

Microbial cell surface hydrophobicity (15, 19) is probably the most studied microbial cell surface characteristic measured due to its ubiquitously accepted role in microbial adhesion to surfaces (1, 20, 23). Yet, the electrostatic charge properties of microbial cell surfaces also play a role in microbial adhesion (17, 21). Although by far most microbial cell surfaces are negatively charged (4, 14), positively charged organisms that adhere tenaciously to negatively charged surfaces have been identified (2, 14); furthermore, positively charged domains that stimulate attraction to negatively charged substrata have been suggested to exist on microbial cell surfaces (1).
However, the electrophoretic mobilities of individual organisms within single-strain microbial populations need not necessarily be the same. Usually, electrophoretic mobilities are reported as one mean value (μmean), being at best supplemented with a standard deviation from different mean values obtained with separately cultured strains (SDcultures) but the standard deviations from values for individual microorganisms within a pure culture may be more relevant (10). This is illustrated in Fig. 1, which shows the results of three electrophoretic mobility measurements of separately cultured Escherichia coli Hu734 cells. Each measurement of the mean electrophoretic mobility (μi) comprises values for at least 120 individual bacteria and represents a Gaussian distribution with its own population standard deviation (SDpopulation) from values for individual bacteria in the culture. SDpopulations, although probably more relevant than the standard deviation from values for cultures, are seldom reported in the literature. Sometimes, even subpopulations exist within a single-strain culture possessing distinctly different electrophoretic mobilities, as illustrated in the example given in Fig. 2. Due to inadequate experimental methods, such heterogeneities in electrophoretic mobilities of microbial populations have been largely neglected in the literature.
FIG. 1.

Electrophoretic mobility distributions of E. coli Hu734 organisms suspended in 10 mM potassium phosphate (pH 6.3). The three distributions (μ1, μ2, and μ3) were measured on separately cultured bacterial strains, and each has its own SDpopulation. Typically, the μmean of μ1, μ2, and μ3 is presented in the literature as an electrophoretic mobility of −2.1 (±0.7) × 10−8 m2 V−1 s−1, with a standard deviation, SDcultures, from the three measurements.
FIG. 2.

Bimodal electrophoretic mobility distribution of Pseudomonas aeruginosa #3, suspended in phosphate-buffered saline (pH 7.0).
Since such distributions in electrophoretic mobilities have a major impact on microbial interactions with surfaces, we here briefly describe how electrophoretic mobility distributions can be measured using microelectrophoresis, review three published applications, and discuss the impact of potential population heterogeneity for microbial adhesion studies.
MICROELECTROPHORESIS AND THE MEASUREMENT OF ELECTROPHORETIC MOBILITY DISTRIBUTIONS
In microelectrophoresis, microorganisms are suspended in a liquid phase. A flow chamber, which can be rectangular or cylindrical, is subsequently filled with this suspension, and a voltage between 75 and 150 V is applied over the flow chamber (13). Negatively charged microorganisms are then attracted to the positive electrode, and positively charged organisms are attracted to the negative electrode. The velocity at which an organism travels is a direct measure of its electrophoretic mobility. In so-called first-generation instruments, the velocities of individual organisms were clocked manually, enabling accurate measurement of electrophoretic mobility distributions at the expense of extremely time-consuming experiments (18). Second-generation instruments, like the well-known PenKem Lazer Zee Meter 501, allow us to measure only the μmean of a given population when it is operated in its standard mode. In its standard mode, moving organisms are viewed through a rotating mirror, and by matching the rotational speed of the mirror, the average velocity of the suspended microorganisms is counterbalanced, after which the rotational speed of the mirror can be used to calculate the population electrophoretic mobility, but the SDpopulation cannot be obtained.
Recently, Glynn et al. (10) erroneously disqualified this generation of instruments to measure more biologically relevant electrophoretic mobility distributions with an SDpopulation. By arresting the mirror and using computer-aided image analysis, a variety of methods exists (18, 24) through which second-generation instruments can be upgraded to extremely fast third-generation instruments, enabling the measurement of electrophoretic mobility distributions. In our experience, as a result of the direct observation, upgraded second-generation instruments provide the most reliable instruments to this end. Some applications, showing the advantages and additional possibilities of measuring electrophoretic mobility distributions rather than mean population values, will now be discussed.
ELECTROPHORETIC MOBILITY DISTRIBUTIONS OF SINGLE-STRAIN BACTERIAL CULTURES
Electrophoretic heterogeneity of lactobacilli after serial passaging.
A Lactobacillus acidophilus RC14 pure culture was serially passaged up to 50 times in liquid growth medium (9) and examined by electron microscopy after ruthenium red-uranyl acetate staining and by microelectrophoresis. In 10 mM potassium phosphate solution, pH 5.0, the primary isolate had a single, nearly zero electrophoretic mobility of 0.20 × 10−8 m2 V−1 s−1. In cultures with P values of both 20 and 50, a more negatively charged subpopulation with an electrophoretic mobility of −1.33 × 10−8 m2 V−1 s−1 existed next to the virtually uncharged population. Electron microscopy demonstrated that, in cultures of the primary isolate, all organisms had a thick stainable layer as their outermost cell surface but that, in serially passaged cultures, 31 to 42% of all organisms were devoid of such a stainable layer (Fig. 3). This result corresponded numerically with the electrophoretic mobility distributions measured, and the fraction of bald organisms could be identified with the more negatively charged subpopulation (9).
FIG. 3.
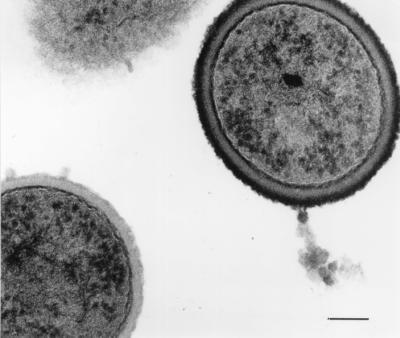
Electron micrograph of ruthenium red-uranyl acetate-stained (12) L. acidophilus RC14 cells after being subcultured 20 times. Individual organisms with a thick stainable layer as their outermost cell surface can be observed next to organisms devoid of a stainable layer (micrograph reprinted from the Journal of Microbiological Methods [9] with permission from Elsevier Science). The bar denotes 0.15 μm.
Electrophoretic mobility distributions of Treponema denticola ATCC 33520 and hemagglutination.
Exponential- and stationary-phase pure cultures of T. denticola ATCC 33520 displayed two distinct pH-dependent electrophoretic mobilities (6). Adsorption of a culture of T. denticola with an excess of erythrocytes removed a large portion of the more negatively charged subpopulation, which was the minority population, suggesting that this portion of the spirochete population was actually adhering to the erythrocytes through attractive electrostatic interactions (6).
Heterogeneous electrophoretic mobilities of subgingival bacteria.
Eleven out of 12 fresh clinical isolates of gram-negative Porphyromonas gingivalis, Prevotella intermedia, and Actinobacillus actinomycetemcomitans and of gram-positive Peptostreptococcus micros displayed heterogeneous populations with respect to pH-dependent electrophoretic mobilities (5). For the gram-negative strains, the more negatively charged subpopulation was in the majority, while the Peptostreptococcus micros strains appeared to be composed mainly of a less negatively charged subpopulation. Vesicles of P. gingivalis and Prevotella intermedia displayed the same heterogeneity, while an A. actinomycetemcomitans strain passaged several times in fluid medium lost its fimbriae and became homogeneous with respect to charge. It was hypothesized (5) that the ability of single-strain bacterial populations to express multiple electrophoretic mobilities could be useful to the organism in its adherence in a complex environment, like a subgingival pocket, where there are oral hard and soft tissues and other microbial cell surfaces present (7).
In conclusion, the display of multiple electrophoretic mobilities in single-strain microbial populations can be excellently studied using microelectrophoresis. Although the occurrence of multiple electrophoretic mobilities in single-strain microbial populations has been largely neglected in current literature, it probably occurs in isolates obtained from environments as diverse as the urogenital tract and the oral cavity and it has an established role in the adhesion and survival of the strains displaying this trait. The impact of population heterogeneity occurring in single-strain microbial cultures may be far-reaching, particularly in microbial adhesion to surfaces, as demonstrated above, and it may relate to poorly understood aspects of, e.g., microbial adhesion to hydrocarbons (MATH) (15), as will be explained below.
POPULATION HETEROGENEITY AND ADHESION TO HYDROCARBONS
Hydrophobicity is by definition the degree of dislike of a surface for water (3, 22), but since the pioneering work of Rosenberg et al. (19), it is invariably associated with the ability of microorganisms to adhere to hydrocarbons in an aqueous suspension. Adhesion, however, whether to hydrocarbons in aqueous suspension or to any other surface, is always an interplay of all factors involved in the process (17), including not only hydrophobicity but also electrostatic charge interactions (21), and influences of structural cell surface properties (11). MATH analysis can be carried out as originally proposed by Rosenberg et al. (19), but Rosenberg himself criticized the assay as not being sufficiently quantitative (15). In the kinetic mode of MATH analysis, a microbial suspension is vortexed for different periods of time with a hydrocarbon phase and the optical density of the aqueous phase is measured as a function of the vortexing time (Fig. 4). This process is opposed to that of the original MATH assay, in which only one point in time is taken and the optical density after a single vortexing round is taken as a measure of the cell surface hydrophobicity. In the kinetic mode, linear-regression analysis is carried out to derive an initial microbial removal rate by the hydrocarbon as a measure of hydrophobicity. Poorly understood aspects of the kinetic MATH assay are the observations that, for some strains, all organisms in the aqueous suspension finally adhere to the hydrocarbon phase after prolonged vortexing, like for Streptococcus salivarius HB in Fig. 4, but that, for other strains, a fraction of all suspended organisms remain in suspension, regardless of the duration of vortexing (see data for Streptococcus oralis J22 in Fig. 4).
FIG. 4.

Optical densities as a function of vortexing time for S. salivarius HB (●) and S. oralis J22 (▴) in MATH analysis. Hexadecane was used as the hydrocarbon phase, while bacteria were suspended in 10 mM potassium phosphate, pH 5.0. Note that for S. salivarius HB, the optical density is reduced essentially to zero but that for S. oralis J22, a sizeable optical density remains, also after prolonged vortexing.
Medrzycka (16) has demonstrated that hydrocarbon droplets in aqueous suspensions are negatively charged. It has been indicated that all different types of hydrocarbons employed in MATH are negatively charged in the common buffer systems used for MATH (3). Thus, electrophoretic mobility is a factor in MATH as well (8, 21, 22). Mechanistically, it has been demonstrated that, above the isoelectric point of a microbial strain, adhesion to hydrocarbons occurs despite electrostatic repulsion, provided that the organisms have sufficiently high intrinsic hydrophobicity, but that, below their isoelectric point, organisms experience a minor electrostatic attraction to hydrocarbon droplets in aqueous suspensions (21). Consequently, it is suggested that the fraction of organisms in a suspension unable to adhere to the hydrocarbon droplets even after prolonged vortexing represents a more negatively charged subpopulation in the culture.
CONCLUSIONS
Microbial cultures should preferentially be examined for potential population heterogeneity prior to any adhesion study to prevent erroneous conclusions being drawn due to the existence of subpopulations with different surface properties in single-strain cultures. The measurement of electrophoretic mobility distributions seems ideal to this end.
REFERENCES
- 1.Bos R, van der Mei H C, Busscher H J. Physico-chemistry of initial microbial adhesive interactions—its mechanisms and methods for study. FEMS Microbiol Rev. 1999;23:179–229. doi: 10.1111/j.1574-6976.1999.tb00396.x. [DOI] [PubMed] [Google Scholar]
- 2.Busscher H J, Bellon-Fontaine M N, Mozes N, van der Mei H C, Sjollema J, Cerf O, Rouxhet P G. Deposition of Leuconostoc mesenteroides and Streptococcus thermophilus to solid substrata in a parallel plate flow cell. Biofouling. 1990;2:55–63. [Google Scholar]
- 3.Busscher H J, van de Belt-Gritter B, van der Mei H C. Implications of microbial adhesion to hydrocarbons for evaluating cell surface hydrophobicity. 1. Zeta potentials of hydrocarbon droplets. Colloids Surf B. 1995;5:111–116. [Google Scholar]
- 4.Busscher H J, Bos R, van der Mei H C, Handley P S. Physicochemistry of microbiol adhesion from an overall approach to the limits. In: Baszkin A, Norde W, editors. Physical chemistry of biological surfaces. New York, N.Y: Marcel Dekker; 2000. pp. 431–458. [Google Scholar]
- 5.Cowan M M, van der Mei H C, Stokroos I, Busscher H J. Heterogeneity of surfaces of subgingival bacteria as detected by zeta potential measurements. J Dent Res. 1992;71:1803–1806. doi: 10.1177/00220345920710110701. [DOI] [PubMed] [Google Scholar]
- 6.Cowan M M, Mikx F H M, Busscher H J. Electrophoretic mobility and hemagglutination of Treponema denticola ATCC33520. Colloids Surf B. 1994;2:407–410. [Google Scholar]
- 7.Ganeshkumar N, Hughes C V, Weiss E I. Coaggregation in dental plaque formation. In: Busscher H J, Evans L V, editors. Oral biofilms and plaque control. India: Harwood Academic Publishers; 1998. pp. 125–144. [Google Scholar]
- 8.Geertsema-Doornbusch G I, van der Mei H C, Busscher H J. Microbial cell surface hydrophobicity. The involvement of electrostatic interactions in microbial adhesion to hydrocarbon (MATH) J Microbiol Methods. 1993;18:61–68. [Google Scholar]
- 9.Geertsema-Doornbusch G I, Noordmans J, Bruce A W, Reid G, Khoury A E, van der Mei H C, Busscher H J. Quantitation of microbial cell surface heterogeneity by microelectrophoresis and electron microscopy—application to lactobacilli after serial passaging. J Microbiol Methods. 1994;19:269–277. [Google Scholar]
- 10.Glynn J R, Belongia B M, Arnold R G, Ogden K L, Baygents J C. Capillary electrophoresis measurements of electrophoretic mobility for colloidal particles of biological interest. Appl Environ Microbiol. 1998;64:2572–2577. doi: 10.1128/aem.64.7.2572-2577.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Handley P S. Structure, composition and function of surface structures on oral bacteria. Biofouling. 1990;2:239–264. [Google Scholar]
- 12.Handley P S. Negative staining. In: Mozes N, Handley P S, Busscher H J, Rouxhet P G, editors. Microbial cell surface analysis—structural and physicochemical methods. New York, N.Y: VCH Publishers; 1991. pp. 63–68. [Google Scholar]
- 13.James A M. Charge properties of microbial surfaces. In: Mozes N, Handley P S, Busscher H J, Rouxhet P G, editors. Microbial cell surface analysis—structural and physicochemical methods. New York, N.Y: VCH Publishers; 1991. pp. 221–262. [Google Scholar]
- 14.Jucker B A, Harms H, Zehnder A J B. Adhesion of the positively charged bacterium Stenotrophomona (Xanthomonas) maltophilia 70401 to glass and Teflon. J Bacteriol. 1996;178:5472–5479. doi: 10.1128/jb.178.18.5472-5479.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lichtenberg D, Rosenberg M, Scharfman N, Ofek I. A kinetic approach to bacterial adherence to hydrocarbons. FEMS Microbiol Lett. 1985;4:141–146. [Google Scholar]
- 16.Medrzycka K B. The effect of particle concentration in extremely dilute solutions. Colloid Polym Sci. 1991;269:85–90. [Google Scholar]
- 17.Mozes N, Marchal F, Hermesse M P, van Haecht J L, Reulieux L, Leonard A J, Rouxhet P G. Immobilization of microorganisms by a support, interplay of electrostatic and non-electrostatic interactions. Biotechnol Bioeng. 1987;30:439–450. doi: 10.1002/bit.260300315. [DOI] [PubMed] [Google Scholar]
- 18.Noordmans J, Kempen J, Busscher H J. Automated image analysis to determine zeta potential distributions in particulate microelectrophoresis. J Colloid Interface Sci. 1993;156:394–399. [Google Scholar]
- 19.Rosenberg M, Rosenberg E, Judes H, Weiss E. Bacterial adherence to hydrocarbons and to surfaces in the oral cavity. FEMS Microbiol Lett. 1983;20:1–5. [Google Scholar]
- 20.Rutter P R, Vincent B. The adhesion of microorganisms to surfaces: physico-chemical aspects. In: Berkeley R C W, Lynch J M, Melling J, Rutter P R, Vincent B, editors. Microbial adhesion to surfaces. London, United Kingdom: Ellis Horwood Limited; 1980. pp. 79–91. [Google Scholar]
- 21.van der Mei H C, de Vries J, Busscher H J. Hydrophobic and electrostatic cell surface properties of thermophilic dairy streptococci. Appl Environ Microbiol. 1993;59:4305–4312. doi: 10.1128/aem.59.12.4305-4312.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.van der Mei H C, van de Belt-Gritter B, Busscher H J. Implications of microbial adhesion to hydrocarbons for evaluating cell surface hydrophobicity. 2. Adhesion mechanisms. Colloids Surf B. 1995;5:117–126. [Google Scholar]
- 23.van Loosdrecht M C M, Lyklema J, Norde W, Schraa G, Zehnder A J B. The role of bacterial cell wall hydrophobicity in adhesion. Appl Environ Microbiol. 1987;53:1893–1897. doi: 10.1128/aem.53.8.1893-1897.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wit P J, Noordmans J, Busscher H J. Tracking of colloidal particles using microscopic image sequence analysis. Application to particulate microelectrophoresis and particle deposition. Colloids Surf A. 1997;125:85–92. [Google Scholar]