholar]
- 133.Mitmesser S., Combs M. Prebiotics: Inulin and Other Oligosaccharides. Elsevier Inc.; Amsterdam, The Netherlands: 2017. [Google Scholar]
- 134.Lin S.-H., Chou L.-M., Chien Y.-W., Chang J.-S., Lin C.-I. Prebiotic Effects of Xylooligosaccharides on the Improvement of Microbiota Balance in Human Subjects. Gastroenterol. Res. Pract. 2016;2016:5789232. doi: 10.1155/2016/5789232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Parajó J.C., Garrote G., Cruz J.M., Dominguez H. Production of xylooligosaccharides by autohydrolysis of lignocellulosic materials. Trends Food Sci. Technol. 2004;15:115–120. doi: 10.1016/j.tifs.2003.09.009. [DOI] [Google Scholar]
- 136.Scott K.P., Martin J.C., Duncan S.H., Flint H.J. Prebiotic stimulation of human colonic butyrate-producing bacteria and bifidobacteria, in vitro. FEMS Microbiol. Ecol. 2014;87:30–40. doi: 10.1111/1574-6941.12186. [DOI] [PubMed] [Google Scholar]
- 137.Tuohy K., Rouzaud G., Bruck W., Gibson G. Modulation of the Human Gut Microflora Towards Improved Health Using Prebiotics—Assessment of Efficacy. Curr. Pharm. Des. 2005;11:75–90. doi: 10.2174/1381612053382331. [DOI] [PubMed] [Google Scholar]
- 138.Modler H.W. Bifidogenic factors—Sources, metabolism and applications. Int. Dairy J. 1994;4:383–407. doi: 10.1016/0958-6946(94)90055-8. [DOI] [Google Scholar]
- 139.Sabater-Molina M., Larqué E., Torrella F., Zamora S. Dietary fructooligosaccharides and potential benefits on health. J. Physiol. Biochem. 2009;65:315–328. doi: 10.1007/BF03180584. [DOI] [PubMed] [Google Scholar]
- 140.Carabin I.G., Flamm W.G. Evaluation of safety of inulin and oligofructose as dietary fiber. Regul. Toxicol. Pharmacol. 1999;30:268–282. doi: 10.1006/rtph.1999.1349. [DOI] [PubMed] [Google Scholar]
- 141.Elison E., Vigsnaes L.K., Rindom Krogsgaard L., Rasmussen J., Sørensen N., McConnell B., Hennet T., Sommer M.O.A., Bytzer P. Oral supplementation of healthy adults with 2’-O-fucosyllactose and lacto-N-neotetraose is well tolerated and shifts the intestinal microbiota. Br. J. Nutr. 2016;116:1356–1368. doi: 10.1017/S0007114516003354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Chen L., Liu R., Qin C., Meng Y., Zhang J., Wang Y., Xu G. Sources and intake of resistant starch in the Chinese diet. Asia Pac. J. Clin. Nutr. 2010;19:274–282. [PubMed] [Google Scholar]
- 143.Brouns F., Theuwissen E., Adam A., Bell M., Berger A., Mensink R.P. Cholesterol-lowering properties of different pectin types in mildly hyper-cholesterolemic men and women. Eur. J. Clin. Nutr. 2012;66:591–599. doi: 10.1038/ejcn.2011.208. [DOI] [PubMed] [Google Scholar]
- 144.Salminen S., Salminen E. Lactulose, lactic acid bacteria, intestinal microecology and mucosal protection. Scand. J. Gastroenterol. Suppl. 1997;222:45–48. doi: 10.1080/00365521.1997.11720717. [DOI] [PubMed] [Google Scholar]
- 145.Barreteau H., Delattre C., Michaud P. Production of oligosaccharides as promising new food additive generation. Food Technol. Biotechnol. 2006;44:323–333. [Google Scholar]
- 146.Denji K.A., Mansour M.R., Akrami R., Ghobadi S., Jafarpour S.A., Mirbeygi S.K. Effect of dietary prebiotic mannan oligosaccharide (mos) on growth performance, intestinal microflora, body composition, haematological and blood serum biochemical parameters of rainbow trout (oncorhynchus mykiss) juveniles. J. Fish. Aquat. Sci. 2015;10:255–265. doi: 10.3923/jfas.2015.255.265. [DOI] [Google Scholar]
- 147.Klatt N.R., Canary L.A., Sun X., Vinton C.L., Funderburg N.T., Morcock D.R., Quiñones M., Deming C.B., Perkins M., Hazuda D.J., et al. Probiotic/prebiotic supplementation of antiretrovirals improves gastrointestinal immunity in SIV-infected macaques. J. Clin. Investig. 2013;123:903–907. doi: 10.1172/JCI66227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Champagne C.P. Some Technological Challenges in the Addition of Probiotic Bacteria to Foods. Springer; New York, NY, USA: 2009. [Google Scholar]
- 149.Akutko K., Stawarski A. Probiotics, prebiotics and synbiotics in inflammatory bowel diseases. J. Clin. Med. 2021;10:2466. doi: 10.3390/jcm10112466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Cunningham M., Azcarate-Peril M.A., Barnard A., Benoit V., Grimaldi R., Guyonnet D., Holscher H.D., Hunter K., Manurung S., Obis D., et al. Shaping the Future of Probiotics and Prebiotics. Trends Microbiol. 2021;29:667–685. doi: 10.1016/j.tim.2021.01.003. [DOI] [PubMed] [Google Scholar]
- 151.Wan X., Guo H., Liang Y., Zhou C., Liu Z., Li K., Niu F., Zhai X., Wang L. The physiological functions and pharmaceutical applications of inulin: A review. Carbohydr. Polym. 2020;246:116589. doi: 10.1016/j.carbpol.2020.116589. [DOI] [PubMed] [Google Scholar]
- 152.Rezende E.S.V., Lima G.C., Naves M.M.V. Dietary fibers as beneficial microbiota modulators: A proposal classification by prebiotic categories. Nutrition. 2021;89:111217. doi: 10.1016/j.nut.2021.111217. [DOI] [PubMed] [Google Scholar]
- 153.Yu X., Gurry T., Nguyen L.T.T., Richardson H.S., Alm E.J. Prebiotics and Community Composition Influence Gas Production of the Human Gut Microbiota. MBio. 2020;11:e00217-20. doi: 10.1128/mBio.00217-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Kleessen B., Sykura B., Zunft H.J., Blaut M. Effects of inulin and lactose on fecal microflora, microbial activity, and bowel habit in elderly constipated persons. Am. J. Clin. Nutr. 1997;65:1397–1402. doi: 10.1093/ajcn/65.5.1397. [DOI] [PubMed] [Google Scholar]
- 155.Tuohy K.M., Finlay R.K., Wynne A.G., Gibson G.R. A Human Volunteer Study on the Prebiotic Effects of HP-Inulin—Faecal Bacteria Enumerated Using Fluorescent In Situ Hybridisation (FISH) Anaerobe. 2001;7:113–118. doi: 10.1006/anae.2001.0368. [DOI] [Google Scholar]
- 156.Roland N., Nugon-Baudon L., Andrieux C., Szylit O. Comparative study of the fermentative characteristics of inulin and different types of fibre in rats inoculated with a human Whole faecal flora. Br. J. Nutr. 1995;74:239–249. doi: 10.1079/BJN19950127. [DOI] [PubMed] [Google Scholar]
- 157.Flores-Maltos D.A., Mussatto S.I., Contreras-Esquivel J.C., Rodríguez-Herrera R., Teixeira J.A., Aguilar C.N. Biotechnological production and application of fructooligosaccharides. Crit. Rev. Biotechnol. 2016;36:259–267. doi: 10.3109/07388551.2014.953443. [DOI] [PubMed] [Google Scholar]
- 158.Xiao J., Sakaguchi E., Bai G. Short-term supplementation with dietary fructooligosaccharide and dietary mannitol elevated the absorption of calcium and magnesium in adult rats. Czech J. Anim. Sci. 2016;61:281–289. doi: 10.17221/44/2015-CJAS. [DOI] [Google Scholar]
- 159.Sangeetha P.T., Ramesh M.N., Prapulla S.G. Recent trends in the microbial production, analysis and application of Fructooligosaccharides. Trends Food Sci. Technol. 2005;16:442–457. doi: 10.1016/j.tifs.2005.05.003. [DOI] [Google Scholar]
- 160.Maathuis A.J.H., van den Heuvel E.G., Schoterman M.H.C., Venema K. Galacto-Oligosaccharides Have Prebiotic Activity in a Dynamic In Vitro Colon Model Using a 13C-Labeling Technique. J. Nutr. 2012;142:1205–1212. doi: 10.3945/jn.111.157420. [DOI] [PubMed] [Google Scholar]
- 161.Davis L.M.G., Martínez I., Walter J., Goin C., Hutkins R.W. Barcoded pyrosequencing reveals that consumption of galactooligosaccharides results in a highly specific bifidogenic response in humans. PLoS ONE. 2011;6:e25200. doi: 10.1371/journal.pone.0025200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Van Loo J., Cummings J., Delzenne N., Englyst H., Franck A., Hopkins M., Kok N., Macfarlane G., Newton D., Quigley M., et al. Functional food properties of non-digestible oligosaccharides: A consensus report from the ENDO project (DGXII AIRII-CT94-1095) Br. J. Nutr. 1999;81:121–132. doi: 10.1017/s0007114599000252. [DOI] [PubMed] [Google Scholar]
- 163.Tran N.T., Tang Y., Li Z., Zhang M., Wen X., Ma H., Li S. Galactooligosaccharides and Resistant Starch Altered Microbiota and Short-Chain Fatty Acids in an in vitro Fermentation Study Using Gut Contents of Mud Crab (Scylla paramamosain) Front. Microbiol. 2020;11:1352. doi: 10.3389/fmicb.2020.01352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Burokas A., Arboleya S., Moloney R.D., Peterson V.L., Murphy K., Clarke G., Stanton C., Dinan T.G., Cryan J.F. Targeting the Microbiota-Gut-Brain Axis: Prebiotics Have Anxiolytic and Antidepressant-like Effects and Reverse the Impact of Chronic Stress in Mice. Biol. Psychiatry. 2017;82:472–487. doi: 10.1016/j.biopsych.2016.12.031. [DOI] [PubMed] [Google Scholar]
- 165.Schmidt K., Cowen P.J., Harmer C.J., Tzortzis G., Errington S., Burnet P.W.J. Prebiotic intake reduces the waking cortisol response and alters emotional bias in healthy volunteers. Psychopharmacology. 2015;232:1793–1801. doi: 10.1007/s00213-014-3810-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Ray C., Kerketta J.A., Rao S., Patel S., Dutt S., Arora K., Pournami F., Bhushan P. Human Milk Oligosaccharides: The Journey Ahead. Int. J. Pediatr. 2019;2019:2390240. doi: 10.1155/2019/2390240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.LoCascio R.G., Ninonuevo M.R., Freeman S.L., Sela D.A., Grimm R., Lebrilla C.B., Mills D.A., German J.B. Glycoprofiling of bifidobacterial consumption of human milk oligosaccharides demonstrates strain specific, preferential consumption of small chain glycans secreted in early human lactation. J. Agric. Food Chem. 2007;55:8914–8919. doi: 10.1021/jf0710480. [DOI] [PubMed] [Google Scholar]
- 168.Bunesova V., Lacroix C., Schwab C. Fucosyllactose and L-fucose utilization of infant Bifidobacterium longum and Bifidobacterium kashiwanohense. BMC Microbiol. 2016;16:248. doi: 10.1186/s12866-016-0867-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Gibson G.R., Wang X. Regulatory effects of bifidobacteria on the growth of other colonic bacteria. J. Appl. Bacteriol. 1994;77:412–420. doi: 10.1111/j.1365-2672.1994.tb03443.x. [DOI] [PubMed] [Google Scholar]
- 170.Collado M.C., Cernada M., Baüerl C., Vento M., Pérez-Martínez G. Microbial ecology and host-microbiota interactions during early life stages. Gut Microbes. 2012;3:352–365. doi: 10.4161/gmic.21215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Schwab C., Ruscheweyh H.J., Bunesova V., Pham V.T., Beerenwinkel N., Lacroix C. Trophic interactions of infant bifidobacteria and eubacterium hallii during L-fucose and fucosyllactose degradation. Front. Microbiol. 2017;8:95. doi: 10.3389/fmicb.2017.00095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Weichert S., Jennewein S., Hüfner E., Weiss C., Borkowski J., Putze J., Schroten H. Bioengineered 2′-fucosyllactose and 3-fucosyllactose inhibit the adhesion of Pseudomonas aeruginosa and enteric pathogens to human intestinal and respiratory cell lines. Nutr. Res. 2013;33:831–838. doi: 10.1016/j.nutres.2013.07.009. [DOI] [PubMed] [Google Scholar]
- 173.Reverri E.J., Devitt A.A., Kajzer J.A., Baggs G.E., Borschel M.W. Review of the clinical experiences of feeding infants formula containing the human milk oligosaccharide 2′-fucosyllactose. Nutrients. 2018;10:1346. doi: 10.3390/nu10101346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Holscher H.D., Caporaso J.G., Hooda S., Brulc J.M., Fahey G.C.J., Swanson K.S. Fiber supplementation influences phylogenetic structure and functional capacity of the human intestinal microbiome: Follow-up of a randomized controlled trial. Am. J. Clin. Nutr. 2015;101:55–64. doi: 10.3945/ajcn.114.092064. [DOI] [PubMed] [Google Scholar]
- 175.Hooda S., Boler B.M.V., Serao M.C.R., Brulc J.M., Staeger M.A., Boileau T.W., Dowd S.E., Fahey G.C.J., Swanson K.S. 454 pyrosequencing reveals a shift in fecal microbiota of healthy adult men consuming polydextrose or soluble corn fiber. J. Nutr. 2012;142:1259–1265. doi: 10.3945/jn.112.158766. [DOI] [PubMed] [Google Scholar]
- 176.Toporovski M.S., de Morais M.B., Abuhab A., Crippa Júnior M.A. Effect of polydextrose/fructooligosaccharide mixture on constipation symptoms in children aged 4 to 8 years. Nutrients. 2021;13:1634. doi: 10.3390/nu13051634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Costabile A., Fava F., Röytiö H., Forssten S.D., Olli K., Klievink J., Rowland I.R., Ouwehand A.C., Rastall R.A., Gibson G.R., et al. Impact of polydextrose on the faecal microbiota: A double-blind, crossover, placebo-controlled feeding study in healthy human subjects. Br. J. Nutr. 2012;108:471–481. doi: 10.1017/S0007114511005782. [DOI] [PubMed] [Google Scholar]
- 178.Tachon S., Zhou J., Keenan M., Martin R., Marco M.L. The intestinal microbiota in aged mice is modulated by dietary resistant starch and correlated with improvements in host responses. FEMS Microbiol. Ecol. 2013;83:299–309. doi: 10.1111/j.1574-6941.2012.01475.x. [DOI] [PubMed] [Google Scholar]
- 179.Brüssow H. Microbiota and healthy ageing: Observational and nutritional intervention studies. Microb. Biotechnol. 2013;6:326–334. doi: 10.1111/1751-7915.12048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Charrier J.A., Martin R.J., McCutcheon K.L., Raggio A.M., Goldsmith F., Goita M., Senevirathne R.N., Brown I.L., Pelkman C., Zhou J., et al. High fat diet partially attenuates fermentation responses in rats fed resistant starch from high-amylose maize. Obesity. 2013;21:2350–2355. doi: 10.1002/oby.20362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Gullón B., Gómez B., Martínez-Sabajanes M., Yáñez R., Parajó J.C., Alonso J.L. Pectic oligosaccharides: Manufacture and functional properties. Trends Food Sci. Technol. 2013;30:153–161. doi: 10.1016/j.tifs.2013.01.006. [DOI] [Google Scholar]
- 182.Gómez B., Gullón B., Yáñez R., Schols H., Alonso J.L. Prebiotic potential of pectins and pectic oligosaccharides derived from lemon peel wastes and sugar beet pulp: A comparative evaluation. J. Funct. Foods. 2016;20:108–121. doi: 10.1016/j.jff.2015.10.029. [DOI] [Google Scholar]
- 183.Leijdekkers A.G.M., Aguirre M., Venema K., Bosch G., Gruppen H., Schols H.A. In vitro fermentability of sugar beet pulp derived oligosaccharides using human and pig fecal inocula. J. Agric. Food Chem. 2014;62:1079–1087. doi: 10.1021/jf4049676. [DOI] [PubMed] [Google Scholar]
- 184.Gibson G., Angus F. Leatherhead Ingredients Handbook: Prebiotics and Probiotics. Springer; New York, NY, USA: Wiley–Blackwell; Hoboken, NJ, USA: 2000. [Google Scholar]
- 185.Terada A., Hara H., Kato S., Kimura T., Fujimori I., Hara K., Maruyama T., Mitsuoka T. Effect of Lactosucrose (4G-β-D-Galactosylsucrose) on Fecal Flora and Fecal Putrefactive Products of Cats. J. Vet. Med. Sci. 1993;55:291–295. doi: 10.1292/jvms.55.291. [DOI] [PubMed] [Google Scholar]
- 186.Ballongue J., Schumann C., Quignon P. Effects of lactulose and lactitol on colonic microflora and enzymatic activity. Scand. J. Gastroenterol. Suppl. 1997;222:41–44. doi: 10.1080/00365521.1997.11720716. [DOI] [PubMed] [Google Scholar]
- 187.García-Cayuela T., Díez-Municio M., Herrero M., Martínez-Cuesta M.C., Peláez C., Requena T., Moreno F.J. Selective fermentation of potential prebiotic lactose-derived oligosaccharides by probiotic bacteria. Int. Dairy J. 2014;38:11–15. doi: 10.1016/j.idairyj.2014.03.012. [DOI] [Google Scholar]
- 188.Li W., Yu S., Zhang T., Jiang B., Stressler T., Fischer L., Mu W. Efficient Biosynthesis of Lactosucrose from Sucrose and Lactose by the Purified Recombinant Levansucrase from Leuconostoc mesenteroides B-512 FMC. J. Agric. Food Chem. 2015;63:9755–9763. doi: 10.1021/acs.jafc.5b03648. [DOI] [PubMed] [Google Scholar]
- 189.Maruyama T., Terada A., Ogata Y., Fujita K., Ishigami H., Hara K., Pet T., Co F., Kaijo T., Shiba B. Effective Dose of Lactosucrose on Fecal Flora and Fecal Metabolites of Humans vitro by bifidobacteria and lactobacilli, and to a limited degree by the The effects of dietary lactosucrose on the fecal flora and fecal metab- olites were studied in eight he. Bifidobact. Microflora. 1994;13:51–63. [Google Scholar]
- 190.Nath A., Haktanirlar G., Varga Á., Molnár M.A., Albert K., Galambos I., Koris A., Vatai G. Biological Activities of Lactose-Derived Prebiotics and Symbiotic with Probiotics on Gastrointestinal System. Medicina. 2018;54:18. doi: 10.3390/medicina54020018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Díez-Municio M., Herrero M., Olano A., Moreno F.J. Synthesis of novel bioactive lactose-derived oligosaccharides by microbial glycoside hydrolases. Microb. Biotechnol. 2014;7:315–331. doi: 10.1111/1751-7915.12124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Kihara M., Ohba K., Sakata T. Trophic effect of dietary lactosucrose on intestinal tunica muscularis and utilization of this sugar by gut microbes in red seabream Pagrus major, a marine carnivorous teleost, under artificial rearing. Comp. Biochem. Physiol. Part A Physiol. 1995;112:629–634. doi: 10.1016/0300-9629(95)02037-3. [DOI] [Google Scholar]
- 193.Kihara M., Sakata T. Production of short-chain fatty acids and gas from various oligosaccharides by gut microbes of carp (Cyprinus carpio L.) in micro-scale batch culture. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002;132:333–340. doi: 10.1016/S1095-6433(02)00029-6. [DOI] [PubMed] [Google Scholar]
- 194.Kihara M., Sakata T. Influences of incubation temperature and various saccharides on the production of organic acids and gases by gut microbes of rainbow trout Oncorhynchus mykiss in a micro-scale batch culture. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2001;171:441–447. doi: 10.1007/s003600100190. [DOI] [PubMed] [Google Scholar]
- 195.Kamel R., Afifi S.M., Kassem I.A.A., Elkasabgy N.A., Farag M.A. Arabinoxylan and rhamnogalacturonan mucilage: Outgoing and potential trends of pharmaceutical, environmental, and medicinal merits. Int. J. Biol. Macromol. 2020;165:2550–2564. doi: 10.1016/j.ijbiomac.2020.10.175. [DOI] [PubMed] [Google Scholar]
- 196.Hopkins M.J., Englyst H.N., Macfarlane S., Furrie E., Macfarlane G.T., McBain A.J. Degradation of Cross-Linked and Non-Cross-Linked Arabinoxylans by the Intestinal Microbiota in Children. Appl. Environ. Microbiol. 2003;69:6354–6360. doi: 10.1128/AEM.69.11.6354-6360.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Glitsø L.V., Gruppen H., Schols H.A., Højsgaard S., Sandström B., Bach Knudsen K.E. Degradation of rye arabinoxylans in the large intestine of pigs. J. Sci. Food Agric. 1999;79:961–969. doi: 10.1002/(SICI)1097-0010(19990515)79:7<961::AID-JSFA311>3.0.CO;2-1. [DOI] [Google Scholar]
- 198.Terada A., Hara H., Sakamoto J., Sato N., Takagi S., Mitsuoka T., Mino R., Hara K., Fujimori I., Yamada T. Effects of Dietary Supplementation with Lactosucrose (4G-β-D-Galactosylsucrose) on Cecal Flora, Cecal Metabolites, and Performance in Broiler Chickens. Poult. Sci. 1994;73:1663–1672. doi: 10.3382/ps.0731663. [DOI] [PubMed] [Google Scholar]
- 199.Finegold S.M., Li Z., Summanen P.H., Downes J., Thames G., Corbett K., Dowd S., Krak M., Heber D. Xylooligosaccharide increases bifidobacteria but not lactobacilli in human gut microbiota. Food Funct. 2014;5:436–445. doi: 10.1039/c3fo60348b. [DOI] [PubMed] [Google Scholar]
- 200.Chung Y.-C., Hsu C.-K., Ko C.-Y., Chan Y.-C. Dietary intake of xylooligosaccharides improves the intestinal microbiota, fecal moisture, and pH value in the elderly. Nutr. Res. 2007;27:756–761. doi: 10.1016/j.nutres.2007.09.014. [DOI] [Google Scholar]
- 201.Berger K., Burleigh S., Lindahl M., Bhattacharya A., Patil P., Stålbrand H., Nordberg Karlsson E., Hållenius F., Nyman M., Adlercreutz P. Xylooligosaccharides Increase Bifidobacteria and Lachnospiraceae in Mice on a High-Fat Diet, with a Concomitant Increase in Short-Chain Fatty Acids, Especially Butyric Acid. J. Agric. Food Chem. 2021;69:3617–3625. doi: 10.1021/acs.jafc.0c06279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Hsu C.K., Liao J.W., Chung Y.C., Hsieh C.P., Chan Y.C. Xylooligosaccharides and fructooligosaccharides affect the intestinal microbiota and precancerous colonic lesion development in rats. J. Nutr. 2004;134:1523–1528. doi: 10.1093/jn/134.6.1523. [DOI] [PubMed] [Google Scholar]
- 203.Roberfroid M.B., Van Loo J.A.E., Gibson G.R. The Bifidogenic Nature of Chicory Inulin and Its Hydrolysis Products. J. Nutr. 1998;128:11–19. doi: 10.1093/jn/128.1.11. [DOI] [PubMed] [Google Scholar]
- 204.Walker A.W., Duncan S.H., Carol McWilliam Leitch E., Child M.W., Flint H.J. pH and peptide supply can radically alter bacterial populations and short-chain fatty acid ratios within microbial communities from the human colon. Appl. Environ. Microbiol. 2005;71:3692–3700. doi: 10.1128/AEM.71.7.3692-3700.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Duncan S.H., Louis P., Thomson J.M., Flint H.J. The role of pH in determining the species composition of the human colonic microbiota. Environ. Microbiol. 2009;11:2112–2122. doi: 10.1111/j.1462-2920.2009.01931.x. [DOI] [PubMed] [Google Scholar]
- 206.Guariguata L., Whiting D.R., Hambleton I., Beagley J., Linnenkamp U., Shaw J.E. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res. Clin. Pract. 2014;103:137–149. doi: 10.1016/j.diabres.2013.11.002. [DOI] [PubMed] [Google Scholar]
- 207.Sherwin R.S., Anderson R.M., Buse J.B., Chin M.H., Eddy D., Fradkin J., Ganiats T.G., Ginsberg H., Kahn R., Nwankwo R., et al. The prevention or delay of type 2 diabetes. Diabetes Care. 2003;26((Suppl. 1)):S62–S69. doi: 10.2337/diacare.26.2007.s62. [DOI] [PubMed] [Google Scholar]
- 208.Nathan D.M. Long-term complications of diabetes mellitus. N. Engl. J. Med. 1993;328:1676–1685. doi: 10.1056/NEJM199306103282306. [DOI] [PubMed] [Google Scholar]
- 209.Ley R.E., Turnbaugh P.J., Klein S., Gordon J.I. Human gut microbes associated with obesity. Nature. 2006;444:1022–1023. doi: 10.1038/4441022a. [DOI] [PubMed] [Google Scholar]
- 210.Moya-Pérez A., Luczynski P., Renes I.B., Wang S., Borre Y., Anthony Ryan C., Knol J., Stanton C., Dinan T.G., Cryan J.F. Intervention strategies for cesarean section-induced alterations in the microbiota-gut-brain axis. Nutr. Rev. 2017;75:225–240. doi: 10.1093/nutrit/nuw069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Rabot S., Membrez M., Bruneau A., Gérard P., Harach T., Moser M., Raymond F., Mansourian R., Chou C.J. Germ-free C57BL/6J mice are resistant to high-fat-diet-induced insulin resistance and have altered cholesterol metabolism. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2010;24:4948–4959. doi: 10.1096/fj.10-164921. [DOI] [PubMed] [Google Scholar]
- 212.Yde C.C., Jensen H.M., Christensen N., Servant F., Lelouvier B., Lahtinen S., Stenman L.K., Airaksinen K., Kailanto H.-M. Polydextrose with and without Bifidobacterium animalis ssp. lactis 420 drives the prevalence of Akkermansia and improves liver health in a multi-compartmental obesogenic mice study. PLoS ONE. 2021;16:e0260765. doi: 10.1371/journal.pone.0260765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Snelson M., de Pasquale C., Ekinci E.I., Coughlan M.T. Gut microbiome, prebiotics, intestinal permeability and diabetes complications. Best Pract. Res. Clin. Endocrinol. Metab. 2021;35:101507. doi: 10.1016/j.beem.2021.101507. [DOI] [PubMed] [Google Scholar]
- 214.Tajabadi-Ebrahimi M., Sharifi N., Farrokhian A., Raygan F., Karamali F., Razzaghi R., Taheri S., Asemi Z. A Randomized Controlled Clinical Trial Investigating the Effect of Synbiotic Administration on Markers of Insulin Metabolism and Lipid Profiles in Overweight Type 2 Diabetic Patients with Coronary Heart Disease. Exp. Clin. Endocrinol. Diabetes Off. J. Ger. Soc. Endocrinol. Ger. Diabetes Assoc. 2017;125:21–27. doi: 10.1055/s-0042-105441. [DOI] [PubMed] [Google Scholar]
- 215.Li H.Y., Zhou D.D., Gan R.Y., Huang S.Y., Zhao C.N., Shang A., Xu X.Y., Li H. Bin Effects and mechanisms of probiotics, prebiotics, synbiotics, and postbiotics on metabolic diseases targeting gut microbiota: A narrative review. Nutrients. 2021;13:3211. doi: 10.3390/nu13093211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Vezza T., Abad-Jiménez Z., Marti-Cabrera M., Rocha M., Víctor V.M. Microbiota-mitochondria inter-talk: A potential therapeutic strategy in obesity and type 2 diabetes. Antioxidants. 2020;9:848. doi: 10.3390/antiox9090848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Kim Y.A., Keogh J.B., Clifton P.M. Probiotics, prebiotics, synbiotics and insulin sensitivity. Nutr. Res. Rev. 2018;31:35–51. doi: 10.1017/S095442241700018X. [DOI] [PubMed] [Google Scholar]
- 218.Pereira D.I.A., Gibson G.R. Effects of consumption of probiotics and prebiotics on serum lipid levels in humans. Crit. Rev. Biochem. Mol. Biol. 2002;37:259–281. doi: 10.1080/10409230290771519. [DOI] [PubMed] [Google Scholar]
- 219.Costa G.T., Guimarães S.B., Sampaio H.A.C. Fructo-oligosaccharide effects on blood glucose. An overview [Efeitos dos fruto-oligossacar{í}deos no controle glic{ê}mico. revis{ã}o] Acta Cir. Bras. 2012;27:279–282. doi: 10.1590/S0102-86502012000300013. [DOI] [PubMed] [Google Scholar]
- 220.Dehghan P., Pourghassem Gargari B., Asghari Jafar-abadi M. Oligofructose-enriched inulin improves some inflammatory markers and metabolic endotoxemia in women with type 2 diabetes mellitus: A randomized controlled clinical trial. Nutrition. 2014;30:418–423. doi: 10.1016/j.nut.2013.09.005. [DOI] [PubMed] [Google Scholar]
- 221.Letexier D., Diraison F., Beylot M. Addition of inulin to a moderately high-carbohydrate diet reduces hepatic lipogenesis and plasma triacylglycerol concentrations in humans. Am. J. Clin. Nutr. 2003;77:559–564. doi: 10.1093/ajcn/77.3.559. [DOI] [PubMed] [Google Scholar]
- 222.Aliasgharzadeh A., Khalili M., Mirtaheri E., Pourghassem Gargari B., Tavakoli F., Abbasalizad Farhangi M., Babaei H., Dehghan P. A Combination of Prebiotic Inulin and Oligofructose Improve Some of Cardiovascular Disease Risk Factors in Women with Type 2 Diabetes: A Randomized Controlled Clinical Trial. Adv. Pharm. Bull. 2015;5:507–514. doi: 10.15171/apb.2015.069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Jackson K.G., Taylor G.R.J., Clohessy A.M., Williams C.M. The effect of the daily intake of inulin on fasting lipid, insulin and glucose concentrations in middle-aged men and women. Br. J. Nutr. 1999;82:23–30. doi: 10.1017/S0007114599001087. [DOI] [PubMed] [Google Scholar]
- 224.Dewulf E.M., Cani P.D., Claus S.P., Fuentes S., Puylaert P.G.B., Neyrinck A.M., Bindels L.B., De Vos W.M., Gibson G.R., Thissen J.P., et al. Insight into the prebiotic concept: Lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut. 2013;62:1112–1121. doi: 10.1136/gutjnl-2012-303304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Schellekens H., Torres-Fuentes C., van de Wouw M., Long-Smith C.M., Mitchell A., Strain C., Berding K., Bastiaanssen T.F.S., Rea K., Golubeva A.V., et al. Bifidobacterium longum counters the effects of obesity: Partial successful translation from rodent to human. EBioMedicine. 2021;63:103176. doi: 10.1016/j.ebiom.2020.103176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Everard A., Lazarevic V., Derrien M., Girard M., Muccioli G.G., Neyrinck A.M., Possemiers S., Van Holle A., François P., de Vos W.M., et al. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes. 2011;60:2775–2786. doi: 10.2337/db11-0227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Sangwan V., Tomar S.K., Ali B., Singh R.R.B., Singh A.K. Hypoglycaemic effect of galactooligosaccharides in alloxan-induced diabetic rats. J. Dairy Res. 2015;82:70–77. doi: 10.1017/S0022029914000582. [DOI] [PubMed] [Google Scholar]
- 228.Vulevic J., Juric A., Tzortzis G., Gibson G.R. A mixture of trans-galactooligosaccharides reduces markers of metabolic syndrome and modulates the fecal microbiota and immune function of overweight adults. J. Nutr. 2013;143:324–331. doi: 10.3945/jn.112.166132. [DOI] [PubMed] [Google Scholar]
- 229.Albarracín M., Weisstaub A.R., Zuleta Á., Mandalunis P., González R.J., Drago S.R. Effects of extruded whole maize, polydextrose and cellulose as sources of fibre on calcium bioavailability and metabolic parameters of growing Wistar rats. Food Funct. 2014;5:804–810. doi: 10.1039/c3fo60424a. [DOI] [PubMed] [Google Scholar]
- 230.Maki K.C., Pelkman C.L., Finocchiaro E.T., Kelley K.M., Lawless A.L., Schild A.L., Rains T.M. Resistant starch from high-amylose maize increases insulin sensitivity in overweight and obese men. J. Nutr. 2012;142:717–723. doi: 10.3945/jn.111.152975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Bodinham C.L., Smith L., Thomas E.L., Bell J.D., Swann J.R., Costabile A., Russell-Jones D., Umpleby A.M., Robertson M.D. Efficacy of increased resistant starch consumption in human type 2 diabetes. Endocr. Connect. 2014;3:75–84. doi: 10.1530/EC-14-0036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Bianchi G., Ronchi M., Marchesini G. Effect of lactulose on carbohydrate metabolism and diabetes mellitus. Scand. J. Gastroenterol. Suppl. 1997;222:62–64. doi: 10.1080/00365521.1997.11720721. [DOI] [PubMed] [Google Scholar]
- 233.Lopez H.W., Levrat M.A., Guy C., Messager A., Demigné C., Rémésy C. Effects of soluble corn bran arabinoxylans on cecal digestion, lipid metabolism, and mineral balance (Ca, Mg) in rats. J. Nutr. Biochem. 1999;10:500–509. doi: 10.1016/S0955-2863(99)00036-4. [DOI] [PubMed] [Google Scholar]
- 234.Lu Z.X., Walker K.Z., Muir J.G., O’Dea K. Arabinoxylan fibre improves metabolic control in people with Type II diabetes. Eur. J. Clin. Nutr. 2004;58:621–628. doi: 10.1038/sj.ejcn.1601857. [DOI] [PubMed] [Google Scholar]
- 235.Imaizumi K., Nakatsu Y., Sato M., Sedarnawati Y., Sugano M. Effects of xylooligosaccharides on blood glucose, serum and liver lipids and-cecum short-chain fatty acids in diabetic rats. Agric. Biol. Chem. 1991;55:199–205. doi: 10.1080/00021369.1991.10870553. [DOI] [Google Scholar]
- 236.Kim S., Rhee I., Joo G., Ha H. Effects of dietary xylooligosaccharide on lipid levels of serum in rats fed high cholesterol diet. J. Korean Soc. Food Sci. Nutr. 1998;27:945–951. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.