

Abstract
Most rapid diagnostic tests for Plasmodium falciparum malaria target the Histidine-Rich Proteins 2 and 3 (HRP2 and HRP3). Deletions of the hrp2 and hrp3 genes result in false-negative tests and are a threat for malaria control. A novel assay for molecular surveillance of hrp2/hrp3 deletions was developed based on droplet digital PCR (ddPCR). The assay quantifies hrp2, hrp3, and a control gene with very high accuracy. The theoretical limit of detection was 0.33 parasites/µl. The deletion was reliably detected in mixed infections with wild-type and hrp2-deleted parasites at a density of >100 parasites/reaction. For a side-by-side comparison with the conventional nested PCR (nPCR) assay, 248 samples were screened in triplicate by ddPCR and nPCR. No deletions were observed by ddPCR, while by nPCR hrp2 deletion was observed in 8% of samples. The ddPCR assay was applied to screen 830 samples from Kenya, Zanzibar/Tanzania, Ghana, Ethiopia, Brazil, and Ecuador. Pronounced differences in the prevalence of deletions were observed among sites, with more hrp3 than hrp2 deletions. In conclusion, the novel ddPCR assay minimizes the risk of false-negative results (i.e., hrp2 deletion observed when the sample is wild type), increases sensitivity, and greatly reduces the number of reactions that need to be run.
Research organism: P. falciparum
Introduction
In 2019, over 200 million cases of malaria and over 400,000 deaths were recorded (World Health Organisation, 2020). Plasmodium falciparum remains the primary cause of malaria in humans. Fast and accurate diagnosis and treatment of clinical episodes are key components of malaria control. Diagnosis is commonly performed either by light microscopy, or rapid diagnostic tests (RDTs). RDTs are lateral flow devices that detect parasite proteins in human blood through immunohistochemistry. Light microscopy requires basic lab infrastructure and skilled microscopists. In contrast, RDTs require minimal infrastructure and training, and results are available within approximately 15 min at a cost of less than 1 USD per test. RDTs are the only field-deployable diagnostic tool available at peripheral health centers and for community screening to diagnose asymptomatic infections, for example through reactive case detection (RCD; Stuck et al., 2020). In 2016, over 300 million RDTs were used by malaria control programs (WHO, 2017).
The most sensitive RDTs for P. falciparum rely on the detection of Histidine-Rich Protein 2 (HRP2) (Li et al., 2017). HRP2 is a highly expressed secreted protein, and thus an ideal target for diagnosis. It is also the target for a new generation of ultra-sensitive RDTs with a limit of detection of <100 parasites/µl (Acquah et al., 2021; Hofmann et al., 2019). While alternative RDTs detecting other proteins, for example parasite lactase dehydrogenase (pLDH), or aldolase, are available, they are generally less sensitive (World Health Organisation, 2016a; World Health Organisation, 2018). HRP2-based RDTs can also detect HRP3, a structurally similar protein sharing multiple epitopes with HRP2.
In 2010, a report revealed that a large proportion of P. falciparum parasites in Peru did not carry the hrp2 gene (Gamboa et al., 2010), and thus could not be detected by HRP2-based RDTs. Since then, an increasing number of reports from Latin America (Maltha et al., 2012; Sáenz et al., 2015; Murillo Solano et al., 2015; Dorado et al., 2016; Akinyi Okoth et al., 2015), Africa (Amoah et al., 2016; Koita et al., 2012; Parr et al., 2017; Berhane et al., 2018; Menegon et al., 2017; Bharti et al., 2016; Li et al., 2015; Kumar et al., 2013), and Asia Johora et al., 2017; Pati et al., 2018 found varying proportions of parasites with hrp2 deletion, reaching up to 80% of clinical cases in certain hospitals in Eritrea (Berhane et al., 2018). In addition, hrp3 can be deleted. The deletion of either hrp2 or hrp3, or both genes, has no known impact on parasite fitness. RDTs can yield a positive result if hrp2 is deleted but hrp3 is expressed, but the sensitivity of the RDT is lower in this case (Kong et al., 2021).
Molecular surveillance to assess the frequency of hrp2 and hrp3 deletion is crucial to decide whether HRP2-based RDTs can be used (Cheng et al., 2014). The WHO recommends to use alternative diagnostics if the prevalence of hrp2 deletion is above 5% (World Health Organisation, 2016b). At this level, the number of tests that are false-negative tests because of hrp2 deletion will exceed the number of tests that are false-negative tests because alternative diagnostics offer a lower sensitivity (World Health Organisation, 2016b). The prevalence of hrp2 deletion has been found to differ substantially within countries (Murillo Solano et al., 2015; Parr et al., 2017), thus the choice of diagnostics might need to be adapted at subnational level. The feasibility of such approaches depends on the organization of the national control program, barriers to transmission within-country, and other factors. Where low levels of deletions are present, HRP2-based RDTs remain a highly useful tool for diagnosis.
Deletion screening has been classically done using nested PCR (nPCR) followed by gel electrophoresis (Cheng et al., 2014). Absence of a band is interpreted as deletion. False-negative results could occur when PCR conditions are suboptimal, or when parasite density is low and amplification is stochastic. To overcome this limitation, threefold repetition of the nested hrp2 PCR and a control PCR (e.g., msp2 or glurp) is recommended (Cheng et al., 2014). As a result, for each sample 12 PCRs need to be run. Deletion status might remain unresolved if results differ among replicates. As an additional problem, multiple clone P. falciparum infections are common in most endemic settings (Lopez and Koepfli, 2021; Grignard et al., 2020). In case of a multiple clone infection with a wild-type parasite and a parasite carrying the deletion, the wild-type parasite will result in a band on the gel when using the nPCR assay. While multiple clone infections will result in a positive RDT (if the density of the wild-type strain is sufficiently high), their presence can mask the presence of deletions, resulting in an underestimation of the frequency of deletion (Watson et al., 2019). More recently, quantitative PCR (qPCR) protocols for hrp2/hrp3 deletion typing were published (Grignard et al., 2020; Schindler et al., 2019; Kreidenweiss et al., 2019), greatly enhancing throughput. However, when parasite densities are low, considerable variation in quantification is observed between replicates (Koepfli et al., 2016). While quantification is accurate in the case of clinical samples that are high density, accurate estimates of the population frequency of deletions might require typing of asymptomatic, low-density samples. As a result of the technical challenges for accurate typing, maps of hrp2 deletion frequency remain scattered and incomplete.
We have developed a novel method for the typing of hrp2 and hrp3 deletions based on droplet digital PCR (ddPCR). ddPCR yields highly accurate quantification of parasites (Koepfli et al., 2016). In a ddPCR experiment, the reaction volume is partitioned into approximately 15,000 microdroplets, which are then subject to endpoint PCR. Each droplet function as an individual PCR reaction, with amplification occurring if the droplet contains template DNA. ddPCR offers multiple benefits over qPCR for quantification. Template DNA concentration can be derived directly from the number of positive and negative droplets using Poisson statistics. No external standards are required. Quantification is reliable as long as positive and negative droplets are separated, even if amplification is suboptimal, for example in the case of PCR inhibitors. Amplification efficiency, which is crucial for accurate quantification by qPCR, is thus not relevant in a ddPCR experiment. Using two different probes, two targets can be quantified in a single reaction well, for example a control gene and a target gene. Because the target DNA of each assay is partitioned into separate droplets (with the exception of a few droplets partitioned into the same droplet in case of high-density infections), no competition between assays occurs, thus circumventing a common problem of multiplexed qPCR assays. As each droplet with a template can be considered a ‘within-well replicate’, a single well offers the sensitivity and specificity of a large number of replicates by nPCR or qPCR. The risk of false-negative results (i.e., no amplification when the template is present) is thus minimal compared to nPCR or qPCR. A sample negative for hrp2/3 and the control gene (e.g., due to pipetting error) will not be classified as carrying a deletion and will be excluded from any calculations on deletion frequency in a population. The novel assay greatly reduces the number of reactions to be run compared to gel-based assays, showed high sensitivity and accuracy, and can detect the deletion in mixed infections. The assay was extensively validated using culture strains, and field samples from Kenya, Zanzibar/Tanzania, Ethiopia, Ghana, Brazil, and Ecuador.
Results
Assay development and validation
New primers and probes were developed for ddPCR assays for hrp2 exon 1, hrp2 exon 2, hrp3, and tRNA (Table 1). Upon optimization of the annealing temperature, clear separation between negative and positive droplets was obtained across a wide range of parasite densities. Samples ranging from 1.3 to 49,000 parasites/µl were successfully typed, with samples at densities of >10,000 parasites/µl diluted in H2O. Representative examples are shown in Figure 1, Figure 1—figure supplement 1, and Figure 1—figure supplement 2. No positive droplets for hrp2, hrp3, or hrp2 exon 1 were observed in case of deletion, while the separation between negative droplets and those positive for tRNA remained clear (Figure 1C, Figure 1—figure supplement 1, and Figure 1—figure supplement 2). In case DNA degradation, loss of DNA during extraction, or no DNA added to the reaction (e.g., pipetting error), no signal will be observed for tRNA. In such a case, the sample will be excluded from calculations of hrp2/3 deletion frequency.
Table 1. Primer and probe sequences.
| Assay | Sequence 5′–3′ | |
|---|---|---|
| hrp2 exon 2 forward | CATTTTTAAATGCTTTTTTATTTTTATATAG | |
| hrp2 exon 2 | hrp2 exon 2 reverse | CTTGAGTTTCGTGTAATAATCTC |
| hrp2 exon 2 probe | FAM-CGCATTTAATAATAACTTGTGTAGCAAAAATGC-BHQ-1 | |
| hrp2 exon 1 forward | ATATTTATACATTTTTGTTATTATTTCTTTTTC | |
| hrp2 exon 1 | hrp2 exon 1 reverse | CGTTATCTAACAAAAGTACGGAG |
| hrp2 exon 1 probe | FAM-CAAAAACGGCAGCGGATAATACTT-BHQ-1 | |
| hrp3 forward | ATGCTAATCACGGATTTCATTTTA | |
| hrp3 | hrp3 reverse | ATCGTCATGGTGAGAATCATC |
| hrp3 probe | FAM-CCTTCACGATAACAATTCCCATACTTTAC-BHQ-1 | |
| tRNA forward | CATCAAATGAAGATTTAACAAGAG | |
| tRNA | tRNA reverse | CTTTTTGATTCTATAGTTTCATCTTTATG |
| tRNA probe | HEX-CTACCTCAGAACAACCATTATGTGCT-BHQ-1 |
Figure 1. Examples of hrp2 exon 2 deletion typing by droplet digital PCR (ddPCR).
Droplets positive for hrp2 are shown in blue (top left of each panel). Droplets positive for tRNA are shown in green (bottom right). Droplets positive for hrp2 and tRNA are shown in orange (top right). Negative droplets (for both hrp2 and tRNA) are shown in gray (bottom left). (A) Wild-type infection of medium density (304 parasites/µl, 2 µl DNA used for experiment). Approximately 600 droplets are positive each for hrp2 and tRNA, and 36 for both targets. (B) Wild-type sample of low-density (15 parasites/µl): 32 and 29 droplets are positive for hrp2 and tRNA, respectively. (C) hrp2 deletion (1135 parasites/µl): Droplets are positive for tRNA, but no droplets are positive for hrp2. (D) Mixed infection with wild-type parasites and parasites carrying hrp2 deletion (overall 299 parasites/µl). Only 15 droplets are positive for hrp2, but 598 droplets are positive for tRNA.

Figure 1—figure supplement 1. Examples of hrp3 droplet digital PCR (ddPCR) assays.

Figure 1—figure supplement 2. Examples of hrp2 exon 1 droplet digital PCR (ddPCR) assays.

To evaluate the reproducibility and the limit of reliable detection, 248 samples from asymptomatic carriers in Kenya were typed for the hrp2 exon 2 assay in triplicate. Geometric mean density was 95 parasites/µl, and 47/248 samples were at densities <10 parasites/µl. By ddPCR, 235/248 (94.6%) of samples met inclusion criteria of ≥2 droplets positive for tRNA in all three replicates. Highly similar quantification among replicates was observed (Figure 2A). For each sample, the highest and lowest value of hrp2 copies/µl was recorded. Correlation among technical replicates was very high (n = 235, R2 = 0.990). Likewise, for each sample, the highest and lowest value of tRNA copies was recorded, and correlation was very high (n = 235, R2 = 0.991). The coefficient of variation (CV) across the triplicates was calculated. CV was 17.7% for the hrp2 assay, and 17.1% for the tRNA assay. As no deletions were observed in this sample set, highly similar quantification of hrp2 and tRNA was expected, and CV between hrp2 and tRNA in the same reaction could be calculated. Mean CV between hrp2 and tRNA was 11.8%. CV was lower among samples of a higher density of >100 parasites/µl (n = 126) at 11.5% for hrp2 assay, 10.1% for the tRNA assay, and 4.7% for the within-well hrp2/tRNA comparison (n = 126). The lower CV within wells than among replicates shows that variation in pipetting, resulting slightly different numbers of template added to each replicate reaction, is the main source of variation, while within-well quantification of hrp2 and tRNA is highly accurate.
Figure 2. Validation of assay.
(A) Samples typed in triplicate for the hrp2 exon 2/tRNA assay. Representative examples of different parasite densities are shown. For each sample, the quantification of hrp2 exon 2, and of tRNA is shown. Results from the same run are connected by a dashed line. (B) Mixtures of 3D7 (wild type) and Dd2 (hrp2 deletion). Mixtures were run in triplicate at densities of 20–1000 parasites/reaction, and at ratios of 0–100% Dd2. The expected proportion of hrp2 to tRNA copies corresponds to the proportion of 3D7 in the mixture. The observed proportion reflects the expected proportion closely for all mixtures at 1000 and 100 parasites/reaction.

Figure 2—figure supplement 1. Mixtures of 3D7 (wild type) and 11,140 (hrp3 deletion).

For the hrp3/tRNA assay, CV was calculated among 96 triplicates. CV was 19.5% for the hrp3 assay, and 20.7% for the tRNA assay. Mean CV between hrp3 and tRNA across all three replicates was 16.8%.
No deletions were detected in the 248 samples from western Kenya. That is, no samples with droplets for tRNA but no droplets for hrp2 were observed, even though density was low in many samples. In the ddPCR, approximately two-thirds of the reaction volume was transformed into droplets. Applying the threshold of two droplets positive for tRNA, three template genomes were required per reaction well to reach that threshold. Up to 9 µl of template DNA could be added to one reaction when primers and probes are kept at 100 µM. Thus, the theoretical limit of detection was 0.33 parasites/µl (three templates in 9 µl of DNA, of which 6 µl are partitioned into droplets). If ≤5 droplets are positive for tRNA and a deletion is observed, it is recommended to repeat the sample.
Detection of mixed infections with hrp2- or hrp3-negative and wild-type parasites
To test the ability of the assay to detect deletions when only a proportion of all parasites in an isolate carry the deletion, experimental mixtures were prepared with DNA from parasite culture of 3D7 (wild type) and Dd2 (hrp2 deletion). Mixtures were prepared at ratios from 0% to 100% Dd2, and at densities of 10–500 parasites/µl. At densities of 200–1000 parasites/reaction, the quantification by ddPCR represented the mixture ratio with high accuracy (Figure 2B). Whenever 40% or more of parasites carried the deletion, the observed ratio was clearly below 100%. In cases where only a small proportion of all parasites carried the deletion (20% Dd2 vs. 80% 3D7), the difference in quantification of hrp2 and tRNA was too small to observe the deletion. Likewise, at densities of 20 parasites/reaction the ratio did not accurately reflect the experimental mixture. Similar results were obtained for mixtures of parasites with hrp3 deletion and wild type. At densities >20 parasites/reaction, the mixture was reflected with high accuracy (Figure 2—figure supplement 1).
The ability to detect a minority clone carrying hrp2 at a lower frequency (e.g., at 1%) in a sample dominated by a clone carrying the deletion will depend on the overall parasite density. At a density of 1000 parasites/reaction 10 droplets are expected to be positive for hrp2, thus the wild-type parasite will be detected. At an overall density of 50 parasites/reaction, <1 droplet is expected to be for hrp2, thus the clone will be missed.
The results were corroborated by screening of 739 field samples from five countries. In wild-type samples, a very similar quantification of hrp2 and tRNA was expected. Unless samples carried a clear deletion (i.e., no or very little hrp2 signal), in all samples with densities of >50 copies/µl, the ratio of hrp2 to tRNA copies was above 0.6 (Figure 3). With increasing parasite density, the ratio got closer to 1.
Figure 3. Ratios of hrp2 to tRNA copies in 684 field samples.
With increasing parasite density (x-axis), the ratio becomes close to 1. Deletions (with no wild-type parasites present) have a ratio of 0. Dashed lines show a ratio of hrp2 to tRNA copies of 0.6, and 50 parasites/µl. Mixed infection can be reliably detected at densities >50 parasites/µl, and if >40% of parasites carry the deletion.

Figure 3—figure supplement 1. Ratios of hrp2 to tRNA copies in field samples from Zanzibar (n = 91).
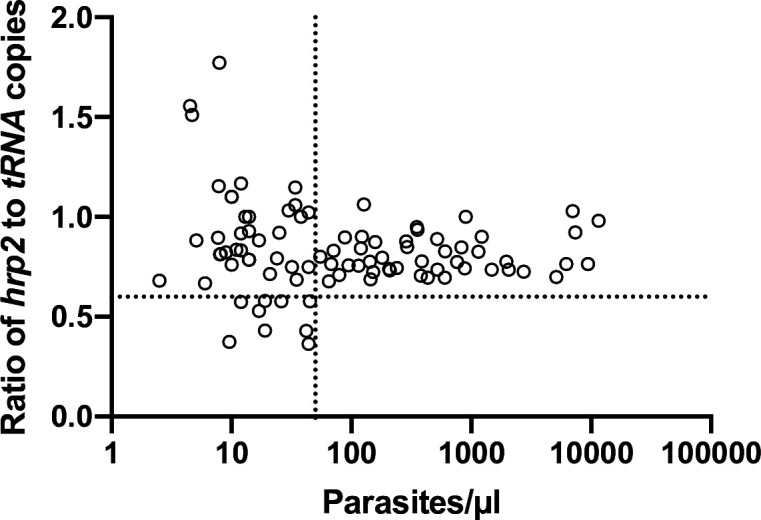
A lower ratio of hrp2 to tRNA copies was observed in samples from Zanzibar, where the mean ratio of hrp2/tRNA copies was 0.82 in absence of any samples that carried hrp2 deletion (Figure 3—figure supplement 1). The effect might be caused by sampling and storage procedures. Blood samples from Zanzibar were collected on filter paper, and stored at ambient temperature for over 3 years prior to extraction. Possibly, this could have resulted in DNA degradation, that affected hrp2 more than tRNA.
Comparison of ddPCR to gel-based nPCR
The hrp2 exon 2 ddPCR assay was compared to the classical nPCR assay with visualization of products on agarose gel in 248 asymptomatic infections from western Kenya. The density of asymptomatic infections is often low and thus amplification by PCR can be stochastic. All assays were run in triplicate, that is hrp2 exon 2/tRNA by ddPCR, hrp2 nPCR, and msp2 nPCR.
Samples were included in the analysis if the PCR for the control gene was positive in all three replicates, that is if >2 droplets were positive for tRNA in the ddPCR assay, or a band was detected for msp2 in all three replicates. For the gel-based assay, 85.5% (212/248) samples met inclusion criteria (Table 2). Among those positive, for 144/212 samples a band for hrp2 was observed in all three replicates. A band in two replicates was observed for 34 samples, and a band in a single replicate for 17 samples. For 17 samples, despite obtaining a band for msp2 in all three replicates, no band for hrp2 was observed. These 17 samples would thus be classified as hrp2 deletion, resulting in a prevalence of deletion of 8.0% (17/212). Figure 4 shows representative examples of results of the nPCR and ddPCR assays.
Table 2. Comparison between nested PCR and droplet digital PCR (ddPCR)-based hrp2 deletion typing.
| Nested PCR | ddPCR | |
|---|---|---|
| Total samples | 248 | 248 |
| Met inclusion criteria | 212 (85.5%) | 235 (94.8%) |
| Deletion in 3/3 replicates | 17 | 0 |
| Deletion in 2/3 replicates | 17 | 0 |
| Deletion in 1/3 replicates | 34 | 2 |
| No deletion | 144 | 233 |
| Prevalence of deletion | 8.0% (17/212) | 0% (0/235) |
Figure 4. Comparison of nested PCR (nPCR) and droplet digital PCR (ddPCR) for hrp2 deletion typing.

Representative examples of results obtained by hrp2 and msp2 nPCR, and by ddPCR. The expected size of the hrp2 band is 228 bp. A band is visible in samples 3, 4, and 5, but no band is visible in samples 1, 2, 6, and 7. msp2 was run as control for the nPCR assay. msp2 is a size polymorphic gene with amplicons ranging from approximately 200–500 bp. Bands are observed for all samples, and multiple bands are observed in case of polyclonal infections. L = 100 bp DNA ladder (New England BioLabs). Samples were run in triplicate, and the same results were obtained all three times. By ddPCR, no deletions were observed in any samples. For samples 1 and 7, the hrp2 exon 2/tRNA ddPCR plot is shown. Droplets are visible for both targets, thus no deletion is observed.
By ddPCR, the criteria for inclusion (≥2 droplets for tRNA) were met by 94.8% (235/248) of samples. Among them, ≥2 droplets for hrp2 were detected in all three replicates in 233/235 samples. In two samples in only two of the three replicates ≥2 droplets were positive for hrp2. Both of these samples had one replicate with 1 positive droplet for hrp2, and five positive droplets for tRNA. In none of the samples all three or two out of three replicates were negative for hrp2. Thus, the prevalence of deletion by ddPCR was 0%.
hrp2 and hrp3 deletions in Africa and South America
The new ddPCR assay was applied to screen for deletions in 830 samples from Kenya, Zanzibar, Ethiopia, Ghana, Brazil, and Ecuador. The frequency of deletions for all loci and sites is given in Table 3.
Table 3. hrp2 and hrp3 deletions in Africa and South America.
| Site | Deletions | |||||
|---|---|---|---|---|---|---|
| N | hrp2 exon 2 | hrp2 exon 1 | hrp3 | hrp2 + hrp3* | Mixed† | |
| Kenya | 241 | 0% (0/241) | 0% (0/241) | 0% (0/241) | 0% (0/241) | |
| Zanzibar | 91 | 0% (0/91) | 0% (0/91) | 0% (0/91) | 0% (0/91) | |
| Ethiopia | 47 | 2.1% (1/47) | 2.1% (1/47) | 74.5% (35/47) | 2.1% (1/47) | 1× hrp3 |
| Ghana | 223 | 0% (0/226) | 0% (0/223) | 0.4% (1/223) | 0% (0/170) | 1× hrp2, 3× hrp3 |
| Brazil | 187 | 46.5% (87/187) | NA | 62.0% (116/187) | 46.0% (86/187) | 2× hrp2, 2× hrp3 |
| Ecuador | 41 | 0% (0/41) | 0% (0/41) | 53.7% (22/41) | 0% (0/39) | |
Samples with deletions of hrp2 and hrp3. Note that these samples are also counted as deletions in the columns for hrp2, and for hrp3 (e.g., in Brazil 87 samples carried hrp2 deletion, of which 86 also carried hrp3 deletion).
Samples with only a proportion of all parasites carrying the deletion.
From Kenya, 241 samples were typed and no deletions of hrp2 or hrp3 were observed. In Zanzibar, no deletions were observed among 91 samples. Among 223 samples from Ghana, 1 hrp2 deletion/wild-type mixed sample was detected (ratio of hrp2 to tRNA copies was <0.6, and parasite density >50 parasites/µl), 1 hrp3 deletion, and 3 samples with hrp3 deletion/wild-type mixes.
In Ethiopia, 47 samples met inclusion criteria. One sample carried a deletion of hrp2 exons 1 and 2, resulting in a frequency of deletion of 2.1%. hrp3 deletion was observed in 35/47 (74.5%) samples, and one sample carried a mixed infection with wild type/hrp3 deletion. The sample with hrp2 deletion was among the samples with hrp3 deletion, that is both genes were deleted.
From Brazil, 187 samples were screened. Eighty-seven samples carried a deletion of hrp2. Two additional samples carried mixed infections with wild-type parasites and hrp2 deletion. Deletion of hrp3 was observed in 116/187 (62.0%) samples, and 86/187 (46.0%) samples carried deletions of hrp2 and hrp3. No hrp2 deletions were observed in Ecuador, but 22/41 (53.7%) samples carried hrp3 deletions.
Discussion
Molecular surveillance of the extent of hrp2 and hrp3 deletions is a high priority task to select the optimal tools for P. falciparum diagnosis. The novel assay based on ddPCR yielded highly accurate results, and was able to detect mixed infections with wild-type parasites and parasites carrying hrp2 deletion. The good performance of the assay was shown by typing samples from six countries. The samples reflected a range of parasite densities, from low-density asymptomatic infections to high-density clinical infections, and sites represented a range in the frequency of hrp2 and/or hrp3 deletions. Across over 800 field samples screened, <10 samples needed to be repeated because of poor separation between negative and positive droplets, or failure of droplet generation.
The assay can be used to screen samples for P. falciparum positivity based on the result for tRNA, and hrp2 or hrp3 deletion in a single assay. Alternatively, it can be used to type samples prior determined P. falciparum positive by microscopy or another molecular assay. High sensitivity is required for accurate typing of low-density infections. We validated a threshold of two droplets positive for tRNA as the limit of detection. When 9 µl of DNA are used as template, this results in a theoretical limit of detection of 0.33 parasites/µl. Sensitivity can be further increased by concentrating the DNA prior to typing.
The ddPCR assay showed increased sensitivity and specificity to type low-density infections compared to nPCR. Out of 248 asymptomatic samples from western Kenya, 95% could be analyzed by ddPCR, but only 85% by nPCR. More importantly, results on deletion status differed substantially. By ddPCR, no deletions were observed. By nPCR, no band was observed for hrp2 in 8% of samples that had a positive band for the control gene (msp2) in all three replicates. Thus, the frequency of deletion was above the 5% threshold, and it would be erroneously recommended to discontinue the use of HRP2-based RDTs. The frequency of deletion by nPCR was similar to a previous study conducted in a nearby site in western Kenya that found hrp2 deletion in 8/89 samples using nPCR (Beshir et al., 2017). The direct comparison of ddPCR and nPCR in a large number of samples points to a possible overestimation of hrp2 deletion frequency by studies relying on nPCR. Further systematic comparison with qPCR-based assays (Grignard et al., 2020; Kreidenweiss et al., 2019; Schindler et al., 2019) will be required to determine the sensitivity and specificity of each assay.
In almost all transmission settings, polyclonal P. falciparum infections are frequent (Lopez and Koepfli, 2021). Using a gel-based assay for deletion typing, in a mixed infection with a wild-type parasite and a parasite carrying a deletion, the wild-type parasite will produce a band and mask the deletion. These infections can be detected by RDT, but depending on the proportion of polyclonal infections, they can result in pronounced underestimation of the true frequency of deletion (Watson et al., 2019). The highly accurate quantification by ddPCR allows detection of mixed infections. Based on experimental mixtures of 3D7 and Dd2, and field samples, mixed infections were reliably detected when at least 40% of parasites carried the deletion, and at densities above 100 parasites per reaction. Using a well-working qPCR assay with an amplification efficacy of 100%, a similar difference between hrp2 and tRNA copy numbers would result in less than half a cycle difference. This is within the normal variation of technical replicates (Koepfli et al., 2016; Hindson et al., 2013), and thus such mixed infections could not be detected by qPCR.
Twenty-nine samples from Zanzibar and 16 from Ghana had tested negative by HRP2-based RDT despite high density of 100 to >10,000 parasites/µl (Stuck et al., 2020), yet no hrp2 deletions were observed in these populations. False-negative RDT results might be caused due to incorrect handling, prozone effect (Gillet et al., 2009), or sequence variation without deletion of the hrp2 gene (Nderu et al., 2019). The finding corroborates the importance of molecular typing. Studies comparing microscopy and RDT results can give important clues for the presence of deletions (Berhane et al., 2017), but molecular typing is required for confirmation (Berhane et al., 2018).
Though not reported in the literature, deletion of hrp2 exon 1 only, but not exon 2, could result in a lack of expression of HRP2. In this scenario, the hrp2 exon 2 assay would not show any deletion, yet HRP2-based RDTs would show a false-negative result. In this study, no discrepancy was observed between deletion status for hrp2 exons 1 and 2. Thus, for future surveillance, it is recommended to only type samples for hrp2 exon 2 and hrp3. In case of negative RDTs despite no hrp2 exon 2 deletion, typing for exon 1 can be done.
No hrp2 deletions were found in Zanzibar, and Kenya, and one mixed infection in Ghana. The data from Ghana contrasts an earlier study that reported a frequency of deletion of >30% (Amoah et al., 2016). A single hrp2 deletion was found in southwestern Ethiopia. This is in stark contrast to very high levels of deletion in western Ethiopia (Alemayehu et al., 2021), Eritrea (Berhane et al., 2018), and Sudan (Prosser et al., 2021). The results corroborate the need for studies assessing hrp2 deletion and selection of diagnostic tools at subnational level.
Contrasting findings were obtained from the sample sets from South America, with very high levels of deletion in Brazil, and no deletions in Ecuador. Brazil and Ecuador share no borders, and the amount of human migration is limited. The results add to the heterogeneous pattern of hrp2/hrp3 deletion in South America, with high frequency of hrp2 deletion in the Amazon (Gamboa et al., 2010; Murillo Solano et al., 2015; Góes et al., 2020), but low levels among the Pacific coast (Sáenz et al., 2015; Murillo Solano et al., 2015). An outbreak of parasites with hrp2 deletion at the Peruvian Pacific coast was caused by infections imported from the Amazon (Baldeviano et al., 2015). Brazil is committed to P. falciparum elimination, and the samples typed originated from the main hotspot of transmission (Ferreira and Castro, 2016). RDTs remain relatively little used in Brazil. Microscopy remains the diagnostic method of choice, and RDTs are mostly used in remote areas, for example, populations from Amerindian Reserves and some traditional riverine communities with no access to conventional microscopy. The frequency of hrp2 deletion in Brazil clearly exceeds the 5% threshold, thus it is recommended that no HRP2-based RDTs are used.
To determine whether the frequency of the deletion exceeds the threshold of 5%, the WHO recommends systematic surveillance and typing of a minimum of 370 per site (World Health Organization, 2020). Limitations of the molecular assays available for typing have been a major hindrance to type that number of samples in many sites where hrp2 or hrp3 deletion is suspected. Even a low proportion of false-negative results could impact the decision to discontinue HRP2-based RDTs. As a result of the scarcity of field data, the spatiotemporal dynamics of hrp2 deletion in parasite populations, drivers of the deletion, potential fitness costs, and clinical consequences remain poorly understood. Results from simulation studies suggest that the use of hrp2-based RDTs selects for hrp2-negative parasites, in particular, if transmission levels are low and a large proportion of all infections turn clinical and result in treatment seeking (Gatton et al., 2017; Watson et al., 2017). While digital PCR is less common than nPCR or qPCR, it has been successfully established in malaria-endemic countries. The assay is run in 96-well format; thus throughput is comparable to qPCR, and assay setup is similarly straightforward. While costs of digital PCR instruments and reagents are moderately higher than for qPCR, they have declined recently.
In conclusion, the novel, high-throughput, highly sensitive and specific ddPCR assay will facilitate molecular surveillance for hrp2 and hrp3 deletion, and thus aid the selection of diagnostic tests to accelerate malaria control and elimination. Data obtained using this assay will help to understand the evolutionary processes underlying the de novo emergence and spread of the deletion.
Materials and methods
Digital PCR assays
Four novel ddPCR assays were developed. One assay targets the conserved first 120 bp of hrp2 exon 2, and thus is located directly adjacent to the histidine-rich repeats. Different breakpoints for the hrp2 deletion have been described (Cheng et al., 2014). The novel primers for exon 2 are located in a region that is deleted in all known deletion variants, thus irrespective of the specific breakpoint, hrp2 deletion will be detected. The second assay targets hrp2 exon 1. While exon 1 does not contain antigens that are recognized by RDTs, the deletion of this exon 1 would prevent proper expression of the protein. The third assay targets hrp3. The assay amplifies a 101-bp segment in the center of exon 2. Each assay was multiplexed with an assay targeting serine-tRNA ligase (PF3D7_0717700, herein referred to as ‘tRNA’). tRNA is a conserved, essential single copy gene, that is frequently used as reference for gene expression assays (Friedrich et al., 2014). In wild-type infections not carrying a deletion, the copy numbers of tRNA and hrp2 or hrp3 are identical.
Novel primers and probes were developed for all assays (Table 1). Assay conditions are given in Supplementary file 1. Across over >3000 genomes available through MalariaGen and PlasmoDB, no SNPs were recorded in primer and probe sequences, thus the assay can be used for the screening of samples of global origin. Assay conditions were optimized to achieve maximal separation between positive and negative droplets (Figure 1, Figure 1—figure supplement 1, Figure 1—figure supplement 2).
3D7 and Dd2 parasite culture strain mixtures
In order to determine the ability to detect mixed clone infections with only one strain carrying the deletion, experimental mixtures were made from two well-characterized laboratory strains, 3D7 (no deletions), and Dd2 (carrying the hrp2 deletion). Each strain was cultured separately, DNA extracted, and quantified by ddPCR using the hrp2/tRNA ligase assay. No hrp2 was detected in Dd2. Mixtures were prepared with a concentration (of both strains combined) of 10, 50, 100, and 500 parasites/µl, and with a 3D7 to Dd2 ratio of 100:0, 80:20, 60:40, 40:60, 20:80, and 0:100. Each mixture was run in triplicate using the hrp2 exon 2/tRNA ligase assay. The ratio of hrp2 to tRNA copy numbers was compared to expected values.
nPCR assay for hrp2
In order to compare the ddPCR assay directly to the established nPCR assay, 248 samples from asymptomatic carriers in western Kenya were run by the hrp2 exon 2 assay and the hrp2 nPCR in triplicate. Assay conditions for the nPCR followed published protocols (Akinyi et al., 2013; Abdallah et al., 2015) and are given in Supplementary file 1.
Field samples
Field samples were screened from Kenya, Zanzibar in the United Republic of Tanzania, Ethiopia, Ghana, Brazil, and Ecuador (summarized in Table 4). Samples were determined P. falciparum positive either by microscopy (Brazil, Ecuador), 18S qPCR (Rosanas-Urgell et al., 2010 Ethiopia), or varATS qPCR (Hofmann et al., 2015; Zanzibar, Kenya, Ghana).
Table 4. Field samples included in this study.
| Site | N | Year of collection | Type of diagnosis* | Type of infection | Sample collection method |
|---|---|---|---|---|---|
| Kenya | 248 | 2019 | qPCR | Asymptomatic | Whole blood |
| Zanzibar/Tanzania | 91 | 2017–2018 | qPCR | Asymptomatic | Filter paper |
| Ethiopia | 47 | 2016 | qPCR | Clinical + asymptomatic | Filter paper |
| Ghana | 213 | 2020 | qPCR | Clinical | Whole blood |
| Brazil | 187 | 2010–2013 | Microscopy | Clinical | Whole blood |
| Ecuador | 41 | 2019–2020 | Microscopy | Clinical | Filter paper |
qPCR, quantitative PCR.
While samples might have been screened by other diagnostic methods, the screening listed was used as inclusion criteria for this study.
Samples from Kenya (n = 248) were from Chulaimbo and Homa Bay in western Kenya close to Lake Victoria. Samples were collected in a cross-sectional survey including individuals of all ages from January to August 2019. Finger-prick samples were collected in EDTA microtainers and infections detected by qPCR. Overall P. falciparum prevalence across both sites was 16% (Oduma et al., 2021). No diagnosis by RDT or microscopy was done.
Samples from Zanzibar (n = 91) had been collected in the frame of a study on RCD from May 2017 to October 2018 (Stuck et al., 2020). Asymptomatic infections identified through RCD were typed. During RCD, infections were diagnosed by HRP2/pLDH-based RDT (SD BIOLINE Malaria Ag Pf HRP2/pLHD), a blood spot was collected on filter paper, and infections diagnosed by qPCR (Stuck et al., 2020). Prevalence (not including index cases) was 0.8% by RDT and 2.4% by qPCR. All samples with a density (by qPCR) of >100 parasites/µl were selected for hrp2/hrp3 deletion typing, irrespective of RDT result.
Samples from Ethiopia (n = 47) included clinical cases and asymptomatic individuals sampled in June to November 2016 from Jimma Zone, Oromia Region. P. falciparum prevalence was 4% by microscopy and 8.3% by qPCR (Zemene et al., 2018). No RDT screening was conducted.
From Ghana, two sets of samples were typed. The first set (n = 11) was collected in June–September of 2017 from febrile school children aged 5–14 years (Dieng et al., 2019). The second set (n = 212) was collected in Mankranso and Agona Hospitals in the Ashanti region from febrile patients from September to December 2020.
In Brazil, samples (n = 187) were collected in Cruzeiro do Sul, Upper Juruá Valley, northwestern Brazil, in 2010–2013. This is the country’s main malaria hotspot, which accounted for nearly 15% of all P. falciparum infections in Brazil at the time of the study. Samples were collected from clinical patients 4–73 years of age (mean, 26.6) enrolled for a drug efficacy trial (Ladeia-Andrade et al., 2016). Only baseline samples from patients with microscopy- and PCR-confirmed P. falciparum infection were included in this study. Even though these samples had been collected nearly a decade ago and might not reflect the current status of hrp2/hrp3 deletion, they were included in light of the known high levels of hrp2/hrp3 deletion in Latin America to confirm the ability of the assays to reliably detect deletions in field samples (Góes et al., 2020). Because of little template volume available, Brazilian samples were not screened for hrp2 exon 1 deletion.
In Ecuador, samples (n = 41) were collected from clinical patients from March 2019 to April 2020. The samples were collected in Esmeraldas and Carchi Provinces in the north-west of the country, where most P. falciparum cases of Ecuador are reported. All infections were confirmed by microscopy and collected as blood spots in filter paper. Three samples were collected from travelers coming from the Pacific coast in Colombia but diagnosed in Ecuador.
Data are reported as parasites/µl, representing the number of P. falciparum genomes per µl eluted DNA. Samples collection and DNA extraction procedures are different among studies (e.g., whole blood vs. filter paper collected, different kits used for extraction). As a result, the conversion from parasites per µl blood to genomes per µl eluted DNA differs among sample sets. For the present study, these differences are not relevant, as for the validation of PCR assays, the number of genomes per µl input DNA is crucial.
Data analysis
For the analysis of the ddPCR data, the following criteria were applied: A minimum of two droplets positive for tRNA were required to include a sample in data analysis. Samples were repeated if a deletion was observed but ≤5 droplets were positive for tRNA. If >5 droplets are positive for tRNA, the probability of a false-negative result for hrp2 or hrp3 (i.e., no positive droplet in a wild-type infection) is less than 1:500. Data for all samples and assays are provided in Supplementary file 2. The CV of samples run in replicates, and of different targets within the same tube, was calculated as the standard deviation divided by the mean.
Acknowledgements
We thank all study participants providing blood samples and the study teams and health center personnel who supported sample collection. We thank Michael T Ferdig and Katelyn M Vendrely for providing culture strain DNA for the 3D7/Dd2 experimental mixtures. Financial Disclosure Statement: This work was supported by NIH grants R21AI137891 awarded to CK, and U19 AI129326, D43 TW001505 awarded to GY (https://www.nih.gov/). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Funding Statement
The funders had no role in study design, data collection, and interpretation, or the decision to submit the work for publication.
Contributor Information
Cristian Koepfli, Email: ckoepfli@nd.edu.
Bavesh D Kana, University of the Witwatersrand, South Africa.
Bavesh D Kana, University of the Witwatersrand, South Africa.
Funding Information
This paper was supported by the following grants:
National Institutes of Health R21 AI137891 to Cristian Koepfli.
National Institutes of Health D43 TW001505 to Guiyun Yan.
National Institutes of Health U19 AI129326 to Guiyun Yan.
Additional information
Competing interests
No competing interests declared.
Author contributions
Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Investigation, Writing - review and editing.
Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing - original draft, Writing - review and editing.
Ethics
Informed written consent was obtained from all study participants or their parents or legal guardians prior to sample collection. The study was approved by the University of Notre Dame Institutional Review Board (approvals 18-08-4803, 19-04-5321, and 18-12-5029), the Institutional Scientific and Ethical Review boards of the Noguchi Memorial Institute of Medical Research, University of Ghana, the Committee on Human Research, Publication and Ethics, School of Medical Science, Kwame Nkrumah University of Science and Technology, Kumasi (CHRPE/AP/375/20), the Zanzibar Medical Research Ethics Committee (ZAMREC/0001/Feb/17), the Institutional Review Board of Tulane University (17-993573), the Institutional Review Board of the Ifakara Health Institute (003-2017), the Ethics Commission of North-western and Central Switzerland (Req-2017-00162), the Institutional Review Board of Institute of Health, Jimma University, Ethiopia (RPGC/486/06), Maseno University Ethics Review Committee (MUERC protocol number 00456), the Ethics Committee for Research in Human Beings of the Pontificia Universidad Católica del Ecuador (CEISH-571-2018), the Ministry of Public Health of Ecuador (MSP-DIS-2019-004-O), and the institutional review board of Oswaldo Cruz Foundation, Brazil (no. 022/2009).
Additional files
Data availability
All data are provided in supplementary file S3.
References
- Abdallah JF, Okoth SA, Fontecha GA, Torres REM, Banegas EI, Matute ML, Bucheli STM, Goldman IF, de Oliveira AM, Barnwell JW, Udhayakumar V. Prevalence of pfhrp2 and pfhrp3 gene deletions in Puerto Lempira, Honduras. Malaria Journal. 2015;14:19. doi: 10.1186/s12936-014-0537-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Acquah FK, Donu D, Obboh EK, Bredu D, Mawuli B, Amponsah JA, Quartey J, Amoah LE. Diagnostic performance of an ultrasensitive HRP2-based malaria rapid diagnostic test kit used in surveys of afebrile people living in Southern Ghana. Malaria Journal. 2021;20:125. doi: 10.1186/s12936-021-03665-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akinyi S, Hayden T, Gamboa D, Torres K, Bendezu J, Abdallah JF, Griffing SM, Quezada WM, Arrospide N, De Oliveira AM, Lucas C, Magill AJ, Bacon DJ, Barnwell JW, Udhayakumar V. Multiple genetic origins of histidine-rich protein 2 gene deletion in Plasmodium falciparum parasites from Peru. Scientific Reports. 2013;3:2797. doi: 10.1038/srep02797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akinyi Okoth S, Abdallah JF, Ceron N, Adhin MR, Chandrabose J, Krishnalall K, Huber CS, Goldman IF, Macedo de Oliveira A, Barnwell JW, Udhayakumar V. Variation in Plasmodium falciparum Histidine-Rich Protein 2 (Pfhrp2) and Plasmodium falciparum Histidine-Rich Protein 3 (Pfhrp3) Gene Deletions in Guyana and Suriname. PLOS ONE. 2015;10:e0126805. doi: 10.1371/journal.pone.0126805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alemayehu GS, Blackburn K, Lopez K, Cambel Dieng C, Lo E, Janies D, Golassa L. Detection of high prevalence of Plasmodium falciparum histidine-rich protein 2/3 gene deletions in Assosa zone, Ethiopia: implication for malaria diagnosis. Malaria Journal. 2021;20:109. doi: 10.1186/s12936-021-03629-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amoah LE, Abankwa J, Oppong A. Plasmodium falciparum histidine rich protein-2 diversity and the implications for PfHRP 2: based malaria rapid diagnostic tests in Ghana. Malaria Journal. 2016;15:101. doi: 10.1186/s12936-016-1159-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baldeviano GC, Okoth SA, Arrospide N, Gonzalez RV, Sánchez JF, Macedo S, Conde S, Tapia LL, Salas C, Gamboa D, Herrera Y, Edgel KA, Udhayakumar V, Lescano AG. Molecular Epidemiology of Plasmodium falciparum Malaria Outbreak, Tumbes, Peru, 2010-2012. Emerging Infectious Diseases. 2015;21:797–803. doi: 10.3201/eid2105.141427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berhane A, Russom M, Bahta I, Hagos F, Ghirmai M, Uqubay S. Rapid diagnostic tests failing to detect Plasmodium falciparum infections in Eritrea: an investigation of reported false negative RDT results. Malaria Journal. 2017;16:105. doi: 10.1186/s12936-017-1752-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berhane A, Anderson K, Mihreteab S, Gresty K, Rogier E, Mohamed S, Hagos F, Embaye G, Chinorumba A, Zehaie A, Dowd S, Waters NC, Gatton ML, Udhayakumar V, Cheng Q, Cunningham J. Major Threat to Malaria Control Programs by Plasmodium falciparum Lacking Histidine-Rich Protein 2, Eritrea. Emerging Infectious Diseases. 2018;24:462–470. doi: 10.3201/eid2403.171723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beshir KB, Sepúlveda N, Bharmal J, Robinson A, Mwanguzi J, Busula AO, de Boer JG, Sutherland C, Cunningham J, Hopkins H. Plasmodium falciparum parasites with histidine-rich protein 2 (pfhrp2) and pfhrp3 gene deletions in two endemic regions of Kenya. Scientific Reports. 2017;7:14718. doi: 10.1038/s41598-017-15031-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bharti PK, Chandel HS, Ahmad A, Krishna S, Udhayakumar V, Singh N. Prevalence of pfhrp2 and/or pfhrp3 Gene Deletion in Plasmodium falciparum Population in Eight Highly Endemic States in India. PLOS ONE. 2016;11:e0157949. doi: 10.1371/journal.pone.0157949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng Q, Gatton ML, Barnwell J, Chiodini P, McCarthy J, Bell D, Cunningham J. Plasmodium falciparum parasites lacking histidine-rich protein 2 and 3: a review and recommendations for accurate reporting. Malaria Journal. 2014;13:283. doi: 10.1186/1475-2875-13-283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dieng CC, Gonzalez L, Pestana K, Dhikrullahi SB, Amoah LE, Afrane YA, Lo E. Contrasting Asymptomatic and Drug Resistance Gene Prevalence of Plasmodium falciparum in Ghana: Implications on Seasonal Malaria Chemoprevention. Genes. 2019;10:E538. doi: 10.3390/genes10070538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dorado EJ, Okoth SA, Montenegro LM, Diaz G, Barnwell JW, Udhayakumar V, Murillo Solano C. Genetic Characterisation of Plasmodium falciparum Isolates with Deletion of the pfhrp2 and/or pfhrp3 Genes in Colombia: The Amazon Region, a Challenge for Malaria Diagnosis and Control. PLOS ONE. 2016;11:e0163137. doi: 10.1371/journal.pone.0163137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferreira MU, Castro MC. Challenges for malaria elimination in Brazil. Malaria Journal. 2016;15:284. doi: 10.1186/s12936-016-1335-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedrich O, Reiling SJ, Wunderlich J, Rohrbach P. Assessment of Plasmodium falciparum PfMDR1 transport rates using Fluo-4. Journal of Cellular and Molecular Medicine. 2014;18:1851–1862. doi: 10.1111/jcmm.12313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gamboa D, Ho M-F, Bendezu J, Torres K, Chiodini PL, Barnwell JW, Incardona S, Perkins M, Bell D, McCarthy J, Cheng Q. A large proportion of P. falciparum isolates in the Amazon region of Peru lack pfhrp2 and pfhrp3: implications for malaria rapid diagnostic tests. PLOS ONE. 2010;5:e8091. doi: 10.1371/journal.pone.0008091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gatton ML, Dunn J, Chaudhry A, Ciketic S, Cunningham J, Cheng Q. Implications of Parasites Lacking Plasmodium falciparum Histidine-Rich Protein 2 on Malaria Morbidity and Control When Rapid Diagnostic Tests Are Used for Diagnosis. The Journal of Infectious Diseases. 2017;215:1156–1166. doi: 10.1093/infdis/jix094. [DOI] [PubMed] [Google Scholar]
- Gillet P, Mori M, Van Esbroeck M, Van den Ende J, Jacobs J. Assessment of the prozone effect in malaria rapid diagnostic tests. Malaria Journal. 2009;8:271. doi: 10.1186/1475-2875-8-271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Góes L, Chamma-Siqueira N, Peres JM, Nascimento JM, Valle S, Arcanjo AR, Lacerda M, Blume L, Póvoa M, Viana G. Evaluation of Histidine-Rich Proteins 2 and 3 Gene Deletions in Plasmodium falciparum in Endemic Areas of the Brazilian Amazon. International Journal of Environmental Research and Public Health. 2020;18:E123. doi: 10.3390/ijerph18010123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grignard L, Nolder D, Sepúlveda N, Berhane A, Mihreteab S, Kaaya R, Phelan J, Moser K, van Schalkwyk DA, Campino S, Parr JB, Juliano JJ, Chiodini P, Cunningham J, Sutherland CJ, Drakeley C, Beshir KB. A novel multiplex qPCR assay for detection of Plasmodium falciparum with histidine-rich protein 2 and 3 (pfhrp2 and pfhrp3) deletions in polyclonal infections. EBioMedicine. 2020;55:e102757. doi: 10.1016/j.ebiom.2020.102757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hindson CM, Chevillet JR, Briggs HA, Gallichotte EN, Ruf IK, Hindson BJ, Vessella RL, Tewari M. Absolute quantification by droplet digital PCR versus analog real-time PCR. Nature Methods. 2013;10:1003–1005. doi: 10.1038/nmeth.2633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hofmann N, Mwingira F, Shekalaghe S, Robinson LJ, Mueller I, Felger I. Ultra-sensitive detection of Plasmodium falciparum by amplification of multi-copy subtelomeric targets. PLOS Medicine. 2015;12:e1001788. doi: 10.1371/journal.pmed.1001788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hofmann NE, Antunes Moniz C, Holzschuh A, Keitel K, Boillat-Blanco N, Kagoro F, Samaka J, Mbarack Z, Ding XC, González IJ, Genton B, D’Acremont V, Felger I. Diagnostic Performance of Conventional and Ultrasensitive Rapid Diagnostic Tests for Malaria in Febrile Outpatients in Tanzania. The Journal of Infectious Diseases. 2019;219:1490–1498. doi: 10.1093/infdis/jiy676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johora FT, Hossain ME, Alam MS, Rahman R, Kibria MG, Nima MK, Haque R, Mohon AN, Hougard T. Case Report: A Case of Plasmodium falciparum hrp2 and hrp3 Gene Mutation in Bangladesh. The American Journal of Tropical Medicine and Hygiene. 2017;97:1155–1158. doi: 10.4269/ajtmh.16-0884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koepfli C, Nguitragool W, Hofmann NE, Robinson LJ, Ome-Kaius M, Sattabongkot J, Felger I, Mueller I. Sensitive and accurate quantification of human malaria parasites using droplet digital PCR (ddPCR. Scientific Reports. 2016;6:39183. doi: 10.1038/srep39183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koita OA, Doumbo OK, Ouattara A, Tall LK, Konaré A, Diakité M, Diallo M, Sagara I, Masinde GL, Doumbo SN, Dolo A, Tounkara A, Traoré I, Krogstad DJ. False-negative rapid diagnostic tests for malaria and deletion of the histidine-rich repeat region of the hrp2 gene. The American Journal of Tropical Medicine and Hygiene. 2012;86:194–198. doi: 10.4269/ajtmh.2012.10-0665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kong A, Wilson SA, Ah Y, Nace D, Rogier E, Aidoo M. HRP2 and HRP3 cross-reactivity and implications for HRP2-based RDT use in regions with Plasmodium falciparum HRP2 gene deletions. Malaria Journal. 2021;20:207. doi: 10.1186/s12936-021-03739-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreidenweiss A, Trauner F, Rodi M, Koehne E, Held J, Wyndorps L, Manouana GP, McCall M, Adegnika AA, Lalremruata A, Kremsner PG, Fendel R, Sandri TL. Monitoring the threatened utility of malaria rapid diagnostic tests by novel high-throughput detection of Plasmodium falciparum hrp2 and hrp3 deletions: A cross-sectional, diagnostic accuracy study. EBioMedicine. 2019;50:14–22. doi: 10.1016/j.ebiom.2019.10.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar N, Pande V, Bhatt RM, Shah NK, Mishra N, Srivastava B, Valecha N, Anvikar AR. Genetic deletion of HRP2 and HRP3 in Indian Plasmodium falciparum population and false negative malaria rapid diagnostic test. Acta Tropica. 2013;125:119–121. doi: 10.1016/j.actatropica.2012.09.015. [DOI] [PubMed] [Google Scholar]
- Ladeia-Andrade S, de Melo GNP, de Souza-Lima R, Salla LC, Bastos MS, Rodrigues PT, Luz FC, Ferreira MU. No Clinical or Molecular Evidence of Plasmodium falciparum Resistance to Artesunate-Mefloquine in Northwestern Brazil. The American Journal of Tropical Medicine and Hygiene. 2016;95:148–154. doi: 10.4269/ajtmh.16-0017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li P, Xing H, Zhao Z, Yang Z, Cao Y, Li W, Yan G, Sattabongkot J, Cui L, Fan Q. Genetic diversity of Plasmodium falciparum histidine-rich protein 2 in the China-Myanmar border area. Acta Tropica. 2015;152:26–31. doi: 10.1016/j.actatropica.2015.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li B, Sun Z, Li X, Li X, Wang H, Chen W, Chen P, Qiao M, Mao Y. Performance of pfHRP2 versus pLDH antigen rapid diagnostic tests for the detection of Plasmodium falciparum: a systematic review and meta-analysis. Archives of Medical Science. 2017;13:541–549. doi: 10.5114/aoms.2017.67279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez L, Koepfli C. Systematic review of Plasmodium falciparum and Plasmodium vivax polyclonal infections: Impact of prevalence, study population characteristics, and laboratory procedures. PLOS ONE. 2021;16:e0249382. doi: 10.1371/journal.pone.0249382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maltha J, Gamboa D, Bendezu J, Sanchez L, Cnops L, Gillet P, Jacobs J. Rapid diagnostic tests for malaria diagnosis in the Peruvian Amazon: impact of pfhrp2 gene deletions and cross-reactions. PLOS ONE. 2012;7:e43094. doi: 10.1371/journal.pone.0043094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menegon M, L’Episcopia M, Nurahmed AM, Talha AA, Nour BYM, Severini C. Identification of Plasmodium falciparum isolates lacking histidine-rich protein 2 and 3 in Eritrea. Infection, Genetics and Evolution. 2017;55:131–134. doi: 10.1016/j.meegid.2017.09.004. [DOI] [PubMed] [Google Scholar]
- Murillo Solano C, Akinyi Okoth S, Abdallah JF, Pava Z, Dorado E, Incardona S, Huber CS, Macedo de Oliveira A, Bell D, Udhayakumar V, Barnwell JW. Deletion of Plasmodium falciparum Histidine-Rich Protein 2 (pfhrp2) and Histidine-Rich Protein 3 (pfhrp3) Genes in Colombian Parasites. PLOS ONE. 2015;10:e0131576. doi: 10.1371/journal.pone.0131576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nderu D, Kimani F, Thiong’o K, Karanja E, Akinyi M, Too E, Chege W, Nambati E, Meyer CG, Velavan TP. Plasmodium falciparum histidine-rich protein (PfHRP2 and 3) diversity in Western and Coastal Kenya. Scientific Reports. 2019;9:1709. doi: 10.1038/s41598-018-38175-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oduma CO, Ogolla S, Atieli H, Ondigo BN, Lee MC, Githeko AK, Dent AE, Kazura JW, Yan G, Koepfli C. Increased investment in gametocytes in asymptomatic Plasmodium falciparum infections in the wet season. BMC Infectious Diseases. 2021;21:44. doi: 10.1186/s12879-020-05761-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parr JB, Verity R, Doctor SM, Janko M, Carey-Ewend K, Turman BJ, Keeler C, Slater HC, Whitesell AN, Mwandagalirwa K, Ghani AC, Likwela JL, Tshefu AK, Emch M, Juliano JJ, Meshnick SR. Pfhrp2-Deleted Plasmodium falciparum Parasites in the Democratic Republic of the Congo: A National Cross-sectional Survey. The Journal of Infectious Diseases. 2017;216:36–44. doi: 10.1093/infdis/jiw538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pati P, Dhangadamajhi G, Bal M, Ranjit M. High proportions of pfhrp2 gene deletion and performance of HRP2-based rapid diagnostic test in Plasmodium falciparum field isolates of Odisha. Malaria Journal. 2018;17:394. doi: 10.1186/s12936-018-2502-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prosser C, Gresty K, Ellis J, Meyer W, Anderson K, Lee R, Cheng Q. Plasmodium falciparum Histidine-Rich Protein 2 and 3 Gene Deletions in Strains from Nigeria, Sudan, and South Sudan. Emerging Infectious Diseases. 2021;27:471–479. doi: 10.3201/eid2702.191410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosanas-Urgell A, Mueller D, Betuela I, Barnadas C, Iga J, Zimmerman PA, del Portillo HA, Siba P, Mueller I, Felger I. Comparison of diagnostic methods for the detection and quantification of the four sympatric Plasmodium species in field samples from Papua New Guinea. Malaria Journal. 2010;9:361. doi: 10.1186/1475-2875-9-361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sáenz FE, Morton LC, Okoth SA, Valenzuela G, Vera-Arias CA, Vélez-Álvarez E, Lucchi NW, Castro LE, Udhayakumar V. Clonal population expansion in an outbreak of Plasmodium falciparum on the northwest coast of Ecuador. Malaria Journal. 2015;13 Suppl 1:497. doi: 10.1186/s12936-015-1019-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schindler T, Deal AC, Fink M, Guirou E, Moser KA, Mwakasungula SM, Mihayo MG, Jongo SA, Chaki PP, Abdulla S, Valverde PCM, Torres K, Bijeri JR, Silva JC, Hoffman SL, Gamboa D, Tanner M, Daubenberger C. A multiplex qPCR approach for detection of pfhrp2 and pfhrp3 gene deletions in multiple strain infections of Plasmodium falciparum. Scientific Reports. 2019;9:13107. doi: 10.1038/s41598-019-49389-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stuck L, Fakih BS, Al-Mafazy AWH, Hofmann NE, Holzschuh A, Grossenbacher B, Bennett A, Cotter C, Reaves E, Ali A, der Horst T, Felger I, Hetzel MW, Yukich J. Malaria infection prevalence and sensitivity of reactive case detection in Zanzibar. International Journal of Infectious Diseases. 2020;97:337–346. doi: 10.1016/j.ijid.2020.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watson OJ, Slater HC, Verity R, Parr JB, Mwandagalirwa MK, Tshefu A, Meshnick SR, Ghani AC. Modelling the drivers of the spread of Plasmodium falciparum hrp2 gene deletions in sub-Saharan Africa. eLife. 2017;6:e25008. doi: 10.7554/eLife.25008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watson OJ, Verity R, Ghani AC, Garske T, Cunningham J, Tshefu A, Mwandagalirwa MK, Meshnick SR, Parr JB, Slater HC. Impact of seasonal variations in Plasmodium falciparum malaria transmission on the surveillance of pfhrp2 gene deletions. eLife. 2019;8:e40339. doi: 10.7554/eLife.40339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WHO World Malaria Report 2017. WHO 2017
- World Health Organisation False-negative RDT results and implications of new reports of P. falciparum histidine-rich protein 2/3 gene deletions. World Health Organisation 2016a
- World Health Organisation P. falciparum hrp2/3 gene deletions. Conclusions and recommendations of a Technical Consultation. World Health Organisation 2016b
- World Health Organisation Malaria Rapid Diagnostic Test Performance. Results of WHO Product Testing of Malaria RDTs 2018
- World Health Organisation World Malaria Report. World Health Organisation 2020
- World Health Organization Surveillance template protocol for pfhrp2/pfhrp3 gene deletions. World Health Organization 2020
- Zemene E, Koepfli C, Tiruneh A, Yeshiwondim AK, Seyoum D, Lee M-C, Yan G, Yewhalaw D. Detection of foci of residual malaria transmission through reactive case detection in Ethiopia. Malaria Journal. 2018;17:390. doi: 10.1186/s12936-018-2537-5. [DOI] [PMC free article] [PubMed] [Google Scholar]