

Abstract
Cancer has been an insurmountable problem in the history of medical science. The uncontrollable proliferation of cancer cells is one of cancer’s main characteristics, which is closely associated with abnormal mitosis. Targeting mitosis is an effective method for cancer treatment. This review summarizes several natural products with anti-tumor effects related to mitosis, focusing on targeting microtubulin, inducing DNA damage, and modulating mitosis-associated kinases. Furthermore, the main disadvantages of several typical compounds, including drug resistance, toxicity to non-tumor tissues, and poor aqueous solubility and pharmacokinetic properties, are also discussed, together with strategies to address them. Improved understanding of cancer cell mitosis and natural products may pave the way to drug development for the treatment of cancer.
Keywords: Tumor, mitosis, natural products, mechanism, pharmacology
Introduction
According to data reported by International Agency for Research on Cancer, approximately 19.3 million new cancer cases and 10 million cancer deaths occurred worldwide in 20201. Although effective drugs for cancer treatment are continually being developed, cancer mortality is increasing each year2. Compared with normal cells, cancer cells have characteristics of uncontrollable proliferation, evasion of growth inhibitors, invasion and metastasis, and evasion of immune surveillance, thus making cancer difficult to cure and resulting in poor prognosis3. The uncontrollable proliferation of cancer cells, a major characteristic of cancer, is closely associated with abnormal mitosis. The mitosis of cancer cells is often unregulated, and cell division is accelerated by cancer cell evasion of growth inhibitory factors4. Targeting proliferation or mitosis are effective methods for cancer treatment. Many natural products exert favorable anti-tumor effects through regulating mitosis. The classical anti-mitotic drugs derived from natural products, including periwinkle alkaloids or paclitaxel-like drugs and their derivatives, have a wide range of clinical uses5.
Microtubules are involved in segregation during mitosis in eukaryotic cells. Microtubules are regulated primarily by polymerization kinetics, and their polymerization occurs through a nucleation-extension mechanism6. Targeting microtubules is currently used to prevent tumor cell mitosis and retard tumor development in clinical therapy7. DNA damage is directly associated with normal mitotic processes. If DNA damage is not repaired before mitosis occurs, the probability of chromosomal mutations is greatly increased, and the damaged DNA can enter daughter cells through faulty mitosis, and eventually induce cellular lesions or death8,9. Thus, promoting DNA damage in cancer cells and inhibiting repair can decrease cancer cell proliferation or promote cancer cell death. The growth of cancer cells is faster than that of normal cells, because cancer cells can avoid the inhibition of growth factors and unscheduled re-entry into the cell cycle10. Beyond microtubules and DNA damage, mitosis-associated kinases are another important aspect of mitosis that can be targeted. These kinases include cyclin-dependent kinases (CDKs) and aurora kinases (AURKs). Numerous CDKs regulate cell mitosis through binding cyclin proteins. The first CDK inhibitor to enter clinical studies was flavopiridol, derived from the native Indian plant rohitukine11. AURKs are major serine/threonine kinases in mitosis, which are involved in several events such as centrosome maturation and segregation, spindle assembly and maintenance, chromosome segregation, and cytoplasmic segregation. Studies have shown that inhibition of AURK expression or activity inhibits the proliferation, migration, and invasion of cancer cells12. Studies are identifying an increasing number of natural products that downregulate AURKs, and the modification of AURKA inhibitors may be a promising research direction.
This review summarizes several natural products with anti-tumor effects related to mitosis, focusing on targeting microtubulin, DNA damage, and mitosis-associated kinases. We also briefly summarize the mechanisms of mitosis targeting by these natural products in Table 1, and clinically developed antitumor drugs targeting mitosis derived from natural products in Table 2. Improved understanding of cancer cell mitosis and natural products may pave the way to further drug development for the treatment of cancer.
Table 1.
Summary of anti-tumor natural products targeting mitosis
| Herbal medicine | Source | Chemical structure | Target | Mechanism | Indications | References |
|---|---|---|---|---|---|---|
| Paclitaxel (Taxol) | Taxus brevifolia Nutt. |

|
Microtubules | Binds the paclitaxel site and causes microtubule polymerization | Breast cancer, ovarian cancer, head and neck cancer, lung cancer, etc. | 18–22 |
| Vincristine | Catharanthus roseus (L.) G. Don |

|
Microtubules | Binds the vincristine site and depolymerizes microtubules | Acute lymphoblastic leukemia, Hodgkin’s disease, reticulocytic sarcoma, breast cancer, etc. | 23,25,29 |
| Colchicine | Colchicum autumnale L. |
|
Microtubules | Binds colchicine sites and depolymerizes microtubules | Gout | 33,34 |
| Podophyllotoxin | Sinopodophyllumhexandrum (Royle) Ying |
|
Microtubules | Binds colchicine sites and depolymerizes microtubules | Condyloma acuminatum, acute granulocytic leukemia, chronic eosinophilic leukemia, small cell lung cancer, etc. | 37–40 |
| Aplyronine A | Aplysia kurodai |

|
Microtubules | Microtubule depolymerization | – | 41,42,45,46 |
| Pironetin | Streptomyces |

|
Microtubules | Binds alpha-microtubule protein and depolymerizes microtubules | – | 48,49 |
| Camptothecin | Camptotheca acuminata Decne. |

|
DNA damage | Topoisomerase I↓ | Stomach cancer, esophageal cancer, colorectal cancer, primary liver cancer, lung cancer, etc. | 54,61 |
| Piperine | Piper Nigrum L. |

|
DNA damage | ROS↑, tetraspiral DNA molecule↑, DNA polymerase β↓ | Convulsions, epilepsy, etc. | 65–69,71–73 |
| Sulforaphane | Brassica oleracea L.var.italic Planch. |

|
DNA damage | ROS↑, HDACs↓, γH2AX↑ | Chronic obstructive pulmonary disease | 74–80 |
| Resveratrol | Veratrum grandiflorum Loes. |

|
Cell cycle block | DDR↑, S phase block, apoptosis↑ | Atherosclerosis, coronary artery disease, ischemic heart disease, hyperlipidemia, breast cancer, etc. | 81–84,86–89 |
| Licochalcone A | Glycyrrhiza uralensis Fisch. |

|
Cell cycle block | Cyclin B1↓, CDK1↓, G2/M phase block | – | 91 |
| Hinokitiol | Chamaecyparisobtusa var. formosana (Hayata) Rehd. |
|
Cell cycle block | P53↑, D1-CDK4↓, G0/G1 phase block CDK7↓, Cyclin D1↓, Cyclin A2↓, S phase block |
Inhibition of Staphylococcus aureus | 92,93 |
| Cryptotanshinone | Salvia miltiorrhizaBge. |
|
Cell cycle block | Cyclin A/D↓, CDK2/4↓, G0/G1 phase block | Coronary heart disease, angina pectoris, etc. | 94 |
| Genistein | Euchresta japonica Hook. f. ex Regel |

|
Cell cycle block | Cyclin A/B↓, CDK2/CDC2↓, G2/M phase block p15↑, CDK↓, p21↑, Cyclin D1↓ |
– | 95–99 |
| Berberine | Coptis chinensis Franch. |

|
Cell cycle block | Cyclin D1/D2/E↓, CDK2/4/6↓, G1 phase block | Gastroenteritis, bacterial dysentery and other intestinal infections, conjunctivitis, suppurative otitis media, etc. | 100,101 |
| Ascleposide | Saussurea lappa |

|
Cell cycle block | Cyclin D1/A↓, CDK4↓, p21/27↓, G2/M phase block |
– | 102–104 |
| Garcinol | Garcinia |

|
Cell cycle block | Cyclin D1/D3↓, CDK2/4↓ Cyclin E↑, CDK6↑, p21/27↑, G1 phase block |
– | 108 |
| Curcumin | Curcuma longa L. |

|
Aurora kinase | AURKA↓, Histone H3↓, monopolar spindle↑ | Liver cancer, liver fibrosis, hyperlipidemia, Alzheimer’s disease, atherosclerosis, etc. | 124,125 |
| Tanshinone I | Salvia miltiorrhizaBge. |
|
Aurora kinase | AURKA↓ | Angina pectoris, coronary artery disease, etc. | 126–128 |
| Gossypin | Hibiscus vitifolius |

|
Aurora kinase | Binding ATP pocket of AURKA, AURKA↓ | – | 129–131 |
↑: promotes, ↓: inhibits.
Table 2.
Summary of clinical drugs derived from natural products that target mitosis
| Medicine | Source | Mechanisms | Indications | Approval status | References |
|---|---|---|---|---|---|
| Taxol | Paclitaxel | Maintains microtubule protein stability by inhibiting microtubule depolymerization and further inhibits cell mitosis | Ovarian cancer, breast cancer, lung cancer, head and neck cancer, esophageal cancer, germ cell tumors, endometrial cancer, lymphoma, bladder cancer, etc. | First marketed in 1992 | 24–26 |
| Abraxane | Metastatic breast cancer that has failed combination chemotherapy or that has recurred within 6 months after adjuvant chemotherapy | First marketed in 2005 | 27,28 | ||
| Cynviloq | Breast cancer, non-small cell lung cancer, etc. | First marketed in 2007 | – | ||
| Paclical | Lung cancer, breast cancer, ovarian cancer, etc. | First marketed in 2015 | 29 | ||
| Liporaxel | Breast cancer, stomach cancer, etc. | First marketed in 2016 | 32 | ||
| Oraxol | Metastatic breast cancer, etc. | In phase III clinical trials | 33,34 | ||
| Taxotere | Breast cancer and non-small cell lung cancer, etc. | First marketed in 1996 | 35,36 | ||
| Jevtana | Prostate cancer, etc. | First marketed in 2010 | – | ||
| Larotaxel | Breast cancer, etc. | In phase III clinical trials | – | ||
| Milataxel | Breast cancer, colorectal cancer, etc. | In phase II clinical trials | – | ||
| Ortataxel | Breast cancer, etc. | In phase II clinical trials | – | ||
|
Vinblastine
Sulfate |
Vinblastine | Prevents the formation of spindle microtubules by inhibiting microtubule protein polymerization, causing mitosis to stop at mid-phase | Malignant lymphoma, testicular tumor, choriocarcinoma, breast cancer, etc. | First marketed in 1958 | 45,46 |
|
Vincristine
Sulfate |
Vincristine | Leukemia, lymphoma, etc. | First marketed in 1962 | 45,46 | |
|
Vindesine
Sulfate |
Vinblastine | Non-small cell lung cancer, small cell lung cancer, malignant lymphoma, breast cancer, esophageal cancer, malignant melanoma, etc. | First marketed in 1980 | 45,46 | |
|
Vinorelbine
Ditartrate |
Vinblastine | Non-small cell lung cancer, breast cancer, etc. | First marketed in 1989 | 45,46 | |
| Javlor | Vinblastine | Urothelial carcinoma, etc. | First marketed in 2009 | 47–49 | |
| DPT | Podophyllotoxin | – | Non-small cell lung cancer, breast cancer, stomach cancer, etc. | In phase I clinical trials | 61,62 |
| Irinotecan | Camptothecin | Specifically inhibits DNA TopoI, which promotes double-stranded DNA deconvolution, thereby inhibiting DNA synthesis and achieving heterotypic cell growth | Colorectal cancer, lung cancer, breast cancer, pancreatic cancer, etc. | First marketed in 1994 | 92,93 |
| Topotecan | Small cell lung cancer, metastatic advanced ovarian cancer that has failed first-line chemotherapy, etc. | First marketed in 1996 | 92,93 | ||
| Hydroxycam-ptothecin | Liver cancer, colorectal cancer, lung cancer, leukemia, etc. | In phase III clinical trials | 94,95 | ||
| Exatecan | Breast cancer, ovarian cancer, colorectal cancer, etc. | In phase I clinical trials | 94,95 | ||
| Rubitecan | Most solid tumors, blood cancers, etc. | In phase III clinical trials | 96 | ||
| Flavopiridol | Rohitukine | Inhibits mitosis by inhibiting cyclin-dependent kinases, thus leading to cell cycle arrest | Esophageal cancer, lung cancer, prostate cancer, etc. | In phase II clinical trials | 167,168 |
| UCN-01 | Staurosporine | Lymphoma, etc. | In phase II clinical trials | 169 | |
| Midostaurin | Acute myeloid leukemia containing FLT-3 mutation, etc. | First marketed in 2017 | 170 | ||
| Roscovitine | Olomucine | Breast cancer, nasopharyngeal carcinoma, etc. | In phase I clinical trials | 171,172 | |
| Seliciclib | Breast cancer, nasopharyngeal carcinoma, etc. | In phase I clinical trials | 171,172 |
Microtubule
Microtubules in eukaryotic cells are polar cytoskeletal structures with crucial roles in cell mitosis, intracellular transport, and many other key cellular events. The cortex of microtubules is often composed of 13 protofilaments, mainly comprising tubulin heterodimers (α- and β-tubulin)13. Each heterodimer is arranged in a head-to-tail combination, thus forming directional microtubules and giving rise to the polar character of microtubules. The terminal with β-tubulin is the plus end, and the terminal with α-tubulin is the minus end6. Microtubules are hollow tubes that span the entire cell, and facilitate mitosis or intracellular transport13. Microtubules compose the spindle and are involved in all processes of mitosis. The kinetochore microtubules of the spindle emanate from the pole at one end and bind the centromeric chromatin at the other end14. Chromosomes that undergo replication have two mitoses, and bind two microtubules facing opposite spindle poles, thus achieving biorientation15. The aberrant expression of tubulin or microtubule-regulating proteins often occurs during tumor development7. For example, the high expression of β-microtubulin, particularly the βIII isoform, promotes tumor differentiation and metastasis. In addition, elevated levels of βIII-microtubulin affect resistance to broad-spectrum drugs and non-microtubule-targeting drugs16,17. Natural products targeting microtubule proteins are shown in Figure 1.
Figure 1 .

Natural products targeting microtubule proteins. Microtubules are composed of 2 types of microtubule proteins, α- and β-tubules, in a directed arrangement, which further form the spindle. The movement of microtubules is regulated, and inhibition or stabilization of microtubules interferes with mitosis. Paclitaxel is a microtubule stabilizer that binds the Taxane site of β-microtubule proteins. Vincristine, colchicine, podophyllotoxin, and aplyronine A are microtubule destabilizers that bind different sites of β-microtubule proteins. Pironetin acts on α-microtubule proteins and exerts microtubule depolymerizing effects.
Paclitaxel (Taxol)
Paclitaxel is a secondary metabolite isolated from the bark of the Pacific yew Taxusbrevifolia. Paclitaxel promotes the assembly of tubulin into microtubules and prevents the depolymerization of microtubules, thus further promoting the formation of stable microtubules from microtubulin, and effectively blocking the cell cycle at G2/M phase, preventing mitosis, and ultimately inhibiting proliferation or promoting apoptosis of cancer cells18. Recent research has shown that paclitaxel does not directly inhibit cell mitosis but instead induces the production of multipolar mitotic spindles that mediate unbalanced or mis-segregated chromosome distribution19,20. This abnormality leads to the formation of aneuploid progeny, loss of important chromosomes in the tumor cells, and ultimately cell death20. This mechanism partly explains the reason for paclitaxel insensitivity. Some clinical studies have shown that only approximately half of patients with breast cancer treated with paclitaxel have tumor shrinkage or disappearance. In patients who do not respond to paclitaxel, no clinically relevant paclitaxel is detected, and the mitosis of tumor cells does not cease21,22. In paclitaxel-sensitive patients, multipolar spindles form in tumor cells. When the abnormal multipolar spindle is maintained, the tumor cells die. After restoration of bipolar spindles, cancer cells usually become more resistant to paclitaxel.
Although paclitaxel is a highly successful antitumor drug in clinical practice, its disadvantages, such as poor water solubility, poor targeting, and strong non-selective toxic effects, have greatly hindered the development of new paclitaxel drugs23. Taxol, a first-generation paclitaxel preparation, has poor solubility, a short half-life, a long injection time, low bioavailability, and a tendency to cause allergic reactions24–26. Several new dosage forms of paclitaxel are available on the market. For example, albumin-bound paclitaxel, whose trade name is Abraxane, requires only half an hour to inject and can be used to treat breast cancer27,28. In 2007, Samyang launched Cynviloq, the first paclitaxel micelle formulation marketed worldwide. This dosage form has not only extended the half-life but also improved drug solubility. Paclical, which has a much higher drug loading than Cynviloq, was introduced to treat expanded indications29. Paclitaxel liposomes for injection are yet another formulation of paclitaxel that can be used as a first-line chemotherapy for ovarian cancer, or in combination with cisplatin30,31. In 2016, the Korean drug regulatory agency launched the world’s first paclitaxel oral solution (Liporaxel) for gastric cancer32. Subsequently, Hanmi Pharmaceuticals developed Oraxol, an innovative oral formulation of paclitaxel combined with ansquetta mesylate. Clinical studies have shown that Oraxol locally inhibits the activity of p-GP in the intestinal tract, thus improving drug efficacy while decreasing adverse effects elsewhere in the body33,34. After the development of new dosage forms of paclitaxel, paclitaxel derivatives followed. Sanofi developed Taxotere to address the problems of poor solubility and allergic reactions of paclitaxel, but this drug tends to cause permanent hair loss35,36. In China, in addition to Taxotere, which has been marketed, Jevtana, Larotaxel, Milataxel, and Ortataxel are under clinical trials.
Vincristine
Vincristine is a potent agent with well-defined effects on hematologic disorders and several types of solid tumors. As a microtubule-destabilizing drug, vincristine impairs the microtubule network by binding the Vinca domain in the β-tubulin subunit, thus inhibiting the polymerization of microtubules and arresting mitosis in phase M37. Vincristine forms a wedge at the dimer interface between α- and β-microtubulin while bending the microtubule growth axis38. Low concentrations restrain mitosis without depolymerization of the spindle in HeLa cells, whereas high concentrations induce rearrangement of spindle microtubules and centrosome rupture, thus arresting mitosis in M phase39. The transcription factor MYC, a key regulator of cellular proliferation, has been reported to be associated with highly aggressive cancers and poor prognosis in patients with cancer40. MYC overexpression induces mitotic spindle assembly defects through microtubule nucleation and organization41. Phosphorylation of MYC occurs in mitosis, but a fraction of non-phosphorylated MYC binds the microtubules of the mitotic spindle42,43. Vincristine has been found to destabilize wild-type MYC during mitosis. Furthermore, a strong inhibitory effect of vincristine has been observed in a population of lymphoma cells with high MYC expression44. The above results indicate that vincristine may be a good candidate in the treatment of MYC-driven malignancies.
Vincristines are typical antitumor drugs that target microtubules; the 4 main members are vinblastine, vincristine, vinorelbine, and vindesine45,46. Because the bioavailability of these 4 drugs is very low, they are administered mainly intravenously as sulfate and tartaric acid in clinical practice. Vinflunine (Javlor), a semi-synthetic derivative of periwinkle alkaloids, is a novel bifluorinated microtubule inhibitor for the treatment of bladder cancer that has failed to respond to chemotherapy47–49.
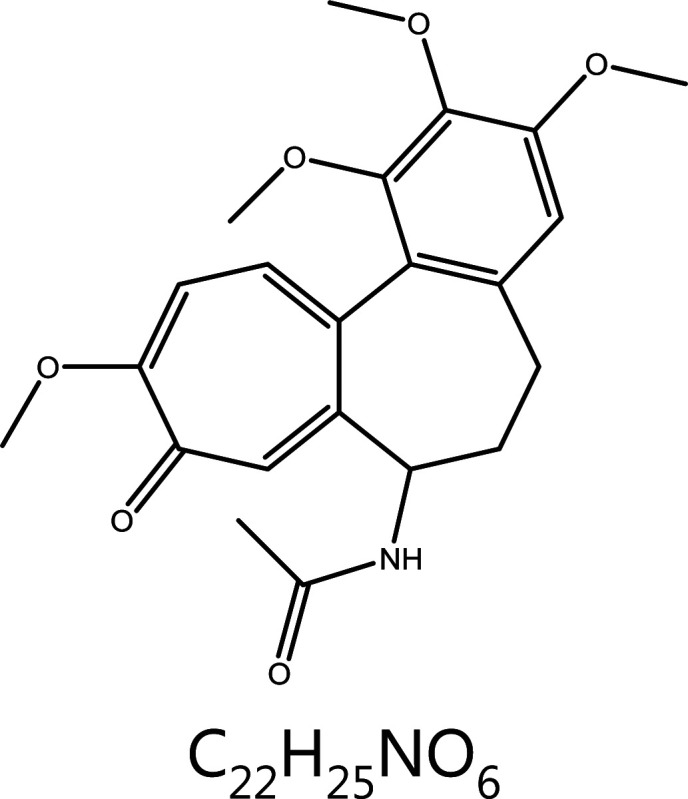
Colchicine
Colchicine, another microtubule-targeting agent (MTA), is a tricyclic alkaloid extracted from plants of the genus Colchicum (autumn crocus). This classical anti-mitotic drug binds soluble tubulin at colchicine site between α- and β-subunits, thus irreversibly forming tubulin-colchicine complexes, which induce many conformational changes in tubulin, disrupt further microtubule polymerization, and finally block mitosis at metaphase and induce cell death50. A detailed kinetic analysis of the inhibitory interaction has shown that tubulin-colchicine complexes bind the ends of the microtubules and prevent further addition to microtubules by sterically blocking terminal tubulin dimer growth50. The mitotic effect of colchicine mediated cell division is only temporary, and when the colchicine inside cells is metabolized, mitosis continues. In addition, the sensitivity of cells to colchicine varies according to its concentration. At low concentrations, colchicine arrests microtubule growth, whereas at higher concentrations, it promotes microtubule depolymerisation50. The severe toxicity of colchicine at high doses in normal tissues limits its clinical use as an anti-tumor agent51,52. Because colchicine has high anti-proliferative activity and promising clinical prospects, many colchicine analogues have been synthesized to avoid adverse effects53. However, because the effective dose of colchicine is close to the toxic dose, it is not commonly used clinically52. The main clinical applications of colchicine are currently the treatment of ventilation and cardiovascular diseases54. Because of the high toxicity of colchicine, experimental studies have synthesized derivatives with substituted functional groups on the a-ring (methoxy), b-ring (acetamide), and c-ring (methoxy) for potentiation and detoxification55.
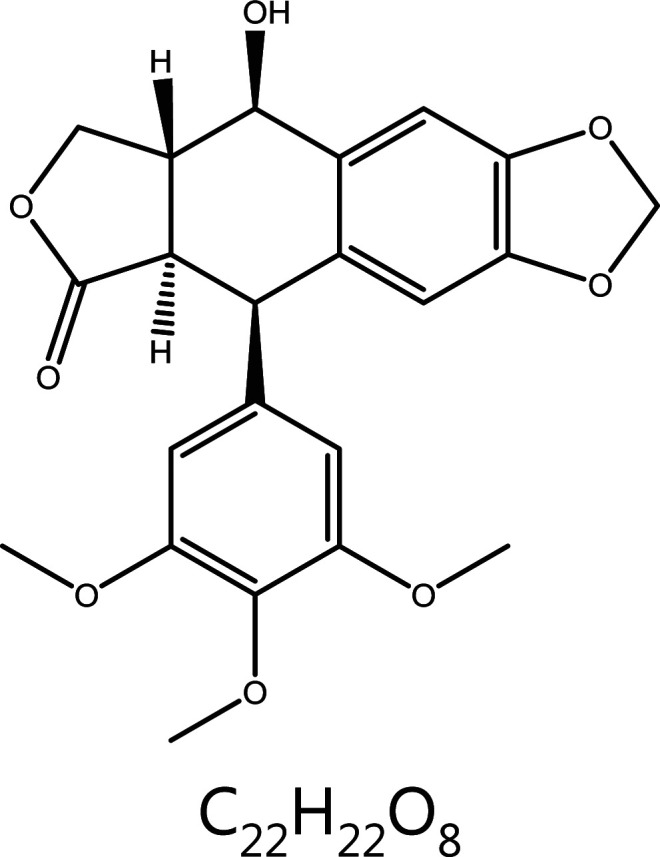
Podophyllotoxin
Podophyllotoxin, an aryltetralin-type lignan isolated from species of Podophyllum, is widely used to treat a variety of cancers including liver cancer, lung cancer, and neuroblastoma56. Similarly to colchicine, podophyllotoxin binds the colchicine site at the interface of α- and β-tubulin, inhibits the assembly of tubulin into microtubules, blocks the cell cycle in G2/M phase, and finally leads to mitosis arrest57,58. However, clinical therapy is severely limited because of systemic toxicity effects. Recently, modification of podophyllotoxin has enabled clinical application; the most clinically used drugs are etoposide and teniposide, which are glycosylated at the C-4 position59,60. Because podophyllotoxin is an effective targeted microtubule drug, drug development has continued. Deoxypodophyllotoxin (DPT) injection is currently under development in China in phase I trials. Various in vitro experimental studies on DPT have indicated its strong killing effect on a variety of human-derived tumor cells61. In vivo experiments have shown that DPT is effective against a variety of drug-resistant tumor strains, thus making it a valuable new antitumor drug62.
Aplyronine A (ApA)
ApA, a marine natural product originally isolated from Aplysiakurodai sea hares, exerts antitumor activity by inducing interactions between actin and microtubulin63–65. Actin is a class of globular multifunctional proteins that form microfilaments and are essential in diverse cellular processes, including movement and contraction. The dynamic balance between microtubules and actin is highly important for the stable morphology and mitosis of mammalian cells66. ApA specifically induces microtubule breakdown in the presence of actin. Conformation analysis of ApA on actin has indicated that the C13 methoxy group of ApA not only directly interacts with Glu334 of actin through the C9 hydroxyl group but also interacts with Arg147 through water molecules66. One study has indicated that ApA and actin synergistically inhibit microtubule polymerization67. ApA binds actin inserts its aliphatic tail into the actin molecule and binds the hydrophobic cleft consisting of actin subdomains 1 and 3, thus creating an actin-ApA complex68. The complex may further interact with microtubule protein heterodimers (released from the cytoplasm) and form a ternary complex, which microtubule polymerization65.
Pironetin
Pironetin, a Streptomyces polyketide metabolite, is a microtubule destabilizer that blocks cell mitosis in G2/M phase by inhibiting microtubule protein polymerization69. Unlike most MTAs, including paclitaxel, vincristine, colchicine, laulimalide, and medensin, pironetin is the only agent crystallographically characterized to bind solely to α-tubulin. Both α- and β-microtubulin bind GTP, but only β-microtubulin has GTPase activity and can hydrolyze GTP to GDP. Consequently, drugs binding the β-subunit are rapidly resistant. In contrast, the α-subunit has no GTPase activity and is not affected by mutations in β-microtubulin. Drugs targeting α-microtubulin have high clinical value. Pironetin inhibits the binding of radiolabeled vinblastine to tubulin, and has a stronger affinity for tubulin than vinblastine70. Researchers have found that pironetin disrupts the longitudinal contacts between microtubulin heterodimers and that the disruption may be the basis for promoting microtubule depolymerization. A re-examination of the molecular structure of pironetin has confirmed that pironetin covalently binds Cys316 of α-microtubulin, thus refuting the previous conclusion that it covalently binds Lys35270,71. The residues on both sides of Cys316 on the S8 chain differ from the side chain structures of Cys316 and Leu318, which are necessary for the binding of pironetin71. The distortion of the precise position of Glu254 and the resultant lower GTPase activity of β-tubulin may be an additional mechanism through which pironetin destabilizes the assembly of tubulin into microtubules71.
MTAs interact with tubulin, thus leading to cell death by altering microtubule dynamics. Natural MTAs, such as paclitaxel and vincristine, have long been used in cancer treatment72. We summarized the effects and mechanism of natural products targeting microtubulin, including paclitaxel, vincristine, colchicine, podophyllotoxin, ApA, and pironetin. Paclitaxel is a representative microtubule stabilizer that can result in the formation of aneuploid progeny by inducing multilevel spindle production, thus leading to tumor cell death. Others drugs include microtubule inhibitors, which block microtubule polymerization and disrupt microtubules and consequently mitosis. The special compound is pironetin, a recently discovered inhibitor of α-microtubulin, which disrupts the longitudinal contacts between microtubule protein heterodimers, thereby promoting microtubule depolymerization. Although MTAs are potent anti-tumoral drugs, their resistance and dose-limiting toxicity limit their clinical efficacy. To address the shortcomings of microtubule-targeted drugs, researchers have proposed the concept of dual-target microtubulin inhibitors, in which microtubulin inhibitors are combined with other antitumor drugs to achieve synergistic effects73,74. In recent years, to overcome the resistance and toxic effects of microtubulin inhibitors, particularly those developed from natural products, researchers have gradually focused on the development of dual-targeted drugs, such as paclitaxel dual-targeted nano-drugs75–78. The development of dual-targeted microtubule protein inhibitors is an area of great promise.
DNA damage
DNA damage is a permanent alteration of DNA nucleotide sequences caused by endogenous or exogenous factors, including spontaneous damage to DNA molecules, by-products of cellular metabolism, exogenous free radicals, and topological changes triggered by physical or chemical damage. DNA damage has been recognized as the underlying cause of many diseases, including cancer. Types of DNA damage include nucleotide mutations, substitutions, deletions, insertions, bulky adducts, single-strand breaks, and double-stand breaks (DSBs)79,80. DNA damage leads to genomic instability and induces functional mutations in oncogenes or tumor-suppressor genes, which are a causative factor for tumorigenesis81. DNA damage evokes responses by multiple repair mechanisms and signaling pathways. The DNA damage response (DDR) involves various intra- and inter-cellular signaling events and enzymatic activities that lead to cell-cycle arrest, regulation of DNA replication, and DNA damage repair82. Because genomic instability is a pervasive characteristic of tumor cells, DDR defects can serve as therapeutic targets. DNA damaging chemotherapies have been used in clinical cancer chemotherapy for decades, and include drugs that induce covalent crosslinks between DNA bases, drugs that attach alkyl groups to bases, and drugs that cause single-strand breaks or DSBs by targeting topoisomerase (Topo) I or II enzymes83. The natural products targeting DNA damage are shown in Figure 2.
Figure 2.

Natural products targeting DNA damage. DNA damage and repair (DDR) are closely associated with mitosis, and many natural products have been found to interfere with DNA damage or DDR. Camptothecin (CPT) directly binds DNA as a topoisomerase inhibitor. Sulforaphane (SFN) inhibits HDAC activity and enhances acetyltransferase expression, thus further causing DNA damage. Resveratrol (RSV) induces ROS accumulation, thereby leading to base double strand breakage. Piperine (PIP) inhibits the expression of DNA polymerase and prevents DDR. PIP also enhances GST, QR, and UDP-GT, and indirectly decreases DNA damage.
Camptothecin (CPT)
CPT is a pentacyclic alkaloid first isolated from Camptotheca acuminate. CPT is a typical TopoI inhibitor that induces S and G2 phase arrest. DNA TopoI resolves the tension caused during the winding and release of the DNA helix by forming a cleavage complex (TopoI covalent complex) that produces a transient DNA single-strand break, thus repairing the double-helix structure of DNA. CPT binds at the interface between TopoI and DNA and inhibits TopoI by binding to the Topo I-DNA complex84. TopoI inhibition of CPT is required for binding the TopoI/DNA covalent complex and forming a ternary complex that prevents rejoining of the broken DNA85,86. Early studies have demonstrated that CPT acts mainly on the DNA synthesis phase (S phase). CPT also affects pre-G1 phase and late-G2 phase, to a lesser extent than S phase87. The targets of CPT vary by concentration: low concentrations of CPT act on G2/M phase, whereas increasing concentrations block more cells in S phase88. Treatment with 25-nM CPT delays replication by several hours, whereas 1-μM CPT leads to a persistent replication block and massive cell death. Nanomolar concentrations (low doses) of CPT have been found to stimulate replication-fork slowing and reversal, whereas at high doses (micromolar levels), DSB formation is prevalent89. Mitotic arrest deficient 2 (Mad2), a mitotic checkpoint protein, is responsible for verifying the attachment of the mitophagus to the spindle90. CPT upregulates Mad2 through the Jun N-terminal kinase (JNK)-mediated SP1 pathway, and promotes microtubule protein polymerization and autophagy, thus leading to mitotic arrest and cell death91. Because of the poor solubility and severe toxicity of CPT, more effective CPT derivatives have been developed. Topotecan and irinotecan, water-soluble analogs, have been approved for treating common human cancer types92,93. Irinotecan and topotecan are semi-synthetic derivatives of CPT; irinotecan is the first-line drug for advanced colorectal cancer, and topotecan is a drug indicated for the treatment of breast cancer94,95. The CPT derivatives exhibit strong antitumor activity and low toxic adverse effects. Other marketed CPT derivatives, including hydroxylCPT and exatecan, have been tested in clinical trials and found to strongly block the effects of DNA synthesis in tumor cells96,97. Rubitecan, a second-generation topoisomerase inhibitor, is used to treat blood cancers and a variety of solid tumors98.
Piperine (PIP)
PIP is an acinnamamide alkaloid extracted from Piper nigrum and Piper longum, a commonly used spice99. PIP consists of 2 heterocycles linked by an aliphatic diene system, a structure that enables PIP to modulate multiple targets100. PIP exerts anti-tumor effects through a variety of mechanisms, such as inducing G1 cell cycle arrest101, inducing abnormal DNA structure, inducing ROS production and consequently disrupting redox homeostasis102, inhibiting tumor angiogenesis103, and inducing tumor cell autophagy.104. PIP directly binds the minor grooves of DNA105. Recent research has indicated that PIP shows specificity for G-quadruplex DNA and binds the DNA sequence of the c-myc promoter region (Pu24T). Thus, the abnormal DNA tetraspiral DNA molecule structure causes apoptosis106. In addition, in melanoma cells, PIP decreases the expression of DNA polymerase β, an enzyme that plays a role in the repair of DSBs, and prevents the repair of DNA damage. PIP also activates checkpoint kinase 1 (Chk1) and causes G1 phase cell cycle arrest107. Furthermore, PIP is well known for its ability to regulate metabolism enzymes, such as cytochrome P450. In an experimental model of benzo(a)pyrene [B(a)p]-induced lung cancer, PIP has been found to enhance the expression of detoxification enzymes, such as glutenzyme detoxification enzyme (GST), anthracene reductase (QR), and UDP-borate transferase (UDP-GT), thus indirectly decreasing DNA damage108.
Sulforaphane (SFN)
SFN, an isothiocyanate found in Brassicaceae vegetables, particularly broccoli, is recognized for its antioxidant capacity and anticancer activity109. SFN exhibits cytotoxic activity through several mechanisms including inducing detoxifying enzyme production, inducing ROS production, cell cycle arrest, apoptosis, and epigenetic regulation. Researchers have found that SFN inhibits histone deacetylase (HDAC) activity and enhances acetyltransferase expression, thus further causing DNA damage. Importantly, colon cancer cells are far more susceptible than non-cancer cells to SNF-induced DNA damage110. Myzak et al.111 first reported that SFN inhibits the activity of HDAC itself without altering the protein level of HDAC and β-catenin. HDAC inhibition has also been reported to disrupt the cell cycle in the G2 phase and interfere with the mitotic spindle checkpoint112, effects also caused by SFN. SFN upregulates CDK1 and cyclin B1 in HepG2 cells, which form a complex responsible for the cell cycle transition from G2 to mitosis. The cyclin B1-CDK1 maintenance induced by SFN causes misalignment of chromosomes to the spindle, thus arresting mitosis at prometaphase113. SFN-induced ROS generation is another important mechanism causing DNA damage and tumor cell apoptosis. For example, SFN inhibits and disrupts redox homeostasis by depleting glutathione (GSH), thereby inducing ROS production. The increase in expression of γH2AX, a DNA damage marker, after treatment with SFN, indicates the occurrence of DNA damage and a subsequent G0/G1 phase block in esophageal squamous cell carcinoma cells114. In fact, SFN exerts dichotomous effects in ROS production. SFN is also an indirect ROS scavenger through enhancing nuclear factor E2-related factor 2 (Nrf2) activity. SFN promotes the nuclear accumulation of Nrf2, which acts as a transcriptional activator via antioxidant response elements115.
Resveratrol (RSV)
RSV, a natural polyphenolic compound found in foods including grapes, berries, peanuts, and red wine, has potent anti-inflammatory, anti-bacterial, and anti-oxidant activities, and also has preventive and therapeutic effects on various cancers, as demonstrated in clinical trials116,117. In human head and neck squamous cell carcinoma cells, RSV induces DNA damage, mainly targeting mutant cancer cells without affecting normal cells, a phenomenon that is associated with Smad4118. In colon cancer models, RSV single treatment induces DDR correlated with S-phase delay and apoptosis; however, prolonged treatments lead to stable resistance which is triggered by ROS over-production. Research has indicated the importance of RSV-induced ROS production and the indirect DNA-damaging effects, as well as the long-term adaptation of cancer cells toward oxidative stress119. In extra nodal natural killer cell/T-cell lymphoma, an aggressive lymphoma with poor prognosis, RSV blocks the cell cycle in S-phase by activating the DDR pathway via upregulating the expression of Zta (also referred to as BZLF1, ZEBRA, EB1) of Epstein-Barr virus. The mechanism of DDR activation may involve an ATM/Chk2/p53-dependent pathway120,121. Etoposide is an antitumor agent that acts on DNA TopoII, thus causing DNA strand breaks; this chemotherapeutic agent impedes DNA repair122. Importantly, RSV synergistically activates the DDR pathway with etoposide, thus suppressing cell proliferation and promoting apoptosis123. In addition, studies have shown that RSV acts synergistically with etoposide, a DNA TopoII inhibitor, thereby causing DSBs, activating the DDR, and promoting apoptosis124.
DNA damage occurs in gene mutations and cancer development, and subsequently further activates the DDR to repair the damage. If the repair is not completed before mitosis, the damage is passed on to daughter cells and may lead to cellular senescence, apoptosis, and even cancer. Genomic instability is a feature of cancer cells that makes them susceptible to DNA damage. Targeting DNA damage and DDR inhibition holds promise for exploiting this defect in cancer cells, and the development of DNA damage inducers and targeted inhibition of DDR may lead to favorable anti-tumor treatments. The roles of natural products in DNA damage are diverse. Some compounds tend to induce ROS accumulation, thus leading to base double bond breakage; examples include PIP, carotenoids, and RSV. These compounds also indirectly cause DNA damage in tumor cells by regulating other proteins; their effects include the inhibitory effect of PIP on DNA polymerase β and the down-regulation of deacetylase by carotenoids. CPT plays a crucial role in the induction of DNA damage via directly binding DNA as a typical topoisomerase inhibitor. These natural products indirectly affect the mitosis of tumor cells by promoting DNA damage, thus leading to cell death. However, the specific mechanisms of most natural products in DNA damage remain unclear, and the identification of specific molecular targets is lacking.
Mitosis-associated kinases
In addition to targeting microtubules and targeting DNA damage, as described above, another highly important strategy is targeting mitosis-associated kinases. Mitosis cannot proceed properly without many protein phosphorylation events, which are regulated by protein kinases125,126. The protein kinases described in this review are CDK and AURK, which are frequently overexpressed in cancer cells and result in uncontrolled proliferation. In addition, recent studies have found that many natural products directly control the mitotic process of cancer cells by regulating these 2 kinases.
CDKs
CDKs drive the cell cycle primarily through chemotaxis on serine/threonine proteins, and act synergistically with cyclins, important factors in the regulation of the cell cycle. The cell division cycle is a strictly controlled process that leads to the duplication of genomic DNA and the reproduction of 2 daughter cells. The cell cycle is divided into 4 main phases: (1) G1 (gap 1) phase, known as prophase synthesis, which involves the synthesis of RNA and ribosomes in preparation for S phase; (2) S (synthesis) phase, involving synthesis of DNA and histones; (3) G2 (gap 2) phase, involving late DNA synthesis and preparation for mitosis; and (4) M (mitosis) phase, in which the cell divides the duplicated DNA and cellular components equally between daughter cells127. The cell cycle involves numerous CDKs, which form complexes with cyclin proteins and subsequently regulate cell mitosis128. Specific CDK–cyclin complexes drive various events in a sequential and orderly manner. During G1 phase, cyclin D senses mitogenic signals, its expression increases and it binds CDK4/6; the complex formed subsequently phosphorylates and inactivates the retinoblastoma (RB) family proteins. The partial inactivation of RB proteins allows for the expression of E-type cyclins (E1 and E2), which bind CDK2. CDK2–cyclin E complexes further phosphorylate and completely inactivate RBs, thereby releasing more E2F (a transcription factor) and activating the G1/S checkpoint129,130. CDK2 is subsequently activated by cyclin A, then drives the transition from S to G2 phase. In S phase, the accumulatied CDK2-cyclin A complex phosphorylates tricarboxylic acid (TCA) cyclase, thus enhancing glycolysis and providing energy for DNA replication129,131. Finally, CDK1 is activated by cyclin A and consequently promotes mitotic entry. After nuclear envelope rupture, cyclin A is degraded, thereby promoting the formation of the CDK1-cyclin B complex, which drives the cell cycle to mitosis128,129.
The cell cycle is commonly dysregulated in neoplasia, and the specific transcription of cell cycle proteins is usually aberrantly activated; cells subsequently undergo continued proliferation or unscheduled re-entry into the cell cycle132. In most tumors, CCND family members (encoding cyclin D) and CDK4/6 are overexpressed, thus eliminating RB-mediated cell cycle inhibition and leading to uncontrolled cell proliferation133,134. In addition, tumor development is usually accompanied by the up-regulation of cyclin E, which has a high affinity toward CDK2. The role of cyclin E in tumor development has been fully confirmed, whereas the role of CDK2 has not yet been elucidated135. Many studies have indicated that CDK2 inhibition retards malignancies, such as melanoma, ovarian cancer, and diffuse large B-cell lymphoma. The expression of cell cycle-related proteins is altered to some extent in most cancer cells, such as cyclin A, cell division cycle gene7 (CDC7), cyclin B-CDK1, and cyclin H-CDK7136. Natural products targeting CDKs are shown in Figure 3.
Figure 3.

Natural products regulating the cell cycle. An organized cell cycle is the basis for cell division. Many natural products act as cell cycle blockers via targeting cyclin proteins and CDKs. Licorice, hinokitiol, cryptotanshinone (CTS), berberine, ascleposide, and garcinol block the cell cycle in G0/G1 phase. Genistein blocks the cell cycle in G2/M phase and promotes apoptosis. Hinokitiol also decreases CDK7, cyclin D1, and cyclin A2, thereby blocking S phase.
Licorice
Glycyrrhiza uralensis Fisch (licorice) is a versatile herb whose active coupounds are mainly flavonoids and triterpene saponins. Licorice has anti-tumor effects through modulating the cell cycle or the immune microenvironment. A previous report has indicated that licorice inhits the proliferation of H1975 non-small cell lung cancer (NSCLC) cells and MCF-7 breast cancer cells in a dose-dependent manner, potentially through the down-regulation of cyclin D1-CDK4 complex expression, and blocks tumor cells in G0/G1 phase. The down-regulation of cyclin D1-CDK4 increases antigen presentation and induces infiltration of CD8+ T cells into tumors, thereby exerting anti-tumor effects137. Licochalcone A, a flavonoid isolated from licorice with anti-tumor properties, has been reported to inhibit the proliferation of HepG2 cells by terminating the cell cycle at the G2/M transition, as characterized by the decreased expression of cyclin B1 and CDK1138.
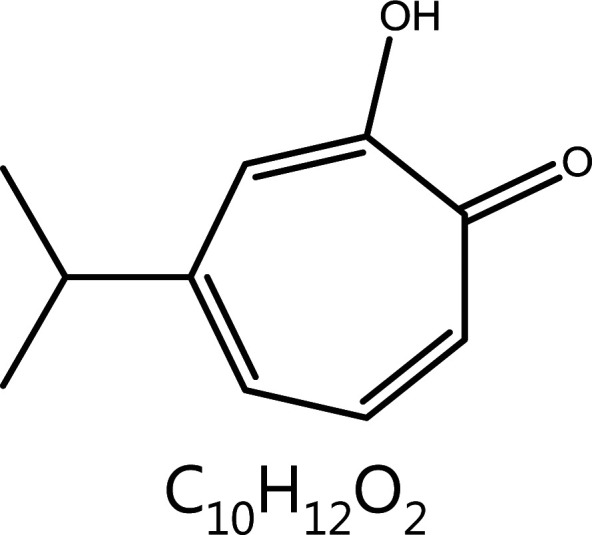
Hinokitiol
Hinokitiol, a natural monosteroidal compound with chro-phenolic ketone structure extracted from the trunks of Chamaecyparisobtusa, is often used as a potent fungicide. In recent years, hinokitiol has been found to have notable anti-tumor activity, including inhibiting tumor cell migration, promoting cell apoptosis, and inducing autophagy, thus highlighting its great potential for treating tumors. Hinokitiol treatment of endometrial carcinoid cells increases p53, decreases cyclin D1 and CDK4, and blocks cell growth in G0/G1 phase139. In human hepatocellular carcinoma cells, hinobinol mainly causes a decrease in CDK7, cyclin D1, and cyclin A2, thus blocking the S phase140.
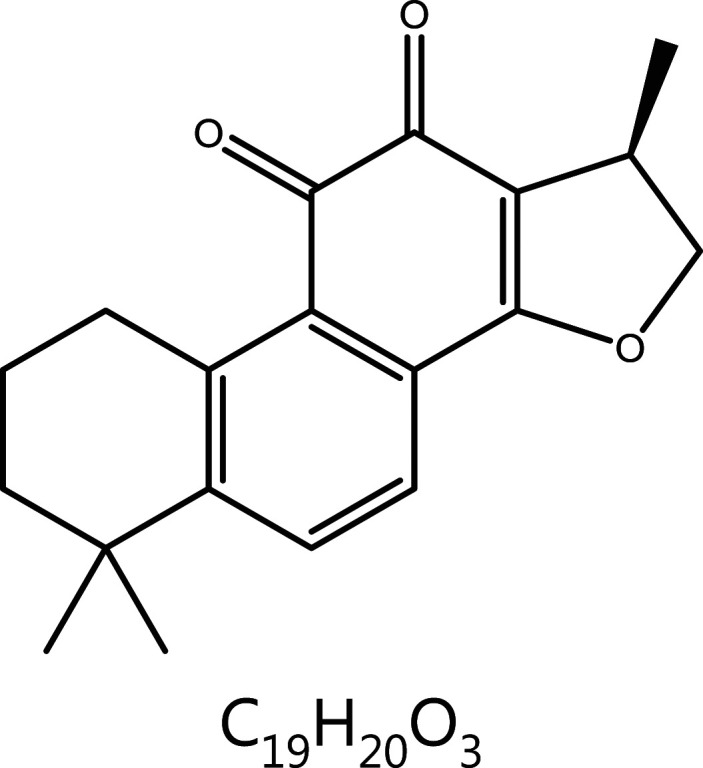
Cryptotanshinone (CTS)
Cryptotanshinone (CTS), a lipophilic compound extracted from Salvia miltiorrhiza Bunge, shows anti-tumor activity. In vitro experiments have indicated that CTS inhibits the viability of human NSCLC cells (A549 and H460), partly via down-regulating cyclin A/D and CDK2/4, thus blocking the cell cycle in G0/G1 phase141. Furthermore, CTS down-regulates IAP family proteins and apoptosis inhibitors, and promotes apoptosis, thus indicating a close relationship between CTS-induced cell cycle arrest and apoptosis.
Genistein
Genistein, an isoflavone extracted from Genista tinctoria Linn, exerts anti-tumor effects through apoptosis, cell cycle arrest, and inhibition of angiogenesis142. Genistein promotes apoptosis by inhibiting the activation of the PI3K/Akt signaling pathway. Cyclin A/B downregulation and binding CDK2 and CDC2 then block the cell cycle in G2/M phase and promote apoptosis143. In addition, in other tumor types, genistein indirectly plays a different role as a CDK inhibitor. In uterine smooth muscle tumors, genistein promotes expression of p15, a CDK inhibitor144. In colorectal cancer, the action of genistein is associated with the elimination of CDK2 phosphorylation145. In ovarian cancer, genistein mainly inhibits cyclin D1 expression but enhances p21 expression146.
Berberine
Berberine is a natural quaternary alkaloid isolated from Coptis chinensis Franch, with anti-inflammatory, anti-oxidant, and anti-tumor effects147. In vitro, berberine has been found to significantly decrease the expression of cyclin D1, D2, and E in DU145 human prostate cancer cells, with the most pronounced effect on cyclin D1. The down-regulation of CDK2, CDK4, and particularly CDK6 has also been observed148. The inhibition of cyclin D and CDKs by berberine indicates inhibition of of cyclin D-CDK6 complex formation, thus blocking the cell cycle in G1 phase.
Ascleposide
Ascleposide, a natural cardiac steroid lactone isolated from Saussurea lappa, has been found to inhibit DU145 prostate cancer cell growth149. Ascleposide downregulates the expression of cyclin D1, cyclin A, and CDK4 in a dose-dependent manner, and also downregulaets c-Myc (an early identified oncogenes) in human prostate cancer cells150. C-myc indirectly regulates the cell cycle through the dissociation of the CDK repressor protein p21 from the CDK4/6-cyclin D complex151. Therefore, ascleposide exerts indirect and direct effects on cell cycle regulation, and decreases the G1 distribution of cells while increasing the proportion of cells in G2/M and sub-G1 phase150. The p21 and p27 Cip/Kip proteins are crucial CDK-interacting protein/kinase inhibitory proteins in mitotis152. Interestingly, ascleposide downregulates these Cip/Kip proteins rather than upregulating them150, possibly because p21 and p27 resist the apoptotic response in cancer cells153,154, thus explaining the ascleposide-induced p21 and p27 downregulation, and anticancer activity in CRPC cells150.
Others
Garcinol, a polyisoprene benzophenone extracted from the fruit of Garcinia indica, exerts anti-inflammatory and anti-cancer activities. Garcinol induces decreases in cyclin D1, D3, CDK2, and CDK4, and increases in cyclin E and CDK6 in human NSCLC cells (H1299)155. In addition, Garcinol upregulates the CDK repressor proteins p21Waf1/cip1 and p27Kip1, thus inhibiting cyclin-CDK complex assembly. P21Waf1/cip1 knockdown results in a complete cell cycle block in G1 phase, thus indicating that the garcinol-induced G1 block in H1299 cells is achieved by upregulation of p21Waf1/cip1 protein expression. However, the effect is unclear in H460 cells155. Therefore, the mechanism underlying garcinol-induced cell cycle arrest requires further investigation. Similar G1 arrest by 13-O-acetylsolstitialin A and Tetrandrine has been observed, both of which act on the cyclin D1-CDK4 complex: 13-O-acetylsolstitialin A decreases the expression of cyclin D1 and CDK4156, whereas Tetrandrine binds the cyclin D1-CDK4 complex and forms hydrogen bonds157.
Many natural products also induce G2/M arrest, but their related molecular mechanisms are not well studied. Examples include Polygonatum cyrtonema Hua158, Paris chinensis dioscin159, Lycorisaurea agglutinin160, Strychni Semen (ESS)161, and 6-shogaol162. Some cause clear S phase arrest by downregualting cyclin A and CDK2 (Juglanthraquinone C163 and tea polyphenols164) or activating CDK repressor proteins (Litchi chinensis Sonn165–167 and Gynostemma L168).
Normal progression of the cell cycle is necessary to ensure that mitosis occurs without error. The disruption of cell cycle control is a common feature in human cancers169. Many natural products can block the cell cycle, including licorice, Hinokitiol, and Cryptotanshinone. However, most studies have been limited to examining their effects on cyclins and kinases, whereas in-depth molecular studies are lacking. Moreover, the effects of most compounds have not been confirmed in vivo.
In recent years, clinical development of natural product-derived CDK inhibitors has become more difficult, although increasing natural products have been discovered. Flavopiridol, the first inhibitor of CDKs to enter clinical trials worldwide, is a semi-synthetic flavonoid derivative170. However, in phase II clinical trials, researchers have found that it did not show single drug activity in a variety of cancers but showed very significant dose-limiting toxicity171. Consequently, Flavopiridol has not been marketed to date. UCN-01 is a derivative of staurosporine that is currently in phase II clinical trials. It binds protein kinase competitively with ATP and has a good selective effect on kinases172. Modification of the structure of astrosporin has led to the discovery of a derivative with multi-target kinase inhibitory activity, midostaurin, which was successfully marketed in 2017173. Roscovitine and seliciclib are tautomers derived from the natural product olomoucine. Seliciclib has stronger CDK inhibitory activity than Roscovitine, and it blocks RNA polymerase II phosphorylation by inhibiting CDK7 and CDK9 activity174,175. Currently, seliciclib has entered phase I and II clinical trials as a drug candidate.
Aurora kinase
The AURK family is an important class of serine/threonine kinases responsible for the regulation of cell mitosis. Its 3 members are known as Aurora-A (AURKA), Aurora-B (AURKB), and Aurora-C (AURKC) in mammalian cells. AURKA is responsible for regulating mitotic spindle assembly and centrosome segregation, as well as facilitating the transition of cells from G2 to M phase. AURKB is the catalytic component of the chromosome pericentromere complex, which regulates chromatin protein modifications, and promotes chromosome segregation and cytoplasmic division. AURKC is expressed in testis cells and is thought to be important for chromosome segregation during meiosis. AURKA and AURKB play key roles in mitosis.
These kinases are associated with tumorigenesis and are often overexpressed in tumor cells. For example, AURKA is usually highly expressed and actively repairs damaged DNA, thus prompting a return to mitosis. AURKA overexpression constitutively activates CDK1 and abolishes the G2/M DNA damage checkpoint in cancer cells176. In addition, AURKA promotes the proliferation and migration of cancer cells during mitotic interphase. AURKB is also a mitotic driver, is overexpressed in a variety of tumor tissues, and promotes tumor invasion. Although AURKC executes unique roles in meiosis, those roles are not discussed here. Given that AURK overexpression is associated with tumorigenesis, these kinases have been potential targets for cancer therapy, and several AURK inhibitors have been developed. In recent years, an increasing number of natural products have shown similar anti-tumor effects to AURK inhibitors. Natural products targeting AURKs are shown in Figure 4.
Figure 4.

Natural products for targeting AURKs to inhibit mitosis. Aurora kinase (AURKs) are involved in mitotic processes, and down-regulation of AURKs in cancer cells inhibits mitosis. Curcumin inhibits AURKA and changes the bipolar spindle to unipolar, thus inhibiting the continuation of mitosis. Gossypin, as well as 2 active components of eriocaulon sieboldianum (HPDL and QGGP), binds the ATP pocket of AURKA, thereby depleting AURKA.
Curcumin
Curcumin, a hydrophobic polyphenol extracted from rhizome herbs such as Zingiberaceae Martinov and AraceaeJuss, has a broad spectrum of biological and pharmacological activities, including anti-inflammatory and anti-tumor effects. Curcumin has been found to decrease the AURKA protein and kinase activity in human breast chemoresistant nonmetastatic MCF-7 and highly metastatic cancer MDA-MB-231 cells. The effects of curcumin are similar to those of AURKA siRNA treatment, including monopolar spindle formation, S and G2/M arrest, and diminished cell division177. In human bladder cancer cells, curcumin down-regulates the promoter activity of AURKA, thus inhibiting AURKA at the transcriptional level. In addition, curcumin inhibits the phosphorylation of AURKA and its downstream target histone H3 along with the formation of a monopolar spindle. Eventually, G2/M phase arrest occurs, thus decreasing cell division178. In summary, the mitotic inhibitory effects of curcumin on cancer cells may be closely associated with the down-regulation of AURKA. In addition to targeting AURKA, curcumin inhibits mitosis by promoting DNA damage, including DNA double-strand breaks179, and activation of DDR and pathways180. Thus, curcumin is an anti-tumor natural product worthy of in-depth study.
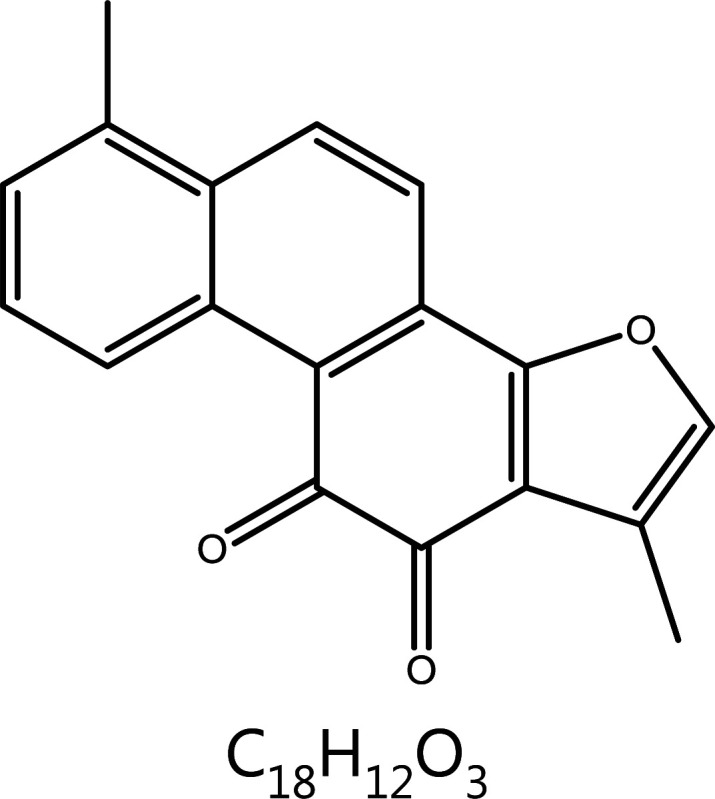
Tanshinone I
Danshen (Salvia miltiorrhiza Bunge) has been widely used in China for centuries to treat coronary heart diseases. Tanshinones such as cryptotanshinone (CT), tanshinone IIA (T2A), and tanshinone I (T1), are diterpene compounds in Danshen that have been shown to possess anti-tumor activity. Among the studied tanshinones, T1 has strong anti-tumor effects partially attributed to AURKA. In lung cancer, T1 significantly inhibits cell proliferation and tumor growth, but this effect is dramatically eliminated by AURKA knockdown181. T1 has shown similar effects in prostate cancer182 and breast cancer183. In particular, in breast cancer cells (MDA-MB231), only T1, but not other tanshinones, shows a potent inhibitory effect on cell growth; therefore, T1 causes cell cycle arrest in both estrogen-dependent and estrogen-independent cells. T1 regulates the expression of cyclin D, CDK4 and cyclin B, and inhibits the expression of AURKA. Tanshinones appear to have few adverse effects on normal mammary epithelial cells183. In summary, AURKA is an important target for tanshinones.
Gossypin
Gossypin, a flavone originally isolated from Hibiscus vitifolius, has many pharmacological activities, including anti-angiogenesis, anti-inflammation, and anti-tumor functions184. Computational docking results have indicated that gossypin directly binds the ATP pocket of AURKA, thus exerting inhibitory effects on AURKA. In human gastric cancer cells, Gossypin inhibits AURKA and induces a G2/M phase block, thereby leading to apoptosis185. A similar binding ATP pocket of AURKA has been found in 2 active components of Eriocaulon sieboldianum (HPDL and QGGP) leading to G2/M phase arrest in HepG2 cells186.
Others
P3, a high molecular weight compound isolated from a marine sponge called Crambetailliezi, selectively inhibits AURKA and AURKB, and decreases the mitotic index and apoptotic phenotype of U-2 OS sarcoma cells187.
Some TCM formulas or single herbs have also shown inhibitory effects on AURKA, although the active compounds remains unclear. San Huang Decoction (SHD), a Chinese herbal formula made from 5 Chinese herbs, has been widely used to treat metabolic syndrome for centuries and has been used for clinical treatment of patients with breast cancer188. SHD has been found to inhibit the proliferaiton of MCF-7 and MDA-MB-231 human breast cancer cells, an effect associated with the down-regulation of AURKA189. In addition, by targeting AURKA, SHD may inhibit tumor angiogenesis in breast cancer190. Hedyotis diffusa Willd. (H) and Scutellaria barbata D.Don (S) are 2 ancient anti-cancer Chinese herbs that are used in combined “HS” treatment in prostate cancer190. HS inhibits the G2 to M phase transformation of prostate cancer cells by decreasing the protein and mRNA levels of AURKA, CDK1, PLK1, and Cyclin B1190. The active components and molecular mechanisms of these herbs require more extensive research.
AURKs are crucial in the spindle assembly checkpoint, alignment of metaphase chromosomes, and chromosomal biorientation. AURKA and AURKB play key roles in mitosis, and some natual products targeting AURKA and AURKB exert anti-tumor effects in vitro. Curcumin, tanshinone I, and Gossypin act on AURKA, and P3 acts on AURKA and AURKB in vitro. Some TCM formulas, such as SHD, show similar effects on AURKA. However, the mechanism and efficacy in vivo require further in-depth study.
Althrough natural products with AURKA inhibitory effects are gradually being discovered, no AURKA inhibitors have been marketed to date or entered clinical trials, owing to the multi-target effects, weak selectivity, and strong toxic adverse effects of natural products. Currently, highly selective AURKA inhibitors have become a focus of new drug development and clinical research.
Other mitosis-related kinases
Polo-like kinase 1 (PLK1), a new target for mitosis discovered in recent years, belongs to a group of serine/threonine kinases widely found in eukaryotic cells, and is mainly regulated by G1-phase transcriptional repressors and G2-phase transcriptional activators191. Recent studies have revealed that the overexpression of PLK1 not only promotes multiple defects in mitosis but also leads to the accumulation of chromosomal instability, thus favoring aneuploidy and tumor formation192. PLK1 is a promising target for inhibiting mitosis in cancer cells. Scutellaria baicalensis Georgi has strong anti-PLK1 activity, and its main active components, baicalin and baicalin, covalently bind PLK1 via Cys133, thus providing a basis for further development of drugs targeting PLK1193. In vitro experiments with Glucocappasalin have demonstrated its ability to inhibit PLK1 expression in HeLa cells and to induce cell cycle G2/M phase arrest194. In addition, Jerantinine B195, caffeic acid phenethyl ester,196 and catechins197 have anti-PLK1 effects in various types of cancer cells.
Monopolar spindle1 (Mps1) is a serine/threonine kinase that ensures proper orientation of sister chromatids on the mitotic spindle, mainly through the spindle assembly checkpoint198,199. Treatment of mitotic colorectal cancer cell lines with Mps1 inhibitors results in cell death200. T-LAK cell-originated protein kinase (TOPK) is a serine/threonine kinase in the mitogen-activated protein kinase kinase (MAPKK) family that plays a role in cell cycle and mitotic progression201. Although little research has been performed on the natural products that regulate Mps1 and TOPK, these kinases are of great interest as promising targets for mitosis, and the discovery of regulatory natural products is believed to provide a reliable basis for clinical trials.
Discussion
Mitosis is an essential and conserved process in the life cycle of mammalian cells, and is uncontrolled in cancer cells. Thus, interfering with mitosis was an early concept for cancer therapy that has been demonstrated to be successful in clinical settings. In recent years, natural products have attracted increasing attention in cancer therapy, owing to their structural and target diversity. This review summarized the clinical uses of experimental anti-tumor natural products targeting multiple elements during mitosis, including microtubules, the DDR, CDKs, and AURKs. Although many anti-tumor natural products have achieved good clinical effects—including paclitaxel, CPTs, and vincristine—drug resistance, poor targeting, and poor druggability are important factors limiting the clinical application or drug development.
Drug resistance is a major problem in clinical settings. A typical representative of resistance is paclitaxel, a drug that issensitive to βIII-microtubulin16. Studies have confirmed that the high expression of βIII in tumor cells is an important reason for paclitaxel resistance, in cancers including lung cancer, ovarian cancer, breast cancer, and gastric cancer. Overexpression of tubulin-βIII decreases the ability of paclitaxel to suppress microtubule dynamics, thus resulting in resistance202. Another study on the cytotoxicity of HT29-D4 paclitaxel human colorectal cancer cells has demonstrated that undifferentiated and differentiated cancer cells differ in their sensitivity to paclitaxel, owing to altered βIII-microtubule protein expression. Furthermore, the selective overexpression of tubulin αII and tubulin βI and βIV accounts for the resistance of vincristine203, in accordance with the conclusion that differential overexpression of the tubulin isotype is a cause of drug resistance. Resistance to CPTs, DNA TopoI inhibitors, remains not completely clear. The structural alteration of TopoI and cellular response alterations in CPT-DNA-ternary complex formation are the 2 factors in drug resistence204. A growing body of research is focusing on the therapeutic targets to ameliorate drug resistance. For example, in the treatment of triple-negative breast cancer, paclitaxel resistance can be reversed by decreasing LIN9 expression, thus suggesting that paclitaxel resistance in triple-negative breast cancer can be reversed by combining bromo- and extra-terminal domain inhibitors with paclitaxel. In addition, compared with β-tubulin-targeted MTA, α-targeted natural products (such as Pironetin) have better selectivity and therefore are in urgent need of development.
Another crucial factor limiting clinical application or use in other therapeutic areas is toxicity toward normal cells. A major problem in using MTA in cancer treatment is its high systemic toxicity, including bone marrow suppression, gastrointestinal bleeding, hair loss, and tingling of the hands and feet. For example, the major toxicities caused by paclitaxel are neutropenia and peripheral neuropathies. Colchicine has been used to treat gout for many years, but its high toxicity at anti-tumor doses, possibly because of its quasi-irreversible binding tubulin, has limited its anticancer therapy development. Because of colchicine’s severe toxicity, many colchicine analogues have been developed to decrease toxicity and maintain efficacy. Studies have shown that modifying colchicine substituents alters binding affinity, and thus the inhibition of tubulin polymerization and cytotoxicity. A simple analog “AC,” retaining only the A and C-ring of colchicine, binds tubulin instantaneously and reversibly, owing to the low activation energy of colchicine–tubulin binding50. Different functional groups, such as amides and thioamides, have been introduced in colchicine. An A-ring derivative, 4-halo-(-)-colchicine shows strong activity against human cancer cells, including lung adenocarcinoma A549, colon adenocarcinoma HT29, and HCT116, with the high selectivity and the low toxicity205.
The aqueous insolubility or poor pharmacokinetic properties of some natural products have limited their clinical efficacy. Structural modification is one method to increase solubility. For example, the low aqueous solubility and bioavailability of CPT has been largely addressed with chemically modified topotecan and irinocand. In addition to structural modification of natural products, novel delivery systems, such as nanoparticles, provide another way to address current challenges. Various nanoparticles have been developed for delivery of anti-tumor active compounds206. Researchers have developed PCL-TPGS (caprolactone-Tocopherol Polyethylene Glycol Succ)-loaded paclitaxel, and the area under the curve of paclitaxel increased about 2.7-fold, the t1/2 increased, and the clearance rate decreased compared to free paclitaxel207. Similarly, compared with free curcumin, phospholipid-, chitosan- or lipid nanocapsule-loaded curcumin show greater areas under the curve and Cmax, and are more efficiently delivered into tumor cells208–210. Moreover, beyond improving the solubility and pharmacokinetic properties, some nanoparticles further enhance the stability and selectivity of drugs, thereby increasing the efficacy or decreasing the toxicity.
In the face of problems associated with the development of clinical drugs of natural product origin, researchers have modified dosage forms and structures to increase efficacy and decrease toxicity. Regarding clinical drugs targeting microtubules, the classical example is paclitaxel, although its insolubility and drug resistance are a major challenge. Several new dosage forms and new structures have been developed to solve the problem during a 50-year-long research process. The emergence of upgraded dosage forms, such as albumin paclitaxel, paclitaxel liposomes, paclitaxel micelles, paclitaxel nanoformulations, and paclitaxel oral formulations, has expanded the indications for paclitaxel drugs. Beyond dosage form modification, many structural modifications to paclitaxel have been developed, such as cabazitaxel, docetaxel, milataxel, ortataxel, salutaxel, simotaxel, and paclitaxel ceribate. The development path of paclitaxel provides a good example for other natural products to solve the clinical development problems of natural products through dosage form modification and structural modification. Beyond the problems of high toxicity, low bioavailability, poor solubility, and high drug resistance of some natural products, the clinical drug development of natural products also faces the problem of unclear multiple targets. Although the multi-target characteristics of natural products can decrease drug resistance, they complicate mechanistic studies. Currently, many new techniques for target prediction are available, such as reverse molecular docking and 3D pharmacophore modeling techniques211,212. Wei et al.213 have used the parameters of protein-ligand conformation interaction patterns as important indicators for target prediction and screening; this new strategy for target prediction effectively improved the accuracy of small molecule target identification.
In summary, many anti-tumor natural products have been introduced to date, and their anti-tumor activity and molecular targets are receiving increasing attention. Here, we focused on the inhibition of mitosis in tumor cells. Paclitaxel, CPT, and vincristine are typical examples of natural anti-tumor drugs, which have favorable anti-mitosis effects. As the mechanisms of these drugs have been explored, new discoveries have provided clinical advantages. The potential of natural products in anti-tumor drug development is enormous. Simultaneously, natural products have disadvantages such as toxicity and low efficacy, which must be improved in the future. Currently, drug development is increasingly becoming inclined toward structural modification and dosage modification of natural products with significant antitumor effects, and many new drug delivery systems have been developed to expand the drug delivery capacity and improve drug targeting. This review is intended to provide ideas for researchers to discover or develop more potent and selective anti-cancer agents.
Acknowledgements
We thank Professor Wenwei Hu (Cancer Institute of New Jersey, USA) for linguistic assistance during the preparation of this manuscript.
Grant support
This work was supported by the National Key Research and Development Program of China (Grant No: 2021YFE0203100), National Natural Science Foundation of China (Grant Nos: 81873089, 81603253 and 81973570), Innovation Team and Talents Cultivation Program of National Administration of Traditional Chinese Medicine (Grant No: ZYYCXTD-C-202009), and Innovation Team and Talents Cultivation Program of National Administration of Traditional Chinese Medicine (Grant No: ZYYCXTD-D-202002).
Conflict of interest statement
No potential conflicts of interest are disclosed.
Author contributions
Conceived and designed the analysis: Haiyang Yu, Weiling Pu.
Collected the data: Manru Huang, Caiyan Liu.
Contributed data or analysis tools: Manru Huang, Yingying Shao, Shiyue Zhou.
Performed the analysis: Caiyan Liu, Gaoyong Hu, Shuangshuang Yin.
Wrote the paper: Manru Huang, Caiyan Liu.
References
- 1.Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality Worldwide for 36 Cancers in 185 Countries. CA: a Cancer Journal For Clinicians. 2021;71:209–49. doi: 10.3322/caac.21660. [DOI] [PubMed] [Google Scholar]
- 2.Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394–424. doi: 10.3322/caac.21492. [DOI] [PubMed] [Google Scholar]
- 3.Trosko JE. On the potential origin and characteristics of cancer stem cells. Carcinogenesis. 2021;42:905–12. doi: 10.1093/carcin/bgab042. [DOI] [PubMed] [Google Scholar]
- 4.McIntosh JR. Mitosis. Cold Spring Harbor Perspect Biol. 2016;8(9):a023218. doi: 10.1101/cshperspect.a023218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Steinmetz MO, Prota AE. Microtubule-targeting agents: strategies to hijack the Cytoskeleton. Trends Cell Biol. 2018;28:776–92. doi: 10.1016/j.tcb.2018.05.001. [DOI] [PubMed] [Google Scholar]
- 6.Gudimchuk NB, McIntosh JR. Regulation of microtubule dynamics, mechanics and function through the growing tip. Nat Rev Mol Cell Biol. 2021;22:777–95. doi: 10.1038/s41580-021-00399-x. [DOI] [PubMed] [Google Scholar]
- 7.Jordan MA, Wilson L. Microtubules as a target for anticancer drugs. Nat Rev Cancer. 2004;4:253–65. doi: 10.1038/nrc1317. [DOI] [PubMed] [Google Scholar]
- 8.Lezaja A, Altmeyer M. Dealing with DNA lesions: when one cell cycle is not sufficient. Curr Opin Cell Biol. 2021;70:27–36. doi: 10.1016/j.ceb.2020.11.001. [DOI] [PubMed] [Google Scholar]
- 9.Orthwein A, Fradet-Turcotte A, Noordermeer SM, Canny MD, Brun CM, Strecker J, et al. Mitosis inhibits DNA double-strand break repair to guard against telomere fusions. Science. 2014;344:189–93. doi: 10.1126/science.1248024. [DOI] [PubMed] [Google Scholar]
- 10.Williams GH, Stoeber K. The cell cycle and cancer. J Pathol. 2012;226:352–64. doi: 10.1002/path.3022. [DOI] [PubMed] [Google Scholar]
- 11.Sedlacek HH. Mechanisms of action of flavopiridol. Crit Rev Oncol/Hematol. 2001;38:139–70. doi: 10.1016/s1040-8428(00)00124-4. [DOI] [PubMed] [Google Scholar]
- 12.Du R, Huang C, Liu K, Li X, Dong Z. Targeting AURKA in Cancer: molecular mechanisms and opportunities for cancer therapy. Mol Cancer. 2021;20:15. doi: 10.1186/s12943-020-01305-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Goodson HV, Jonasson EM. Microtubules and microtubule-associated proteins. Cold Spring Harb Perspect Biol. 2018;10:a022608. doi: 10.1101/cshperspect.a022608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kops G, Gassmann R. Crowning the kinetochore: the fibrous corona in chromosome segregation. Trends Cell Biol. 2020;30:653–67. doi: 10.1016/j.tcb.2020.04.006. [DOI] [PubMed] [Google Scholar]
- 15.Magidson V, O’Connell CB, Loncarek J, Paul R, Mogilner A, Khodjakov A. The spatial arrangement of chromosomes during prometaphase facilitates spindle assembly. Cell. 2011;146:555–67. doi: 10.1016/j.cell.2011.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kavallaris M. Microtubules and resistance to tubulin-binding agents. Nat Rev Cancer. 2010;10:194–204. doi: 10.1038/nrc2803. [DOI] [PubMed] [Google Scholar]
- 17.Khwaja S, Kumar K, Das R, Negi AS. Microtubule associated proteins as targets for anticancer drug development. Bioorg Chem. 2021;116:105320. doi: 10.1016/j.bioorg.2021.105320. [DOI] [PubMed] [Google Scholar]
- 18.Horwitz SB. Mechanism of action of taxol. Trends Pharmacol Sci. 1992;13:134–36. doi: 10.1016/0165-6147(92)90048-b. [DOI] [PubMed] [Google Scholar]
- 19.Scribano CM, Wan J, Esbona K, Tucker JB, Lasek A, Zhou AS, et al. Chromosomal instability sensitizes patient breast tumors to multipolar divisions induced by paclitaxel. Sci Transl Med. 2021;13:eabd4811. doi: 10.1126/scitranslmed.abd4811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Weaver BA. How Taxol/paclitaxel kills cancer cells. Mol Biol Cell. 2014;25:2677–81. doi: 10.1091/mbc.E14-04-0916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Budd GT, Barlow WE, Moore HC, Hobday TJ, Stewart JA, Isaacs C, et al. SWOG S0221: a phase III trial comparing chemotherapy schedules in high-risk early-stage breast cancer. J Clin Oncol. 2015;33:58–64. doi: 10.1200/JCO.2014.56.3296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fountzilas G, Dafni U, Dimopoulos MA, Koutras A, Skarlos D, Papakostas P, et al. A randomized phase III study comparing three anthracycline-free taxane-based regimens, as first line chemotherapy, in metastatic breast cancer: a Hellenic Cooperative Oncology Group study. Breast Cancer Res Treat. 2009;115:87–99. doi: 10.1007/s10549-008-0047-9. [DOI] [PubMed] [Google Scholar]
- 23.Ma Y, Yu S, Ni S, Zhang B, Kung ACF, Gao J, et al. Targeting strategies for enhancing paclitaxel specificity in chemotherapy. Front Cell Devel Biol. 2021;9:626910. doi: 10.3389/fcell.2021.626910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mu L, Feng SS. A novel controlled release formulation for the anticancer drug paclitaxel (Taxol): PLGA nanoparticles containing vitamin E TPGS. J Control Release. 2003;86:33–48. doi: 10.1016/s0168-3659(02)00320-6. [DOI] [PubMed] [Google Scholar]
- 25.Fenyvesi F, Kiss T, Fenyvesi E, Szente L, Veszelka S, Deli MA, et al. Randomly methylated beta-cyclodextrin derivatives enhance taxol permeability through human intestinal epithelial Caco-2 cell monolayer. J Pharm Sci. 2011;100:4734–44. doi: 10.1002/jps.22666. [DOI] [PubMed] [Google Scholar]
- 26.Runowicz CD, Wiernik PH, Einzig AI, Goldberg GL, Horwitz SB. Taxol in ovarian cancer. Cancer. 1993;71:1591–96. doi: 10.1002/cncr.2820710442. [DOI] [PubMed] [Google Scholar]
- 27.Moreno-Aspitia A, Perez EA. North Central Cancer Treatment Group N0531: Phase II Trial of weekly albumin-bound paclitaxel (ABI-007; Abraxane) in combination with gemcitabine in patients with metastatic breast cancer. Clin Breast Cancer. 2005;6:361–4. doi: 10.3816/CBC.2005.n.042. [DOI] [PubMed] [Google Scholar]
- 28.Albumin-bound paclitaxel (Abraxane) for advanced breast cancer. Med Lett Drugs Ther. 2005;47:39–40. [PubMed] [Google Scholar]
- 29.Bernabeu E, Cagel M, Lagomarsino E, Moretton M, Chiappetta DA. Paclitaxel: What has been done and the challenges remain ahead. Int J Pharm. 2017;526:474–95. doi: 10.1016/j.ijpharm.2017.05.016. [DOI] [PubMed] [Google Scholar]
- 30.Zhang J, Pan Y, Shi Q, Zhang G, Jiang L, Dong X, et al. Paclitaxel liposome for injection (Lipusu) plus cisplatin versus gemcitabine plus cisplatin in the first-line treatment of locally advanced or metastatic lung squamous cell carcinoma: a multicenter, randomized, open-label, parallel controlled clinical study. Cancer Commun (Lond) 2022;42:3–16. doi: 10.1002/cac2.12225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ye L, He J, Hu Z, Dong Q, Wang H, Fu F, et al. Antitumor effect and toxicity of Lipusu in rat ovarian cancer xenografts. Food Chem Toxicol. 2013;52:200–6. doi: 10.1016/j.fct.2012.11.004. [DOI] [PubMed] [Google Scholar]
- 32.Yang JI, Jin B, Kim SY, Li Q, Nam A, Ryu MO, et al. Antitumour effects of Liporaxel (oral paclitaxel) for canine melanoma in a mouse xenograft model. Vet Comp Oncol. 2020;18:152–60. doi: 10.1111/vco.12540. [DOI] [PubMed] [Google Scholar]
- 33.Shanmugam S, Im HT, Sohn YT, Kim YI, Park JH, Park ES, et al. Enhanced oral bioavailability of paclitaxel by solid dispersion granulation. Drug Dev Ind Pharm. 2015;41:1864–76. doi: 10.3109/03639045.2015.1018275. [DOI] [PubMed] [Google Scholar]
- 34.Lee KW, Lee KH, Zang DY, Park YI, Shin DB, Kim JW, et al. Phase I/II study of weekly oraxol for the second-line treatment of patients with metastatic or recurrent gastric cancer. Oncologist. 2015;20:896–7. doi: 10.1634/theoncologist.2015-0202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Jedličková H, Vokurka S, Vojtíšek R, Malečková A. Alopecia and hair damage induced by oncological therapy. Klin Onkol. 2019;32:353–9. doi: 10.14735/amko2019353. [DOI] [PubMed] [Google Scholar]
- 36.Kong TH, Han SM, Seo YJ. Chondritis of the ear after docetaxel-carboplatin chemotherapy. J Oncol Pharm Pract. 2019;25:975–9. doi: 10.1177/1078155218762629. [DOI] [PubMed] [Google Scholar]
- 37.Groth-Pedersen L, Ostenfeld MS, Høyer-Hansen M, Nylandsted J, Jäättelä M. Vincristine induces dramatic lysosomal changes and sensitizes cancer cells to lysosome-destabilizing siramesine. Cancer Res. 2007;67:2217–25. doi: 10.1158/0008-5472.CAN-06-3520. [DOI] [PubMed] [Google Scholar]
- 38.Stanton RA, Gernert KM, Nettles JH, Aneja R. Drugs that target dynamic microtubules: a new molecular perspective. Med Res Rev. 2011;31:443–81. doi: 10.1002/med.20242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Jordan MA, Thrower D, Wilson L. Effects of vinblastine, podophyllotoxin and nocodazole on mitotic spindles. implications for the role of microtubule dynamics in mitosis. J Cell Sci. 1992;102(Pt 3):401–16. doi: 10.1242/jcs.102.3.401. [DOI] [PubMed] [Google Scholar]
- 40.Ahmadi SE, Rahimi S, Zarandi B, Chegeni R, Safa M. MYC: a multipurpose oncogene with prognostic and therapeutic implications in blood malignancies. J Hematol Oncol. 2021;14:121. doi: 10.1186/s13045-021-01111-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rohrberg J, Van de Mark D, Amouzgar M, Lee JV, Taileb M, Corella A, et al. MYC dysregulates mitosis, revealing cancer vulnerabilities. Cell Rep. 2020;30:3368–82.e3367. doi: 10.1016/j.celrep.2020.02.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Koch HB, Zhang R, Verdoodt B, Bailey A, Zhang CD, Yates JR, et al. Large-scale identification of c-MYC-associated proteins using a combined TAP/MudPIT approach. Cell Cycle. 2007;6:205–17. doi: 10.4161/cc.6.2.3742. [DOI] [PubMed] [Google Scholar]
- 43.Sjostrom SK, Finn G, Hahn WC, Rowitch DH, Kenney AM. The Cdk1 complex plays a prime role in regulating N-myc phosphorylation and turnover in neural precursors. Dev Cell. 2005;9:327–38. doi: 10.1016/j.devcel.2005.07.014. [DOI] [PubMed] [Google Scholar]
- 44.Becker S, Kiecke C, Schafer E, Sinzig U, Deuper L, Trigo-Mourino P, et al. Destruction of a microtubule-bound MYC reservoir during mitosis contributes to vincristine’s anticancer activity. Mol Cancer Res. 2020;18:859–72. doi: 10.1158/1541-7786.MCR-19-1203. [DOI] [PubMed] [Google Scholar]
- 45.Takigawa N, Tanimoto M. [Vinca alkaloid and MDR1] Gan To Kagaku Ryoho. 2008;35:1086–89. [PubMed] [Google Scholar]
- 46.Mondal A, Gandhi A, Fimognari C, Atanasov AG, Bishayee A. Alkaloids for cancer prevention and therapy: current progress and future perspectives. Eur J Pharmacol. 2019;858:172472. doi: 10.1016/j.ejphar.2019.172472. [DOI] [PubMed] [Google Scholar]
- 47.Frampton JE, Moen MD. Vinflunine. Drugs. 2010;70:1283–93. doi: 10.2165/11204970-000000000-00000. [DOI] [PubMed] [Google Scholar]
- 48.Yun-San Yip A, Yuen-Yuen Ong E, Chow LW-C. Vinflunine: clinical perspectives of an emerging anticancer agent. Expert Opin Investig Drugs. 2008;17:583–91. doi: 10.1517/13543784.17.4.583. [DOI] [PubMed] [Google Scholar]
- 49.Bernardini A, Duenas M, Martin-Soberon MC, Rubio C, Suarez-Cabrera C, Ruiz-Palomares R, et al. Genomic landscape of vinflunine response in metastatic urothelial cancer. Cancers (Basel) 2022;14:378. doi: 10.3390/cancers14020378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Bhattacharyya B, Panda D, Gupta S, Banerjee M. Anti-mitotic activity of colchicine and the structural basis for its interaction with tubulin. Med Res Rev. 2008;28:155–83. doi: 10.1002/med.20097. [DOI] [PubMed] [Google Scholar]
- 51.Eleftheriou G, Bacis G, Fiocchi R, Sebastiano R. Colchicine-induced toxicity in a heart transplant patient with chronic renal failure. Clin Toxicol (Phila) 2008;46:827–30. doi: 10.1080/15563650701779703. [DOI] [PubMed] [Google Scholar]
- 52.Finkelstein Y, Aks SE, Hutson JR, Juurlink DN, Nguyen P, Dubnov-Raz G, et al. Colchicine poisoning: the dark side of an ancient drug. Clin Toxicol (Phila) 2010;48:407–14. doi: 10.3109/15563650.2010.495348. [DOI] [PubMed] [Google Scholar]
- 53.Krzywik J, Aminpour M, Maj E, Mozga W, Wietrzyk J, Tuszyński JA, et al. New series of double-modified colchicine derivatives: synthesis, cytotoxic effect and molecular docking. Molecules. 2020;25:3540. doi: 10.3390/molecules25153540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Robinson PC, Terkeltaub R, Pillinger MH, Shah B, Karalis V, Karatza E, et al. Consensus statement regarding the efficacy and safety of long-term low-dose colchicine in gout and cardiovascular disease. Am J Med. 2022;135:32–8. doi: 10.1016/j.amjmed.2021.07.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Dubey KK, Kumar P, Labrou NE, Shukla P. Biotherapeutic potential and mechanisms of action of colchicine. Crit Rev Biotechnol. 2017;37:1038–47. doi: 10.1080/07388551.2017.1303804. [DOI] [PubMed] [Google Scholar]
- 56.Antúnez-Mojica M, Rodríguez-Salarichs J, Redondo-Horcajo M, León A, Barasoain I, Canales Á, et al. Structural and biochemical characterization of the interaction of tubulin with potent natural analogues of podophyllotoxin. J Nat Prod. 2016;79:2113–21. doi: 10.1021/acs.jnatprod.6b00428. [DOI] [PubMed] [Google Scholar]
- 57.Giri A, Lakshmi Narasu M. Production of podophyllotoxin from Podophyllum hexandrum: a potential natural product for clinically useful anticancer drugs. Cytotechnology. 2000;34:17–26. doi: 10.1023/A:1008138230896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Sackett DL. Podophyllotoxin, steganacin and combretastatin: natural products that bind at the colchicine site of tubulin. Pharmacol Therapeutics. 1993;59:163–228. doi: 10.1016/0163-7258(93)90044-e. [DOI] [PubMed] [Google Scholar]
- 59.Zhao Y, Li D, Wei M, Du R, Yan Z. The ester derivatives obtained by C-ring modification of podophyllotoxin induce apoptosis and inhibited proliferation in PC-3M cells via down-regulation of PI3K/Akt signaling pathway. Bioorg Med Chem Lett. 2021;46:128174. doi: 10.1016/j.bmcl.2021.128174. [DOI] [PubMed] [Google Scholar]
- 60.Zhao W, Cong Y, Li HM, Li S, Shen Y, Qi Q, et al. Challenges and potential for improving the druggability of podophyllotoxin-derived drugs in cancer chemotherapy. Nat Prod Rep. 2021;38:470–88. doi: 10.1039/d0np00041h. [DOI] [PubMed] [Google Scholar]
- 61.Gamage CDB, Park SY, Yang Y, Zhou R, Taş İ, Bae WK, et al. Deoxypodophyllotoxin exerts anti-cancer effects on colorectal cancer cells through induction of apoptosis and suppression of tumorigenesis. Int J Mol Sci. 2019;20:2612. doi: 10.3390/ijms20112612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Zang X, Wang G, Cai Q, Zheng X, Zhang J, Chen Q, et al. A Promising microtubule inhibitor deoxypodophyllotoxin exhibits better efficacy to multidrug-resistant breast cancer than paclitaxel via avoiding efflux transport. Drug Metab Dispos. 2018;46:542–51. doi: 10.1124/dmd.117.079442. [DOI] [PubMed] [Google Scholar]
- 63.Futaki K, Takahashi M, Tanabe K, Fujieda A, Kigoshi H, Kita M. Synthesis and biological activities of aplyronine a analogues toward the development of antitumor protein-protein interaction inducers between actin and tubulin: conjugation of the C1-C9 macrolactone part and the C24-C34 side chain. ACS Omega. 2019;4:8598–613. doi: 10.1021/acsomega.9b01099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Yamada K, Ojika M, Kigoshi H, Suenaga K. Aplyronine A, a potent antitumour macrolide of marine origin, and the congeners aplyronines B-H: chemistry and biology. Nat Prod Rep. 2009;26:27–43. doi: 10.1039/b800263k. [DOI] [PubMed] [Google Scholar]
- 65.Kita M, Kigoshi H. Marine natural products that interfere with multiple cytoskeletal protein interactions. Nat Prod Rep. 2015;32:534–42. doi: 10.1039/c4np00129j. [DOI] [PubMed] [Google Scholar]
- 66.Chakrabarti KR, Hessler L, Bhandary L, Martin SS. Molecular Pathways: New signaling considerations when targeting cytoskeletal balance to reduce tumor growth. Clin Cancer Res. 2015;21:5209–14. doi: 10.1158/1078-0432.CCR-15-0328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Kita M, Hirayama Y, Yoneda K, Yamagishi K, Chinen T, Usui T, et al. Inhibition of microtubule assembly by a complex of actin and antitumor macrolide aplyronine A. J Am Chem Soc. 2013;135:18089–95. doi: 10.1021/ja406580w. [DOI] [PubMed] [Google Scholar]
- 68.Hirata K, Muraoka S, Suenaga K, Kuroda T, Kato K, Tanaka H, et al. Structure basis for antitumor effect of aplyronine a. J Mol Biol. 2006;356:945–54. doi: 10.1016/j.jmb.2005.12.031. [DOI] [PubMed] [Google Scholar]
- 69.Torijano-Gutierrez S, Vilanova C, Diaz-Oltra S, Murga J, Falomir E, Carda M, et al. The mechanism of the interactions of pironetin analog/combretastatin A-4 hybrids with tubulin. Arch Pharm (Weinheim) 2015;348:541–7. doi: 10.1002/ardp.201500106. [DOI] [PubMed] [Google Scholar]
- 70.Usui T, Watanabe H, Nakayama H, Tada Y, Kanoh N, Kondoh M, et al. The anticancer natural product pironetin selectively targets Lys352 of alpha-tubulin. Chem Biol. 2004;11:799–806. doi: 10.1016/j.chembiol.2004.03.028. [DOI] [PubMed] [Google Scholar]
- 71.Yang J, Wang Y, Wang T, Jiang J, Botting CH, Liu H, et al. Pironetin reacts covalently with cysteine-316 of α-tubulin to destabilize microtubule. Nat Commun. 2016;7:12103. doi: 10.1038/ncomms12103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Castro-Gamero AM, Pezuk JA, Brassesco MS, Tone LG. G2/M inhibitors as pharmacotherapeutic opportunities for glioblastoma: the old, the new, and the future. Cancer Biol Med. 2018;15:354–74. doi: 10.20892/j.issn.2095-3941.2018.0030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Tanabe K. Microtubule depolymerization by kinase inhibitors: unexpected findings of dual inhibitors. Int J Mol Sci. 2017;18:2508. doi: 10.3390/ijms18122508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Stanković T, Dinić J, Podolski-Renić A, Musso L, Burić SS, Dallavalle S, et al. Dual inhibitors as a new challenge for cancer multidrug resistance treatment. Curr Med Chem. 2019;26:6074–106. doi: 10.2174/0929867325666180607094856. [DOI] [PubMed] [Google Scholar]
- 75.Yang H-W, Hua M-Y, Liu H-L, Tsai R-Y, Chuang C-K, Chu P-C, et al. Cooperative dual-activity targeted nanomedicine for specific and effective prostate cancer therapy. ACS Nano. 2012;6:1795–805. doi: 10.1021/nn2048526. [DOI] [PubMed] [Google Scholar]
- 76.Madamsetty VS, Pal K, Dutta SK, Wang E, Mukhopadhyay D. Targeted dual intervention-oriented drug-encapsulated (DIODE) nanoformulations for improved treatment of pancreatic cancer. Cancers (Basel) 2020;12:1189. doi: 10.3390/cancers12051189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Houdaihed L, Evans JC, Allen C. In vivo evaluation of dual-targeted nanoparticles encapsulating paclitaxel and everolimus. Cancers (Basel) 2019;11:752. doi: 10.3390/cancers11060752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Grojean M, Schwarz MA, Schwarz JR, Hassan S, von Holzen U, Zhang C, et al. Targeted dual inhibition of c-Met/VEGFR2 signalling by foretinib improves antitumour effects of nanoparticle paclitaxel in gastric cancer models. J Cell Mol Med. 2021;25:4950–61. doi: 10.1111/jcmm.16362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.O’Connor MJ. Targeting the DNA damage response in cancer. Mol Cell. 2015;60:547–60. doi: 10.1016/j.molcel.2015.10.040. [DOI] [PubMed] [Google Scholar]
- 80.Curtin NJ. DNA repair dysregulation from cancer driver to therapeutic target. Nat Rev Cancer. 2012;12:801–17. doi: 10.1038/nrc3399. [DOI] [PubMed] [Google Scholar]
- 81.Gillman R, Lopes Floro K, Wankell M, Hebbard L. The role of DNA damage and repair in liver cancer. Biochim Biophys Acta Rev Cancer. 2021;1875:188493. doi: 10.1016/j.bbcan.2020.188493. [DOI] [PubMed] [Google Scholar]
- 82.Sun S, Osterman MD, Li M. Tissue specificity of DNA damage response and tumorigenesis. Cancer Biol Med. 2019;16:396–414. doi: 10.20892/j.issn.2095-3941.2019.0097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Lord CJ, Ashworth A. The DNA damage response and cancer therapy. Nature. 2012;481:287–94. doi: 10.1038/nature10760. [DOI] [PubMed] [Google Scholar]
- 84.Liu LF, Desai SD, Li TK, Mao Y, Sun M, Sim SP. Mechanism of action of camptothecin. Ann N Y Acad Sci. 2000;922:1–10. doi: 10.1111/j.1749-6632.2000.tb07020.x. [DOI] [PubMed] [Google Scholar]
- 85.Leteurtre F, Fesen M, Kohlhagen G, Kohn KW, Pommier Y. Specific interaction of camptothecin, a topoisomerase I inhibitor, with guanine residues of DNA detected by photoactivation at 365 nm. Biochemistry. 1993;32:8955–62. doi: 10.1021/bi00085a029. [DOI] [PubMed] [Google Scholar]
- 86.Li F, Jiang T, Li Q, Ling X. Camptothecin (CPT) and its derivatives are known to target topoisomerase I (Top1) as their mechanism of action: did we miss something in CPT analogue molecular targets for treating human disease such as cancer? Am J Cancer Res. 2017;7:2350–94. [PMC free article] [PubMed] [Google Scholar]
- 87.Tsao YP, D’Arpa P, Liu LF. The involvement of active DNA synthesis in camptothecin-induced G2 arrest: altered regulation of p34cdc2/cyclin B. Cancer Res. 1992;52:1823–29. [PubMed] [Google Scholar]
- 88.Wang X, Tanaka M, Krstin S, Peixoto HS, Moura CCM, Wink M. Cytoskeletal interference - a new mode of action for the anticancer drugs camptothecin and topotecan. Eur J Pharmacol. 2016;789:265–74. doi: 10.1016/j.ejphar.2016.07.044. [DOI] [PubMed] [Google Scholar]
- 89.Ray Chaudhuri A, Hashimoto Y, Herrador R, Neelsen KJ, Fachinetti D, Bermejo R, et al. Topoisomerase I poisoning results in PARP-mediated replication fork reversal. Nat Struct Mol Biol. 2012;19:417–23. doi: 10.1038/nsmb.2258. [DOI] [PubMed] [Google Scholar]
- 90.Saitoh S, Ishii K, Kobayashi Y, Takahashi K. Spindle checkpoint signaling requires the mis6 kinetochore subcomplex, which interacts with mad2 and mitotic spindles. Mol Biol Cell. 2005;16:3666–77. doi: 10.1091/mbc.E05-01-0014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Dilshara MG, Jayasooriya R, Karunarathne W, Choi YH, Kim GY. Camptothecin induces mitotic arrest through Mad2-Cdc20 complex by activating the JNK-mediated Sp1 pathway. Food Chem Toxicol. 2019;127:143–55. doi: 10.1016/j.fct.2019.03.026. [DOI] [PubMed] [Google Scholar]
- 92.Liu YQ, Li WQ, Morris-Natschke SL, Qian K, Yang L, Zhu GX, et al. Perspectives on biologically active camptothecin derivatives. Med Res Rev. 2015;35:753–89. doi: 10.1002/med.21342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Creemers GJ, Lund B, Verweij J. Topoisomerase I inhibitors: topotecan and irenotecan. Cancer Treat Rev. 1994;20:73–96. doi: 10.1016/0305-7372(94)90011-6. [DOI] [PubMed] [Google Scholar]
- 94.de Man FM, Goey AKL, van Schaik RHN, Mathijssen RHJ, Bins S. Individualization of irinotecan treatment: a review of pharmacokinetics, pharmacodynamics, and pharmacogenetics. Clin Pharmacokinet. 2018;57:1229–54. doi: 10.1007/s40262-018-0644-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Saraf S, Jain A, Hurkat P, Jain SK. Topotecan liposomes: a visit from a molecular to a therapeutic platform. Crit Rev Ther Drug Carrier Syst. 2016;33:401–32. doi: 10.1615/CritRevTherDrugCarrierSyst.2016015926. [DOI] [PubMed] [Google Scholar]
- 96.Li Y, Luo W, Zhang H, Wang C, Yu C, Jiang Z, et al. Antitumor mechanism of hydroxycamptothecin via the metabolic perturbation of ribonucleotide and deoxyribonucleotide in human colorectal carcinoma cells. Molecules (Basel, Switzerland) 2021;26:4902. doi: 10.3390/molecules26164902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Ulukan H, Swaan PW. Camptothecins: a review of their chemotherapeutic potential. Drugs. 2002;62:2039–57. doi: 10.2165/00003495-200262140-00004. [DOI] [PubMed] [Google Scholar]
- 98.Clark JW. Rubitecan. Expert Opin Investig Drugs. 2006;15:71–9. doi: 10.1517/13543784.15.1.71. [DOI] [PubMed] [Google Scholar]
- 99.Selvendiran K, Singh JPV, Sakthisekaran D. In vivo effect of piperine on serum and tissue glycoprotein levels in benzo(a)pyrene induced lung carcinogenesis in Swiss albino mice. Pulm Pharmacol Ther. 2006;19:107–11. doi: 10.1016/j.pupt.2005.04.002. [DOI] [PubMed] [Google Scholar]
- 100.Elimam DM, Elgazar AA, Bonardi A, Abdelfadil M, Nocentini A, El-Domany RA, et al. Natural inspired piperine-based sulfonamides and carboxylic acids as carbonic anhydrase inhibitors: Design, synthesis and biological evaluation. Eur J Med Chem. 2021;225:113800. doi: 10.1016/j.ejmech.2021.113800. [DOI] [PubMed] [Google Scholar]
- 101.Lai LH, Fu QH, Liu Y, Jiang K, Guo QM, Chen QY, et al. Piperine suppresses tumor growth and metastasis in vitro and in vivo in a 4T1 murine breast cancer model. Acta Pharmacol Sin. 2012;33:523–30. doi: 10.1038/aps.2011.209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Guo L, Yang Y, Sheng Y, Wang J, Ruan S, Han C. Mechanism of piperine in affecting apoptosis and proliferation of gastric cancer cells via ROS-mitochondria-associated signalling pathway. J Cell Mol Med. 2021;25:9513–22. doi: 10.1111/jcmm.16891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Doucette CD, Hilchie AL, Liwski R, Hoskin DW. Piperine, a dietary phytochemical, inhibits angiogenesis. J Nutr Biochem. 2013;24:231–39. doi: 10.1016/j.jnutbio.2012.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Rather RA, Bhagat M. Cancer Chemoprevention and piperine: molecular mechanisms and therapeutic opportunities. Front Cell Dev Biol. 2018;6:10. doi: 10.3389/fcell.2018.00010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Vindelov LL. Flow microfluorometric analysis of nuclear DNA in cells from solid tumors and cell suspensions. A new method for rapid isolation and straining of nuclei. Virchows Arch B Cell Pathol. 1977;24:227–42. [PubMed] [Google Scholar]
- 106.Tawani A, Amanullah A, Mishra A, Kumar A. Evidences for Piperine inhibiting cancer by targeting human G-quadruplex DNA sequences. Sci Rep. 2016;6:39239. doi: 10.1038/srep39239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Fofaria NM, Kim S-H, Srivastava SK. Piperine causes G1 phase cell cycle arrest and apoptosis in melanoma cells through checkpoint kinase-1 activation. PLoS One. 2014;9:e94298. doi: 10.1371/journal.pone.0094298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Selvendiran K, Banu SM, Sakthisekaran D. Oral supplementation of piperine leads to altered phase II enzymes and reduced DNA damage and DNA-protein cross links in Benzo(a)pyrene induced experimental lung carcinogenesis. Mol Cell Biochem. 2005;268:141–7. doi: 10.1007/s11010-005-3702-z. [DOI] [PubMed] [Google Scholar]
- 109.Calcabrini C, Maffei F, Turrini E, Fimognari C. Sulforaphane potentiates anticancer effects of doxorubicin and cisplatin and mitigates their toxic effects. Front Pharmacol. 2020;11:567. doi: 10.3389/fphar.2020.00567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Rajendran P, Kidane AI, Yu TW, Dashwood WM, Bisson WH, Lohr CV, et al. HDAC turnover, CtIP acetylation and dysregulated DNA damage signaling in colon cancer cells treated with sulforaphane and related dietary isothiocyanates. Epigenetics. 2013;8:612–23. doi: 10.4161/epi.24710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Myzak MC, Karplus PA, Chung FL, Dashwood RH. A novel mechanism of chemoprotection by sulforaphane: inhibition of histone deacetylase. Cancer Res. 2004;64:5767–74. doi: 10.1158/0008-5472.CAN-04-1326. [DOI] [PubMed] [Google Scholar]
- 112.Eckschlager T, Plch J, Stiborova M, Hrabeta J. Histone deacetylase inhibitors as anticancer drugs. Int J Mol Sci. 2017;18:1414. doi: 10.3390/ijms18071414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Dos Santos P, Machado ART, De Grandis RA, Ribeiro DL, Tuttis K, Morselli M, et al. Transcriptome and DNA methylation changes modulated by sulforaphane induce cell cycle arrest, apoptosis, DNA damage, and suppression of proliferation in human liver cancer cells. Food Chem Toxicol. 2020;136:111047. doi: 10.1016/j.fct.2019.111047. [DOI] [PubMed] [Google Scholar]
- 114.Zheng K, Ma J, Wang Y, He Z, Deng K. Sulforaphane inhibits autophagy and induces exosome-mediated paracrine senescence via regulating mTOR/TFE3. Mol Nutr Food Res. 2020;64:e1901231. doi: 10.1002/mnfr.201901231. [DOI] [PubMed] [Google Scholar]
- 115.Xie H, Chun FKH, Rutz J, Blaheta RA. Sulforaphane impact on Reactive Oxygen Species (ROS) in Bladder Carcinoma. Int J Molr Sci. 2021;22:5938. doi: 10.3390/ijms22115938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Rauf A, Imran M, Butt MS, Nadeem M, Peters DG, Mubarak MS. Resveratrol as an anti-cancer agent: a review. Crit Rev Food Sci Nutr. 2018;58:1428–47. doi: 10.1080/10408398.2016.1263597. [DOI] [PubMed] [Google Scholar]
- 117.Singh AP, Singh R, Verma SS, Rai V, Kaschula CH, Maiti P, et al. Health benefits of resveratrol: evidence from clinical studies. Med Res Rev. 2019;39:1851–91. doi: 10.1002/med.21565. [DOI] [PubMed] [Google Scholar]
- 118.Tyagi A, Gu M, Takahata T, Frederick B, Agarwal C, Siriwardana S, et al. Resveratrol selectively induces DNA Damage, independent of Smad4 expression, in its efficacy against human head and neck squamous cell carcinoma. Clin Cancer Res. 2011;17:5402–11. doi: 10.1158/1078-0432.CCR-11-1072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Colin DJ, Limagne E, Ragot K, Lizard G, Ghiringhelli F, Solary É, et al. The role of reactive oxygen species and subsequent DNA-damage response in the emergence of resistance towards resveratrol in colon cancer models. Cell Death Dis. 2014;5:e1533. doi: 10.1038/cddis.2014.486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Yamaguchi M, Suzuki R, Oguchi M. Advances in the treatment of extranodal NK/T-cell lymphoma, nasal type. Blood. 2018;131:2528–40. doi: 10.1182/blood-2017-12-791418. [DOI] [PubMed] [Google Scholar]
- 121.Sui X, Zhang C, Zhou J, Cao S, Xu C, Tang F, et al. Resveratrol inhibits Extranodal NK/T cell lymphoma through activation of DNA damage response pathway. J Exp Clin Cancer Res. 2017;36:133. doi: 10.1186/s13046-017-0601-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Hande KR. Etoposide: four decades of development of a topoisomerase II inhibitor. Eur J Cancer (Oxford, England: 1990) 1998;34:1514–21. doi: 10.1016/s0959-8049(98)00228-7. [DOI] [PubMed] [Google Scholar]
- 123.Amiri F, Zarnani AH, Zand H, Koohdani F, Jeddi-Tehrani M, Vafa M. Synergistic anti-proliferative effect of resveratrol and etoposide on human hepatocellular and colon cancer cell lines. Eur J Pharmacol. 2013;718:34–40. doi: 10.1016/j.ejphar.2013.09.020. [DOI] [PubMed] [Google Scholar]
- 124.Montecucco A, Biamonti G. Cellular response to etoposide treatment. Cancer Lett. 2007;252:9–18. doi: 10.1016/j.canlet.2006.11.005. [DOI] [PubMed] [Google Scholar]
- 125.Otto T, Sicinski P. Cell cycle proteins as promising targets in cancer therapy. Nat Rev Cancer. 2017;17:93–115. doi: 10.1038/nrc.2016.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Damodaran AP, Vaufrey L, Gavard O, Prigent C. Aurora a kinase is a priority pharmaceutical target for the treatment of cancers. Trends Pharmacol Sci. 2017;38:687–700. doi: 10.1016/j.tips.2017.05.003. [DOI] [PubMed] [Google Scholar]
- 127.Dang F, Nie L, Wei W. Ubiquitin signaling in cell cycle control and tumorigenesis. Cell Death Differ. 2021;28:427–38. doi: 10.1038/s41418-020-00648-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Matthews HK, Bertoli C, de Bruin RAM. Cell cycle control in cancer. Nat Rev Mol Cell Biol. 2021;23:74–88. doi: 10.1038/s41580-021-00404-3. [DOI] [PubMed] [Google Scholar]
- 129.Liu J, Peng Y, Wei W. Cell cycle on the crossroad of tumorigenesis and cancer therapy. Trends Cell Biol. 2021;32:30–44. doi: 10.1016/j.tcb.2021.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Hwang HC, Clurman BE. Cyclin E in normal and neoplastic cell cycles. Oncogene. 2005;24:2776–86. doi: 10.1038/sj.onc.1208613. [DOI] [PubMed] [Google Scholar]
- 131.Moiseeva TN, Bakkenist CJ. Regulation of the initiation of DNA replication in human cells. DNA Repair (Amst) 2018;72:99–106. doi: 10.1016/j.dnarep.2018.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Bai J, Li Y, Zhang G. Cell cycle regulation and anticancer drug discovery. Cancer Biol Med. 2017;14:348–62. doi: 10.20892/j.issn.2095-3941.2017.0033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Alvarez-Fernandez M, Malumbres M. Mechanisms of sensitivity and resistance to CDK4/6 inhibition. Cancer Cell. 2020;37:514–29. doi: 10.1016/j.ccell.2020.03.010. [DOI] [PubMed] [Google Scholar]
- 134.Weinstein JN, Collisson EA, Mills GB, Shaw KRM, Ozenberger BA, Ellrott K, et al. The Cancer Genome Atlas Pan-Cancer analysis project. Nat Genet. 2013;45:1113–20. doi: 10.1038/ng.2764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Porter DC, Zhang N, Danes C, McGahren MJ, Harwell RM, Faruki S, et al. Tumor-specific proteolytic processing of cyclin E generates hyperactive lower-molecular-weight forms. Mol Cell Biol. 2001;21:6254–69. doi: 10.1128/MCB.21.18.6254-6269.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Suski JM, Braun M, Strmiska V, Sicinski P. Targeting cell-cycle machinery in cancer. Cancer Cell. 2021;39:759–78. doi: 10.1016/j.ccell.2021.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Zhu J, Huang R, Yang R, Xiao Y, Yan J, Zheng C, et al. Licorice extract inhibits growth of non-small cell lung cancer by down-regulating CDK4-Cyclin D1 complex and increasing CD8+ T cell infiltration. Cancer Cell Int. 2021;21:529. doi: 10.1186/s12935-021-02223-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Wang J, Zhang Y-S, Thakur K, Hussain SS, Zhang J-G, Xiao G-R, et al. Licochalcone A from licorice root, an inhibitor of human hepatoma cell growth via induction of cell apoptosis and cell cycle arrest. Food Chem Toxicol. 2018;120:407–17. doi: 10.1016/j.fct.2018.07.044. [DOI] [PubMed] [Google Scholar]
- 139.Chen HY, Cheng WP, Chiang YF, Hong YH, Ali M, Huang TC, et al. Hinokitiol exhibits antitumor properties through induction of ROS-mediated apoptosis and p53-driven cell-cycle arrest in endometrial cancer cell lines (Ishikawa, HEC-1A, KLE) Int J Mol Sci. 2021;22:8268. doi: 10.3390/ijms22158268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Zhang G, He J, Ye X, Zhu J, Hu X, Shen M, et al. β-Thujaplicin induces autophagic cell death, apoptosis, and cell cycle arrest through ROS-mediated Akt and p38/ERK MAPK signaling in human hepatocellular carcinoma. Cell Death Dis. 2019;10:255. doi: 10.1038/s41419-019-1492-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Kim SA, Kang OH, Kwon DY. Cryptotanshinone induces cell cycle arrest and apoptosis of NSCLC cells through the PI3K/Akt/GSK-3β pathway. Int J Mol Sci. 2018;19:2739. doi: 10.3390/ijms19092739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Spagnuolo C, Russo GL, Orhan IE, Habtemariam S, Daglia M, Sureda A, et al. Genistein and cancer: current status, challenges, and future directions. Adv Nutr. 2015;6:408–19. doi: 10.3945/an.114.008052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Park C, Cha H-J, Lee H, Hwang-Bo H, Ji SY, Kim MY, et al. Induction of G2/M cell cycle arrest and apoptosis by genistein in human bladder cancer T24 cells through inhibition of the ROS-dependent PI3k/Akt signal transduction pathway. Antioxidants (Basel) 2019;8:327. doi: 10.3390/antiox8090327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Di X, Andrews DM, Tucker CJ, Yu L, Moore AB, Zheng X, et al. A high concentration of genistein down-regulates activin A, Smad3 and other TGF-β pathway genes in human uterine leiomyoma cells. Exp Mol Med. 2012;44:281–92. doi: 10.3858/emm.2012.44.4.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Park JH, Oh EJ, Choi YH, Kang CD, Kang HS, Kim DK, et al. Synergistic effects of dexamethasone and genistein on the expression of Cdk inhibitor p21WAF1/CIP1 in human hepatocellular and colorectal carcinoma cells. Int J Oncol. 2001;18:997–1002. doi: 10.3892/ijo.18.5.997. [DOI] [PubMed] [Google Scholar]
- 146.Hwang KA, Kang NH, Yi BR, Lee HR, Park MA, Choi KC. Genistein, a soy phytoestrogen, prevents the growth of BG-1 ovarian cancer cells induced by 17β-estradiol or bisphenol A via the inhibition of cell cycle progression. Int J Oncol. 2013;42:733–40. doi: 10.3892/ijo.2012.1719. [DOI] [PubMed] [Google Scholar]
- 147.Wang K, Feng X, Chai L, Cao S, Qiu F. The metabolism of berberine and its contribution to the pharmacological effects. Drug Metab Rev. 2017;49:139–57. doi: 10.1080/03602532.2017.1306544. [DOI] [PubMed] [Google Scholar]
- 148.Mantena SK, Sharma SD, Katiyar SK. Berberine, a natural product, induces G1-phase cell cycle arrest and caspase-3-dependent apoptosis in human prostate carcinoma cells. Mol Cancer Ther. 2006;5:296–308. doi: 10.1158/1535-7163.MCT-05-0448. [DOI] [PubMed] [Google Scholar]
- 149.Zhang RR, Tian HY, Tan YF, Chung TY, Sun XH, Xia X, et al. Structures, chemotaxonomic significance, cytotoxic and Na(+),<softenter>K(+)-ATPase inhibitory activities of new cardenolides from Asclepias curassavica. Org Biomol Chem. 2014;12:8919–29. doi: 10.1039/c4ob01545b. [DOI] [PubMed] [Google Scholar]
- 150.Leu WJ, Wang CT, Hsu JL, Chen IS, Chang HS, Guh JH. Ascleposide, a natural cardenolide, induces anticancer signaling in human castration-resistant prostatic cancer through Na(+) /K(+) -ATPase internalization and tubulin acetylation. Prostate. 2020;80:305–18. doi: 10.1002/pros.23944. [DOI] [PubMed] [Google Scholar]
- 151.Bretones G, Delgado MD, Leon J. Myc and cell cycle control. Biochim Biophys Acta. 2015;1849:506–16. doi: 10.1016/j.bbagrm.2014.03.013. [DOI] [PubMed] [Google Scholar]
- 152.Asghar U, Witkiewicz AK, Turner NC, Knudsen ES. The history and future of targeting cyclin-dependent kinases in cancer therapy. Nat Rev Drug Discov. 2015;14:130–46. doi: 10.1038/nrd4504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Suzuki A, Kawano H, Hayashida M, Hayasaki Y, Tsutomi Y, Akahane K. Procaspase 3/p21 complex formation to resist fas-mediated cell death is initiated as a result of the phosphorylation of p21 by protein kinase A. Cell Death Differ. 2000;7:721–8. doi: 10.1038/sj.cdd.4400706. [DOI] [PubMed] [Google Scholar]
- 154.Eymin B, Haugg M, Droin N, Sordet O, Dimanche-Boitrel MT, Solary E. p27Kip1 induces drug resistance by preventing apoptosis upstream of cytochrome c release and procaspase-3 activation in leukemic cells. Oncogene. 1999;18:1411–18. doi: 10.1038/sj.onc.1202437. [DOI] [PubMed] [Google Scholar]
- 155.Yu SY, Liao CH, Chien MH, Tsai TY, Lin JK, Weng MS. Induction of p21(Waf1/Cip1) by garcinol via downregulation of p38-MAPK signaling in p53-independent H1299 lung cancer. J Agric Food Chem. 2014;62:2085–95. doi: 10.1021/jf4037722. [DOI] [PubMed] [Google Scholar]
- 156.Shahrestanaki MK, Bagheri M, Ghanadian M, Aghaei M, Jafari SM. Centaurea cyanus extracted 13-O-acetylsolstitialin A decrease Bax/Bcl-2 ratio and expression of cyclin D1/Cdk-4 to induce apoptosis and cell cycle arrest in MCF-7 and MDA-MB-231 breast cancer cell lines. J Cell Biochem. 2019;120:18309–319. doi: 10.1002/jcb.29141. [DOI] [PubMed] [Google Scholar]
- 157.Li J, Wang Q, Wang Z, Cui N, Yang B, Niu W, et al. Tetrandrine inhibits colon carcinoma HT-29 cells growth via the Bcl-2/Caspase 3/PARP pathway and G1/S phase. Biosci Rep. 2019;39:BSR20182109. doi: 10.1042/BSR20182109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Li L, Thakur K, Cao YY, Liao BY, Zhang JG, Wei ZJ. Anticancerous potential of polysaccharides sequentially extracted from Polygonatum cyrtonema Hua in Human cervical cancer Hela cells. Int J Biol Macromol. 2020;148:843–50. doi: 10.1016/j.ijbiomac.2020.01.223. [DOI] [PubMed] [Google Scholar]
- 159.Gao LL, Li FR, Jiao P, Yang MF, Zhou XJ, Si YH, et al. Paris chinensis dioscin induces G2/M cell cycle arrest and apoptosis in human gastric cancer SGC-7901 cells. World J Gastroenterol. 2011;17:4389–95. doi: 10.3748/wjg.v17.i39.4389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Li CY, Wang Y, Wang HL, Shi Z, An N, Liu YX, et al. Molecular mechanisms of Lycoris aurea agglutinin-induced apoptosis and G2 /M cell cycle arrest in human lung adenocarcinoma A549 cells, both in vitro and in vivo. Cell Prolif. 2013;46:272–82. doi: 10.1111/cpr.12034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Lee SM, Kwon JI, Choi YH, Eom HS, Chi GY. Induction of G2/M arrest and apoptosis by water extract of Strychni Semen in human gastric carcinoma AGS cells. Phytother Res. 2008;22:752–8. doi: 10.1002/ptr.2355. [DOI] [PubMed] [Google Scholar]
- 162.Qi LW, Zhang Z, Zhang CF, Anderson S, Liu Q, Yuan CS, et al. Anti-Colon Cancer Effects of 6-Shogaol Through G2/M Cell Cycle Arrest by p53/p21-cdc2/cdc25A Crosstalk. Am J Chin Med. 2015;43:743–56. doi: 10.1142/S0192415X15500469. [DOI] [PubMed] [Google Scholar]
- 163.Yao Y, Zhang YW, Sun LG, Liu B, Bao YL, Lin H, et al. Juglanthraquinone C, a novel natural compound derived from Juglans mandshurica Maxim, induces S phase arrest and apoptosis in HepG2 cells. Apoptosis. 2012;17:832–41. doi: 10.1007/s10495-012-0722-5. [DOI] [PubMed] [Google Scholar]
- 164.Wang J, Pan Y, Hu J, Ma Q, Xu Y, Zhang Y, et al. Tea polyphenols induce S phase arrest and apoptosis in gallbladder cancer cells. Braz J Med Biol Res. 2018;51:e6891. doi: 10.1590/1414-431X20176891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Ibrahim SR, Mohamed GA. Litchi chinensis: medicinal uses, phytochemistry, and pharmacology. J Ethnopharmacol. 2015;174:492–513. doi: 10.1016/j.jep.2015.08.054. [DOI] [PubMed] [Google Scholar]
- 166.Hsu CP, Lin CC, Huang CC, Lin YH, Chou JC, Tsia YT, et al. Induction of apoptosis and cell cycle arrest in human colorectal carcinoma by Litchi seed extract. J Biomed Biotechnol. 2012;2012:341479. doi: 10.1155/2012/341479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Guo H, Luo H, Yuan H, Xia Y, Shu P, Huang X, et al. Litchi seed extracts diminish prostate cancer progression via induction of apoptosis and attenuation of EMT through Akt/GSK-3β signaling. Sci Rep. 2017;7:41656. doi: 10.1038/srep41656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Ma J, Hu X, Liao C, Xiao H, Zhu Q, Li Y, et al. Gypenoside L inhibits proliferation of liver and esophageal cancer cells by inducing senescence. Molecules. 2019;24:1054. doi: 10.3390/molecules24061054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Swanton C. Cell-cycle targeted therapies. Lancet Oncol. 2004;5:27–36. doi: 10.1016/s1470-2045(03)01321-4. [DOI] [PubMed] [Google Scholar]
- 170.Zhai S, Senderowicz AM, Sausville EA, Figg WD. Flavopiridol, a novel cyclin-dependent kinase inhibitor, in clinical development. Ann Pharmacother. 2002;36:905–11. doi: 10.1345/aph.1A162. [DOI] [PubMed] [Google Scholar]
- 171.Blagosklonny MV. Flavopiridol, an inhibitor of transcription: implications, problems and solutions. Cell cycle (Georgetown, Tex) 2004;3:1537–42. doi: 10.4161/cc.3.12.1278. [DOI] [PubMed] [Google Scholar]
- 172.Zhao B, Bower MJ, McDevitt PJ, Zhao H, Davis ST, Johanson KO, et al. Structural basis for Chk1 inhibition by UCN-01. J Biol Chem. 2002;277:46609–15. doi: 10.1074/jbc.M201233200. [DOI] [PubMed] [Google Scholar]
- 173.Kim ES. Midostaurin: first global approval. Drugs. 2017;77:1251–59. doi: 10.1007/s40265-017-0779-0. [DOI] [PubMed] [Google Scholar]
- 174.MacCallum DE, Melville J, Frame S, Watt K, Anderson S, Gianella-Borradori A, et al. Seliciclib (CYC202, R-Roscovitine) induces cell death in multiple myeloma cells by inhibition of RNA polymerase II-dependent transcription and down-regulation of Mcl-1. Cancer Res. 2005;65:5399–407. doi: 10.1158/0008-5472.CAN-05-0233. [DOI] [PubMed] [Google Scholar]
- 175.Khalil HS, Mitev V, Vlaykova T, Cavicchi L, Zhelev N. Discovery and development of Seliciclib. how systems biology approaches can lead to better drug performance. J Biotechnol. 2015;202:40–9. doi: 10.1016/j.jbiotec.2015.02.032. [DOI] [PubMed] [Google Scholar]
- 176.Stark GR, Taylor WR. Control of the G2/M transition. Mol Biotechnol. 2006;32:227–48. doi: 10.1385/MB:32:3:227. [DOI] [PubMed] [Google Scholar]
- 177.Ke CS, Liu HS, Yen CH, Huang GC, Cheng HC, Huang CY, et al. Curcumin-induced Aurora-A suppression not only causes mitotic defect and cell cycle arrest but also alters chemosensitivity to anticancer drugs. J Nutr Biochem. 2014;25:526–39. doi: 10.1016/j.jnutbio.2014.01.003. [DOI] [PubMed] [Google Scholar]
- 178.Liu HS, Ke CS, Cheng HC, Huang CY, Su CL. Curcumin-induced mitotic spindle defect and cell cycle arrest in human bladder cancer cells occurs partly through inhibition of aurora A. Mol Pharmacol. 2011;80:638–46. doi: 10.1124/mol.111.072512. [DOI] [PubMed] [Google Scholar]
- 179.Schwarz K, Dobiasch S, Nguyen L, Schilling D, Combs SE. Modification of radiosensitivity by Curcumin in human pancreatic cancer cell lines. Sci Rep. 2020;10:3815. doi: 10.1038/s41598-020-60765-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Mosieniak G, Sliwinska MA, Przybylska D, Grabowska W, Sunderland P, Bielak-Zmijewska A, et al. Curcumin-treated cancer cells show mitotic disturbances leading to growth arrest and induction of senescence phenotype. Int J Biochem Cell Biol. 2016;74:33–43. doi: 10.1016/j.biocel.2016.02.014. [DOI] [PubMed] [Google Scholar]
- 181.Li Y, Gong Y, Li L, Abdolmaleky HM, Zhou J-R. Bioactive tanshinone I inhibits the growth of lung cancer partly via downregulation of Aurora A function. Mol Carcinog. 2013;52:535–43. doi: 10.1002/mc.21888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Gong Y, Li Y, Lu Y, Li L, Abdolmaleky H, Blackburn GL, et al. Bioactive tanshinones in Salvia miltiorrhiza inhibit the growth of prostate cancer cells in vitro and in mice. Int J Cancer. 2011;129:1042–52. doi: 10.1002/ijc.25678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Gong Y, Li Y, Abdolmaleky HM, Li L, Zhou J-R. Tanshinones inhibit the growth of breast cancer cells through epigenetic modification of Aurora A expression and function. PLoS One. 2012;7:e33656. doi: 10.1371/journal.pone.0033656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Anand P, Kunnumakkara AB, Newman RA, Aggarwal BB. Bioavailability of curcumin: problems and promises. Mol Pharm. 2007;4:807–18. doi: 10.1021/mp700113r. [DOI] [PubMed] [Google Scholar]
- 185.Wang L, Wang X, Chen H, Zu X, Ma F, Liu K, et al. Gossypin inhibits gastric cancer growth by direct targeting of AURKA and RSK2. Phytother Res. 2019;33:640–50. doi: 10.1002/ptr.6253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Fan Y, Lu H, Ma H, Feng F, Hu X, Zhang Q, et al. Bioactive compounds of Eriocaulon sieboldianum blocking proliferation and inducing apoptosis of HepG2 cells might be involved in Aurora kinase inhibition. Food Funct. 2015;6:3746–59. doi: 10.1039/c5fo00371g. [DOI] [PubMed] [Google Scholar]
- 187.Nguyen TN, Feizbakhsh O, Sfecci E, Baratte B, Delehouze C, Garcia A, et al. Kinase-based screening of marine natural extracts leads to the identification of a cytotoxic high molecular weight metabolite from the mediterranean sponge Crambe tailliezi. Mar Drugs. 2019;17:569. doi: 10.3390/md17100569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Liu Q, Jiao Z, Liu Y, Li Z, Shi X, Wang W, et al. Chemical profiling of San-Huang decoction by UPLC-ESI-Q-TOF-MS. J Pharm Biomed Anal. 2016;131:20–32. doi: 10.1016/j.jpba.2016.07.036. [DOI] [PubMed] [Google Scholar]
- 189.Xu Y, Chen X, Chen X, Bian W, Yao C, Zhang X, et al. San Huang Decoction downregulates Aurora kinase A to inhibit breast cancer cell growth and enhance chemosenstivity to anti-tumor drugs. Pathol Res Pract. 2016;212:696–703. doi: 10.1016/j.prp.2016.05.004. [DOI] [PubMed] [Google Scholar]
- 190.Hnit SST, Yao M, Xie C, Ge G, Bi L, Jin S, et al. Transcriptional regulation of G2/M regulatory proteins and perturbation of G2/M Cell cycle transition by a traditional Chinese medicine recipe. J Ethnopharmacol. 2020;251:112526. doi: 10.1016/j.jep.2019.112526. [DOI] [PubMed] [Google Scholar]
- 191.Shakeel I, Basheer N, Hasan GM, Afzal M, Hassan MI. Polo-like Kinase 1 as an emerging drug target: structure, function and therapeutic implications. J Drug Target. 2021;29:168–84. doi: 10.1080/1061186X.2020.1818760. [DOI] [PubMed] [Google Scholar]
- 192.Gheghiani L, Wang L, Zhang Y, Moore XTR, Zhang J, Smith SC, et al. PLK1 induces chromosomal instability and overrides cell-cycle checkindicates drive tumorigenesis. Cancer Res. 2021;81:1293–307. doi: 10.1158/0008-5472.CAN-20-1377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Liang H, Liu H, Kuang Y, Chen L, Ye M, Lai L. Discovery of targeted covalent natural products against PLK1 by herb-based screening. J Chem Inf Model. 2020;60:4350–58. doi: 10.1021/acs.jcim.0c00074. [DOI] [PubMed] [Google Scholar]
- 194.Xu G, Yan X, Hu Z, Zheng L, Ding K, Zhang Y, et al. Glucocappasalin induces G2/M-Phase arrest, apoptosis, and autophagy pathways by targeting CDK1 and PLK1 in cervical carcinoma cells. Front Pharmacol. 2021;12:671138. doi: 10.3389/fphar.2021.671138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Qazzaz ME, Raja VJ, Lim KH, Kam TS, Lee JB, Gershkovich P, et al. In vitro anticancer properties and biological evaluation of novel natural alkaloid jerantinine B. Cancer Lett. 2016;370:185–97. doi: 10.1016/j.canlet.2015.10.013. [DOI] [PubMed] [Google Scholar]
- 196.Ren X, Liu J, Hu L, Liu Q, Wang D, Ning X. Caffeic acid phenethyl ester inhibits the proliferation of HEp2 cells by regulating Stat3/Plk1 pathway and inducing S phase arrest. Biol Pharm Bull. 2019;42:1689–93. doi: 10.1248/bpb.b19-00315. [DOI] [PubMed] [Google Scholar]
- 197.Shan HM, Shi Y, Quan J. Identification of green tea catechins as potent inhibitors of the polo-box domain of polo-like kinase 1. ChemMedChem. 2015;10:158–63. doi: 10.1002/cmdc.201402284. [DOI] [PubMed] [Google Scholar]
- 198.Yu L, Lang Y, Hsu CC, Chen WM, Chiang JC, Hsieh JT, et al. Mitotic phosphorylation of tumor suppressor DAB2IP maintains spindle assembly checkpoint and chromosomal stability through activating PLK1-Mps1 signal pathway and stabilizing mitotic checkpoint complex. Oncogene. 2022;41:489–501. doi: 10.1038/s41388-021-02106-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Xie Y, Wang A, Lin J, Wu L, Zhang H, Yang X, et al. Mps1/TTK: a novel target and biomarker for cancer. J Drug Target. 2017;25:112–8. doi: 10.1080/1061186X.2016.1258568. [DOI] [PubMed] [Google Scholar]
- 200.Zhang X, Ling Y, Guo Y, Bai Y, Shi X, Gong F, et al. Mps1 kinase regulates tumor cell viability via its novel role in mitochondria. Cell Death Dis. 2016;7:e2292. doi: 10.1038/cddis.2016.193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Herbert KJ, Ashton TM, Prevo R, Pirovano G, Higgins GS. T-LAK cell-originated protein kinase (TOPK): an emerging target for cancer-specific therapeutics. Cell Death Dis. 2018;9:1089. doi: 10.1038/s41419-018-1131-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Kamath K, Wilson L, Cabral F, Jordan MA. BetaIII-tubulin induces paclitaxel resistance in association with reduced effects on microtubule dynamic instability. J Biol Chem. 2005;280:12902–907. doi: 10.1074/jbc.M414477200. [DOI] [PubMed] [Google Scholar]
- 203.Sirotnak FM, Danenberg KD, Chen J, Fritz F, Danenberg PV. Markedly decreased binding of vincristine to tubulin in vinca alkaloid-resistant Chinese hamster cells is associated with selective overexpression of alpha and beta tubulin isoforms. Biochem Biophys Res Commun. 2000;269:21–4. doi: 10.1006/bbrc.2000.2246. [DOI] [PubMed] [Google Scholar]
- 204.Beretta GL, Gatti L, Perego P, Zaffaroni N. Camptothecin resistance in cancer: insights into the molecular mechanisms of a DNA-damaging drug. Curr Med Chem. 2013;20:1541–65. doi: 10.2174/0929867311320120006. [DOI] [PubMed] [Google Scholar]
- 205.Ghawanmeh AA, Al-Bajalan HM, Mackeen MM, Alali FQ, Chong KF. Recent developments on (-)-colchicine derivatives: synthesis and structure-activity relationship. Eur J Med Chem. 2020;185:111788. doi: 10.1016/j.ejmech.2019.111788. [DOI] [PubMed] [Google Scholar]
- 206.Abdifetah O, Na-Bangchang K. Pharmacokinetic studies of nanoparticles as a delivery system for conventional drugs and herb-derived compounds for cancer therapy: a systematic review. Int J Nanomedicine. 2019;14:5659–77. doi: 10.2147/IJN.S213229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Bernabeu E, Helguera G, Legaspi MJ, Gonzalez L, Hocht C, Taira C, et al. Paclitaxel-loaded PCL-TPGS nanoparticles: in vitro and in vivo performance compared with Abraxane®. Colloids Surf B Biointerfaces. 2014;113:43–50. doi: 10.1016/j.colsurfb.2013.07.036. [DOI] [PubMed] [Google Scholar]
- 208.Shukla M, Jaiswal S, Sharma A, Srivastava PK, Arya A, Dwivedi AK, et al. A combination of complexation and self-nanoemulsifying drug delivery system for enhancing oral bioavailability and anticancer efficacy of curcumin. Drug Dev Ind Pharm. 2017;43:847–61. doi: 10.1080/03639045.2016.1239732. [DOI] [PubMed] [Google Scholar]
- 209.Lollo G, Ullio-Gamboa G, Fuentes E, Matha K, Lautram N, Benoit JP. In vitro anti-cancer activity and pharmacokinetic evaluation of curcumin-loaded lipid nanocapsules. Mater Sci Eng C Mater Biol Appl. 2018;91:859–67. doi: 10.1016/j.msec.2018.06.014. [DOI] [PubMed] [Google Scholar]
- 210.Ramaswamy S, Dwarampudi LP, Kadiyala M, Kuppuswamy G, Reddy KVVS, Kumar CKA, et al. Formulation and characterization of chitosan encapsulated phytoconstituents of curcumin and rutin nanoparticles. Int J Biol Macromol. 2017;104(Pt B):1807–12. doi: 10.1016/j.ijbiomac.2017.06.112. [DOI] [PubMed] [Google Scholar]
- 211.Ruiz-Moreno AJ, Dömling A, Velasco-Velázquez MA. Reverse docking for the identification of molecular targets of anticancer compounds. Methods Mol Biol. 2021;2174:31–43. doi: 10.1007/978-1-0716-0759-6_4. [DOI] [PubMed] [Google Scholar]
- 212.Toplak Z, Hendrickx LA, Gubic S, Mozina S, Zegura B, Stern A, et al. 3D pharmacophore-based discovery of novel KV10.1 inhibitors with antiproliferative activity. Cancers (Basel) 2021;13:1244. doi: 10.3390/cancers13061244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Wei X, Wu X, Cheng Z, Wu Q, Cao C, Xu X, et al. Botanical drugs: a new strategy for structure-based target prediction. Brief Bioinform. 2022;23:bbab425. doi: 10.1093/bib/bbab425. [DOI] [PubMed] [Google Scholar]