

Abstract
Background:
The genetic factors that modulate risk for developing lung cancer have not been fully defined. Here, we sought to determine the prevalence and clinical significance of germline pathogenic/likely pathogenic variants (PV) in patients with advanced lung cancer.
Methods:
We studied clinical and tumor characteristics of germline PV in 5118 patients who underwent prospective genomic profiling using paired tumor–normal tissue samples in 468 cancer genes.
Results:
Germline PV in high/moderate penetrance genes were observed in 222 (4.3%) patients; of these, 193 patients had PV in DNA damage repair (DDR) pathway genes including BRCA2 (n=54), CHEK2 (n=30) and ATM (n=26) that showed high rate of biallelic inactivation in tumors. BRCA2 heterozygotes with lung adenocarcinoma were more likely to be never smokers and had improved survival compared to non-carriers. Fourteen patients with germline PV in lung cancer predisposing genes (TP53, EGFR, BAP1 and MEN1) were diagnosed at younger age compared to non-carriers, and of tumor suppressors, 75% demonstrated bi-allelic inactivation in tumors. Significantly higher proportion of germline PV in high/moderate penetrance genes were detected in high-risk patients who had either a family history of any cancer, multiple primary tumors or early age at diagnosis compared to unselected patients (10.5% vs 4.1%; p-value=1.7e-04).
Conclusions:
These data underscore the biological and clinical importance of germline mutations in highly penetrant DDR genes as a risk factor for lung cancer.
Impact:
The family members of lung cancer patients harboring PV in cancer predisposing genes should be referred for genetic counseling and may benefit from proactive surveillance.
Keywords: Germline genetics, Lung cancer, Genotype-Phenotype association, Cancer Genomics
INTRODUCTION
Lung cancer is the leading cause of cancer mortality worldwide, with a five-year survival rate of 15%(1). Small-cell lung cancer (SCLC) accounts for 15%–17% of cases, and the various histologies that comprise non-small cell lung cancer (NSCLC including lung adenocarcinoma and squamous cell carcinoma) account for 85%(2). SCLC and NSCLC differ in risk factors, somatic mutational landscapes, invasiveness, and response to treatment. While tobacco smoking is a primary cause for both categories of lung cancer(3,4), only about 15% of lifelong smokers develop the disease, indicating there is significant individual variation in susceptibility. The smoking prevalence is much lower among adenocarcinoma patients compared to squamous and small cell lung cancer patients. Additionally, an estimated 10–25% of lung cancer cases occur in never-smokers (5) highlighting the potential role of genetic predisposition in lung cancer etiology. Lung cancer is significantly more common among those with a positive family history (odds ratio =1.57–2.7) and this is particularly true for young-onset lung cancers(6). Genetic background or ancestry may also influence the somatic evolution of lung cancer(7). Rare familial forms of lung cancer have been associated with germline pathogenic or likely pathogenic variants (germline PV) of EGFR, TP53, RB1, CDC147, BAP1, HER2, PARK2, and YAP1(8–10). Several lung cancer susceptibility loci with small effect sizes were identified using genome wide association studies (GWAS)(11–13). Recent studies reported that 2.5–4.5% of lung cancer patients carried germline PV that have been linked to cancer risk in Mendelian syndromes(14,15). Using whole exome sequencing, we and others have recently identified rare, deleterious variants in ATM, BRCA2 and Fanconi Anemia genes that associate with lung cancer risk (16) (17) (18). The increased use of clinical sequencing for both tumor profiling and germline testing has incidentally detected clinically actionable germline PV in lung cancer patients (19,20). While genetic risk models have been applied to inherited breast, ovarian, pancreatic, and prostate cancer(20–23), the clinicopathologic significance of germline PV in lung cancer has not been elucidated.
Here, we report the prevalence and clinical implications of germline PV in 5118 patients with lung cancer who underwent paired tumor-matched normal tissue sample clinical sequencing using the MSK-IMPACT assay(24). We performed integrated germline-somatic mutation data analyses to identify germline PV in lung cancer patients and correlated to individual and familial outcomes data to assess clinical and biological significance. Our study defined a subset of lung cancer patients at hereditary risk for a range of tumor types, including cases whose kindreds show clustering of TP53, ATM, or BRCA1/2 associated tumor types, as well as rarer subsets with lung tumor predispositions due to inherited mutations of EGFR, MEN1 or BAP1.
MATERIALS AND METHODS
Study Samples
In total, we analyzed 5118 patients with lung cancer at Memorial Sloan Kettering Cancer Center (MSKCC) who underwent tumor and normal DNA sequencing as part of their clinical care utilizing MSK-IMPACT (24)(Integrated Mutation Profiling of Actionable Cancer Targets) to interrogate 468 cancer-related genes from January 2014 until August 2019. All patients provided written informed consent and the study was approved by the Memorial Sloan Kettering Cancer Center Institutional Review Board (NCT01775072) and conducted in accordance with recognized ethical guidelines. Baseline clinical data including gender, histological subtypes, age at diagnosis (binned into five-year intervals), self-reported ancestry/ethnicity, smoking status (current/prior smoker versus never-smoker), and stage (metastatic or not) for all patients were obtained from institutional electronic medical records (EMR). A selected 152 patients who had either a family history of any cancer, personal history of multiple primary cancers or early age at diagnosis (≤55 years old) provided consent to return of germline results for up to 88 cancer predisposition genes tested on the targeted gene panel including all cancer susceptibility genes identified in American College of Medical Genetics and Genomics (ACMG) guidelines(25). The detailed clinical data and family history of any cancer were available for high-risk patients’ analysis. Patients with ≥2 primary cancers were identified through EMR database query, manually reviewed, and classified as multiple primaries based on IARC rules(26). Patients who received the platinum-based chemotherapy drugs cisplatin, carboplatin, or both, with or without other chemotherapies were identified through automated EMR database query.
Sequencing Analysis and Variant Interpretation
Tumor and blood lymphocyte DNA from patients were sequenced using the MSK-IMPACT platform interrogating up to 468 cancer-associated genes. The analytical methods including germline and somatic variant calling were performed as described in Srinivasan et.al (27) and Jonsson P. et, al. (28). The analytical approach is represented in the flow diagram (Supp. Figure S1). All patients were anonymized prior to germline variant pathogenicity assessment and linking of germline, somatic, and clinical data for the integrated analyses. The pathogenicity assessment for germline variants with <1% population allele frequency in the genome aggregate (gnomAD r2.1.1)(29) database was performed according to ACMG criteria(25). The germline variants annotated as pathogenic/likely pathogenic in CilnVar (January 2020)(25,30) were manually curated and only high confidence (ClinVar gold star >=2) pathogenic variants were reported here. Here, we identified and reported germline pathogenic variants in the full cohort including patients from European, Ashkenazi Jewish, Asian, African, and Others/unknown ancestries. We excluded germline variants likely derived from clonal hematopoiesis or from tumor-derived circulating cell-free DNA as previously described(28). We further classified germline PV according to the current disease modeling into high-penetrance (relative risk (RR) of disease>4); moderate-penetrance (RR 2–4); low-penetrance (RR<2); uncertain penetrance; or associated with an autosomal recessive condition as previously described (20). The common truncating variant BRCA2 p.Lys3326Ter and missense variant ATM p. Leu2307Phe reported in lung cancer GWAS as a risk allele(12,13) and CHEK2* p.Ile157Thr reported as protective allele(12) were excluded in the current analysis.
High-risk Lung Cancer Patients’ Analysis
The high-risk patients were defined as a lung cancer patient who had either a strong family history of any cancers, or personal history of multiple primary tumors, or early age at diagnosis (≤55 years). We compared the prevalence of germline pathogenic variants aggregated by genes with high/moderate or low penetrance, genes with uncertain significance or recessive between high-risk patients and unselected patients and performed Fisher’s exact test to report the odds ratio and p-value.
Loss of Heterozygosity in Tumor and Somatic Driver Alteration Association Test
Tumor sequence data were analyzed as previously described to detect somatic mutations, small insertions and deletions (indels), DNA copy number alterations, and select translocations(31). Loss of heterozygosity (LOH) in the tumor at the locus of the detected germline PV was evaluated using the FACETS algorithm(32). Modeling of zygosity and filtering of these calls was performed as previously described(27,28). The zygosity for each germline variant in tumor was determined to be heterozygous, loss of the wild type (WT) allele or loss of the mutant allele. Patients harboring germline PV were considered to have biallelic inactivation if either loss of WT allele was observed or a second somatic truncating mutation was detected in their tumors. Since our method could not phase a second somatic truncating mutation with a germline variant, we considered the zygosity of a germline variant as indeterminate if two or more detectable somatic pathogenic events (e.g., patients with germline PV who had both loss of WT allele and a somatic truncating mutation in tumors were excluded). We estimated the rate of biallelic inactivation as the number of germline mutation carriers who had biallelic inactivation compared to total number of evaluable germline mutation carriers (after excluding indeterminate cases). We next assessed the enrichment of biallelic inactivation (either loss of WT allele or a second somatic truncating mutation) in patients carrying germline PV compared to patients without any germline PV (germline PV absent) and carrying non-pathogenic variants such as benign/likely benign variants that did not have any functional consequence (benign/likely benign variants annotated in ClinVAR database(January 2020)(25,30)). The distribution of biallelic inactivation in germline benign/likely benign variants was determined in the corresponding tumor in a manner identical to that of pathogenic variants as described above. We annotated somatic mutations as oncogenic or likely oncogenic using OncoKB, a precision oncology knowledge base(28). We performed all analyses separately for histological subtypes (lung adenocarcinoma or lung squamous cell carcinoma) since there are significant differences in mutational landscapes between them. We compared frequency of key somatic driver mutations, oncogenic fusions, and copy number alterations in germline PV carriers and compared with patients without any germline mutations.
Statistical and Survival Analysis
The binned clinical data were anonymized before linking to genomic data. Clinical characteristics of patients with germline PV aggregated by gene were compared with those without any germline PV using Fisher’s exact test. To assess the associations between germline PV carrier status and clinical characteristics or somatic driver mutations, logistic regression was used for binary phenotypes. All patients (including Ashkenazi Jewish patients) were analyzed for association tests and self-reported ancestry was used as covariate in the multivariate model. Multiple linear regression was performed for quantitative traits to adjust for clinical variables including sex, self-reported ancestry (European, Asian, African or others), smoking status (never vs current/former smoker), age at diagnosis and tumor stage (metastatic yes/no). All analyses were performed separately for patients with lung adenocarcinoma, lung squamous cell carcinoma and small cell lung cancer. In total, 15 genes with ≥5 patients carrying germline PV were tested for association analysis, and using Bonferroni correction for multiple testing, the results with p-value < 0.003 were reported as statistically significant.
Overall survival (OS) data was available and analyzed for 1876 patients with lung adenocarcinoma. OS was measured from the date of diagnosis to the date of death and censored at the date of last follow-up for survivors. Survival times were binned for the purpose of anonymization such that each time interval had more than 15 patients. To assess association between OS and germline PV status, univariate Cox regression was performed with likelihood ratio tests. Because survival times were binned to preserve anonymization of data, point estimates for survival times were chosen randomly within the time interval bins. 100 replicates were performed to assure stability of estimates (Supp. Figure S2). The proportional hazards assumption was tested with Schoenfeld residuals. Kaplan-Meier curves were utilized to visualize survival differences using the R package “survival” in R software version 3.4.2. Multivariable Cox regression was performed to assess the independent association of germline PV grouped by specific genes with OS using age at diagnosis (binned as 5-year interval time), sex, smoking status (current/former smoker vs never-smoker), stage (metastatic or not), self-reported ancestry (European, Asian, African or others) and platinum-based chemotherapy (platinum-based chemotherapy/non-platinum/none). The overall survival analysis was not possible for the other subtypes because of insufficient sample size.
Estimation of Carrier Frequencies of Germline PV aggregated at gene-level in Population Database
To estimate the carrier frequency of germline PV aggregated at gene-level in general population, we used the genome aggregate (gnomAD r2.1.1) database(29) as described in recent publications(33–35). We annotated germline variants in the gnomAD database as PV using the same variant interpretation pipeline as applied to our patient cohort. The carrier frequency of germline PV in patients with lung adenocarcinoma or with squamous cell carcinoma from European ancestry were computed separately and compared with the estimated carrier frequencies computed using non-Finnish European individuals from gnomAD. We excluded patients with self-reported Ashkenazi Jewish (AJ) ancestry and carriers of AJ and European founder mutations for this comparison (burden analysis).
Data Availability
Raw genomic data for this study were generated at Molecular Diagnostics Service in the Department of Pathology, the Integrated Genomics Operation Core and Marie-Josée and Henry R. Kravis Center for Molecular Oncology at Memorial Sloan Kettering Cancer Center (ClinicalTrials.gov Identifier: NCT01775072). Derived summary data supporting the findings of this study are available from the corresponding author upon request.
RESULTS
Patient Characteristics
We evaluated a cohort of 5118 patients with advanced lung cancer consenting to a research study utilizing tumor-normal clinical sequencing targeted to a panel of 468 cancer-associated genes (MSK-IMPACT assay(24)). Germline-somatic variant and clinical data were anonymized for this study. Clinical characteristics of the patients are summarized in Table 1. Most patients (n=3886; 75.9%) had a diagnosis of lung adenocarcinoma (LUAD); 517 (10.1%) cases had lung squamous cell carcinoma (LUSQ) and 275 (5.4%) cases had small cell lung cancer (SCLC) (Table 1). Smoking data were available for 91% of the patients. Most SCLC and LUSQ patients were current or former smokers (ever-smokers) in our cohort. Among those diagnosed with lung adenocarcinoma, 2611 (67.5%) patients were classified as ever-smokers. A selected 152 patients who had either a family history of any cancer, personal history of multiple primary tumors or early age at diagnosis (≤55 years) were consented to receive both tumor and germline sequencing results and identified as high-risk patients (Supp Table S1). Of these, 79% had a family history of any cancer. The median age at diagnosis for these 152 patients was 64 years compared to 70 years for the unselected cohort (p<0.002).
Table 1:
Patient Characteristics and histological subtypes
| Characteristics | LUAD (n=3886) | LUSQ (n=517) | SCLC (n=275) | Other Histological Subtypes (n=440) |
|---|---|---|---|---|
| Median age at diagnosis (years) | 70 | 75 | 70 | 70 |
| Sex | ||||
| Male | 1509 (38.9%) | 336 (65%) | 142 (51.6%) | 200 (45.5%) |
| Female | 2377 (60.1%) | 181 (35%( | 133 (48.4%) | 240 (55.5%) |
| Smoking History | ||||
| Smoker (current or former) | 2611 (67.2%) | 433 (83.7%) | 203 (73.8%) | 296 (62.2%) |
| Non-smoker | 1006 (25.9%) | 22 (4.2%) | 15 (5.4%) | 71 (16.1%) |
| Unknown/missing data | 269 (6.9%) | 62 (12%) | 57 (20.7%) | 73 (16.6%) |
| Ethnicity (self-reported) | ||||
| European/White/Non-Hispanic | 2430 (62.5%) | 377 (72.9%) | 218 (79.2%) | 293 (66.6%) |
| Ashkenazi Jewish | 643 (16.5%) | 53 (10.2%) | 24 (8.7%) | 54 (12.3%) |
| Asian | 399 (10.3%) | 27 (5.2%) | 15 (5.4%) | 24 (5.4%) |
| African | 191 (4.9%) | 30 (5.8%) | 8 (2.9%) | 34 (7.7%) |
| Others/Unknown | 223 (5.7%) | 30 (5.8%) | 10 (3.6%) | 35 (7.9%) |
| Disease Stage | ||||
| Non- Metastatic | 484 (12.5%) | 44 (8.5%) | 18 (6.5%) | 112 (25.5%) |
| Metastatic | 3341 (86%) | 462 (89.3%) | 251 (91.3%) | 318 (72.2%) |
| Missing data | 61 (1.8%) | 11 (2.1%) | 6 (2.2%) | 10 (2.3%) |
| Personal History of Multiple primary tumors | ||||
| Yes | 1023 (26.3%) | 133 (25.7%) | 58 (21%) | 107 (24.3%) |
Abbreviation: LUAD= Lung Adenocarcinoma, LUSQ= Lung Squamous Cell Carcinoma, SCLC= Small Cell Lung Cancer
Prevalence of Germline Pathogenic Variants (PV)
In our cohort of 5118 patients with lung cancer, 584 harbored one or more germline PV in 49 genes including 462 patients diagnosed with lung adenocarcinoma, 57 patients with squamous cell carcinoma, 29 patients with small cell lung cancer and 34 patients who had other histological subtypes (Supp. Table S2). Of all germline PV carriers, 222 (4.3%) harbored germline PV in high/moderate penetrance genes, 152 harbored uncertain penetrance PV and 210 patients carried low penetrance or recessive PV; 36 patients had 2 or more germline PV (Figure 1a, and Supp. Table S3). Thirty-five patients harbored germline PV in genes previously associated with risk for lung cancer including PARK2 p.Arg275Trp (n=21), TP53 (n=7), BAP1 (n=3), EGFR p.Thr790Met (n=2), and MEN1 (n=2) (Figure 1b). In addition to lung cancer risk genes, germline PV were most frequently observed in DNA damage repair (DDR) pathway genes including high/moderate penetrance genes BRCA2 (n=54), BRCA1 (n=34), CHEK2 (n=30) and ATM (n=26) (Figures 1b, Supp. Table S2). Almost all patients carrying ATM germline PV were diagnosed with lung adenocarcinoma (n=24/25). The frequency of germline PV in BRCA2 was significantly higher in lung squamous cell carcinoma (1.9%) compared to other histologies after adjusting for age at diagnosis, gender, smoking status, stage, and ethnicity (p-value=0.03).
Figure 1:
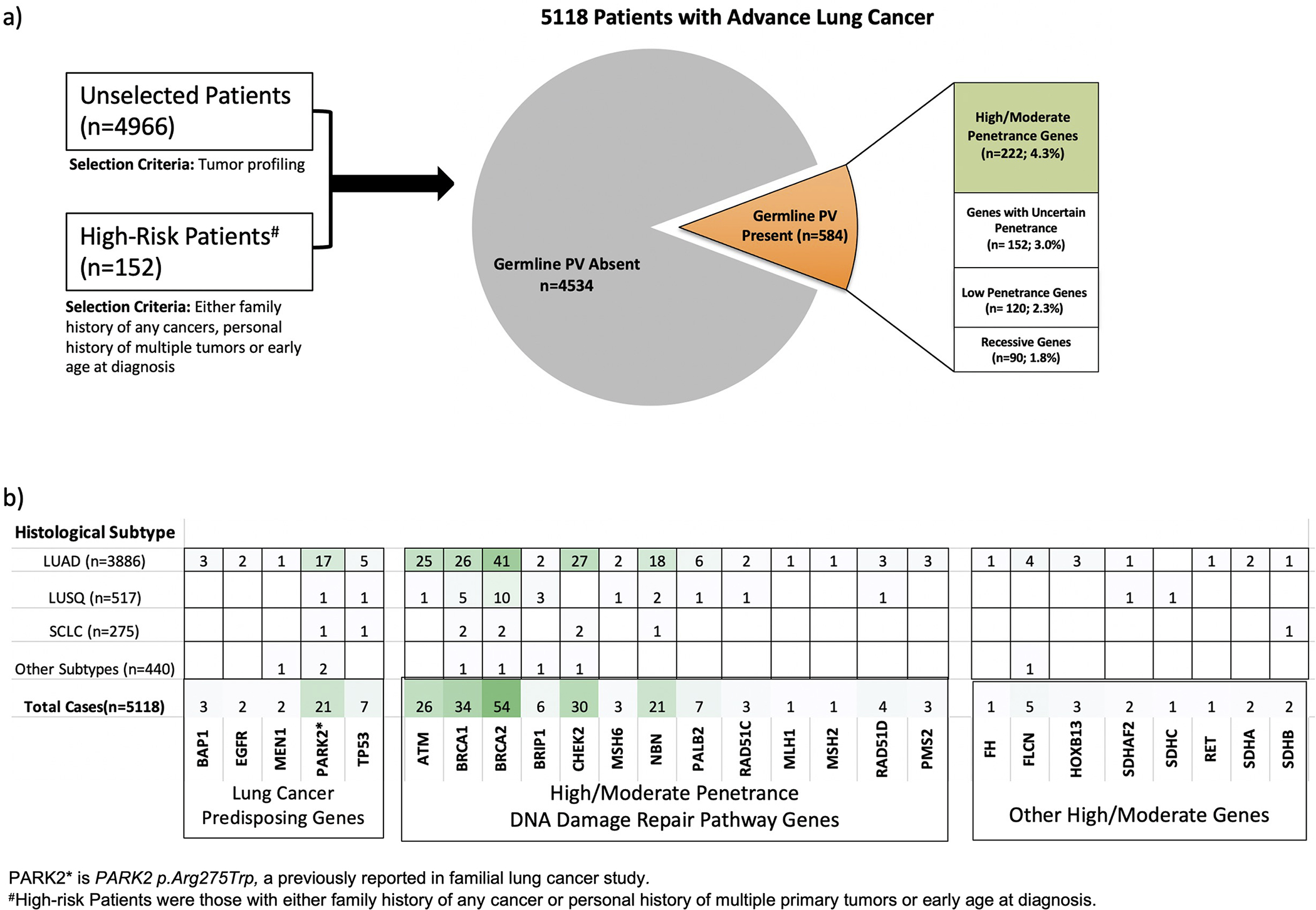
Distribution germline PV in 5118 lung cancer patients. a) The relative frequencies of germline PV in genes classified by penetrance; b) Number of patients carrying germline PV in lung cancer predisposing genes or high/moderate penetrance genes and diagnosed with Lung Adenocarcinoma (LUAD), Lung Squamous Cell Carcinoma (LUSQ), Small Cell Lung Cancer (SCLC) and Other Subtypes.
Among 152 high risk lung cancer patients who had either family history of any cancer, personal history of multiple primary tumors, or early age of diagnosis (≤55 years old), 26 patients harbored germline PV; of these 25 patients and their families were referred for screening and counseling. A significantly higher proportion of germline PV in high/moderate penetrance genes were detected in patients clinically identified as high-risk compared to patients in the unselected cohort (10.5% vs 4.1%; odds ratio=3.0, 95% CI= 1.7–5.2, p-value=1.7e-04) (Supp. Table S4).
Somatic Characteristics of Germline PV Carriers
The biological impact of germline PV on lung tumors was assessed using Knudson’s two-hit hypothesis for bi-allelic inactivation(36). Biallelic inactivation of germline PV was considered if either the wild type (WT) allele was lost in the tumor (loss of heterozygosity) or a second somatic truncating mutation was observed. We excluded germline PV carriers with two or more detectable somatic pathogenic events in the same gene (e.g., presence of both somatic truncating mutation and loss of WT allele) to eliminate the bias that biallelic inactivation may be resulted by somatic mutations. Among patients with lung adenocarcinoma, biallelic inactivation was observed in 75% of patients harboring germline PV in high-penetrance lung cancer predisposing tumor suppressors (TP53, BAP1, and MEN1 in Figure 2a). Biallelic inactivation in four DDR genes (ATM, BRCA2, CHEK2, and MRE11A) was observed in greater than 40% of carriers whereas only 14% of BRCA1 germline PV carriers showed biallelic inactivation (Figure 2a) in patients with lung adenocarcinoma. ATM and CHEK2 had significantly higher rate of biallelic inactivation in lung adenocarcinoma patients harboring germline PV compared to the patients harboring benign/likely benign variants (56.5% vs 26.2% in ATM and 51.8% vs 20.4% in CHEK2; p-values <0.005 statistically significant after correcting for multiple testing; Figure 2b). In the stratified analysis using smoking history of lung adenocarcinoma patients, we observed that among never smokers, BRCA2 showed marginally higher rate of biallelic inactivation in patients carrying germline PV compared to patients with benign/likely benign variants at the same locus (47.3% vs 25.4%, p-value=0.06: Supp. Table S5). The biallelic inactivation in lung squamous cell carcinoma patients carrying germline PV in TP53, RAD51D, BRIP1 and BRCA2 were observed in 60% of the patients (Figure 2c).
Figure 2:
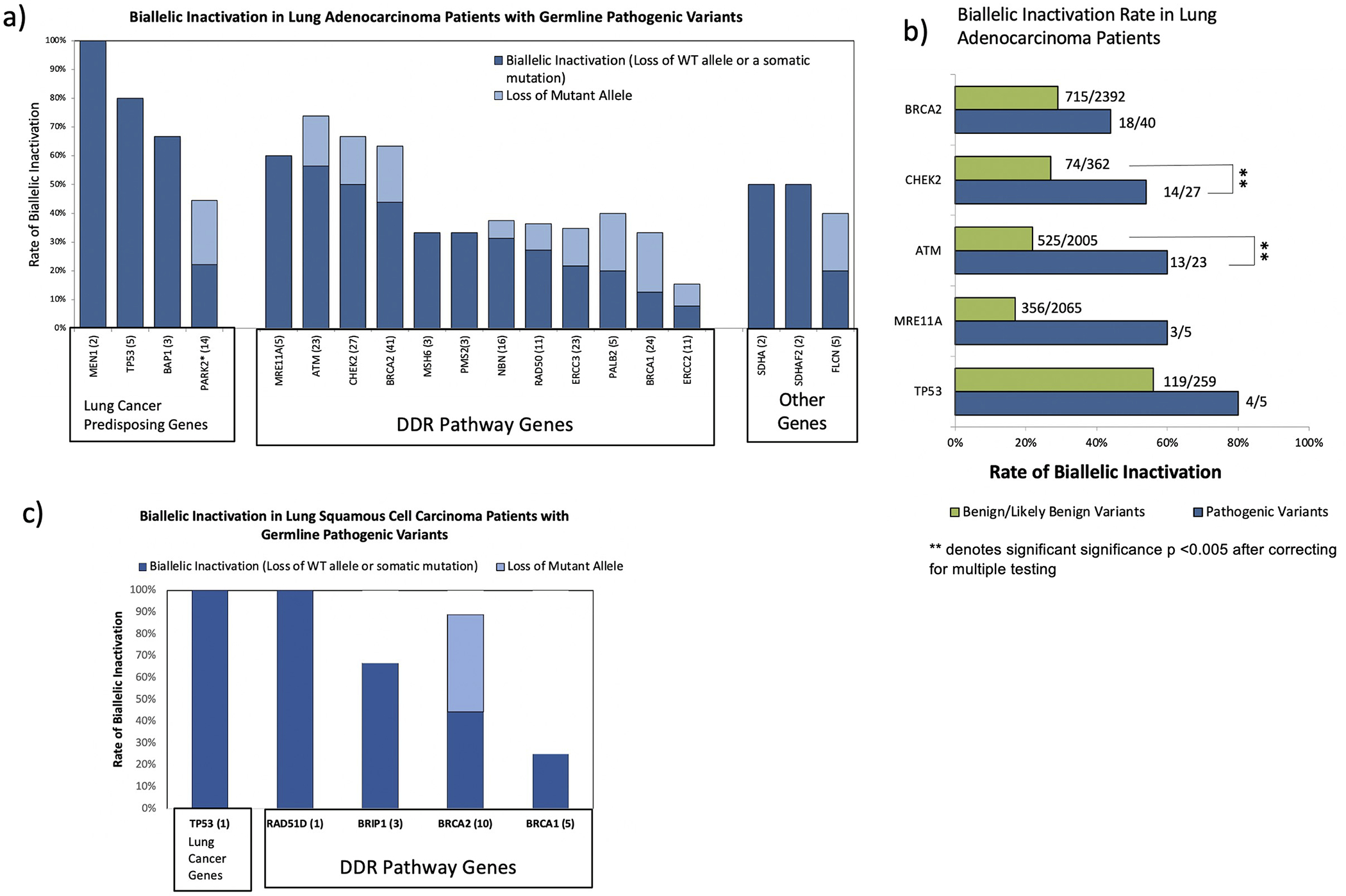
Distribution of biallelic inactivation in patients with germline PV. a) Biallelic inactivation rate in patients with lung adenocarcinoma; b) Biallelic inactivation rate in patients with germline PV compared to biallelic inactivation rate in patients harboring benign/likely benign variants in the same genes; c) Biallelic inactivation rate in patients with lung squamous cell carcinoma
Tumor mutation burden and the frequencies of somatic driver mutations or somatic copy number alterations were not significantly different between patients with and without germline PV after correcting for multiple testing.
Association with Clinical Characteristics and Overall Survival
Twenty-eight lung adenocarcinoma patients who carried germline PV in lung cancer risk genes (TP53, BAP1, MEN1, EGFR and PARK2) had significantly younger age at diagnosis compared to patients without any germline PV, after adjusting for gender, smoking status, ethnicity, and tumor stage (p-value=0.0008, Figure 3a). Excess of germline PV in BRCA1 and BRCA2 were observed in lung adenocarcinoma patients who were never smokers compared to current/former smokers, based on a multivariate model (adjusted odds ratio=1.3, 95% CI=1.1–1.6 p-value <0.001, Figure 3b). The germline PV status was not associated with other clinical features in patients with lung adenocarcinoma or other histologies. We observed that the prevalence of germline PV in patients from Ashkenazi Jewish ancestry was significantly higher than that observed in patients from European, Asian, or African ancestry; reflective of presence of founder mutations in patients from Ashkenazi Jewish ancestry (Supp. Table S6 and Supp. Table S7)
Figure 3.
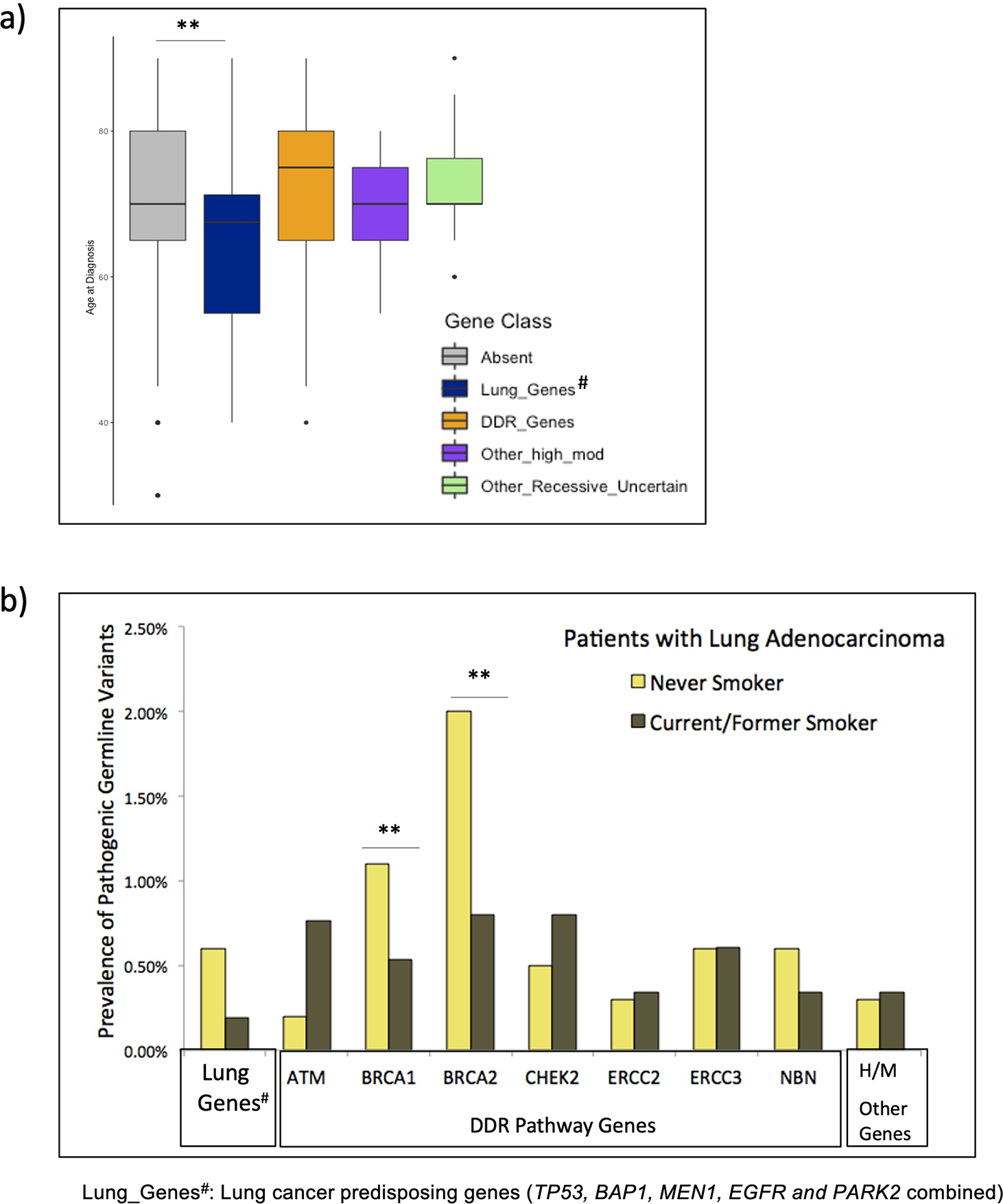
Association of germline PV with clinical characteristics in lung adenocarcinoma patients. a) Association with age of cancer diagnosis; b) Association with smoking history (never-smoker vs current/former smokers)
An exploratory overall survival (OS) analysis was performed for 1876 advanced stage lung adenocarcinoma patients using carrier status for germline PV. The mutational status of high/moderate penetrance genes or DDR pathway genes (combined) was not associated with OS. However, gene-centric analysis showed that carrier status for PV in BRCA2 was associated with improved OS (HR=0.29, 95%CI=0.07–0.99, p-value = 0.03, Figure 4a). In multivariable Cox regression analysis stratified by smoking history, which did not satisfy the proportional hazards assumption (Supp. Table S8), and adjusted by age at diagnosis, sex, tumor stage (metastatic yes/no), self-reported ancestry/ethnicity, and receipt of platinum-based chemotherapy (cisplatin or carboplatin treated/other chemotherapy/none), the BRCA2 germline PV carriers were marginally associated with OS (adjusted HR=0.26, 95% CI=0.07–1.0, p-value=0.06) (Figure 4b); additional studies are needed. The germline mutational status of other genes including ATM, BRCA1, or CHEK2 was not associated with OS (Supp. Figure S3).
Figure 4:
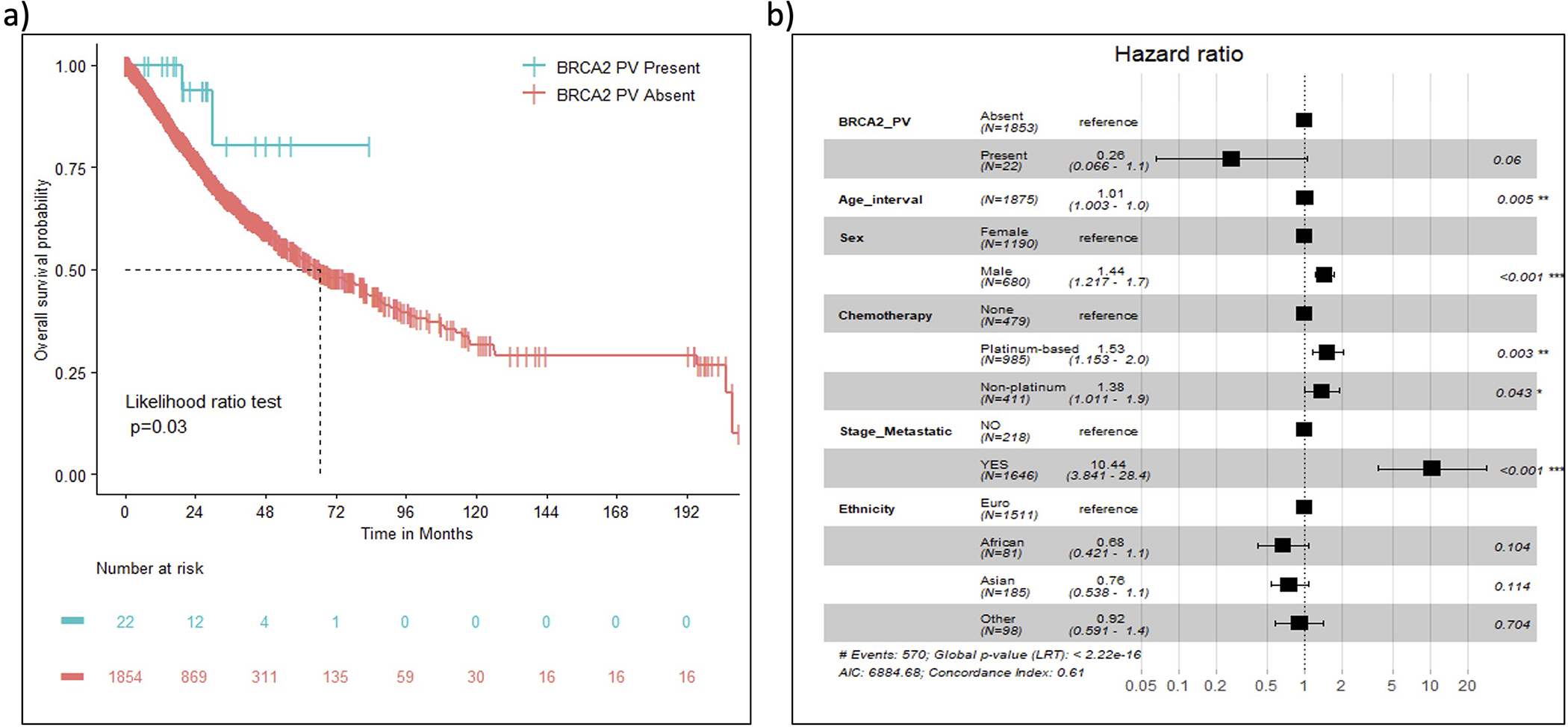
Overall survival analysis in patients with advance lung adenocarcinoma a) Kaplan-Meier survival curves comparing carriers of germline PV in BRCA2 (n=22) and non-carriers(n=1854); b) Multivariate Cox proportional hazard regression analyses
Comparison to population database
We observed that the carrier frequencies of germline PV in genes ATM and BRCA2 in patients with lung adenocarcinoma from European ancestry were statistically higher than expected frequencies observed in individuals from non-Finnish European population in gnomAD database (odds ratio=2.3; 95% CI=1.3–3.6 for ATM and odds ratio= 2.2, 95% CI=1.3–3.4 for BRCA2; p-values =0.002, Supp. Table S9). We also observed higher than expected frequency of germline PV in BRCA2 in lung squamous cell carcinoma patients compared to individuals from non-Finnish European population in gnomAD population database (33,37) (odds ratio=5.8, 95% CI=2.3–12.2, p-value=0.0003) (Supp. Table S9).
DISCUSSION
Here, we report a large integrated analysis of germline and somatic mutations in 5118 patients with advanced lung cancer, identifying germline PV in high or moderate penetrance genes in 4.3% of patients. Significantly increased prevalence of germline PV in high/moderate penetrance genes was observed in patients with either a family of any cancers, personal history of multiple primary tumors or early age of diagnosis (≤55 years age). In our cohort, patients harboring germline PV in lung cancer predisposing genes (TP53, EGFR, BAP1, and MEN1) had earlier age at diagnosis compared to patients without germline PV; 75% of these with germline PV in tumor suppressor gene (TP53, BAP1, and MEN1) demonstrated bi-allelic inactivation in their tumors. Germline PV in BAP1 a mesothelioma susceptibility gene(38) was identified in 3 lung adenocarcinoma patients with two of them demonstrating loss of heterozygosity in their tumors – suggesting a role of BAP1 in a subset of lung adenocarcinoma.
The most frequently mutated genes were associated with DDR pathway including ATM, BRCA1, BRCA2 and CHEK2 that were typically associated with breast, ovarian, pancreatic, prostate, and other cancer susceptibility(39–42). Our data support previous findings that ATM and BRCA2 constitute lung cancer susceptibility genes (13,16,18,43). The observation that among lung adenocarcinoma cases, 56.5% of ATM germline PV carriers, 51.8% of CHEK2 germline PV carriers and 45% of BRCA2 germline PV carriers demonstrated bi-allelic inactivation in their tumors supports their biological roles in lung cancer etiology. We observed higher than expected frequencies of germline PV in ATM and BRCA2 in patients with lung adenocarcinoma; and BRCA2 in patients with lung squamous cell carcinoma. BRCA1 and BRCA2 heterozygotes with lung adenocarcinoma were more likely to be never-smokers. Lung adenocarcinoma patients with germline BRCA2 mutations had an improved overall survival presumably due to improved outcome following platinum therapies, however, additional studies are needed due to limited sample size to clarify the interactions between germline PV and treatment response in lung cancer patients. Recently, a report of 87 patients with small cell lung cancer demonstrated that the subset with inherited mutations in DDR genes including BRCA2 had more favorable responses to platinum-based chemotherapy(43). Our findings also suggested that germline PV in BRCA1 and BRCA2 in lung cancer patients might have distinct consequences on lung tumors. While 193 lung cancer patients harbored germline PV in high/moderate penetrance genes in DDR pathway, it also remains unclear if such patients will benefit from trials of targeted therapies, as observed for patients with DDR deficiencies and breast, ovarian, pancreatic, prostate, and other cancer types (28).
The observed prevalence of all germline PV in the current NSCLC patient cohort (n=584; 11.4%) is somewhat higher than that reported in the larger pan-cancer study led by TCGA (8%)(44) and similar prevalence in small cell lung cancer was reported in a recent study(43). Given that the TCGA study was primarily based on early stage resected cancers, the difference may reflect an ascertainment bias given the high proportion of lung cancer patients with advanced metastatic disease referred for tumor profiling in our cohort; we have previously shown an enrichment in germline PV in metastatic compared to non-metastatic tumors of various types (20). A limitation of the current study was the incomplete annotation of family history typically assessed at time of genetic counseling, as the vast majority of lung cancer patients reported here, in contrast to patients with other cancer types seen at our center (20), did not receive germline findings or follow-up genetic counseling. Sequencing analysis was also limited to 468 cancer-associated genes, however this included 88 known ACMG cancer predisposition genes. We utilized publicly available population data (gnomAD r2.1.1) to compute the estimated carrier frequencies of germline PV by gene as previously described in recent publications(33–35), however we acknowledge the limitation of such methods due to technical differences in sequencing platforms/bioinformatic pipelines and the presence of population stratification as confounders. We reported survival analysis for only 1876 patients with advanced lung adenocarcinoma due to limited data availability. Additional studies are warranted to address the effects of germline PV status on response to therapy and clinical outcome.
In conclusion, our study systemically characterized the germline PV in cancer predisposing genes in patients with lung cancer, emphasizing the role of genetic counseling for unaffected family members who may benefit from proactive surveillance. The study expands the known lung cancer susceptibility genes to include ATM and BRCA2 in addition to TP53, EGFR, and MEN1, and provides supporting evidence for BAP1, a gene previously associated with mesothelioma. Additional research is warranted to further establish the criteria for genetic counseling in lung cancer patients to improve precision oncology practices. Screening with low-dose CT may be incorporated into the care of the high-risk patients with lung cancer and their unaffected relatives carrying germline mutations independent of smoking history. Our study also demonstrates the biological importance of germline mutations in DDR genes in patients with advance lung cancer. Such patients may benefit from personalized therapeutic modalities as several DDR inhibitors have been developed and are either approved for other cancer types (e.g., PARP inhibitors) or are in clinical trials. Additional studies will provide a better understanding of response to such therapies in lung cancer cohorts defined by germline PV status. The identification of germline PV in highly penetrant, clinically actionable genes in 4.3% of patients with lung cancer, particularly those with early age at diagnosis (≤55 years), multiple primary tumors, or a family history of cancer, is important given the high global incidence of lung cancer and the potential benefits of cascade testing for relatives(45) to guide surveillance for early cancer detection.
Supplementary Material
Acknowledgments:
We would like to thank all individuals who participated in this study. We gratefully acknowledge the members of the Molecular Diagnostics Service in the Department of Pathology and the use of the Integrated Genomics Operation Core and Clinical Genetics Service at Memorial Sloan Kettering Cancer Center. K. Offit received the grants from Robert and Kate Niehaus Center for Inherited Cancer Genomics, the Andrew Sabin Family Foundation, and the Sharon Levine Corzine Research Fund; MSKCC is supported by the NCI Core grant P30 CA008748; C.I. Amos is a research scholar of the Cancer Prevention and Research Institute of Texas and supported by RR170048 and U19CA203654; M.F. Berger received the funding NIH/NCI 1R01CA227534-01A; D. R. Jones received the NIH/NCI awards 1R01 CA217169 and 1R01 CA234617; D. B. Solit received the grant Cycle for Survival and Marie-Josée and Henry R. Kravis Center for Molecular Oncology; and Z.H. Gümüş received award from the LUNGevity Foundation Fund.
Conflict of interest disclosure statement:
Disclosures: MGZ has received consulting fees from Ikena and honoraria for CME content from PER and Medscape. Dr. Zauderer employer, Memorial Sloan Kettering, receives research funding from the DOD, NIH, PRECOG, GSK, Epizyme, Polaris, Sellas Life Sciences, Bristol Myers Squibb, Millenium/Takeda, Curis and Atara for research conducted by MGZ. None of this funding is related to the current work. Dr. Zauderer serves as the Chair of the Board of Directors of the Mesothelioma Applied Research Foundation, an uncompensated position. MDH reports grants from BMS; and personal fees from Achilles; Adagene; Adicet; Arcus; AstraZeneca; Blueprint; BMS; DaVolterra; Eli Lilly; Genentech/Roche; Genzyme/Sanofi; Janssen; Immunai; Instil Bio; Mana Therapeutics; Merck; Mirati; Natera; Pact Pharma; Shattuck Labs; and Regeneron; as well as equity options from Factorial, Immunai, Shattuck Labs, Arcus, and Avail Bio. A patent filed by Memorial Sloan Kettering related to the use of tumor mutational burden to predict response to immunotherapy (PCT/US2015/062208) is pending and licensed by PGDx. Subsequent to the completion of this work, MDH began as an employee (and equity holder) at AstraZeneca. KAC serves in advisory board of Astra Zeneca, MJH Life Sciences, MSD Ireland, GSK Ireland Nextcure, receives funding and educational support from MSD Ireland, Immunogen, Pfizer and Roche Ireland. YLL discloses research funding from GSK, REPARE and AstraZeneca unrelated to this work. MER reports honoraria from Research to Practice, Intellisphere, Physicians’ Education Resource, and MyMedEd; consulting or advisory for Artios Pharma (uncompensared), AstraZeneca (uncompensated), Change Healthcare, Daiichi-Sankyo (uncompensated), Epic Sciences (uncompensated), Merck (uncompensated), Pfizer (uncompensated), Tempus Lab (uncompensated), Zenith Pharma (uncompensated); research funding from Breast Cancer Research Foundation, AstraZeneca (Institution), Merck (Institution), and Pfizer (institution); and other support from AstraZeneca (editorial services) and Pfizer (editorial services). DBS has consulted with/received honoraria from Pfizer, Loxo/Lilly Oncology, Vividion Therapeutics, FORE Therapeutics, Scorpion Therapeutics and BridgeBio. MFB reports consulting fees from Eli Lilly and PetDx, both unrelated to the current work. SML and KO are co-founders of AnaNeo Therapeutics, Inc, which does not bear on the work presented. CMR has consulted regarding oncology drug development with AbbVie, Amgen, Astra Zeneca, Daiichi Sankyo, Epizyme, Genentech/Roche, Ipsen, Jazz, Kowa, Lilly, Merck, and Syros. He serves on the scientific advisory boards of Bridge Medicines, Earli, and Harpoon Therapeutics. PS is currently an employee of Natera Inc. BST reports advisory board activities for Boehringer Ingelheim and Loxo Oncology at Lilly and research support from Genentech, all unrelated to the current work. All other authors declare no potential conflicts of interest.
References:
- 1.Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin 2018;68(6):394–424 doi 10.3322/caac.21492. [DOI] [PubMed] [Google Scholar]
- 2.Travis WD, Brambilla E, Nicholson AG, Yatabe Y, Austin JHM, Beasley MB, et al. The 2015 World Health Organization Classification of Lung Tumors: Impact of Genetic, Clinical and Radiologic Advances Since the 2004 Classification. J Thorac Oncol 2015;10(9):1243–60 doi 10.1097/JTO.0000000000000630. [DOI] [PubMed] [Google Scholar]
- 3.Bade BC, Dela Cruz CS. Lung Cancer 2020: Epidemiology, Etiology, and Prevention. Clin Chest Med 2020;41(1):1–24 doi 10.1016/j.ccm.2019.10.001. [DOI] [PubMed] [Google Scholar]
- 4.Burris JL, Studts JL, DeRosa AP, Ostroff JS. Systematic Review of Tobacco Use after Lung or Head/Neck Cancer Diagnosis: Results and Recommendations for Future Research. Cancer Epidemiol Biomarkers Prev 2015;24(10):1450–61 doi 10.1158/1055-9965.EPI-15-0257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Rivera GA, Wakelee H. Lung Cancer in Never Smokers. Adv Exp Med Biol 2016;893:43–57 doi 10.1007/978-3-319-24223-1_3. [DOI] [PubMed] [Google Scholar]
- 6.Brennan P, Hainaut P, Boffetta P. Genetics of lung-cancer susceptibility. Lancet Oncol 2011;12(4):399–408 doi 10.1016/S1470-2045(10)70126-1. [DOI] [PubMed] [Google Scholar]
- 7.Carrot-Zhang J, Soca-Chafre G, Patterson N, Thorner AR, Nag A, Watson J, et al. Genetic ancestry contributes to somatic mutations in lung cancers from admixed Latin American populations. Cancer Discov 2020. doi 10.1158/2159-8290.CD-20-1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Xiong D, Wang Y, Kupert E, Simpson C, Pinney SM, Gaba CR, et al. A recurrent mutation in PARK2 is associated with familial lung cancer. Am J Hum Genet 2015;96(2):301–8 doi 10.1016/j.ajhg.2014.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Liu Y, Kheradmand F, Davis CF, Scheurer ME, Wheeler D, Tsavachidis S, et al. Focused Analysis of Exome Sequencing Data for Rare Germline Mutations in Familial and Sporadic Lung Cancer. J Thorac Oncol 2016;11(1):52–61 doi 10.1016/j.jtho.2015.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Walpole S, Pritchard AL, Cebulla CM, Pilarski R, Stautberg M, Davidorf FH, et al. Comprehensive Study of the Clinical Phenotype of Germline BAP1 Variant-Carrying Families Worldwide. J Natl Cancer Inst 2018;110(12):1328–41 doi 10.1093/jnci/djy171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.McKay JD, Hung RJ, Han Y, Zong X, Carreras-Torres R, Christiani DC, et al. Large-scale association analysis identifies new lung cancer susceptibility loci and heterogeneity in genetic susceptibility across histological subtypes. Nat Genet 2017;49(7):1126–32 doi 10.1038/ng.3892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wang Y, McKay JD, Rafnar T, Wang Z, Timofeeva MN, Broderick P, et al. Rare variants of large effect in BRCA2 and CHEK2 affect risk of lung cancer. Nat Genet 2014;46(7):736–41 doi 10.1038/ng.3002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ji X, Mukherjee S, Landi MT, Bosse Y, Joubert P, Zhu D, et al. Protein-altering germline mutations implicate novel genes related to lung cancer development. Nat Commun 2020;11(1):2220 doi 10.1038/s41467-020-15905-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Parry EM, Gable DL, Stanley SE, Khalil SE, Antonescu V, Florea L, et al. Germline Mutations in DNA Repair Genes in Lung Adenocarcinoma. J Thorac Oncol 2017;12(11):1673–8 doi 10.1016/j.jtho.2017.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hu X, Yang D, Li Y, Li L, Wang Y, Chen P, et al. Prevalence and clinical significance of pathogenic germline BRCA1/2 mutations in Chinese non-small cell lung cancer patients. Cancer Biol Med 2019;16(3):556–64 doi 10.20892/j.issn.2095-3941.2018.0506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Esai Selvan M, Zauderer MG, Rudin CM, Jones S, Mukherjee S, Offit K, et al. Inherited Rare, Deleterious Variants in ATM Increase Lung Adenocarcinoma Risk. J Thorac Oncol 2020. doi 10.1016/j.jtho.2020.08.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Esai Selvan M, Klein RJ, Gumus ZH. Rare, Pathogenic Germline Variants in Fanconi Anemia Genes Increase Risk for Squamous Lung Cancer. Clin Cancer Res 2019;25(5):1517–25 doi 10.1158/1078-0432.CCR-18-2660. [DOI] [PubMed] [Google Scholar]
- 18.Liu Y, Xia J, McKay J, Tsavachidis S, Xiao X, Spitz MR, et al. Rare deleterious germline variants and risk of lung cancer. NPJ Precis Oncol 2021;5(1):12 doi 10.1038/s41698-021-00146-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lincoln SE, Nussbaum RL, Kurian AW, Nielsen SM, Das K, Michalski S, et al. Yield and Utility of Germline Testing Following Tumor Sequencing in Patients With Cancer. JAMA Netw Open 2020;3(10):e2019452 doi 10.1001/jamanetworkopen.2020.19452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mandelker D, Zhang L, Kemel Y, Stadler ZK, Joseph V, Zehir A, et al. Mutation Detection in Patients With Advanced Cancer by Universal Sequencing of Cancer-Related Genes in Tumor and Normal DNA vs Guideline-Based Germline Testing. JAMA 2017;318(9):825–35 doi 10.1001/jama.2017.11137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Nicolosi P, Ledet E, Yang S, Michalski S, Freschi B, O’Leary E, et al. Prevalence of Germline Variants in Prostate Cancer and Implications for Current Genetic Testing Guidelines. JAMA Oncol 2019;5(4):523–8 doi 10.1001/jamaoncol.2018.6760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Shindo K, Yu J, Suenaga M, Fesharakizadeh S, Cho C, Macgregor-Das A, et al. Deleterious Germline Mutations in Patients With Apparently Sporadic Pancreatic Adenocarcinoma. J Clin Oncol 2017;35(30):3382–90 doi 10.1200/JCO.2017.72.3502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Stadler ZK, Maio A, Chakravarty D, Kemel Y, Sheehan M, Salo-Mullen E, et al. Therapeutic Implications of Germline Testing in Patients With Advanced Cancers. J Clin Oncol 2021:JCO2003661 doi 10.1200/JCO.20.03661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Cheng DT, Prasad M, Chekaluk Y, Benayed R, Sadowska J, Zehir A, et al. Comprehensive detection of germline variants by MSK-IMPACT, a clinical diagnostic platform for solid tumor molecular oncology and concurrent cancer predisposition testing. BMC Med Genomics 2017;10(1):33 doi 10.1186/s12920-017-0271-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Richards S, Aziz N, Bale S, Bick D, Das S, Gastier-Foster J, et al. Standards and guidelines for the interpretation of sequence variants: a joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet Med 2015;17(5):405–24 doi 10.1038/gim.2015.30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Working Group R International rules for multiple primary cancers (ICD-0 third edition). Eur J Cancer Prev 2005;14(4):307–8 doi 10.1097/00008469-200508000-00002. [DOI] [PubMed] [Google Scholar]
- 27.Srinivasan P, Bandlamudi C, Jonsson P, Kemel Y, Chavan SS, Richards AL, et al. The context-specific role of germline pathogenicity in tumorigenesis. Nat Genet 2021;53(11):1577–85 doi 10.1038/s41588-021-00949-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jonsson P, Bandlamudi C, Cheng ML, Srinivasan P, Chavan SS, Friedman ND, et al. Tumour lineage shapes BRCA-mediated phenotypes. Nature 2019;571(7766):576–9 doi 10.1038/s41586-019-1382-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wang Q, Pierce-Hoffman E, Cummings BB, Karczewski KJ, Alföldi J, Francioli LC, Gauthier LD, Hill AJ, O’Donnell-Luria AH, Genome Aggregation Database (gnomAD) Production Team, Genome Aggregation Database (gnomAD) Consortium, & MacArthur, D. G.. Landscape of multi-nucleotide variants in 125,748 human exomes and 15,708 genomes. bioRxiv 2019:p. 573378. [Google Scholar]
- 30.Landrum MJ, Lee JM, Benson M, Brown G, Chao C, Chitipiralla S, et al. ClinVar: public archive of interpretations of clinically relevant variants. Nucleic Acids Res 2016;44(D1):D862–8 doi 10.1093/nar/gkv1222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zehir A, Benayed R, Shah RH, Syed A, Middha S, Kim HR, et al. Mutational landscape of metastatic cancer revealed from prospective clinical sequencing of 10,000 patients. Nat Med 2017;23(6):703–13 doi 10.1038/nm.4333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Shen R, Seshan VE. FACETS: allele-specific copy number and clonal heterogeneity analysis tool for high-throughput DNA sequencing. Nucleic Acids Res 2016;44(16):e131 doi 10.1093/nar/gkw520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Guo MH, Plummer L, Chan YM, Hirschhorn JN, Lippincott MF. Burden Testing of Rare Variants Identified through Exome Sequencing via Publicly Available Control Data. Am J Hum Genet 2018;103(4):522–34 doi 10.1016/j.ajhg.2018.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Pritchard CC, Mateo J, Walsh MF, De Sarkar N, Abida W, Beltran H, et al. Inherited DNA-Repair Gene Mutations in Men with Metastatic Prostate Cancer. N Engl J Med 2016;375(5):443–53 doi 10.1056/NEJMoa1603144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Carlo MI, Ravichandran V, Srinavasan P, Bandlamudi C, Kemel Y, Ceyhan-Birsoy O, et al. Cancer Susceptibility Mutations in Patients With Urothelial Malignancies. J Clin Oncol 2020;38(5):406–14 doi 10.1200/JCO.19.01395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Knudson AG Jr. Mutation and cancer: statistical study of retinoblastoma. Proc Natl Acad Sci U S A 1971;68(4):820–3 doi 10.1073/pnas.68.4.820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wang Q, Pierce-Hoffman E, Cummings BB, Alfoldi J, Francioli LC, Gauthier LD, et al. Landscape of multi-nucleotide variants in 125,748 human exomes and 15,708 genomes. Nat Commun 2020;11(1):2539 doi 10.1038/s41467-019-12438-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Testa JR, Cheung M, Pei J, Below JE, Tan Y, Sementino E, et al. Germline BAP1 mutations predispose to malignant mesothelioma. Nat Genet 2011;43(10):1022–5 doi 10.1038/ng.912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Girard E, Eon-Marchais S, Olaso R, Renault AL, Damiola F, Dondon MG, et al. Familial breast cancer and DNA repair genes: Insights into known and novel susceptibility genes from the GENESIS study, and implications for multigene panel testing. Int J Cancer 2019;144(8):1962–74 doi 10.1002/ijc.31921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Jara L, Dubois K, Gaete D, de Mayo T, Ratkevicius N, Bravo T, et al. Variants in DNA double-strand break repair genes and risk of familial breast cancer in a South American population. Breast Cancer Res Treat 2010;122(3):813–22 doi 10.1007/s10549-009-0709-2. [DOI] [PubMed] [Google Scholar]
- 41.Zhao F, Shang Y, Zeng C, Gao D, Li K. Association of single nucleotide polymorphisms of DNA repair genes in NER pathway and susceptibility to pancreatic cancer. Int J Clin Exp Pathol 2015;8(9):11579–86. [PMC free article] [PubMed] [Google Scholar]
- 42.Zhang XJ, Liu P, Zhu F. Polymorphisms of DNA repair-related genes with susceptibility and prognosis of prostate cancer. Genet Mol Res 2014;13(2):4419–24 doi 10.4238/2014.January.24.20. [DOI] [PubMed] [Google Scholar]
- 43.Tlemsani C NT, et al. Whole-exome sequencing reveals germline-mutated small cell lung cancer subtype with favorable response to DNA repair-targeted therapies. Science Translational Medicine 2021. doi 10.1126/scitranslmed.abc7488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Huang KL, Mashl RJ, Wu Y, Ritter DI, Wang J, Oh C, et al. Pathogenic Germline Variants in 10,389 Adult Cancers. Cell 2018;173(2):355–70 e14 doi 10.1016/j.cell.2018.03.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Offit K, Tkachuk KA, Stadler ZK, Walsh MF, Diaz-Zabala H, Levin JD, et al. Cascading After Peridiagnostic Cancer Genetic Testing: An Alternative to Population-Based Screening. J Clin Oncol 2020;38(13):1398–408 doi 10.1200/JCO.19.02010. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Raw genomic data for this study were generated at Molecular Diagnostics Service in the Department of Pathology, the Integrated Genomics Operation Core and Marie-Josée and Henry R. Kravis Center for Molecular Oncology at Memorial Sloan Kettering Cancer Center (ClinicalTrials.gov Identifier: NCT01775072). Derived summary data supporting the findings of this study are available from the corresponding author upon request.