

Abstract
Objective
Juvenile fibromyalgia (FM) is a prevalent chronic pain condition affecting children and adolescents worldwide during a critical period of brain development. To date, no published studies have addressed the pathophysiology of juvenile FM. This study was undertaken to characterize gray matter volume (GMV) alterations in juvenile FM patients for the first time, and to investigate their functional and clinical relevance.
Methods
Thirty‐four female adolescents with juvenile FM and 38 healthy adolescents underwent a structural magnetic resonance imaging examination and completed questionnaires assessing core juvenile FM symptoms. Using voxel‐based morphometry, we assessed between‐group GMV differences and associations between GMV and functional disability, fatigue, and pain interference in juvenile FM. We also studied whether validated brain patterns predicting pain, cognitive control, or negative emotion were amplified/attenuated in juvenile FM patients and whether structural alterations reported in adult FM were replicated in adolescents with juvenile FM.
Results
Compared to controls, juvenile FM patients showed GMV reductions in the anterior midcingulate cortex (aMCC) region (family‐wise error corrected P [P FWE‐corr] = 0.04; estimated with threshold‐free cluster enhancement [TFCE]; n = 72) associated with pain. Within the juvenile FM group, patients reporting higher functional disability had larger GMV in inferior frontal regions (P FWE‐corr = 0.006; TFCE estimated; n = 34) linked to affective, self‐referential, and language‐related processes. Last, GMV reductions in juvenile FM showed partial overlap with findings in adult FM, specifically for the anterior/posterior cingulate cortices (P = 0.02 and P = 0.03, respectively; n = 72).
Conclusion
Pain‐related aMCC reductions may be a structural hallmark of juvenile FM, whereas alterations in regions involved in emotional, self‐referential, and language‐related processes may predict disease impact on patients’ well‐being. The partial overlap between juvenile and adult FM findings strengthens the importance of early symptom identification and intervention to prevent the transition to adult forms of the disease.
INTRODUCTION
Juvenile fibromyalgia (FM) affects 2–6% of children and adolescents, mostly females, and is characterized by widespread musculoskeletal pain and other debilitating symptoms, such as functional disability and fatigue. Juvenile FM symptoms often persist into adulthood, highlighting the importance of early recognition and intervention (1, 2). Research has provided insight into the clinical characteristics of juvenile FM (2, 3), but there is a research gap regarding disease pathophysiology. A recent review provided the first robust evidence of central hyperexcitability in children with chronic pain and preliminarily suggested altered cortical nociceptive processing (4). Additionally, a functional magnetic resonance imaging (fMRI) study showed that adolescents with idiopathic pain had decreased activation in the thalamus and precentral and middle frontal gyri during pain processing (5).
Conversely, neuroimaging research in adult FM has flourished over the past 2 decades, suggesting that chronic pain symptoms are associated with brain alterations involving multiple circuits and functional domains (6, 7, 8, 9, 10, 11). At the structural level, meta‐analyses in adult FM have consistently found gray matter volume (GMV) reductions in regions involved in affective and cognitive dimensions of pain, such as the anterior midcingulate cortex (aMCC), the medial prefrontal cortex (mPFC), and the default mode network (6, 7). Such reductions have been in part correlated with age, leading researchers to suggest that atrophy in older patients with FM may result from long‐term exposure to nociceptive input (7, 12, 13). The opposite was hypothesized for younger patients, for whom recurrent overengagement of pain modulatory circuits could result in hypertrophy earlier in the disease course followed by atrophy later in life (12, 13). Importantly, GMV loss in adult FM has been identified beyond pain‐processing regions in brain circuits involved in self‐referential processing, executive function, and emotion regulation (14), highlighting the importance of studying whole‐brain structural alterations. Moreover, studies have shown a strong correspondence between structural and functional brain abnormalities, specifically in cingulate and dorsal mPFC regions (15, 16). Therefore, studies combining structural findings with functional meta‐analytic decoding, or validated brain patterns predicting relevant functional domains, may help identify a link between structural abnormalities and their functional relevance in patients with chronic pain.
Pathophysiologic findings at an early disease stage could inform interventions that may more effectively target the multidimensional symptom constellation in juvenile FM. Along these lines, fMRI research in adult FM has linked pain catastrophizing with increased primary somatosensory cortex–anterior insula functional connectivity, which normalized after cognitive behavioral therapy (17). Likewise, it has shown that alterations in the descending pain inhibition pathway improved with regular exercise (18). Therefore, we hypothesize that, if replicated in youth with juvenile FM, such alterations could be modulated with similar treatment strategies as in adults. New insights regarding whether distinct pathophysiologic findings predict different treatment response trajectories may help tailor first treatment actions and most appropriate treatment combinations in a personalized manner. Such knowledge might also contribute to the development of new psychobiologically informed treatments, which could potentially alter the trajectory of juvenile FM symptoms and foster a healthier transition into adulthood (19).
In this study, we investigated GMV abnormalities in adolescent girls with juvenile FM and associations with disease‐related functional interference. We combined voxel‐based morphometry and functional meta‐analytic decoding of structural data to provide insight into potential functional roles of structural alterations in juvenile FM. Based on findings in adult FM (6, 7), we hypothesized that 1) compared to healthy adolescents, juvenile FM patients would show alterations in the cingulate and medial prefrontal cortices and the parahippocampal gyrus, and 2) within the juvenile FM group, individual differences in functional disability, fatigue, and pain interference would be associated with structural abnormalities in circuits involved in emotional processing and cognitive modulation of pain and emotion (anterior insula, prefrontal cortices, limbic, and striatal regions) (20, 21). We also investigated whether 3 validated brain patterns involving medial prefrontal regions and predicting pain, cognitive control, or negative emotion (22) were amplified or attenuated in juvenile FM. Given that amplified pain is a core juvenile FM symptom, we anticipated between‐group differences specifically for the pain‐predictive pattern. Finally, we tested whether structural alterations reported in adult FM were replicated in adolescents with juvenile FM to potentially identify a link between structural abnormalities in juvenile and adult forms of the disease.
PATIENTS AND METHODS
Participants
This study included 34 adolescent girls diagnosed as having juvenile FM (mean ± SD age 16.37 ± 1.07 years [range 13–18 years]) and 38 healthy adolescent girls (mean ± SD age 15.89 ± 1.32 years [range 13–18 years]). The sample size was decided assuming similar power as previous studies that assessed structural differences between adults with FM and healthy controls using voxel‐based morphometry (studies included in meta‐analyses [6,7]). We enrolled female participants exclusively because most chronic pain conditions occurring in adolescence predominantly affect girls (23) and because boys with juvenile FM are sufficiently rare as to raise questions about the potential for a distinct mechanism (24). Inclusion and exclusion criteria are detailed in the Supplementary Methods, available on the Arthritis & Rheumatology website at http://onlinelibrary.wiley.com/doi/10.1002/art.42073. Participants provided informed assent, and their parents or legal guardians provided written informed consent. The study protocol and consent forms were approved by the Institutional Review Board of the Cincinnati Children's Hospital Medical Center (study ID: 2017‐7771) in compliance with the Declaration of Helsinki.
Clinical measures
Developmentally appropriate and validated self‐report measures were used to assess disability, fatigue, and pain interference. Participants self‐reported their functional disability (physical difficulty experienced when doing their daily activities) using the 15‐item Functional Disability Inventory (FDI) (25) and their fatigue symptoms over the past week using the 10‐item Patient‐Reported Outcomes Measurement Information System (PROMIS) Pediatric Fatigue‐Short Form (26). Additionally, juvenile FM patients completed the Brief Pain Inventory (BPI) (27), from which we extracted mean pain intensity and pain interference scores. Healthy adolescents did not complete the BPI because they were selected based on having no pain.
Imaging data acquisition
MRI data acquisition is detailed in the Supplementary Methods, available on the Arthritis & Rheumatology website at http://onlinelibrary.wiley.com/doi/10.1002/art.42073.
Data analysis
Statistical analyses of demographic and clinical variables
Differences between juvenile FM patients and healthy adolescents with regard to sociodemographic and clinical variables were analyzed by chi‐square and 2‐sample t‐tests, using SPSS software version 26 (IBM).
Voxel‐based morphometry protocol
Voxel‐based morphometry quantifies gray matter at a voxelwise, whole‐brain level (28), allowing for a comprehensive measurement of gray matter throughout the brain. In this study, structural data were preprocessed using MatLab‐R2021a (MathWorks) and SPM12 (UCL). First, a trained researcher (MS) reviewed all images to ensure they were free from acquisition artifacts and magnetic field inhomogeneities. Then, images were preprocessed using a standard pipeline combining voxel‐based morphometry with Diffeomorphic Anatomical Registration Through Exponentiated Lie Algebra algorithm (DARTEL) (29) and including 4 main steps: tissue segmentation, normalization, modulation, and Gaussian smoothing. After segmentation, the resulting rigidly transformed versions of gray matter were normalized using DARTEL. Specifically, images were iteratively matched to a template generated from their own mean to generate a series of templates with increasing resolution. Native space gray matter images were then registered to the highest resolution template within a high‐dimensional diffeomorphic framework. Since the DARTEL method creates a study‐specific template and then spatially normalizes individual images to this template, it provides better registration, specifically in the boundaries of gray and white matter, than previous voxel‐based morphometry methods (29).
Spatially normalized tissue maps were modulated by the Jacobian determinants from the corresponding flow fields to restore the volumetric information lost during the high‐dimensional spatial registration. This modulation step multiplies each voxel by the relative change in volume, which allows for a comparison of absolute GMV corrected for individual brain size (30). Only normalized and modulated images were transformed to a standard Montreal Neurological Institute (MNI) template and resliced to 1.5 mm. Therefore, volume computations were not affected by this final transformation to MNI space. Similarly, multiple neuroimaging studies have used this template in adolescents (for example, ref. 31). Moreover, using a standard MNI template enables us to compare our findings with the results of previous studies that used this template (for example, ref. 12). Finally, images were smoothed with an 8‐mm full‐width at half‐maximum isotropic Gaussian kernel. A trained researcher (MS) reviewed all processed images to ensure image quality.
Statistical analyses of brain structural data
To assess regional GMV differences between juvenile FM patients and controls, we estimated a t‐test model with SPM12. As a standard procedure in voxel‐based morphometry analyses (32), we added age and total GMV as nuisance variables, and excluded all voxels with a gray matter value of <0.2 (maximum value 1).
To test associations between regional GMV and clinical variables in juvenile FM patients, we performed multiple regression models in SPM12 with the following independent variables of interest in each separate model: 1) functional disability score from the FDI, 2) fatigue symptoms, measured with the PROMIS form, and 3) pain interference score from the BPI. Each model included age and total GMV as nuisance covariates and GMV as the dependent variable. We excluded voxels with a gray matter value of <0.2.
To correct for multiple comparisons, we performed, for each analysis, a voxelwise nonparametric permutation test with 5,000 random permutations using the threshold‐free cluster enhancement (TFCE) approach (33) implemented in the SPM‐TFCEr214 toolbox (dbm.neuro.uni‐jena.de/tfce), and the whole‐brain significance threshold was set at a family‐wise error corrected P value (PFWE‐corr) of less than 0.05. The TFCE approach was introduced to increase the sensitivity of voxel‐based analysis (33) and is currently recommended and widely used in voxel‐based morphometry studies (34). Compared to voxelwise or clusterwise inferences, TFCE inference improves power and validity and relies on minimal assumptions about data distribution (33). For completion, imaging results were also explored at an uncorrected threshold of P < 0.001 and a cluster extent (Ke) >50 voxels.
Neurosynth meta‐analytic decoding
To interpret the functional role of structural regions differing in GMV between juvenile FM patients and controls, or associated with clinical symptoms in the juvenile FM group, we performed meta‐analytic decoding using the Neurosynth database (35) (neurosynth.org). This meta‐analytic strategy leverages the power of large data sets to compute whole‐brain distributions for psychological terms. Thus, it allows performing unbiased reverse‐inferences to identify terms consistently associated with a particular brain coordinate across neuroimaging studies (35). We uploaded the unthresholded t‐maps produced in our study onto the Neurosynth database and decoded them, which returned psychological and anatomic terms associated with the spatial pattern of each analysis (Supplementary Table 1, available on the Arthritis & Rheumatology website at http://onlinelibrary.wiley.com/doi/10.1002/art.42073, shows a list of the 50 Neurosynth terms most correlated with each t‐map and their Pearson's correlation coefficients.) For interpretation purposes, we selected the 10 psychological terms (i.e., referring to psychological functions) most correlated with each map and displayed them in Figures 1 and 2 (36).
Figure 1.
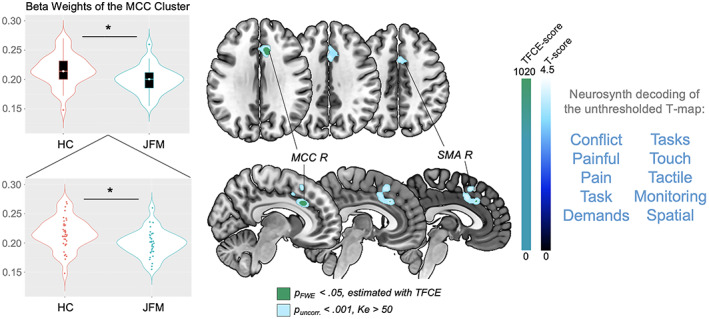
Regions of significantly reduced gray matter volume (GMV) in adolescents with juvenile fibromyalgia (JFM) compared to healthy controls (HCs). Left, Violin plots of the beta weights of the anterior midcingulate cortex (aMCC) cluster in healthy controls and patients with juvenile FM. In the top panel, each box represents the 25th to 75th percentiles. Lines inside the boxes represent the median. Lines outside the boxes represent the 10th and 90th percentiles. Circles indicate outliers. In the bottom panel, circles indicate individual subjects. * = P < 0.01. Middle, Brain maps showing significant between‐group GMV differences (contrast: juvenile FM patients < healthy controls). Results are presented at a significance level of family‐wise error corrected P (P FWE‐corr) < 0.05, estimated using the threshold‐free cluster enhancement (TFCE) approach, and uncorrected P (P uncorr) < 0.001 with a cluster extent (Ke) >50 voxels. Right, The 10 functional annotations most associated with the unthresholded t‐map, obtained by meta‐analytic decoding using Neurosynth. R = right; SMA = supplementary motor area.
Figure 2.
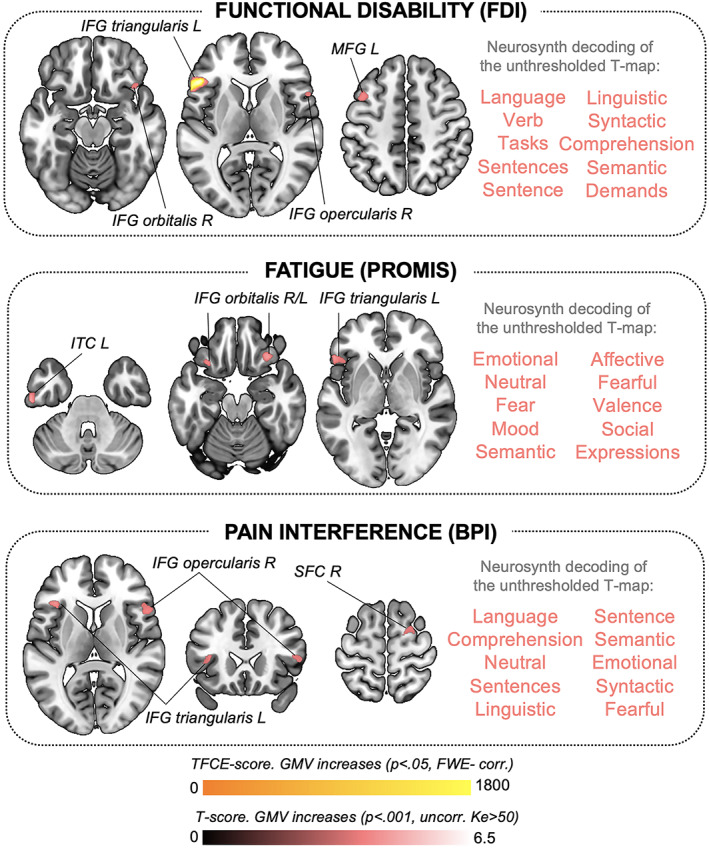
GMV changes associated with clinical symptoms in adolescents with juvenile FM. Brain maps show significant correlations between GMV and clinical symptoms reflecting disease‐related functional interference in adolescents with juvenile FM, including functional disability as measured by the Functional Disability Inventory (FDI), fatigue as measured by Patient‐Reported Outcomes Measurement Information System (PROMIS), and pain interference as measured by the Brief Pain Inventory (BPI). Results are presented at a significance level of P FWE‐corr < 0.05 (GMV increases shown in yellow) and P uncorr < 0.001 with Ke >50 voxels (GMV increases shown in red). We did not observe negative correlations at the P FWE‐corr or P uncorr level. The 10 functional annotations most associated with the corresponding unthresholded t‐map, obtained by meta‐analytic decoding using Neurosynth, are shown to the right of each brain map. L = left; IFG = inferior frontal gyrus; MFG = middle frontal gyrus; ITC = inferior temporal cortex; SFC = superior frontal cortex (see Figure 1 for other definitions).
Evaluation of patterns predicting pain, cognitive control, and negative emotion in juvenile FM
Our voxel‐based morphometry analysis showed that juvenile FM patients had reduced GMV in the right aMCC. To study the functional organization of the medial frontal cortex (MFC) surrounding this region, we tested whether the similarity between our structural maps and previously validated brain patterns predicting either pain, cognitive control, or negative emotion across 18 studies and specifically involving the aMCC–dorsal MFC (22) differed between juvenile FM patients and healthy adolescents. The maps developed by Kragel et al (22) corresponding to the partial least squares regression coefficients for the different functional domains being predicted are available at the Canlab repository (github.com/canlab/Neuroimaging_Pattern_Masks/tree/master/Multivariate_signature_patterns/2018_Kragel_MFC_Generalizability). We extracted the weighted maps corresponding to pain, cognitive control, and negative emotion in the dorsomedial aspect of the frontal cortex including anterior and posterior MCC and dorsal MFC. For each subject, we computed the dot product between our preprocessed structural images, regressed by age and total GMV, and pain, cognitive control, and negative emotion–weighted maps. Dot product was calculated using publicly available code at the Canlab repository (canlab.github.io) running on MatLab. We then ran 2‐sample t‐tests using SPSS to assess between‐group differences in pattern similarity for each weighted map.
Evaluation of GMV differences in regions found altered in adults with FM
To identify potentially overlapping alterations between adult and juvenile forms of FM, we assessed GMV alterations in regions of interest (ROIs) identified based on previous meta‐analytic evidence in adult FM patients. The selection and construction of ROIs is detailed in the Supplementary Methods, available on the Arthritis & Rheumatology website at http://onlinelibrary.wiley.com/doi/10.1002/art.42073. For each subject, we computed the mean parameter estimates (beta values representing GMV for each subject after removing age and total GMV effects) within each of the 6 final ROIs. Finally, we assessed between‐group differences in mean parameter estimates for each ROI using 2‐sample t‐tests in SPSS.
RESULTS
Demographic and clinical variables
Adolescents with juvenile FM and healthy controls did not differ in terms of sex (all female), age, race, household income, or caregiver education level (all P > 0.1) (Table 1). As anticipated, juvenile FM patients reported significantly higher functional disability (t = 13.89, P < 0.001) and fatigue symptoms (t = 12.30, P < 0.001) (Table 1). Since controls were selected based on having no pain (numerical rating scales for pain of 0), mean pain intensity and pain interference were only assessed in juvenile FM participants (mean ± SD intensity 5.43 ± 1.42; mean ± SD interference 4.29 ± 1.55), indicating moderate intensity and mild‐to‐moderate interference. Medication details are presented in Table 1.
Table 1.
Demographic and clinical characteristics of the patients with juvenile FM and healthy controls*
| Controls (n = 38) | Patients with juvenile FM (n = 34) | t/LRV | |
|---|---|---|---|
| Demographic characteristic | |||
| Age, years | 15.89 ± 1.32 | 16.37 ± 1.07 | −1.67 |
| Race, no. Caucasian/non‐Caucasian | 35/3 | 32/2 | 0.11† |
| Annual household income (scale 1–6)‡ | 5.13 ± 2.07 | 5.09 ± 2.02 | 0.09 |
| Education level of caregiver 1 (scale 1–5)§ | 3.97 ± 0.88 | 3.88 ± 0.91 | 0.43 |
| Education level of caregiver 2 (scale 1–5)§ | 3.74 ± 0.78 | 3.63 ± 0.98 | 0.55 |
| Brain | |||
| Total GMV volume | 787.70 ± 59.87 | 785.50 ± 53.17 | 0.16 |
| Total WMV | 384.27 ± 40.42 | 399.49 ± 36.89 | −1.66 |
| Clinical variables | |||
| Functional disability (FDI) | 0.5 ± 1.25 | 22.74 ± 9.26 | −13.89¶ |
| Fatigue (PROMIS) | 14.28 ± 6.64 | 35.23 ± 6.87 | −12.30¶ |
| Pain interference (BPI) | – | 4.29 ± 1.55 | – |
| Average pain intensity | – | 5.43 ± 1.42 | – |
| Medication, no. (%) | |||
| Pain‐related drugs# | – | 12 (35.29) | – |
| Psychiatric drugs** | – | 15 (44.12) | – |
| Gastrointestinal drugs†† | – | 7 (20.59) | – |
| Melatonin | – | 3 (8.82) | – |
| Antihistamines | – | 3 (8.82) | – |
| Vitamins/iron supplements | – | 3 (8.82) | – |
| Birth control | 2 (5.26) | 2 (5.88) | – |
| Hypertension treatment | 1 (2.63) | – | – |
| Statins | 1 (2.63) | – | – |
Except where indicated otherwise, values are the mean ± SD. FM = fibromyalgia; GMV = gray matter volume; WMV = white matter volume; FDI = Functional Disability Inventory; PROMIS = Patient‐Reported Outcomes Measurement Information System; BPI = Brief Pain Inventory.
Likelihood ratio value (LRV).
Annual household income is shown using a scale of 1–6, where 1 = <$24,999; 2 = $25,000 to $49,999; 3 = $50,000 to $74,999; 4 = $75,000 to $99,000; 5 = $100,000 to $124,999; and 6 = >$125,000.
Caregiver education level is shown using a scale of 1–5, where 1 = less than high school; 2 = high school/GED; 3 = partial college or trade school; 4 = college graduate; 5 = postgraduate degree.
Significant values (P < 0.001).
# Pain‐related drugs included antiepileptic drugs, nonsteroidal antiinflammatory drugs, muscle relaxants, acetaminophen, and/or acetylsalicylic acid.
Psychiatric drugs included antidepressants, anxiolytics, and attention deficit hyperactivity disorder drugs.
Gastrointestinal drugs included antiacids, antireflux drugs, and constipation drugs.
Whole‐brain voxel‐based morphometry analyses
Reductions in GMV in juvenile FM
The groups did not differ in total GMV (P = 0.87). Compared to controls, juvenile FM patients had significantly less regional GMV in a cluster of the right aMCC (P FWE‐corr = 0.04; TFCE estimated). At the uncorrected threshold (P < 0.001; Ke >50 voxels), this cluster extended to the left aMCC and bilateral supplementary motor area. Neurosynth meta‐analytic decoding revealed that the unthresholded t‐map for this contrast was related to, among others, the functional terms “pain,” “painful,” and “conflict” (Figure 1 and Table 2). However, correlations between the beta weights of the FWE‐corrected aMCC cluster and clinical variables (functional disability, fatigue, pain interference, and pain intensity) did not reveal any significant association (all P > 0.3) (Supplementary Table 2, available on the Arthritis & Rheumatology website at http://onlinelibrary.wiley.com/doi/10.1002/art.42073), which suggests that, in our sample, GMV reductions in this area do not track clinical severity but instead identify the category of juvenile FM patients as compared with healthy adolescents. Similarly, beta weights of the aMCC cluster did not correlate with symptom duration (r = 0.078; P = 0.677), which indicates that this alteration does not depend on how long patients have been experiencing juvenile FM symptoms.
Table 2.
Results of the whole‐brain voxel‐based morphometry analyses*
| Brain region | x, y, z† | T | Cluster size, voxels‡ | |
|---|---|---|---|---|
| Structural results at P FWE‐corr < 0.05, estimated with TFCE | ||||
| GMV differences between adolescents with juvenile FM and controls | ||||
| Contrast: juvenile FM < controls | aMCC R | 9, 22, 30 | 1018.43§ | 113 |
| Correlations between GMV and clinical symptoms in adolescents with juvenile FM | ||||
| Functional disability (FDI) | ||||
| ↑GMV | IFG pars triangularis L | −54, 22, 4 | 1784.78§ | 340 |
| Structural results at P uncorr < 0.001, Ke >50 voxels | ||||
| GMV differences between adolescents with juvenile FM and controls | ||||
| Contrast: juvenile FM < controls | aMCC R | 9, 22, 30 | 4.14 | 983 |
| Correlations between GMV and clinical symptoms in adolescents with juvenile FM | ||||
| Functional disability (FDI) | ||||
| ↑GMV | IFG pars triangularis L | −54, 22, 4 | 6.47 | 805 |
| ↑GMV | IFG pars opercularis R | 57, 14, 15 | 4.35 | 242 |
| ↑GMV | MFG L | −42, 12, 48 | 4.13 | 191 |
| ↑GMV | IFG pars orbitalis R | 42, 21, −14 | 3.81 | 117 |
| Fatigue (PROMIS) | ||||
| ↑GMV | Inferior temporal cortex L | −60, −16, −33 | 5.03 | 115 |
| ↑GMV | IFG pars triangularis L | −54, 26, −3 | 4.83 | 317 |
| ↑GMV | IFG pars orbitalis R | 32, 27, −21 | 4.63 | 366 |
| ↑GMV | IFG pars triangularis L | −50, 28, 9 | 4.54 | 82 |
| ↑GMV | IFG pars orbitalis L | −30, 21, −20 | 4.18 | 76 |
| Pain interference (BPI) | ||||
| ↑GMV | IFG pars triangularis L | −38, 26, 10 | 4.97 | 157 |
| ↑GMV | IFG pars opercularis R | 52, 16, 12 | 4.21 | 264 |
| ↑GMV | Superior frontal cortex R | 28, −3, 66 | 4.11 | 154 |
P FWE‐corr = family wise error rate–corrected P value; GMV = gray matter volume; FM = fibromyalgia; aMCC = anterior midcingulate cortex; R = right; FDI = Functional Disability Inventory; IFG = inferior frontal gyrus; L = left; P uncorr = uncorrected P value; MFG = middle frontal gyrus; PROMIS = Patient‐Reported Outcomes Measurement Information System; BPI = Brief Pain Inventory.
Anatomic coordinates (x, y, z) are given in Montreal Neurological Institute space.
Voxel size 1.5 × 1.5 × 1.5 mm.
Threshold‐free cluster enhancement (TFCE) values.
Correlations between GMV and clinical juvenile FM symptoms
Functional disability correlated with increased GMV in the left inferior frontal gyrus (IFG) (P FWE‐corr = 0.006; TFCE estimated). At the uncorrected level (P < 0.001; Ke >50 voxels), functional disability was linked to larger volumes in the bilateral IFG and the left middle frontal gyrus. Additionally, fatigue correlated with increased GMV in the bilateral IFG and the left inferior temporal cortex. Pain interference was associated with larger GMV in the left and right IFG and the right superior frontal cortex. These findings are detailed in Table 2. Figure 2 shows the anatomy of the findings and the associated meta‐analytic terms based on Neurosynth decoding of each unthresholded, symptom‐predicting t‐map. Notably, t‐maps reflecting GMV increases associated with functional disability, fatigue, and pain interference in juvenile FM patients were strongly linked to emotional and/or language production processes.
Importantly, the symptom–GMV associations described above remained significant after including symptom duration as an additional nuisance variable, along with age and total GMV, in the multiple regression models. Thus, these associations are independent of symptom duration. As a post hoc analysis, we built an additional model with symptom duration as a predictor variable, GMV as the dependent variable, and age and total GMV as nuisance covariates. We found no significant associations at the corrected level (P FWE‐corr < 0.05; TFCE estimated). At the uncorrected threshold (P < 0.001, Ke >50 voxels), symptom duration correlated with GMV decreases in right‐lateralized clusters in temporoparietal areas (Supplementary Table 3, available on the Arthritis & Rheumatology website at http://onlinelibrary.wiley.com/doi/10.1002/art.42073). According to Neurosynth decoding, the corresponding t‐map was most associated with theory of mind and language‐related processes (Supplementary Figure 1, available on the Arthritis & Rheumatology website at http://onlinelibrary.wiley.com/doi/10.1002/art.42073).
Reduced similarity between brain structure and a validated pain‐predictive pattern involving the aMCC–dorsal MFC in juvenile FM
We tested whether validated brain patterns predicting either pain, cognitive control, or negative emotion, specifically in the aMCC–dorsal MFC region (22), differed between juvenile FM patients and controls. We found between‐group differences only in the pain pattern, which is consistent with the location of GMV between‐group differences and the fact that pain is a core complaint in juvenile FM. Specifically, the dot product between structural data and the pain–weighted map was significantly lower in juvenile FM patients compared to controls within the aMCC–dorsal MFC mask (t = 2.47, P = 0.016) (Figure 3). Thus, the anatomic aMCC pattern resembled the functional pain pattern significantly less in juvenile FM patients than in controls. We found no differences for the patterns predicting cognitive control or negative emotion (all P > 0.4).
Figure 3.
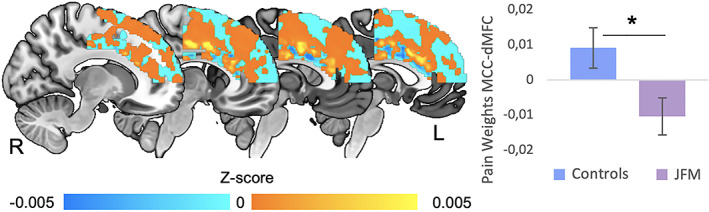
Differences in the similarity between brain structure and a validated pain‐predictive pattern in anterior midcingulate cortex (aMCC)–dorsal medial frontal cortex (dMFC) circuits in patients with juvenile fibromyalgia (JFM) compared to healthy adolescents. The anatomic aMCC–dMFC pattern resembled the functional pain pattern significantly less in juvenile FM patients than in healthy adolescents (t = 2.47, P = 0.016). We found no between‐group differences in the patterns predicting cognitive control and negative emotion within the aMCC–dMFC (all P > 0.05). Bars show the mean ± SEM. * = P < 0.02.
GMV differences between adolescents with juvenile FM and healthy adolescents within meta‐analytic regions altered in adults with FM
Consistent with meta‐analytic findings in adults (6, 7), GMV reductions in 2 ROIs, located in the left anterior cingulate cortex (ACC) and the right posterior cingulate cortex (PCC), were replicated in adolescents with juvenile FM (P = 0.02 and P = 0.03, respectively) (Figure 4A). Neurosynth meta‐analytic decoding showed that the left ACC ROI was associated with functional terms such as “pain,” “autobiographical,” and “mentalizing” (the ability to understand the mental state of oneself or others), whereas the right PCC ROI was associated with “suppressed,” “declines,” and “resting state.” The remaining ROIs extracted from the meta‐analyses (6, 7), located in the cerebellum, the dorsal ACC, and the fusiform and parahippocampal gyri, did not show evidence of alteration in juvenile FM patients (all P > 0.4), as opposed to what has been reported in adults with FM (see Figure 4B). Notably, among our sample, 2 juvenile FM participants had Chiari malformations, which is no higher than might be expected based on population prevalence (37).
Figure 4.
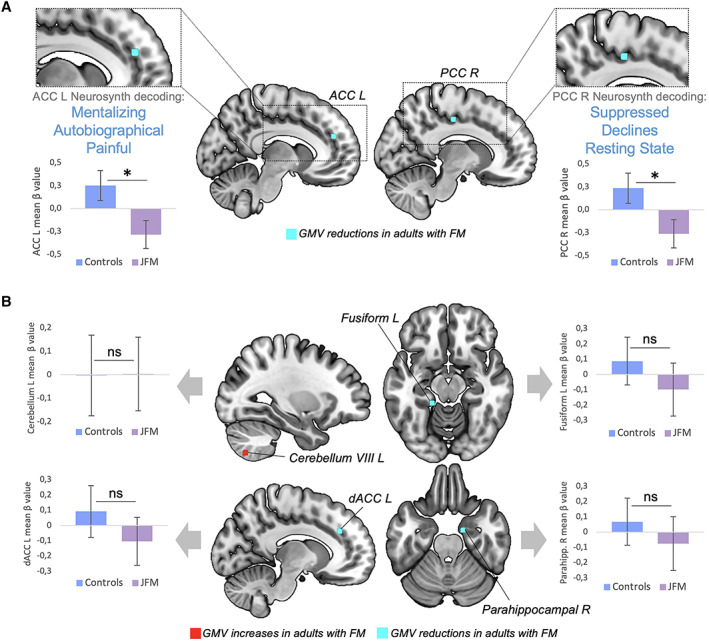
Replication in juvenile fibromyalgia (JFM) of meta‐analytic gray matter volume (GMV) alterations observed in adults with FM. A, Peaks of GMV altered in both adults with FM and adolescents with juvenile FM (all P < 0.03). Notably, the GMV at these peaks was decreased in both adult and juvenile forms of FM. Three associated functional annotations, obtained by meta‐analytic decoding using Neurosynth, are shown for each region. B, Peaks of GMV altered in adults with FM but not in adolescents with juvenile FM (all P > 0.4). Bars show the mean ± SEM. * = P < 0.03. L = left; R = right; dACC = dorsal anterior cingulate cortex; PCC = posterior cingulate cortex; NS = not significant (P > 0.05).
DISCUSSION
To our knowledge, this is the first study assessing brain structural alterations in juvenile FM. Results indicate that decreased GMV in the aMCC may be a key feature of juvenile FM. This region is a core element of central acute pain processing (9, 10, 38, 39, 40), involved in affective encoding, cognitive interpretation, anticipation, and response selection (41). Consistent with these findings, meta‐analytic decoding revealed that among all t‐maps generated in our study, the one for aMCC was the only one associated with the terms “pain” and “painful.” Moreover, results showed that the anatomic aMCC pattern resembled the validated aMCC–dorsal MFC pain predictive pattern—and not the negative emotion or cognitive control ones—significantly less in juvenile FM patients than in healthy adolescents.
We also found that adolescents with juvenile FM exhibited GMV reductions in 2 of the 3 cingulate regions (i.e., ACC, PCC) that meta‐analyses reported to be decreased in adult FM patients. Previous studies have shown that repetitive painful stimulation leads to aMCC gray matter increases in healthy subjects (42), whereas patients with chronic pain display GMV reductions in this region (6, 7). Our findings do not support the hypothesis that younger patients may show increased GMV as a consequence of overengaging pain modulatory systems (12, 13). Instead, results suggest that reductions in the cingulate cortex, mainly in the aMCC, may be a structural hallmark of both adult and juvenile forms of FM, independent of symptom duration. Longitudinal studies are warranted to determine whether such alterations predate chronic pain onset or reflect an early pain‐driven alteration. Notably, other GMV alterations reported in adults were not replicated in juvenile FM patients, which may be due to the limited power of the present investigation or may suggest that they appear later in life as a result of progression of chronic pain or medication exposure.
The aMCC is not selective for pain (43, 44). Research has linked activity in this area with multiple functions, including attention, cognitive control, reward‐based learning, decision‐making, and emotional and social processing (for review, see refs. 41 and 43). To deepen our understanding of the functional contributions of the MFC surrounding the region where we found GMV reductions in juvenile FM patients, we assessed how this structural pattern resembled functional patterns predicting pain, cognitive control, and negative emotion within the aMCC–dorsal MFC (22), and how this similarity differed between groups. In adolescents with juvenile FM, the anatomic aMCC pattern resembled the pain pattern significantly less than in controls, which may suggest that the anatomic hallmark of nociceptive pain processing has been attenuated to some degree in juvenile FM patients.
Whether such findings reflect the effects of excessive engagement of the pain‐specific functional pattern or a reorganization of acute nociceptive processing brain circuits in patients remains to be elucidated. This finding indicates that pattern similarity reductions in patients occur beyond the cluster showing GMV reductions and reflects an overall reduction of the normal pain predictive pattern in patients. Moreover, it supports the idea of altered organization of aMCC–dorsal MFC circuits. Future functional studies using experimental pain tasks are warranted to test the extent to which this alteration replicates during pain processing to further interpret this novel finding. Taken together, alterations of the pain pattern in juvenile FM patients, along with meta‐analytic decoding findings specifically mentioning the terms “pain” and “painful,” suggest that this abnormality may be related to pain processing and decision‐making/evaluative aspects in the context of pain and less with other emotional/affective/cognitive processes.
Regarding regions associated with individual differences in patients’ clinical symptoms reflecting pain‐related suffering and diminished functional ability, we found a significant association between functional disability and the left ventrolateral prefrontal cortex (vlPFC). Meta‐analyses have linked the left vlPFC with language processes during emotion regulation, suggesting that it might support active reinterpretation of the meaning of emotional stimuli and facilitate the selection of appropriate reappraisals (45). Likewise, the vlPFC has been associated with retrieval of semantic autobiographical memories and self‐related judgments (46). Consistent with these findings, at the uncorrected level, we found that other symptoms reflecting the impact that juvenile FM has on patients’ functional ability (i.e., functional disability, fatigue, and pain interference) all correlated with GMV increases in the vlPFC. Taken together, these findings suggest that juvenile FM patients with higher levels of pain‐related suffering and impairment show augmented GMV in brain circuits involved in instantiating representations of the self and the world through language.
Adding further support to this interpretation, at the uncorrected level, fatigue correlated with increased volumes in the left inferior temporal cortex, linked to visual and mnemonic processing (47), and functional disability and pain interference correlated with larger volumes in the right superior frontal cortex, involved in self‐focused reappraisal (48), and in the left middle frontal gyrus, associated with attention reorienting (49). Notably, these associations were independent of symptom duration. Neurosynth decoding confirmed that these variables were associated with a brain pattern related to emotional, self‐related judgment, and language‐related processes. Future studies should test whether alterations in the nature, recurrence, and valence of patients’ narratives about themselves and the world may predict greater levels of suffering and disability in juvenile FM and to which extent vlPFC circuits mediate such associations.
From a neurodevelopmental perspective, frontal GMV decreases after age 11 years in healthy girls because of synaptic pruning (50). This maturation occurs earlier in ventral than in dorsal regions (50). Thus, vlPFC volume increases associated with juvenile FM impairment may reflect a link between these symptoms and developmental immaturity in frontal circuits specializing in emotional appraisal and regulation, which reinforces the need to consider therapeutic strategies that target these circuits, which may have the potential to reverse alterations before they become hard‐wired and to mitigate the functional and psychosocial impact of pain‐related symptoms on the life of adolescents with juvenile FM.
Finally, symptom duration was associated with reduced GMV in temporoparietal areas—at an uncorrected level—including theory of mind– and language‐related regions. The findings suggest that not only disease‐related disability, but also juvenile FM duration may be linked to alterations in regions mediating mentalizing and emotional awareness through language. Last, our between‐group and symptom duration–related findings taken together do not support the hypothesis of hypertrophy in young FM patients as a group (12, 13). However, juvenile FM patients with greater symptom severity showed GMV increases; thus, future studies with larger samples are warranted to assess whether different structural correlates underlie distinct patient clusters identified based on symptom severity.
This study has some notable limitations. First, we enrolled only females; thus, our findings cannot be generalized to male patients—although they are quite rare. Future studies are warranted to examine between‐sex differences in juvenile FM–related structural alterations and the ages at which these changes occur. Second, since this is the first study assessing the structural abnormalities of juvenile FM, replication of our findings in independent samples is crucial to determine their robustness and translational utility. Likewise, the brain–symptom severity correlations presented here, although providing a cohesive picture of the alterations and opening venues for future research, are still preliminary, based on exploratory thresholds, and need replication. Nevertheless, such correlational findings highlight the importance of studying individual differences to characterize different patient profiles. Although medication regimen was stable for at least 3 weeks before the MRI assessment (no modifications in type, dose, or intake regimen), medication could have acted as a confounding factor. Last, despite the efforts to recruit diverse patient profiles, our sample had a low representation of subjects of different races and ethnicities and those with low socioeconomic status. Future community‐oriented clinical research is needed to overcome the systematically high proportion of White participants with medium‐high socioeconomic status in research samples.
In conclusion, this study provides the first evidence of structural alterations in adolescents with juvenile FM. Our findings suggest that pain‐related GMV reductions in the aMCC are common to juvenile FM patients as a group, whereas alterations in regions involved in affective, self‐relevant memory, and language processes predict disease impact on clinical variables related to patients’ well‐being. Taken together, the findings reinforce the need to combine pain‐specific, sensory therapies with therapies aimed at promoting cognitive regulation of pain, negative affect, and potentially pervasive self‐related narratives patients may hold of themselves. Also, the findings indicate partial overlap in the structural circuitry compromised in both juvenile and adult FM, potentially establishing a link between juvenile and adult forms of the disease and strengthening the need for early, neurobiologically oriented interventions to prevent the transition from juvenile to adult FM.
AUTHOR CONTRIBUTIONS
All authors were involved in drafting the article or revising it critically for important intellectual content, and all authors approved the final version to be published. Dr. Suñol had full access to all of the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis.
Study conception and design
Suñol, Maloney, Ting, Kashikar‐Zuck, Coghill, López‐Solà.
Acquisition of data
Payne, Tong.
Analysis and interpretation of data
Suñol, Coghill, López‐Solà.
Supporting information
Disclosure Form
Appendix S1: Supporting Information
ACKNOWLEDGMENTS
The authors gratefully thank Matt Lanier, Kaley Bridgewater, Kelsey Murphy, Brynne Williams, and Lacey Haas (Imaging Research Center, Department of Radiology, Cincinnati Children's Hospital Medical Center) for their contributions to MRI data collection.
Supported by the National Institute of Arthritis and Musculoskeletal and Skin Diseases, NIH (grants R01‐AR‐074795 and P30‐AR‐076316). Dr. Lopez‐Sola's work was supported by the Serra Hunter Programme of the Generalitat de Catalunya.
Author disclosures are available at https://onlinelibrary.wiley.com/action/downloadSupplement?doi=10.1002%2Fart.42073&file=art42073‐sup‐0001‐Disclosureform.pdf.
Contributor Information
Maria Suñol, Email: msunol@ub.edu.
Marina López‐Solà, Email: mlopezsola@ub.edu.
REFERENCES
- 1. Ting TV, Barnett K, Lynch‐Jordan A, Whitacre C, Henrickson M, Kashikar‐Zuck S. 2010 American College of Rheumatology adult fibromyalgia criteria for use in an adolescent female population with juvenile fibromyalgia. J Pediatr 2016;169:181–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Kashikar‐Zuck S, Ting TV, Arnold LM, Bean J, Powers SW, Graham TB, et al. Cognitive behavioral therapy for the treatment of juvenile fibromyalgia: a multisite, single‐blind, randomized, controlled clinical trial. Arthritis Rheum 2012;64:297–305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Kashikar‐Zuck S, Lynch AM, Slater S, Graham TB, Swain NF, Noll RB. Family factors, emotional functioning, and functional impairment in juvenile fibromyalgia syndrome. Arthritis Rheum 2008;59:1392–98. [DOI] [PubMed] [Google Scholar]
- 4. Pas R, Ickmans K, Van Oosterwijck S, Van der Cruyssen K, Foubert A, Leysen L, et al. Hyperexcitability of the central nervous system in children with chronic pain: a systematic review. Pain Med 2018;19:2504–14. [DOI] [PubMed] [Google Scholar]
- 5. Molina J, Amaro E Jr, da Rocha LG, Jorge L, Santos FH, Len CA. Functional resonance magnetic imaging (fMRI) in adolescents with idiopathic musculoskeletal pain: a paradigm of experimental pain. Pediatr Rheumatol Online J 2017;15:81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Lin C, Lee SH, Weng HH. Gray matter atrophy within the default mode network of fibromyalgia: a meta‐analysis of voxel‐based morphometry studies. Biomed Res Int 2016;7296125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Shi H, Yuan C, Dai Z, Ma H, Sheng L. Gray matter abnormalities associated with fibromyalgia: a meta‐analysis of voxel‐based morphometric studies. Semin Arthritis Rheum 2016;46:330–7. [DOI] [PubMed] [Google Scholar]
- 8. Napadow V, Kim J, Clauw DJ, Harris RE. Decreased intrinsic brain connectivity is associated with reduced clinical pain in fibromyalgia. Arthritis Rheum 2012;64:2398–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. López‐Solà M, Pujol J, Wager TD, Garcia‐Fontanals A, Blanco‐Hinojo L, Garcia‐Blanco S, et al. Altered functional magnetic resonance imaging responses to nonpainful sensory stimulation in fibromyalgia patients. Arthritis Rheumatol 2014;66:3200–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. López‐Solà M, Woo CW, Pujol J, Deus J, Harrison BJ, Monfort J, et al. Towards a neurophysiological signature for fibromyalgia. Pain 2017;158:34–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Pujol J, Macià D, Garcia‐Fontanals A, Blanco‐Hinojo L, López‐Solà M, Garcia‐Blanco S, et al. The contribution of sensory system functional connectivity reduction to clinical pain in fibromyalgia. Pain 2014;155:1492–503. [DOI] [PubMed] [Google Scholar]
- 12. Ceko M, Bushnell MC, Fitzcharles MA, Schweinhardt P. Fibromyalgia interacts with age to change the brain. Neuroimage Clin 2013;3:249–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Moayedi M, Weissman‐Fogel I, Salomons TV, Crawley AP, Goldberg MB, Freeman BV, et al. Abnormal gray matter aging in chronic pain patients. Brain Res 2012;1456:82–93. [DOI] [PubMed] [Google Scholar]
- 14. Bushnell MC, Ceko M, Low LA. Cognitive and emotional control of pain and its disruption in chronic pain. Nat Rev Neurosci 2013;14:502–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Jin F, Zheng P, Liu H, Guo H, Sun Z. Functional and anatomical connectivity‐based parcellation of human cingulate cortex. Brain Behav 2018;8:e01070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Tovar DT, Chavez RS. Large‐scale functional coactivation patterns reflect the structural connectivity of the medial prefrontal cortex. Soc Cogn Affect Neurosci 2021;16:875–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Lazaridou A, Kim J, Cahalan CM, Loggia ML, Franceschelli O, Berna C, et al. Effects of cognitive‐behavioral therapy (CBT) on brain connectivity supporting catastrophizing in fibromyalgia. Clin J Pain 2017;33:215–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Ellingson LD, Stegner AJ, Schwabacher IJ, Koltyn KF, Cook DB. Exercise strengthens central nervous system modulation of pain in fibromyalgia. Brain Sci 2016;6:8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Kashikar‐Zuck S, King C, Ting TV, Arnold LM. Juvenile fibromyalgia: different from the adult chronic pain syndrome? Curr Rheumatol Rep 2016;18:19. [DOI] [PubMed] [Google Scholar]
- 20. Lutz J, Jäger L, de Quervain D, Krauseneck T, Padberg F, Wichnalek M, et al. White and gray matter abnormalities in the brain of patients with fibromyalgia: a diffusion‐tensor and volumetric imaging study. Arthritis Rheum 2008;58:3960–69. [DOI] [PubMed] [Google Scholar]
- 21. Martucci KT, Mackey SC. Neuroimaging of pain: human evidence and clinical relevance of central nervous system processes and modulation. Anesthesiology 2018;128:1241–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Kragel PA, Kano M, Van Oudenhove L, Ly HG, Dupont P, Rubio A, et al. Generalizable representations of pain, cognitive control, and negative emotion in medial frontal cortex. Nat Neurosci 2018;21:283–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Fillingim RB, King CD, Ribeiro‐Dasilva MC, Rahim‐Williams B, Riley JL III. Sex, gender, and pain: a review of recent clinical and experimental findings. J Pain 2009;10:447–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Mogil JS. Qualitative sex differences in pain processing: emerging evidence of a biased literature. Nat Rev Neurosci 2020;21:353–65. [DOI] [PubMed] [Google Scholar]
- 25. Walker LS, Greene JW. The functional disability inventory: measuring a neglected dimension of child health status. J Pediatr Psychol 1991;16:39–58. [DOI] [PubMed] [Google Scholar]
- 26. PROMIS . PROMIS Instrument Development and Psychometric Evaluation Scientific Standards Version 2.0. 2013.
- 27. Daut RL, Cleeland CS, Flanery RC. Development of the Wisconsin brief pain questionnaire to assess pain in cancer and other diseases. Pain 1983;17:197–210. [DOI] [PubMed] [Google Scholar]
- 28. Ashburner J, Friston KJ. Voxel‐based morphometry–the methods. Neuroimage 2000;11:805–21. [DOI] [PubMed] [Google Scholar]
- 29. Ashburner J. A fast diffeomorphic image registration algorithm. Neuroimage 2007;38:95–113. [DOI] [PubMed] [Google Scholar]
- 30. Yaxu Y, Ren Z, Ward J, Jiang Q. Atypical brain structures as a function of gray matter volume (GMV) and gray matter density (GMD) in young adults relating to autism spectrum traits. Front Psychol 2020;11:523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Kennedy JT, Astafiev SV, Golosheykin S, Korucuoglu O, Anokhin AP. Shared genetic influences on adolescent body mass index and brain structure: a voxel‐based morphometry study in twins. Neuroimage 2019;199:261–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Barnes J, Ridgway GR, Bartlett J, Henley SM, Lehmann M, Hobbs N, et al. Head size, age and gender adjustment in MRI studies: a necessary nuisance? Neuroimage 2010;53:1244–55. [DOI] [PubMed] [Google Scholar]
- 33. Smith SM, Nichols TE. Threshold‐free cluster enhancement: addressing problems of smoothing, threshold dependence and localisation in cluster inference. Neuroimage 2009;44:83–98. [DOI] [PubMed] [Google Scholar]
- 34. Li H, Nickerson LD, Nichols TE, Gao JH. Comparison of a non‐stationary voxelation‐corrected cluster‐size test with TFCE for group‐Level MRI inference. Hum Brain Mapp 2017;38:1269–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Yarkoni T, Poldrack RA, Nichols TE, Van Essen DC, Wager TD. Large‐scale automated synthesis of human functional neuroimaging data. Nat Methods 2011;8:665–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Koban L, Jepma M, López‐Solà M, Wager TD. Different brain networks mediate the effects of social and conditioned expectations on pain. Nat Commun 2019;10:4096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Kahn EN, Muraszko KM, Maher CO. Prevalence of Chiari I malformation and syringomyelia. Neurosurg Clin N Am 2015;26:501–7. [DOI] [PubMed] [Google Scholar]
- 38. Coghill RC, Talbot JD, Evans AC, Meyer E, Gjedde A, Bushnell MC, et al. Distributed processing of pain and vibration by the human brain. J Neurosci 1994;14:4095–108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Coghill RC, Sang CN, Maisog JM, Iadarola MJ. Pain intensity processing within the human brain: a bilateral, distributed mechanism. J Neurophysiol 1999;82:1934–43. [DOI] [PubMed] [Google Scholar]
- 40. Wager TD, Atlas LY, Lindquist MA, Roy M, Woo CW, Kross E. An fMRI‐based neurologic signature of physical pain. N Engl J Med 2013;368:1388–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Vogt BA. Midcingulate cortex: structure, connections, homologies, functions and diseases. J Chem Neuroanat 2016;74:28–46. [DOI] [PubMed] [Google Scholar]
- 42. Teutsch S, Herken W, Bingel U, Schoell E, May A. Changes in brain gray matter due to repetitive painful stimulation. Neuroimage 2008;15:42:845–9. [DOI] [PubMed] [Google Scholar]
- 43. Shackman AJ, Salomons TV, Slagter HA, Fox AS, Winter JJ, Davidson RJ. The integration of negative affect, pain and cognitive control in the cingulate cortex. Nat Rev Neurosci 2011;12:154–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Silvestrini N, Chen JI, Piché M, Roy M, Vachon‐Presseau E, Woo CW, et al. Distinct fMRI patterns colocalized in the cingulate cortex underlie the after‐effects of cognitive control on pain. Neuroimage 2020;217:116898. [DOI] [PubMed] [Google Scholar]
- 45. Buhle JT, Silvers JA, Wager TD, Lopez R, Onyemekwu C, Kober H, et al. Cognitive reappraisal of emotion: a meta‐analysis of human neuroimaging studies. Cereb Cortex 2014;24:2981–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Denny BT, Kober H, Wager TD, Ochsner KN. A meta‐analysis of functional neuroimaging studies of self‐ and other judgments reveals a spatial gradient for mentalizing in medial prefrontal cortex. J Cogn Neurosci 2012;24:1742–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Ranganath C. Working memory for visual objects: complementary roles of inferior temporal, medial temporal, and prefrontal cortex. Neuroscience 2006;139:277–89. [DOI] [PubMed] [Google Scholar]
- 48. Falquez R, Couto B, Ibanez A, Freitag MT, Berger M, Arens EA, et al. Detaching from the negative by reappraisal: the role of right superior frontal gyrus (BA9/32). Front Behav Neurosci 2014;8:165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Japee S, Holiday K, Satyshur MD, Mukai I, Ungerleider LG. A role of right middle frontal gyrus in reorienting of attention: a case study. Front Syst Neurosci 2015;9:23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Lenroot RK, Giedd JN. Brain development in children and adolescents: insights from anatomical magnetic resonance imaging. Neurosci Biobehav Rev 2006;30:718–29. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Disclosure Form
Appendix S1: Supporting Information