

Abstract
Androgen deprivation therapy (ADT) is the standard of care treatment for advance stage prostate cancer. Treatment with ADT develops resistance in multiple ways leading to the development of castration-resistant prostate cancer (CRPC). Present research establishes that prostate cancer stem-like cells (CSCs) play a central role in the development of treatment resistance followed by disease progression. Prostate CSCs are capable of self-renewal, differentiation, and regenerating tumor heterogeneity. The stemness properties in prostate CSCs arise due to various factors such as androgen receptor mutation and variants, epigenetic and genetic modifications leading to alteration in the tumor microenvironment, changes in ATP-binding cassette (ABC) transporters, and adaptations in molecular signaling pathways. ADT reprograms prostate tumor cellular machinery leading to the expression of various stem cell markers such as Aldehyde Dehydrogenase 1 Family Member A1 (ALDH1A1), Prominin 1 (PROM1/CD133), Indian blood group (CD44), SRY-Box Transcription Factor 2 (Sox2), POU Class 5 Homeobox 1(POU5F1/Oct4), Nanog and ABC transporters. These markers indicate enhanced self-renewal and stemness stimulating CRPC evolution, metastatic colonization, and resistance to antiandrogens. In this review, we discuss the role of ADT in prostate CSCs differentiation and acquisition of CRPC, their isolation, identification and characterization, as well as the factors and pathways contributing to CSCs expansion and therapeutic opportunities.
Keywords: Prostate cancer, second-generation antiandrogens, cancer stem cells, castration resistance prostate cancer, androgen deprivation therapy
INTRODUCTION
Prostate cancer is the most commonly diagnosed cancer in the United States, and approximately 50% of the men diagnosed with advance stage prostate cancer undergo androgen deprivation therapy (ADT)[1]. US Food and Drug Administration approved ADT to treat metastatic prostate cancer as a neo-adjuvant in post-radiation therapy[2]. ADT has also been accepted as the first-line treatment for prostate tumors that have extended to the lymph nodes, and biochemical recurrence followed by prostate-specific antigen (PSA) resurgence, asymptomatic metastatic and locally advanced disease[3]. Since androgen receptor (AR) is essential for the function, survival, and differentiation of prostatic tissue, ADT reduces androgens necessary to block cancer progression[4]. Accumulating data suggests that androgens are important players in the human body in maintaining physiological functions[5-6]. Androgen receptor holds a key function in prostatic epithelial cells’ growth and proliferation in response to testosterone[7]. ADT comprises the use of the first-generation antiandrogens such as bicalutamide, nilutamide or flutamide that solely targets AR translocation to the nucleus and prevent downstream signaling. The second-generation antiandrogens viz. enzalutamide, apalutamide and darolutamide, as well as inhibitors of androgen biosynthesis such as abiraterone acetate, improve upon this mechanism[8]. Changes in the function of AR signaling result in tumor suppression to tumor promotion, where the disease eventually progresses to the emergence of castration-resistant prostate cancer (CRPC)[9]. Accumulated evidence suggests that ADT has a significant role in the management of metastatic prostate cancer; it reduces complications and enhances overall survival[2,10-11]. In a study based on Cochrane meta-analysis, the neo-adjuvant ADT with radical prostatectomy significantly improves adverse histopathologic parameters such as surgical margin or pathologic tumor stage[12]. Studies reported that prostate cancer patients undergoing radiotherapy together with ADT treatment further increases the probability of disease-free survival[12-13]. This combinatorial treatment is well accepted and highlighted in the guidelines of the American Urological Association and European Association of Urology[14-15]. However, numerous studies indicate that ADT is associated with a multitude of side effects that can impact the quality of life[16-18]. These include fatigue, loss of libido, arterial stiffness, erectile dysfunction, hot flushes, new-onset diabetes mellitus, altered body composition, osteoporosis and induced skeletal complications, and cognitive decline[16-18]. Some recent findings also demonstrated that ADT treatment might increase cardiovascular-mediated morbidity and mortality[19-20].
Prostate cancer resistance during ADT treatment is reported in in vitro models of recurrence and CRPC patients[21]. CRPC development is linked with genes associated with AR signaling, both at transcription and translation levels[21]. A study including multiple isogenic tumor xenograft models demonstrated increased AR expression in recurrent tumor samples compared to paired androgen-sensitive samples[22]. AR stabilization alters the rate of post-translational modifications and interaction with heat shock proteins which ultimately modulates normal cellular physiology[23]. Studies also reported that stabilization of AR is positively associated with ADT resistance, which may be linked with CRPC[4,24]. Phosphorylation at particular sites and enrichment of growth factors reactivate AR, which further increases prostate cancer proliferation under low androgen levels[25]. Apart from AR stabilization and phosphorylation, the mutation in AR gene is also associated with resistance development[26]. A point mutation was reported in the lymph node of a patient with metastatic prostate cancer and causes amino acid substitution at position 878, threonine to alanine[27]. This amino acid substitution results in response to non-androgen hormones that enhance resistance to ADT treatment. Some other reported mutations are W742C, H875Y/T, F877L and L702H in response to resistance development against enzalutamide, nilutamide, flutamide and bicalutamide[28-29]. Other factors such as different AR splice variant expression, aberrant glucocorticoids and glucocorticoid receptor expression, impairment of DNA repair pathway, miRNAs, cellular metabolism, and alterations in enzymatic and signaling pathways tightly support resistance development in response to ADT[30-31].
Emerging research implicates that cancer stem cells (CSCs) are key to the development of therapeutic resistance, and studies have also established a link to ADT relapse tumors[32]. Subsequent work has shown that ADT increases the enrichment of CSCs populations which are inherently treatment-resistant having the ability to promote CRPC[33]. The CSC hypothesis is an emerging model that describes several molecular characteristics of cancer. CSCs facilitate the development of a cellular hierarchy, maintain a CSC rich-pool, differentiate into a proliferating progeny, and assist the formation of a heterogeneous tumor[34]. As a result, there is a strong consensus that CSCs are the cells of origin in cancer and have the propensity of cancers to relapse, metastasize, and develop resistance to conventional therapies[34]. In this review, we describe the role of ADT in CSC differentiation during CRPC acquisition, their isolation, identification and characterization. Outlining the underlying mechanisms triggered by ADT and highlighting potential CSCs targets could aid in the development of future therapeutic strategies in prostate cancer patients leading to improved outcomes.
PROSTATE CANCER STEM-LIKE CELLS
Stem cell markers in prostate cancer
Normal stem cells possess the intrinsic capacity for unlimited replicative potential as well as differentiation into all lineages of mature cells required for tissue and/or organ maintenance[35]. In general, cancer cells possess cellular heterogeneity and inherent genetic instability, which makes them immortal in characteristics. CSCs are commonly defined as cells that demonstrate characteristics similar to a normal stem cell, including lack of senescence, self-renewal capacity and pluripotency[36]. This type of cancer cell possesses the ability to develop a cellular hierarchy, facilitating the maintenance of a CSC pool while differentiating into a proliferating progeny, enabling the formation and growth of a heterogeneous tumor.
The prostate gland consists of basal (undifferentiated), differentiated and neuroendocrine cells[37]. The basal cells are androgen-independent and express cytokeratin 5, cytokeratin 14, Tumor Protein P63, and Cell Surface Glycoprotein (CD44) markers[38-40]. They also express much less AR, PSA and prostatic acid phosphatase (PAP)[38-40]. Differentiated cells include glandular epithelial and secretory luminal cells, which express AR, PSA, PAP, cytokeratin 8 and cytokeratin 18[41]. The neuroendocrine cells are androgen-independent and do not secrete PSA[42]. Prostate cancer stem cells are androgen-independent cells that give rise to androgen-sensitive progenitor cells[38]. These progenitor cells in the presence of androgens differentiate into androgen-dependent cells. A number of research studies indicate that CSCs are considered as the cells of origin in cancer and have been linked to tumorigenesis, treatment resistance and cancer relapse[41].
Several research studies identified prostate CSCs genes that are important for self-renewal, pluripotency, resistance and serve as markers for identification[43]. Stem cell antigen-1, aldehyde dehydrogenases (ALDH), CD133 (PROM1), trophoblast cell surface antigen 2 and CD44 are markers to identify prostate CSCs in the basal compartment[44]. The other common markers include CD44, CD24, and CD49 Antigen-Like Family Member D (also known as Integrin Subunit Alpha 4), which have been tightly associated with prostate CSCs[45]. Another study identified a rare prostate cancer stem cell maker, KIT Proto-Oncogene, Receptor Tyrosine Kinase (KIT) in adult mouse prostatic stem cell population, which possess cancer stem cell-like features such as differentiation and self-renewal[46]. The prostate CSCs enriched in CD133+ cells isolated from established primary human prostate cancer cell lines and in alpha(2)beta(1)-integrin subunit phenotype identified as genetically unstable with clonogenic formation and invasive potential[47]. Yu et al.[48] have identified high ALDH1 activity in LNCaP and PC-3 prostate cancer cells associated with CSC-like properties; in particular, ALDHhi/CD44+ cells possess a high clonogenic function and tumorigenic potential. A study conducted on prostate cancer tissue specimens indicated Oct4, Sox2 and Nanog genes as prostate CSC markers[49]. Collins et al.[50] further demonstrate in the mouse xenograft model that elevated levels of ALDH support stemness in cells; indeed, ALDHhi/CD44+/α2β1+ cells are enhanced during castration and were critical in the development of antiandrogen resistance. Further, cells possessing similar phenotypes were isolated from clinical specimens and analyzed for self-renewal and spheroid formation. The outcome showed that ALDHhi/CD44+/α2β1+ cells significantly support cell proliferation and colony formation[51]. A study performed on patient biopsies samples (Gleason score range 5-6) suggested that CD133+/CD44+/ATP-binding cassette sub-family G member 2 (ABCG2)+/CD24− cells actively participate in spheroid formation[52]. In addition, primary tumor cells containing Enhancer of Zeste 2 Polycomb Repressive Complex 2 Subunit (EZH2+)/E-cadherin- markers are highly associated with tumor recurrence [Table 1][53].
Table 1.
Prostate cancer stem cells markers
| Pathways | Markers names | Gene symbol | Ref. |
| Tumor progression | KIT proto-oncogene, receptor tyrosine kinase | CD117/c-kit | [43,54-56] |
| Prominin 1 | CD133 | ||
| Indian blood group | CD44 | ||
| α2β1 integrin | ITGB1 | ||
| Integrin Subunit Alpha 6 | α6 integrin | ||
| C-X-C motif chemokine receptor 4 | CXCR4 | ||
| Epithelial cell adhesion molecule | EPCAM | ||
| Cytokeratin 5 | KRT5 | ||
| Kallikrein related peptidase 3 | KLK3/PSA | ||
| Tumor-associated calcium signal transducer 2 | Trop2 | ||
| Activated leukocyte cell adhesion molecule | ALCAM | ||
| Aldehyde dehydrogenase 1 family member A1 | ALDH1 | ||
| Transglutaminase 2 | TG2 | ||
| Enhancer of zeste 2 polycomb repressive complex 2 subunit | EZH2 | ||
| Metastatic colonization and growth |
KIT proto-oncogene, receptor tyrosine kinase | CD117/c | [43,56] |
| C-X-C motif chemokine receptor | CXCR4 | ||
| Epithelial cell adhesion molecule | EPCAM | ||
| E-cadherin/ cadherin 1 | CDH1 | ||
| Indian blood group | CD44 | ||
| Enhancer of zeste 2 polycomb repressive complex 2 subunit | EZH2 | ||
| Recurrence and therapeutic resistance |
KIT proto-oncogene, receptor tyrosine kinase | CD117/c-kit | [43,54-56] |
| α2β1 integrin | ITGB1 | ||
| Integrin subunit alpha 6 | ITGA6 | ||
| E-cadherin/ cadherin 1 | CDH1 | ||
| Epithelial cell adhesion molecule | EPCAM | ||
| C-X-C motif chemokine receptor 4 | CXCR4 | ||
| Enhancer of zeste 2 polycomb repressive complex 2 subunit | EZH2 | ||
| Aldehyde dehydrogenase 1 family member A1 | ALDH1 | ||
| Transglutaminase 2 | TG2 | ||
| Activated leukocyte cell adhesion molecule | CD166/ALCAM | ||
| Kallikrein related peptidase 3 | KLK3/PSA | ||
| Androgen receptor splice variant 7 | AR-V7 | ||
| ATP binding cassette subfamily G member 2 (junior blood group) | ABCG2 | ||
| Self-renewal capacity | Prominin 1 | CD133 | [43,55] |
| Cytokeratin 5 | KRT5 | ||
| Kallikrein related peptidase 3 | KLK3/PSA | ||
| Aldehyde dehydrogenase 1 family member A1 | ALDH1 | ||
| Activated leukocyte cell adhesion molecule | CD166/ALCAM | ||
| C-X-C motif chemokine receptor 4 | CXCR4 | ||
| Tumor-associated calcium signal transducer 2 | Trop2 | ||
| Integrin subunit alpha 6 | α6 integrin | ||
| α2β1 integrin | ITGB1 | ||
| Indian blood group | CD44 | ||
| Stemness gene expression | Prominin 1 | CD133 | [43,54] |
| Indian blood group | CD44 | ||
| E-cadherin/ cadherin 1 | CDH1 | ||
| Kallikrein related peptidase 3 | KLK3/PSA | ||
| Aldehyde dehydrogenase 1 family member A1 | ALDH1 | ||
| Enhancer of zeste 2 polycomb repressive complex 2 subunit | EZH2 |
Molecular pathways in the generation of prostate cancer stem-like cells post ADT
AR is a key transcription factor involved in androgen-dependent prostate cancer growth. Targeting AR with the first-generation antiandrogens does not inhibit inter or intramolecular N-C interactions required for the nuclear localization[57]. At the diagnosis of metastatic CRPC, the common genomic alteration event found in AR is amplification and AR mutation[58]. These AR genomic alterations dysregulate the signaling pathway in patients and demonstrate a compensatory resistance mechanism via increasing AR expression in response to the potent AR inhibition by enzalutamide, which results in diminished efficacy of treatment overtime[59]. In a systemic study, exome sequencing of 150 metastatic CRPC biopsy specimens demonstrated 63% of AR mutation and amplification in comparison to 440 primary prostate cancer tissues[60]. Apart from AR mutation, AR variants such as ARV7 were also reported for resistance development and support androgen-independent growth of prostate cancer cells[61]. Prostate cancer patients who underwent ADT showed hematopoiesis from pluripotent stem cells, PI3K/AKT signaling, ERK/MAPK signaling, and Wnt/β-catenin signaling, and the role of Nanog in mammalian embryonic stem cell pluripotency signaling pathways were overrepresented [Figure 1]. This information revealed that the genomic alteration in AR either by amplification or mutation tends to increase the expression of associated stem cell markers.
Figure 1.

Canonical signaling pathway overrepresented in prostate cancer patients under ADT. These pathways play a critical role in driving cancer stem-like cell phenotype. The red bar represents the genes upregulated and green is downregulated and overlaid with IPA database. The X-axis in the graph represents the signaling pathways while the Y-axis showed -log (P-value) and percentage (%). ADT: Androgen deprivation therapy
With reference to hematopoiesis, pluripotent stem cells, the expression of transmembrane receptors genes, which includes CD4, CD247, CD3E, CD8A, CSF3, CXCL8, and family members of immunoglobulin heavy constant gamma proteins (IGHA1, IGHD, IGHG1, IGHG2, IGHG3, IGHM) and the expression of cytokines such as IL6 and IL10 were increased in patients undergoing ADT [Table 2].
Table 2.
List of genes, location, types, along with fold changes value associated with hematopoiesis pluripotent stem cells
| Gene symbol | Gene name | Fold change in ADT | Location | Type(s) |
| CD4 | CD4 molecule | 1.829 | Plasma membrane | transmembrane receptor |
| CD247 | CD247 molecule | 1.437 | Plasma membrane | transmembrane receptor |
| CD3E | CD3e molecule | 1.597 | Plasma membrane | transmembrane receptor |
| CD8A | CD8a molecule | 1.469 | Plasma membrane | other |
| CSF3 | colony stimulating factor 3 | 5.074 | Extracellular space | cytokine |
| CXCL8 | C-X-C motif chemokine ligand 8 | 3.395 | Extracellular space | cytokine |
| FCER1G | Fc fragment of IgE receptor Ig | 1.815 | Plasma membrane | transmembrane receptor |
| IGHA1 | immunoglobulin heavy constant alpha 1 | 3.2 | Extracellular space | other |
| IGHD | immunoglobulin heavy constant delta | 2.58 | Extracellular space | other |
| IGHG1 | immunoglobulin heavy constant gamma 1 (G1m marker) | 2.139 | Extracellular space | other |
| IGHG2 | immunoglobulin heavy constant gamma 2 (G2m marker) | 3.362 | Plasma membrane | other |
| IGHG3 | immunoglobulin heavy constant gamma 3 (G3m marker) | 2.922 | Extracellular space | other |
| IGHM | immunoglobulin heavy constant mu | 2.946 | Plasma membrane | transmembrane receptor |
| IL6 | interleukin 6 | 6.333 | Extracellular space | cytokine |
| IL10 | interleukin 10 | 1.764 | Extracellular space | cytokine |
| LIF | LIF interleukin 6 family cytokine | 2.636 | Extracellular space | cytokine |
Isolation, identification and characterization of prostate cancer stem cells
Extensive work has shown that CSCs from primary prostate tumors or established cancer cell lines can be isolated from a heterogeneous population using cell surface markers, such as CD44 and CD133, aldehyde dehydrogenase (ALDH) activity using the ALDEFLUOR assay, and Hoechst dye to identify the side population[62-63]. Isolation of prostate CSCs has been performed by several groups. The first CSC isolation was performed from patients undergoing radical prostatectomy[64]. These CSCs exhibited a significant capacity for self-renewal and the ability to regenerate the phenotypically mixed populations of non-clonogenic cells. The prostate CSCs were isolated using the CD44/α2β1high/CD133+ phenotype and demonstrated high clonogenic and invasive capacity of basal cell origin with high levels of genetic instability[64-65]. A study by Rajasekhar et al.[66] observed that CSCs expressing human pluripotent stem cell marker TRA-60-1+ /CD151+ /CD166+ possess high self-renewal and differentiation ability, and were able to reiterate tumor heterogeneity in serial in vivo transplantations. More recent studies have identified that a majority of prostate tumors harbor prostate-specific TMPRSS2 gene and the ERG oncogene (TMPRSS2: ERG) gene fusion which could be used as a stem cell marker with high specificity providing ERG-driven survival advantages[67]. Moreover, ALDH1high prostate cancer cells have been shown to exhibit several CSC characteristics such as clonogenicity, migration, tumorigenicity, and propensity to form metastases in vivo[68]. Another method to enrich the CSC population is developing tumorspheres in cell culture with a higher in vivo tumor incidence rate[69].
Our group and others have demonstrated fluorescence-activated cell sorting and magnetic-activated cell sorting utilizing various human prostate cancer cell lines[70-71]. Other studies showed that CD44 and CD133 were associated with high Nanog expression in prostate carcinoma cell lines[71]. Nanog has shown to be predominantly expressed from the NanogP8 pseudogene in a panel of prostate carcinoma cells including DU145, LNCaP, and PC-3 and primary prostate carcinoma cells. NanogP8 expression was enriched several folds in CD133+ and CD133+/CD44+ CSCs compared to non-CSCs[71]. Human prostate cancer PC-3 cells displayed high CD44+/CD133+ CSC-like features including enhanced tumor sphere formation and elevated Nanog levels. Similarly, CD117+/ABCG2+ cells isolated from 22Rv1 prostate cancer cells overexpress the core stem cell transcription factors, Nanog, Oct3/4, and Sox2, and the CSC marker CD133[72]. A recent study from our group has demonstrated high expression of ALDH1high, Oct4 and Sox2 in clinical prostate cancer specimens undergoing ADT, compared to grade-matched controls[73].
Most standard therapies for prostate cancer primarily affect cancer cells, but CSCs undergo G0/G1 phase cell cycle arrest and remain static, thus evading cell death from chemotherapeutic drugs[74]. Experimental data also suggest that CSCs are resistant to conventional chemotherapy and radiation and may be the cells responsible for disease recurrence and/or progression[75]. A study showed that CD133+ had a high capacity to proliferate in vitro and have AR+ phenotype[76]. These CD133+ cells form branched spheroids structure in a 3D culture system and generate prostatic-like acini in vivo[76]. Hence, the drug-resistant characteristic of CSCs is useful to isolate and identify CSCs. Previous studies have shown that radiotherapy combined with hypoxic culture can also be used to enrich CSCs population[77].
Therapeutic opportunities for prostate cancer stem-like cells
Prostate cancer patients undergo treatment therapy such as radiotherapy or chemotherapy, resulting in shrinkage of tumors[78]. However, after therapy, some cells accumulate genetic/epigenetic changes that result in loss of control on self-renewal potential. These cells, referred to as prostate CSCs, reprogram the tumor environment to their benefit, supporting increased survival, self-renewal, and tumor recurrence[64]. Research showed that cellular immunotherapy has some beneficial role in the treatment of prostate cancer[79]. The T cell-based immunotherapy showed a positive response to prostate cancer patients with metastatic CRPC and increased the overall survival[79]. A research group prepared an immunogenic peptide derived from dendritic cells sensitized to CD44 and EpCAM followed by co-culture with the expanded peripheral blood lymphocyte (PBL)-derived cytokine-induced killer cell[80]. This study showed that dendritic cells- cytokine-induced killer cells exhibit remarkable cytotoxicity against prostate cancer stem-like cells-enriched prostate spheroids both in vitro and in vivo[80]. In addition to these findings, several other cellular events impart growth advantages to CSCs. In this context, various signaling pathways such as Wnt/β-catenin, hedgehog, NF-κB and Notch; ABC transporters and tumor microenvironment could be the putative target(s) for prostate CSCs[81] [Figure 2].
Figure 2.
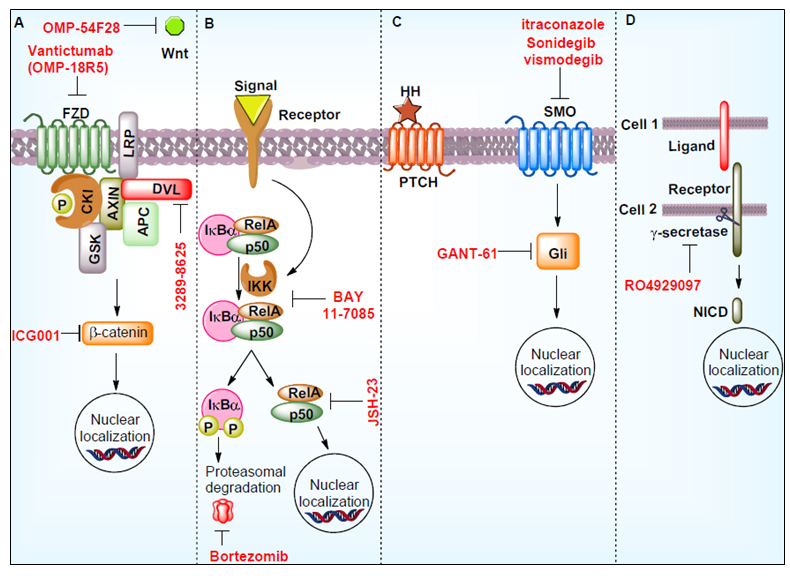
Signaling pathways regulating CSCs and the use of inhibitors in suppressing these pathways. These molecules thereof could be developed as potential therapeutics. The denotes in the figure are: Hedgehog signaling pathway (HH), notch intracellular domain (NICD), phosphorylation (P), smoothened (SMO), Wnt signaling pathways (Wnt), nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha (IkBα), RELA Proto-Oncogene (RelA), Frizzled (FZD), casein kinase I (CKI), Axin (AXIN), APC Regulator of WNT Signaling Pathway (APC), Dishevelled (Dvl), Glycogen Synthase Kinase (GSK), IkappaB Kinase (IKK).
Wnt signaling pathway is involved in various cellular processes and is crucial for cell fate determination, cell polarity, cell migration, neural patterning and organogenesis during embryonic development. Wnt pathway is also associated with the maintenance of stem cells in a self-renewing state[82]. A study demonstrated that Wnt signaling activation is oncogenic in the prostate and supports CRPC growth in vivo[83]. This study also showed that increased Wnt signaling induces neuroendocrine differentiation, epithelial-mesenchymal transition and drives stem cell-like features to prostate cancer cells. A number of small molecule inhibitors and monoclonal antibodies have been tested to inhibit the Wnt signaling pathway. Wnt signaling inhibitors such as 3289-8625, Foxy-5, and OMP-54F28 have been reported to inhibit prostate cancer cell growth[84-86]. A porcupine (palmitoylation of Wnt ligands) inhibitor LGK974 combined with docetaxel and paclitaxel also showed remarkable effectiveness on solid tumors[64]. A monoclonal antibody vantictumab (OMP-18R5) blocks canonical Wnt signaling pathways and inhibits prostate cancer progression[87].
Hedgehog signaling pathways play an important role in the development of prostate cancer. Hedgehog signaling targets genes involved in prostate CSCs survival, proliferation, and metastasis[88]. This signaling also enhanced the overexpression of ABC transporters in prostate cancer cells[88]. Hedgehog signaling inhibitor sonidegib (LDE-225) suppresses the key genes including Oct4, Nanog, c-Myc, and Sox2 involved in self-renewal and stemness potential[89]. Gli transcription factor inhibitor GANT-61 inhibits PTCH1 expression and tumor growth in vivo[90]. Other Hedgehog signaling inhibitors such as vismodegib, itraconazole and orteronel either alone or in combination and/or surgery inhibit prostate cancer growth[64].
Upregulation of the NF-κB pathway has been observed in cancer stem cells[91]. Various studies also demonstrated that NF-κB signaling was upregulated in prostate cancer cells and associated with increased progression, chemotherapy resistance, metastasis and recurrence[92]. Several clinical trials have been performed by targeting the NF-κB signaling pathway. Clinical trial NCT01695473 used PI3K inhibitor BKM120, which acts downstream of NF-κB in high-risk localized prostate cancer patients[93]. NCT00118092 clinical trial was performed using heat shock protein 90 inhibitor 17-(allylamino)-17-demethoxygeldanamycin to treat metastatic prostate cancer patients[94]. Aspirin, a reported drug for inflammation-regulated cancer, was utilized in clinical trial NCT02757365[95]. This trial demonstrated that aspirin suppresses CRPC progression.
Notch signaling pathways are well known for their contribution to self-renewal, differentiation, resistance and stemness development[96]. Interaction of Notch receptor and ligand facilitates NICD production through gamma-secretase, which translocates to the nucleus and initiates transcription of self-renewal, stemness, and other CSC development associated genes[97]. In this context, a number of gamma-secretase inhibitory agents were identified. A clinical trial (NCT01200810) was performed in prostate cancer patients using gamma-secretase inhibitor RO4929097 with bicalutamide[98]. This trial compares PSA expression with time after surgery/radiation and combined treatment[98].
Alteration in ABC transporter genes and tumor microenvironment tightly regulates cellular physiology and transcriptomic machinery in prostate CSCs[99]. The tumor microenvironment plays a decisive role in regulating CSCs progression[100]. It also facilitates abnormal cancer signaling pathways, epithelial-mesenchymal transition, invasion, etc. Overexpression of ABC transporters exports therapeutic drugs outside the cells, which makes them resistant to the drug[101]. Several lines of research have been performed to target the ABC transporters. An ABC transporter efflux inhibitor verapamil inhibits prostate cancer proliferation by inhibiting the potassium ion channel[102]. Cyclosporin A, another ABC transporter inhibitor, inactivates NFATc1 (nuclear factor of activated T-cells) in biochemical recurrence and CRPC[103].
CONCLUSION
Despite thorough research for mechanisms leading to CRPC as a result of resistance to antiandrogens in the past few decades, our understanding remains limited. ADT causes complex alterations within tumors in terms of factors and pathways as well as its epigenetics and genetics affecting the tumor microenvironment. Several research studies showed that aberrant cellular signaling, generation of AR variants, and AR mutation support the development of drug resistance. The accumulative effects of these factors also contribute to the generation of prostate cancer stem-like cells. CSCs are the reservoir of cancer cells that exhibit surface markers such as ALDH, CD133 and CD44, possessing properties of self-renewal and the ability to reestablish the heterogeneous tumor cell population promoting metastatic colonization, self-renewal, and recurrence. Research showed that targeting prostate CSCs could be a better strategy for the treatment of CRPC. In this direction, various small molecule inhibitors, antibodies and other combinatorial treatments have been evaluated in various clinical trials. The outcome demonstrated that these therapies increased overall survival in prostate cancer patients. However, the lifespan increment of prostate cancer patients is still challenging for clinicians as all these drug therapies become resistant after a certain time of treatment.
DECLARATIONS
Authors’ contributions
Made substantial contributions to conception and writing of the manuscript: Kushwaha PP, Verma S, Kumar S, Gupta S
Developed the figures and tables: Kushwaha PP, Verma S
Made substantial contributions to conception and design of the manuscript and performed data analysis and interpretation: Kushwaha PP, Verma S
Provided administrative, technical, and material support: Gupta S
Availability of data and materials
Not applicable.
Financial support and sponsorship
This work was supported by the Department of Defense Grants W81XWH-18-1-0618, W81XWH-19-1-0720 to S.G. Acknowledgement is due to Indian Council of Medical Research, India and Department of Science and Technology, India for providing financial support and DST-India for providing Departmental grant to the Department of Biochemistry, Central University of Punjab, Bathinda, India in the form of DST-FIST grant to SK.
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2022.
References
- 1.Gunner C, Gulamhusein A, Rosario DJ. The modern role of androgen deprivation therapy in the management of localised and locally advanced prostate cancer. J Clin Urol. 2016;9:24–9. doi: 10.1177/2051415816654048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Crawford ED, Heidenreich A, Lawrentschuk N, et al. Androgen-targeted therapy in men with prostate cancer: evolving practice and future considerations. Prostate Cancer Prostatic Dis. 2019;22:24–38. doi: 10.1038/s41391-018-0079-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Perera M, Roberts MJ, Klotz L, et al. Intermittent versus continuous androgen deprivation therapy for advanced prostate cancer. Nat Rev Urol. 2020;17:469–81. doi: 10.1038/s41585-020-0335-7. [DOI] [PubMed] [Google Scholar]
- 4.Karantanos T, Corn PG, Thompson TC. Prostate cancer progression after androgen deprivation therapy: mechanisms of castrate resistance and novel therapeutic approaches. Oncogene. 2013;32:5501–11. doi: 10.1038/onc.2013.206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Saad F, Bögemann M, Suzuki K, Shore N. Treatment of nonmetastatic castration-resistant prostate cancer: focus on second-generation androgen receptor inhibitors. Prostate Cancer Prostatic Dis. 2021;24:323–34. doi: 10.1038/s41391-020-00310-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Formaggio N, Rubin MA, Theurillat JP. Loss and revival of androgen receptor signaling in advanced prostate cancer. Oncogene. 2021;40:1205–16. doi: 10.1038/s41388-020-01598-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Perlmutter MA, Lepor H. Androgen deprivation therapy in the treatment of advanced prostate cancer. Rev Urol. 2007;9:S3–8. [PMC free article] [PubMed] [Google Scholar]
- 8.Vaishampayan UN, Heilbrun LK, Monk P 3rd, et al. Clinical efficacy of enzalutamide vs bicalutamide combined with androgen deprivation therapy in men with metastatic hormone-sensitive prostate cancer: a randomized clinical trial. JAMA Netw Open. 2021;4:e2034633. doi: 10.1001/jamanetworkopen.2020.34633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Spratt DE, Malone S, Roy S, et al. Prostate radiotherapy with adjuvant androgen deprivation therapy (ADT) improves metastasis-free survival compared to neoadjuvant ADT: an individual patient meta-analysis. J Clin Oncol. 2021;39:136–44. doi: 10.1200/JCO.20.02438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Thoma C. Prostate cancer: 2-year ADT improves survival in patients receiving high-dose radiotherapy. Nat Rev Urol. 2015;12:181. doi: 10.1038/nrurol.2015.43. [DOI] [PubMed] [Google Scholar]
- 11.Kim M, Song C, Jeong IG, et al. Androgen deprivation therapy during and after post-prostatectomy radiotherapy in patients with prostate cancer: a case control study. BMC Cancer. 2018;18:271. doi: 10.1186/s12885-018-4189-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Shore ND. Current and future management of locally advanced and metastatic prostate cancer. Rev Urol. 2020;22:110–23. [PMC free article] [PubMed] [Google Scholar]
- 13.Singh AB. Physiology and pathophysiology of male sex hormones. Trends in Endocrinology & Metabolism. 2004;15:408–9. doi: 10.1016/j.tem.2004.09.001. [DOI] [Google Scholar]
- 14.Merlet J, Racine C, Moreau E, Moreno SG, Habert R. Male fetal germ cells are targets for androgens that physiologically inhibit their proliferation. Proc Natl Acad Sci U S A. 2007;104:3615–20. doi: 10.1073/pnas.0611421104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mizokami A, Koh E, Izumi K, et al. Prostate cancer stromal cells and LNCaP cells coordinately activate the androgen receptor through synthesis of testosterone and dihydrotestosterone from dehydroepiandrosterone. Endocr Relat Cancer. 2009;16:1139–55. doi: 10.1677/ERC-09-0070. [DOI] [PubMed] [Google Scholar]
- 16.Barnes KA, Ball LE, Galvão DA, Newton RU, Chambers SK. Nutrition care guidelines for men with prostate cancer undergoing androgen deprivation therapy: do we have enough evidence? Prostate Cancer Prostatic Dis. 2019;22:221–34. doi: 10.1038/s41391-018-0099-9. [DOI] [PubMed] [Google Scholar]
- 17.Higano CS. Side effects of androgen deprivation therapy: monitoring and minimizing toxicity. Urology. 2003;61:32–8. doi: 10.1016/s0090-4295(02)02397-x. [DOI] [PubMed] [Google Scholar]
- 18.Nguyen PL, Alibhai SM, Basaria S, et al. Adverse effects of androgen deprivation therapy and strategies to mitigate them. Eur Urol. 2015;67:825–36. doi: 10.1016/j.eururo.2014.07.010. [DOI] [PubMed] [Google Scholar]
- 19.Álvarez-Maestro M, Eguibar A, Chanca P, et al. Androgen deprivation therapy in patients with prostate cancer increases serum levels of thromboxane A2: cardiovascular Implications. Front Cardiovasc Med. 2021;8:653126. doi: 10.3389/fcvm.2021.653126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lee HY, Chen HL, Teoh JY, et al. Abiraterone and enzalutamide had different adverse effects on the cardiovascular system: a systematic review with pairwise and network meta-analyses. Prostate Cancer Prostatic Dis. 2021;24:244–52. doi: 10.1038/s41391-020-00275-3. [DOI] [PubMed] [Google Scholar]
- 21.Alumkal JJ, Sun D, Lu E, et al. Transcriptional profiling identifies an androgen receptor activity-low, stemness program associated with enzalutamide resistance. Proc Natl Acad Sci U S A. 2020;117:12315–23. doi: 10.1073/pnas.1922207117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Montgomery RB, Mostaghel EA, Vessella R, et al. Maintenance of intratumoral androgens in metastatic prostate cancer: a mechanism for castration-resistant tumor growth. Cancer Res. 2008;68:4447–54. doi: 10.1158/0008-5472.CAN-08-0249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Li J, Fu X, Cao S, et al. Membrane-associated androgen receptor (AR) potentiates its transcriptional activities by activating heat shock protein 27 (HSP27) J Biol Chem. 2018;293:12719–29. doi: 10.1074/jbc.RA118.003075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.He Y, Wei T, Ye Z, et al. A noncanonical AR addiction drives enzalutamide resistance in prostate cancer. Nat Commun. 2021;12:1521. doi: 10.1038/s41467-021-21860-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Shu SK, Liu Q, Coppola D, Cheng JQ. Phosphorylation and activation of androgen receptor by Aurora-A. J Biol Chem. 2016;291:22854. doi: 10.1074/jbc.A110.121129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sumiyoshi T, Mizuno K, Yamasaki T, et al. Clinical utility of androgen receptor gene aberrations in circulating cell-free DNA as a biomarker for treatment of castration-resistant prostate cancer. Sci Rep. 2019;9:4030. doi: 10.1038/s41598-019-40719-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wadosky KM, Koochekpour S. Molecular mechanisms underlying resistance to androgen deprivation therapy in prostate cancer. Oncotarget. 2016;7:64447–70. doi: 10.18632/oncotarget.10901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jernberg E, Bergh A, Wikström P. Clinical relevance of androgen receptor alterations in prostate cancer. Endocr Connect. 2017;6:R146–61. doi: 10.1530/EC-17-0118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Watson PA, Arora VK, Sawyers CL. Emerging mechanisms of resistance to androgen receptor inhibitors in prostate cancer. Nat Rev Cancer. 2015;15:701–11. doi: 10.1038/nrc4016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Huang Y, Jiang X, Liang X, Jiang G. Molecular and cellular mechanisms of castration resistant prostate cancer. Oncol Lett. 2018;15:6063–76. doi: 10.3892/ol.2018.8123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Culig Z. Molecular mechanisms of enzalutamide resistance in prostate cancer. Curr Mol Biol Rep. 2017;3:230–5. doi: 10.1007/s40610-017-0079-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Huang H, Wang C, Liu F, et al. Reciprocal network between cancer stem-like cells and macrophages facilitates the progression and androgen deprivation therapy resistance of prostate cancer. Clin Cancer Res. 2018;24:4612–26. doi: 10.1158/1078-0432.CCR-18-0461. [DOI] [PubMed] [Google Scholar]
- 33.Moltzahn F, Thalmann GN. Cancer stem cells in prostate cancer. Transl Androl Urol. 2013;2:242–53. doi: 10.3978/j.issn.2223-4683.2013.09.06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Plaks V, Kong N, Werb Z. The cancer stem cell niche: how essential is the niche in regulating stemness of tumor cells? Cell Stem Cell. 2015;16:225–38. doi: 10.1016/j.stem.2015.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kim J, Koo BK, Knoblich JA. Human organoids: model systems for human biology and medicine. Nat Rev Mol Cell Biol. 2020;21:571–84. doi: 10.1038/s41580-020-0259-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Batlle E, Clevers H. Cancer stem cells revisited. Nat Med. 2017;23:1124–34. doi: 10.1038/nm.4409. [DOI] [PubMed] [Google Scholar]
- 37.Prajapati A, Gupta S, Mistry B, Gupta S. Prostate stem cells in the development of benign prostate hyperplasia and prostate cancer: emerging role and concepts. Biomed Res Int. 2013;2013:107954. doi: 10.1155/2013/107954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Collins AT, Maitland NJ. Prostate cancer stem cells. Eur J Cancer. 2006;42:1213–8. doi: 10.1016/j.ejca.2006.01.037. [DOI] [PubMed] [Google Scholar]
- 39.Xin L, Lawson DA, Witte ON. The Sca-1 cell surface marker enriches for a prostate-regenerating cell subpopulation that can initiate prostate tumorigenesis. Proc Natl Acad Sci U S A. 2005;102:6942–7. doi: 10.1073/pnas.0502320102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rizzo S, Attard G, Hudson DL. Prostate epithelial stem cells. Cell Prolif. 2005;38:363–74. doi: 10.1111/j.1365-2184.2005.00356.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Heer R. Hunterian Lecture. Characterisation of human prostate epithelial progenitor differentiation in response to androgens. Ann R Coll Surg Engl. 2011;93:424–8. doi: 10.1308/10.1308/147870811x589245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Butler W, Huang J. Neuroendocrine cells of the prostate: histology, biological functions, and molecular mechanisms. Precis Clin Med. 2021;4:25–34. doi: 10.1093/pcmedi/pbab003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Harris KS, Kerr BA. Prostate cancer stem cell markers drive progression, therapeutic resistance, and bone metastasis. Stem Cells Int. 2017;2017:8629234. doi: 10.1155/2017/8629234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Zhang K, Zhou S, Wang L, et al. Current stem cell biomarkers and their functional mechanisms in prostate cancer. Int J Mol Sci. 2016;17:1163. doi: 10.3390/ijms17071163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Li S, Goncalves KA, Lyu B, Yuan L, Hu GF. Chemosensitization of prostate cancer stem cells in mice by angiogenin and plexin-B2 inhibitors. Commun Biol. 2020;3:26. doi: 10.1038/s42003-020-0750-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kerr BA, Miocinovic R, Smith AK, et al. CD117⁺ cells in the circulation are predictive of advanced prostate cancer. Oncotarget. 2015;6:1889–97. doi: 10.18632/oncotarget.2796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Chen X, Li Q, Liu X, et al. Defining a population of stem-like human prostate cancer cells that can generate and propagate castration-resistant prostate cancer. Clin Cancer Res. 2016;22:4505–16. doi: 10.1158/1078-0432.CCR-15-2956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yu C, Yao Z, Dai J, et al. ALDH activity indicates increased tumorigenic cells, but not cancer stem cells, in prostate cancer cell lines. In Vivo. 2011;25:69–76.. [PubMed] [Google Scholar]
- 49.Miyazawa K, Tanaka T, Nakai D, Morita N, Suzuki K. Immunohistochemical expression of four different stem cell markers in prostate cancer: high expression of NANOG in conjunction with hypoxia-inducible factor-1α expression is involved in prostate epithelial malignancy. Oncol Lett. 2014;8:985–92. doi: 10.3892/ol.2014.2274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Collins AT, Berry PA, Hyde C, Stower MJ, Maitland NJ. Prospective identification of tumorigenic prostate cancer stem cells. Cancer Res. 2005;65:10946–51. doi: 10.1158/0008-5472.CAN-05-2018. [DOI] [PubMed] [Google Scholar]
- 51.Clark DW, Palle K. Aldehyde dehydrogenases in cancer stem cells: potential as therapeutic targets. Ann Transl Med. 2016;4:518. doi: 10.21037/atm.2016.11.82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Castellón EA, Valenzuela R, Lillo J, et al. Molecular signature of cancer stem cells isolated from prostate carcinoma and expression of stem markers in different Gleason grades and metastasis. Biol Res. 2012;45:297–305. doi: 10.4067/S0716-97602012000300011. [DOI] [PubMed] [Google Scholar]
- 53.Rhodes DR, Sanda MG, Otte AP, Chinnaiyan AM, Rubin MA. Multiplex biomarker approach for determining risk of prostate-specific antigen-defined recurrence of prostate cancer. J Natl Cancer Inst. 2003;95:661–8. doi: 10.1093/jnci/95.9.661. [DOI] [PubMed] [Google Scholar]
- 54.Ugolkov AV, Eisengart LJ, Luan C, Yang XJ. Expression analysis of putative stem cell markers in human benign and malignant prostate. Prostate. 2011;71:18–25. doi: 10.1002/pros.21217. [DOI] [PubMed] [Google Scholar]
- 55.Trerotola M, Rathore S, Goel HL, et al. CD133, Trop-2 and α2β1 integrin surface receptors as markers of putative human prostate cancer stem cells. Am J Transl Res. 2010;2:135–44. [PMC free article] [PubMed] [Google Scholar]
- 56.Matsika A, Srinivasan B, Day C, et al. Cancer stem cell markers in prostate cancer: an immunohistochemical study of ALDH1, SOX2 and EZH2. Pathology. 2015;47:622–8. doi: 10.1097/PAT.0000000000000325. [DOI] [PubMed] [Google Scholar]
- 57.Narayanan R. Therapeutic targeting of the androgen receptor (AR) and AR variants in prostate cancer. Asian J Urol. 2020;7:271–83. doi: 10.1016/j.ajur.2020.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ku SY, Gleave ME, Beltran H. Towards precision oncology in advanced prostate cancer. Nat Rev Urol. 2019;16:645–54. doi: 10.1038/s41585-019-0237-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Guerrero J, Alfaro IE, Gómez F, Protter AA, Bernales S. Enzalutamide, an androgen receptor signaling inhibitor, induces tumor regression in a mouse model of castration-resistant prostate cancer. Prostate. 2013;73:1291–305. doi: 10.1002/pros.22674. [DOI] [PubMed] [Google Scholar]
- 60.Ojo D, Lin X, Wong N, Gu Y, Tang D. Prostate cancer stem-like cells contribute to the development of castration-resistant prostate cancer. Cancers (Basel) 2015;7:2290–308. doi: 10.3390/cancers7040890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Yu B, Liu Y, Luo H, et al. Androgen receptor splicing variant 7 (ARV7) inhibits docetaxel sensitivity by inactivating the spindle assembly checkpoint. J Biol Chem. 2021;296:100276. doi: 10.1016/j.jbc.2021.100276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Li T, Su Y, Mei Y, et al. ALDH1A1 is a marker for malignant prostate stem cells and predictor of prostate cancer patients’ outcome. Lab Invest. 2010;90:234–44. doi: 10.1038/labinvest.2009.127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Efremov YR, Proskurina AS, Potter EA, et al. Cancer stem cells: emergent nature of tumor emergency. Front Genet. 2018;9:544. doi: 10.3389/fgene.2018.00544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Leão R, Domingos C, Figueiredo A, et al. Cancer stem cells in prostate cancer: implications for targeted therapy. Urol Int. 2017;99:125–36. doi: 10.1159/000455160. [DOI] [PubMed] [Google Scholar]
- 65.Xu T, He K, Wang L, Goldkorn A. Prostate tumor cells with cancer progenitor properties have high telomerase activity and are rapidly killed by telomerase interference. Prostate. 2011;71:1390–400. doi: 10.1002/pros.21355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Rajasekhar VK, Studer L, Gerald W, Socci ND, Scher HI. Tumour-initiating stem-like cells in human prostate cancer exhibit increased NF-κB signalling. Nat Commun. 2011;2:162. doi: 10.1038/ncomms1159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.John J, Powell K, Conley-Lacomb MK, Chinni SR. TMPRSS2-ERG Fusion gene expression in prostate tumor cells and its clinical and biological significance in prostate cancer progression. J Cancer Sci Ther. 2012;4:94–101. doi: 10.4172/1948-5956.1000119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Nishida S, Hirohashi Y, Torigoe T, et al. Prostate cancer stem-like cells/cancer-initiating cells have an autocrine system of hepatocyte growth factor. Cancer Sci. 2013;104:431–6. doi: 10.1111/cas.12104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Zhang C, Yang Z, Dong DL, et al. 3D culture technologies of cancer stem cells: promising ex vivo tumor models. J Tissue Eng. 2020;11:2041731420933407. doi: 10.1177/2041731420933407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Vander Griend DJ, Karthaus WL, Dalrymple S, et al. The role of CD133 in normal human prostate stem cells and malignant cancer-initiating cells. Cancer Res. 2008;68:9703–11. doi: 10.1158/0008-5472.CAN-08-3084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Kanwal R, Shukla S, Walker E, Gupta S. Acquisition of tumorigenic potential and therapeutic resistance in CD133+ subpopulation of prostate cancer cells exhibiting stem-cell like characteristics. Cancer Lett. 2018;430:25–33. doi: 10.1016/j.canlet.2018.05.014. [DOI] [PubMed] [Google Scholar]
- 72.Santaliz-Ruiz LE, Xie X, Old M, Teknos TN, Pan Q. Emerging role of nanog in tumorigenesis and cancer stem cells. Int J Cancer. 2014;135:2741–8. doi: 10.1002/ijc.28690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Verma S, Shankar E, Kalayci FNC, et al. Androgen deprivation induces transcriptional reprogramming in prostate cancer cells to develop stem cell-like characteristics. Int J Mol Sci. 2020;21:9568. doi: 10.3390/ijms21249568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Sun Y, Liu Y, Ma X, Hu H. The influence of cell cycle regulation on chemotherapy. Int J Mol Sci. 2021;22:6923. doi: 10.3390/ijms22136923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Qiu H, Fang X, Luo Q, Ouyang G. Cancer stem cells: a potential target for cancer therapy. Cell Mol Life Sci. 2015;72:3411–24. doi: 10.1007/s00018-015-1920-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Glumac PM, LeBeau AM. The role of CD133 in cancer: a concise review. Clin Transl Med. 2018;7:18. doi: 10.1186/s40169-018-0198-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Yang L, Shi P, Zhao G, et al. Targeting cancer stem cell pathways for cancer therapy. Signal Transduct Target Ther. 2020;5:8. doi: 10.1038/s41392-020-0110-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hurwitz MD. Chemotherapy and radiation for prostate cancer. Transl Androl Urol. 2018;7:390–8. doi: 10.21037/tau.2018.03.07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Yeku O, Slovin SF. Immune therapy for prostate cancer. Cancer J. 2016;22:334–41. doi: 10.1097/PPO.0000000000000223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wang Z, Li Y, Wang Y, et al. Targeting prostate cancer stem-like cells by an immunotherapeutic platform based on immunogenic peptide-sensitized dendritic cells-cytokine-induced killer cells. Stem Cell Res Ther. 2020;11:123. doi: 10.1186/s13287-020-01634-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Han L, Shi S, Gong T, Zhang Z, Sun X. Cancer stem cells: therapeutic implications and perspectives in cancer therapy. Acta Pharmaceutica Sinica B. 2013;3:65–75. doi: 10.1016/j.apsb.2013.02.006. [DOI] [Google Scholar]
- 82.Zhang K, Guo Y, Wang X, et al. WNT/β-catenin directs self-renewal symmetric cell division of hTERThigh prostate cancer stem cells. Cancer Res. 2017;77:2534–47. doi: 10.1158/0008-5472.CAN-16-1887. [DOI] [PubMed] [Google Scholar]
- 83.Zhang Z, Cheng L, Li J, et al. Inhibition of the Wnt/β-catenin pathway overcomes resistance to enzalutamide in castration-resistant prostate cancer. Cancer Res. 2018;78:3147–62. doi: 10.1158/0008-5472.CAN-17-3006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Le PN, McDermott JD, Jimeno A. Targeting the Wnt pathway in human cancers: therapeutic targeting with a focus on OMP-54F28. Pharmacol Ther. 2015;146:1–11. doi: 10.1016/j.pharmthera.2014.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Schneider JA, Logan SK. Revisiting the role of Wnt/β-catenin signaling in prostate cancer. Mol Cell Endocrinol. 2018;462:3–8. doi: 10.1016/j.mce.2017.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Kelsey R. Prostate cancer: foxy-5 in prostate cancer model. Nat Rev Urol. 2017;14:638. doi: 10.1038/nrurol.2017.160. [DOI] [PubMed] [Google Scholar]
- 87.Raman S, Beilschmidt M, To M, et al. Structure-guided design fine-tunes pharmacokinetics, tolerability, and antitumor profile of multispecific frizzled antibodies. Proc Natl Acad Sci U S A. 2019;116:6812–7. doi: 10.1073/pnas.1817246116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Singh SK, Gordetsky JB, Bae S, et al. Selective targeting of the hedgehog signaling pathway by PBM nanoparticles in docetaxel-resistant prostate cancer. Cells. 2020;9:1976. doi: 10.3390/cells9091976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Fu J, Rodova M, Nanta R, et al. NPV-LDE-225 (Erismodegib) inhibits epithelial mesenchymal transition and self-renewal of glioblastoma initiating cells by regulating miR-21, miR-128, and miR-200. Neuro Oncol. 2013;15:691–706. doi: 10.1093/neuonc/not011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Gonnissen A, Isebaert S, McKee CM, et al. The hedgehog inhibitor GANT61 sensitizes prostate cancer cells to ionizing radiation both in vitro and in vivo. Oncotarget. 2016;7:84286–98. doi: 10.18632/oncotarget.12483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Rinkenbaugh AL, Baldwin AS. The NF-κB pathway and cancer stem cells. Cells. 2016;5:16. doi: 10.3390/cells5020016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Kaltschmidt C, Banz-Jansen C, Benhidjeb T, et al. A role for NF-κB in organ specific cancer and cancer stem cells. Cancers (Basel) 2019;11:655. doi: 10.3390/cancers11050655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Pietzak EJ, Eastham JA. Neoadjuvant treatment of high-risk, clinically localized prostate cancer prior to radical prostatectomy. Curr Urol Rep. 2016;17:37. doi: 10.1007/s11934-016-0592-4. [DOI] [PubMed] [Google Scholar]
- 94.Heath EI, Hillman DW, Vaishampayan U, et al. A phase II trial of 17-allylamino-17-demethoxygeldanamycin in patients with hormone-refractory metastatic prostate cancer. Clin Cancer Res. 2008;14:7940–6. doi: 10.1158/1078-0432.CCR-08-0221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Vidal AC, Howard LE, Moreira DM, et al. Aspirin, NSAIDs, and risk of prostate cancer: results from the REDUCE study. Clin Cancer Res. 2015;21:756–62. doi: 10.1158/1078-0432.CCR-14-2235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Ranganathan P, Weaver KL, Capobianco AJ. Notch signalling in solid tumours: a little bit of everything but not all the time. Nat Rev Cancer. 2011;11:338–51. doi: 10.1038/nrc3035. [DOI] [PubMed] [Google Scholar]
- 97.Takebe N, Miele L, Harris PJ, et al. Targeting notch, hedgehog, and Wnt pathways in cancer stem cells: clinical update. Nat Rev Clin Oncol. 2015;12:445–64. doi: 10.1038/nrclinonc.2015.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Stein MN, DiPaola RS, Mayer TM, et al. A randomized phase II study of bicalutamide (BIC) followed by placebo or gamma secretase inhibitor RO4929097 (RO492) in men with rising PSA. J Clin Oncol. 2012;30:219. doi: 10.1200/jco.2012.30.5_suppl.219. [DOI] [Google Scholar]
- 99.Begicevic RR, Falasca M. ABC transporters in cancer stem cells: beyond chemoresistance. Int J Mol Sci. 2017;18:2362. doi: 10.3390/ijms18112362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Liao CP, Adisetiyo H, Liang M, Roy-Burman P. Cancer stem cells and microenvironment in prostate cancer progression. Horm Cancer. 2010;1:297–305. doi: 10.1007/s12672-010-0051-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Giddings EL, Champagne DP, Wu MH, et al. Mitochondrial ATP fuels ABC transporter-mediated drug efflux in cancer chemoresistance. Nat Commun. 2021;12:2804. doi: 10.1038/s41467-021-23071-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Rybalchenko V, Prevarskaya N, Van Coppenolle F, et al. Verapamil inhibits proliferation of LNCaP human prostate cancer cells influencing K+ channel gating. Mol Pharmacol. 2001;59:1376–87. doi: 10.1124/mol.59.6.1376. [DOI] [PubMed] [Google Scholar]
- 103.Kawahara T, Kashiwagi E, Ide H, et al. The role of NFATc1 in prostate cancer progression: cyclosporine a and tacrolimus inhibit cell proliferation, migration, and invasion. Prostate. 2015;75:573–84. doi: 10.1002/pros.22937. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.