

Summary
Adult cortical areas consist of specialized cell types and circuits that support unique higher-order cognitive functions. How this regional diversity develops from an initially uniform neuroepithelium has been the subject of decades of seminal research, and emerging technologies, including single-cell transcriptomics, provide a new perspective on area-specific molecular diversity. Here we review the early developmental processes that underlie cortical arealization, including both cortex intrinsic and extrinsic mechanisms as embodied by the protomap and protocortex hypotheses, respectively. We propose an integrated model of serial homology whereby intrinsic genetic programs and local factors establish early transcriptomic differences between excitatory neurons destined to give rise to broad “proto-regions”, while activity-dependent mechanisms lead to progressive refinement and formation of sharp boundaries between functional areas. Finally, we explore the potential of these basic developmental processes to inform our understanding of the emergence of functional neural networks and circuit abnormalities in neurodevelopmental disorders.
Introduction
The cerebral cortex is responsible for many of the higher-level cognitive functions in humans including language, perception, decision-making, and motor planning. Anatomically, the cerebral cortex can be broadly divided into neocortex (or isocortex) and allocortex, which includes structures such as the hippocampus and olfactory cortex and comprises a relatively larger proportion of cortical area in lower mammals such as mice. Neocortex is defined by the presence, at some point during development, of six distinct anatomical layers parallel to the cortical surface with alternating cell density (layers I, III, and V being relatively cell-sparse and layers II, IV, and VI being relatively cell-dense). It remains unclear whether the neocortex truly represents a phylogenetically newer anatomic structure (Northcutt and Kaas, 1995) and so the term “isocortex” may be more appropriate.
At the cellular level, the cerebral cortex is composed of approximately 16 billion neurons, accounting for more than 80% of the total brain mass (Azevedo et al., 2009). The astonishing diversity of neuronal types has been appreciated since the days of Santiago Ramon y Cajal, and yet we still lack a comprehensive overview of neuronal diversity in the cerebral cortex, in particular how morphological, molecular, and physiological diversity relates to functional areas of the cortex. Even less is known about the developmental processes that give rise to these diverse cell types. Recent advances in molecular profiling, including those with single cell resolution, have created the opportunity to revisit these developmental processes at the level of genes and regulatory pathways (Lein et al., 2017). Here we review the current evidence and potential mechanisms of cortical arealization with a particular focus on human cortical development, although studies using model organisms are also included.
Manifestations of Cortical Arealization in the Adult
Differences in cortical cytoarchitecture including abrupt changes in cell number, density and lamination, and the appearance of morphologically distinct cell types have long been recognized (Brodmann, 1909; Defelipe et al., 1999; von Economo and Koskinas, 1925). These histological differences have been used to define anatomical areal subdivisions and often correspond to specialized functional areas (Bayer and Altman, 1991; Elston, 2003; Hof and Nimchinsky, 1992). Across the cortex, the density of neurons varies by as much as two-fold, primarily due to an increase in the density of upper (“supragranular”) cortical layer neurons in the caudal and medial areas (Charvet et al., 2015). Several of the most distinct area-specific lamination patterns are highlighted in Figure 1A. One of the most striking examples is the abrupt transition from secondary visual cortex to primary visual cortex, where a single layer IV abruptly splits into three sublayers. In other cortical areas, including the primary motor cortex, layer IV is not apparent in histological sections. Similarly, the insular cortex shows a unique lamination pattern in which layer VI is split into sublaminae which are continuous with the adjacent claustrum (Watson and Puelles, 2017). Even within a single functional area, such as primary motor cortex, the cell and neuron density may vary in a topographic manner. and these differences are thought to reflect changes in the cortical circuitry (Young et al., 2013a).
Figure 1: Arealization of the human cerebral cortex.

A) Classical cytoarchitectonic areas described by (Brodmann, 1909). B) Areal differences in local microcircuit architecture between granular and agranular cortices modified from (Beul and Hilgetag, 2015; Shipp, 2005). C) Hierarchical organization between cortical areas, inferred and/or modified from (Badre and Nee, 2018; Felleman and Van Essen, 1991; Ventre-Dominey, 2014).
At the cellular level, systematic studies mapping diversity in the brain have emphasized the remarkable conservation of basic cell types across cortical areas (Harris and Shepherd, 2015). For example, although primary motor cortex has no obvious layer 4 on routine histological sections, excitatory neurons with connectivity profiles typical of L4 neurons have recently been described (Yamawaki et al., 2014), suggesting a conservation of basic cell types and circuit motifs across cortical areas even with diverse cytoarchitecture and function. Across functional areas, variations in cellular composition are hypothesized to contribute to area-specific variations of microcircuits. Notably, specialized cell types with distinct morphologies and localization include the “Betz” cells in layer V of primary motor cortex and “von Economo” neurons in layer V of certain areas in the frontal, insular and anterior cingulate (von Economo and Koskinas, 1925). In rodents, spiny stellate neurons are much more abundant in layer 4 of the primary somatosensory cortex than pyramidal neurons (Egger et al., 1999; Woolsey et al., 1975), while the reverse is true in the visual cortex (Peters and Kara, 1985; Saez and Friedlander, 2009; Scala et al., 2019). Regional variations in the numbers of many other types of neurons and receptors have been documented throughout the cerebral cortex (Ding et al., 2016; Hendry et al., 1987; Palomero-Gallagher and Zilles, 2017; Schleicher et al., 2000; Xu et al., 2010). Ongoing large-scale efforts that seek to comprehensively characterize the cellular composition and connectivity of cortical areas (Gouwens et al., 2018; Jiang et al., 2015; Markram et al., 2015) have the potential to systematically compare cellular composition and local microcircuitry that may underlie functional variations across cortical areas and give rise to their higher-level hierarchical organization (Figure 1B–1C).
At the molecular level, initial bulk transcriptomic studies did not reveal major differences in gene expression between cortical areas in adult humans (Khaitovich et al., 2004; Roth et al., 2006; Xu et al., 2018), consistent with the idea that different areas may utilize a conserved set of basic cell types. Nonetheless, limited transcriptional differences were still identified between cortical areas in the adult human brain (Hawrylycz et al., 2015, 2012), including a handful of genes that show abrupt changes in expression at the boundary between the primary and secondary visual cortex (visual cortical areas 17 and 18) (Zeng et al., 2012). Because bulk gene expression studies rely on pooling transcripts from millions of cells, cellular composition differences can significantly confound the discovery of cell type-specific gene expression profiles, particularly for rare cell populations or closely related subtypes (Kelley and Oldham, 2015). To overcome this limitation, single-cell RNA sequencing has recently emerged as a powerful tool for data-driven classification of cell type homology across anatomical areas and species (Aevermann et al., 2018; Hodge et al., 2018). Using this approach, two recent studies in the adult mouse have compared neurons across anatomical regions of the cerebral cortex and revealed that molecularly-defined excitatory neuron subtypes segregate according to the area from which they were sampled, while interneuron subtypes were shared across areas (Saunders et al., 2018; Tasic et al., 2018). Although this level of coverage has not yet been achieved in the adult human, initial characterization using in situ hybridization (Zeng et al., 2012) and single-nucleus RNA-sequencing (Hodge et al., 2018; Lake et al., 2018) indicate that differences in gene expression may also define area- or region-specific subtypes of excitatory neurons in the human cerebral cortex.
The presence of transcriptomically distinct cell types in different cortical areas may impact area-specific computations at multiple levels including the input-output function of individual neurons (London and Hausser, 2005) and the formation of local and long-distance connections (Averbeck et al., 2009; Harris and Shepherd, 2015; Markov et al., 2013). Testing these predictions will require further validation using emerging technologies to relate molecular cell types to classical definitions based on morphology and physiology. Techniques such as Patch-seq now enable combined recovery of morphology, physiology, and gene expression in the same cells (Cadwell et al., 2016; Fuzik et al., 2016; Scala et al., 2019), and will facilitate generation of a more complete inventory of cell types in the brain. In addition, profiling of epigenetic states at single cell resolution provides an additional molecular perspective on cell type classification (Gray et al., 2017; Luo et al., 2017), and may facilitate the generation of novel tools for visualizing, targeting, and replacing cell types in the brain (Graybuck et al., 2019; Hrvatin et al., 2019; Mich et al., 2019).
Overview of Cortical Development
The cytoarchitectonic and molecular features that distinguish cortical areas discussed above represent only a few examples sampled from a vast body of work. Despite the multitude of well-characterized differences between cortical areas in the adult brain, many of these differences are not apparent during development. The central question of how cortical areas form during development has been the subject of studies that span several decades. In this review, we focus on discussing early developmental events that establish broad distinctions among the major cortical regions. Two hypotheses have been postulated to explain how the apparently uniform processes of proliferation and differentiation during development give rise to the area-specific variations of the adult cortex. First, the “protomap” hypothesis theorizes that progenitors are pre-patterned to give rise to region-specific neurons with distinct molecular, physiological, and connectivity features (Bishop et al., 2000; Rakic, 1988; Rubenstein and Rakic, 1999). Second, the “protocortex” hypothesis suggests that the cortex is initially homogeneous and develops area-specific features only in response to extrinsic signals, most notably thalamic input (Van der Loos and Woolsey, 1973; Creutzfeldt, 1977; O’Leary, 1989). In the following sections, we will discuss the evidence behind both theories and highlight how novel molecular techniques are beginning to shed light on these long-standing questions.
Progenitors of the Cerebral Cortex
Before the onset of neurogenesis, the neuroepithelium is composed of neuroepithelial stem (NES) cells which undergo symmetric divisions that expand the neuroepithelial surface area and determine the size of the ‘founder’ population of neurogenic ventricular radial glia (vRG) cells, whose cell bodies are located in the ventricular zone (VZ). Already at these early stages, morphogen gradients, including FGF expressed by the anterior neural ridge, Wnt and BMP signaling from the roof plate, and Sfrp1 and TGFa from the ventral pallium, induce the expression of patterning transcription factors in the neuroepithelium and vRG cells (Bayatti et al., 2008; Mallamaci and Stoykova, 2006; Rubenstein et al., 1998). These patterning factors subsequently induce the expression of transcription factors that execute region-specific neurodevelopmental programs and control the cell fate of neurons born in different brain regions. In the neocortex, these transcription factors include EMX1, EMX2, PAX6, LHX2, and COUP-TF1 (Figure 2A). Many of these factors are expressed preferentially in the vRG cells and their loss of function results in changes in the cortical area map, indicating that patterning transcription factors may contribute to the “protomap” formation (O’Leary et al., 2007).
Figure 2: Timing of neurodevelopmental events in the cerebral cortex.

A) Key patterning centers set up morphogen and transcription factor gradients across the developing neuroepithelium to influence area-specific cell fates. B) Schematic comparison of the mouse and human cerebral cortical development at peak stages of neurogenesis. While the processes are broadly conserved, several important differences can be highlighted: prominent expansion of the outer subventricular zone (OSVZ) and of the outer radial glia population which resides therein (Fietz et al., 2010; Hansen et al., 2010; Reillo et al., 2011); expansion of the subplate (SP), which is associated with the expansion of the axonal plexus, but not necessarily cell numbers (Duque et al., 2016); expansion of the upper layer excitatory neuron types (Hodge et al., 2018); discontinuation of the radial glia scaffold (Nowakowski et al., 2016); emergence of persisting Cajal-Retzius cells (Meyer and Gonzalez-Gomez, 2018); expansion of axonal plexus in the OSVZ and the emergence of multilaminar axonal-cellular compartment (Zunic Isasegi et al., 2018); early arrival of thalamocortical plexus in the cortical anlage (Marin-Padilla, 1983). C) Broad comparison of key neurogenesis periods across species, including data from (Clancy et al., 2001; Rakic, 1974). Embryonic days (E) next to species names indicate approximate length of gestation.
Around post-conception week (pcw) 12 of human development, a distinct anatomical zone called the outer subventricular zone (OSVZ) forms and is populated by proliferative progenitor cells (Rakic and Sidman, 1968a, 1968b; Sidman and Rakic, 1973; Smart et al., 2002), including cells that are derived from vRG and retain neural stem cell potential, but adopt a distinct morphology, dynamic cell behavior, and molecular identity (Figure 2B). These cells are called outer radial glia (oRG) (Hansen et al., 2010; Pollen et al., 2015; Thomsen et al., 2016), and are also referred to as basal radial glia (Fietz et al., 2010; Reillo et al., 2011). It is currently unknown whether the same molecular patterning seen across vRG cells is relayed to the oRG cells. Across the cortical neuroepithelium, the abundance of oRG cells varies, with abundant oRG cell populations found underneath the prospective gyri (Nowakowski et al., 2016; Smart et al., 2002), consistent with the hypothesis that basal progenitors may play an important role in gyrification of the cerebral cortex (Kriegstein et al., 2006; Reillo et al., 2011). Control of the cell cycle duration and rate of cell cycle re-entry of oRG cells may play an important role in regulating the tangential extent of individual areas, as well as area-specific cytoarchitectonics and neuronal numbers (Lukaszewicz et al., 2005).
The molecular mechanisms regulating the timing of oRG cell production are unclear but are thought to involve well-established developmental signaling pathways including LIF and Shh (Martmez-Martmez et al., 2016; Matsumoto et al., 2017; Stahl et al., 2013; Wang et al., 2016; Watanabe et al., 2017). During human supragranular layer production, the vast majority of pia-contacting radial glia fibers originate from cells located in the OSVZ (deAzevedo et al., 2003; Nowakowski et al., 2016; Figure 2B). Neither mouse nor ferret cortical development involves such a dramatic developmental transition of the radial glia scaffold, and in these species continuous radial glia fiber connections exist between ventricular and pial surfaces of the cortex even at the end of neurogenesis (Reillo et al., 2011). In the developing ferret, generation of oRG cells is confined to a narrow time window during deep layer neurogenesis (Martmez-Martmez et al., 2016) and involves asymmetric division of vRG cells with one (basal) daughter cell inheriting the pia-contacting fiber (LaMonica et al., 2013). The daughter cell that remains in contact with the ventricle begins to regrow the basal fiber but does not reach the pial surface and is unable to support radial migration to the cortical plate (LaMonica et al., 2013; Nowakowski et al., 2016). These “truncated” radial glia cells emerge sharply around pcw 14–14.5 in humans and acquire a distinct molecular and morphological phenotype (Nowakowski et al., 2016; Rakic, 1978).
The absolute increase in surface area and number of cortical neurons combined with the selective expansion of frontal and parieto-temporal cortical regions is thought to underlie the emergence of advanced cognitive abilities in humans including perception, language, and reasoning (Geschwind and Rakic, 2013). Because of the central role of radial glia fibers in supporting neuronal migration to the cortical plate (Rakic, 1972), the total number of pia-contacting radial fibers directly predicts the total cortical surface area (Rakic, 1988). Expansive divisions of vRG and NES cells could serve as a fundamental mechanism for regulating the size of any given cortical area (Rakic, 1988). Consistent with this hypothesis, functional experiments in mice promoting NES and vRG expansion, preventing their elimination, or delaying NES differentiation into vRG cells have been shown to result in an expansion of the tangential surface area of the cortical neuroepithelium (Beattie et al., 2017; Chenn and Walsh, 2002, 2003; Haydar et al., 1999; Kuida et al., 1996; Li et al., 2017; Sahara and O’Leary, 2009). In addition, programmed cell death as a mechanism regulating cortical surface area may be underappreciated, although reports of the extent of cell elimination during normal cortical development vary depending on analysis method (Kuida et al., 1996; Rakic and Zecevic, 2000; Wilkie et al., 2004). To what extent these pathways converge and interplay with classical mechanisms of organ size control remains largely unclear (Kunche et al., 2016; Tumaneng et al., 2012). Whether oRG cells directly contribute to the tangential expansion of the cortical surface area also remains unclear, but experiments inducing overproduction or proliferation of OSVZ progenitors in ferrets have suggested a role for this cell type in regulating cortical surface area in mammals (Matsumoto et al., 2017; Nonaka-Kinoshita et al., 2013). Given that the majority of oRG cells are generated at later stages of development than vRG cells, expansive divisions of the oRG cell population would be predicted to disproportionately expand the surface area and the number of neurons in the upper layers of the cortex relative to deep layers, but evidence for such expansion is currently limited (Borrell, 2018; Nowakowski et al., 2016; Richman et al., 1975; Stahl et al., 2013; Wang et al., 2016). Further studies into the mechanisms of cortical area expansion could advance our understanding of selective expansion of specific cortical areas in humans compared to non-human primates.
During peak excitatory neurogenesis, two distinct modes of neuronal production have been described. The direct neurogenesis pathway involves neuronal differentiation directly from asymmetric division of vRG cells and is believed to be more prevalent at early stages of cortical development, at least in mice (Miyata et al., 2001; Noctor et al., 2001). The second mode of neuronal differentiation involves an intermediate progenitor cell (IPC) that divides in the subventricular zone (SVZ). IPCs may re-enter the cell cycle to divide, usually symmetrically, and produce postmitotic neurons (Haubensak et al., 2004; Noctor et al., 2004; Figure 3). In contrast to radial glia, IPCs do not maintain continuous connections with the pial or ventricular surfaces and undergo a stationary division. They also tend to adopt a multipolar morphology, although recent studies suggest that IPCs may also adopt unipolar or bipolar morphologies in primates (Betizeau et al., 2013; Kriegstein and Noctor, 2004; Pollen et al., 2015).
Figure 3: Maturation and differentiation in the cortex.

A) Left: schematic representing radial glia maturation from neuroepithelial stem cells (NESC), followed by their differentiation into astrocytes. Right: schematic representing sequential production of cortical layers from radial glia in the mouse. B) in contrast, human cortical development involves an expanded diversity of radial glia with distinct maturation trajectories (left). Right: neurogenesis in the human cortex occurs in the ventricular zone early in development, and progressively shifts towards the outer subventricular zone. VZ-ventricular zone, SVZ - subventricular zone, ISVZ- inner subventricular zone, OSVZ- outer subventricular zone, tRG-truncated radial glia, vRG- ventricular radial glia, NESC- neuroepithelial stem cells, IPC- intermediate progenitor cells, oRG- outer radial glia, WM- white matter, LI-VI - cortical layer I-VI.
The role of intermediate progenitors in the cortex remains an area of active research, but recent studies suggest that this cell type may play an important role in cell fate decisions in the developing cortex. For example, the IPC population may be more heterogeneous than previously appreciated, and some IPCs generated early in development may be maintained in a quiescent state until upper layer neurogenesis (Mihalas et al., 2016). IPCs are believed to play a critical role in regulating the number of excitatory neurons in the cerebral cortex through transit amplification of the neuronal output from radial glia (Pontious et al., 2008), and may contribute to variations in cortical thickness across areas (Pontious et al., 2008), suggesting that they too may have an essential role in determining cortical area-specific neuronal composition. Consistent with these findings, there are areal differences in the fraction of neurons that are derived from IPCs, particularly in layers 4 and 5 (Vasistha et al., 2015). Other roles have also been attributed to this cell type, including promoting tangential migration of interneurons to the cortex (Sessa et al., 2010) and contributing to the maintenance of vRGs (Yoon et al., 2008). Interestingly, human IPCs located in the outer subventricular zone (OSVZ) (Smart et al., 2002) appear to have a substantially increased proliferative capacity, are able to undergo multiple rounds of expansive division (Hansen et al., 2010; Kriegstein et al., 2006), and are hypothesized to adopt diverse morphologies and possibly also differentiate into radial glia (Betizeau et al., 2013). Many interesting questions remain, such as whether IPCs in different cortical areas show distinct molecular properties or fate potential, and, if so, how area-specific identities are passed from radial glia to IPCs.
Differentiation and Maturation of Excitatory Neurons
The developmental time-window of neurogenesis varies widely across species. In mice, neurons are generated between embryonic days 11 and 17 (Caviness et al., 1995), whereas in the human cortex, neuronal production commences around pcw 6 and extends through mid-gestation, possibly continuing into the third trimester (Malik et al., 2013; Rash et al., 2019; Figure 2C). Newborn neurons utilize radial glia fibers to arrive at the cortical plate (Rakic, 1988), and their journey is orchestrated by a complex network of molecular factors and signaling pathways (Marin and Rubenstein, 2003), which will not be reviewed here. Neurons arising from the same group of progenitors migrate radially to occupy nearby positions within the adult cerebral cortex (Rakic, 1972, 1978), leading Rakic to propose the Radial Unit Hypothesis of cortical expansion, whereby changes in the number of founder cells affect the number of radial units without substantially altering the number of neurons within each unit (Rakic, 2008).
It is estimated that radial migration in the developing macaque cortex takes approximately 3–7 days in early development (at E46), and longer than 7 days around E90 (Rakic, 1975). Lineage tracing studies have shown that the majority of excitatory neurons migrate radially, with some tangential dispersion (Gertz and Kriegstein, 2015; Reillo et al., 2011; Torii et al., 2009). Tangential migration of excitatory neurons has been demonstrated in multiple species, including the mouse, where tangentially-migrating neurons contribute to the development of the subiculum (Britanova et al., 2006). Tangential dispersion is more prominent in gyrencephalic animals such as the ferret, where both increased dispersion of radial glial fibers (Reillo et al., 2011) and sequential migration of excitatory neurons along adjacent radial glial fibers (Gertz and Kriegstein, 2015) are thought to contribute to cortical surface area expansion.
Cajal-Retzius and subplate cells are the first neurons that populate the cerebral cortex, and their neurogenesis occurs prior to the generation of excitatory neurons (Angevine and Sidman, 1961; Hevner et al., 2003; Meyer et al., 2000; Price et al., 1997). However, a recent study suggested that a second wave of Cajal-Retzius cells may emerge around mid-gestation in humans (Meyer and Gonzalez-Gomez, 2018). Both of these waves are generated not just locally in the cortex, but also in other brain regions, with cells migrating tangentially to the cortex (Griveau et al., 2010; Pedraza et al., 2014). In many species, including the mouse, Cajal-Retzius cells and subplate neurons largely disappear during early development (Griveau et al., 2010, but see also Chowdhury et al., 2010 and Hoerder-Suabedissen and Molnar, 2013). In humans, however, Cajal-Retzius cells persist in the outermost layers of the cortex, while subplate neurons give rise to the interstitial white matter neurons (Kostovic and Rakic, 1980; Marin-Padilla, 2015).
The vast majority of excitatory neurons destined for layers II-VI are generated in an ‘inside out’ fashion, such that early-born neurons contribute to the deepest layers of the cortex, followed by subsequent waves of neurogenesis destined to progressively more superficial layers (Angevine and Sidman, 1961; Figure 3). Although the exact role of cortical layering is still unclear (Adesnik and Naka, 2018), laminar position strongly correlates with projection cell type and connectivity profile (Harris and Shepherd, 2015). Upon arrival at the cortical plate, excitatory neurons undergo further maturation and can be classified according to their laminar position, gene expression profile, and projection type. Classically, three broad classes of projection neurons have been described for cortical excitatory neurons (Harris and Shepherd, 2015). Corticothalamic neurons project to the thalamus, their cell bodies occupy layer VI, and they express the transcription factor TBR1. Pyramidal tract neurons project to the spinal cord, but often possess collateral projections to the thalamus and the medulla (Economo et al., 2018). These cells mainly occupy layer V, and express transcription factors FEZF2 and BCL11B (previously known as Ctip2). Intratelencephalic (IT) neurons project to targets within the forebrain, including the striatum, ipsi- or contralateral hemisphere of the cortex. IT neurons are located in layers II-VI and are characterized by the expression of Satb2. Excellent reviews have provided detailed descriptions of the molecular markers of neurons in different layers, and projection classes (Harris and Shepherd, 2015; Lodato and Arlotta, 2015; Molyneaux et al., 2007). In the adult human, layer-specific dissections of the cortex followed by single-nucleus sequencing have identified human-specific markers of cortical layers in the medial temporal gyrus (Hodge et al., 2018), as well as new and previously described markers of layer identity (Zeng et al., 2012). Interestingly, most molecularly-defined cell types in the adult human cortex appear to be present in more than one layer, suggesting that a combination of molecular identity and layer position may interact to generate a wide range of cell types both within and across cortical areas.
Mechanisms regulating the decision of cortical neurons to adopt a particular laminar fate remain unclear. A bulk RNA-sequencing study spanning prenatal to young adult time periods in macaques found that a number of genes are differentially expressed between prefrontal cortex and occipital cortex, and that markers of layer identity emerge between E80 and E90 and are fully defined by birth (Bakken et al., 2016). In both macaque and human, birthdates of excitatory neurons roughly correlate with upper versus deep layer neuronal identity, but distinctions within these groups are not refined until much later (Bakken et al., 2016; Miller et al., 2014). By contrast, in the mouse, a number of birth dating experiments have shown laminar identity is more precisely determined by neuronal birth date (Greig et al., 2013; Telley et al., 2016).
Development of inhibitory cortical interneurons
Cortical inhibitory interneurons are extraordinarily diverse in terms of their morphology and physiology, and their numbers have increased relative to excitatory neurons over the course of evolution (Defelipe, 2011) along with the appearance of unique interneuron subtypes in humans (Boldog et al., 2018; DeFelipe et al., 2002). Cortical interneurons are generated in the medial and caudal ganglionic eminences, as well as the preoptic area, and migrate tangentially to integrate into local cortical circuits (Faux et al., 2012; Harwell et al., 2015; Mayer et al., 2015). While some studies suggest that clonally related interneurons are not randomly distributed in the cortex (Brown et al., 2011; Sultan et al., 2016), it is unclear whether this is functionally relevant given the wide spatial distribution (1500 μm average inter-soma distance) of clonally related interneurons, which also cross functional boundaries (Mayer et al., 2016; Turrero Garcia et al., 2016). While in most species interneuron migration occurs embryonically, recent studies in the human cortex suggest an extended period of interneuron production which continues through 7 months after birth, at least in the frontal lobe (Paredes et al., 2016). Parvalbumin-positive (PV+) interneurons form dense connections with excitatory neurons leading to the postulation of a “blanket inhibition” hypothesis (Beutler and Kuhl, 1986; Fino and Yuste, 2011; Packer and Yuste, 2011). Yet, neighboring PV+ interneurons are differentially recruited into neuronal ensembles and are thought to participate in complex cognitive computations (Lagler et al., 2016). A recent study in mice has shown that certain circuits between inhibitory interneurons and excitatory neurons form transiently during development and may contribute to normal formation of mature local and thalamocortical microcircuits (Marques-Smith et al., 2016).
Several studies have shed light on the developmental mechanisms guiding interneuron migration and wiring. Analysis of the Fezf2 mutant mouse, in which deep-layer subcerebral excitatory projection neurons are missing, shows a mislamination of deep-layer interneurons while upper-layer interneurons remain unaffected (Lodato et al., 2011). In a complementary study, the Satb2 knockout model, in which callosal projection neurons are misspecified, shows a specific mislamination of upper-layer interneurons, while deep-layer interneurons are unaffected (Wester et al., 2019). In addition, induction of specific types of cortical excitatory neurons in an ectopic, subcortical location results in recruitment of interneurons of the correct cortical layer identity (Lodato et al., 2011), suggesting that extrinsic cues from excitatory neurons, not birthdate, is a key determinant of appropriate layer-specific interneuron migration and positioning. In addition to the precise cell type-specific wiring diagram described among different morphologically distinct subtypes of interneurons (Jiang et al., 2015), recent studies also suggest that interneurons form synaptic connections with different subtypes of excitatory neurons in a cell type-specific manner. For example, parvalbumin-positive interneurons in layer 5 of the prefrontal cortex preferentially inhibit corticofugal projection neurons over neighboring callosal neurons (Lee et al., 2014). Similarly, in the medial entorhinal cortex, parvalbumin-positive and cannabinoid type-1-receptor-positive interneurons in layer 2 preferentially synapse onto calbindin-positive callosal projection neurons while avoiding neighboring reelin-positive projection neurons that project to the dentate gyrus (Varga et al., 2010; Armstrong et al., 2016). Fate conversion experiments in which postmitotic layer 2/3 callosal projection neurons are reprogrammed to subcortically-projecting neurons recruit perisomatic input from parvalbumin-positive interneurons to levels similar to that of endogenous subcortically-projecting neurons found in layer 5 despite their ectopic location in layer 2/3 (Ye et al., 2015). These findings suggest that the identity of projection neurons, at least in part, dictates the establishment of appropriate inhibitory synapses in a class-specific manner.
Interneurons have been proposed to control the rate of maturation of cortical circuits and plasticity. One recent study has suggested a role for inhibitory interneurons in activity-dependent plasticity in adult mice (Adler et al., 2019). Heterochronic transplantation experiments in which embryonic interneurons are transplanted into juvenile and adult cortex have shown that interneurons, and particularly MGE-derived interneurons, can reopen ocular dominance plasticity well beyond the critical period (Davis et al., 2015; Southwell et al., 2010). However, the opposite experiment of transplanting mature interneurons to an embryonic circuit to determine whether it can precipitate faster than normal maturation has not yet been performed, and it remains unclear whether interneurons act as bidirectional master regulators of ocular dominance plasticity.
Interneurons are disproportionately affected by apoptosis compared to excitatory neurons, with an estimated loss of ~30% of interneurons during early postnatal development compared to ~12% of excitatory neurons (Wong et al., 2018). This process is regulated at least in part by activity-dependent mechanisms (Priya et al., 2018). Interestingly, deep-layer interneurons die earlier than interneurons of the upper layers, and waves of interneuron apoptosis are preceded by projection neuron apoptosis suggesting that these two events may be linked (Wong et al., 2018). Furthermore, projection neuron activity was shown to promote the survival of interneurons via negative regulation of PTEN (Wong et al., 2018). These findings suggest that one mechanism by which excitatory neurons orchestrate the assembly of region-specific cellular components could be via regulation of inhibitory interneuron survival.
Molecular correlates of areal identity
Molecular signatures of positional identity are apparent from an early developmental stage. In the embryonic and early postnatal mouse, a number of genes show differential expression across cortical regions (Bishop et al., 2000). For example, while Cad8 is broadly expressed across the entire cortex within layer 5, it is also highly expressed in layer 2/3 in rostral cortical areas (Nakagawa et al., 1999; Suzuki et al., 1997). Cad6 expression is enriched in the dorsal and lateral developing isocortex corresponding to the prospective somatosensory and auditory areas but is lowly expressed, especially in layers 4 and 5, in the extreme rostral isocortex (Nakagawa et al., 1999; Suzuki et al., 1997). RORB (formerly known as RZR-beta) and p75 (also known as NGFR) show opposing graded expression patterns across the isocortex, with RORB expressed in regions rostral to the developing visual cortex while p75 expression is found only in the subplate of caudal regions. (Mackarehtschian et al., 1999; Rubenstein et al., 1999). EphA7 is expressed throughout most of the developing isocortex but at much lower levels in intermediate regions corresponding to the somatosensory area (Rubenstein et al., 1999). Finally, ephrin-A5 is expressed most highly in the dorsolateral isocortex corresponding to the somatosensory area (Mackarehtschian et al., 1999). These are just a few of the many examples of area-specific gene expression patterns, and the major findings from these and other studies are summarized in Table 1.
Table 1.
Area-enriched genes and knock out phenotypes.
| Gene | Organism | Expression Pattern | Mouse Knockout Cortical Phenotype | Relevant References |
|---|---|---|---|---|
| AUTS2 | Human; Mouse | Frontal cortex | Neonatal lethal in mouse. Defects in neuronal migration, axonal elongation and neuritogenesis in mouse embryos | Bedogni et al., 2010a, 2010b; Hori et al., 2014; Srinivasan et al., 2012 |
| BCL6 | Mouse | Somatosensory cortex at birth. Rostral in adulthood | Unknown | Leamey et al., 2008 |
| BHLHB5 | Mouse | Postmitotic neurons of primary sensory areas (visual, somatosensory and auditory cortex) Layers II-V | Aberrant gene expression and structural organization of somatosensory and caudal motor cortices | Joshi et al., 2008 |
| BLN2 | Human | High rostro-caudal gradient | Unknown | Lambert et al., 2011 |
| C13ORF38 | Human | High expression in temporal lobe | Unknown | Pletikos et al., 2014 |
| CBLN2 | Human; Mouse | High-to-low rostro-caudal gradient | Unknown | Johnson et al., 2009; Miura et al., 2006 |
| CDH6 | Mouse | High-to-low caudal-rostral gradient in postmitotic neurons | Unknown | Terakawa et al., 2013 |
| CDH8 | Ferret; Human; Macaque; Mouse | Frontal cortex | Autism-like behavior; reduced neuronal complexity | Bernier et al., 2014; Friedman et al., 2015; de Juan Romero et al., 2015 |
| CLMP | Human | High-to-low rostro-caudal in human. No areal differences in macaque | Unknown | Pletikos et al., 2014 |
| CNTN4 | Human | High rostro-caudal gradient | Unknown | Lambert et al., 2011 |
| CNTNAP2 | Human | High-to-low rostro-caudal gradient | Reduced spine density and levels of GluA1 of AMPA receptors in spines | Johnson et al., 2009; Lambert et al., 2011; Varea et al., 2015 |
| CPNE8 | Human | Frontal cortex, upper layer excitatory neurons | Unknown | Lambert et al., 2011; Nowakowski et al., 2017 |
| CRYM | Mouse | High-to-low caudal-rostral gradient | Unknown | Bedogni et al., 2010b; Leamey et al., 2008 |
| CUX1 | Mouse | Higher in somatosensory than in motor cortex in layers 2–4 | Aberrant dendritic morphology of upper-layers projection neurons | Cubelos et al., 2015; Nieto et al., 2004 |
| DIAP3 | Mouse | Subcerebral projection neurons of the motor cortex | Unknown | Arlotta et al., 2005 |
| DKK3 | Mouse | Visual cortex | No obvious phenotype in cortex | (Barrantes et al., 2006; Leamey et al., 2008) |
| DUSP6 | Mouse | Somatosensory cortex | Unknown | Leamey et al., 2008 |
| EOMES | Ferret; Human | High-to-low caudal-rostral gradient. No gradient is observed in mouse. | Microcephaly and behavioral defects. Reduction of upper layer projection neurons | Arnold et al., 2008; de Juan Romero et al., 2015; Nowakowski et al., 2017 |
| EPHA3 | Macaque | High-to-low caudal-rostral gradient in macaque subventricular and intermediate zones | No obvious phenotype | Donoghue and Rakic, 1999; Vaidya et al., 2003 |
| EPHA5 | Mouse | Somatosensory cortex | Formation of abnormal cell aggregates and abnormal spine morphology | Cooper et al., 2009; Das et al., 2016 |
| EPHA6 | Mouse; Macaque | Cortical plate of developing occipital cortex | Formation of abnormal cell aggregates and abnormal spine morphology | Bedogni et al., 2010b; Das et al., 2016; Donoghue and Rakic, 1999 |
| EPHA7 | Macaque; Mouse | High-to-low rostro-caudal in mouse. Caudal-rostral gradient in macaque early development (E65) | Dendrites are longer and more complex | Clifford et al., 2014; Donoghue and Rakic, 1999 |
| EMX2 | Human; Mouse | High-to-low medial-lateral gradient | Reduction of caudal-medial identity | Gulisano et al., 1996; Ip et al., 2010; Mallamaci and Stoykova, 2006 |
| FGF17 | Mouse | Frontal cortex | Reduction of dorsal frontal cortex; rostral shift in cortical sensory areas | Cholfin and Rubenstein, 2007 |
| FGRF2 | Ferret; Mouse | High-to-low caudal-rostral gradient in mouse ventricular zone and ferret outer subventricular zone | Reduction of projection neurons in medial Prefrontal cortex observed in conditional knockout in radial glia cells | de Juan Romero et al., 2015; Stevens et al., 2010 |
| FGRF3 | Ferret; Human; Mouse | High-to-low caudal-rostral gradient | Reduction of cortical volume | de Juan Romero et al., 2015; Moldrich et al., 2011; Nowakowski et al., 2017 |
| FLRT3 | Mouse | Visual cortex | Altered distribution of projection neurons during development | Leamey et al., 2008; Seiradake et al., 2014 |
| HCRTR2 | Human | Frontal cortex, deep layer excitatory neurons | Unknown | Nowakowski et al., 2017 |
| ID2 | Mouse | In intermediate and caudal cortex is expressed in layer 5. In frontal cortex is expressed in layer 2/3 | Unknown | Bulfone et al., 1995 |
| IGFBP4 | Mouse | Subcerebral projection neurons of the motor cortex | Unknown | Arlotta et al., 2005 |
| IGSF4A | Mouse | Visual cortex | Unknown | Leamey et al., 2008 |
| KBTBD9 | Mouse | Somatosensory cortex | Unknown | Leamey et al., 2008 |
| LHX2 | Ferret; Mouse | High-to-low caudomedial-rostrolateral gradient during development | Barrel cores and barrel walls in somatosensory cortex fail to form. | Chou et al., 2009; de Juan Romero et al., 2015; Shetty et al., 2013 |
| LIX1 | Mouse | Subcerebral projection neurons of the visual cortex | Unknown | Moeller et al., 2002 |
| LMO3 | Mouse | Parietal cortex | Behavioral defects | Bulchand et al., 2003;Savarese et al., 2014 |
| LMO4 | Human Mouse | Postmitotic neurons of motor cortex and higher order sensory areas | Disrupted molecular identity and connectivity of rostral motor cortex | Cederquist et al., 2013; Lambert et al., 2011 |
| LUZP2 | Mouse | Somatosensory cortex | Unknown | Leamey et al., 2008 |
| NHLH1 | Mouse | High-to-low caudal-rostral gradient | No obvious phenotype | Elsen et al., 2013; Kruger and Braun, 2002 |
| NPN2 | Mouse | High-to-low caudal-rostral gradient | Subcerebral neurons of the visual cortex fail to prune their axons | Low et al., 2008 |
| NPY | Human | High-to-low caudal-rostral gradient. No specificity in mouse | Unknown | Johnson et al., 2009 |
| NR1F2 | Ferret, Mouse, Rat | High-to-low rostro caudal gradient | Unknown | Miyashita-Lin et al., 1999; Rowell et al., 2010;Schaeren-Wiemers et al., 1997 |
| NR2F1 | Human; Mouse | High-to-low caudal-rostral gradient in cortical progenitors and postmitotic neurons | Expansion of motor cortex at expense of sensory areas | Armentano et al., 2006; Nowakowski et al., 2017 |
| NR2F2 | Human; Macaque | High expression in temporal lobe | Unknown | Pletikos et al., 2014 |
| NR4A2 | Human | Parieto-temporal cortex | Unknown | Lambert et al., 2011 |
| ODZ1 | Mouse | High-to-low rostro caudal gradient in layer 4 and subplate neurons | Unknown | Li et al., 2006 |
| ODZ2 | Human; Mouse | Layer 5 neurons of visual cortex | Binocular circuits fail to form | Leamey et al., 2008; Li et al., 2006; Nowakowski et al., 2017; Young et al., 2013b |
| ODZ3 | Mouse | Deep layer neurons of the visual cortex | Formation of eye dominance domains in the visual cortex | Leamey et al., 2008; Li et al., 2006; Merlin et al., 2013 |
| ODZ4 | Mouse | High-to-low caudal-rostral gradient | Hypomyelination of the central nervous system | Leamey et al., 2008; Li et al., 2006; Suzuki et al., 2012 |
| P75 | Rat; Mouse | High-to-low caudal-rostral gradient | Increased excitability of neurons, enhanced cholinergic innervation into visual cortex | von Bohlen und Halbach and von Bohlen und Halbach, 2016; Gibon et al., 2015; Mackarehtschian et al., 1999; Mallamaci et al., 2000 |
| PAX6 | Mouse | High-to-low rostro-caudal latero-medial gradient | Perinatal lethal. Conditional mutant has a dramatic decrease of upper layer neurons, increase of deep layer neurons reaching the marginal zone and increase number of oligodendrocytes. Phenotype is particularly pronounced in frontal cortex | Stoykova et al., 2000; Tuoc et al., 2009 |
| PCDH8 | Mouse | High-to-low caudal-rostral gradient. No specificity in mouse | No obvious phenotype | Bedogni et al., 2010b; Yamamoto et al., 2000 |
| PENK | Human | High rostro-caudal gradient | Unknown | Lambert et al., 2011 |
| PLXA3 | Mouse | High-to-low caudal-rostral gradient early postnatally | No obvious defect. However, in double knockout with Plxa4 the pruning phenotype of Plxa4 is increased | Low et al., 2008 |
| PLXA4 | Mouse | High-to-low caudal-rostral gradient early postnatally | Subcerebral neurons of the visual cortex fail to prune their axons | Low et al., 2008 |
| PLXND1 | Macaque; Mouse | High-to-low caudal-rostral gradient in postmitotic neurons | Hem-derived Cajal-Retzius cells reach more dorsal neocortical regions | Bribián et al., 2014; Watakabe et al., 2006 |
| SATB2 | Human | High-to-low rostro-caudal, co-expressing with BCL11B in deep layers until sharp boundary segregates expression with SATB2 in upper layers | Upper layer callosal projection neurons fail to form | Alcamo et al., 2008; Britanova et al., 2008; Nowakowski et al., 2017 |
| SEMA3E | Mouse; Macaque | High-to-low caudal-rostral gradient in postmitotic neurons | Hem-derived Cajal-Retzius cells reach more dorsal neocortical regions | Bribián et al., 2014; Watakabe et al., 2006 |
| SLN | Human | High rostro-caudal gradient | Unknown | Lambert et al., 2011 |
| SOX9 | Human | Parieto-temporal cortex | Unknown | Lambert et al., 2011 |
| SP8 | Mouse | High-to-low anterior-medial to posterior-lateral gradient in cortical progenitors | Unknown | Sahara et al., 2007 |
| TBR1 | Mouse | High-to-low rostro-caudal | Expression of caudal genes in frontal cortex | Bedogni et al., 2010b |
| THBS1 | Human | High rostro-caudal gradient | Unknown | Lambert et al., 2011 |
| TSHZ2 | Mouse | Occipital cortex | Unknown | Bedogni et al., 2010b |
| USP6 | Mouse | Somatosensory cortex | Unknown | Leamey et al., 2008 |
| VSTM2L | Human | High rostro-caudal gradient | Unknown | Lambert et al., 2011 |
| WNT7B | Human; Macaque | High-to-low caudal-rostral gradient in postmitotic neurons | Unknown | Ozair et al., 2018; Pletikos et al., 2014 |
| WSCD1 | Mouse | High-to-low rostro-caudal in layer 6 neurons and subplate | Unknown | Bedogni et al., 2010b |
The first systematic characterization of areas in human development was performed using exon arrays to explore a number of rostral, temporal and occipital cortical regions as well as subcortical brain regions during mid-gestation (Johnson et al., 2009). Hundreds of genes were found to be differentially expressed between rostral and caudal brain regions, and samples from motor and somatosensory cortices showed an intermediate expression profile, consistent with the idea that a gradient of morphogen and gene expression patterns may give rise to frontal, central and occipital cortical areas (O’Leary et al., 2007). Additionally, distinct transcriptional programs were observed within regions of the developing prefrontal cortex, most notably in the orbital prefrontal cortex (Johnson et al., 2009), indicating that transcriptional mechanisms may also be engaged for intra-areal refinement. Overall, examination of cortical areas during development has shown that the majority of expressed genes in the human brain are differentially expressed across brain regions or time, but these differences universally diminish over the course of development (Kang et al., 2011). Similar trends were observed in the macaque, with the most dynamic changes occurring early in postnatal development (Bakken et al., 2015). A pair of recent studies that surveyed 16 brain regions transcriptomically across developmental time in the human and macaque found a “cup shaped pattern” of development, with large regional differences early in development narrowing, and re-emerging during childhood at which point these transcriptional changes were paired with epigenetic restructuring (Li et al., 2018; Zhu et al., 2018). Interestingly, bulk tissue studies have revealed only limited transcriptional distinctions between cortical areas in the adult human brain as discussed earlier (Hawrylycz et al., 2015, 2012; Khaitovich et al., 2004; Roth et al., 2006; Silbereis et al., 2016).
Single-cell sequencing approaches are providing new insights into the processes of early cortical arealization. For example, single-cell RNA-sequencing has identified hundreds of differentially expressed genes between cells of the same broad class between developing prefrontal (PFC) and primary visual (V1) cortical areas (Nowakowski et al., 2017). Although positional differences in gene expression were apparent at all stages of lineage progression (radial glia, intermediate progenitors, and postmitotic neurons) they became amplified over the course of neuronal differentiation, with only a few transcription factors differentially expressed at the level of radial glia and hundreds of transcripts distinguishing postmitotic neurons. In addition to previously identified markers for developing PFC and V1, novel candidates restricted to specific subtypes of cells in PFC or V1 were identified using this single-cell approach, including CPNE8 in the upper layers of PFC and HCRTR2 in subplate neurons of V1. These transcriptomic signatures of positional identity precede those of laminar specification (Nowakowski et al., 2017), suggesting that molecular correlates of laminar identity emerge later in development. Furthermore, there appeared to be a narrow transition zone between rostral and caudal gene expression patterns, suggesting that rostral and caudal lineages give rise to discrete cell types rather than representing opposite ends of a developmental gradient. The early divergence of area-specific excitatory neurons is consistent with a protomap hypothesis in which multiple excitatory neuron lineages emerge simultaneously across the developing cortex. It is possible that a limited set of transcription factors differentially expressed in radial glia initiate divergent intrinsic gene regulatory programs which are later amplified in an area-specific manner upon differentiation.
The ability to query chromatin state at the single-cell level is a new frontier that expands our ability to identify epigenetic variations across cell types and brain regions (Graybuck et al., 2019; Hrvatin et al., 2019; Mich et al., 2019). Recent work in the adult human shows a strong association between chromatin state and brain region (Lake et al., 2018). In addition, chromatin state strongly correlates with cell type identity (Luo et al., 2017), suggesting that analysis of chromatin or epigenetic states of individual cell types could further elucidate the regulatory processes underlying cell fate specification (Gray et al., 2017). Along these lines, a recent single-cell transcriptomic study in developing mouse showed that the temporal progression of apical progenitor states is epigenetically regulated by the Polycomb repressor complex PRC2 (Telley et al., 2019). Extending these studies across developmental time points and brain regions in humans will further illuminate the extent to which a protomap might be encoded in progenitor cells.
Extrinsic mechanisms of cortical arealization
In addition to early patterning of the cortical neuroepithelium (i.e. protomap) and intrinsic genetic mechanisms regulating neurogenesis, post-mitotic factors, such as signaling molecules expressed by the subplate or released by the thalamocortical axons, may further refine the gene regulatory networks that give rise to individual cell types (Barber et al., 2015; Callaway and Borrell, 2011; Kraushar et al., 2015; Ozair et al., 2018; Pouchelon et al., 2014). One such example is signaling via Cajal-Retzius cells which may also contribute to cortical arealization by adopting a subtype-specific distribution that influences the identity of local excitatory neurons postmitotically (Barber et al., 2015; Borello and Pierani, 2010). A large body of work indicates a substantial role for thalamic input in promoting cortical arealization. Here we will discuss early events in arealization and refer readers to a number of excellent reviews on the role of thalamic inputs in formation of more refined maps during critical periods of postnatal development (Bednar and Wilson, 2016; Feldman and Brecht, 2005; Inan and Crair, 2007).
The thalamus is a subcortical structure of the diencephalon that receives and modulates sensory input from the periphery, as well as inputs from the cortex, cerebellum, and basal ganglia, and projects predominantly back to the cortex in an area-and modality-specific manner (Haber and Calzavara, 2009; Figure 4A). In the developing mouse, thalamic neurons begin to send out their projections around E12, and the first thalamocortical axons reach the cortical subplate around E14, where they pause for several days before invading the cortical plate to synapse in Layer 4. Axons originating from different thalamic nuclei retain their topographic mapping by differentially responding to axon guidance cues, especially as they traverse the striatum, and segregate to their appropriate target cortical area (Molnár and Blakemore, 1995; Molnar et al., 1998; Bielle et al., 2011a, 2011b; Shimogori and Grove, 2005; Figure 4A–B). Upon arrival at the cortical primordium, thalamocortical axons have been shown to form transient synapses with subplate cells (Molliver et al., 1973), although the exact role of these connections is unknown. During human development, projections from the thalamus are present in the cortical neuroepithelium even before the onset of neurogenesis, suggesting that thalamocortical afferents may play an even more important role in patterning early cortical progenitors (Alzu’bi et al., 2019; Marin-Padilla, 1983). In animal models, two main approaches have been undertaken to investigate the contribution of thalamocortical inputs to areal specification: 1) manipulating the quantitative amount of thalamocortical input while preserving the qualitative type of input or “modality” and 2) rerouting sensory inputs to different thalamic nuclei. These studies have been the subject of recent excellent reviews (Simi and Studer, 2018), and we will discuss here only a handful of notable examples from each of these categories.
Figure 4: Mechanisms of arealization.
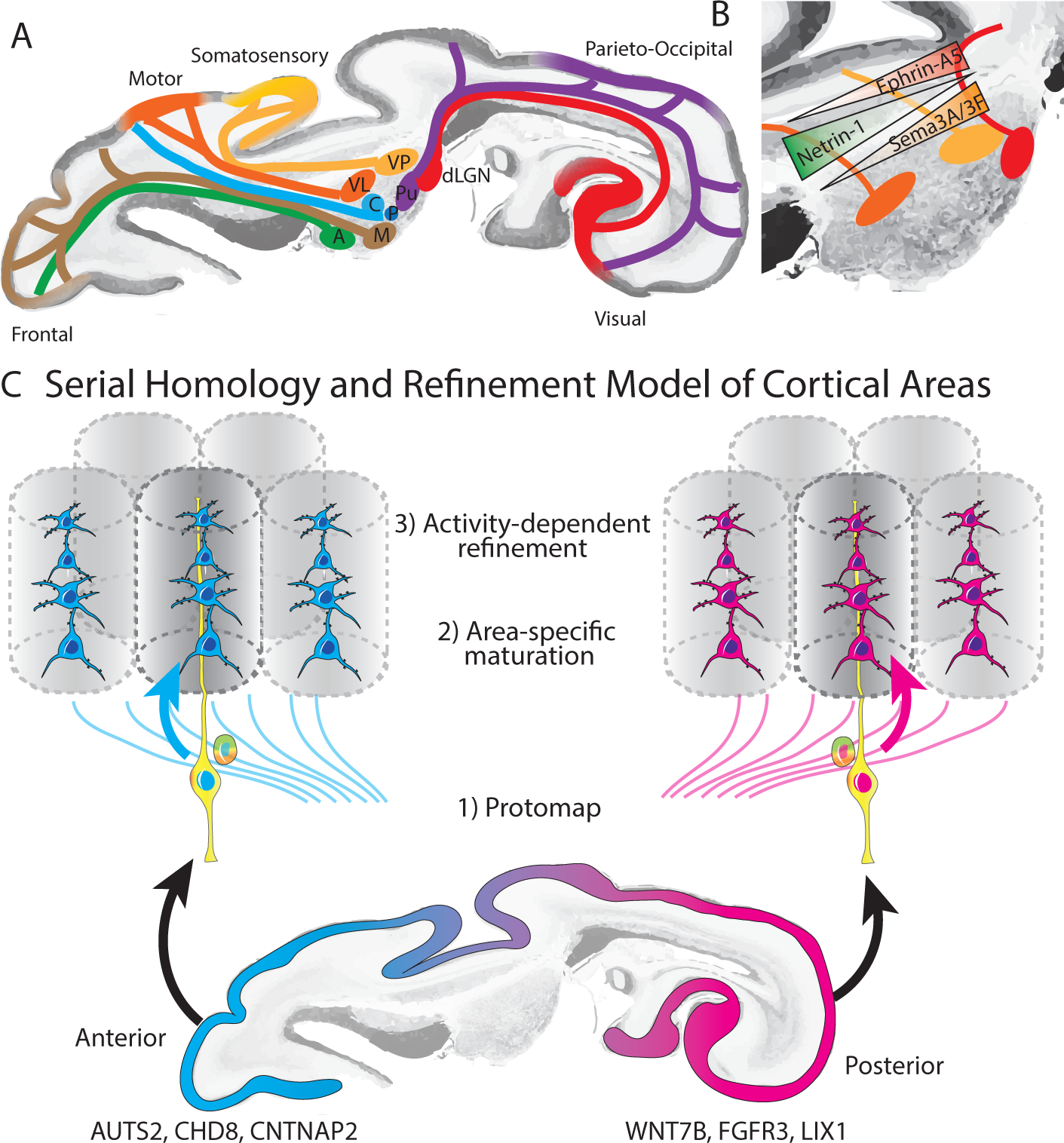
A) Early development of thalamocortical tracts provides anatomical basis for modality specific responses in the cortex, activity dependent changes in cortical area size and functional circuit development. B) Morphogen gradients contribute to shaping thalamic areal specification. C) Serial homology and refinement model, in which area-specific gene expression programs establish an initial “protomap” which is further refined by area-specific maturation signals and activity-dependent processes to generate the final mature cortical areas.
Experimentally increasing or decreasing the size of the dorsal lateral geniculate nucleus (dLGN, the major thalamic nucleus involved in the visual pathway) via genetic manipulation of SHH signaling in mice results in a corresponding change in the size of the primary visual cortex (Vue et al., 2013). Conversely, experimental closure of the eye (‘enucleation’) in ferrets during early postnatal development results in reduced proliferation of OSVZ radial glia, and a reduction of both dLGN and primary visual cortex size (Reillo et al., 2011). Thalamus and cortex explant co-culture studies have demonstrated that thalamocortical axons express FGF and are able to regulate radial glia proliferation (Dehay et al., 2001). Given the previously discussed role of the radial glial scaffold in regulating the tangential surface area of cortex, these findings suggest a possible mechanism whereby thalamocortical input selectively regulates the size of cortical areas. In addition, a recent study found that prenatal thalamocortical calcium waves are required for appropriate barrel organization in mouse somatosensory cortex (Antón-Bolaños et al., 2019), further underscoring the role of extrinsic factors in regulating the development of the cerebal cortex.
Enucleation experiments performed in monkey by removal of the retina during prenatal development (before E80) also resulted in a dramatic reduction of the size of the primary visual area (striate cortex, ‘area 17’) and changes in cortical folding (Dehay et al., 1989; Rakic, 1988), although when performed at later stages of development (after E80), more modest changes in the size of the striate cortex were observed (Dehay et al., 1996a). It has been proposed that depriving dLGN input to the area 17 neurons that would normally receive it would result in the formation of a hybrid ‘area X’ with features intermediate between area 17 and the adjacent extrastriate area 18 (Rakic, 1988, 1991), suggesting a role for both genetically predetermined and area-specific thalamic inputs in development of area 17. However, other studies have found that this region (which they refer to as default extrastriate cortex or DEC instead of ‘area X ‘) is cytoarchitectonically indistinguishable from normal extrastriate area 18 (Dehay et al., 1996b). More recently, molecular tools have enabled targeted silencing of neuronal activity in distinct thalamic nuclei to further probe the role of activity in shaping cortical circuits and arealization. For example, synchronized calcium waves propagating through gap junctions across cells in each of the modality-specific thalamus nuclei are necessary for appropriate specification of cortical area size (Moreno-Juan et al., 2017).
Studies on the role of modality-specific input on cortical development and function have supported the idea that different cortical areas can perform similar fundamental computations. In these experiments, misrouting retinal axons to the main thalamic somatosensory nucleus (Metin and Frost, 1989) or auditory nucleus (Roe et al., 1992; Sur et al., 1988) in newborn animals led to development of orientation selectivity in non-visual areas. Furthermore, rerouting retinal input onto auditory pathways leads to the development of orientation modules and long-range horizontal connectivity patterns reminiscent of those seen in native primary visual cortex (Sharma et al., 2000), suggesting that the pattern of afferent input profoundly alters intracortical connectivity and contributes to cortical area map formation. A recent study showed that following enucleation in the mouse, cortical layer V neurons aberrantly project to first-order dLGN instead of the expected higher-order superior colliculus (Grant et al., 2016), which raises the possibility that sensory experience is also important for the development of appropriate transthalamic cortico-cortical circuits. Moreover, it appears that manipulations of thalamocortical input can alter the terminal specification of neuronal identity. While interrogating the specification of layer IV neurons in the somatosensory cortex, primary S1 neurons were respecified to molecularly and functionally resemble secondary S2 neurons when their thalamic input nucleus was ablated (Pouchelon et al., 2014). Similarly, ablation of geniculocortical afferents prevents the development of patterned gene expression that distinguish higher order visual areas from V1 (Chou et al., 2013). Elimination of all thalamocortical projections does not entirely abolish the development of area-specific gene expression domains (Miyashita-Lin et al., 1999), although the formation of sharp boundaries between cortical areas depends on thalamocortical input (Vue et al., 2013).
Integration of intrinsic and extrinsic inputs
As noted above, projections from the thalamus are present in the cortical neuroepithelium even before the onset of neurogenesis during human development, making it difficult to experimentally tease apart the relative contributions of genetic predisposition of progenitors (i.e. protomap) and thalamocortical inputs (i.e. protocortex) in areal specification. A few studies have investigated the degree of functional interchangeability, or competency, of cells located in different developing brain regions when transplanted to a heterotopic location that receives an alternate source of thalamic input. Cortical transplantation experiments in rodents have shown different results depending on the embryological age of the donor cortex and specific cortical areas examined. When occipital cortex progenitors are harvested at embryonic day 12 (E12) and grafted into the parietal somatosensory cortical area of newborn hosts, they adopt the pattern of reciprocal thalamic connections and cytoarchitectonic organization of the heterotopic area (Gaillard et al., 2003). Even late embryonic visual cortex (E17-E18) progenitors transplanted into neonatal somatosensory cortex develop structures resembling barrel fields (Schlaggar and O’Leary, 1991), although the degree of similarity to native somatosensory cortex in terms of connectivity and function was not examined. In contrast, progenitors harvested from the embryonic occipital cortex at E16 and transplanted into neonatal motor cortex have been shown to retain occipital-like connectivity patterns in line with their site of origin (Ebrahimi-Gaillard et al., 1994). Mouse embryonic stem cell-derived neurons transplanted into newborn mouse frontal cortex tend to follow the projection patterns of visual and limbic areas (Gaspard et al., 2008), and cortical injury models in the adult rodent brain have shown that damaged visual circuits, but not motor circuits, can be reestablished by transplanting stem cell-derived excitatory neurons (Falkner et al., 2016; Michelsen et al., 2015). Together, these results suggest that cortical areal identity, at least between motor and sensory areas, is to some degree prespecified during embryonic development and influences the ability of neurons to integrate into functional circuits. However, because of technical challenges associated with these experiments, many questions remain about the efficiency of integration, the role of maturation stage, and regional variation that may affect the efficiency of heterotopic transplantation.
To reconcile the long standing models of cortical arealization embodied by the protomap and protocortex hypotheses (Van der Loos and Woolsey, 1973; Creutzfeldt, 1977; O’Leary et al., 2007; Shatz, 1992; Woolsey and Van der Loos, 1970), we propose a model of serial homology in which intrinsic and extrinsic factors both contribute to the establishment of area-specific cell types and circuits (Figure 4C). According to this model, intrinsic genetic programs and local secreted factors establish initial “proto-regions,” while activity-dependent mechanisms lead to progressive refinement and the formation of sharp boundaries between functional areas in the mature isocortex. One implication of this model could be that of developmental constructivism. Genetic pre-programming of protoregions may be necessary to maintain region-specific regulatory programs underlying neurogenesis throughout evolution. On the other hand, unlike the determinate development of C. elegans, where developmental lineage history of each cell can be precisely mapped across individuals, mammalian nervous system development is indeterminate, and emerges from many parallel and homologous lineages. One evolutionary advantage of this model is that of size control, because individual neural stem cells can integrate extracellular signals to control the number of cell divisions they undergo, and ultimately the number of neurons they produce. However, this developmental pattern poses new challenges, including that of terminal fate selection. By retaining some degree of fate plasticity, the identity of post-mitotic neurons can be later refined by activity, offering a solution to the challenges associated with indeterminacy of developmental lineage. Further research is needed into the interaction between gene regulatory networks and extrinsic signals to provide more complete understanding of developmental mechanisms that give rise to the molecular differences between homologous subtypes of excitatory neurons, and local cortical circuitry that ultimately supports cortical function.
Development of cortical circuits
The emergence of cognitive capabilities necessitates the formation of appropriate synaptic connections and functional networks during development. Despite significant advances in our understanding of the molecular programs that generate diverse neuronal cell types, the developmental programs that shape neural networks and direct synaptic specificity in the developing cerebral cortex remain largely unknown, but are thought to involve an array of trans-synaptic cell adhesion molecules (Sudhof, 2018) as well as synaptic activity (Kwon and Sabatini, 2011; Oh et al., 2016).
In the macaque, the earliest synapses form in the cortex around E50 in the subplate and the marginal zone, while the first synapses in layer IV of the cortical plate emerge around E65 (Bourgeois and Rakic, 1993; Rakic et al., 1986, 1994). Peak synaptic numbers are reached during early postnatal development (approximately 2 months after birth), followed by elimination of the overproduced synapses (Changeux and Danchin). In the developing human cortex, synapses in the marginal zone are present as early as pcw 5 (Molliver et al., 1973; Zecevic, 1998). While synaptogenesis may occur very early in human development, mature membrane properties are not seen until pcw 14 and cells capable of firing repetitive action potentials are not seen until around pcw 18 (Moore et al., 2009). This maturation of excitatory neurons coincides with the onset of spontaneous activity at pcw 18 (Moore et al., 2011), but precedes the emergence of the earliest thalamocortical synapses in layer IV of the visual cortex, which occurs around pcw 19 (Hevner, 2000). While synaptogenesis and pruning in humans follows a similar trajectory across cortical areas, the total synaptic density peaks at around 3 months after birth in sensory areas, and at around 15 months after birth in prefrontal areas (Huttenlocher and Dabholkar, 1997). The specificity and permanence of these early connections formed among excitatory neurons during development is not well understood.
Connectivity across the prospective areas also emerges early in development and has been most extensively studied in the context of the development of cat visual cortex (Callaway and Katz, 1990, 1991, 1992; Dehay et al., 1984, 1988a; Innocenti and Clarke, 1984; Innocenti et al., 1988; Katz and Callaway, 1992; Price and Blakemore, 1985a). Injections of neuronal tracers revealed many more projections during development than in adult stages, suggesting an initial overproduction of cortico-cortical connections, followed by selective loss of specific connections (Price and Blakemore, 1985a, 1985b). However, studies in macaque have not revealed similar trends (Barone et al., 1995, 1996; Dehay et al., 1988b), suggesting that in primates lasting cortico-cortical connections may develop without forming transient exuberant projections, or that these connections had already been pruned at the time points examined (E97 and later). Additional work is needed to expand our understanding of the molecular mechanisms and developmental timeline of intra- and inter-areal circuit assembly.
Modular organization of cortical circuits
The concept of an elementary computational module of the mammalian isocortex was first introduced by Lorente de No (Lorente de No, 1933, 1949) based on anatomical studies in which he described “vertical cylinders” in the adult cortex, within which he precociously hypothesized “the whole process of transmission of impulses from the afferent fiber to the efferent axon may be accomplished.” These narrow columnar structures seen in adult cortex are now referred to as anatomical minicolumns (Buxhoeveden and Casanova, 2002) and are hypothesized to represent the vestige of ontogenetic columns seen in the developing cortical plate (Rakic, 1972). Radially oriented bundles of dendritic fibers have also been recognized as a repetitive structure in the cerebral cortex (Fleischhauer et al., 1972; Peters and Walsh, 1972) that corresponds to target-specific projections of callosal neurons (Lev and White, 1997). Dendrites in individual bundles have been shown to originate from multiple minicolumns (Peters and Kara, 1987), providing a cytoarchitectonic framework for converging information processed by many neighboring minicolumns (Innocenti and Vercelli, 2010). Functional columnar organization of the cerebral cortex was famously demonstrated by Hubel and Wiesel in the primary visual cortex (Hubel and Wiesel, 1959), and by Mountcastle in the primary somatosensory cortex (Mountcastle, 1957) of cats, where both groups found that columns, or slabs, of cells aligned perpendicularly to the cortical surface have similar feature selectivity for sensory stimuli in the environment (i.e. orientation preference in V1 or somatotopic position in S1). These findings suggest that the partitioning of the cortex into functional columns or modules is a recurrent organizational theme across species and brain regions.
Connectivity between excitatory neurons in adult cortex is highly nonrandom (Yoshimura et al., 2005), suggestive of the presence of underlying modular organization. For example, excitatory neurons in layer 5 tend to form interdigitating subnetworks composed of a few dozen highly interconnected neurons each (Perin et al., 2011). In layer 2/3 of mouse primary visual cortex, where neurons with diverse orientation tuning preferences are arranged in a predominantly mixed salt-and-pepper configuration (Ohki et al., 2005; but see also Ringach et al., 2016), excitatory neurons with similar feature selectivity tend to be preferentially connected (Ko et al., 2011) and have stronger synapses (Cossell et al., 2015) compared to pairs of neurons with orthogonal tuning preferences, consistent with the idea that excitatory neurons are organized into functional subnetworks within the local microcircuit.
It has long been speculated that ontogenetic columns may serve as building blocks for later assembly of functional columns (von Bonin and Mehler, 1971; Buxhoeveden and Casanova, 2002; Mountcastle, 1997). However, the link between ontogenetic and anatomical minicolumns remains speculative, and a precise definition of a universal functional column is also absent, leading many to question the validity of the link between developmental and functional columns (Horton and Adams, 2005; Molnar, 2013). Across primate species there is up to three-fold variation in neuronal density, and this number shows no correlation with total cortical area or total number of neurons (Herculano-Houzel et al., 2008), suggesting that models assuming a constant number of neurons within a given cortical area across primates (Rockel et al., 1980) are largely implausible. Nevertheless, studies in mice have found that vertically aligned pairs of neurons derived from a common progenitor are more likely to be synaptically connected (Cadwell et al., 2019; Yu et al., 2009) and have similar feature selectivity (Li et al., 2012; Ohtsuki et al., 2012) compared to nearby unrelated neurons, although the effect size varies substantially between studies (Smith and Fitzpatrick, 2012). The mechanism by which clonally related neurons form specific synapses is thought to involve transient gap junction coupling during migration (Yu et al., 2012), although other mechanisms such as shared expression of cell adhesion molecules have also been proposed (Tarusawa et al., 2016). While these studies suggest that cell lineage may play a role in establishing a blueprint of functional cortical circuits, larger functional columns such as orientation columns are necessarily comprised of several ontogenetic columns (Rakic, 2008), and ontogenetic columns themselves are likely derived from polyclonal proliferative units (Rakic, 1988). Moreover, it remains unknown whether preferential lineage-dependent connectivity and function persists in adult animals or becomes completely overwritten by experience-dependent plasticity (Blakemore and Cooper, 1970), and whether a similar bias in connectivity and function exists among clonally related neurons in primate cortex has not yet been explored.
Circuit dysfunction in neurodevelopmental disorders
A more detailed understanding how area-specific cell types and circuits emerge may provide novel insights into the pathogenesis of neurodevelopmental disorders characterized by abnormal cortical circuit assembly. Autism spectrum disorder (ASD) is perhaps one of the best studied diseases in this category and highlights both the importance and complexity of relating early arealization to phenotypes that emerge later in life. Syndromic forms of autism are often associated with macrocephaly and epilepsy (Winden et al., 2018). Brain size in idiopathic autism is slightly reduced at birth, increases dramatically during the first year of life, and then plateaus such that in adulthood the majority of patients are within the normal range (Redcay and Courchesne, 2005). Similarly, patients with idiopathic autism have increased numbers of neurons in the prefrontal cortex (Courchesne et al., 2011) and abnormal laminar architecture in prefrontal and temporal cortical areas (Stoner et al., 2014). A limited case series comparing cytoarchitectonic features between autistic and normal brains found that anatomical minicolumns are narrower and more disorganized in autistic brains compared to controls (Buxhoeveden et al., 2006). Noninvasive imaging and recording techniques such as electroencephalography and magnetoencephalography suggest that local connections are increased in ASD, whereas long range connections are reduced (O’Reilly et al., 2017; Wass, 2011). Early studies using transcranial magnetic stimulation suggest that plasticity is also impaired in patients with high-functioning autism and Asperger syndrome (Jung et al., 2013). Overall, these studies suggest that early developmental mechanisms that establish proper area-specific neuronal cell types, lamination patterns, and connectivity may be disrupted in ASD and other neurodevelopmental disorders and play a critical role in the disease pathogenesis.
The genetic instabilities that evolutionarily gave rise to the developmental programs of the primate cerebral cortex may also have built-in vulnerabilities, as noted by the identification of key gene expression networks that decrease in expression over time during macaque brain development (Bakken et al., 2015). One such network enriched for genes implicated in ASD may be differentially expressed when compared to networks in human development (Bakken et al., 2015). Transcriptional comparisons of bulk sequencing gene networks and genes that have been characterized as dysregulated in autism found an enrichment of these networks during the early and peak stages of neurogenesis in subsets of excitatory neurons, specifically in the prefrontal cortex (Willsey et al., 2013). More recently, a comparative study found that the genes implicated in ASD and schizophrenia show unique prenatal expression patterns in humans compared to nonhuman primates (Zhu et al., 2018). These findings raise the hypothesis that area-specific neurons may have selective vulnerability and causal roles in the emergence of neurodevelopmental disorders. Notably, even subtle changes in the shape of the dendritic tree or the expression of certain ion channels in a subset of area-specific excitatory neurons could fundamentally alter the way neurons integrate synaptic inputs (Branco and Hausser, 2011). Mechanisms that give rise to cortical areas in normal development could go awry in neurodevelopmental disorders, and further work is necessary to understand how this happens, what genes are involved, and whether therapeutic interventions might be effective.
Biologically-inspired neural networks
The hierarchical organization of the cortex has inspired in silico models of network architecture. Biologically-inspired networks serve as two-way streets, simultaneously informing both computational and experimental viewpoints (Camacho et al., 2018; Cox and Dean, 2014). Recent advances in machine learning using deep neural networks have revolutionized the artificial intelligence field in applications such as object and voice recognition (reviewed in (LeCun et al., 2015)). Artificial neural networks (ANNs) are vaguely inspired by the hierarchical organization of cortical areas in the mammalian cerebral cortex, specifically the visual processing stream (van Gerven and Bohte, 2017; McCulloch and Pitts, 1990). Most ANNs are initiated using a set of initially random weights between individual nodes, and the weights are modified by backpropagation of an error signal (i.e. “cost function’) in an iterative process known as gradient descent. Backpropagation of error in biological systems was originally thought to be implausible; however recent studies suggest that the brain could implement the error back-propagation algorithm used by artificial neural networks (Marblestone et al., 2016; Whittington and Bogacz, 2019). For example, by simply re-framing the ANN nodes as a set of dendritic events, and incorporating lateral input from local interneurons, a recent study provides a potential mechanism by which neurons could implement back-propagation by increasing or decreasing synaptic strength of individual synapses within the same cell (Sacramento et al., 2018), such as might occur via synaptic tagging (Frey and Morris, 1997). As we continue to develop a better understanding of the biophysical properties of individual cells and cortical microcircuits, additional biologically plausible implementations of error back-propagation and other algorithms used by ANNs may emerge.
The performance of ANNs has recently approached or even surpassed that of humans in some tasks, in particular image recognition (Silver et al., 2016, 2017). ANNs also appear to recapitulate the general organization of functional cortical areas. For example, the layers of deep convolutional neural networks mirror the hierarchical organization of the visual object recognition pathway (Guclu and van Gerven, 2015; Marcus et al., 2014). However, these approaches still fall short of human cognition in their ability to generalize beyond the training data and truly grasp abstract concepts, as highlighted by their failure in the face of adversarial input and the requirement for large amounts of training data (Goodfellow et al., 2014; Kurakin et al., 2016). One promising approach is to integrate unsupervised or generative models, which do not rely on labeled training data, to intuitively learn underlying patterns in the data such as nuisance variables (Patel et al., 2015). Interestingly, recent theoretical work has also shown that incorporation of a modular architecture termed “matrix capsules,” inspired by the modular organization of the cerebral cortex, may significantly improve the accuracy and robustness of deep learning algorithms (Hinton et al., 2018). Large-scale efforts are underway to discover additional principles of information processing in biological brains that can be translated into meaningful advances in machine learning, such as the Machine Intelligence from Cortical Networks project funded by the Intelligence Advanced Research Projects Activity (Cepelewicz, 2016). Besides areal differences in circuit architecture, subtle changes in the implementation of cost functions or error signals could be mediated by neuromodulatory inputs such as acetylcholine or dopamine and could contribute to area-specific computations (Marblestone et al., 2016). An improved understanding of the differences in network architecture between functionally distinct cortical areas, and the developmental programs that give rise to these circuits, may provide additional insights into the core features of biological networks that enable robust and efficient learning.
Concluding remarks
Research spanning several decades has revealed substantial area-specific variations in cellular composition, lamination pattern, and circuit organization of the adult cerebral cortex. Recently, advances in single cell transcriptomics have enabled unbiased comparisons of molecular signatures of homologous cell types across areas and suggest an updated view of serial homology during cortical development in which molecular distinctions across broad “proto-regions” are established early in development, and are further refined into functional areas via activity-dependent mechanisms. So far, these biological processes have largely been explored in only a few major areas, and therefore additional studies are needed to determine what features are present as early in development as neurogenesis, and what activity-dependent changes in cell identity and cytoarchitecture maturation coincide with emergence of specialized local microcircuits. The proposed model of serial homology with progressive refinement provides a more nuanced perspective of cortical arealization and may contribute to a more principled understanding of normal and pathological neural circuits.
Acknowledgem ents
We thank John Rubenstein, Alex Pollen, Ryan Delgado, Andreas Tolias, and Adam Marblestone for careful reading of the manuscript and for providing helpful comments. This work was supported by a gift from the Schmidt Futures and the Bowes Foundation (to T.J.N.). A.B was supported by the National Institutes of Health (NIH), National Institute of Neurological Disorders and Stroke (K99 NS111731). M.A.M.-R. was supported by the NIH, Natinal Center for Advancing Translational Sciences (TL1 TR001871).
Footnotes
Declaration of Interests
The authors declare no competing interests.
References
- Adesnik H, and Naka A (2018). Cracking the Function of Layers in the Sensory Cortex. Neuron 100, 1028–1043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adler A, Zhao R, Shin ME, Yasuda R, and Gan W-B (2019). Somatostatin-Expressing Interneurons Enable and Maintain Learning-Dependent Sequential Activation of Pyramidal Neurons. Neuron 102, 202–216.e7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aevermann BD, Novotny M, Bakken T, Miller JA, Diehl AD, Osumi-Sutherland D, Lasken RS, Lein ES, and Scheuermann RH (2018). Cell type discovery using single-cell transcriptomics: implications for ontological representation. Hum. Mol. Genet 27, R40–R47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alcamo EA, Chirivella L, Dautzenberg M, Dobreva G, Farinas I, Grosschedl R, and McConnell SK (2008). Satb2 regulates callosal projection neuron identity in the developing cerebral cortex. Neuron 57, 364–377. [DOI] [PubMed] [Google Scholar]
- Alzu’bi A, Homman-Ludiye J, Bourne JA, and Clowry GJ (2019). Thalamocortical Afferents Innervate the Cortical Subplate much Earlier in Development in Primate than in Rodent. Cereb. Cortex 29, 1706–1718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Angevine JB, and Sidman RL (1961). Autoradiographic study of cell migration during histogenesis of cerebral cortex in the mouse. Nature 192, 766–768. [DOI] [PubMed] [Google Scholar]
- Antón-Bolaños N, Sempere-Ferràndez A, Guillamón-Vivancos T, Martini FJ, Pérez-Saiz L, Gezelius H, Filipchuk A, Valdeolmillos M, and López-Bendito G (2019). Prenatal activity from thalamic neurons governs the emergence of functional cortical maps in mice. Science eaav7617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arlotta P, Molyneaux BJ, Chen J, Inoue J, Kominami R, and Macklis JD (2005). Neuronal subtype-specific genes that control corticospinal motor neuron development in vivo. Neuron 45, 207–221. [DOI] [PubMed] [Google Scholar]
- Armentano M, Filosa A, Andolfi G, and Studer M (2006). COUP-TFI is required for the formation of commissural projections in the forebrain by regulating axonal growth. Development 133, 4151–4162. [DOI] [PubMed] [Google Scholar]
- Armstrong C, Wang J, Yeun Lee S, Broderick J, Bezaire MJ, Lee SH, and Soltesz I (2016). Target-selectivity of parvalbumin-positive interneurons in layer II of medial entorhinal cortex in normal and epileptic animals. Hippocampus 26, 779–793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnold SJ, Huang G-J, Cheung AFP, Era T, Nishikawa S-I, Bikoff EK, Molnár Z, Robertson EJ, and Groszer M (2008). The T-box transcription factor Eomes/Tbr2 regulates neurogenesis in the cortical subventricular zone. Genes Dev. 22, 2479–2484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Averbeck BB, Battaglia-Mayer A, Guglielmo C, and Caminiti R (2009). Statistical analysis of parieto-frontal cognitive-motor networks. J. Neurophysiol 102, 1911–1920. [DOI] [PubMed] [Google Scholar]
- Azevedo FAC, Carvalho LRB, Grinberg LT, Farfel JM, Ferretti REL, Leite REP, Filho WJ, Lent R, and Herculano-Houzel S (2009). Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. J. Comp. Neurol [DOI] [PubMed] [Google Scholar]
- Badre D, and Nee DE (2018). Frontal Cortex and the Hierarchical Control of Behavior. Trends Cogn. Sci 22, 170–188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bakken TE, Miller JA, Luo R, Bernard A, Bennett JL, Lee C-K, Bertagnolli D, Parikshak NN, Smith KA, Sunkin SM, et al. (2015). Spatiotemporal dynamics of the postnatal developing primate brain transcriptome. Hum. Mol. Genet 24, 4327–4339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bakken TE, Miller JA, Ding S-L, Sunkin SM, Smith KA, Ng L, Szafer A, Dalley RA, Royall JJ, Lemon T, et al. (2016). A comprehensive transcriptional map of primate brain development. Nature 535, 367–375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barber M, Arai Y, Morishita Y, Vigier L, Causeret F, Borello U, Ledonne F, Coppola E, Contremoulins V, Pfrieger FW, et al. (2015). Migration Speed of Cajal-Retzius Cells Modulated by Vesicular Trafficking Controls the Size of Higher-Order Cortical Areas. Curr. Biol 25, 2466–2478. [DOI] [PubMed] [Google Scholar]
- Barone P, Dehay C, Berland M, Bullier J, and Kennedy H (1995). Developmental remodeling of primate visual cortical pathways. Cereb. Cortex 5, 22–38. [DOI] [PubMed] [Google Scholar]
- Barone P, Dehay C, Berland M, and Kennedy H (1996). Role of directed growth and target selection in the formation of cortical pathways: prenatal development of the projection of area V2 to area V4 in the monkey. J. Comp. Neurol 374, 1–20. [DOI] [PubMed] [Google Scholar]
- Barrantes I. del B., Montero-Pedrazuela A, Guadaño-Ferraz A, Obregon M-J, Martinez de Mena R, Gailus-Durner V, Fuchs H, Franz TJ, Kalaydjiev, Klempt M, et al. (2006). Generation and characterization of dickkopf3 mutant mice. Mol. Cell. Biol 26, 2317–2326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayatti N, Sarma S, Shaw C, Eyre JA, Vouyiouklis DA, Lindsay S, and Clowry GJ (2008). Progressive loss of PAX6, TBR2, NEUROD and TBR1 mRNA gradients correlates with translocation of EMX2 to the cortical plate during human cortical development. Eur. J. Neurosci 28, 1449–1456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayer SA, and Altman J (1991). Neocortical Development, Volume 1 (New York: Raven Press; ). [Google Scholar]
- Beattie R, Postiglione MP, Burnett LE, Laukoter S, Streicher C, Pauler FM, Xiao G, Klezovitch O, Vasioukhin V, Ghashghaei TH, et al. (2017). Mosaic Analysis with Double Markers Reveals Distinct Sequential Functions of Lgl1 in Neural Stem Cells. Neuron 94, 517–533.e3. [DOI] [PubMed] [Google Scholar]
- Bednar JA, and Wilson SP (2016). Cortical Maps. Neuroscientist 22, 604–617. [DOI] [PubMed] [Google Scholar]
- Bedogni F, Hodge RD, Nelson BR, Frederick EA, Shiba N, Daza RA, and Hevner RF (2010a). Autism susceptibility candidate 2 (Auts2) encodes a nuclear protein expressed in developing brain regions implicated in autism neuropathology. Gene Expr. Patterns 10, 9–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bedogni F, Hodge RD, Elsen GE, Nelson BR, Daza RAM, Beyer RP, Bammler TK, Rubenstein JLR, and Hevner RF (2010b). Tbr1 regulates regional and laminar identity of postmitotic neurons in developing neocortex. Proc. Natl. Acad. Sci. U. S. A 107, 13129–13134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernier R, Golzio C, Xiong B, Stessman HA, Coe BP, Penn O, Witherspoon K, Gerdts J, Baker C, Vulto-van Silfhout AT, et al. (2014). Disruptive CHD8 mutations define a subtype of autism early in development. Cell 158, 263–276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Betizeau M, Cortay V, Patti D, Pfister S, Gautier E, Bellemin-Menard A, Afanassieff M, Huissoud C, Douglas RJ, Kennedy H, et al. (2013). Precursor Diversity and Complexity of Lineage Relationships in the Outer Subventricular Zone of the Primate. Neuron 80, 442–457. [DOI] [PubMed] [Google Scholar]
- Beul SF, and Hilgetag CC (2015). Towards a “canonical” agranular cortical microcircuit. Front. Neuroanat 8, 165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beutler E, and Kuhl W (1986). Characteristics and significance of the reverse glucose-6-phosphate dehydrogenase reaction. J. Lab. Clin. Med 107, 502–507. [PubMed] [Google Scholar]
- Bielle F, Marcos-Mondéjar P, Leyva-Díaz E, Lokmane L, Mire E, Mailhes C, Keita M, García N, Tessier-Lavigne M, Garel S, et al. (2011a). Emergent growth cone responses to combinations of Slit1 and Netrin 1 in thalamocortical axon topography. Curr. Biol 21, 1748–1755. [DOI] [PubMed] [Google Scholar]
- Bielle F, Marcos-Mondejar P, Keita M, Mailhes C, Verney C, Nguyen Ba-Charvet K, Tessier-Lavigne M, Lopez Bendito G, and Garel S (2011b). Slit2 activity in the migration of guidepost neurons shapes thalamic projections during development and evolution. Neuron 69, 1085–1098. [DOI] [PubMed] [Google Scholar]
- Bishop KM, Goudreau G, and O’Leary DD (2000). Regulation of area identity in the mammalian neocortex by Emx2 and Pax6. Science 288, 344–349. [DOI] [PubMed] [Google Scholar]
- Blakemore C, and Cooper GF (1970). Development of the brain depends on the visual environment. Nature 228, 477–478. [DOI] [PubMed] [Google Scholar]
- von Bohlen und Halbach V, and von Bohlen und Halbach O (2016). Deletion of p75NTR enhances the cholinergic innervation pattern of the visual cortex. Vis. Neurosci 33, E012. [DOI] [PubMed] [Google Scholar]
- Boldog E, Bakken TE, Hodge RD, Novotny M, Aevermann BD, Baka J, Borde S, Close JL, Diez-Fuertes F, Ding S-L, et al. (2018). Transcriptomic and morphophysiological evidence for a specialized human cortical GABAergic cell type. Nat. Neurosci 21, 1185–1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Bonin G, and Mehler WR (1971). On columnar arrangement of nerve cells in cerebral cortex. Brain Res. 27, 1–9. [DOI] [PubMed] [Google Scholar]
- Borello U, and Pierani A (2010). Patterning the cerebral cortex: traveling with morphogens. Curr. Opin. Genet. Dev 20, 408–415. [DOI] [PubMed] [Google Scholar]
- Borrell V (2018). How Cells Fold the Cerebral Cortex. J. Neurosci 38, 776–783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bourgeois JP, and Rakic P (1993). Changes of synaptic density in the primary visual cortex of the macaque monkey from fetal to adult stage. J. Neurosci 13, 2801–2820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Branco T, and Häusser M (2011). Synaptic integration gradients in single cortical pyramidal cell dendrites. Neuron 69, 885–892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bribián A, Nocentini S, Llorens F, Gil V, Mire E, Reginensi D, Yoshida Y, Mann F, and del Río JA (2014). Sema3E/PlexinD1 regulates the migration of hem-derived Cajal-Retzius cells in developing cerebral cortex. Nat. Commun 5, 4265. [DOI] [PubMed] [Google Scholar]
- Britanova O, Alifragis P, Junek S, Jones K, Gruss P, and Tarabykin V (2006). A novel mode of tangential migration of cortical projection neurons. Dev. Biol 298, 299–311. [DOI] [PubMed] [Google Scholar]
- Britanova O, de Juan Romero C, Cheung A, Kwan KY, Schwark M, Gyorgy A, Vogel T, Akopov S, Mitkovski M, Agoston D, et al. (2008). Satb2 is a postmitotic determinant for upper-layer neuron specification in the neocortex. Neuron 57, 378–392. [DOI] [PubMed] [Google Scholar]
- Brodmann K (1909). Vergleichende Lokalisationslehre der Grosshirnrinde. J. Nerv. Ment. Dis [Google Scholar]
- Brown KN, Chen S, Han Z, Lu C-H, Tan X, Zhang X-J, Ding L, Lopez-Cruz A, Saur D, Anderson SA, et al. (2011). Clonal Production and Organization of Inhibitory Interneurons in the Neocortex. Science (80-.) 334, 480–486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bulchand S, Subramanian L, and Tole S (2003). Dynamic spatiotemporal expression of LIM genes and cofactors in the embryonic and postnatal cerebral cortex. Dev. Dyn 226, 460–469. [DOI] [PubMed] [Google Scholar]
- Bulfone A, Smiga SM, Shimamura K, Peterson A, Puelles L, and Rubenstein JL (1995). T-brain-1: a homolog of Brachyury whose expression defines molecularly distinct domains within the cerebral cortex. Neuron 15, 63–78. [DOI] [PubMed] [Google Scholar]
- Buxhoeveden DP, and Casanova MF (2002). The minicolumn hypothesis in neuroscience. Brain 125, 935–951. [DOI] [PubMed] [Google Scholar]
- Buxhoeveden DP, Semendeferi K, Buckwalter J, Schenker N, Switzer R, and Courchesne E (2006). Reduced minicolumns in the frontal cortex of patients with autism. Neuropathol. Appl. Neurobiol 32, 483–491. [DOI] [PubMed] [Google Scholar]
- Cadwell CR, Palasantza A, Jiang X, Berens P, Deng Q, Yilmaz M, Reimer J, Shen S, Bethge M, Tolias KF, et al. (2016). Electrophysiological, transcriptomic and morphologic profiling of single neurons using Patch-seq. Nat. Biotechnol [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cadwell CR, Scala F, Fahey PG, Kobak D, Sinz FH, Johnsson P, Li S, Cotton RJ, Sandberg R, Berens P, et al. (2019). Cell type composition and circuit organization of neocortical radial clones. BioRxiv 526681. [Google Scholar]
- Callaway EM, and Borrell V (2011). Developmental Sculpting of Dendritic Morphology of Layer 4 Neurons in Visual Cortex: Influence of Retinal Input. J. Neurosci 31, 7456–7470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callaway EM, and Katz LC (1990). Emergence and refinement of clustered horizontal connections in cat striate cortex. J. Neurosci 10, 1134–1153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callaway EM, and Katz LC (1991). Effects of binocular deprivation on the development of clustered horizontal connections in cat striate cortex. Proc. Natl. Acad. Sci. U. S. A 88, 745–749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callaway EM, and Katz LC (1992). Development of axonal arbors of layer 4 spiny neurons in cat striate cortex. J. Neurosci 12, 570–582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Camacho DM, Collins KM, Powers RK, Costello JC, and Collins JJ (2018). Next-Generation Machine Learning for Biological Networks. Cell 173, 1581–1592. [DOI] [PubMed] [Google Scholar]
- Caviness VS, Takahashi T, and Nowakowski RS (1995). Numbers, time and neocortical neuronogenesis: a general developmental and evolutionary model. Trends Neurosci. 18, 379–383. [DOI] [PubMed] [Google Scholar]
- Cederquist GY, Azim E, Shnider SJ, Padmanabhan H, and Macklis JD (2013). Lmo4 establishes rostral motor cortex projection neuron subtype diversity. J. Neurosci 33, 6321–6332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cepelewicz J (2016). The U.S. Government Launches a $100-Million “Apollo Project of the Brain.” Sci. Am [Google Scholar]
- Changeux JP, and Danchin A Selective stabilisation of developing synapses as a mechanism for the specification of neuronal networks. Nature 264, 705–712. [DOI] [PubMed] [Google Scholar]
- Charvet CJ, Cahalane DJ, and Finlay BL (2015). Systematic, cross-cortex variation in neuron numbers in rodents and primates. Cereb. Cortex 25, 147–160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chenn A, and Walsh CA (2002). Regulation of Cerebral Cortical Size by Control of Cell Cycle Exit in Neural Precursors. Science (80-.) 297, 365–369. [DOI] [PubMed] [Google Scholar]
- Chenn A, and Walsh CA (2003). Increased neuronal production, enlarged forebrains and cytoarchitectural distortions in beta-catenin overexpressing transgenic mice. Cereb. Cortex 13, 599–606. [DOI] [PubMed] [Google Scholar]
- Cholfin JA, and Rubenstein JLR (2007). Patterning of frontal cortex subdivisions by Fgf17. Proc. Natl. Acad. Sci. U. S. A 104, 7652–7657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou S-J, Perez-Garcia CG, Kroll TT, and O’Leary DDM (2009). Lhx2 specifies regional fate in Emxl lineage of telencephalic progenitors generating cerebral cortex. Nat. Neurosci 12, 1381–1389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou S-J, Babot Z, Leingartner A, Studer M, Nakagawa Y, and O’Leary DDM (2013). Geniculocortical Input Drives Genetic Distinctions Between Primary and Higher-Order Visual Areas. Science (80-.) 340, 1239–1242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chowdhury TG, Jimenez JC, Bomar JM, Cruz-Martin A, Cantle JP, and Portera-Cailliau C (2010). Fate of Cajal-Retzius neurons in the postnatal mouse neocortex. Front. Neuroanat 4, 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clancy B, Darlington RB, and Finlay BL (2001). Translating developmental time across mammalian species. Neuroscience, 105, 7–17. [DOI] [PubMed] [Google Scholar]
- Clifford MA, Athar W, Leonard CE, Russo A, Sampognaro PJ, Van der Goes M-S, Burton DA, Zhao X, Lalchandani RR, Sahin M, et al. (2014). EphA7 signaling guides cortical dendritic development and spine maturation. Proc. Natl. Acad. Sci. U. S. A 111, 4994–4999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper MA, Crockett DP, Nowakowski RS, Gale NW, and Zhou R (2009). Distribution of EphA5 receptor protein in the developing and adult mouse nervous system. J. Comp. Neurol 514, 310–328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cossell L, Iacaruso MF, Muir DR, Houlton R, Sader EN, Ko H, Hofer SB, and Mrsic-Flogel TD (2015). Functional organization of excitatory synaptic strength in primary visual cortex. Nature 518, 399–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Courchesne E, Mouton PR, Calhoun ME, Semendeferi K, Ahrens-Barbeau C, Hallet MJ, Barnes CC, and Pierce K (2011). Neuron number and size in prefrontal cortex of children with autism. JAMA 306, 2001–2010. [DOI] [PubMed] [Google Scholar]
- Cox DD, and Dean T (2014). Neural Networks and Neuroscience-Inspired Computer Vision. Curr. Biol 24, R921–R929. [DOI] [PubMed] [Google Scholar]
- Creutzfeldt OD (1977). Generality of the functional structure of the neocortex. Naturwissenschaften 64, 507–517. [DOI] [PubMed] [Google Scholar]
- Cubelos B, Briz CG, Esteban-Ortega GM, and Nieto M (2015). Cux1 and Cux2 selectively target basal and apical dendritic compartments of layer II-III cortical neurons. Dev. Neurobiol 75, 163–172. [DOI] [PubMed] [Google Scholar]
- Das G, Yu Q, Hui R, Reuhl K, Gale NW, and Zhou R (2016). EphA5 and EphA6: regulation of neuronal and spine morphology. Cell Biosci. 6, 48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis MF, Figueroa Velez DX, Guevarra RP, Yang MC, Habeeb M, Carathedathu MC, and Gandhi SP (2015). Inhibitory Neuron Transplantation into Adult Visual Cortex Creates a New Critical Period that Rescues Impaired Vision. Neuron 86, 1055–1066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- deAzevedo LC, Fallet C, Moura-Neto V, Daumas-Duport C, Hedin-Pereira C, and Lent R (2003). Cortical radial glial cells in human fetuses: depth-correlated transformation into astrocytes. J. Neurobiol 55, 288–298. [DOI] [PubMed] [Google Scholar]
- Defelipe J (2011). The evolution of the brain, the human nature of cortical circuits, and intellectual creativity. Front. Neuroanat 5, 29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Defelipe J, González-Albo MC, Del Río MR, and Elston GN (1999). Distribution and patterns of connectivity of interneurons containing calbindin, calretinin, and parvalbumin in visual areas of the occipital and temporal lobes of the macaque monkey. J. Comp. Neurol 412, 515–526. [DOI] [PubMed] [Google Scholar]
- DeFelipe J, Alonso-Nanclares L, and Arellano JI (2002). Microstructure of the neocortex: comparative aspects. J. Neurocytol 31, 299–316. [DOI] [PubMed] [Google Scholar]
- Dehay C, Bullier J, and Kennedy H (1984). Transient projections from the fronto-parietal and temporal cortex to areas 17, 18 and 19 in the kitten. Exp. Brain Res 57, 208–212. [DOI] [PubMed] [Google Scholar]
- Dehay C, Kennedy H, and Bullier J (1988a). Characterization of transient cortical projections from auditory, somatosensory, and motor cortices to visual areas 17, 18, and 19 in the kitten. J. Comp. Neurol 272, 68–89. [DOI] [PubMed] [Google Scholar]
- Dehay C, Kennedy H, Bullier J, and Berland M (1988b). Absence of interhemispheric connections of area 17 during development in the monkey. Nature 331, 348–350. [DOI] [PubMed] [Google Scholar]
- Dehay C, Horsburgh G, Berland M, Killackey H, and Kennedy H (1989). Maturation and connectivity of the visual cortex in monkey is altered by prenatal removal of retinal input. Nature 337, 265–267. [DOI] [PubMed] [Google Scholar]
- Dehay C, Giroud P, Berland M, Killackey HP, and Kennedy H (1996a). Phenotypic characterisation of respecified visual cortex subsequent to prenatal enucleation in the monkey: Development of acetylcholinesterase and cytochrome oxidase patterns. J. Comp. Neurol 376, 386–402. [DOI] [PubMed] [Google Scholar]
- Dehay C, Giroud P, Berland M, Killackey H, and Kennedy H (1996b). Contribution of thalamic input to the specification of cytoarchitectonic cortical fields in the primate: effects of bilateral enucleation in the fetal monkey on the boundaries, dimensions, and gyrification of striate and extrastriate cortex. J. Comp. Neurol 367, 70–89. [DOI] [PubMed] [Google Scholar]
- Dehay C, Savatier P, Cortay V, and Kennedy H (2001). Cell-cycle kinetics of neocortical precursors are influenced by embryonic thalamic axons. J. Neurosci 21, 201–214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding SL, Royall JJ, Sunkin SM, Ng L, Facer BAC, Lesnar P, Guillozet-Bongaarts A, McMurray B, Szafer A, Dolbeare TA, et al. (2016). Comprehensive cellular-resolution atlas of the adult human brain. J. Comp. Neurol [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donoghue MJ, and Rakic P (1999). Molecular gradients and compartments in the embryonic primate cerebral cortex. Cereb. Cortex 9, 586–600. [DOI] [PubMed] [Google Scholar]
- Duque A, Krsnik Z, Kostović I, and Rakic P (2016). Secondary expansion of the transient subplate zone in the developing cerebrum of human and nonhuman primates. Proc. Natl. Acad. Sci. U. S. A 113, 9892–9897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebrahimi-Gaillard A, Guitet J, Garnier C, and Roger M (1994). Topographic distribution of efferent fibers originating from homotopic or heterotopic transplants: heterotopically transplanted neurons retain some of the developmental characteristics corresponding to their site of origin. Brain Res. Dev. Brain Res 77, 271–283. [DOI] [PubMed] [Google Scholar]
- Economo MN, Viswanathan S, Tasic B, Bas E, Winnubst J, Menon V, Graybuck LT, Nguyen TN, Smith KA, Yao Z, et al. (2018). Distinct descending motor cortex pathways and their roles in movement. Nature 563, 79–84. [DOI] [PubMed] [Google Scholar]
- von Economo CF, and Koskinas GN (1925). Die cytoarchitektonik der hirnrinde des erwachsenen menschen (Wien and Berlin: J. Springer; ). [Google Scholar]
- Egger V, Feldmeyer D, and Sakmann B (1999). Coincidence detection and changes of synaptic efficacy in spiny stellate neurons in rat barrel cortex. Nat. Neurosci 2, 1098–1105. [DOI] [PubMed] [Google Scholar]
- Elsen GE, Hodge RD, Bedogni F, Daza RAM, Nelson BR, Shiba N, Reiner SL, and Hevner RF (2013). The protomap is propagated to cortical plate neurons through an Eomes-dependent intermediate map. Proc. Natl. Acad. Sci. U. S. A 110, 4081–4086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elston GN (2003). Cortex, Cognition and the Cell: New Insights into the Pyramidal Neuron and Prefrontal Function. Cereb. Cortex [DOI] [PubMed] [Google Scholar]
- Falkner S, Grade S, Dimou L, Conzelmann K-K, Bonhoeffer T, Gotz M, and Hubener M (2016). Transplanted embryonic neurons integrate into adult neocortical circuits. Nature 539, 248–253. [DOI] [PubMed] [Google Scholar]
- Faux C, Rakic S, Andrews W, and Britto JM (2012). Neurons on the Move: Migration and Lamination of Cortical Interneurons. Neurosignals 20, 168–189. [DOI] [PubMed] [Google Scholar]
- Feldman DE, and Brecht M (2005). Map plasticity in somatosensory cortex. Science 310, 810–815. [DOI] [PubMed] [Google Scholar]
- Felleman DJ, and Van Essen DC (1991). Distributed hierarchical processing in the primate cerebral cortex. Cereb. Cortex 1, 1–47. [DOI] [PubMed] [Google Scholar]
- Fietz SA, Kelava I, Vogt J, Wilsch-Brauninger M, Stenzel D, Fish JL, Corbeil D, Riehn A, Distler W, Nitsch R, et al. (2010). OSVZ progenitors of human and ferret neocortex are epithelial-like and expand by integrin signaling. Nat. Neurosci 13, 690–699. [DOI] [PubMed] [Google Scholar]
- Fino E, and Yuste R (2011). Dense inhibitory connectivity in neocortex. Neuron 69, 1188–1203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fleischhauer K, Petsche H, and Wittkowski W (1972). Vertical bundles of dendrites in the neocortex. Z. Anat. Entwicklungsgesch 136, 213–223. [DOI] [PubMed] [Google Scholar]
- Frey U, and Morris RG (1997). Synaptic tagging and long-term potentiation. Nature 385, 533–536. [DOI] [PubMed] [Google Scholar]
- Friedman LG, Riemslagh FW, Sullivan JM, Mesias R, Williams FM, Huntley GW, and Benson DL (2015). Cadherin-8 expression, synaptic localization, and molecular control of neuronal form in prefrontal corticostriatal circuits. J. Comp. Neurol 523, 75–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuzik J, Zeisel A, Máté Z, Calvigioni D, Yanagawa Y, Szabó G, Linnarsson S, and Harkany T (2016). Integration of electrophysiological recordings with single-cell RNA-seq data identifies neuronal subtypes. Nat. Biotechnol 34, 175–183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaillard A, Nasarre C, and Roger M (2003). Early (E12) cortical progenitors can change their fate upon heterotopic transplantation. Eur. J. Neurosci 17, 1375–1383. [DOI] [PubMed] [Google Scholar]
- Gaspard N, Bouschet T, Hourez R, Dimidschstein J, Naeije G, van den Ameele J, Espuny-Camacho I, Herpoel A, Passante L, Schiffmann SN, et al. (2008). An intrinsic mechanism of corticogenesis from embryonic stem cells. Nature 455, 351–357. [DOI] [PubMed] [Google Scholar]
- Gertz CC, and Kriegstein AR (2015). Neuronal Migration Dynamics in the Developing Ferret Cortex. J. Neurosci 35, 14307–14315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Gerven M, and Bohte S (2017). Editorial: Artificial Neural Networks as Models of Neural Information Processing. Front. Comput. Neurosci 11, 114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geschwind DH, and Rakic P (2013). Cortical Evolution: Judge the Brain by Its Cover. Neuron 80, 633–647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibon J, Buckley SM, Unsain N, Kaartinen V, Seguela P, and Barker PA (2015). proBDNF and p75NTR Control Excitability and Persistent Firing of Cortical Pyramidal Neurons. J. Neurosci 35, 9741–9753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodfellow IJ, Shlens J, and Szegedy C (2014). Explaining and Harnessing Adversarial Examples.
- Gouwens NW, Sorensen SA, Berg J, Lee C, Jarsky T, Ting J, Sunkin SM, Feng D, Anastassiou C, Barkan E, et al. (2018). Classification of electrophysiological and morphological types in mouse visual cortex. BioRxiv 368456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grant E, Hoerder-Suabedissen A, and Molnar Z (2016). The Regulation of Corticofugal Fiber Targeting by Retinal Inputs. Cereb. Cortex 26, 1336–1348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray LT, Yao Z, Nguyen TN, Kim TK, Zeng H, and Tasic B (2017). Layer-specific chromatin accessibility landscapes reveal regulatory networks in adult mouse visual cortex. Elife 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graybuck LT, Sedeño-Cortés A, Nguyen TN, Walker M, Szelenyi E, Lenz G, Sieverts L, Kim TK, Garren E, Kalmbach B, et al. (2019). Prospective, brain-wide labeling of neuronal subclasses with enhancer-driven AAVs. BioRxiv 525014. [Google Scholar]
- Greig LC, Woodworth MB, Galazo MJ, Padmanabhan H, and Macklis JD (2013). Molecular logic of neocortical projection neuron specification, development and diversity. Nat. Rev. Neurosci 14, 755–769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griveau A, Borello U, Causeret F, Tissir F, Boggetto N, Karaz S, and Pierani A (2010). A novel role for Dbx1- derived Cajal-Retzius cells in early regionalization of the cerebral cortical neuroepithelium. PLoS Biol. 8, e1000440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guclu U, and van Gerven MAJ (2015). Deep Neural Networks Reveal a Gradient in the Complexity of Neural Representations across the Ventral Stream. J. Neurosci 35, 10005–10014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gulisano M, Broccoli V, Pardini C, and Boncinelli E (1996). Emx1 and Emx2 show different patterns of expression during proliferation and differentiation of the developing cerebral cortex in the mouse. Eur. J. Neurosci 8, 1037–1050. [DOI] [PubMed] [Google Scholar]
- Haber SN, and Calzavara R (2009). The cortico-basal ganglia integrative network: the role of the thalamus. Brain Res. Bull 78, 69–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansen DV, Lui JH, Parker PRL, and Kriegstein AR (2010). Neurogenic radial glia in the outer subventricular zone of human neocortex. Nature 464, 554–561. [DOI] [PubMed] [Google Scholar]
- Harris KD, and Shepherd GMG (2015). The neocortical circuit: themes and variations. Nat. Neurosci 18, 170–181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harwell CC, Fuentealba LC, Gonzalez-Cerrillo A, Parker PRL, Gertz CC, Mazzola E, Garcia MT, Alvarez-Buylla A, Cepko CL, and Kriegstein AR (2015). Wide Dispersion and Diversity of Clonally Related Inhibitory Interneurons. Neuron 87, 999–1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haubensak W, Attardo A, Denk W, and Huttner WB (2004). Neurons arise in the basal neuroepithelium of the early mammalian telencephalon: a major site of neurogenesis. Proc. Natl. Acad. Sci. U. S. A 101, 3196–3201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawrylycz M, Miller JA, Menon V, Feng D, Dolbeare T, Guillozet-Bongaarts AL, Jegga AG, Aronow BJ, Lee C-K, Bernard A, et al. (2015). Canonical genetic signatures of the adult human brain. Nat. Neurosci 18, 1832–1844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawrylycz MJ, Lein ES, Guillozet-Bongaarts AL, Shen EH, Ng L, Miller JA, van de Lagemaat LN, Smith KA, Ebbert A, Riley ZL, et al. (2012). An anatomically comprehensive atlas of the adult human brain transcriptome. Nature 489, 391–399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haydar TF, Kuan CY, Flavell RA, and Rakic P (1999). The Role of Cell Death in Regulating the Size and Shape of the Mammalian Forebrain. Cereb. Cortex 9, 621–626. [DOI] [PubMed] [Google Scholar]
- Hendry SH, Schwark HD, Jones EG, and Yan J (1987). Numbers and proportions of GABA-immunoreactive neurons in different areas of monkey cerebral cortex. J. Neurosci 7, 1503–1519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herculano-Houzel S, Collins CE, Wong P, Kaas JH, and Lent R (2008). The basic nonuniformity of the cerebral cortex. Proc. Natl. Acad. Sci. U. S. A 105, 12593–12598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hevner RF (2000). Development of connections in the human visual system during fetal mid-gestation: a DiI-tracing study. J. Neuropathol. Exp. Neurol 59, 385–392. [DOI] [PubMed] [Google Scholar]
- Hevner RF, Neogi T, Englund C, Daza RAM, and Fink A (2003). Cajal-Retzius cells in the mouse: transcription factors, neurotransmitters, and birthdays suggest a pallial origin. Brain Res. Dev. Brain Res 141, 39–53. [DOI] [PubMed] [Google Scholar]
- Hinton GE, Sabour S, and Frosst N (2018). Matrix capsules with EM routing. In International Conference on Learning Representations, p. [Google Scholar]
- Hodge RD, Bakken TE, Miller JA, Smith KA, Barkan ER, Graybuck LT, Close JL, Long B, Penn O, Yao Z, et al. (2018). Conserved cell types with divergent features between human and mouse cortex. BioRxiv 384826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoerder-Suabedissen A and Molnar Z (2013). Molecular diversity of early-born subplate neurons. Cereb Cortex 23, 1473–1483. [DOI] [PubMed] [Google Scholar]
- Hof PR, and Nimchinsky EA (1992). Regional distribution of neurofilament and calcium-binding proteins in the cingulate cortex of the macaque monkey. Cereb. Cortex 2, 456–467. [DOI] [PubMed] [Google Scholar]
- Hori K, Nagai T, Shan W, Sakamoto A, Taya S, Hashimoto R, Hayashi T, Abe M, Yamazaki M, Nakao K, et al. (2014). Cytoskeletal regulation by AUTS2 in neuronal migration and neuritogenesis. Cell Rep. 9, 2166–2179. [DOI] [PubMed] [Google Scholar]
- Horton JC, and Adams DL (2005). The cortical column: a structure without a function. Philos. Trans. R. Soc. Lond. B. Biol. Sci 360, 837–862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hrvatin S, Tzeng CP, Nagy MA, Stroud H, Koutsioumpa C, Wilcox OF, Griffith EC, and Greenberg ME (2019). PESCA: A scalable platform for the development of cell-type-specific viral drivers. BioRxiv 570895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hubel DH, and Wiesel TN (1959). Receptive fields of single neurones in the cat’s striate cortex. J. Physiol 148, 574–591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huttenlocher PR, and Dabholkar AS (1997). Regional differences in synaptogenesis in human cerebral cortex. J. Comp. Neurol 387, 167–178. [DOI] [PubMed] [Google Scholar]
- Inan M, and Crair MC (2007). Development of cortical maps: perspectives from the barrel cortex. Neuroscientist 13, 49–61. [DOI] [PubMed] [Google Scholar]
- Innocenti GM, and Clarke S (1984). The organization of immature callosal connections. J. Comp. Neurol 230, 287–309. [DOI] [PubMed] [Google Scholar]
- Innocenti GM, and Vercelli A (2010). Dendritic bundles, minicolumns, columns, and cortical output units. Front. Neuroanat 4, 11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Innocenti GM, Berbel P, and Clarke S (1988). Development of projections from auditory to visual areas in the cat. J. Comp. Neurol 272, 242–259. [DOI] [PubMed] [Google Scholar]
- Ip BK, Wappler I, Peters H, Lindsay S, Clowry GJ, and Bayatti N (2010). Investigating gradients of gene expression involved in early human cortical development. J. Anat 217, 300–311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang X, Shen S, Cadwell CR, Berens P, Sinz F, Ecker AS, Patel S, and Tolias AS (2015). Principles of connectivity among morphologically defined cell types in adult neocortex. Science (80-.) 350, aac9462–aac9462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson MB, Kawasawa YI, Mason CE, Krsnik Z, Coppola G, Bogdanovic D, Geschwind DH, Mane SM, State MW, and Sestan N (2009). Functional and evolutionary insights into human brain development through global transcriptome analysis. Neuron 62, 494–509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joshi PS, Molyneaux BJ, Feng L, Xie X, Macklis JD, and Gan L (2008). Bhlhb5 regulates the postmitotic acquisition of area identities in layers II-V of the developing neocortex. Neuron 60, 258–272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Juan Romero C, Bruder C, Tomasello U, Sanz-Anquela JM, and Borrell V (2015). Discrete domains of gene expression in germinal layers distinguish the development of gyrencephaly. EMBO J. 34, 1859–1874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jung NH, Janzarik WG, Delvendahl I, Munchau A, Biscaldi M, Mainberger F, Baumer T, Rauh R, and Mall V (2013). Impaired induction of long-term potentiation-like plasticity in patients with high-functioning autism and Asperger syndrome. Dev. Med. Child Neurol 55, 83–89. [DOI] [PubMed] [Google Scholar]
- Kang HJ, Kawasawa YI, Cheng F, Zhu Y, Xu X, Li M, Sousa AMM, Pletikos M, Meyer KA, Sedmak G, et al. (2011). Spatio-temporal transcriptome of the human brain. Nature 478, 483–489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katz LC, and Callaway EM (1992). Development of local circuits in mammalian visual cortex. Annu. Rev. Neurosci 15, 31–56. [DOI] [PubMed] [Google Scholar]
- Kelley KW, and Oldham MC (2015). Transcriptional architecture of the human brain. Nat. Neurosci 18, 1699–1701. [DOI] [PubMed] [Google Scholar]
- Khaitovich P, Muetzel B, She X, Lachmann M, Hellmann I, Dietzsch J, Steigele S, Do H-H, Weiss G, Enard W, et al. (2004). Regional patterns of gene expression in human and chimpanzee brains. Genome Res. 14, 1462–1473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ko H, Hofer SB, Pichler B, Buchanan KA, Sjostrom PJ, and Mrsic-Flogel TD (2011). Functional specificity of local synaptic connections in neocortical networks. Nature 473, 87–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kostovic I, and Rakic P (1980). Cytology and time of origin of interstitial neurons in the white matter in infant and adult human and monkey telencephalon. J. Neurocytol 9, 219–242. [DOI] [PubMed] [Google Scholar]
- Kraushar ML, Viljetic B, Wijeratne HRS, Thompson K, Jiao X, Pike JW, Medvedeva V, Groszer M, Kiledjian M, Hart RP, et al. (2015). Thalamic WNT3 Secretion Spatiotemporally Regulates the Neocortical Ribosome Signature and mRNA Translation to Specify Neocortical Cell Subtypes. J. Neurosci 35, 10911–10926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kriegstein AR, and Noctor SC (2004). Patterns of neuronal migration in the embryonic cortex. Trends Neurosci. 27, 392–399. [DOI] [PubMed] [Google Scholar]
- Kriegstein A, Noctor S, and Martinez-Cerdeno V (2006). Patterns of neural stem and progenitor cell division may underlie evolutionary cortical expansion. Nat. Rev. Neurosci 7, 883–890. [DOI] [PubMed] [Google Scholar]
- Kruger M, and Braun T (2002). The Neuronal Basic Helix-Loop-Helix Transcription Factor NSCL-1 Is Dispensable for Normal Neuronal Development. Mol. Cell. Biol 22, 792–800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuida K, Zheng TS, Na S, Kuan C, Yang D, Karasuyama H, Rakic P, and Flavell RA (1996). Decreased apoptosis in the brain and premature lethality in CPP32-deficient mice. Nature 384, 368–372. [DOI] [PubMed] [Google Scholar]
- Kunche S, Yan H, Calof AL, Lowengrub JS, and Lander AD (2016). Feedback, Lineages and Self-Organizing Morphogenesis. PLoS Comput. Biol 12, e1004814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurakin A, Goodfellow I, and Bengio S (2016). Adversarial examples in the physical world.
- Kwon H-B, and Sabatini BL (2011). Glutamate induces de novo growth of functional spines in developing cortex. Nature 474, 100–104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lagler M, Ozdemir AT, Lagoun S, Malagon-Vina H, Borhegyi Z, Hauer R, Jelem A, and Klausberger T (2016). Divisions of Identified Parvalbumin-Expressing Basket Cells during Working Memory-Guided Decision Making. Neuron 91, 1390–1401. [DOI] [PubMed] [Google Scholar]
- Lake BB, Chen S, Sos BC, Fan J, Kaeser GE, Yung YC, Duong TE, Gao D, Chun J, Kharchenko PV, et al. (2018). Integrative single-cell analysis of transcriptional and epigenetic states in the human adult brain. Nat. Biotechnol 36, 70–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lambert N, Lambot M-A, Bilheu A, Albert V, Englert Y, Libert F, Noel J-C, Sotiriou C, Holloway AK, Pollard KS, et al. (2011). Genes expressed in specific areas of the human fetal cerebral cortex display distinct patterns of evolution. PLoS One 6, e17753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LaMonica BE, Lui JH, Hansen DV, and Kriegstein AR (2013). Mitotic spindle orientation predicts outer radial glial cell generation in human neocortex. Nat. Commun 4, 1665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leamey CA, Glendining KA, Kreiman G, Kang N-D, Wang KH, Fassler R, Sawatari A, Tonegawa S, and Sur M (2008). Differential gene expression between sensory neocortical areas: potential roles for Ten_m3 and Bcl6 in patterning visual and somatosensory pathways. Cereb. Cortex 18, 53–66. [DOI] [PubMed] [Google Scholar]
- LeCun Y, Bengio Y, and Hinton G (2015). Deep learning. Nature 521, 436–444. [DOI] [PubMed] [Google Scholar]
- Lee AT, Gee SM, Vogt D, Patel T, Rubenstein JL, and Sohal VS (2014). Pyramidal neurons in prefrontal cortex receive subtype-specific forms of excitation and inhibition. Neuron 81, 61–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lein ES, Belgard TG, Hawrylycz M, and Molnar Z (2017). Transcriptomic perspectives on neocortical structure, development, evolution, and disease. Annu Rev Neurosci 40, 629–652. [DOI] [PubMed] [Google Scholar]
- Lev DL, and White EL (1997). Organization of pyramidal cell apical dendrites and composition of dendritic clusters in the mouse: emphasis on primary motor cortex. Eur. J. Neurosci 9, 280–290. [DOI] [PubMed] [Google Scholar]
- Li H, Bishop KM, and O’Leary DDM (2006). Potential target genes of EMX2 include Odz/Ten-M and other gene families with implications for cortical patterning. Mol. Cell. Neurosci 33, 136–149. [DOI] [PubMed] [Google Scholar]
- Li M, Santpere G, Imamura Kawasawa Y, Evgrafov OV, Gulden FO, Pochareddy S, Sunkin SM, Li Z, Shin Y, Zhu Y, et al. (2018). Integrative functional genomic analysis of human brain development and neuropsychiatric risks. Science (80-.) 362, eaat7615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Y, Lu H, Cheng P, Ge S, Xu H, Shi S-H, and Dan Y (2012). Clonally related visual cortical neurons show similar stimulus feature selectivity. Nature 486, 118–121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Y, Muffat J, Omer A, Bosch I, Lancaster MA, Sur M, Gehrke L, Knoblich JA, and Jaenisch R (2017). Induction of Expansion and Folding in Human Cerebral Organoids. Cell Stem Cell 20, 385–396.e3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lodato S, and Arlotta P (2015). Generating Neuronal Diversity in the Mammalian Cerebral Cortex. Annu. Rev. Cell Dev. Biol 31, 699–720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lodato S, Rouaux C, Quast KB, Jantrachotechatchawan C, Studer M, Hensch TK, and Arlotta P (2011). Excitatory projection neuron subtypes control the distribution of local inhibitory interneurons in the cerebral cortex. Neuron 69, 763–779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- London M, and Hausser M (2005). DENDRITIC COMPUTATION. Annu. Rev. Neurosci 28, 503–532. [DOI] [PubMed] [Google Scholar]
- Van der Loos H, and Woolsey TA (1973). Somatosensory cortex: structural alterations following early injury to sense organs. Science 179, 395–398. [DOI] [PubMed] [Google Scholar]
- Lorente de No R (1933). Studies on the structure of the cerebral cortex. I. Area entorhinalis. J Psychol Neurol 45, 381–438. [Google Scholar]
- Lorente de Nó R (1949). Cerebral cortex: architecture, intracortical connections, motor projections. In Physiology of the Nervous System, Fulton JF, ed. (New York, NY: Oxford University Press; ), pp. 288–312. [Google Scholar]
- Low LK, Liu X-B, Faulkner RL, Coble J, and Cheng H-J (2008). Plexin signaling selectively regulates the stereotyped pruning of corticospinal axons from visual cortex. Proc. Natl. Acad. Sci. U. S. A 105, 8136–8141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lukaszewicz A, Savatier P, Cortay V, Giroud P, Huissoud C, Berland M, Kennedy H, and Dehay C (2005). G1 phase regulation, area-specific cell cycle control, and cytoarchitectonics in the primate cortex. Neuron 47, 353–364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo C, Keown CL, Kurihara L, Zhou J, He Y, Li J, Castanon R, Lucero J, Nery JR, Sandoval JP, et al. (2017). Single-cell methylomes identify neuronal subtypes and regulatory elements in mammalian cortex. Science 357, 600–604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackarehtschian K, Lau CK, Caras I, and McConnell SK (1999). Regional differences in the developing cerebral cortex revealed by ephrin-A5 expression. Cereb. Cortex 9, 601–610. [DOI] [PubMed] [Google Scholar]
- Malik S, Vinukonda G, Vose LR, Diamond D, Bhimavarapu BBR, Hu F, Zia MT, Hevner R, Zecevic N, and Ballabh P (2013). Neurogenesis continues in the third trimester of pregnancy and is suppressed by premature birth. J. Neurosci 33, 411–423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mallamaci A, and Stoykova A (2006). Gene networks controlling early cerebral cortex arealization. Eur. J. Neurosci 23, 847–856. [DOI] [PubMed] [Google Scholar]
- Mallamaci A, Muzio L, Chan C-H, Parnavelas J, and Boncinelli E (2000). Area identity shifts in the early cerebral cortex of Emx2−/− mutant mice. Nat. Neurosci 3, 679–686. [DOI] [PubMed] [Google Scholar]
- Marblestone AH, Wayne G, and Kording KP (2016). Toward an Integration of Deep Learning and Neuroscience. Front. Comput. Neurosci 10, 94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marcus G, Marblestone A, and Dean T (2014). The atoms of neural computation. Science (80-.) 346, 551–552. [DOI] [PubMed] [Google Scholar]
- Marin-Padilla M (1983). Structural organization of the human cerebral cortex prior to the appearance of the cortical plate. Anat. Embryol. (Berl) 168, 21–40. [DOI] [PubMed] [Google Scholar]
- Marin-Padilla M (2015). Human cerebral cortex Cajal-Retzius neuron: development, structure and function. A Golgi study. Front. Neuroanat 9, 21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marin O, and Rubenstein JLR (2003). Cell migration in the forebrain. Annu. Rev. Neurosci 26, 441–483. [DOI] [PubMed] [Google Scholar]
- Markov NT, Ercsey-Ravasz M, Van Essen DC, Knoblauch K, Toroczkai Z, and Kennedy H (2013). Cortical High-Density Counterstream Architectures. Science (80-.) 342, 1238406–1238406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Markram H, Muller E, Ramaswamy S, Reimann MW, Abdellah M, Sanchez CA, Ailamaki A, Alonso-Nanclares L, Antille N, Arsever S, et al. (2015). Reconstruction and Simulation of Neocortical Microcircuitry. Cell 163, 456–492. [DOI] [PubMed] [Google Scholar]
- Marques-Smith A, Lyngholm D, Kaufmann A-K, Stacey JA, Hoerder-Suabedissen A, Becker EBE, Wilson MC, Molnar Z, and Butt SJB (2016). A Transient Translaminar GABAergic Interneuron Circuit Connects Thalamocortical Recipient Layers in Neonatal Somatosensory Cortex. Neuron 89, 536–549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martínez-Martínez MÁ, De Juan Romero C, Fernández V, Cárdenas A, Götz M, and Borrell V (2016). A restricted period for formation of outer subventricular zone defined by Cdh1 and Trnp1 levels. Nat. Commun 7, 11812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsumoto N, Shinmyo Y, Ichikawa Y, and Kawasaki H (2017). Gyrification of the cerebral cortex requires FGF signaling in the mammalian brain. Elife 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayer C, Jaglin XH, Cobbs LV, Bandler RC, Streicher C, Cepko CL, Hippenmeyer S, and Fishell G (2015). Clonally Related Forebrain Interneurons Disperse Broadly across Both Functional Areas and Structural Boundaries. Neuron 87, 989–998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayer C, Bandler RC, and Fishell G (2016). Lineage Is a Poor Predictor of Interneuron Positioning within the Forebrain. Neuron 92, 45–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCulloch WS, and Pitts W (1990). A logical calculus of the ideas immanent in nervous activity. 1943. Bull. Math. Biol 52, 99–115; discussion 73–97. [PubMed] [Google Scholar]
- Merlin S, Horng S, Marotte LR, Sur M, Sawatari A, and Leamey CA (2013). Deletion of Ten-m3 induces the formation of eye dominance domains in mouse visual cortex. Cereb. Cortex 23, 763–774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Metin C, and Frost DO (1989). Visual responses of neurons in somatosensory cortex of hamsters with experimentally induced retinal projections to somatosensory thalamus. Proc. Natl. Acad. Sci. U. S. A 86, 357–361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer G, and González-Gómez M (2018). The Subpial Granular Layer and Transient Versus Persisting Cajal-Retzius Neurons of the Fetal Human Cortex. Cereb. Cortex 28, 2043–2058. [DOI] [PubMed] [Google Scholar]
- Meyer G, Schaaps JP, Moreau L, and Goffinet AM (2000). Embryonic and early fetal development of the human neocortex. J. Neurosci 20, 1858–1868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mich JK, Hess EE, Graybuck LT, Somasundaram S, Miller JA, Ding Y, Shapovalova NV, Fong O, Yao S, Mortrud M, et al. (2019). Epigenetic landscape and AAV targeting of human neocortical cell classes. BioRxiv 555318. [Google Scholar]
- Michelsen KA, Acosta-Verdugo S, Benoit-Marand M, Espuny-Camacho I, Gaspard N, Saha B, Gaillard A, and Vanderhaeghen P (2015). Area-specific reestablishment of damaged circuits in the adult cerebral cortex by cortical neurons derived from mouse embryonic stem cells. Neuron 85, 982–997. [DOI] [PubMed] [Google Scholar]
- Mihalas AB, Elsen GE, Bedogni F, Daza RAM, Ramos-Laguna KA, Arnold SJ, and Hevner RF (2016). Intermediate Progenitor Cohorts Differentially Generate Cortical Layers and Require Tbr2 for Timely Acquisition of Neuronal Subtype Identity. Cell Rep. 16, 92–105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller JA, Ding S-L, Sunkin SM, Smith KA, Ng L, Szafer A, Ebbert A, Riley ZL, Royall JJ, Aiona K, et al. (2014). Transcriptional landscape of the prenatal human brain. Nature 508, 199–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miura E, Iijima T, Yuzaki M, and Watanabe M (2006). Distinct expression of Cbln family mRNAs in developing and adult mouse brains. Eur. J. Neurosci 24, 750–760. [DOI] [PubMed] [Google Scholar]
- Miyashita-Lin EM, Hevner R, Wassarman KM, Martinez S, and Rubenstein JL (1999). Early neocortical regionalization in the absence of thalamic innervation. Science 285, 906–909. [DOI] [PubMed] [Google Scholar]
- Miyata T, Kawaguchi A, Okano H, and Ogawa M (2001). Asymmetric inheritance of radial glial fibers by cortical neurons. Neuron 31, 727–741. [DOI] [PubMed] [Google Scholar]
- Moeller C, Yaylaoglu MB, Alvarez-Bolado G, Thaller C, and Eichele G (2002). Murine Lix1, a novel marker for substantia nigra, cortical layer 5, and hindbrain structures. Brain Res. Gene Expr. Patterns 1, 199–203. [DOI] [PubMed] [Google Scholar]
- Moldrich RX, Mezzera C, Holmes WM, Goda S, Brookfield SJ, Rankin AJ, Barr E, Kurniawan N, Dewar D, Richards LJ, et al. (2011). Fgfr3 regulates development of the caudal telencephalon. Dev. Dyn 240, 1586–1599. [DOI] [PubMed] [Google Scholar]
- Molliver ME, Kostović I, and van der Loos H (1973). The development of synapses in cerebral cortex of the human fetus. Brain Res. 50, 403–407. [DOI] [PubMed] [Google Scholar]
- Molnár Z (2013). Cortical Columns. In Comprehensive Developmental Neuroscience: Neural Circuit Development and Function in the Healthy and Diseased Brain, Rubenstein JLR, and Rakic P, eds. (London: Academic Press; ), pp. 109–129. [Google Scholar]
- Molnár Z, Adams R, and Blakemore C (1998). Mechanisms underlying the establishment of thalamocortical connections in the rat. J Neurosci 18, 5723–5745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molnar Z and Blakemore C (1995). How do thalamic axons find their way to the cortex? Trends Neurosci 18, 389–397. [DOI] [PubMed] [Google Scholar]
- Molyneaux BJ, Arlotta P, Menezes JRL, and Macklis JD (2007). Neuronal subtype specification in the cerebral cortex. Nat. Rev. Neurosci 8, 427–437. [DOI] [PubMed] [Google Scholar]
- Moore AR, Filipovic R, Mo Z, Rasband MN, Zecevic N, and Antic SD (2009). Electrical Excitability of Early Neurons in the Human Cerebral Cortex during the Second Trimester of Gestation. Cereb. Cortex 19, 1795–1805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore AR, Zhou W-L, Jakovcevski I, Zecevic N, and Antic SD (2011). Spontaneous electrical activity in the human fetal cortex in vitro. J. Neurosci 31, 2391–2398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moreno-Juan V, Filipchuk A, Anton-Bolanos N, Mezzera C, Gezelius H, Andres B, Rodriguez-Malmierca L, Susin R, Schaad O, Iwasato T, et al. (2017). Prenatal thalamic waves regulate cortical area size prior to sensory processing. Nat. Commun 8, 14172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mountcastle V (1997). The columnar organization of the neocortex. Brain 120, 701–722. [DOI] [PubMed] [Google Scholar]
- Mountcastle VB (1957). Modality and topographic properties of single neurons of cat’s somatic sensory cortex. J. Neurophysiol 20, 408–434. [DOI] [PubMed] [Google Scholar]
- Nakagawa Y, Johnson JE, and O’Leary DD (1999). Graded and areal expression patterns of regulatory genes and cadherins in embryonic neocortex independent of thalamocortical input. J. Neurosci 19, 10877–10885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieto M, Monuki ES, Tang H, Imitola J, Haubst N, Khoury SJ, Cunningham J, Gotz M, and Walsh CA (2004). Expression of Cux-1 and Cux-2 in the subventricular zone and upper layers II-IV of the cerebral cortex. J. Comp. Neurol 479, 168–180. [DOI] [PubMed] [Google Scholar]
- Noctor SC, Flint AC, Weissman TA, Dammerman RS, and Kriegstein AR (2001). Neurons derived from radial glial cells establish radial units in neocortex. Nature 409, 714–720. [DOI] [PubMed] [Google Scholar]
- Noctor SC, Martḿez-Cerdeño V, Ivic L, and Kriegstein AR (2004). Cortical neurons arise in symmetric and asymmetric division zones and migrate through specific phases. Nat. Neurosci 7, 136–144. [DOI] [PubMed] [Google Scholar]
- Nonaka-Kinoshita M, Reillo I, Artegiani B, Martínez-Martínez MÁ, Nelson M, Borrell V, and Calegari F (2013). Regulation of cerebral cortex size and folding by expansion of basal progenitors. EMBO J. 32, 1817–1828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Northcutt RG, and Kaas JH (1995). The emergence and evolution of mammalian neocortex. Trends Neurosci. 18, 373–379. [DOI] [PubMed] [Google Scholar]
- Nowakowski TJ, Pollen AA, Sandoval-Espinosa C, and Kriegstein AR (2016). Transformation of the Radial Glia Scaffold Demarcates Two Stages of Human Cerebral Cortex Development. Neuron 91, 1219–1227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nowakowski TJ, Bhaduri A, Pollen AA, Alvarado B, Mostajo-Radji MA, Di Lullo E, Haeussler M, Sandoval-Espinosa C, Liu SJ, Velmeshev D, et al. (2017). Spatiotemporal gene expression trajectories reveal developmental hierarchies of the human cortex. Science (80-.) 358, 1318–1323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Leary DD (1989). Do cortical areas emerge from a protocortex? Trends Neurosci. 12, 400–406. [DOI] [PubMed] [Google Scholar]
- O’Leary DDM, Chou S-J, and Sahara S (2007). Area patterning of the mammalian cortex. Neuron 56, 252–269. [DOI] [PubMed] [Google Scholar]
- O’Reilly C, Lewis JD, and Elsabbagh M (2017). Is functional brain connectivity atypical in autism? A systematic review of EEG and MEG studies. PLoS One 12, e0175870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh WC, Lutzu S, Castillo PE, and Kwon H-B (2016). De novo synaptogenesis induced by GABA in the developing mouse cortex. Science 353, 1037–1040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohki K, Chung S, Ch’ng YH, Kara P, and Reid RC (2005). Functional imaging with cellular resolution reveals precise micro-architecture in visual cortex. Nature 433, 597–603. [DOI] [PubMed] [Google Scholar]
- Ohtsuki G, Nishiyama M, Yoshida T, Murakami T, Histed M, Lois C, and Ohki K (2012). Similarity of visual selectivity among clonally related neurons in visual cortex. Neuron 75, 65–72. [DOI] [PubMed] [Google Scholar]
- Ozair MZ, Kirst C, van den Berg BL, Ruzo A, Rito T, and Brivanlou AH (2018). hPSC Modeling Reveals that Fate Selection of Cortical Deep Projection Neurons Occurs in the Subplate. Cell Stem Cell 23, 60–73.e6. [DOI] [PubMed] [Google Scholar]
- Packer AM, and Yuste R (2011). Dense, unspecific connectivity of neocortical parvalbumin-positive interneurons: a canonical microcircuit for inhibition? J. Neurosci 31, 13260–13271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palomero-Gallagher N, and Zilles K (2017). Cortical layers: Cyto-, myelo-, receptor- and synaptic architecture in human cortical areas. Neuroimage. [DOI] [PubMed] [Google Scholar]
- Paredes MF, James D, Gil-Perotin S, Kim H, Cotter JA, Ng C, Sandoval K, Rowitch DH, Xu D, McQuillen PS, et al. (2016). Extensive migration of young neurons into the infant human frontal lobe. Science (80-.) 354, aaf7073–aaf7073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patel AB, Nguyen T, and Baraniuk RG (2015). A Probabilistic Theory of Deep Learning.
- Pedraza M, Hoerder-Suabedissen A, Albert-Maestro MA, Molnar Z, and De Carlos JA (2014). Extracortical origin of some murine subplate cell populations. Proc. Natl. Acad. Sci. U. S. A 111, 8613–8618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perin R, Berger TK, and Markram H (2011). A synaptic organizing principle for cortical neuronal groups. Proc. Natl. Acad. Sci 108, 5419–5424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peters A, and Kara DA (1985). The neuronal composition of area 17 of rat visual cortex. I. The pyramidal cells. J. Comp. Neurol 234, 218–241. [DOI] [PubMed] [Google Scholar]
- Peters A, and Kara DA (1987). The neuronal composition of area 17 of rat visual cortex. IV. The organization of pyramidal cells. J. Comp. Neurol 260, 573–590. [DOI] [PubMed] [Google Scholar]
- Peters A, and Walsh TM (1972). A study of the organization of apical dendrites in the somatic sensory cortex of the rat. J. Comp. Neurol 144, 253–268. [DOI] [PubMed] [Google Scholar]
- Pletikos M, Sousa AMM, Sedmak G, Meyer KA, Zhu Y, Cheng F, Li M, Kawasawa YI, and S estan N (2014). Temporal Specification and Bilaterality of Human Neocortical Topographic Gene Expression. Neuron 81, 321–332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollen AA, Nowakowski TJ, Chen J, Retallack H, Sandoval-Espinosa C, Nicholas CR, Shuga J, Liu SJ, Oldham MC, Diaz A, et al. (2015). Molecular Identity of Human Outer Radial Glia during Cortical Development. Cell 163, 55–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pontious A, Kowalczyk T, Englund C, and Hevner RF (2008). Role of Intermediate Progenitor Cells in Cerebral Cortex Development. Dev. Neurosci 30, 24–32. [DOI] [PubMed] [Google Scholar]
- Pouchelon G, Gambino F, Bellone C, Telley L, Vitali I, Luscher C, Holtmaat A, and Jabaudon D (2014). Modality-specific thalamocortical inputs instruct the identity of postsynaptic L4 neurons. Nature 511, 471–474. [DOI] [PubMed] [Google Scholar]
- Price DJ, and Blakemore C (1985a). The postnatal development of the association projection from visual cortical area 17 to area 18 in the cat. J. Neurosci 5, 2443–2452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price DJ, and Blakemore C (1985b). Regressive events in the postnatal development of association projections in the visual cortex. Nature 316, 721–724. [DOI] [PubMed] [Google Scholar]
- Price DJ, Aslam S, Tasker L, and Gillies K (1997). Fates of the earliest generated cells in the developing murine neocortex. J. Comp. Neurol 377, 414–422. [PubMed] [Google Scholar]
- Priya R, Paredes MF, Karayannis T, Yusuf N, Liu X, Jaglin X, Graef I, Alvarez-Buylla A, and Fishell G (2018). Activity Regulates Cell Death within Cortical Interneurons through a Calcineurin-Dependent Mechanism. Cell Rep. 22, 1695–1709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rakic P (1972). Mode of cell migration to the superficial layers of fetal monkey neocortex. J. Comp. Neurol 145, 61–83. [DOI] [PubMed] [Google Scholar]
- Rakic P (1974). Neurons in rhesus monkey visual cortex: systematic relation between time of origin and eventual disposition. Science 183, 425–427. [DOI] [PubMed] [Google Scholar]
- Rakic P (1975). Timing of major ontogenetic events in the visual cortex of the rhesus monkey. UCLA Forum Med. Sci 3–40. [DOI] [PubMed] [Google Scholar]
- Rakic P (1978). Neuronal migration and contact guidance in the primate telencephalon. Postgrad. Med. J 54 Suppl 1, 25–40. [PubMed] [Google Scholar]
- Rakic P (1988). Specification of cerebral cortical areas. Science 241, 170–176. [DOI] [PubMed] [Google Scholar]
- Rakic P (1991). Experimental manipulation of cerebral cortical areas in primates. Philos. Trans. R. Soc. Lond. B. Biol. Sci 331, 291–294. [DOI] [PubMed] [Google Scholar]
- Rakic P (2008). Radial Unit Hypothesis of Neocortical Expansion. pp. 30–45. [DOI] [PubMed]
- Rakic P, and Sidman RL (1968a). Supravital DNA synthesis in the developing human and mouse brain. J. Neuropathol. Exp. Neurol 27, 246–276. [PubMed] [Google Scholar]
- Rakic P, and Sidman RL (1968b). Autoradiographic study of supravital DNA synthesis in fetal human brain. J. Neuropathol. Exp. Neurol 27, 139–140. [PubMed] [Google Scholar]
- Rakic S, and Zecevic N (2000). Programmed cell death in the developing human telencephalon. Eur. J. Neurosci 12, 2721–2734. [DOI] [PubMed] [Google Scholar]
- Rakic P, Bourgeois JP, Eckenhoff MF, Zecevic N, and Goldman-Rakic PS (1986). Concurrent overproduction of synapses in diverse regions of the primate cerebral cortex. Science 232, 232–235. [DOI] [PubMed] [Google Scholar]
- Rakic P, Bourgeois J-P, and Goldman-Rakic PS (1994). Synaptic development of the cerebral cortex: implications for learning, memory, and mental illness. In Progress in Brain Research, pp. 227–243. [DOI] [PubMed] [Google Scholar]
- Rash BG, Duque A, Morozov YM, Arellano JI, Micali N, and Rakic P (2019). Gliogenesis in the outer subventricular zone promotes enlargement and gyrification of the primate cerebrum. Proc. Natl. Acad. Sci. U. S. A 116, 7089–7094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Redcay E, and Courchesne E (2005). When is the brain enlarged in autism? A meta-analysis of all brain size reports. Biol. Psychiatry 58, 1–9. [DOI] [PubMed] [Google Scholar]
- Reillo I, de Juan Romero C, Garcia-Cabezas MA, and Borrell V (2011). A role for intermediate radial glia in the tangential expansion of the mammalian cerebral cortex. Cereb. Cortex 21, 1674–1694. [DOI] [PubMed] [Google Scholar]
- Richman DP, Stewart RM, Hutchinson JW, and Caviness VS (1975). Mechanical model of brain convolutional development. Science 189, 18–21. [DOI] [PubMed] [Google Scholar]
- Ringach DL, Mineault PJ, Tring E, Olivas ND, Garcia-Junco-Clemente P, and Trachtenberg JT (2016). Spatial clustering of tuning in mouse primary visual cortex. Nat. Commun 7, 12270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rockel AJ, Hiorns RW, and Powell TP (1980). The basic uniformity in structure of the neocortex. Brain 103, 221–244. [DOI] [PubMed] [Google Scholar]
- Roe AW, Pallas SL, Kwon YH, and Sur M (1992). Visual projections routed to the auditory pathway in ferrets: receptive fields of visual neurons in primary auditory cortex. J. Neurosci 12, 3651–3664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roth RB, Hevezi P, Lee J, Willhite D, Lechner SM, Foster AC, and Zlotnik A (2006). Gene expression analyses reveal molecular relationships among 20 regions of the human CNS. Neurogenetics 7, 67–80. [DOI] [PubMed] [Google Scholar]
- Rowell JJ, Mallik AK, Dugas-Ford J, and Ragsdale CW (2010). Molecular analysis of neocortical layer structure in the ferret. J. Comp. Neurol 518, 3272–3289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rubenstein JL, and Rakic P (1999). Genetic control of cortical development. Cereb. Cortex 9, 521–523. [DOI] [PubMed] [Google Scholar]
- Rubenstein JL, Shimamura K, Martinez S, and Puelles L (1998). Regionalization of the prosencephalic neural plate. Annu. Rev. Neurosci 21, 445–477. [DOI] [PubMed] [Google Scholar]
- Rubenstein JL, Anderson S, Shi L, Miyashita-Lin E, Bulfone A, and Hevner R (1999). Genetic control of cortical regionalization and connectivity. Cereb. Cortex 9, 524–532. [DOI] [PubMed] [Google Scholar]
- Sacramento J, Costa RP, Bengio Y, and Senn W (2018). Dendritic cortical microcircuits approximate the backpropagation algorithm.
- Saez I, and Friedlander MJ (2009). Synaptic output of individual layer 4 neurons in guinea pig visual cortex. J. Neurosci 29, 4930–4944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sahara S, and O’Leary DDM (2009). Fgf10 Regulates Transition Period of Cortical Stem Cell Differentiation to Radial Glia Controlling Generation of Neurons and Basal Progenitors. Neuron 63, 48–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sahara S, Kawakami Y, Izpisua Belmonte JC, and O’Leary DDM (2007). Sp8 exhibits reciprocal induction with Fgf8 but has an opposing effect on anterior-posterior cortical area patterning. Neural Dev. 2, 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saunders A, Macosko EZ, Wysoker A, Goldman M, Krienen FM, de Rivera H, Bien E, Baum M, Bortolin L, Wang S, et al. (2018). Molecular Diversity and Specializations among the Cells of the Adult Mouse Brain. Cell 174, 1015–1030.e16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savarese A, Zou ME, Kharazia V, Maiya R, and Lasek AW (2014). Increased behavioral responses to ethanol in Lmo3 knockout mice. Genes, Brain Behav. 13, 777–783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scala F, Kobak D, Shan S, Bernaerts Y, Laturnus S, Cadwell CR, Hartmanis L, Froudarakis E, Castro J, Tan ZH, et al. (2019). Neocortical layer 4 in adult mouse differs in major cell types and circuit organization between primary sensory areas. BioRxiv 507293. [Google Scholar]
- Schaeren-Wiemers N, Andre E, Kapfhammer JP, and Becker-Andre M (1997). The expression pattern of the orphan nuclear receptor RORbeta in the developing and adult rat nervous system suggests a role in the processing of sensory information and in circadian rhythm. Eur. J. Neurosci 9, 2687–2701. [DOI] [PubMed] [Google Scholar]
- Schlaggar BL, and O’Leary DD (1991). Potential of visual cortex to develop an array of functional units unique to somatosensory cortex. Science 252, 1556–1560. [DOI] [PubMed] [Google Scholar]
- Schleicher A, Amunts K, Geyer S, Kowalski T, Schormann T, Palomero-Gallagher N, and Zilles K (2000). A stereological approach to human cortical architecture: identification and delineation of cortical areas. J. Chem. Neuroanat 20, 31–47. [DOI] [PubMed] [Google Scholar]
- Seiradake E, del Toro D, Nagel D, Cop F, Hartl R, Ruff T, Seyit-Bremer G, Harlos K, Border EC, Acker-Palmer A, et al. (2014). FLRT structure: balancing repulsion and cell adhesion in cortical and vascular development. Neuron 84, 370–385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sessa A, Mao C-A, Colasante G, Nini A, Klein WH, and Broccoli V (2010). Tbr2-positive intermediate (basal) neuronal progenitors safeguard cerebral cortex expansion by controlling amplification of pallial glutamatergic neurons and attraction of subpallial GABAergic interneurons. Genes Dev. 24, 1816–1826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma J, Angelucci A, and Sur M (2000). Induction of visual orientation modules in auditory cortex. Nature 404, 841–847. [DOI] [PubMed] [Google Scholar]
- Shatz CJ (1992). Dividing up the neocortex. Science (80-.) 258, 237–238. [DOI] [PubMed] [Google Scholar]
- Shetty AS, Godbole G, Maheshwari U, Padmanabhan H, Chaudhary R, Muralidharan B, Hou P-S, Monuki ES, Kuo H-C, Rema V, et al. (2013). Lhx2 regulates a cortex-specific mechanism for barrel formation. Proc. Natl. Acad. Sci. U. S. A 110, E4913–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimogori T, and Grove EA (2005). Fibroblast growth factor 8 regulates neocortical guidance of area-specific thalamic innervation. J. Neurosci 25, 6550–6560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shipp S (2005). The importance of being agranular: a comparative account of visual and motor cortex. Philos. Trans. R. Soc. Lond. B. Biol. Sci 360, 797–814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidman RL, and Rakic P (1973). Neuronal migration, with special reference to developing human brain: a review. Brain Res. 62, 1–35. [DOI] [PubMed] [Google Scholar]
- Silbereis JC, Pochareddy S, Zhu Y, Li M, and Sestan N (2016). The Cellular and Molecular Landscapes of the Developing Human Central Nervous System. Neuron 89, 248–268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silver D, Huang A, Maddison CJ, Guez A, Sifre L, van den Driessche G, Schrittwieser J, Antonoglou I, Panneershelvam V, Lanctot M, et al. (2016). Mastering the game of Go with deep neural networks and tree search. Nature 529, 484–489. [DOI] [PubMed] [Google Scholar]
- Silver D, Schrittwieser J, Simonyan K, Antonoglou I, Huang A, Guez A, Hubert T, Baker L, Lai M, Bolton A, et al. (2017). Mastering the game of Go without human knowledge. Nature 550, 354–359. [DOI] [PubMed] [Google Scholar]
- Simi A, and Studer M (2018). Developmental genetic programs and activity-dependent mechanisms instruct neocortical area mapping. Curr. Opin. Neurobiol 53, 96–102. [DOI] [PubMed] [Google Scholar]
- Smart IHM, Dehay C, Giroud P, Berland M, and Kennedy H (2002). Unique Morphological Features of the Proliferative Zones and Postmitotic Compartments of the Neural Epithelium Giving Rise to Striate and Extrastriate Cortex in the Monkey. Cereb. Cortex 12, 37–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith GB, and Fitzpatrick D (2012). Specifying Cortical Circuits: A Role for Cell Lineage. Neuron 75, 4–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Southwell DG, Froemke RC, Alvarez-Buylla A, Stryker MP, and Gandhi SP (2010). Cortical plasticity induced by inhibitory neuron transplantation. Science 327, 1145–1148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Srinivasan K, Leone DP, Bateson RK, Dobreva G, Kohwi Y, Kohwi-Shigematsu T, Grosschedl R, and McConnell SK (2012). A network of genetic repression and derepression specifies projection fates in the developing neocortex. Proc. Natl. Acad. Sci. U. S. A 109, 19071–19078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stahl R, Walcher T, De Juan Romero C, Pilz GA, Cappello S, Irmler M, Sanz-Aquela JM, Beckers J, Blum R, Borrell V, et al. (2013). Trnp1 regulates expansion and folding of the mammalian cerebral cortex by control of radial glial fate. Cell 153, 535–549. [DOI] [PubMed] [Google Scholar]
- Stevens HE, Smith KM, Maragnoli ME, Fagel D, Borok E, Shanabrough M, Horvath TL, and Vaccarino FM (2010). Fgfr2 is required for the development of the medial prefrontal cortex and its connections with limbic circuits. J. Neurosci 30, 5590–5602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoner R, Chow ML, Boyle MP, Sunkin SM, Mouton PR, Roy S, Wynshaw-Boris A, Colamarino SA, Lein ES, and Courchesne E (2014). Patches of disorganization in the neocortex of children with autism. N. Engl. J. Med 370, 1209–1219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoykova A, Treichel D, Hallonet M, and Gruss P (2000). Pax6 modulates the dorsoventral patterning of the mammalian telencephalon. J. Neurosci 20, 8042–8050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sudhof TC (2018). Towards an Understanding of Synapse Formation. Neuron 100, 276–293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sultan KT, Han Z, Zhang X-J, Xianyu A, Li Z, Huang K, and Shi S-H (2016). Clonally Related GABAergic Interneurons Do Not Randomly Disperse but Frequently Form Local Clusters in the Forebrain. Neuron 92, 31–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sur M, Garraghty PE, and Roe AW (1988). Experimentally induced visual projections into auditory thalamus and cortex. Science 242, 1437–1441. [DOI] [PubMed] [Google Scholar]
- Suzuki N, Fukushi M, Kosaki K, Doyle AD, de Vega S, Yoshizaki K, Akazawa C, Arikawa-Hirasawa E, and Yamada Y (2012). Teneurin-4 is a novel regulator of oligodendrocyte differentiation and myelination of small-diameter axons in the CNS. J. Neurosci 32, 11586–11599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki SC, Inoue T, Kimura Y, Tanaka T, and Takeichi M (1997). Neuronal circuits are subdivided by differential expression of type-II classic cadherins in postnatal mouse brains. Mol. Cell. Neurosci 9, 433–447. [DOI] [PubMed] [Google Scholar]
- Tarusawa E, Sanbo M, Okayama A, Miyashita T, Hirayama T, Hirabayashi T, Hasegawa S, Kaneko R, Toyoda S, Kobayashi T, et al. (2016). Establishment of high reciprocal connectivity between clonally related cortical neurons is regulated by the Dnmt3b DNA methyltransferase and clustered protocadherins. BMC Biol 14, 103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tasic B, Yao Z, Graybuck LT, Smith KA, Nguyen TN, Bertagnolli D, Goldy J, Garren E, Economo MN, Viswanathan S, et al. (2018). Shared and distinct transcriptomic cell types across neocortical areas. Nature 563, 72–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Telley L, Govindan S, Prados J, Stevant I, Nef S, Dermitzakis E, Dayer A, and Jabaudon D (2016). Sequential transcriptional waves direct the differentiation of newborn neurons in the mouse neocortex. Science 351, 1443–1446. [DOI] [PubMed] [Google Scholar]
- Telley L, Agirman G, Prados J, Amberg N, Fiévre S, Oberst P, Bartolini G, Vitali I, Cadilhac C, Hippenmeyer S, et al. (2019). Temporal patterning of apical progenitors and their daughter neurons in the developing neocortex. Science 364, pii: eaav2522. [DOI] [PubMed] [Google Scholar]
- Terakawa YW, Inoue YU, Asami J, Hoshino M, and Inoue T (2013). A sharp cadherin-6 gene expression boundary in the developing mouse cortical plate demarcates the future functional areal border. Cereb. Cortex 23, 2293–2308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomsen ER, Mich JK, Yao Z, Hodge RD, Doyle AM, Jang S, Shehata SI, Nelson AM, Shapovalova NV, Levi BP, et al. (2016). Fixed single-cell transcriptomic characterization of human radial glial diversity. Nat. Methods 13, 87–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torii M, Hashimoto-Torii K, Levitt P, and Rakic P (2009). Integration of neuronal clones in the radial cortical columns by EphA and ephrin-A signalling. Nature 461, 524–528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tumaneng K, Russell RC, and Guan K-L (2012). Organ Size Control by Hippo and TOR Pathways. Curr. Biol 22, R368–R379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tuoc TC, Radyushkin K, Tonchev AB, Pinon MC, Ashery-Padan R, Molnar Z, Davidoff MS, and Stoykova A (2009). Selective cortical layering abnormalities and behavioral deficits in cortex-specific Pax6 knock-out mice. J. Neurosci 29, 8335–8349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turrero Garcia M, Mazzola E, and Harwell CC (2016). Lineage Relationships Do Not Drive MGE/PoA-Derived Interneuron Clustering in the Brain. Neuron 92, 52–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vaidya A, Pniak A, Lemke G, and Brown A (2003). EphA3 Null Mutants Do Not Demonstrate Motor Axon Guidance Defects. Mol. Cell. Biol 23, 8092–8098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varea O, Martin-de-Saavedra MD, Kopeikina KJ, Schurmann B, Fleming HJ, Fawcett-Patel JM, Bach A, Jang S, Peles E, Kim E, et al. (2015). Synaptic abnormalities and cytoplasmic glutamate receptor aggregates in contactin associated protein-like 2/Caspr2 knockout neurons. Proc. Natl. Acad. Sci. U. S. A 112, 6176–6181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varga C, Lee SY, and Soltesz I (2010). Target-selective GABAergic control of entorhinal cortex. Nat Neurosci 13, 822–824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vasistha NA, García-Moreno F, Arora S, Cheung AFP, Arnold SJ, Robertson EJ, and Molnar Z (2015). Cortical and Clonal Contribution of Tbr2 Expressing Progenitors in the Developing Mouse Brain. Cereb. Cortex 25, 3290–3302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ventre-Dominey J (2014). Vestibular function in the temporal and parietal cortex: distinct velocity and inertial processing pathways. Front. Integr. Neurosci 8, 53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vue TY, Lee M, Tan YE, Werkhoven Z, Wang L, and Nakagawa Y (2013). Thalamic Control of Neocortical Area Formation in Mice. J. Neurosci 33, 8442–8453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L, Hou S, and Han Y-G (2016). Hedgehog signaling promotes basal progenitor expansion and the growth and folding of the neocortex. Nat. Neurosci 19, 888–896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wass S (2011). Distortions and disconnections: Disrupted brain connectivity in autism. Brain Cogn. 75, 18–28. [DOI] [PubMed] [Google Scholar]
- Watakabe A, Ohsawa S, Hashikawa T, and Yamamori T (2006). Binding and complementary expression patterns of semaphorin 3E and plexin D1 in the mature neocortices of mice and monkeys. J. Comp. Neurol 499, 258–273. [DOI] [PubMed] [Google Scholar]
- Watanabe M, Buth JE, Vishlaghi N, de la Torre-Ubieta L, Taxidis J, Khakh BS, Coppola G, Pearson CA, Yamauchi K, Gong D, et al. (2017). Self-Organized Cerebral Organoids with Human-Specific Features Predict Effective Drugs to Combat Zika Virus Infection. Cell Rep. 21, 517–532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watson C, and Puelles L (2017). Developmental gene expression in the mouse clarifies the organization of the claustrum and related endopiriform nuclei. J. Comp. Neurol 525, 1499–1508. [DOI] [PubMed] [Google Scholar]
- Wester JC, Mahadevan V, Rhodes CT, Calvigioni D, Venkatesh S, Maric D, Hunt S, Yuan X, Zhang Y, Petros TJ, et al. (2019). Neocortical Projection Neurons Instruct Inhibitory Interneuron Circuit Development in a Lineage-Dependent Manner. Neuron. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whittington JCR, and Bogacz R (2019). Theories of Error Back-Propagation in the Brain. Trends Cogn. Sci 23, 235–250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilkie AL, Jordan SA, Sharpe JA, Price DJ, and Jackson IJ (2004). Widespread tangential dispersion and extensive cell death during early neurogenesis in the mouse neocortex. Dev. Biol 267, 109–118. [DOI] [PubMed] [Google Scholar]
- Willsey AJ, Sanders SJ, Li M, Dong S, Tebbenkamp AT, Muhle RA, Reilly SK, Lin L, Fertuzinhos S, Miller JA, et al. (2013). Coexpression networks implicate human midfetal deep cortical projection neurons in the pathogenesis of autism. Cell 155, 997–1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winden KD, Ebrahimi-Fakhari D, and Sahin M (2018). Abnormal mTOR Activation in Autism. Annu. Rev. Neurosci 41, 1–23. [DOI] [PubMed] [Google Scholar]
- Wong FK, Bercsenyi K, Sreenivasan V, Portalés A, Fernández-Otero M, and Marín O (2018). Pyramidal cell regulation of interneuron survival sculpts cortical networks. Nature 557, 668–673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woolsey TA, and Van der Loos H (1970). The structural organization of layer IV in the somatosensory region (SI) of mouse cerebral cortex. The description of a cortical field composed of discrete cytoarchitectonic units. Brain Res. 17, 205–242. [DOI] [PubMed] [Google Scholar]
- Woolsey TA, Dierker ML, and Wann DF (1975). Mouse SmI cortex: qualitative and quantitative classification of golgi-impregnated barrel neurons. Proc. Natl. Acad. Sci. U. S. A 72, 2165–2169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu C, Li Q, Efimova O, He L, Tatsumoto S, Stepanova V, Oishi T, Udono T, Yamaguchi K, Shigenobu S, et al. (2018). Human-specific features of spatial gene expression and regulation in eight brain regions. Genome Res. 28,1097–1110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu X, Roby KD, and Callaway EM (2010). Immunochemical characterization of inhibitory mouse cortical neurons: three chemically distinct classes of inhibitory cells. J. Comp. Neurol 518, 389–404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamamoto A, Kemp C, Bachiller D, Geissert D, and De Robertis EM (2000). Mouse paraxial protocadherin is expressed in trunk mesoderm and is not essential for mouse development. Genesis 27, 49–57. [PubMed] [Google Scholar]
- Yamawaki N, Borges K, Suter BA, Harris KD, and Shepherd GMG (2014). A genuine layer 4 in motor cortex with prototypical synaptic circuit connectivity. Elife 3, e05422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye Z, Mostajo-Radji MA, Brown JR, Rouaux C, Tomassy GS, Hensch TK, and Arlotta P (2015). Instructing Perisomatic Inhibition by Direct Lineage Reprogramming of Neocortical Projection Neurons. Neuron 88, 475–483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoon K-J, Koo B-K, Im S-K, Jeong H-W, Ghim J, Kwon M, Moon J-S, Miyata T, and Kong Y-Y (2008). Mind Bomb 1-Expressing Intermediate Progenitors Generate Notch Signaling to Maintain Radial Glial Cells. Neuron 58, 519–531. [DOI] [PubMed] [Google Scholar]
- Yoshimura Y, Dantzker JLM, and Callaway EM (2005). Excitatory cortical neurons form fine-scale functional networks. Nature 433, 868–873. [DOI] [PubMed] [Google Scholar]
- Young NA, Collins CE, and Kaas JH (2013a). Cell and neuron densities in the primary motor cortex of primates. Front. Neural Circuits 7, 30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young TR, Bourke M, Zhou X, Oohashi T, Sawatari A, Fässler R, and Leamey CA (2013b). Ten-m2 is required for the generation of binocular visual circuits. J. Neurosci 33, 12490–12509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu Y-C, Bultje RS, Wang X, and Shi S-H (2009). Specific synapses develop preferentially among sister excitatory neurons in the neocortex. Nature 458, 501–504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu Y-C, He S, Chen S, Fu Y, Brown KN, Yao X-H, Ma J, Gao KP, Sosinsky GE, Huang K, et al. (2012). Preferential electrical coupling regulates neocortical lineage-dependent microcircuit assembly. Nature 486, 113–117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zecevic N (1998). Synaptogenesis in layer I of the human cerebral cortex in the first half of gestation. Cereb. Cortex 8, 245–252. [DOI] [PubMed] [Google Scholar]
- Zeng H, Shen EH, Hohmann JG, Oh SW, Bernard A, Royall JJ, Glattfelder KJ, Sunkin SM, Morris JA, Guillozet-Bongaarts AL, et al. (2012). Large-scale cellular-resolution gene profiling in human neocortex reveals species-specific molecular signatures. Cell 149, 483–496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu Y, Sousa AMM, Gao T, Skarica M, Li M, Santpere G, Esteller-Cucala P, Juan D, Ferrández-Peral L, Gulden FO, et al. (2018). Spatiotemporal transcriptomic divergence across human and macaque brain development. Science 362, eaat8077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Žunic Išasegi I, Radoš M, Krsnik Ž, Radoš M, Benjak V, and Kostović I (2018). Interactive histogenesis of axonal strata and proliferative zones in the human fetal cerebral wall. Brain Struct. Funct 223, 3919–3943. [DOI] [PMC free article] [PubMed] [Google Scholar]