

Abstract
Background
Current therapies for alcohol-induced liver injury are of limited efficacy and associated with significant side effects. With the proposed pathophysiology of alcohol-induced liver injury to be related to deranged gut microbiota, we hypothesized that probiotics would have beneficial effects in attenuating alcohol-induced liver injury.
Methods
Twenty-four male Sprague-Dawley rats were divided into 4 groups: control group, alcohol group, Lactobacillus plantarum group, and mixed-strain probiotics group. After 4 weeks, all rats were sacrificed, and blood samples were analyzed for ALT, lipopolysaccharide level (LPS), interleukin 6 (IL-6), and tumor necrosis factor-alpha (TNF-α). Liver tissues were processed for histopathology, malondialdehyde (MDA) level and immunohistochemistry for toll-like receptors 4 (TLR-4). Stool samples were collected, and 16S rRNA sequencing was used to analyze the fecal microbiota.
Results
Liver histopathology showed the presence of significant hepatocyte ballooning in the alcohol group as compared with the control group, and the treatment with L. plantarum or mixed-strain probiotics alleviated these changes. Significant elevation of serum ALT, LPS, IL-6, and TNF-α, hepatic MDA levels, and hepatic TLR-4 expression were observed in alcohol-fed rats as compared with control rats. The administration of L. plantarum or mixed-strain probiotics restored these changes to the levels of control rats. The relative abundance of fecal bacteria at genus level showed a significant reduction in Allobaculum, Romboutsia, Bifidobacterium, and Akkermansia in the alcohol group as compared with the control group. In probiotics-treated rats, significant increases in Allobaculum and Bifidobacterium were observed, while the relative abundance of Romboutsia and Akkermansia was unchanged compared to the alcohol group. A reduction in alpha diversity was observed in alcohol-treated rats, whereas the improvement was noted after probiotic treatment.
Conclusions
The treatment with Lactobacillus, whether as single-, or mixed-strain probiotics, was beneficial in reducing the severity of alcohol-induced liver injury likely through the increase in beneficial bacteria, and the reduction of inflammatory responses, and oxidative stress.
Supplementary Information
The online version contains supplementary material available at 10.1186/s12906-022-03643-9.
Keywords: Alcohol-induced liver injury, Rat model, Probiotics, Microbiota
Introduction
The burden of alcoholic liver disease (ALD) is substantial from both medical and financial aspects. Globally, in 2016, alcohol-attributable liver cirrhosis was responsible for 588,100 deaths and loss of 21,455,000 disability adjusted life years (DALYs) [1]. Alcohol is not only a problem in the Western world, but also in Thailand. The National Household Survey for Substance and Alcohol Use from 2007 reported the prevalence of current drinkers in Thailand of 26.7% with 6.7% of the Thai population being hazardous drinkers [2].
The pathological spectrum of ALD ranges from simple steatosis, steatohepatitis, progressive fibrosis to cirrhosis [3]. Acute alcoholic hepatitis represents an acute on chronic condition in patients with ALD. This condition, if severe, can be associated with high mortality rates up to 65% [4]. The currently available medical therapies for alcoholic hepatitis have limited efficacy. Steroid, which is a mainstay of treatment for severe alcoholic hepatitis, carries significant adverse effects especially the infection risk [5]. There has been an ongoing search for a safe and effective treatment for alcoholic hepatitis but none has achieved such a goal.
Alcohol can alter gut microbiota and cause bacterial overgrowth leading to mucosal inflammation, increased mucosal permeability and bacterial translocation. This derangement in mucosal integrity leads to increased exposure of liver tissues to lipopolysaccharides (LPS) aka endotoxin. LPS then stimulates innate immune receptors, such as Toll-like receptors (TLRs) leading to hepatic stellate cell and Kupffer cell activation. This process starts inflammatory cascades that result in liver injury and fibrosis [6]. Taking the role of gut dysbiosis in ALD into consideration, probiotics appear to be attractive options for the prevention or treatment of alcohol-induced liver injury.
Lactobacillus spp. have been shown in both in vitro and in vivo studies to detoxify alcohol before absorption [7], improve gut epithelial dysfunction [8], reduce intestinal permeability, decrease LPS exposure to the liver and prevent/reduce liver inflammation [9]. A variety of Lactobacillus strains have been used, such as Lactobacillus rhamnosus GG, Lactobacillus acidophilus, Lactobacillus helveticus, heat-killed Lactobacillus brevis SBC8803, and Lactobacillus rhamnosus GG supernatant [10]. In this study, we aimed to evaluate the effect of Lactobacillus plantarum B7 (L. plantarum) on alcohol-induced liver injury in an animal study. Despite being studied in other conditions, such as Helicobacter pylori infection [11, 12], pancreatitis, and post-liver transplantation, L. plantarum has never been tested in alcohol-induced liver injury. Furthermore, L. plantarum has been demonstrated in in vitro studies to exert immunomodulating effects more strongly than other strains of Lactobacillus [12–14]. Moreover, a recent in vitro study from our group showed that the combination of Lactobacillus rhamnosus L34 (L. rhamnosus L34) and Lactobacillus casei L39 (L. casei L39) could inhibit Clostridioides difficile-induced IL-8 and GM-CSF production from HT-29 cells and suppress the activation of phosphorylated NF-kβ [15]. L. rhamnosus L34 has also been shown to inhibit TLR-4 activation and attenuate the severity of gut leakage in a sepsis model [16]. With this knowledge, we hypothesized that L. plantarum and the combination of L. rhamnosus L34 and L. casei L39 might have beneficial effects in reducing alcohol-related liver damage and serve as an alternative treatment or a preventive measure for alcohol-induced liver injury.
The aims of this study were to evaluate the effect of L. plantarum and the combination of L. rhamnosus L34 and L. casei L39 (mixed-strain probiotics) on liver pathology in a rat model of alcohol-induced liver injury and to determine whether probiotics exerted their treatment effects through gut microbiota related changes.
Materials and methods
Experimental protocol
This study was conducted according to the Ethical Principles and Guidelines for the Use of Animals by the National Research Council of Thailand and reported in accordance with Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines. The study protocol was approved by the Animal Care and Use Committee, the Faculty of Medicine, Chulalongkorn University (the permission No. is 035/2561).
As LPS was one of the outcomes of interest in this study, we used LPS levels from a study by Wang and colleagues [17] which also evaluated the effect of probiotics in alcohol-related liver disease to calculate our sample size as described below.
N is a sample size, Zα/2 = 1.96 (based on Z table with the probability of falsely rejecting a true null hypothesis (α) = 0.05), Zβ = 0.84 (based on Z table with the probability of failing to reject a false null hypothesis (β) = 0.80), σ2 = pooled variance, = mean LPS level in alcohol group (0.37 mg/L), = mean LPS level in alcohol + probiotics group (0.17), n1 = sample size in alcohol group (10), s1 = standard deviation in alcohol group (0.17), n2 = sample size in alcohol + probiotics group (10), s2 = standard deviation in alcohol + probiotics group (0.04). Using the above formulas, the sample size in each group was 6.
A total of 24 male Sprague-Dawley rats aged 8 weeks (180-220 g) were purchased from the Nomura Siam International Co., Ltd., Bangkok, Thailand. Rats were housed in a standard room with controlled temperature of 25 ± 1 °C and a 12-hour light-dark cycle. All rats were fed ad libitum with the diet containing 35% of energy from fat, 18% from protein, and 47% from carbohydrate for 4 weeks. The animals were randomly divided into 4 groups (n = 6 in each group):
Control group, rats were fed with distilled water (2.0 mL) via an intragastric tube once a day for 4 weeks.
Alcohol group, rats were fed with 50% ethanol (6.5 g/kg/d) via an intragastric tube twice a day for 4 weeks.
Alcohol and L. plantarum group, rats were fed with 50% ethanol (6.5 g/kg/d) via an intragastric tube twice a day for 4 weeks. Simultaneously, rats were fed with 1 mL of L. plantarum B7 (1 × 108 CFU/mL) through oral gavage once a day for 4 weeks. Probiotics were given 4 hours after the time of alcohol administration to ensure the survival of bacteria.
Alcohol and mixed-strain probiotics, rats were fed with 50% ethanol (6.5 g/kg/d) via an intragastric tube twice a day for 4 weeks. Simultaneously, rats were fed with 1 mL of L. rhamnosus L34 and L. casei L39 (1 × 108 CFU/mL for each bacteria) through oral gavage once a day for 4 weeks. Probiotics were given 4 hours after the time of alcohol administration to ensure the survival of bacteria.
Alcohol and probiotic dosages used in this study were adopted from the results of a pilot study by our team. We did not combine L. plantarum with other strains due to the preliminary research from our laboratory indicating the inhibitory effect of L. plantarum on the growth of other Lactobacillus.
At the end of 4 weeks, all rats were euthanized. Blood and liver samples were collected. Stool samples were obtained at the end of the study for fecal microbiota analysis. Blood samples were analyzed for aspartate aminotransferase (AST), alanine aminotransferase (ALT), lipopolysaccharide level (LPS), interleukin 6 (IL-6), and tumor necrosis factor-alpha (TNF-α). Liver tissues were processed for histopathological exam, malondialdehyde (MDA) levels, and immunohistochemistry for toll-like receptor 4 (TLR-4).
AST and ALT measurement
Blood samples were obtained through cardiac puncture. Serum was then separated by centrifuging the blood at 1560 x g for 30 minutes at 4 °C. ALT and AST levels were measured using Reflotron® Plus. ALT and AST in the serum reacted with reagents within the machine to create color. Reflectance photometer possessed light-emitting diodes (LEDs) that emitted at the key wavelengths of 567, 642 and 951 nm onto the reagent strip. The machine then measured the pattern of reflection according to its depth of color, which was proportional to the concentration of analyte in the sample. The level was expressed as IU/L.
Serum lipopolysaccharide, IL-6 and TNF-α measurement
LPS levels were measured using the Pierce™ Chromogenic Endotoxin Quantitative Kit (Thermo Fisher Scientific, Massachusetts, USA) according to the manufacturer’s protocol. This colorimetric assay uses amebocyte lysates from the blood of horseshoe crab to quantitate endotoxin levels in the serum. Serum LPS levels were expressed as endotoxin units (EU)/mL. Serum levels of IL-6 and TNF-α were measured using commercially available ELISA kits for each cytokine according to the manufacturer’s protocols (R&D Systems, Minneapolis, USA). Both serum IL-6 and TNF-α were expressed as pg/mL.
Hepatic malondialdehyde (MDA) measurement
Tissue homogenization was performed by placing one gram of liver tissue in radioimmunoprecipitation assay buffer (RIPA buffer) and sonicating on ice for 15 seconds. Samples were then centrifuged at 1600 x g for 10 minutes at 4 °C to obtain supernatants. MDA levels were measured from supernatants using a commercial assay kit (Cayman Chemical Company, Michigan, USA) according to the manufacturer’s manual. The test measured the production rate of thiobarbituric acid-reactive substances (TBARS) under an acidic condition at the temperature of 95 °C. The optical density (OD) of supernatant was read at 532 nm wavelength. MDA levels were then obtained by applying the OD reading to a standard curve and expressed as nmol/mg protein.
Liver histopathology
Liver tissues were fixed in 10% formaldehyde for 24-48 hours, embedded in paraffin cassette, and cut into 3-μm-thick slides with microtome. The slides were then deparaffinized and strained with Hematoxylin and Eosin (H&E). Since there was no unified histological scoring system for acute alcoholic hepatitis, we decided to adopt the histological criteria for non-alcoholic steatohepatitis. An experienced pathologist, who was blinded to the experimental groups, examined and graded the severity of pathological changes based on the histopathological scores proposed by Brunt and colleagues [18] which are described below. A total of 5 slices were reviewed in each group.
Hepatic steatosis is scored as follows
0 = less than 5% of total hepatocytes contain fat
1 = 5-33% of total hepatocytes contain fat
2 = 33-66% of total hepatocytes contain fat
3 = more than 66% of total hepatocytes contain fat
Lobular inflammation is scored as follows
1 = None
2 = less than 2 inflammatory foci per 200x field
3 = 2-4 inflammatory foci per 200x field
4 = more than 4 inflammatory foci per 200x field
Hepatic ballooning is scored as follows
0 = None
1 = minimal ballooning
2 = prominent ballooning
Immunohistochemistry for hepatic TLR-4 expression
After examining H&E slides under light microscopy, the pathologist chose the best location for immunohistochemical study. The location on the slide was compared with the location on a paraffin block. The desired area was cut out using a metal cylinder of 4-mm diameter. Each piece was orderly placed in tissue microarray block and labeled. Three 3 μm-thick slides were sliced from the tissue microarray block (1 slide for immunohistochemistry for TLR-4 and 1 slide for H&E stain to confirm the location).
Slides were treated with citrate buffer at pH 6.0 and heated in a microwave for 13 minutes for the antigen retrieval. Endogenous peroxidase activity and nonspecific binding were blocked by incubating slides with 3% hydrogen peroxide for 5 minutes and with 3% normal horse serum for 20 minutes, respectively. After being washed in PBS solution, slides were incubated with primary antibodies for TLR-4 (Affinity Biosciences, Ohio, USA) at the dilution of 1:50 for 60 minutes at a room temperature. After another PBS washing, sections were incubated with the specific secondary antibody for 30 minutes at a room temperature. Following the development of brownish color by diaminobenzidine (DAB), hematoxylin counterstain was performed.
TLR-4 positive cells were Kupffer cells with brown stained cytoplasm. Immunohistochemical slides were scanned using the Aperio ScanScope System (Aperio Technology, California, USA). The ImageScope software (Aperio Technology, California, USA) was used to analyze and quantify TLR-4 positivity. Percent positivity was calculated by the number positive pixels divided by the summation of both positive and negative pixels × 100. Positive and negative pixels were set by using positive and negative control tissues [19].
Fecal microbiota
Fresh stool samples were collected from each rat at the end of the study. Stool collection was performed by separating each rat in a clean container lined with pathogen-free plastic to avoid contamination. Stool samples were then stored at − 80 °C until analysis.
Stool samples were processed and analyzed with the ZymoBIOMICS® Service: Targeted Metagenomic Sequencing (Zymo Research, Irvine, CA). DNA extraction was performed using ZymoBIOMICS®-96 MagBead DNA Kit (Zymo Research, Irvine, CA) according to the manufacturer’s instructions and amplified at the V3-V4 regions of 16 s rRNA gene using polymerase chain reaction (PCR) method. The pooled sequencing library was cleaned up with the Select-a-Size DNA Clean & Concentrator™ (Zymo Research, Irvine, CA), then quantified with TapeStation® (Agilent Technologies, Santa Clara, CA) and Qubit® (Thermo Fisher Scientific, Waltham, WA). DNA sequencing was then performed using Illumina® MiSeq™ with a v3 reagent kit (600 cycles) with 10% PhiX spike-in. Taxonomy assignment was performed using Uclust from QIIME v.1.9.1. Taxonomy was assigned with the Zymo Research Database, a 16S database that is internally designed and curated, as reference. QIIME v.1.9.1 was also used to calculate composition visualization, alpha-diversity, and beta-diversity.
Statistical analyses
The comparison of serum levels of ALT, AST, LPS, IL-6, TNF-α, and hepatic MDA among groups were performed using One-way ANOVA with post-hoc LSD. Serum ALT levels and summation of histopathological scores were compared between groups using Kruskal-Wallis test. In each component of histological changes (steatosis, lobular inflammation, hepatocyte ballooning), we compared the proportion of rats with each score between groups using Fisher’s exact test. Alpha-diversity and Beta-diversity of bacteria were tested using independent T-test and Permutational multivariate analysis of variance (PERMANOVA), respectively. Relative abundance of bacterial composition among groups were compared using nonparametric Mood’s median test. P-value of less than 0.05 was considered statistically significant. All analyses were performed using SPSS version 17 for Windows. Data were presented as median (interquartile range) for microbial composition data and mean ± SD for other variables.
Results
Effects of alcohol and probiotics on liver histopathology
In alcohol-fed rats, significant hepatocyte ballooning was seen on liver histopathology (p = 0.01) with a minimal increase in hepatic steatosis and lobular inflammation compared with the control group (Fig. 1 and Table 1). After treatment with L. plantarum or mixed-strain probiotics, significant reductions in the severity of hepatocyte ballooning were observed in both groups as compared with the alcohol group p < 0.05 for both comparisons). Hepatic steatosis and lobular inflammation scores did not significantly differ between alcohol and treatment groups. It is important to note, however, that steatosis observed in both treatment groups was mostly microvesicular steatosis as compared with macrovesicular steatosis in the alcohol group. Mean summation of pathology scores in control, alcohol, L. plantarum, and mixed-strain groups were 1.3 ± 0.3, 3.3 ± 0.6, 1.3 ± 0.4, 1.7 ± 0.2, respectively (p = 0.004 between control and alcohol groups, p = 0.005 between alcohol and L. plantarum groups, and p = 0.03 between alcohol and mixed strain groups).
Fig. 1.

Liver histopathology in all groups. These are images of liver histopathology using H&E straining at 400x magnification (A, Control group; B, alcohol group; C, L. plantarum group; D, Mixed strain group). Black arrows indicate fat deposition inside hepatocytes and red arrows indicate hepatocyte ballooning
Table 1.
Histopathological scores in all groups
| Group | Steatosis | Inflammation | Ballooning | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 0 | 1 | 2 | 3 | 0 | 1 | 2 | |
| Control | 6 (100%) | – | – | – | – | 5 (83.3%) | 1 (16.7%) | – | 5 (83.3%) | 1 (16.7%) | – |
| Alcohol | 5 (83.3%) | 1 (16.7%) | – | – | – |
3 (50%) |
2 (33.3%) | 1 (16.7%) | – | 3 (50%) | 3 (50%) |
| Alcohol + L. plantarum | 4 (66.7%) | 2 (33.3%) | – | – | 1 (16.7%) | 4 (66.7%) | 1 (16.7%) | – | 6 (100%) | – | – |
| Alcohol + Mixed strains | 5 (83.3%) | 1 (16.7%) | – | – | – | 6 (100%) | – | – | 3 (50%) | 3 (50%) | – |
Values indicate the number of rats with that score in each group
As shown in Fig. 2, TLR-4 positivity was significantly higher in the alcohol group as compared with the control group (% positivity of 2.25 ± 0.77% vs. 0.12 ± 0.12%, respectively, p = 0.02). The administration of mixed-strain probiotics significantly reduced TLR-4 positivity as compared with the alcohol group (% positivity of 0.21 ± 0.10% vs. 2.25 ± 0.77%, respectively, p = 0.02). Although TLR-4 positivity in the L. plantarum group was lower than the alcohol group, the difference was not statistically significant (% positivity of 1.33 ± 0.88% vs. 2.25 ± 0.77%, respectively, p = 0.28). TLR-4 positivity in both treatment groups was similar to that in the control group.
Fig. 2.

Immunohistochemical study and percent positivity for TLR-4 in each group. A. Images of immunohistochemical study for TLR-4. These are images of immunohistochemical study for TLR-4 at 200x magnification (A, Control group; B, alcohol group; C, L. plantarum group; D, Mixed strain group). Red arrows indicate TLR-4 positive cells. B. Bar graph representing percent positivity of TLR-4 in each group. a = p-value< 0.05, ns = non-significant
Effects of alcohol and probiotics on liver enzymes, inflammatory markers and oxidative stress marker
As shown in Table 2 and Supplementary Figs. 1 and 2, significant increases in both ALT and AST were observed in the alcohol group as compared with the control group (ALT, 46.5 ± 5.6 vs. 32.3 ± 1.9 IU/L, respectively, p = 0.02 and AST, 241.2 ± 11.0 vs. 152.0 ± 21.0 IU/L, respectively, p < 0.001). L. plantarum administration significantly reduced both serum ALT (28.2 ± 6.4 IU/L) and AST (125.5 ± 13.4 IU/L) with p-value of less than 0.01 for both parameters when compared with the alcohol group. Likewise, the treatment with mixed-strain probiotics significantly decreased both serum ALT (23.0 ± 2.1 IU/L) and AST (125.6 ± 11.8 IU/L) with p-value of less than 0.01 for both parameters when compared with the alcohol group. ALT and AST levels in both treatment groups were similar to those in the control group.
Table 2.
Changes in serum and liver parameters in all groups
| Parameters | Control | Alcohol | Alcohol + L. plantarum | Alcohol + Mixed strains | p-value |
|---|---|---|---|---|---|
| ALT (IU/L) | 32.3 ± 1.9 | 46.5 ± 5.6 | 28.2 ± 6.4 | 23.0 ± 2.1 | 0.04*, 0.009**, 0.001# |
| AST (IU/L) | 152.0 ± 21.0 | 241.2 ± 11.0 | 125.5 ± 13.4 | 125.6 ± 11.8 | < 0.001*,**,# |
| TNF-α (pg/mL) | 114.1 ± 2.9 | 128.6 ± 0.9 | 115.4 ± 1.5 | 114.7 ± 1.7 | < 0.001*,**,# |
| IL-6 (pg/mL) | 267.8 ± 19.7 | 342.2 ± 15.8 | 275.3 ± 27.3 | 285.8 ± 10.6 | 0.01*, 0.02**, 0.04# |
| MDA (nmol/mg protein) | 1.34 ± 0.05 | 1.52 ± 0.04 | 1.32 ± 0.04 | 1.38 ± 0.03 | 0.005*, 0.002**, 0.03# |
| LPS (EU/mL) | 12.9 ± 4.1 | 27.8 ± 3.0 | 16.4 ± 3.4 | 18.8 ± 3.9 | 0.009*, 0.04**, 0.09# |
All parameters are presented as mean ± SD, *p-value between control and alcohol groups, **p-value between alcohol and L. plantarum groups, #p-value between alcohol and mixed strain groups
As illustrated in Table 2 and Fig. 3A, B, rats in the alcohol group had significantly higher serum TNF-α and IL-6 levels when compared with rats in the control group (TNF-α, 128.6 ± 0.9 vs. 114.1 ± 2.9 pg/mL, respectively, p < 0.001 and IL-6, 342.2 ± 15.8 vs. 267.8 ± 19.7 pg/mL, respectively, p = 0.01). L. plantarum-fed rats had significantly lower serum TNF-α and IL-6 levels than those fed with only alcohol (TNF-α, 115.4 ± 1.5 vs. 128.6 ± 0.9 pg/mL, respectively, p < 0.001 and IL-6, 275.3 ± 27.3 vs. 342.2 ± 15.8 pg/mL, respectively, p = 0.02). Similarly, the administration of mixed-strain probiotics significantly reduced serum TNF-α and IL-6 levels as compared with the alcohol group (TNF-α, 114.7 ± 1.7 vs. 128.6 ± 0.9 pg/mL, respectively, p < 0.001 and IL-6, 285.8 ± 10.6 vs. 342.2 ± 15.8 pg/mL, respectively, p = 0.04). Serum TNF-α and IL-6 levels in both treatment groups were not different from those in the control group.
Fig. 3.
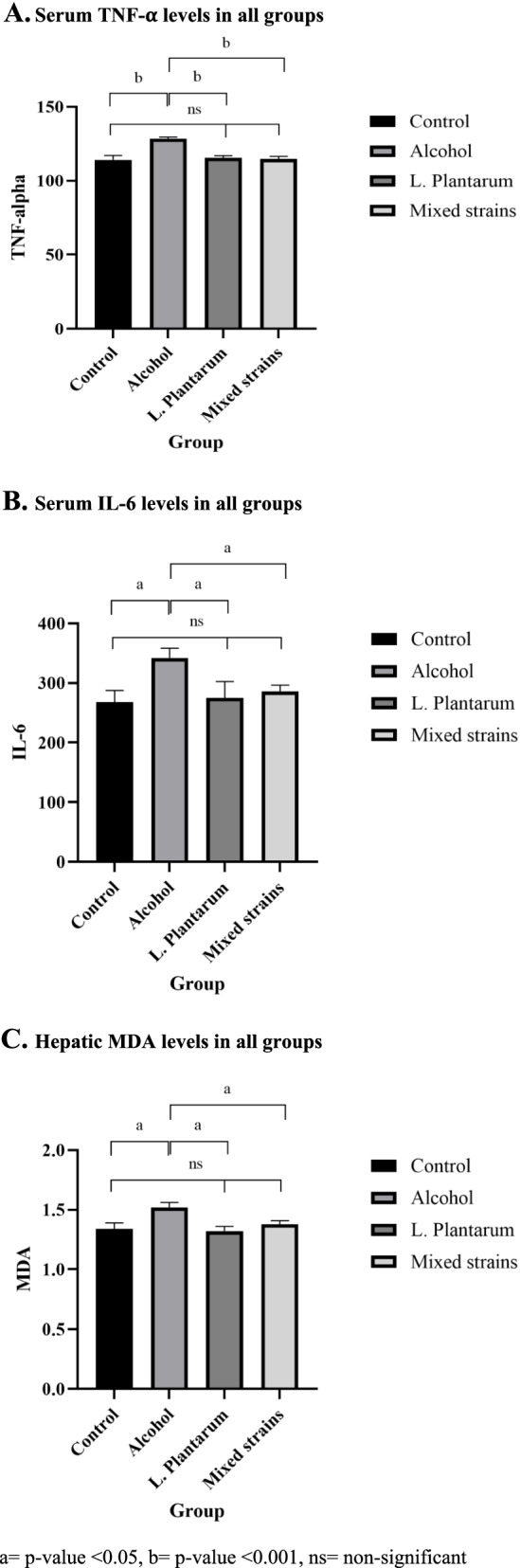
Bar graphs representing liver enzyme, inflammatory marker and hepatic MDA levels in all groups. A. Serum TNF-α levels in all groups. B. Serum IL-6 levels in all groups. C. Hepatic MDA levels in all groups. a = p-value < 0.05, b = p-value < 0.001, ns = non-significant
Levels of hepatic MDA, an oxidative stress marker, significantly increased in the alcohol group as compared with the control group as shown in Table 2 and Fig. 3C (1.52 ± 0.04 vs. 1.34 ± 0.05 nmol/mg protein, respectively, p = 0.005). The administration of L. plantarum and mixed-strain probiotics both decreased hepatic MDA levels significantly as compared with the alcohol group (1.32 ± 0.04 vs. 1.38 ± 0.03 vs. 1.52 ± 0.04 nmol/mg protein, respectively, p = 0.002 for the comparison between L. plantarum and alcohol groups, and p = 0.03 for the comparison between mixed strain and alcohol groups). Hepatic MDA levels in both treatment groups did not significantly differ from those in the control group.
Effects of alcohol and probiotics on serum endotoxin and fecal microbiota
As shown in Table 2 and Supplementary Fig. 3, serum LPS levels were significantly higher in the alcohol group as compared with the control group (27.8 ± 3.0 vs. 12.9 ± 4.1 EU/mL, respectively, p = 0.009). Rats that received L. plantarum had significantly lower levels of serum LPS as compared with the alcohol group (16.4 ± 3.4 vs. 27.8 ± 3.0 EU/mL, respectively, p = 0.04). In the mixed-strain group, serum LPS levels were lower than those in the alcohol group, albeit not statistically significant (18.8 ± 3.9 vs. 27.8 ± 3.0 EU/mL, respectively, p = 0.09). Serum LPS levels in both treatment groups were not different from those in the control group.
We observed that the alpha diversity was highest in the control group (4.37 ± 0.13) and lowest in the alcohol group (3.71 ± 0.19) (Fig. 4A). The alpha diversity was significantly increased after treatment with probiotics (4.14 ± 0.12 for L. plantarum, p = 0.001 and 4.21 ± 0.25 for mixed strains, p = 0.003), when compared with the alcohol group. We also noted that the beta diversity of the control group significantly differed from alcohol and treatment groups (PERMANOVA; p = 0.014), whereas the beta diversity was similar between the two treatment groups but slightly different from the alcohol group (Fig. 4B). Moreover, we observed the taxonomic changes in alcohol-fed rats as compared with control rats (shown in Suppl Fig. 4A-D). At the phylum level, our results showed a significant reduction in Verrucomicrobia [0.04%(0.31) vs. 4.30%(3.90), adjusted p-value = 0.021], and a significant increase in Actinobacteria [10.40%(5.90) vs. 0.50%(0.30), adjusted p-value = 0.003] in the alcohol group as compared with the control group, respectively. The most abundance phyla, Firmicutes were also slightly more enriched in the control group as compared with the alcohol group [79.38%(9.30) vs. 75.25%(4.60), respectively). However, the administration of L. plantarum and mixed-strain probiotics did not result in significant changes in fecal microbial composition as compared with the alcohol group at the phylum level. We further explored whether the microbial community at the genus level was altered after probiotic treatment. In alcohol-fed rats, there was a significant decline in the relative abundance of the genera Akkermansia [0%(0.3) vs. 4.30%(3.90), adjusted p-value = 0.003], Bifidobacterium [0%(0) vs. 8.90%(5.70) adjusted p-value = 0.003], Romboutsia [2.80%(3.90) vs. 25.0%(9.60), adjusted p-value = 0.021], and Allobaculum [4.90%(6.10) vs. 21.80%(6.14), adjusted p-value = 0.003] and an increase in the relative abundance of the genera Marvinbryantia [2.20%(4.90) vs. 0%(0.3), adjusted p-value = 0.021] as compared with rats in the control group, respectively. Moreover, our results showed a significant increase in Allobaculum [32.30%(10.00) in the L. plantarum group and 33.60%(9.80) in the mixed strain group], compared to 4.90%(6.10) in the alcohol group (adjusted p-value = 0.003) and an increase in Bifidobacterium [9.80%(3.80) in the L. plantarum group and 10.50%(4.9) in the mixed strain group], compared to 0%(0) in the alcohol group (adjusted p-value = 0.003).
Fig. 4.

Alpha-diversity (A) and Beta-diversity (B) of gut microbiota in all groups
Discussion
In this study, a daily intragastric administration of 6.5 g/kg/d of alcohol was able to induce liver injury as evidenced by the presence of significant hepatocyte ballooning, micro- and macrovesicular steatosis, and lobular inflammation in conjunction with ALT and AST elevation in alcohol-fed rats. Evidence have shown that oxidative stress plays a role in the development of alcoholic liver disease [20]. Chronic alcohol exposure leads to the increased production of reactive oxygen species as byproducts of alcohol metabolism in hepatocytes and as a result of Kupffer cell activation [20–22]. Similar to other studies, we also found the increase in hepatic MDA levels, an oxidative stress marker, in alcohol-fed rats compared with control rats.
Apart from its effects on the liver, alcohol can activate nuclear factor kappa B (NF-kB) in intestinal cells leading to nitric oxide production and oxidative stress which in turn disrupt intestinal integrity [9, 23, 24]. Alcohol exposure also reduces tight junction protein distribution [17]. As a result of increased intestinal permeability, bacterial translocation occurs. In this study, we demonstrated a significant rise in serum LPS levels, a surrogate marker of bacterial translocation, in alcohol-fed rats as compared with control rats. Our result was consistent with the reports by Grander et al. and Bull-Otterson et al [25, 26]. In conjunction with increased serum LPS levels, the up-regulation of hepatic TLR-4 expression was also observed in alcohol-fed rats in this study. Prior in vitro and in vivo studies have shown that the recognition of LPS by TLR-4 and its co-receptors activates mitogen-activated protein kinase (MAPK) and NF-kB pathways leading to the release of inflammatory cytokines, such IL-6 and TNF-α [13, 27]. Hritz and colleagues demonstrated the increased expression of IL-6, TNF-α, and TLR-4 in alcohol-fed wild-type mice, while these changes were not observed in TLR-4 knock-out mice [27]. Similar to their findings, alcohol-fed rats in our study exhibited higher levels of serum IL-6 and TNF-α than those of control rats.
Fecal microbiota analysis in this study indicated the decrease in absolute abundance and alpha-diversity in alcohol-fed rats as compared with control rats. Similarly, Bull-Otterson and colleagues showed the decrease in bacterial alpha diversity over time in alcohol-fed mice [25]. In contrast to our results, Yan and colleagues demonstrated the increase in total bacterial load in the cecum of alcohol-fed mice using quantitative PCR method, while Lowe and colleagues found the alpha diversity of cecal microbiota to be unaffected by alcohol exposure [28, 29]. Alcohol exposure in our study led to the decrease in relative abundance of phyla Verrucomicrobia, and the increase in relative abundance of phyla Proteobacteria and Actinobacteria. In alcohol-fed rats, there was a decline in the relative abundance of the genera Akkermansia, Bifidobacterium, Romboutsia, and Allobaculum. Similar to our findings, Lowe and colleagues observed the significant enrichment of the phylum Actinobacteria and the reduced abundance of the phylum Verrucomicrobia in alcohol-fed mice. The authors also identified the reduction of genus Akkermansia to be an early marker of alcohol-induced gut microbiota changes [29]. Moreover, Grander et al. found that patients with alcoholic steatohepatitis had the reduced abundance of fecal Akkermansia muciniphila and oral supplement of A. muciniphila could prevent alcohol-induced liver injury in an experimental model of alcoholic liver disease [26]. Akkermansia muciniphila has been shown to increase gut microbiota diversity through short chain fatty acid production and strengthen intestinal barrier function by increasing mucin layer thickness and tight junction protein expression [30, 31]. Bull-Otterson and colleagues reported a reduction in both Bacteroidetes and Firmicutes and an expansion of Proteobacteria and Actinobacteria, while we did not observe significant changes in the relative abundance of either Bacteroidetes or Firmicutes in this study [25]. Differences in the effects of alcohol on bacterial diversity and taxonomic shifts among studies could be from dissimilarities in the source of microbiota (fecal vs. cecal source), animal species (mouse vs. rat), and the type and amount of alcohol exposure (acute vs. chronic vs. binge feeding).
The administration of L. plantarum or the combination of L. rhamnosus L34 and L. casei L39 (mixed-strain probiotics) could reduce the severity of alcohol-induced liver injury, particularly the degree of hepatocyte ballooning, in this study. In conjunction with the histological improvement, we found significant reductions in serum ALT, inflammatory markers (TNF-α and IL-6), and oxidative stress marker (MDA) in probiotics-treated rats. One of the mechanisms of probiotics in the prevention of alcohol-induced liver injury was likely through the reduction of intestinal permeability and bacterial translocation as evidenced by lower levels of serum LPS in probiotics-treated rats. An in vitro study using Caco-2 cells demonstrated that L. plantarum inhibited TNF-α-induced activation of ERK pathway and TNF-α-induced degradation of IkB-α thus preserving intestinal epithelial barrier function [8]. Bull-Otterson et al. found that L. rhamnosus GG (LGG) supplement corrected the reduction in tight junction protein expression, which corresponded with the reduction in plasma LPS levels and hepatic TNF-α expression [25]. In accordance with our results, Forsyth et al. showed that L. rhamnosus supplement improved alcohol-induced steatohepatitis especially the necroinflammatory components along with the reduction in whole gut intestinal permeability, and alcohol-induced oxidative stress in the liver and intestines [9].
The supplement with L. plantarum and mixed-strain probiotics increased the relative abundance of the genera Allobaculum and Bifidobacterium but did not alter other beneficial bacteria. We suspected that probiotics alone might not be sufficient to completely restore the changes in bacterial composition from alcohol exposure. Fecal microbiota transplantation might be required to achieve such a purpose. In agreement with our findings, Bull-Otterson et al. demonstrated that the bacterial composition of alcohol+LGG supplement group was closer to that of the alcohol-fed group than the pair-fed group, although the changes were less prominent in the alcohol+LGG group than the alcohol-fed one. Additional analyses from our study demonstrated that Allobaculum was negatively correlated with the levels of ALT, AST, TNF-α, and MDA suggesting that Allobaculum may be protective against inflammation and oxidative stress. The increase in relative abundance of Allobaculum in both treatment groups might explain the anti-inflammatory effect of probiotics, possibly through the action of short-chain fatty acids produced by Allobaculum [32]. Moreover, the increased abundance of Bifidobacterium, which has previously been shown to improve intestinal integrity [33], might explain the reduction of serum endotoxin and TLR-4 expression in probiotics-treated rats. A mouse model of colitis associated colon cancer showed that probiotic supplement alleviated colitis through the increased abundance of the genera Lactobacillus, Bifidobacterium, and Allobaculum [34].
L. plantarum might also provide protection against alcohol-induced liver injury through other mechanisms. Apart from its effects on intestinal barrier function [8], L. plantarum may attenuate the severity of alcohol-induced liver injury through the reduction in lipid production and LPS-induced inflammatory response in hepatocytes. An in vitro study demonstrated that L. plantarum reduced lipid accumulation in LPS-induced HepG2 cells and its effect was more pronounce than other strains of lactic acid bacteria. Moreover, L. plantarum attenuated the LPS-induced expression of IL-6 and TNF-α by modulating the negative regulators of TLRs and down-regulating p38 MAPK and p65 NF-kB phosphorylation [13]. Furthermore, Lactobacillus spp., such as Lactobacillus fermentum MG590 has been shown to possess alcohol dehydrogenase and acetaldehyde dehydrogenase activities, and L. fermentum MG590 supplement improved hepatocyte viability in alcohol-containing medium in vitro and reduced blood alcohol concentration in alcohol-fed rats [7]. This direct detoxification effect of Lactobacillus reduced the alcohol exposure to the liver thus alleviating the severity of ALD.
However, our study was not without limitations. Since we did not perform fecal microbiota analysis at baseline, we could not state with certainty that fecal microbial changes were not related to temporal shift [35]. However, all rats in this study were purchased from the same vendor at the same time, were housed in the same environment, and were fed the same diet. Therefore, we assumed the fecal microbiota to be similar at baseline in each rat with similar environmental exposure.
Conclusion
Alcohol exposure in rats led to gut dysbiosis, increased bacterial translocation, oxidative stress, liver inflammation, and liver injury. Administration of Lactobacillus plantarum or the combination of Lactobacillus rhamnosus L34 and Lactobacillus casei L39 reduced the severity of alcohol-induced liver injury likely through the strengthening of intestinal integrity, the increase in beneficial bacteria, the anti-inflammatory and anti-oxidant effects.
Supplementary Information
Acknowledgements
We would like to thank the Center of Excellence in Hepatitis and Liver Cancer, Faculty of Medicine Chulalongkorn University and Miss. Thananya Jinato for data visualization in gut microbiota study.
Authors’ contributions
M.C. acquired the funding, conducted the study, performed laboratory tests and statistical analyses, wrote the original draft, and revised the manuscript; K.S. conducted the study, performed laboratory tests, and revised the manuscript; N.C. performed the gut microbiota analysis, wrote the original draft, and revised the manuscript; N.K. reviewed and scored liver pathology, and revised the manuscript; N.W. conducted the study, reviewed and scored liver pathology, and revised the manuscript; P.S. contributed to the conceptualization, supervised the study, and revised the manuscript; S.T. conducted the study, supplied probiotic emulsion for the study, and revised the manuscript; D.W. contributed to the conceptualization, acquired the funding, supervised the study, and revised the manuscript. All authors read and approved the final manuscript.
Funding
This study was funded by the Ratchadapiseksompotch Fund, Faculty of Medicine, Chulalongkorn University, grant number RA62/032 and the Thailand Science Research and Innovation Fund Chulalongkorn University, grant number CU_FRB65_hea (21)_028_30_09. Funders provided monetary support for the study but did not have the role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Declarations
Ethics approval and consent to participate
This study was conducted according to the Ethical Principles and Guidelines for the Use of Animals by the National Research Council of Thailand. The study protocol was approved by the Animal Care and Use Committee, the Faculty of Medicine, Chulalongkorn University (the permission No. is 035/2561).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Rehm J, Shield KD. Global burden of alcohol use disorders and alcohol liver disease. Biomedicines. 2019;7(4):99. doi: 10.3390/biomedicines7040099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Assanangkornchai S, Sam-Angsri N, Rerngpongpan S, Lertnakorn A. Patterns of alcohol consumption in the Thai population: results of the National Household Survey of 2007. Alcohol Alcohol. 2010;45(3):278–285. doi: 10.1093/alcalc/agq018. [DOI] [PubMed] [Google Scholar]
- 3.Mathurin P, Bataller R. Trends in the management and burden of alcoholic liver disease. J Hepatol. 2015;62(1 Suppl):S38–S46. doi: 10.1016/j.jhep.2015.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Drinane MC, Shah VH. Alcoholic hepatitis: diagnosis and prognosis. Clin Liver Dis. 2013;2(2):80–83. doi: 10.1002/cld.164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.EASL Clinical Practice Guidelines Management of alcohol-related liver disease. J Hepatol. 2018;69(1):154–181. doi: 10.1016/j.jhep.2018.03.018. [DOI] [PubMed] [Google Scholar]
- 6.Vassallo G, Mirijello A, Ferrulli A, et al. Review article: alcohol and gut microbiota - the possible role of gut microbiota modulation in the treatment of alcoholic liver disease. Aliment Pharmacol Ther. 2015;41(10):917–927. doi: 10.1111/apt.13164. [DOI] [PubMed] [Google Scholar]
- 7.Kim JH, Kim HJ, Son JH, et al. Effect of Lactobacillus fermentum MG590 on alcohol metabolism and liver function in rats. J Microbiol Biotechnol. 2003;13(6):919–925. [Google Scholar]
- 8.Ko JS, Yang HR, Chang JY, Seo JK. Lactobacillus plantarum inhibits epithelial barrier dysfunction and interleukin-8 secretion induced by tumor necrosis factor-alpha. World J Gastroenterol. 2007;13(13):1962–1965. doi: 10.3748/wjg.v13.i13.1962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Forsyth CB, Farhadi A, Jakate SM, Tang Y, Shaikh M, Keshavarzian A. Lactobacillus GG treatment ameliorates alcohol-induced intestinal oxidative stress, gut leakiness, and liver injury in a rat model of alcoholic steatohepatitis. Alcohol. 2009;43(2):163–172. doi: 10.1016/j.alcohol.2008.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Li F, Duan K, Wang C, McClain C, Feng W. Probiotics and alcoholic liver disease: treatment and potential mechanisms. Gastroenterol Res Pract. 2016;2016:5491465. doi: 10.1155/2016/5491465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sunanliganon C, Thong-Ngam D, Tumwasorn S, Klaikeaw N. Lactobacillus plantarum B7 inhibits Helicobacter pylori growth and attenuates gastric inflammation. World J Gastroenterol. 2012;18(20):2472–2480. doi: 10.3748/wjg.v18.i20.2472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Thiraworawong T, Spinler JK, Werawatganon D, et al. Anti-inflammatory properties of gastric-derived Lactobacillus plantarum XB7 in the context of Helicobacter pylori infection. Helicobacter. 2014;19(2):144–155. doi: 10.1111/hel.12105. [DOI] [PubMed] [Google Scholar]
- 13.Kanmani P, Kim H. Protective effects of lactic acid Bacteria against TLR4 induced inflammatory response in Hepatoma HepG2 cells through modulation of toll-like receptor negative regulators of mitogen-activated protein kinase and NF-kappaB signaling. Front Immunol. 2018;9:1537. doi: 10.3389/fimmu.2018.01537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Vissers YM, Snel J, Zuurendonk PF, Smit BA, Wichers HJ, Savelkoul HF. Differential effects of Lactobacillus acidophilus and Lactobacillus plantarum strains on cytokine induction in human peripheral blood mononuclear cells. FEMS Immunol Med Microbiol. 2010;59(1):60–70. doi: 10.1111/j.1574-695X.2010.00662.x. [DOI] [PubMed] [Google Scholar]
- 15.Boonma P, Spinler JK, Venable SF, Versalovic J, Tumwasorn S. Lactobacillus rhamnosus L34 and Lactobacillus casei L39 suppress Clostridium difficile-induced IL-8 production by colonic epithelial cells. BMC Microbiol. 2014;14:177. doi: 10.1186/1471-2180-14-177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Panpetch W, Chancharoenthana W, Bootdee K, et al. Lactobacillus rhamnosus L34 attenuates gut translocation-induced bacterial Sepsis in murine models of leaky gut. Infect Immun. 2017;86(1):e00700–e00717. doi: 10.1128/IAI.00700-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wang Y, Kirpich I, Liu Y, et al. Lactobacillus rhamnosus GG treatment potentiates intestinal hypoxia-inducible factor, promotes intestinal integrity and ameliorates alcohol-induced liver injury. Am J Pathol. 2011;179(6):2866–2875. doi: 10.1016/j.ajpath.2011.08.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Brunt EM, Janney CG, Di Bisceglie AM, Neuschwander-Tetri BA, Bacon BR. Nonalcoholic steatohepatitis: a proposal for grading and staging the histological lesions. Am J Gastroenterol. 1999;94(9):2467–2474. doi: 10.1111/j.1572-0241.1999.01377.x. [DOI] [PubMed] [Google Scholar]
- 19.Kalra J, Dragowska WH, Bally MB. Using pharmacokinetic profiles and digital quantification of stained tissue microarrays as a medium-throughput, quantitative method for measuring the kinetics of early signaling changes following integrin-linked kinase inhibition in an in vivo model of Cancer. J Histochem Cytochem. 2015;63(9):691–709. doi: 10.1369/0022155415587978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhu H, Jia Z, Misra H, Li YR. Oxidative stress and redox signaling mechanisms of alcoholic liver disease: updated experimental and clinical evidence. J Dig Dis. 2012;13(3):133–142. doi: 10.1111/j.1751-2980.2011.00569.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Venkatraman A, Shiva S, Wigley A, et al. The role of iNOS in alcohol-dependent hepatotoxicity and mitochondrial dysfunction in mice. Hepatology. 2004;40(3):565–573. doi: 10.1002/hep.20326. [DOI] [PubMed] [Google Scholar]
- 22.Kono H, Rusyn I, Yin M, et al. NADPH oxidase-derived free radicals are key oxidants in alcohol-induced liver disease. J Clin Invest. 2000;106(7):867–872. doi: 10.1172/JCI9020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Banan A, Fields JZ, Decker H, Zhang Y, Keshavarzian A. Nitric oxide and its metabolites mediate ethanol-induced microtubule disruption and intestinal barrier dysfunction. J Pharmacol Exp Ther. 2000;294(3):997–1008. [PubMed] [Google Scholar]
- 24.Banan A, Keshavarzian A, Zhang L, et al. NF-kappaB activation as a key mechanism in ethanol-induced disruption of the F-actin cytoskeleton and monolayer barrier integrity in intestinal epithelium. Alcohol. 2007;41(6):447–460. doi: 10.1016/j.alcohol.2007.07.003. [DOI] [PubMed] [Google Scholar]
- 25.Bull-Otterson L, Feng W, Kirpich I, et al. Metagenomic analyses of alcohol induced pathogenic alterations in the intestinal microbiome and the effect of Lactobacillus rhamnosus GG treatment. PLoS One. 2013;8(1):e53028. doi: 10.1371/journal.pone.0053028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Grander C, Adolph TE, Wieser V, et al. Recovery of ethanol-induced Akkermansia muciniphila depletion ameliorates alcoholic liver disease. Gut. 2018;67(5):891–901. doi: 10.1136/gutjnl-2016-313432. [DOI] [PubMed] [Google Scholar]
- 27.Hritz I, Mandrekar P, Velayudham A, et al. The critical role of toll-like receptor (TLR) 4 in alcoholic liver disease is independent of the common TLR adapter MyD88. Hepatology. 2008;48(4):1224–1231. doi: 10.1002/hep.22470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yan AW, Fouts DE, Brandl J, et al. Enteric dysbiosis associated with a mouse model of alcoholic liver disease. Hepatology. 2011;53(1):96–105. doi: 10.1002/hep.24018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lowe PP, Gyongyosi B, Satishchandran A, et al. Alcohol-related changes in the intestinal microbiome influence neutrophil infiltration, inflammation and steatosis in early alcoholic hepatitis in mice. PLoS One. 2017;12(3):e0174544. doi: 10.1371/journal.pone.0174544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Li J, Lin S, Vanhoutte PM, Woo CW, Xu A. Akkermansia Muciniphila protects against atherosclerosis by preventing metabolic Endotoxemia-induced inflammation in Apoe−/− mice. Circulation. 2016;133(24):2434–2446. doi: 10.1161/CIRCULATIONAHA.115.019645. [DOI] [PubMed] [Google Scholar]
- 31.Kim S, Lee Y, Kim Y, et al. Akkermansia muciniphila prevents fatty liver disease, decreases serum triglycerides, and maintains gut homeostasis. Appl Environ Microbiol. 2020;86(7):e03004-19. doi: 10.1128/AEM.03004-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sharma M, Arora I, Stoll ML, et al. Nutritional combinatorial impact on the gut microbiota and plasma short-chain fatty acids levels in the prevention of mammary cancer in Her2/neu estrogen receptor-negative transgenic mice. PLoS One. 2021;15(12):e0234893. doi: 10.1371/journal.pone.0234893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Khailova L, Dvorak K, Arganbright KM, et al. Bifidobacterium bifidum improves intestinal integrity in a rat model of necrotizing enterocolitis. Am J Physiol Gastrointest Liver Physiol. 2009;297(5):G940–G949. doi: 10.1152/ajpgi.00141.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Mendes MCS, Paulino DS, Brambilla SR, Camargo JA, Persinoti GF, Carvalheira JBC. Microbiota modification by probiotic supplementation reduces colitis associated colon cancer in mice. World J Gastroenterol. 2018;24(18):1995–2008. doi: 10.3748/wjg.v24.i18.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.LeBrun ES, Nighot M, Dharmaprakash V, Kumar A, Lo CC, Chain PSG, Ma TY. The Gut Microbiome and Alcoholic Liver Disease: Ethanol Consumption Drives Consistent and Reproducible Alteration in Gut Microbiota in Mice. Life (Basel). 2020;11(1):7. 10.3390/life11010007. [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.