

Abstract
Endometrial cancer (EC) is a common malignant tumor of the female reproductive system. Current treatments such as surgery and long-term hormone therapy are ineffective and have side effects. Danzhi Xiaoyao powder (DXP) can inhibit the growth of EC cells and induce apoptosis, but the pharmacological and molecular mechanisms of anticancer effects are still unclear. In this study, active components and potential targets of DXP were obtained from public databases. Protein effects and regulatory pathways of common targets were analyzed by protein-protein interaction (PPI), GO and KEGG. The results of network pharmacology showed that there are 87 common targets between EC and DXP. GO enrichment analysis showed that these targets were associated with response to oxidative stress, response to nutrient levels, hormone receptor binding and nuclear hormone receptor binding, etc. The results of KEGG analysis indicated that IL-17, TNF, PI3K/AKT, and RAS/RAF/MEK/ERK (ERK) signaling pathway were enriched in the anti-EC of DXP. Additionally, we cultured HEC-1B and KLE cells for validate experiments. DXP showed an inhibition of proliferation, migration, and cell cycle of both cells. Moreover, the expression of RAS, p-RAF, p-MEK, ERK, and p-ERK related proteins were downregulated. In conclusion, DXP might inhibit the proliferation of EC cells via apoptosis. Furthermore, DXP-induced inhibition of EC development might involve RAS/RAF/MEK/ERK pathway.
1. Introduction
Endometrial cancer (EC) is one of the malignant tumors of the female reproductive system, and its prevalence and mortality have rapidly risen worldwide [1, 2]. Globally, there were an estimated 417,000 incident cases and 97,000 deaths in 2020 [3]. The occurrence of EC is usually associated with overexposure to endogenous or exogenous estrogens [4]. Currently, the main treatment methods for EC are surgery and chemotherapy [5]. Chemotherapy drugs have certain toxicity, and due to the evolutionary nature of tumors, tumors cells may differentiate into subclonal cells with certain drug resistance, resulting in reduced antitumor efficacy of these chemotherapy drugs and possible adverse reactions [6, 7]. Therefore, it is important to find drugs with low side effects and better efficacy to improve the condition of EC.
With the development of cellular biotechnology, breakthroughs have been made in the research of anticancer herbal medicines. In the past 20 years, natural antitumor drugs have accounted for 66.29% of the anticancer drugs on the market [8]. Traditional Chinese medicine (TCM) has unique advantages in terms of efficacy and safety. Its reasonable application can help control the development of the disease, reduce the toxic effect of radiotherapy and avoid the recurrence of cancer metastasis to a large extent [9]. Danzhi Xiaoyao powder (DXP) is a formula of TCM including Cortex Moutan, Gardeniae Fructus, Radix Bupleuri, Angelicae Sinensis Radix, Paeoniae Radix Alba, Atractylodes macrocephala Koidz., Poria Cocos (Schw.) Wolf., and Licorice [10]. Studies have found that DXP has antidepressant [11] effects, and some studies have found that DXP has some efficacy against breast cancer and precancerous lesions [12, 13], especially ER-positive breast cancer [13], suggesting that DXP may also have some anticancer effects. Clinical studies have shown that DXP has good therapeutic effects on patients with endometriosis liver depression and blood heat type [14]. Therefore, DXP has the potential to be a drug for the treatment of EC. However, its active components, possible targets, and molecular mechanism have not been elucidated.
TCM plays a therapeutic role by restoring homeostasis in the biological process during disease development through multiple targets [15]. Network pharmacology is a new method to analyze the relationship between drugs, ingredients, targets, and diseases in the network based on biological networks and is gaining importance [16–18]. Therefore, the combining of TCM and network pharmacology is a new approach for the preferential selection of disease-related genes, revealing the association between diseases and drugs and further elucidating the regulatory role of TCM [19, 20].
In this study, network pharmacology combined with in vitro cellular experiment validation was performed to explore the active components, potential targets, and molecular mechanism of the anti-EC effect of DXP.
2. Materials and Methods
2.1. Collection of EC-Related Targets
We searched the Online Mendelian Inheritance in Man (OMIM, https://www.omim.org/) [21], DisGeNET (https://www.disgenet.org/) and GeneCards (https://www.genecards.org/) with the key word “endometrial cancer” to obtain EC-related targets. The duplicate disease targets were merged and deleted.
2.2. Collection of Active Components and Potential Targets of DXP
To collect all the relative components of DXP, we searched the traditional Chinese medicine systems pharmacology database and analysis platform (TCMSP, https://lsp.nwu.edu.cn/tcmsp.php) [22], Traditional Chinese Medicine Integrated Database (TCMID, https://tcm.cz3.nus.edu.sg/group/tcm-id/tcmid) [23], and Herbal Ingredients' Targets Database (HIT, https://lifecenter.sgst.cn/hit) [24], which includes a great deal of information about Chinese herbal medicine. All predicted targets of active components of DXP were searched from TCM-ID, TCMSP, HIT, and Search Tool for Interacting Chemicals (STITCH, https://stitch.embl.de) databases and selected according to the component-target correlation degree above 400 from STITCH database.
2.2.1. Drug-Likeness Screening
In the early stages of drug discovery, drug-likeness is a key consideration in the selection of compounds. A quantitative estimate of drug-likeness (QED), reported by Bickerton et al. in 2012, was used to evaluate a component's drug-likeness [25]. In this work, DXP components with QED ≥ 0.3 were included for further study.
2.2.2. Binomial Statistical Mode Analysis
According to previous studies, a binomial statistical mode was used for secondary screening for active components and potential targets of DXP [26, 27]. In this study, we set targets with P < 0.05 that could be used as main targets for the formula.
2.3. Common Target Screening and Protein-Protein Interaction (PPI) Construction
The gene targets of components of DXP and EC were analyzed by Venny (version 2.1) to obtain the common genes, which were the targets of action of DXP for the treatment of EC.
The PPI network was constructed by importing the common targets to the String database (https://string-db.org/cgi/input.pl), limiting the species to “Homo sapiens.” The results were imported into Cytoscape (version 3.8.0) for further analysis and optimization of the protein interaction network.
2.4. Construction of Herb-Components-Target (H-C-T) Network
An H-C-T network was constructed based on the active components of DXP and common targets. The above information was imported into Cytoscape for further analysis.
2.5. Enrichment Analysis
The clusterProfiler package of R (version 3.15.4) was applied for Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis. The GO analysis enriching for 3 aspects of potential targets is as follows: biological process (BP), cellular component (CC), and molecular function (MF). P < 0.01 was set as a screening threshold and the functional terms and pathways with this value were considered statistically significant and retained.
2.6. Cell Culture
The EC cell line HEC-1B and KLE cells were purchased from the Shanghai Institute for Biological Sciences, Chinese Academy of Sciences. HEC-1B cells were cultured in Dulbecco's Modified Eagle's Medium (DMEM) medium (Gibco, MD, USA) with 10% fetal bovine serum (Gibco); the KLE cells were cultured in DMEM/F12 medium (Gibco) with 10% fetal bovine serum (Gibco), both at 37°C with 95% air and 5% CO2.
2.7. Detection of Cell Viability
HEC-1B and KLE cells (2 × 103/well) were seeded into 96-well plates and cultured overnight at 37°C with 95% air and 5% CO2. Then, cells were treated with different concentrations of DXP (0, 0.05, 0.1, 0.2, 0.4, 0.8, and 1.6 mg/mL) for 24 h and 48 h. Next, cells were incubated with CCK-8 solution (Biosharp, China) for 2 h at 37°C and the absorbance at 450 nm was measured by an automatic microplate reader (BioTek, VT, USA).
2.8. Flow Cytometry
HEC-1B and KLE cells were cultured in 6-well plates at 3 × 105 cells/well and treated with 0.4 or 0.8 mg/mL DXP for 48 h, respectively.
For cell cycle analysis, cells were collected and washed with phosphate-buffered saline (PBS) followed by prechilled 70% ethanol at 4°C for more than 4 h. After fixation, cells were incubated with 100 μg/mL RNase (Nanjing KeyGen Biotech Co., Ltd., China) for 30 min, followed by 50 μg/mL propidium iodide (PI) (Becton-Dickinson, NJ, USA) for 30 min in the dark. Finally, a flow cytometer (Becton-Dickinson) was used to detect the cell cycle of HEC-1B and KLE cells.
For apoptosis analysis, cells were digested with trypsin without EDTA, washed twice with prechilled PBS, and then resuspended in the binding buffer. Next, cells were incubated with Annexin V-FITC (Becton-Dickinson) and PI for 15 min at room temperature under light-proof conditions. Finally, a flow cytometer (Becton-Dickinson) was used to detect the cell apoptosis of HEC-1B and KLE cells.
2.9. Cell Migration
The ability of migration in HEC-1B and KLE cells was detected by transwell assays. HEC-1B and KLE cells were resuspended in serum-free medium. 200 μL·cell suspensions were added to the upper layer of the transwell chamber (Corning, NY, USA). The lower chamber contained 500 μL of serum-containing medium. After 48 h incubation, the cells in the lower chamber were stained with 0.1% crystal violet for 20 min and counted under a 400 × microscope (Olympus, Japan) in 5 random fields.
2.10. Western Blot
The cells were collected and lysed with cell lysis buffer (Beyotime, China) for 30 minutes on ice, then centrifuged at 12000 rpm and 4°C for 10 minutes. The supernatants were collected and the protein concentrations were quantified by the BCA kit (Pierce, IL, USA). The protein samples were separated by the 10% SDS-PAGE gradient gel. The separated protein was transferred onto PVDF membranes. The membranes were blocked with 5% skim milk for 90 min, and incubated with primary antibodies (1 : 1000, Abcam, UK) overnight at 4°C. Following this, they were incubated with secondary antibody (1 : 500,Hangzhou Multi Sciences Co., Ltd., China, GAR007) for 1 h. Finally, the protein bands were detected by an enhanced chemiluminescent (ECL) system (Beyotime). The primary antibodies were listed as below: RAS (ab52939), p-RAF (ab200653), MEK1 (ab32091), p-MEK1 (ab96379), ERK (ab32537), p-ERK (ab131438), and GAPDH (ab9485).
2.11. Statistical Analysis
The results were presented as means ± SD and data comparison of multiple groups was adopted by one-way ANOVA followed by Tukey's test. All statistical analyses were obtained by GraghPad 7.0. The results were considered statistically significant if the P value was less than 0.05.
3. Results
3.1. EC-Related Targets
By retrieving three public databases, 836 EC-related targets were obtained, which included 237 in GeneCards, 197 in OMIM, and 402 in DisGeNET with the key word “endometrial cancer.” A total of 679 targets were screened by combining and deleting the repetitive values.
3.2. Active Components and Potential Targets of DXP
A total of 321 DXP-related components were searched from public databases including HIT, TCMSP, and TCMID databases. The potential targets of DXP were obtained from four public databases (HIT, TCMSP, TCMID, and STITCH), and there were 5,558 targets related to active components of DXP. Then, 274 active components were obtained according to drug-likeness screening (QED ≥ 0.3). Moreover, 589 main targets were obtained from binomial statistical mode analysis. Following it, we retrieved 214 active components of DXP. Among 589 screened components-related targets and 679 EC-related targets, 87 common targets were identified and considered as the key targets for further study (Figure 1). Accordingly, 168 active components were screened and identified as the main active components of DXP that act on EC (Table 1).
Figure 1.
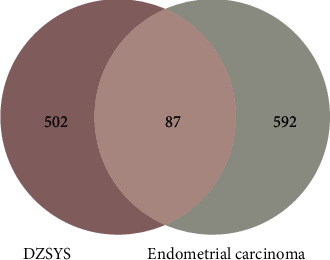
Venn diagram of common targets of danzhi xiaoyao powder (DXP) and endometrial cancer (EC).
Table 1.
Active components of DXP.
| Chemical_name | QED | Chemical_name | QED |
|---|---|---|---|
| Beta-elemene | 0.5799 | Phosphatidic acid | 0.4043 |
| Phenylalanine | 0.5664 | Glabrone | 0.8364 |
| Scopoletin | 0.5425 | Echinatin | 0.6610 |
| Guercetol | 0.5064 | Glabrol | 0.5836 |
| Quercetin | 0.5064 | Nonanoic acid | 0.5775 |
| Ursolic acid | 0.4433 | NON | 0.5672 |
| Calycosin | 0.8850 | (-)-Catechin | 0.5139 |
| Formononetin | 0.8529 | Gancaonin L | 0.4938 |
| Glabridin | 0.8323 | Hemo-sol | 0.4838 |
| Chrysin | 0.8206 | d-limonene | 0.4838 |
| Eugenol | 0.6955 | Alpha-limonene | 0.4838 |
| Thymol | 0.6510 | Stearic acid | 0.3017 |
| Kaempferol | 0.6372 | Catechin | 0.5139 |
| Farnesol | 0.6157 | WLN: QV4 | 0.5826 |
| isoliquiritigenin | 0.5824 | hexanoic acid | 0.5687 |
| Scopoletol | 0.5425 | Pentadecylic acid | 0.4059 |
| Capsaicin | 0.5370 | Myristicin | 0.6887 |
| IPH | 0.5172 | Scatole | 0.4871 |
| Puerarin | 0.4049 | Coumarin | 0.4124 |
| Glucuronic acid | 0.3250 | PAC | 0.6684 |
| Z-ligustilide | 0.6667 | Caprylic acid | 0.5818 |
| Linalool | 0.6172 | Octanoic acid | 0.5818 |
| Salicylic acid | 0.6129 | Oleanolic acid | 0.4460 |
| BUA | 0.5445 | Anethole | 0.6262 |
| L-Carvone | 0.5247 | Vanillin | 0.5173 |
| BuOH | 0.5171 | Gallocatechin | 0.4426 |
| Adenine | 0.5125 | WLN: VHR | 0.4967 |
| Crocetin | 0.5030 | Isorhamnetin | 0.6678 |
| Ethanol | 0.4049 | Geraniol | 0.6172 |
| IFP | 0.3920 | Mairin | 0.4635 |
| glutamate | 0.3835 | GLB | 0.3046 |
| Palmitic acid | 0.3653 | Histidine | 0.4184 |
| Esculetin | 0.3579 | EUG | 0.6993 |
| LPG | 0.3562 | HEX | 0.4628 |
| Alanine | 0.3562 | Vitexin | 0.3497 |
| Deoxygomisin a | 0.7145 | Methyleugenol | 0.6599 |
| Butylated hydroxytoluene | 0.7531 | EB | 0.5161 |
| Apocynin | 0.6736 | 2-Heptanone | 0.5103 |
| Paeonol | 0.5478 | Alpha-glycyrrhetinic acid | 0.5188 |
| Alpha-humulene | 0.4851 | Naphthalene | 0.5114 |
| Stigmasterol | 0.4599 | OYA | 0.3958 |
| Hyacinthin | 0.4290 | DFV | 0.8226 |
| Acetic acid | 0.4199 | Liquiritigenin | 0.8226 |
| Baicalin | 0.3617 | WLN: QVR BVQ | 0.6891 |
| Isovitexin | 0.3497 | 1,2-Benzenedicarboxylic acid | 0.6891 |
| Choline | 0.3405 | Licocoumarone | 0.5799 |
| Tetradecane | 0.3217 | Glabranin | 0.8174 |
| Caffeic acid | 0.4750 | PHPH | 0.5905 |
| Pinocembrin | 0.8226 | Genkwanin | 0.8862 |
| PHB | 0.6129 | 5,4′-Dihydroxy-7-methoxyflavone | 0.8862 |
| 5-Indolol | 0.5651 | PLO | 0.7502 |
| 3,4,5-Trihydroxybenzoic acid | 0.4656 | O-xylene | 0.4767 |
| Gallic acid | 0.4656 | Angelicin | 0.4670 |
| Umbelliferone | 0.3628 | Dodekan | 0.3809 |
| Morusin | 0.6040 | Dodecane | 0.3809 |
| DTY | 0.5110 | Epigallocatechin | 0.4426 |
| Genipin | 0.5093 | Beta-glycyrrhetinic acid | 0.5188 |
| Licochalcone A | 0.4563 | Liquiritin | 0.4838 |
| Beta-sitosterol | 0.4354 | Guasol | 0.5771 |
| Angelicin | 0.4354 | Guaiacol | 0.5771 |
| Zoomaric acid | 0.3256 | BU3 | 0.4701 |
| Uralenic acid | 0.6249 | PYG | 0.4560 |
| 18Beta-glycyrrhetinic acid | 0.5188 | Isoeugenol | 0.7273 |
| Glycyrrhetinic acid | 0.5188 | Ethyl protocatechuate | 0.5494 |
| 18Alpha-glycyrrhetic acid | 0.5188 | Safrol | 0.6221 |
| DBP | 0.4752 | Malic acid | 0.4307 |
| Myristic acid | 0.4490 | Fraxetin | 0.4141 |
| Fructose | 0.3101 | PCR | 0.5390 |
| DEP | 0.6925 | P-cresol | 0.5390 |
| Py | 0.4453 | O-thymol | 0.6510 |
| FER | 0.7180 | Carvacrol | 0.6510 |
| Isoimperatorin | 0.4856 | Licochalcone B | 0.4567 |
| Lauric acid | 0.3925 | Methylglyoxal | 0.3144 |
| Alpha-linolenic acid | 0.3326 | ICO | 0.6690 |
| Succinic acid | 0.5303 | Hexenal | 0.3885 |
| PHA | 0.5664 | Adonitol | 0.3082 |
| L-Ile | 0.4718 | Butal | 0.4435 |
| h-Met-h | 0.4506 | WLN: VH6 | 0.3957 |
| L-valine | 0.4266 | Nonanal | 0.3938 |
| Istidina | 0.4207 | Decanal | 0.3868 |
| L-valin | 0.4120 | PTL | 0.3745 |
| Prolinum | 0.3867 | Trans-2-nonenal | 0.3144 |
| Glutamine | 0.3835 | 2,4-Heptdienal | 0.3076 |
| Protocatechuic acid | 0.5265 | Prunetin | 0.8850 |
3.3. Construction of PPI Network
The 87 common targets of DXP and EC were imported to the String database for analysis, and Cytoscape (version 3.8.0) was used for PPI construction (Figure 2). It contained 87 nodes and 1,587 edges, and the average degree value was 36.5.
Figure 2.
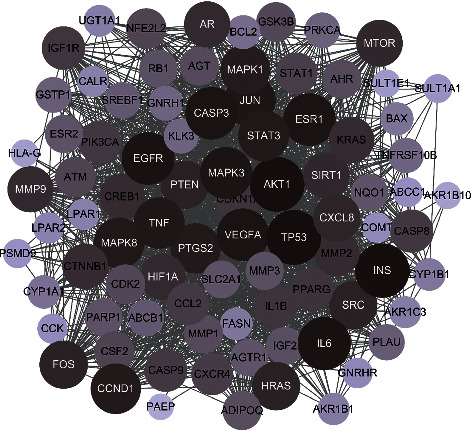
Common target protein-protein interaction (PPI) network between DXP and EC. Each node represents a target. The lines between the nodes represent the association among the different targets. The size and color of the node and its color shade change represent the size of the degree value.
3.4. Construction of Herb-Components-Target (H-C-T) Network
The H-C-T network was obtained through the import of 168 active components and 87 potential common targets of DXP into Cytoscape (version 3.8.0) and is shown in Figure 3. There were 262 nodes and 1159 edges. The node size was proportional to its degree value. The red rhombus stood for different herbs which were contained in DXP. The cyan hexagon represented key targets of DXP acting on EC. Other different colored rhombuses represented the compounds contained in different herbs. Among them, the orange rhombus stood for the components that were contained in many kinds of herbs. The result showed that the top 5 active components nodes with the largest size of degree were quercetin, ethanol, kaempferol, IFP, and palmitic acid, with degree values of 64, 52, 43, 42, and 41, respectively. CASP3, AKT1, CYP1A1, JUN, and PPARG exhibited the highest degree values of 48, 25, 24, 22, and 22, respectively. These active compounds and targets may play a vital role in the effect of DXP against EC.
Figure 3.
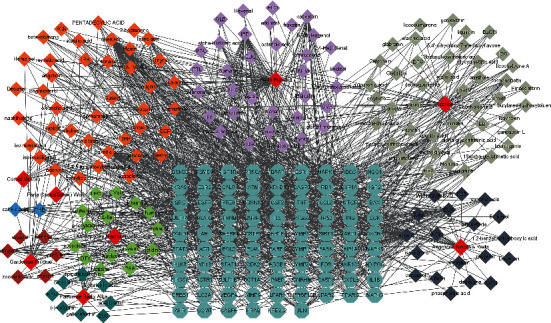
Herb-components-target (H-C-T) network. The red rhombus represents the different herbs contained in DXP. The cyan hexagon represents the key targets of DXP acting on EC. Other different colors rhombuses represent the compounds contained in different herbs. Among them, the orange rhombus represents the components that are contained in many kinds of herbs.
3.5. Enrichment Analysis
In order to illuminate the biological functions and signaling pathways during the process of anti-EC of DXP, we employed the clusterProfiler package of the R language to perform GO and KEGG enrichment analysis on the 87 targets. A total of significant GO 1123 terms were obtained, including 1066 terms of BP, 18 terms of CC, and 39 terms of MF (P < 0.01). The top 15 enriched terms of BP, CC, and MF were selected and visualized. The results showed that the targets were related to response to oxidative stress, response to nutrient levels, response to steroid hormone, and other biological processes; enriched in the transcription factor complex, membrane raft, membrane microdomain, and other cellular components; and related to RNA polymerase II transcription factor binding, hormone receptor binding, ubiquitin-like protein ligase binding, and other molecular functions (Figures 4(a)–4(c)).
Figure 4.
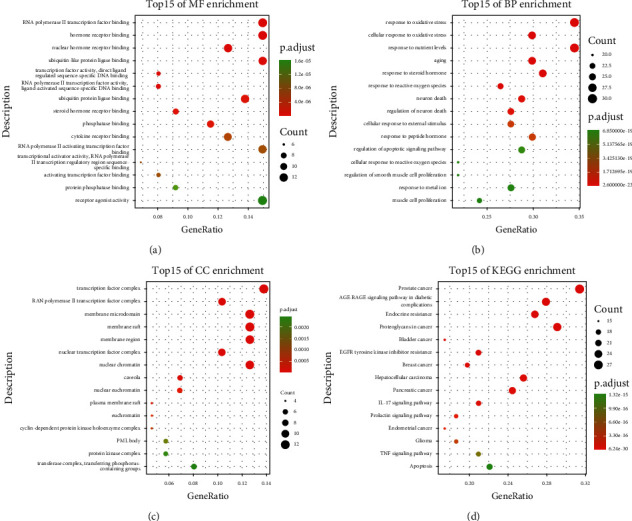
Enrichment analysis of GO and KEGG pathways of key target genes. The size of the circle represents the number of genes, and the color of the circle represents the corrected P value.
In KEGG enrichment analysis, a total of 143 pathways of anti-EC targets of DXP were identified according to the P value less than 0.01. The top 15 significantly enriched pathways with the highest gene counts were shown in Figure 4(d), and the specific information was shown in Table 2. The results showed that the targets may be associated with the AGE-RAGE signaling pathway in diabetic complications, the IL-17 signaling pathway, the TNF signaling pathway, and other signaling pathways. Among them, the endometrial cancer, apoptosis, EGFR tyrosine kinase inhibitor resistance, and proteoglycans in the cancer signaling pathway map showed that the PI3K-AKT and RAS-RAF-MEK-ERK (ERK) signaling pathways may play a crucial role in the treatment of EC with DXP (Figure 5).
Table 2.
Enrichment signaling pathways of KEGG.
| ID | Description | P.adjust | Count |
|---|---|---|---|
| hsa05215 | Prostate cancer | 6.24E − 30 | 27 |
| hsa04933 | AGE-RAGE signaling pathway in diabetic complications | 3.62E − 25 | 24 |
| hsa01522 | Endocrine resistance | 5.68E − 24 | 23 |
| hsa05205 | Proteoglycans in cancer | 7.11E − 19 | 25 |
| hsa05219 | Bladder cancer | 9.54E − 19 | 15 |
| hsa01521 | EGFR tyrosine kinase inhibitor resistance | 1.86E − 18 | 18 |
| hsa05212 | Pancreatic cancer | 2.61E − 17 | 17 |
| hsa05225 | Hepatocellular carcinoma | 2.61E − 17 | 22 |
| hsa05224 | Breast cancer | 2.61E − 17 | 21 |
| hsa04657 | IL-17 signaling pathway | 3.59E − 17 | 18 |
| hsa04917 | Prolactin signaling pathway | 1.39E − 16 | 16 |
| hsa05213 | Endometrial cancer | 1.94E − 16 | 15 |
| hsa05214 | Glioma | 4.10E − 16 | 16 |
| hsa04668 | TNF signaling pathway | 7.33E − 16 | 24 |
| hsa04210 | Apoptosis | 1.32E − 15 | 18 |
Figure 5.
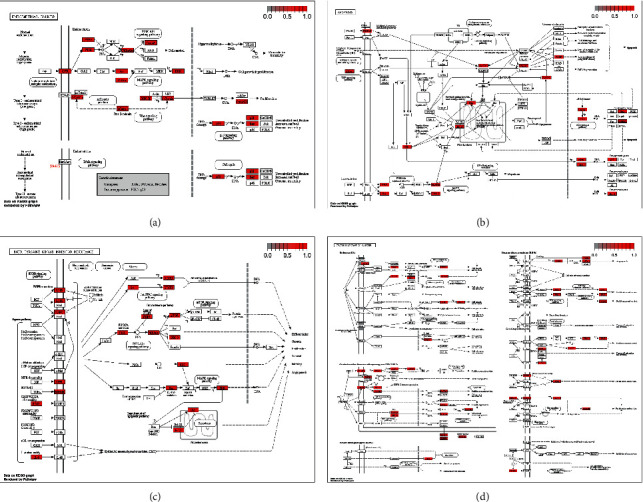
The map of endometrial cancer (a), apoptosis (b), EGFR tyrosine kinase inhibitor resistance (c), and proteoglycans in cancer (d) signaling pathway. The red boxes represent key targets.
The network of target-BP-pathways was obtained by Cytoscape (version 3.8.0) and shown in Figure 6, which contained 105 nodes and 713 edges and included 75 common targets, the top 15 BP terms, and the top 15 signaling pathways. The square stood for disease targets, the triangle stood for signaling pathway, and the arrow represented biological processes.
Figure 6.
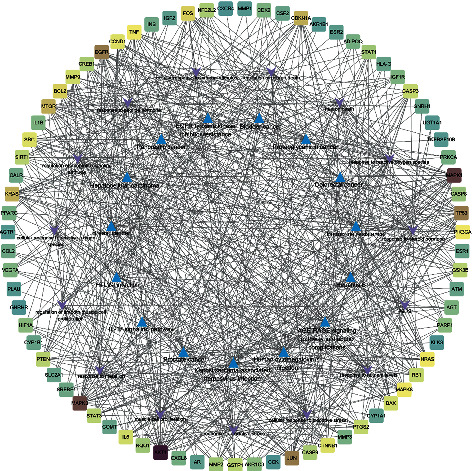
Core target-biological process-signal pathway network. The square represents DXP anti-EC targets, the triangle represents the top 15 signaling pathway, and the arrow represents the top 15 biological processes.
3.6. Correlation Analysis of Targets with BP and Signaling Pathways
To explore the correlation of the hub targets with the signaling pathway and BP more clearly, a hub target network was constituted with the top 10 targets, including TP53 (n = 71), AKT1 (n = 71), INS (n = 70), IL6 (n = 67), EGFR (n = 66), CASP3 (n = 65), ESR1 (n = 65), VEGFA (n = 65), MAPK3 (n = 64), and JUN (n = 63) (Figure 7(a)). The correlations were analyzed by combining them with the top 15 pathways and BP terms. The results showed that AKT1 had a higher correlation with the BP, and its response to oxidative stress was more relevant to the targets (Figure 7(b)). In the analysis of correlation with signaling pathways, AKT1 and MAPK3 were more important to the pathways (Figure 7(c)).
Figure 7.
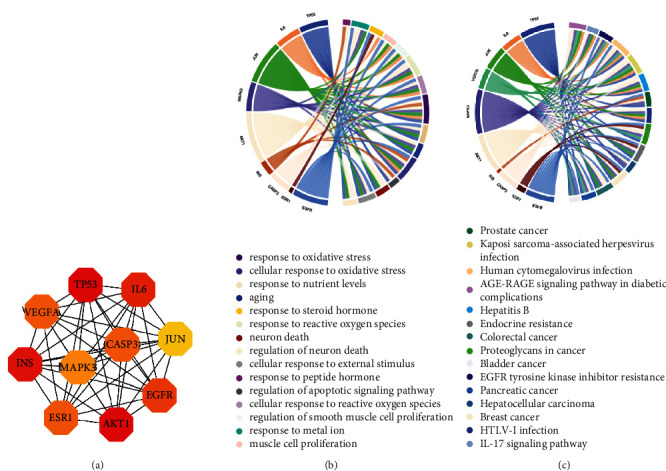
Hub network of top 10 high-degree targets (a), the correlation between the core network and the first 15 BP terms (b), and the correlation between the core network and the first pathway (c).
3.7. DXP Suppresses the Viability, Cell Cycle, and Metastasis While Triggers the Apoptosis of EC Cells
Different concentrations of DXP were used to treat HEC-1B and KLE cells and assay their survival rates. As shown in Figure 8(a), the cell viabilities of both types were decreased with the increasing concentration of the DXP at 24 h and 48 h. The LC50 values were calculated to be 0.717 mg/mL for HEC-1B and 0.603 mg/mL for KLE when DXP treated for 48 h. So, DXP (0.4 mg/mL and 0.8 mg/mL) treated HEC-1B and KLE cells for 48 h were selected in the subsequent experimental.
Figure 8.
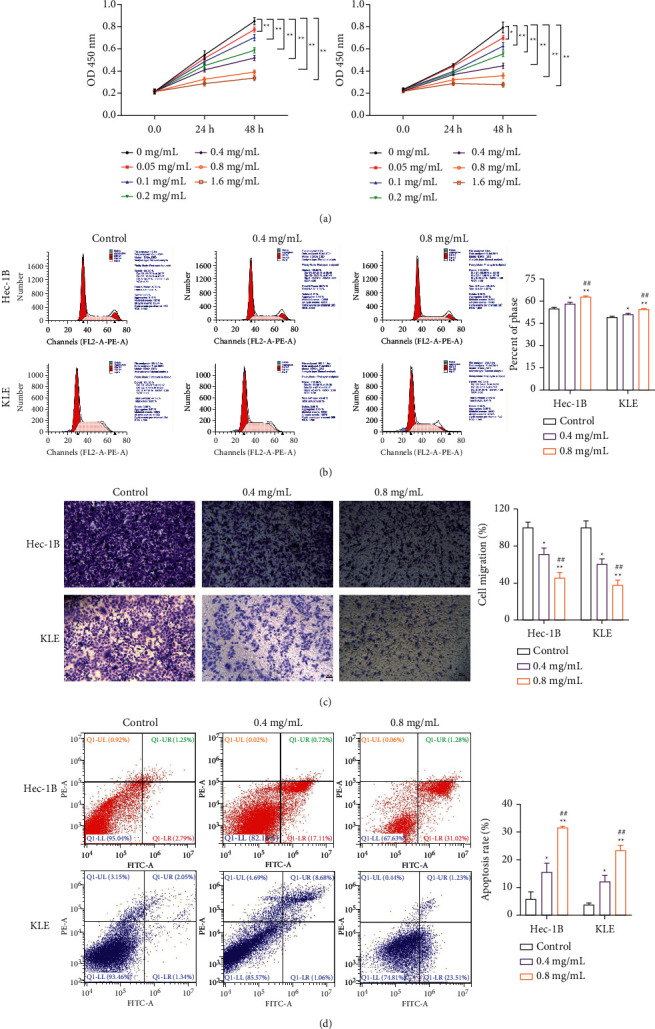
HEC-1B and KLE cells were treated with different concentrations of DXP for cell proliferation (a), cycle (b), migration (c), and apoptosis (d) assays. ∗P < 0.05, ∗∗P < 0.01 vs. Control group; ##P < 0.01 vs. 0.4 mg/mL group.
Cell cycles results showed DXP significantly increased the proportion of G0/G1 phase cells (P < 0.05), and DXP with a high-dose have a better inhibitory effect than low-dose (P < 0.01) (Figure 8(b)). Meanwhile, DXP also inhibited the migration of EC cells. The apoptosis rate in high-dose was significantly higher than low-dose (P < 0.01) (Figure 8(c)). Additionally, the apoptosis rate of EC cells was promoted by DXP addition (P < 0.05) (Figure 8(d)). These results indicated that DXP suppressed the malignant progress of EC cells in vitro.
3.8. DXP Inhibited the Protein Expression of ERK Signaling Pathway
Network pharmacology analysis indicated that the RAS pathway is the most important pathway in the treatment of DXP on EC. Thus, the expression levels of RAS, p-RAF, MEK, p-MEK, ERK, and p-ERK were examined by Western blot. As shown in Figure 9, DXP decreased the expression of RAS, p-RAF, p-MEK, and p-ERK proteins, but had no effect on MEK and ERK protein expression. It suggested that DXP against EC through ERK signaling pathway.
Figure 9.
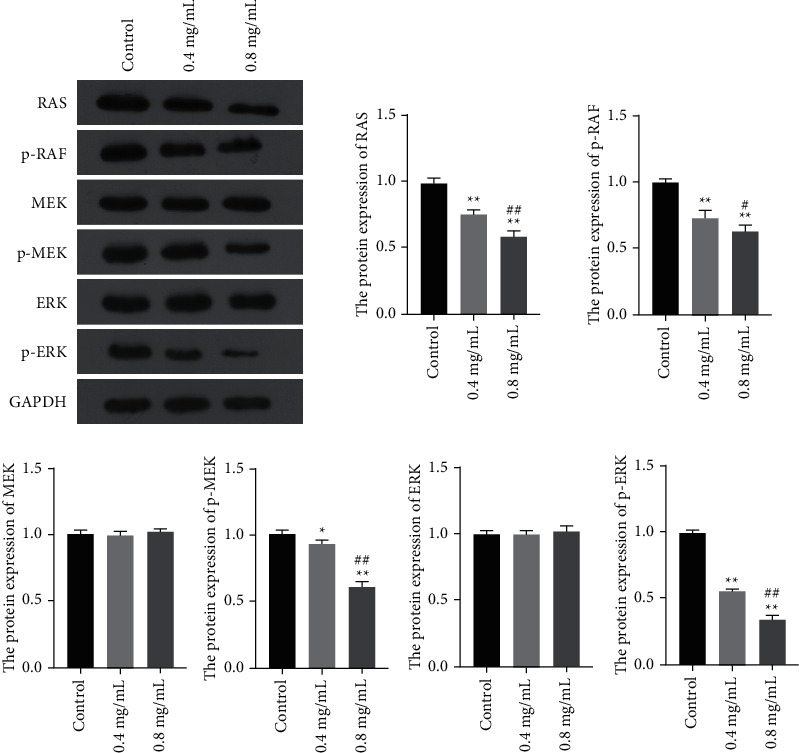
The expression of RAS/RAF/MEK/ERK (ERK) pathway related proteins was detected using Western blot. ∗P < 0.05, ∗∗P < 0.01 vs. Control group; #P < 0.05, ##P < 0.01 vs. 0.4 mg/mL group.
4. Discussion
As an epithelial malignant tumor originating from the endometrium, EC ranks first in the incidence of malignant tumors of the female reproductive system, with a trend of younger incidence [28–30]. In recent years, the extraction of effective antitumor components with low toxicity from TCM has become a new research orientation [31, 32]. DXP is attracting attention for its anticancer and immune-enhancing activities [12]. In this work, we conducted network pharmacology and cellular experiments to identify the active components, core targets, and potential mechanisms of DXP in EC.
In the present study, a total of 168 active components of DXP and 87 DXP-EC targets were screened from public databases. In the H-C-T network, we found that quercetin, associated with the largest number of targets, is the most important component in DXP for EC. Quercetin is a polyhydroxyflavonoid with various biological functions and has antioxidant, anti-inflammatory, and antiviral properties [33]. A New Jersey survey reported that lean women who consume quercetin have a lower risk of developing EC [34]. It has also been found that quercetin inhibits the proliferation of Ishikawa cells (an EC cell line) by downregulating EGF and cyclin D1 [35]. In addition, kaempferol also played a crucial role in the suppression of EC. Kaempferol is a dietary bioflavonoid with anticancer, anti-inflammatory, and antioxidant properties that inhibits human cancer cell proliferation through various mechanisms, including induction of the tumor suppressor p53 and inhibition of Erα [36]. Additionally, Lei's study has substantiated that kaempferol against EC by inducing cell apoptosis, arresting the cell cycle in G2/M phase, suppressing cell invasion and upregulating the m-TOR/PI3K signaling pathway [37]. These reports combined with the present research provide evidence for the therapeutic potential of quercetin and kaempferol in DXP to cure EC.
PPI analysis showed that the hub targets of DXP for EC were TP53, AKT1, INS, IL6, EGFR, CASP3, ESR1, VEGFA, MAPK3, and JUN. To annotate the functions and related pathway of 87 targets, we conducted the GO and KEGG signal pathway analysis. GO analysis results suggested that targets were mainly related to responses to oxidative stress, nutrient levels, steroid hormones, and other biological processes. The KEGG analysis results suggested that the pathways involved in DXP treating EC were mainly related to the IL-17 signaling pathway, TNF signaling pathway, PI3K-AKT signaling pathway, and ERK signaling pathway. Among them, IL-17 is a proinflammatory cytokine and is significantly expressed in a variety of cancers [38–40]. Lai's study demonstrates that IL-17 may regulate chemokines and cytokines in gynecologic cancers through activation of IκBα and phosphorylation of extracellular signal-regulated kinase 1/2 [41]. PI3K-AKT signaling pathway is a key intracellular signaling pathway that regulates cell proliferation and cell cycle, and is closely related to tumorigenesis [42, 43]. Additionally, numerous studies have shown that the PI3K-AKT signaling pathway is involved in the malignant process of EC, such as picropodophyllin inhibiting EC cell proliferation by disrupting the PI3K-AKT signaling pathway [44]; NLRC5 promoting EC progression through activation of the PI3K/AKT pathway [45]. The ERK signaling pathway is part of the mitogen-activated protein kinase (MAPK) signaling pathway, which is involved in various cellular activities, such as cell proliferation, differentiation, apoptosis, inflammation, and other cellular activities, and is closely related to tumor-related immune escape mechanisms [46, 47]. Studies have reported that the ERK signaling pathway is one of the important pathways involved in endometrial carcinogenesis and is involved in tumor cell metabolism, growth, proliferation, survival, and angiogenesis [48].
In vitro experiments were performed to further verify the molecular mechanism of DXP against EC. The cell experiments demonstrated that DXP for the treatment of EC suppressed cell proliferation, arrested the cell cycle in G0/G1 phase, induced cell apoptosis, weakened the ability of cell migration, downregulated RAS expression and RAF, MEK, and ERK phosphorylation levels. These findings verified the predicted result of the network pharmacology approach.
5. Conclusion
In summary, we combined network pharmacology and in vitro experiments to verify the potential mechanism of the bioactive components and targets of DXP in treating EC. We demonstrated that DXP could inhibit the proliferation and migration of EC cells and enhance EC cell apoptosis. Moreover, DXP protects against EC malignant progression via downregulating the ERK signaling pathway. This study provides an effective drug for the treatment of EC. Meanwhile, network pharmacology analysis reveals the molecular mechanisms involved in the treatment of EC with DXP, providing a research basis for subsequent studies.
Data Availability
The data used to support the findings of this study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare that there are no conflicts of interest regarding the publication of this paper.
References
- 1.Siegel R. L., Miller K. D., Jemal A. Cancer statistics, 2018. CA: A Cancer Journal for Clinicians . 2018;68(1):7–30. doi: 10.3322/caac.21442. [DOI] [PubMed] [Google Scholar]
- 2.Brooks R. A., Fleming G. F., Lastra R. R., et al. Current recommendations and recent progress in endometrial cancer. CA: A Cancer Journal for Clinicians . 2019;69(4):258–279. doi: 10.3322/caac.21561. [DOI] [PubMed] [Google Scholar]
- 3.Sung H., Ferlay J., Siegel R. L., et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: A Cancer Journal for Clinicians . 2021;71(3):209–249. doi: 10.3322/caac.21660. [DOI] [PubMed] [Google Scholar]
- 4.Zannoni G. F., Monterossi G., De Stefano I., et al. The expression ratios of estrogen receptor α (ERα) to estrogen receptor β1 (ERβ1) and ERα to ERβ2 identify poor clinical outcome in endometrioid endometrial cancer. Human Pathology . 2013;44(6):1047–1054. doi: 10.1016/j.humpath.2012.09.007. [DOI] [PubMed] [Google Scholar]
- 5.Colombo N., Preti E., Landoni F., et al. Endometrial cancer: ESMO clinical practice guidelines for diagnosis, treatment and follow-up. Annals of Oncology . 2013;24 doi: 10.1093/annonc/mdt353. [DOI] [PubMed] [Google Scholar]
- 6.Zagouri F., Bozas G., Kafantari E., et al. Endometrial cancer: what is new in adjuvant and molecularly targeted therapy? Obstetrics and gynecology international . 2010;2010:11. doi: 10.1155/2010/749579.749579 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Morice P., Leary A., Creutzberg C., Abu-Rustum N., Darai E. Endometrial cancer. Lancet . 2016;387(10023):1094–1108. doi: 10.1016/s0140-6736(15)00130-0. [DOI] [PubMed] [Google Scholar]
- 8.Imran M., Rauf A., Shah Z. A., et al. Chemo-preventive and therapeutic effect of the dietary flavonoid kaempferol: a comprehensive review. Phytotherapy Research . 2019;33(2):263–275. doi: 10.1002/ptr.6227. [DOI] [PubMed] [Google Scholar]
- 9.Ling C. Q., Fan J., Lin H. S., et al. Clinical practice guidelines for the treatment of primary liver cancer with integrative traditional Chinese and western medicine. Journal of integrative medicine . 2018;16(4):236–248. doi: 10.1016/j.joim.2018.05.002. [DOI] [PubMed] [Google Scholar]
- 10.Yang K., Zeng L., Ge J. Exploring the pharmacological mechanism of danzhi xiaoyao powder on ER-positive breast cancer by a network pharmacology approach. Evidence-based Complementary and Alternative Medicine . 2018;2018:20. doi: 10.1155/2018/5059743.5059743 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhu Y. L., Li S. L., Zhu C. Y., Wang W., Zuo W. F., Qiu X. J. Metabolomics analysis of the antidepressant prescription danzhi xiaoyao powder in a rat model of chronic unpredictable mild stress (CUMS) Journal of Ethnopharmacology . 2020;260 doi: 10.1016/j.jep.2020.112832.112832 [DOI] [PubMed] [Google Scholar]
- 12.Li R., Liu L. P., Wang Z., et al. Effect of danzhi xiaoyao san on human breasr cancer cell line MCF-7 in nude mice. Chinese Journal of Experimental Traditional Medical Formulae . 2016;22(2):78–81. [Google Scholar]
- 13.Liao H., Banbury L. K., Leach D. N. Effects and potential mechanisms of danzhi xiaoyao pill on proliferation of MCF-7 human breast cancer cells in vitro. Chinese Journal of Integrative Medicine . 2008;14(2):128–131. doi: 10.1007/s11655-008-0128-y. [DOI] [PubMed] [Google Scholar]
- 14.Tian Y., Schwieters C. D., Opella S. J., Marassi F. M. High quality NMR structures: a new force field with implicit water and membrane solvation for xplor-NIH.R, PR and C-erbB-2 in patients with endometriosis of liver depression and blood heat type. Journal of Biomolecular NMR . 2017;67(1):35–49. doi: 10.1007/s10858-016-0082-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wu L., Wang Y., Nie J., Fan X., Cheng Y. A network pharmacology approach to evaluating the efficacy of Chinese medicine using genome-wide transcriptional expression data. Evidence-based Complementary and Alternative Medicine . 2013;2013:8. doi: 10.1155/2013/915343.915343 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Yuan H., Ma Q., Cui H., et al. How can synergism of traditional medicines benefit from network pharmacology? Molecules . 2017;22(7):p. 1135. doi: 10.3390/molecules22071135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhang Y. Q., Li S. Progress in network pharmacology for modern research of traditional Chinese medicine. Chinese Journal of PharmacologyToxicology . 2015;29(6):883–892. [Google Scholar]
- 18.Liu Z. H., Sun X. B. Network pharmacology: new opportunity for the modernization of traditional Chinese medicine. Yao xue bao = Acta pharmaceutica Sinica . 2012;47(6):696–703. [PubMed] [Google Scholar]
- 19.Yang W., Kang Y., Zhao Q., et al. Herbal formula yangyinjiedu induces lung cancer cell apoptosis via activation of early growth response 1. Journal of Cellular and Molecular Medicine . 2019;23(9):6193–6202. doi: 10.1111/jcmm.14501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Huang J., Guo W., Cheung F., Tan H. Y., Wang N., Feng Y. Integrating network pharmacology and experimental models to investigate the efficacy of coptidis and scutellaria containing huanglian jiedu decoction on hepatocellular carcinoma. American Journal of Chinese Medicine . 2020;48(1):161–182. doi: 10.1142/s0192415x20500093. [DOI] [PubMed] [Google Scholar]
- 21.Amberger J. S., Hamosh A. Searching online mendelian inheritance in man (OMIM): a knowledgebase of human genes and genetic phenotypes. Current Protocols in Bioinformatics . 2017;58 doi: 10.1002/cpbi.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ru J., Li P., Wang J., et al. TCMSP: a database of systems pharmacology for drug discovery from herbal medicines. Journal of Cheminformatics . 2014;6(1):p. 13. doi: 10.1186/1758-2946-6-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Huang L., Xie D., Yu Y., et al. TCMID 2.0: a comprehensive resource for TCM. Nucleic Acids Research . 2018;46:D1117–D1120. doi: 10.1093/nar/gkx1028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ye H., Ye L., Kang H., et al. HIT: linking herbal active ingredients to targets. Nucleic Acids Research . 2011;39:D1055–D1059. doi: 10.1093/nar/gkq1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bickerton G. R., Paolini G. V., Besnard J., Muresan S., Hopkins A. L. Quantifying the chemical beauty of drugs. Nature Chemistry . 2012;4(2):90–98. doi: 10.1038/nchem.1243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Liang X., Li H., Li S. A novel network pharmacology approach to analyse traditional herbal formulae: the Liu-Wei-Di-Huang pill as a case study. Molecular BioSystems . 2014;10(5):1014–1022. doi: 10.1039/c3mb70507b. [DOI] [PubMed] [Google Scholar]
- 27.Yang M., Chen J., Xu L., et al. A network pharmacology approach to uncover the molecular mechanisms of herbal formula ban-xia-xie-xin-tang. Evidence-based Complementary and Alternative Medicine . 2018;2018:22. doi: 10.1155/2018/4050714.4050714 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zhu Y., Shi L., Chen P., Zhang Y., Zhu T. Identification of six candidate genes for endometrial carcinoma by bioinformatics analysis. World Journal of Surgical Oncology . 2020;18(1):p. 161. doi: 10.1186/s12957-020-01920-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.DeSantis C. E., Lin C. C., Mariotto A. B., et al. Cancer treatment and survivorship statistics, 2014. CA: A Cancer Journal for Clinicians . 2014;64(4):252–271. doi: 10.3322/caac.21235. [DOI] [PubMed] [Google Scholar]
- 30.Terzic M., Norton M., Terzic S., Bapayeva G., Aimagambetova G. Fertility preservation in endometrial cancer patients: options, challenges and perspectives. Ecancermedicalscience . 2020;14:p. 1030. doi: 10.3332/ecancer.2020.1030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wang Y., Fu J. Y., Zhang S. Y., et al. Investigation of correlation between traditional Chinese medicine synbrome elements and clinical characteristics of patients with multidrug-resistant pulmonary tuberculosis. Chinese Journal of Antituberculosis . 2020;42(2):115–120. [Google Scholar]
- 32.Lu C. L., Li X., Zhou H. M., et al. Traditional Chinese medicine in cancer care: an overview of 5834 randomized controlled trials published in Chinese. Integrative Cancer Therapies . 2021;20 doi: 10.1177/15347354211031650.153473542110316 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Davoodvandi A., Shabani Varkani M., Clark C. C. T., Jafarnejad S. Quercetin as an anticancer agent: focus on esophageal cancer. Journal of Food Biochemistry . 2020;44(9) doi: 10.1111/jfbc.13374.e13374 [DOI] [PubMed] [Google Scholar]
- 34.Bandera E. V., Williams M. G., Sima C., et al. Phytoestrogen consumption and endometrial cancer risk: a population-based case-control study in New Jersey. Cancer Causes & Control . 2009;20(7):1117–1127. doi: 10.1007/s10552-009-9336-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kaneuchi M., Sasaki M., Tanaka Y., Sakuragi N., Fujimoto S., Dahiya R. Quercetin regulates growth of Ishikawa cells through the suppression of EGF and cyclin D1. International Journal of Oncology . 2003;22(1):159–164. doi: 10.3892/ijo.22.1.159. [DOI] [PubMed] [Google Scholar]
- 36.Luo H., Rankin G. O., Li Z., Depriest L., Chen Y. C. Kaempferol induces apoptosis in ovarian cancer cells through activating p53 in the intrinsic pathway. Food Chemistry . 2011;128(2):513–519. doi: 10.1016/j.foodchem.2011.03.073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lei X., Guo J., Wang Y., et al. Inhibition of endometrial carcinoma by kaempferol is interceded through apoptosis induction, G2/M phase cell cycle arrest, suppression of cell invasion and upregulation of m-TOR/PI3K signalling pathway. Journal of BUON: Official Journal of the Balkan Union of Oncology . 2019;24(4):1555–1561. [PubMed] [Google Scholar]
- 38.Jiang G., Liu C. T., Zhang W. D. IL-17A and GDF15 are able to induce epithelial-mesenchymal transition of lung epithelial cells in response to cigarette smoke. Experimental and Therapeutic Medicine . 2018;16(1):12–20. doi: 10.3892/etm.2018.6145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Molina M. F., Abdelnabi M. N., Fabre T., Shoukry N. H. Type 3 cytokines in liver fibrosis and liver cancer. Cytokine . 2019;124 doi: 10.1016/j.cyto.2018.07.028.154497 [DOI] [PubMed] [Google Scholar]
- 40.Chen R., Chen Q. T., Dong Y. H. Clinical efficacy of apatinib in treating metastatic gastric cancer and its effect on IL-17. Oncology Letters . 2019;17(6):5447–5452. doi: 10.3892/ol.2019.10270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lai T., Wang K., Hou Q., et al. Interleukin 17 induces up-regulation of chemokine and cytokine expression via activation of the nuclear factor κB and extracellular signal–regulated kinase 1/2 pathways in gynecologic cancer cell lines. International Journal of Gynecological Cancer . 2011;21(9):1533–1539. doi: 10.1097/igc.0b013e31822d2abd. [DOI] [PubMed] [Google Scholar]
- 42.Yuan G., Lian Z., Liu Q., et al. Phosphatidyl inositol 3-kinase (PI3K)-mTOR inhibitor PKI-402 inhibits breast cancer induced osteolysis. Cancer Letters . 2019;443:135–144. doi: 10.1016/j.canlet.2018.11.038. [DOI] [PubMed] [Google Scholar]
- 43.Jouali F., Marchoudi N., Talbi S., et al. Detection of PIK3/AKT pathway in moroccan population with triple negative breast cancer. BMC Cancer . 2018;18(1):p. 900. doi: 10.1186/s12885-018-4811-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Dong L., Du M., Lv Q. Picropodophyllin inhibits type I endometrial cancer cell proliferation via disruption of the PI3K/Akt pathway. Acta Biochimica et Biophysica Sinica . 2019;51(7):753–760. doi: 10.1093/abbs/gmz055. [DOI] [PubMed] [Google Scholar]
- 45.Fan Y., Dong Z., Shi Y., Sun S., Wei B., Zhan L. NLRC5 promotes cell migration and invasion by activating the PI3K/AKT signaling pathway in endometrial cancer. Journal of International Medical Research . 2020;48(5) doi: 10.1177/0300060520925352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wu P. K., Becker A., Park J. I. Growth inhibitory signaling of the raf/MEK/ERK pathway. International Journal of Molecular Sciences . 2020;21(15):p. 5436. doi: 10.3390/ijms21155436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Coelho M. A., de Carné Trécesson S., Rana S., et al. Oncogenic RAS signaling promotes tumor immunoresistance by stabilizing PD-L1 mRNA. Immunity . 2017;47(6):1083–1099. doi: 10.1016/j.immuni.2017.11.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Chiu H. C., Li C. J., Yiang G. T., Tsai A. P., Wu M. Y. Epithelial to mesenchymal transition and cell biology of molecular regulation in endometrial carcinogenesis. Journal of Clinical Medicine . 2019;8(4):p. 439. doi: 10.3390/jcm8040439. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.