

Abstract
Objective (s): In this mini-review, we aimed to discuss the Wnt/β-catenin signaling pathway modulation in triple-negative breast cancer, particularly the contribution of lncRNAs and miRNAs in its regulation and their possible entwining role in breast cancer pathogenesis, proliferation, migration, or malignancy.
Background: Malignant tumor formation is very high for breast cancer in women and is a leading cause of death all over the globe. Among breast cancer subtypes, triple-negative breast cancer is rife in premenopausal women, most invasive, and prone to metastasis. Complex pathways are involved in this cancer’s pathogenesis, advancement, and malignancy, including the Wnt/β-catenin signaling pathway. This pathway is conserved among vertebrates and is necessary for sustaining cell homeostasis. It is regulated by several elements such as transcription factors, enhancers, non-coding RNAs (lncRNAs and miRNAs), etc.
Methods: We evaluated lncRNAs and miRNAs differentially expressed in triple-negative breast cancer (TNBC) from the cDNA microarray data set literature survey. Using in silico analyses combined with a review of the current literature, we anticipated identifying lncRNAs and miRNAs that might modulate the Wnt/β-catenin signaling pathway.
Result: The miRNAs and lncRNAs specific to triple-negative breast cancer have been identified based on literature and database searches. Tumorigenesis, metastasis, and EMT were all given special attention. Apart from cross-talk being essential for TNBC tumorigenesis and treatment outcomes, our results indicated eight upregulated and seven downregulated miRNAs and 19 upregulated and three downregulated lncRNAs that can be used as predictive or diagnostic markers. This consolidated information could be useful in the clinic and provide a combined literature resource for TNBC researchers working on the Wnt/β-catenin miRNA/lncRNA axis.
Conclusion: In conclusion, because the Wnt pathway and miRNAs/lncRNAs can modulate TNBC, their intertwinement results in a cascade of complex reactions that affect TNBC and related processes. Their function in TNBC pathogenesis has been highlighted in molecular processes underlying the disease progression.
Keywords: breast cancer, lncRNA, microRNA, wnt/β-catenin, pathogenesis
Introduction
Breast cancer represents the most common type of cancer worldwide, with high morbidity and mortality rates, (Agustsson et al., 2020). Breast cancer shows high heterogeneity, which impacts the clinical course of the disease. Differential expression profiles among patients lead to tumor tissue heterogeneity, resulting in variations in malignant behavior, prognosis, and responsiveness to standard therapies (Padmanaban et al., 2019). This cancer is prevalent in women and is the first leading cause of cancerous death in women. Breast cancer has been progressively increasing in most nations (Ferlay et al., 2015). According to WHO data (https://www.who.int/news-room/fact-sheets/detail/breast-cancer), 2.3 million women were diagnosed with breast cancer in 2020, with about 685,000 fatalities. This year, in the United States alone, around 281,550 cases of invasive and 49,290 cases of non-invasive breast cancer cases were diagnosed in women, according to the American Cancer Society. Most deaths occur due to metastasis instead of the primary tumor in breast cancer (Geng et al., 2014). Metastasis is linked with stem cells, which have characteristics such as self-renewal, differentiation ability, drug resistance, etc. (Phi et al., 2018). These properties favor aggressive behavior in breast cancer stem cells, leading to recurrent and aggressive tumors on and away from the primary site. Upregulation of Wnt/β-catenin signaling has been observed in triple-negative breast cancer (TNBC)/basal-like cancer when collated with other breast cancer subtypes (luminal A, B, and HER2 positive) or normal tissues (Gangrade et al., 2018).
TNBC can be classified into at least six distinct subtypes with differences in clinical behavior and treatment response. TNBC is highly invasive and, due to the non-expression of estrogen receptor, progesterone receptor, or HER-2, it has poor prognosis, high metastatic potential, and is disposed to relapse (Yin et al., 2020). TNBCs are more common in younger and obese women, with premenopausal African American women having the highest prevalence. BRCA1 and BRCA2 gene mutations are identified in approximately 20% of TNBC patients. P53 and Rb1 mutations are also quite common in TNBC tumors (van Barele et al., 2021). Differentially expressed ncRNAs have been found in a variety of human cancers, including breast cancer. Several lncRNA molecules have been linked to tumorigenesis, and their differential expression could constitute a potential new category of biomarkers. The lncRNA HOTAIR (HOX transcript antisense intergenic RNA) was associated with the polycomb repressive complex 2 (PRC2) and the histone demethylation enzyme LSD1 (lysine-specific demethylase 1) in cancer cells, resulting in epigenetic changes that promote tumor development and metastasis (Rodrigues de Bastos and Nagai, 2020). Other circulating lncRNAs, including MALAT1, GAS5, H19, and MEG3, have also been linked to survival and treatment response. LncRNAs have opened up a new field of study for researchers all over the world, and these molecules have been assigned major roles that may have a direct impact on patient survival and therapeutic responsiveness (Gupta et al., 2010). The SPARC gene (secreted protein acidic and rich in cysteine, also known as osteonectin or basement-membrane protein 40) encodes a 32-kDa matricellular glycoprotein involved in a variety of biological activities, including differentiation, proliferation, migration, and adhesion (Zhang et al., 2019).
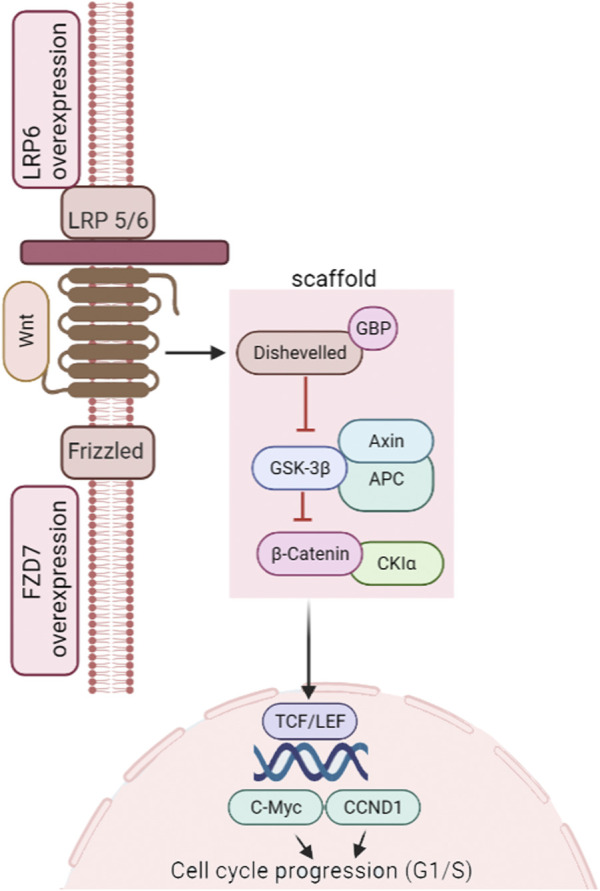
The canonical Wnt/β-catenin signaling pathway contributes to instigation (Figure 1), differentiation, and proliferation of TNBC cells (Zhang et al., 2018), leading to primary tumor formation (Xu et al., 2015), cellular transition of the epithelial-to-mesenchymal (EMT) state, and metastasis (Pohl et al., 2017). Chemoresistance has also been linked with Wnt/β-catenin signaling, with impaired pathways leading to drug-resistant TNBC (Merikhian et al., 2021). This is due to the synergistic contact between the Wnt target gene c-MYC and HIF-1α. This dual gene interaction diminishes cancer cell response to the given drugs. However, knockdown of β-catenin has been reported to cause TNBC cells to respond to doxorubicin or cisplatin (Xu et al., 2015).
FIGURE 1.
Wnt/β-catenin pathway in TNBC instigation. Figure adapted from the KEGG pathway (https://www.genome.jp/pathway/map05224).
This is why the study of this pathway is essential in TNBC, and a comprehensive updated literature review was undertaken to integrate information regarding the Wnt/β-catenin signaling pathway and TNBC.
Wnt/β-Catenin Signaling in TNBC
The Wnt pathway is tangled with low-density lipoprotein receptor-related protein5/6 (LRP5/6) and frizzled (FZD) receptors for its activation (King et al., 2012). If Wnts are insufficient or non-functional, β-catenin pools with a tetrad of proteins (axin, casein kinase 1 (CK1), adenomatous polyposis coli (APC) tumor suppressor, and glycogen synthase kinase-3b (GSK3b). Phosphorylation (by CK1 and GSK3b) is followed by ubiquitination and ultimate degradation of β-catenin (26S proteasome). Conversely, in the presence of the Wnt signal, β-catenin attaches to FZD and LRP5/6 receptors, leading to inhibition of GSK3b and stabilization of cytosolic β-catenin. This β-catenin is then shifted to the nucleus. It associates with T-cell factor/lymphoid-enhancing factor (TCF/LEF) to incite the downstream expression of cell development and cell cycle control genes (MacDonald et al., 2009). Secreted proteins regulate this signaling at the cell surface, where the central modulators are Wnt and R-spondin (Rspo). Inhibitors include Wnt Inhibitory Factor 1 (WIF1), Dickkopf (Dkk), soluble Frizzled-related protein (sFRP), and sclerostin (SOST) (Yao et al., 2011; Danieau et al., 2019). Abnormal Wnt signaling has been implicated in TNBC tumorigenesis (Xu et al., 2015; Mohammadi Yeganeh et al., 2017), stemness, metastasis, and prognosis (Ryu et al., 2020). Dey et al. (2013) identified that patients with dysregulated Wnt/β-catenin signaling had a higher chance of lung and brain metastases. Dysregulation of the canonical pathway is responsible for metastasis in more than half of breast cancer patients as the nucleolar β-catenin level is elevated. However, mutations in the relevant genes are uncommon (Yu et al., 2019). It implies that the role of the β-catenin is indispensable to the Wnt signaling in TNBC advancement, with the dissemination of non-phosphorylated cytoplasmic β-catenin to the nucleus having an imperative role in TNBC metastasis (Breuer et al., 2019; Satriyo et al., 2019).
Green et al. (2013) discovered that Wnt ligands caused enhanced transcription in the majority of TNBC cell lines, while Xu et al. (2015) reported that nuclear accumulation of β-catenin is linked with TNBC characteristics. The role of the key modulators of this pathway in TNBC includes the action of FZD receptors, LRP5/6, Receptor Tyrosine Kinase–Like Orphan Receptors (RORs), and Dead Box Proteins (DDX3 and DDX5) (Pohl et al., 2017). LRP5/6 is crucial in mammary development and is allied with tumorigenesis (Goel et al., 2012). Overexpression of LRP5/6 has been seen in TNBC (Maubant et al., 2018). LRP6 has also been linked with migration and invasion of cells (Ma et al., 2017). Elevated FZD7 expression in TNBC also promotes tumorigenesis (Yang et al., 2011) via transformation-related protein 63 (p63) (Chakrabarti et al., 2014), while FZD8-driven Wnt signaling mediated by c-Myc overexpression drives chemoresistance (Yin et al., 2013). Dead box proteins DDX3 and DDX5 have shown increased EMT in TNBC. DDX3 is associated with increased motility and invasiveness. In comparison, DDX5 is linked with tumorigenesis and cancer cell progression (Moore et al., 2010; Wang et al., 2012; Guturi et al., 2014; Pohl et al., 2017).
Wnt/β-Catenin Signaling in TNBC Stem Cells
Cancer stem cells (CSCs) or cancer stem-like cells within the tumor, being less responsive to environmental stop signals, are responsible for cancer progression, metastasis, chemoresistance, and hence, cancer relapse (Clarke, 2019). They differ from other cancer cells as they use mitochondrial reactive oxygen species (ROS) for respiration, which means higher oxygen consumption, increased mitochondrial mass, and high resistance to DNA damage (Peiris-Pagès et al., 2018; Scatena et al., 2018). Abnormal Wnt/β-catenin signaling is linked with CSC formation and, hence, tumorigenesis, stemness, migration, and chemoresistance in TNBC (Alraouji et al., 2020). Upregulation of β-catenin triggers the CSC phenotype of TNBC, and repression by cadherin leads to suppression of this phenotype (Satriyo et al., 2019). Jang et al. (2015) observed that Wnt/β-catenin pathway genes (WNT1, FZD1, TCF4, and LEF1) are upregulated in CSC-enriched mammospheres in breast cancer, while signaling proteins (LEF1, TCF4, and β-catenin) were increased in high CSC activity, depicting cell fraction, compared to that with low CSC activity. However, recently, Brilliant et al. (2019) reported 11% of Wnt signaling expression in high vs. 33% in the low content of cancer stem cells. In contrast, some researchers have reported inhibiting TNBC CSCs by drugs targeting the Wnt/β-catenin pathway (like hydroxytyrosol) (Cruz-Lozano et al., 2019). Others have reported that β-catenin is also responsible for drug resistance (e.g., doxorubicin resistance) in TNBC CSCs (Xu et al., 2015).
lncRNA and miRNAs Entangled With the Wnt/β-Catenin Pathway in TNBC
According to recent research, more than 90% of the transcripts in the human genome may not be able to code for proteins (Wilusz et al., 2009) but regulate the expression of nearby genes (He and Hannon, 2004; Trzybulska et al., 2018). They are categorized according to their size and function (Trzybulska et al., 2018), with microRNAs (miRNAs) being 19–24 nucleotides long and long non-coding RNAs (lncRNAs) being >200 nucleotides in length. There are other non-coding RNAs such as piwiRNAs, free circulating RNAs, and snoRNAs, but our focus will be on miRNAs and lncRNAs in this review. Long non-coding RNAs are abundant in human cells and play critical roles in a range of biological processes, including cell cycle regulation (Lu et al., 2016), genomic expression (Ballantyne et al., 2016), and cell differentiation (Chen et al., 2016). Increasing evidence has recently indicated that abnormal lncRNA expression is linked with various tumor forms (Mendell, 2016). Researchers have proved that miRNA dysregulation also leads to human cancer via different mechanisms (Karimzadeh et al., 2021), including altered epigenetics (Arif et al., 2020) and abnormal transcriptional control (Müller et al., 2019; Ali Syeda et al., 2020). Apart from performing as oncogenes, where they support proliferative signaling (Miao et al., 2017), invasion (Chen et al., 2017), repelling cell death, eluding progression suppressors, and metastasis, they act as tumor suppressors too (Hong et al., 2019). This has led to their demarcation as potential biomarkers of cancer (Gai et al., 2018; Dai et al., 2019).
miRNA expression in BCSCs and cancer cells signals that they are crucial for promoting characteristics such as stemness and tumorigenesis. Piasecka et al. (2018) reviewed 121 articles demonstrating the role of miRNAs in TNBC. After scanning a plethora of literature, it was revealed that the miRNAs not only serve as predictive markers of TNBC but also have prognostic clinical utility. They assist in attaining CSC properties in TNBC and EMT. Since these properties are conditions for metastasis, miRNAs play an essential role in cell transition to the metastasis stage. Several miRNAs, which are differently expressed in TNBC cells compared to normal cells and entangled with the Wnt/β-catenin pathway, have also been identified (Avery-Kiejda et al., 2014; Goh et al., 2016; Pohl et al., 2017).
More than 70 miRNAs (previously implicated in BC) have shown differential expression in TNBC, targeting 16 genes from the Wnt pathway, causing their increased expression in TNBC metastasis. The impacting miRNAs comprised the miR-17-92 oncogenic cluster members and the miR-200 family, revealing that most miRNAs are not mainly associated with a cancer subtype (Pohl et al., 2017). miR-340 alters CTNNB1, MYC, and ROCK1 gene expression of the Wnt pathway and causes apoptosis in TNBC (Mohammadi Yeganeh et al., 2017). miR-203 expresses higher methylation and is downregulated in TNBC, along with downregulation of the Wnt pathway (Taube et al., 2013). Telonis and Rigoutsos (2018) have identified miR-200c, miR-21, miR-17/92 cluster, and the miR-183/96/182 cluster to be upregulated in TNBC. Wang et al. (2019) demonstrated that miR-125, MiR, MiR-145, MiR-381, MiR-136, and MiR-4324 are associated with the poorest prognosis in TNBC patients. Thus, miRNAs play a critical role in TNBC and are intertwined with the Wnt pathway gene regulation, making them essential players in Wnt-mediated TNBC progression, prognosis, and other outcomes. Previously reported miRNAs with a role in TNBC regulation have been collected in Table 1 below:
TABLE 1.
miRNAs impacting TNBC via the Wnt/β-Catenin axis.
| S. No. | MiRNA | Upregulation or downregulation | Cell line | References |
|---|---|---|---|---|
| 1 | miR-142 | Upregulated | HEK293T, MCF7, and MDA-MB-231 | Isobe et al. (2014) |
| 2 | miR221/222 | Upregulated | MDA-MB-231, MCF7, MDA-MB-468, Hs 578T, HCC1937, and MDA-MB-231, SKBR3, T47D, BT-474, 4T1 | Liu et al. (2018) |
| 3 | miR-124 | Upregulated | BT20, HCC70, 293T | Yang et al. (2020) |
| 4 | miR-125b | Upregulated | MDA-MB-468, MDA-MB-231, MCF-10A, and MCF-7 | Nie et al. (2019) |
| 5 | miR-137 | Downregulated | HCC38, MDA-MB-231, and MDA-MB-468 | Cheng et al. (2019) |
| 6 | miR-29b-1 | Downregulated | MB-231, MDA-MB-468, BT20, and HCC-1395 | Drago-Ferrante et al. (2017) |
| 7 | miR-105 | Upregulated | MB-361, MCF-7, BT-483, AU565, SkBR3, MCF-10A, MB-231 (MDA-MB-231), Hs578T, HCC1599, HCC1806, HCC1937, BT-549, DU4475, and HCC70 | Li et al. (2017) |
| 8 | miR-93 | Upregulated | MB-361, MCF-7, BT-483, AU565, SkBR3, MCF-10A, MB-231 (MDA-MB-231), Hs578T, HCC1599, HCC1806, HCC1937, BT-549, DU4475, and HCC70 | Li et al. (2017) |
| 9 | miR-27a | Upregulated | BT-549, MDA-MB-231, MDA-MB-468, MDA-MB-453, MCF-10A, and DU4475 | Wu et al. (2020) |
| 10 | miR-130a | Downregulated | MDA-MB-468, MDA-MB-231, and MF-10A | Poodineh et al. (2020) |
| 11 | miR-384 | Downregulated | MCF-7; MDA-MB-231 | Wang et al. (2018a) |
| 12 | miR-34a | Downregulated | SUM159PT, mammospheres and Comma-Dβ cells | Bonetti et al. (2019) |
| 13 | miR-374a | Downregulated | MCF7, T47D, BT474, MDA-MB-231, MDA-MB-435, MDA-MB-468, and 4T1 | Cai et al. (2013) |
| 14 | miR-340 | Downregulated | MCF-10A, MDA-MB-231, and HEK 293T | Mohammadi Yeganeh et al. (2017) |
| 15 | miR-218 | Upregulated | MCF-10A, MCF-7, and MDA-MB-231 | Taipaleenmäki et al. (2016) |
LncRNAs may support transcription; aid RNA interference; act as a decoy, peptide, and scaffold; or function as a guide/enhancer RNA (Li et al., 2014). They may deactivate miRNAs in cancer via the “sponge” effect, that is, act as a competing molecule or decoy to attach to miRNAs and perturb them from their target (Fan et al., 2018; Huang et al., 2019) (Figure 2). They have been demarcated as diagnostic and therapeutic targets. Here, we review lncRNA intertwined with the Wnt pathway and TNBC progression, pathogenesis, prognosis, or invasion. LncRNA Lung Cancer–Associated Transcript 1 (LUCAT1) is interlinked with miR-5582 and regulates breast cancer stemness via the Wnt/β-catenin pathway (Zheng et al., 2019). UCA1 promotes EMT (Xiao et al., 2016), while actin filament–associated protein 1 antisense RNA1 (AFAP1-AS1) promotes EMT and tumorigenesis (Zhang et al., 2018). and differentiation antagonizing non-protein coding RNA (DANCR) negatively regulates the Wnt pathway (Li and Zhou, 2018) to uplift tumorigenesis (Tao et al., 2019), EMT, and stemness in TNBC (Zhang and Wang, 2020). HOX Antisense Intergenic RNA (HOTAIR) modulates the Wnt pathway (Li et al., 2016) and leads to metastasis (Collina et al., 2019) and poor prognosis in TNBC via upregulation by miR-146a (Liang et al., 2019).
FIGURE 2.

LncRNA–miRNA gene expression regulation in normal vs. cancer phenotype.
Jiang et al. (2020) reported that DiGeorge Syndrome Critical Region Gene 5 (DGCR5) induces tumorigenesis in TNBC. LncRNA associated with poor prognosis of hepatocellular carcinoma (AWPPH) promotes TNBC growth by upregulating the frizzled homolog 7 (FZD7) ligand of the Wnt pathway (Wang et al., 2018b) and decreased manifestation of lncRNA has been reported to increase the malignant spread of TNBCs (Liu et al., 2017). Long intergenic non-protein coding RNA 1234 (LINC01234) modulates TNBC cell growth, invasion, and EMT positively (Xiao et al., 2021). We also mined lncRNAs with a role in TNBC and impacted by Wnt pathway genes (Table 2) from the lnc2Cancer 3.0 database (Gao et al., 2021).
TABLE 2.
LncRNAs impacting TNBC via the Wnt/β-Catenin axis mined from the lnc2 Cancer database.
| S. No. | Name | Method of identification | Expression pattern | References |
|---|---|---|---|---|
| 1 | ANRIL | qPCR, luciferase reporter assay, RIP. | Upregulated | Xu et al. (2017) |
| 2 | AFAP1-AS1 | qPCR, Western blot, in vitro knockdown etc. | Upregulated | Zhang et al. (2018) |
| 3 | AWPPH | qRT-PCR etc. | Upregulated | Liu et al. (2019) |
| 4 | CCAT1 | qPCR, luciferase reporter assay, Western blot | Upregulated | Han et al. (2019) |
| 5 | DANCR | qPCR, Western blot, in vitro knockdown, RIP etc. | Upregulated | Tang et al. (2018a) |
| 6 | FAM83H-AS1 | qRT-PCR, Western blot | Upregulated | Han et al. (2020) |
| 7 | GAS5 | qPCR, Western blot, Luciferase reporter assay | Downregulated | Li et al. (2018) |
| 8 | H19 | qPCR, Western blot, other | Upregulated | Han et al. (2018) |
| 9 | HOTAIR | Microarray, qPCR etc. | Upregulated | Chen et al. (2015) |
| 10 | LINC00052 | Microarray, qPCR etc. | Downregulated | Lv et al. (2016) |
| 11 | LINC00115 | qRT-PCR, Western blot | Upregulated | Yuan et al. (2020) |
| 12 | LINC00152 | qPCR, Western blot | Upregulated | Wu et al. (2018) |
| 13 | LINC00173 | qPCR | Upregulated | Fan et al. (2020) |
| 14 | LINC01133 | qPCR, Western blot | Upregulated | Tu et al. (2019) |
| 15 | LUCAT1 | qPCR, Western blot, a luciferase reporter assay, in vitro knockdown, RIP | Upregulated | Mou and Wang, (2019) |
| 16 | MALAT1 | Microarray, qPCR etc. | Upregulated | Chen et al. (2015) |
| 17 | NEAT1 | qPCR, Western blot | Upregulated | Shin et al. (2019) |
| 18 | PCAT6 | qPCR, Western blot, luciferase reporter assay, in vitro knockdown etc. | Upregulated | Shi et al. (2020) |
| 19 | PVT1 | qPCR, Western blot, RNAi, other | Upregulated | Wang et al. (2018c) |
| 20 | SOX21-AS1 | qPCR, Western blot, luciferase reporter assay etc | Upregulated | Liu et al. (2020) |
| 21 | XIST | qRT-PCR, Western blot | Downregulated | Li et al. (2020) |
| 22 | ZEB1-AS1 | qRT-PCR, RIP, dual luciferase reporter assay | Upregulated | Luo et al. (2020) |
In addition, lncRNA–miRNA interactions entwining the Wnt pathway have also been noted in TNBC (Volovat et al., 2020). Among these, intranuclear Metastasis-Related Lung Adenocarcinoma Transcript 1 (MALAT1) acts as a sponge of miR-129-5p (Dong et al., 2015), and silencing of this non-coding gene causes a decline in cell propagation and movement, illustrating its role in TNBC pathology (Zuo et al., 2017). AWPPH stimulates cell proliferation and contributes to drug therapy resistance when combined with miRNA-21 and is being exploited as a diagnostic biomarker (Cascione et al., 2013; Liu et al., 2019). At the same time, TUG1 impacts miR-197, prompts NLK expression, and incapacitates the Wnt signaling pathway, making the TNBC cells susceptible to cisplatin therapy (Tang et al., 2018b). AFAP1-AS1 controls miRNA-2110, leading to tumorigenesis and cell invasion (Zuo et al., 2017). The diminished NEF and boosted miRNA-155 levels segregate TNBC patients from controls, suggesting an interlinked modulation prompting enhanced invasion and cell migration in the case of increased miRNA-155 (Song et al., 2019).
Conclusion
This narrative review was centered on TNBC association with the Wnt/β-catenin pathway. Moreover, miRNAs and lncRNAs shown to be specific to triple-negative breast carcinoma were listed, both from literature and database searches. Since the Wnt pathway and miRNAs/lncRNAs can modulate TNBC and their intertwinement forms a cascade of complex reactions that impact TNBC and associated processes, their role in TNBC pathology was highlighted concerning molecular processes underlying disease progression. Particular emphasis was put on tumorigenesis, metastasis, and EMT. Apart from cross-talk critical for TNBC tumorigenesis and treatment outcomes, miRNA and lncRNA can serve as predictive or diagnostic markers, so this consolidated information might be of clinical use and offer a consolidated literature resource to scientists working on the Wnt/β-catenin and miRNA/lncRNA axis of TNBC. They also have a role in chemotherapy resistance. However, information on detailed analyses of Wnt/miRNA/lncRNA axis mechanisms in TNBC is still scant, and more work needs to be carried out to infer the pivotal role of these moieties in TNBC. Owing to the heterogeneity of TNBC, recognition of subgroups or their pathologies based on the varied signatures of this axis could be interesting to explore. Furthermore, knock-out or knock-in functional studies in model organisms could be beneficial to understanding the comprehensive role of this axis in TNBC.
Acknowledgments
We acknowledge the China Postdoctoral Science Foundation for supporting this study.
Author Contributions
Conception/Design: WW, QZ. Collection and assembly of data: XH, WX. Manuscript writing: XH, WW. Final approval of manuscript: WW, QZ, XH, WX.
Funding
This study was supported by the China Postdoctoral Science Foundation Grant (No. 2019M651216).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
- Agustsson A. S., Birgisson H., Agnarsson B. A., Jonsson T., Stefansdottir H., Wärnberg F., et al. (2020). In Situ breast Cancer Incidence Patterns in Iceland and Differences in Ductal Carcinoma In Situ Treatment Compared to Sweden. Sci. Rep. 10 (1), 17623. 10.1038/s41598-020-74134-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ali Syeda Z., Langden S. S. S., Munkhzul C., Lee M., Song S. J. (2020). Regulatory Mechanism of MicroRNA Expression in Cancer. Int. J. Mol. Sci. 21 (5). 10.3390/ijms21051723 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alraouji N. N., Al-Mohanna F. H., Ghebeh H., Arafah M., Almeer R., Al-Tweigeri T., et al. (2020). Tocilizumab Potentiates Cisplatin Cytotoxicity and Targets Cancer Stem Cells in Triple-Negative Breast Cancer. Mol. Carcinog. 59 (9), 1041–1051. 10.1002/mc.23234 [DOI] [PubMed] [Google Scholar]
- Arif K. M. T., Elliott E. K., Haupt L. M., Griffiths L. R. (2020). Regulatory Mechanisms of Epigenetic miRNA Relationships in Human Cancer and Potential as Therapeutic Targets. Cancers (Basel) 12 (10). 10.3390/cancers12102922 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Avery-Kiejda K. A., Braye S. G., Mathe A., Forbes J. F., Scott R. J. (2014). Decreased Expression of Key Tumour Suppressor microRNAs Is Associated with Lymph Node Metastases in Triple Negative Breast Cancer. BMC Cancer 14, 51. 10.1186/1471-2407-14-51 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ballantyne R. L., Zhang X., Nuñez S., Xue C., Zhao W., Reed E., et al. (2016). Genome-wide Interrogation Reveals Hundreds of Long Intergenic Noncoding RNAs that Associate with Cardiometabolic Traits. Hum. Mol. Genet. 25 (14), 3125–3141. 10.1093/hmg/ddw154 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonetti P., Climent M., Panebianco F., Tordonato C., Santoro A., Marzi M. J., et al. (2019). Dual Role for miR-34a in the Control of Early Progenitor Proliferation and Commitment in the Mammary Gland and in Breast Cancer. Oncogene 38 (3), 360–374. 10.1038/s41388-018-0445-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breuer E. K., Fukushiro-Lopes D., Dalheim A., Burnette M., Zartman J., Kaja S., et al. (2019). Potassium Channel Activity Controls Breast Cancer Metastasis by Affecting β-catenin Signaling. Cell Death Dis. 10 (3), 180. 10.1038/s41419-019-1429-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brilliant A., Brilliant Y., Sazonov S., Wnt (2019). WNT, Hedgehog and NOTCH Signaling Pathways in Triple Negative Breast Cancer with High and Low Content of Cancer Stem Cells. Ann. Oncol. 30, iii13–iii14. 10.1093/annonc/mdz095.039 [DOI] [Google Scholar]
- Cai J., Guan H., Fang L., Yang Y., Zhu X., Yuan J., et al. (2013). MicroRNA-374a Activates Wnt/β-Catenin Signaling to Promote Breast Cancer Metastasis. J. Clin. Invest. 123 (2), 566–579. 10.1172/JCI65871 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cascione L., Gasparini P., Lovat F., Carasi S., Pulvirenti A., Ferro A., et al. (2013). Integrated microRNA and mRNA Signatures Associated with Survival in Triple Negative Breast Cancer. PLoS One 8 (2), e55910. 10.1371/journal.pone.0055910 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chakrabarti R., Wei Y., Hwang J., Hang X., Andres Blanco M., Choudhury A., et al. (2014). ΔNp63 Promotes Stem Cell Activity in Mammary Gland Development and Basal-like Breast Cancer by Enhancing Fzd7 Expression and Wnt Signalling. Nat. Cell Biol. 16 (10), 10041–11513. 10.1038/ncb3040 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen C., Li Z., Yang Y., Xiang T., Song W., Liu S. (2015). Microarray Expression Profiling of Dysregulated Long Non-coding RNAs in Triple-Negative Breast Cancer. Cancer Biol. Ther. 16 (6), 856–865. 10.1080/15384047.2015.1040957 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y., Min L., Ren C., Xu X., Yang J., Sun X., et al. (2017). miRNA-148a Serves as a Prognostic Factor and Suppresses Migration and Invasion through Wnt1 in Non-small Cell Lung Cancer. PLoS One 12 (2), e0171751. 10.1371/journal.pone.0171751 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Z., Yu C., Zhan L., Pan Y., Chen L., Sun C. (2016). Lncrna crnde promotes hepatic carcinoma cell proliferation, migration and invasion by suppressing mir-384. Am. J. Cancer Res. 6 (10), 2299–2309. [PMC free article] [PubMed] [Google Scholar]
- Cheng S., Huang Y., Lou C., He Y., Zhang Y., Zhang Q. (2019). FSTL1 Enhances Chemoresistance and Maintains Stemness in Breast Cancer Cells via Integrin β3/Wnt Signaling under miR-137 Regulation. Cancer Biol. Ther. 20 (3), 328–337. 10.1080/15384047.2018.1529101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke M. F. (2019). Clinical and Therapeutic Implications of Cancer Stem Cells. N. Engl. J. Med. 380 (23), 2237–2245. 10.1056/NEJMra1804280 [DOI] [PubMed] [Google Scholar]
- Collina F., Aquino G., Brogna M., Cipolletta S., Buonfanti G., De Laurentiis M., et al. (2019). Lncrna hotair up-regulation is strongly related with lymph nodes metastasis and lar subtype of triple negative breast cancer. J. Cancer 10 (9), 2018–2024. 10.7150/jca.29670 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cruz-Lozano M., González-González A., Marchal J. A., Muñoz-Muela E., Molina M. P., Cara F. E., et al. (2019). Hydroxytyrosol Inhibits Cancer Stem Cells and the Metastatic Capacity of Triple-Negative Breast Cancer Cell Lines by the Simultaneous Targeting of Epithelial-To-Mesenchymal Transition, Wnt/β-Catenin and TGFβ Signaling Pathways. Eur. J. Nutr. 58 (8), 3207–3219. 10.1007/s00394-018-1864-1 [DOI] [PubMed] [Google Scholar]
- Dai W., He J., Zheng L., Bi M., Hu F., Chen M., et al. (2019). miR-148b-3p, miR-190b, and miR-429 Regulate Cell Progression and Act as Potential Biomarkers for Breast Cancer. J. Breast Cancer 22 (2), 219–236. 10.4048/jbc.2019.22.e19 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Danieau G., Morice S., Rédini F., Verrecchia F., Royer B. B. (2019). New Insights about the Wnt/β-Catenin Signaling Pathway in Primary Bone Tumors and Their Microenvironment: A Promising Target to Develop Therapeutic Strategies? Int. J. Mol. Sci. 20 (15). 10.3390/ijms20153751 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dey N., Barwick B. G., Moreno C. S., Ordanic-Kodani M., Chen Z., Oprea-Ilies G., et al. (2013). Wnt Signaling in Triple Negative Breast Cancer Is Associated with Metastasis. BMC Cancer 13, 537. 10.1186/1471-2407-13-537 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong Y., Liang G., Yuan B., Yang C., Gao R., Zhou X. (2015). MALAT1 Promotes the Proliferation and Metastasis of Osteosarcoma Cells by Activating the PI3K/Akt Pathway. Tumour Biol. 36 (3), 1477–1486. 10.1007/s13277-014-2631-4 [DOI] [PubMed] [Google Scholar]
- Drago-Ferrante R., Pentimalli F., Carlisi D., De Blasio A., Saliba C., Baldacchino S., et al. (2017). Suppressive Role Exerted by microRNA-29b-1-5p in Triple Negative Breast Cancer through SPIN1 Regulation. Oncotarget 8 (17), 28939–28958. 10.18632/oncotarget.15960 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan C. N., Ma L., Liu N. (2018). Systematic Analysis of lncRNA-miRNA-mRNA Competing Endogenous RNA Network Identifies Four-lncRNA Signature as a Prognostic Biomarker for Breast Cancer. J. Transl. Med. 16 (1), 264. 10.1186/s12967-018-1640-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan H., Yuan J., Li X., Ma Y., Wang X., Xu B., et al. (2020). LncRNA LINC00173 Enhances Triple-Negative Breast Cancer Progression by Suppressing miR-490-3p Expression. Biomed. Pharmacother. 125, 109987. 10.1016/j.biopha.2020.109987 [DOI] [PubMed] [Google Scholar]
- Ferlay J., Soerjomataram I., Dikshit R., Eser S., Mathers C., Rebelo M., et al. (2015). Cancer Incidence and Mortality Worldwide: Sources, Methods and Major Patterns in GLOBOCAN 2012. Int. J. Cancer 136 (5), E359–E386. 10.1002/ijc.29210 [DOI] [PubMed] [Google Scholar]
- Gai C., Camussi F., Broccoletti R., Gambino A., Cabras M., Molinaro L., et al. (2018). Salivary Extracellular Vesicle-Associated miRNAs as Potential Biomarkers in Oral Squamous Cell Carcinoma. BMC Cancer 18 (1), 439. 10.1186/s12885-018-4364-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gangrade A., Pathak V., Augelli-Szafran C. E., Wei H. X., Oliver P., Suto M., et al. (2018). Preferential Inhibition of Wnt/β-Catenin Signaling by Novel Benzimidazole Compounds in Triple-Negative Breast Cancer. Int. J. Mol. Sci. 19 (5). 10.3390/ijms19051524 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao Y., Shang S., Guo S., Li X., Zhou H., Liu H., et al. (2021). Lnc2Cancer 3.0: an Updated Resource for Experimentally Supported lncRNA/circRNA Cancer Associations and Web Tools Based on RNA-Seq and scRNA-Seq Data. Nucleic Acids Res. 49 (D1), D1251–D1258. 10.1093/nar/gkaa1006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geng S. Q., Alexandrou A. T., Li J. J. (2014). Breast Cancer Stem Cells: Multiple Capacities in Tumor Metastasis. Cancer Lett. 349 (1), 1–7. 10.1016/j.canlet.2014.03.036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goel S., Chin E. N., Fakhraldeen S. A., Berry S. M., Beebe D. J., Alexander C. M. (2012). Both LRP5 and LRP6 Receptors Are Required to Respond to Physiological Wnt Ligands in Mammary Epithelial Cells and Fibroblasts. J. Biol. Chem. 287 (20), 16454–16466. 10.1074/jbc.M112.362137 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goh J. N., Loo S. Y., Datta A., Siveen K. S., Yap W. N., Cai W., et al. (2016). microRNAs in Breast Cancer: Regulatory Roles Governing the Hallmarks of Cancer. Biol. Rev. Camb Philos. Soc. 91 (2), 409–428. 10.1111/brv.12176 [DOI] [PubMed] [Google Scholar]
- Green J. L., La J., Yum K. W., Desai P., Rodewald L. W., Zhang X., et al. (2013). Paracrine Wnt Signaling Both Promotes and Inhibits Human Breast Tumor Growth. Proc. Natl. Acad. Sci. U. S. A. 110 (17), 6991–6996. 10.1073/pnas.1303671110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gupta R. A., Shah N., Wang K. C., Kim J., Horlings H. M., Wong D. J., et al. (2010). Long non-coding rna hotair reprograms chromatin state to promote cancer metastasis. Nature 464 (7291), 1071–1076. 10.1038/nature08975 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guturi K. K., Sarkar M., Bhowmik A., Das N., Ghosh M. K. (2014). DEAD-box Protein P68 Is Regulated by β-catenin/transcription Factor 4 to Maintain a Positive Feedback Loop in Control of Breast Cancer Progression. Breast Cancer Res. 16 (6), 496. 10.1186/s13058-014-0496-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han C., Fu Y., Zeng N., Yin J., Li Q. (2020). LncRNA FAM83H-AS1 Promotes Triple-Negative Breast Cancer Progression by Regulating the miR-136-5p/metadherin axis. Aging (Albany NY) 12 (4), 3594–3616. 10.18632/aging.102832 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han C., Li X., Fan Q., Liu G., Yin J. (2019). CCAT1 Promotes Triple-Negative Breast Cancer Progression by Suppressing miR-218/ZFX Signaling. Aging (Albany NY) 11 (14), 4858–4875. 10.18632/aging.102080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han J., Han B., Wu X., Hao J., Dong X., Shen Q., et al. (2018). Knockdown of lncRNA H19 Restores Chemo-Sensitivity in Paclitaxel-Resistant Triple-Negative Breast Cancer through Triggering Apoptosis and Regulating Akt Signaling Pathway. Toxicol. Appl. Pharmacol. 359, 55–61. 10.1016/j.taap.2018.09.018 [DOI] [PubMed] [Google Scholar]
- He L., Hannon G. J. (2004). MicroRNAs: Small RNAs with a Big Role in Gene Regulation. Nat. Rev. Genet. 5 (7), 522–531. 10.1038/nrg1379 [DOI] [PubMed] [Google Scholar]
- Hong B. S., Ryu H. S., Kim N., Kim J., Lee E., Moon H., et al. (2019). Tumor Suppressor miRNA-204-5p Regulates Growth, Metastasis, and Immune Microenvironment Remodeling in Breast Cancer. Cancer Res. 79 (7), 1520–1534. 10.1158/0008-5472.CAN-18-0891 [DOI] [PubMed] [Google Scholar]
- Huang Q. Y., Liu G. F., Qian X. L., Tang L. B., Huang Q. Y., Xiong L. X. (2019). Long Non-coding RNA: Dual Effects on Breast Cancer Metastasis and Clinical Applications. Cancers (Basel) 11 (11). 10.3390/cancers11111802 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Isobe T., Hisamori S., Hogan D. J., Zabala M., Hendrickson D. G., Dalerba P., et al. (2014). miR-142 Regulates the Tumorigenicity of Human Breast Cancer Stem Cells through the Canonical WNT Signaling Pathway. Elife 3. 10.7554/eLife.01977 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jang G. B., Kim J. Y., Cho S. D., Park K. S., Jung J. Y., Lee H. Y., et al. (2015). Blockade of Wnt/β-Catenin Signaling Suppresses Breast Cancer Metastasis by Inhibiting CSC-like Phenotype. Sci. Rep. 5, 12465. 10.1038/srep12465 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang D., Wang C., He J. (2020). Long Non-coding RNA DGCR5 Incudes Tumorigenesis of Triple-Negative Breast Cancer by Affecting Wnt/β-Catenin Signaling Pathway. J. BUON 25 (2), 702–708. [PubMed] [Google Scholar]
- Karimzadeh M. R., Pourdavoud P., Ehtesham N., Qadbeigi M., Asl M. M., Alani B., et al. (2021). Regulation of DNA Methylation Machinery by Epi-miRNAs in Human Cancer: Emerging New Targets in Cancer Therapy. Cancer Gene Ther. 28 (3-4), 157–174. 10.1038/s41417-020-00210-7 [DOI] [PubMed] [Google Scholar]
- King T. D., Suto M. J., Li Y. (2012). The Wnt/β-Catenin Signaling Pathway: a Potential Therapeutic Target in the Treatment of Triple Negative Breast Cancer. J. Cell Biochem. 113 (1), 13–18. 10.1002/jcb.23350 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li H. Y., Liang J. L., Kuo Y. L., Lee H. H., Calkins M. J., Chang H. T., et al. (2017). miR-105/93-3p Promotes Chemoresistance and Circulating miR-105/93-3p Acts as a Diagnostic Biomarker for Triple Negative Breast Cancer. Breast Cancer Res. 19 (1), 133. 10.1186/s13058-017-0918-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J., Yang S., Su N., Wang Y., Yu J., Qiu H., et al. (2016). Overexpression of long non-coding rna hotair leads to chemoresistance by activating the wnt/β-catenin pathway in human ovarian cancer. Tumour Biol. 37 (2), 2057–2065. 10.1007/s13277-015-3998-6 [DOI] [PubMed] [Google Scholar]
- Li J., Zhou L. (2018). Overexpression of lncrna dancr positively affects progression of glioma via activating wnt/β-catenin signaling. Biomed. Pharmacother. 102, 602–607. 10.1016/j.biopha.2018.03.116 [DOI] [PubMed] [Google Scholar]
- Li S., Zhou J., Wang Z., Wang P., Gao X., Wang Y. (2018). Long Noncoding RNA GAS5 Suppresses Triple Negative Breast Cancer Progression through Inhibition of Proliferation and Invasion by Competitively Binding miR-196a-5p. Biomed. Pharmacother. 104, 451–457. 10.1016/j.biopha.2018.05.056 [DOI] [PubMed] [Google Scholar]
- Li X., Hou L., Yin L., Zhao S. (2020). Lncrna xist interacts with mir-454 to inhibit cells proliferation, epithelial mesenchymal transition and induces apoptosis in triple-negative breast cancer. J. Biosci. 45. 10.1007/s12038-020-9999-7 [DOI] [PubMed] [Google Scholar]
- Li X., Wu Z., Fu X., Han W. (2014). lncRNAs: Insights into Their Function and Mechanics in Underlying Disorders. Mutat. Res. Rev. Mutat. Res. 762, 1–21. 10.1016/j.mrrev.2014.04.002 [DOI] [PubMed] [Google Scholar]
- Liang H., Huang W., Wang Y., Ding L., Zeng L. (2019). Overexpression of MiR-146a-5p Upregulates lncRNA HOTAIR in Triple-Negative Breast Cancer Cells and Predicts Poor Prognosis. Technol. Cancer Res. Treat. 18, 1533033819882949. 10.1177/1533033819882949 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu A. N., Qu H. J., Gong W. J., Xiang J. Y., Yang M. M., Zhang W. (2019). LncRNA AWPPH and miRNA-21 Regulates Cancer Cell Proliferation and Chemosensitivity in Triple-Negative Breast Cancer by Interacting with Each Other. J. Cell Biochem. 120 (9), 14860–14866. 10.1002/jcb.28747 [DOI] [PubMed] [Google Scholar]
- Liu L., Yu D., Shi H., Li J., Meng L. (2017). Reduced lncRNA Aim Enhances the Malignant Invasion of Triple-Negative Breast Cancer Cells Mainly by Activating Wnt/β-catenin/mTOR/PI3K Signaling. Pharmazie 72 (10), 599–603. 10.1691/ph.2017.7547 [DOI] [PubMed] [Google Scholar]
- Liu S., Wang Z., Liu Z., Shi S., Zhang Z., Zhang J., et al. (2018). miR-221/222 Activate the Wnt/β-Catenin Signaling to Promote Triple-Negative Breast Cancer. J. Mol. Cell Biol. 10 (4), 302–315. 10.1093/jmcb/mjy041 [DOI] [PubMed] [Google Scholar]
- Liu X., Song J., Kang Y., Wang Y., Chen A. (2020). Long Noncoding RNA SOX21-AS1 Regulates the Progression of Triple-Negative Breast Cancer through Regulation of miR-520a-5p/ORMDL3 axis. J. Cell Biochem. 121 (11), 4601–4611. 10.1002/jcb.29674 [DOI] [PubMed] [Google Scholar]
- Lu L., Xu H., Luo F., Liu X., Lu X., Yang Q., et al. (2016). Epigenetic Silencing of miR-218 by the lncRNA CCAT1, Acting via BMI1, Promotes an Altered Cell Cycle Transition in the Malignant Transformation of HBE Cells Induced by Cigarette Smoke Extract. Toxicol. Appl. Pharmacol. 304, 30–41. 10.1016/j.taap.2016.05.012 [DOI] [PubMed] [Google Scholar]
- Luo N., Zhang K., Li X., Hu Y. (2020). ZEB1 Induced-Upregulation of Long Noncoding RNA ZEB1-AS1 Facilitates the Progression of Triple Negative Breast Cancer by Binding with ELAVL1 to Maintain the Stability of ZEB1 mRNA. J. Cell Biochem. 121 (10), 4176–4187. 10.1002/jcb.29572 [DOI] [PubMed] [Google Scholar]
- Lv M., Xu P., Wu Y., Huang L., Li W., Lv S., et al. (2016). LncRNAs as New Biomarkers to Differentiate Triple Negative Breast Cancer from Non-triple Negative Breast Cancer. Oncotarget 7 (11), 13047–13059. 10.18632/oncotarget.7509 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma J., Lu W., Chen D., Xu B., Li Y. (2017). Role of Wnt Co-receptor LRP6 in Triple Negative Breast Cancer Cell Migration and Invasion. J. Cell Biochem. 118 (9), 2968–2976. 10.1002/jcb.25956 [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacDonald B. T., Tamai K., He X. (2009). Wnt/beta-catenin Signaling: Components, Mechanisms, and Diseases. Dev. Cell 17 (1), 9–26. 10.1016/j.devcel.2009.06.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maubant S., Tahtouh T., Brisson A., Maire V., Némati F., Tesson B., et al. (2018). LRP5 Regulates the Expression of STK40, a New Potential Target in Triple-Negative Breast Cancers. Oncotarget 9 (32), 22586–22604. 10.18632/oncotarget.25187 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mendell J. T. (2016). Targeting a Long Noncoding RNA in Breast Cancer. N. Engl. J. Med. 374 (23), 2287–2289. 10.1056/NEJMcibr1603785 [DOI] [PubMed] [Google Scholar]
- Merikhian P., Eisavand M. R., Farahmand L. (2021). Triple-negative Breast Cancer: Understanding Wnt Signaling in Drug Resistance. Cancer Cell Int. 21 (1), 419. 10.1186/s12935-021-02107-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miao Y., Zheng W., Li N., Su Z., Zhao L., Zhou H., et al. (2017). MicroRNA-130b Targets PTEN to Mediate Drug Resistance and Proliferation of Breast Cancer Cells via the PI3K/Akt Signaling Pathway. Sci. Rep. 7, 41942. 10.1038/srep41942 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohammadi Yeganeh S., Vasei M., Tavakoli R., Kia V., Paryan M. (2017). The Effect of miR-340 Over-expression on Cell-Cycle-Related Genes in Triple-Negative Breast Cancer Cells. Eur. J. Cancer Care (Engl) 26 (6). 10.1111/ecc.12496 [DOI] [PubMed] [Google Scholar]
- Moore H. C., Jordan L. B., Bray S. E., Baker L., Quinlan P. R., Purdie C. A., et al. (2010). The RNA Helicase P68 Modulates Expression and Function of the Δ133 Isoform(s) of P53, and Is Inversely Associated with Δ133p53 Expression in Breast Cancer. Oncogene 29 (49), 6475–6484. 10.1038/onc.2010.381 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mou E., Wang H. (2019). LncRNA LUCAT1 Facilitates Tumorigenesis and Metastasis of Triple-Negative Breast Cancer through Modulating miR-5702. Biosci. Rep. 39 (9). 10.1042/BSR20190489 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müller S., Glaß M., Singh A. K., Haase J., Bley N., Fuchs T., et al. (2019). IGF2BP1 Promotes SRF-dependent Transcription in Cancer in a m6A- and miRNA-dependent Manner. Nucleic Acids Res. 47 (1), 375–390. 10.1093/nar/gky1012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nie J., Jiang H. C., Zhou Y. C., Jiang B., He W. J., Wang Y. F., et al. (2019). MiR-125b Regulates the Proliferation and Metastasis of Triple Negative Breast Cancer Cells via the Wnt/β-Catenin Pathway and EMT. Biosci. Biotechnol. Biochem. 83 (6), 1062–1071. 10.1080/09168451.2019.1584521 [DOI] [PubMed] [Google Scholar]
- Padmanaban V., Krol I., Suhail Y., Szczerba B. M., Aceto N., Bader J. S., et al. (2019). E-cadherin Is Required for Metastasis in Multiple Models of Breast Cancer. Nature 573 (7774), 439–444. 10.1038/s41586-019-1526-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peiris-Pagès M., Bonuccelli G., Sotgia F., Lisanti M. P. (2018). Mitochondrial Fission as a Driver of Stemness in Tumor Cells: mDIVI1 Inhibits Mitochondrial Function, Cell Migration and Cancer Stem Cell (CSC) Signalling. Oncotarget 9 (17), 13254–13275. 10.18632/oncotarget.24285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phi L. T. H., Sari I. N., Yang Y. G., Lee S. H., Jun N., Kim K. S., et al. (2018). Cancer Stem Cells (CSCs) in Drug Resistance and Their Therapeutic Implications in Cancer Treatment. Stem Cells Int. 2018, 5416923. 10.1155/2018/5416923 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piasecka D., Braun M., Kordek R., Sadej R., Romanska H. (2018). MicroRNAs in Regulation of Triple-Negative Breast Cancer Progression. J. Cancer Res. Clin. Oncol. 144 (8), 1401–1411. 10.1007/s00432-018-2689-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pohl S. G., Brook N., Agostino M., Arfuso F., Kumar A. P., Dharmarajan A. (2017). Wnt Signaling in Triple-Negative Breast Cancer. Oncogenesis 6 (4), e310. 10.1038/oncsis.2017.14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poodineh J., Sirati-Sabet M., Rajabibazl M., Mohammadi-Yeganeh S. (2020). MiR-130a-3p Blocks Wnt Signaling Cascade in the Triple-Negative Breast Cancer by Targeting the Key Players at Multiple Points. Heliyon 6 (11), e05434. 10.1016/j.heliyon.2020.e05434 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodrigues de Bastos D., Nagai M. A. (2020). In Silico analyses Identify lncRNAs: WDFY3-AS2, BDNF-AS and AFAP1-AS1 as Potential Prognostic Factors for Patients with Triple-Negative Breast Tumors. PLoS One 15 (5), e0232284. 10.1371/journal.pone.0232284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ryu W. J., Lee J. D., Park J. C., Cha P. H., Cho Y. H., Kim J. Y., et al. (2020). Destabilization of β-catenin and RAS by Targeting the Wnt/β-Catenin Pathway as a Potential Treatment for Triple-Negative Breast Cancer. Exp. Mol. Med. 52 (5), 832–842. 10.1038/s12276-020-0440-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satriyo P. B., Bamodu O. A., Chen J. H., Aryandono T., Haryana S. M., Yeh C. T., et al. (2019). Cadherin 11 Inhibition Downregulates β-catenin, Deactivates the Canonical WNT Signalling Pathway and Suppresses the Cancer Stem Cell-like Phenotype of Triple Negative Breast Cancer. J. Clin. Med. 8 (2). 10.3390/jcm8020148 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scatena C., Roncella M., Di Paolo A., Aretini P., Menicagli M., Fanelli G., et al. (2018). Doxycycline, an Inhibitor of Mitochondrial Biogenesis, Effectively Reduces Cancer Stem Cells (CSCs) in Early Breast Cancer Patients: A Clinical Pilot Study. Front. Oncol. 8, 452. 10.3389/fonc.2018.00452 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi R., Wu P., Liu M., Chen B., Cong L. (2020). Knockdown of lncRNA PCAT6 Enhances Radiosensitivity in Triple-Negative Breast Cancer Cells by Regulating miR-185-5p/TPD52 Axis. Onco Targets Ther. 13, 3025–3037. 10.2147/OTT.S237559 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shin V. Y., Chen J., Cheuk I. W., Siu M. T., Ho C. W., Wang X., et al. (2019). Long Non-coding RNA NEAT1 Confers Oncogenic Role in Triple-Negative Breast Cancer through Modulating Chemoresistance and Cancer Stemness. Cell Death Dis. 10 (4), 270. 10.1038/s41419-019-1513-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song X., Liu Z., Yu Z. (2019). Lncrna nef is downregulated in triple negative breast cancer and correlated with poor prognosis. Acta Biochim. Biophys. Sin. (Shanghai) 51 (4), 386–392. 10.1093/abbs/gmz021 [DOI] [PubMed] [Google Scholar]
- Taipaleenmäki H., Farina N. H., van Wijnen A. J., Stein J. L., Hesse E., Stein G. S., et al. (2016). Antagonizing miR-218-5p Attenuates Wnt Signaling and Reduces Metastatic Bone Disease of Triple Negative Breast Cancer Cells. Oncotarget 7 (48), 79032–79046. 10.18632/oncotarget.12593 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang J., Zhong G., Zhang H., Yu B., Wei F., Luo L., et al. (2018). Lncrna dancr upregulates pi3k/akt signaling through activating serine phosphorylation of rxra. Cell Death Dis. 9 (12), 1167. 10.1038/s41419-018-1220-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang T., Cheng Y., She Q., Jiang Y., Chen Y., Yang W., et al. (2018). Long Non-coding RNA TUG1 Sponges miR-197 to Enhance Cisplatin Sensitivity in Triple Negative Breast Cancer. Biomed. Pharmacother. 107, 338–346. 10.1016/j.biopha.2018.07.076 [DOI] [PubMed] [Google Scholar]
- Tao W., Wang C., Zhu B., Zhang G., Pang D. (2019). Lncrna dancr contributes to tumor progression via targetting mir-216a-5p in breast cancer: lncrna dancr contributes to tumor progression. Biosci. Rep. 39 (4). 10.1042/BSR20181618 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taube J. H., Malouf G. G., Lu E., Sphyris N., Vijay V., Ramachandran P. P., et al. (2013). Epigenetic Silencing of microRNA-203 Is Required for EMT and Cancer Stem Cell Properties. Sci. Rep. 3, 2687. 10.1038/srep02687 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Telonis A. G., Rigoutsos I. (2018). Race Disparities in the Contribution of miRNA Isoforms and tRNA-Derived Fragments to Triple-Negative Breast Cancer. Cancer Res. 78 (5), 1140–1154. 10.1158/0008-5472.CAN-17-1947 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trzybulska D., Vergadi E., Tsatsanis C. (2018). miRNA and Other Non-coding RNAs as Promising Diagnostic Markers. EJIFCC 29 (3), 221–226. [PMC free article] [PubMed] [Google Scholar]
- Tu Z., Schmöllerl J., Cuiffo B. G., Karnoub A. E. (2019). Microenvironmental Regulation of Long Noncoding RNA LINC01133 Promotes Cancer Stem Cell-like Phenotypic Traits in Triple-Negative Breast Cancers. Stem Cells 37 (10), 1281–1292. 10.1002/stem.3055 [DOI] [PubMed] [Google Scholar]
- van Barele M., Heemskerk-Gerritsen B. A. M., Louwers Y. V., Vastbinder M. B., Martens J. W. M., Hooning M. J., et al. (2021). Estrogens and Progestogens in Triple Negative Breast Cancer: Do They Harm? Cancers (Basel) 13 (11). 10.3390/cancers13112506 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volovat S. R., Volovat C., Hordila I., Hordila D. A., Mirestean C. C., Miron O. T., et al. (2020). MiRNA and LncRNA as Potential Biomarkers in Triple-Negative Breast Cancer: A Review. Front. Oncol. 10, 526850. 10.3389/fonc.2020.526850 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang D., Huang J., Hu Z. (2012). RNA Helicase DDX5 Regulates microRNA Expression and Contributes to Cytoskeletal Reorganization in Basal Breast Cancer Cells. Mol. Cell Proteomics 11 (2), M111–M011932. 10.1074/mcp.M111.011932 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang D. Y., Gendoo D. M. A., Ben-David Y., Woodgett J. R., Zacksenhaus E. (2019). A Subgroup of microRNAs Defines PTEN-Deficient, Triple-Negative Breast Cancer Patients with Poorest Prognosis and Alterations in RB1, MYC, and Wnt Signaling. Breast Cancer Res. 21 (1), 18. 10.1186/s13058-019-1098-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang K., Li X., Song C., Li M. (2018). Lncrna awpph promotes the growth of triple-negative breast cancer by up-regulating frizzled homolog 7 (fzd7). Biosci. Rep. 38 (6). 10.1042/BSR20181223 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Wang L., Wang R., Ye Z., Wang Y., Li X., Chen W., et al. (2018). PVT1 Affects EMT and Cell Proliferation and Migration via Regulating P21 in Triple-Negative Breast Cancer Cells Cultured with Mature Adipogenic Medium. Acta Biochim. Biophys. Sin. (Shanghai) 50 (12), 1211–1218. 10.1093/abbs/gmy129 [DOI] [PubMed] [Google Scholar]
- Wang Y., Zhang Z., Wang J. (2018). MicroRNA-384 Inhibits the Progression of Breast Cancer by Targeting ACVR1. Oncol. Rep. 39 (6), 2563–2574. 10.3892/or.2018.6385 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilusz J. E., Sunwoo H., Spector D. L. (2009). Long Noncoding RNAs: Functional Surprises from the RNA World. Genes Dev. 23 (13), 1494–1504. 10.1101/gad.1800909 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu J., Shuang Z., Zhao J., Tang H., Liu P., Zhang L., et al. (2018). Linc00152 Promotes Tumorigenesis by Regulating DNMTs in Triple-Negative Breast Cancer. Biomed. Pharmacother. 97, 1275–1281. 10.1016/j.biopha.2017.11.055 [DOI] [PubMed] [Google Scholar]
- Wu R., Zhao B., Ren X., Wu S., Liu M., Wang Z., et al. (2020). MiR-27a-3p Targeting GSK3β Promotes Triple-Negative Breast Cancer Proliferation and Migration through Wnt/β-Catenin Pathway. Cancer Manag. Res. 12, 6241–6249. 10.2147/CMAR.S255419 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao C., Wu C. H., Hu H. Z. (2016). LncRNA UCA1 Promotes Epithelial-Mesenchymal Transition (EMT) of Breast Cancer Cells via Enhancing Wnt/beta-Catenin Signaling Pathway. Eur. Rev. Med. Pharmacol. Sci. 20 (13), 2819–2824. [PubMed] [Google Scholar]
- Xiao F., Jiao H., Wu D., Zhang Z., Li S., Guo J. (2021). LINC01234 Aggravates Cell Growth and Migration of Triple-Negative Breast Cancer by Activating the Wnt Pathway. Environ. Toxicol. 36, 1999–2012. 10.1002/tox.23318 [DOI] [PubMed] [Google Scholar]
- Xu J., Prosperi J. R., Choudhury N., Olopade O. I., Goss K. H. (2015). β-Catenin Is Required for the Tumorigenic Behavior of Triple-Negative Breast Cancer Cells. PLoS One 10 (2), e0117097. 10.1371/journal.pone.0117097 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu S. T., Xu J. H., Zheng Z. R., Zhao Q. Q., Zeng X. S., Cheng S. X., et al. (2017). Long non-coding rna anril promotes carcinogenesis via sponging mir-199a in triple-negative breast cancer. Biomed. Pharmacother. 96, 14–21. 10.1016/j.biopha.2017.09.107 [DOI] [PubMed] [Google Scholar]
- Yang L., Wu X., Wang Y., Zhang K., Wu J., Yuan Y. C., et al. (2011). FZD7 Has a Critical Role in Cell Proliferation in Triple Negative Breast Cancer. Oncogene 30 (43), 4437–4446. 10.1038/onc.2011.145 [DOI] [PubMed] [Google Scholar]
- Yang W., Cui G., Ding M., Yang M., Dai D. (2020). MicroRNA-124-3p.1 Promotes Cell Proliferation through Axin1-dependent Wnt Signaling Pathway and Predicts a Poor Prognosis of Triple-Negative Breast Cancer. J. Clin. Lab. Anal. 34 (7), e23266. 10.1002/jcla.23266 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yao H., Ashihara E., Maekawa T. (2011). Targeting the Wnt/β-Catenin Signaling Pathway in Human Cancers. Expert Opin. Ther. Targets 15 (7), 873–887. 10.1517/14728222.2011.577418 [DOI] [PubMed] [Google Scholar]
- Yin L., Duan J. J., Bian X. W., Yu S. C. (2020). Triple-negative Breast Cancer Molecular Subtyping and Treatment Progress. Breast Cancer Res. 22 (1), 61. 10.1186/s13058-020-01296-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin S., Xu L., Bonfil R. D., Banerjee S., Sarkar F. H., Sethi S., et al. (2013). Tumor-initiating Cells and FZD8 Play a Major Role in Drug Resistance in Triple-Negative Breast Cancer. Mol. Cancer Ther. 12 (4), 491–498. 10.1158/1535-7163.MCT-12-1090 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu L., Wang C., Pan F., Liu Y., Ren X., Zeng H., et al. (2019). HePTP Promotes Migration and Invasion in Triple-Negative Breast Cancer Cells via Activation of Wnt/β-Catenin Signaling. Biomed. Pharmacother. 118, 109361. 10.1016/j.biopha.2019.109361 [DOI] [PubMed] [Google Scholar]
- Yuan C., Luo X., Duan S., Guo L. (2020). Long Noncoding RNA LINC00115 Promotes Breast Cancer Metastasis by Inhibiting miR-7. FEBS Open Bio 10 (7), 1230–1237. 10.1002/2211-5463.12842 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang K., Liu P., Tang H., Xie X., Kong Y., Song C., et al. (2018). AFAP1-AS1 Promotes Epithelial-Mesenchymal Transition and Tumorigenesis through Wnt/β-Catenin Signaling Pathway in Triple-Negative Breast Cancer. Front. Pharmacol. 9, 1248. 10.3389/fphar.2018.01248 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Zhang T., Hu H., Yan G., Wu T., Liu S., Chen W., et al. (2019). Long Non-coding RNA and Breast Cancer. Technol. Cancer Res. Treat. 18, 1533033819843889. 10.1177/1533033819843889 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y., Wang X. (2020). Targeting the Wnt/β-Catenin Signaling Pathway in Cancer. J. Hematol. Oncol. 13 (1), 165. 10.1186/s13045-020-00990-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng A., Song X., Zhang L., Zhao L., Mao X., Wei M., et al. (2019). Long Non-coding RNA LUCAT1/miR-5582-3p/TCF7L2 axis Regulates Breast Cancer Stemness via Wnt/β-Catenin Pathway. J. Exp. Clin. Cancer Res. 38 (1), 305. 10.1186/s13046-019-1315-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zuo Y., Li Y., Zhou Z., Ma M., Fu K. (2017). Long Non-coding RNA MALAT1 Promotes Proliferation and Invasion via Targeting miR-129-5p in Triple-Negative Breast Cancer. Biomed. Pharmacother. 95, 922–928. 10.1016/j.biopha.2017.09.005 [DOI] [PubMed] [Google Scholar]