

Abstract
The cell wall of Gram-negative bacteria consists of an inner (cytoplasmic) and outer membrane (OM), separated by a thin peptidoglycan layer. Throughout growth, the outer membrane can bleb to form spherical outer membrane vesicles (OMVs). These OMVs are involved in numerous cellular functions including cargo delivery to host cells and communication with bacterial cells. Recently, the therapeutic potential of OMVs has begun to be explored, including their use as vaccines and drug delivery vehicles. Although OMVs are derived from the OM, it has long been appreciated that the lipid and protein cargo of the OMV differs, often significantly, from that of the OM. More recently, evidence that bacteria can release multiple types of OMVs has been discovered, and evidence exists that size can impact the mechanism of their uptake by host cells. However, studies in this area are limited by difficulties in efficiently separating the heterogeneously sized OMVs. Density gradient centrifugation (DGC) has traditionally been used for this purpose; however, this technique is time-consuming and difficult to scale-up. Size exclusion chromatography (SEC), on the other hand, is less cumbersome and lends itself to the necessary future scale-up for therapeutic use of OMVs. Here, we describe a SEC approach that enables reproducible separation of heterogeneously sized vesicles, using as a test case, OMVs produced by Aggregatibacter actinomycetemcomitans, which range in diameter from less than 150 nm to greater than 350 nm. We demonstrate separation of "large" (350 nm) OMVs and "small" (<150 nm) OMVs, verified by dynamic light scattering (DLS). We recommend SEC-based techniques over DGC-based techniques for separation of heterogeneously sized vesicles due to its ease of use, reproducibility (including user-to-user), and possibility for scale-up.
Introduction
Gram-negative bacteria release vesicles derived from their outer membrane, so-called outer membrane vesicles (OMVs), throughout growth. These OMVs play important roles in cell-to-cell communication, both between bacteria and host as well as between bacterial cells, by carrying a number of important biomolecules, including DNA/RNA, proteins, lipids, and peptidoglycans1,2. In particular, the role of OMVs in bacterial pathogenesis has been extensively studied due to their enrichment in certain virulence factors and toxins3, 4, 5, 6, 7, 8, 9, 10, 11.
OMVs have been reported to range in size from 20 to 450 nm, depending on the parent bacteria and the growth stage, with several types of bacteria releasing heterogeneously sized OMVs8, 12, 13, 14, which also differ in their protein composition and mechanism of host cell entry12. H. pylori released OMVs ranging in diameter from 20 to 450 nm, with the smaller OMVs containing a more homogeneous protein composition than the larger OMVs. Importantly, the two populations of OMVs were observed to be internalized by host cells via different mechanisms12. In addition, we have demonstrated that Aggregatibacter actinomycetemcomitans releases a population of small (<150 nm) OMVs along with a population of large (>350 nm) OMVs, with the OMVs containing a significant amount of a secreted protein toxin, leukotoxin (LtxA)15. While the role of OMV heterogeneity in cellular processes is clearly important, technical difficulties in separating and analyzing distinct populations of vesicles has limited these studies.
In addition to their importance in bacterial pathogenesis, OMVs have been proposed for use in a number of biotechnological applications, including as vaccines and drug delivery vehicles16, 17, 18, 19, 20. For their translational use in such approaches, a clean and monodisperse preparation of vesicles is required. Thus, effective and efficient methods of separation are necessary.
Most commonly, density gradient centrifugation (DGC) is used to separate heterogeneously sized vesicle populations from cellular debris, including flagellae and secreted proteins21; the method has also been reported as an approach to separate heterogeneously sized OMV subpopulations12, 13, 14. However, DGC is time-consuming, inefficient, and highly variable user-to-user22 and is, therefore, not ideal for scale-up. In contrast, size exclusion chromatography (SEC) represents a scalable, efficient, and consistent approach to purify OMVs21, 23, 24. We have found that a long (50-cm), gravity-flow, SEC column, filled with gel filtration medium is sufficient for efficiently purifying and separating subpopulations of OMVs. Specifically, we used this approach to separate A. actinomycetemcomitans OMVs into "large" and "small" subpopulations, as well as to remove protein and DNA contamination. Purification was completed in less than 4 h, and complete separation of the OMV subpopulations and removal of debris was accomplished.
Protocol
1. Preparation of buffers
To prepare the ELISA wash buffer, add 3.94 g Tris-base, 8.77 g NaCl, and 1 g bovine serum albumin (BSA) to 1 L of deionized (DI) water. Add 500 μL polysorbate-20. Adjust the pH to 7.2 using HCl or NaOH.
To prepare the blocking buffer, add 3.94 g Tris-base, 8.77 g NaCl, and 10 g BSA. Add 500 μL polysorbate-20 to 1 L of DI water. Adjust the pH to 7.2 using HCl or NaOH.
-
To prepare the elution buffer (PBS), add 8.01 g NaCl, 2.7 g KCl, 1.42 g Na2HPO4, and 0.24 g KH2PO4 to 1 L DI water. Adjust the pH to 7.4 using HCl or NaOH.
NOTE: A 10x solution of this buffer can be made and diluted with DI water as needed.
2. Preparation of OMV sample
Grow A. actinomycetemcomitans cells to the late exponential phase (optical density at 600 nm of 0.7). Pellet the cells by centrifuging twice at 10,000 x g at 4 °C for 10 min. Filter the supernatant through a 0.45 μm filter.
Concentrate the bacteria-free supernatant using 50 kDa-molecular weight cut-off filters. Ultracentrifuge the concentrated solution at 105,000 x g at 4 °C for 30 min.
Resuspend the pellet in PBS and ultracentrifuge again (105,000 x g at 4 °C for 30 min.) Resuspend the pellet in 2 mL of PBS.
3. Packing the S-1000 column
Mix the stock bottle of gel filtration medium with a glass stir rod and pour out into a glass bottle the volume necessary to fill the column, plus approximately 50% excess (about 135 mL). Let these beads sit until they have settled, and then decant off the excess liquid. Resuspend the beads in elution buffer, so that the final solution is approximately 70% (by volume) gel, 30% buffer. Degas the solution under vacuum.
Mount the glass column vertically using a ring stand and fill with elution buffer to wet the walls of the column. Drain the buffer until there is only about 1 cm of buffer remaining in the column.
Without creating bubbles, carefully pipette beads into the column, filling the column to the top. Continue to drain excess buffer throughout this process. Be sure to not let the beads settle completely before adding additional beads to the top of the column. The column should be packed to a height of about 2 cm below the bottom of the column reservoir.
4. Loading the sample and collecting fractions
Degas the elution buffer under vacuum. Wash the column with two column-volumes (180 mL) of elution buffer.
Allow the remaining buffer to fully enter the column. Once the buffer has reached the top of the gel layer, carefully pipette a 2-mL sample containing OMVs (at a lipid concentration of approximately 100 - 200 nmol/L) onto the surface of the beads, being careful not to disturb any of the beads at the top of the column. Allow the sample to fully enter the gel, that is, when no liquid remains above the gel layer.
Carefully and slowly add elution buffer on top of the gel column. Do not disturb the top layer of the gel, as this will cause sample dilution.
Place a single 50-mL tube under the column and open the column. Collect the first 20 mL of the eluent. Add additional elution buffer to the top of the column, carefully, as needed to ensure the column is never dry.
-
Place a series of 1.5 mL tubes under the column. Start the column and collect a series of 1-mL samples in each tube. As the samples are being collected, continue to add elution buffer to the top of the column, as necessary. Repeat until 96 fractions have been collected. Stop the column.
NOTE: The samples should be stored at −20 °C for long-term storage or 4 °C for short-term storage until further analysis.
To clean the column, run one column volume (90 mL) of 0.1 M NaOH through the column. Run two column volumes (180 mL) of elution buffer through the column.
5. Sample analysis
To measure the lipid concentration in each fraction, pipette 50 μL of each fraction into a single well of a 96-well plate. To each well, add 2.5 μL of lipophilic dye. Incubate for 15 s. Measure the fluorescence intensity on a plate reader with an excitation wavelength of 515 nm and an emission wavelength of 640 nm. To calculate the fraction of all lipid in each sample, sum all of the emission intensities and divide each individual intensity by the total.
- To measure the concentration of a particular protein, pipette 100 μL of each fraction into a single well of an ELISA immuno-plate. Incubate at 25 °C for 3 h.
- Decant the samples. Add 200 μL of ELISA wash buffer to each well and decant. Repeat four times for a total of five washes.
- Add 200 μL of blocking buffer to each well and incubate for 1 h at 25 °C. Decant.
- Incubate plates with 100 μL blocking buffer plus primary antibody (1:10,000 for purified antibody; 1:10 for unpurified antibody) overnight at 4 °C. Decant.
- Add 200 μL of ELISA wash buffer to each well and decant. Repeat four times for a total of five washes.
- Add 100 μL of ELISA wash buffer plus secondary antibody (1:30,000) to each well. Incubate for 1 h at 25 °C.
- Add 200 μL of ELISA wash buffer to each well and decant. Repeat four times for a total of five washes.
- Add 100 μL of the 3,3',5,5'-tetramethylbenzidine (TMB) one-step solution and incubate for 15-30 min or until a blue color develops. Stop the TMB reaction with 50 μL of the stopping solution.
- On a plate reader, read the absorbance of each well at a wavelength of 450 nm.
To measure the total protein concentration, record the absorbance at a wavelength of 280 nm (A280) of each fraction, using a UV-vis spectrophotometer.
A schematic of the protocol is shown in Figure 1.
Figure 1: Schematic of SEC procedure.

The column is packed with degassed gel filtration medium carefully to avoid bubbles and discontinuities, then washed with two column volumes of elution buffer. Next, the sample is carefully pipetted onto the top of the gel, without disrupting gel packing. The column is opened and run until the sample completely enters the gel. At this point, buffer is placed on the top of the column, and the first 20 mL of eluate is collected. Next, a series of 1-mL fractions is collected. These fractions are then placed in a 96-well plate or 96-well immuno-plate for analysis of lipid and protein content.
Representative Results
Figure 2 shows representative results from this method. OMVs produced by A. actinomycetemcomitans strain JP2 were first purified from the culture supernatant using ultracentrifugation15. We previously found that this strain produces two populations of OMVs, one with diameters of about 300 nm and one with diameters of about 100 nm15. To separate these OMV populations, we purified the samples using the SEC protocol described above. Each fraction was analyzed for lipid content using the lipophilic dye and for toxin (LtxA) content using enzyme-linked immunosorbent assay (ELISA) or an immunoblot. The lipid and toxin concentrations are reported as percentages, where "%lipid" indicates what percentage of the total lipid content of the sample is in each fraction and "%toxin" indicates what percentage of the total toxin content of the sample is in each fraction.
Figure 2: Representative results.
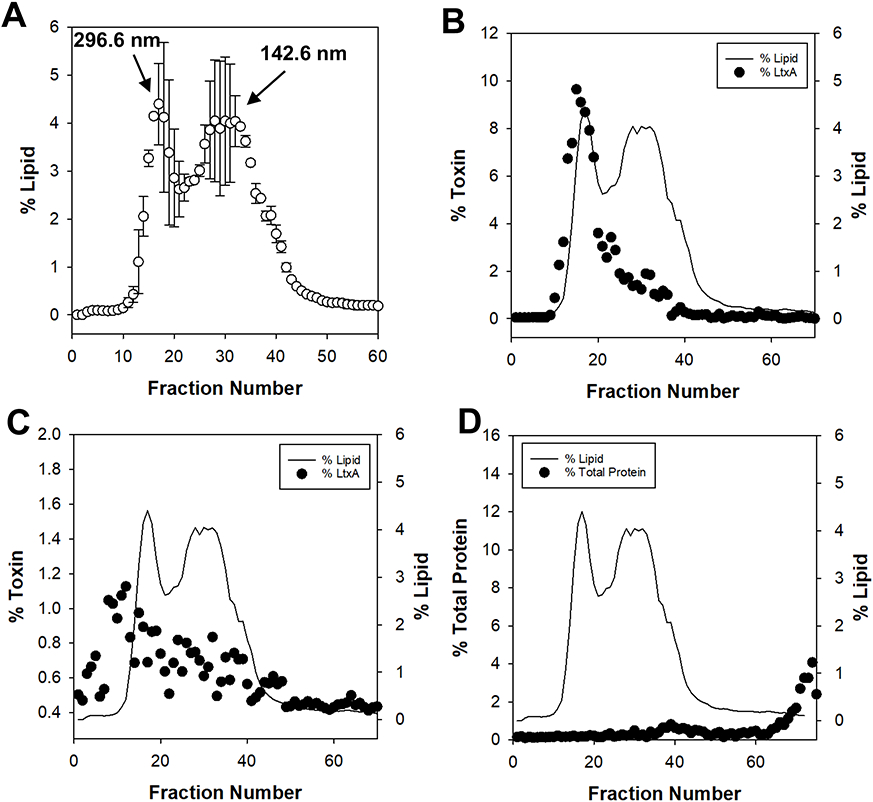
A. actinomycetemcomitans JP2 OMVs were run through the SEC column and each fraction was analyzed for lipid content using a lipophilic dye and toxin (LtxA) content using a monoclonal antibody. (A) The average lipid content of each fraction reported as a percentage of the total lipid content, from three trials. Each data point represents the mean ± standard deviation. (B) The LtxA content of each fraction, reported as a percentage of the total LtxA content, as measured by ELISA with a monoclonal anti-LtxA antibody. (C) The LtxA content of each fraction, reported as a percentage of the total LtxA content, as measured by an immunoblot with a monoclonal anti-LtxA antibody. (D) The total protein content of each fraction, reported as a percentage of the total protein content, as measured by A280. Some of the data are reproduced from Chang et al.26 with permission from John Wiley and Sons Ltd.
Figure 2A shows the averaged lipophilic dye results with standard deviations from three separate purifications, each performed by a different user, demonstrating the reproducibility of this technique. Two distinct lipid peaks are observed, corresponding to "large" OMVs (fraction number 13) and "small" OMVs (fraction number 25). We confirmed the size of the OMVs in these peaks using dynamic light scattering (DLS) and found the mean diameters of the OMVs in fractions 13 and 25 to be 296.6 nm and 142.6 nm, respectively, as shown in Fig. 2A. In comparison, the mean diameter of the OMV sample after ultracentrifugation but before SEC purification was previously found to be 161.0 nm15.
In Figure 2B, the amount of LtxA in each fraction, obtained using ELISA with a monoclonal antibody against LtxA25, is shown overlaid on the lipid concentration from panel A. This technique demonstrates that the toxin is associated primarily with one subpopulation of OMVs. Figure 2C shows the amount of LtxA in each fraction, measured using an immunoblot technique with the same anti-LtxA monoclonal antibody25, overlaid on the lipid concentration from panel A. While the overall trend is similar to what is observed in Figure 2B, the immunoblot approach is much less sensitive than the ELISA technique, resulting in noisier profiles. Figure 2D shows the percentage of the total protein concentration in each fraction, measured using the A280, overlaid on the lipid concentration profile. This panel demonstrates that SEC is able to remove significant amounts of free proteins from the OMV preparations, as evidenced by the high A280 values in fractions greater than 60. (In fact, most of the protein is found in these fractions, which do not contain OMVs, demonstrating that a large amount of protein co-purifies with the OMVs.) In addition, this result shows that the total protein concentration does not necessarily correlate with the concentration of specific proteins. In the case of A. actinomycetemcomtians OMVs, LtxA associates primarily with the population of larger OMVs, while more of the total protein associates with the smaller OMVs.
Together, these representative results demonstrate a number of important features of the SEC protocol for OMV purification. First, the technique is highly reproducible, even between users. Second, the use of a lipophilic dye to detect OMVs in each fraction is a simple and reliable method. Third, to detect specific protein concentrations, ELISA is more robust than an immunoblot. Fourth, SEC is able to remove large amounts of impurities, including proteins and nucleic acids.
Discussion
Here, we have provided a protocol for the simple, fast, and reproducible separation of bacterial OMV subpopulations. Although the technique is relatively straight-forward, there are some steps that must be performed extremely carefully to ensure that efficient separation occurs in the column. First, it is essential that the gel be loaded into the column carefully and slowly to avoid air bubbles. We have observed that leaving the gel at room temperature for several hours before loading the column allows the gel to equilibrate and minimizes bubble formation within the column. When the gel is pipetted into the column, it should be carefully pipetted along the side of the column to minimize turbulence. At all times during loading, excess buffer should be maintained in the column to avoid discontinuities in the settled gel. If a disjunction should occur, add more buffer and pipette up and down to resuspend the gel.
Similarly, loading the column with sample is critically important. Because the sample will become diluted as it passes through the column, the loaded sample should be sufficiently concentrated before separation by SEC. For the A. actinomycetemcomitans OMVs, we have found that a 1-mL sample containing approximately 100-200 nmol/L of lipids is ideal. After the sample is loaded carefully at the top of the column without disrupting the gel layer, the column should be run just until the sample fully enters the gel column. At this point, the column should be stopped so that a layer of buffer can be carefully added to the top of the gel. It is helpful to load only a small volume (~ 1 mL) of buffer, ensuring that the gel layer is not disrupted. Once the sample has been run further into the gel, buffer can be added in larger volumes and the concern with disrupting the gel layer is less of an issue. The column can be reused multiple times, as long as it is maintained in a fully hydrated state and cleaned well (Step 4.6) between runs.
All OMV purification procedures follow the same initial steps that include bacterial growth, removal of bacterial cells, and OMV isolation27. Although this "crude" preparation has commonly been used in OMV studies28, it is becoming increasingly apparent that a subsequent purification step is necessary to remove co-precipitating proteins and other contaminants, as well as to separate OMV subpopulations. In OMV studies, this purification step is commonly completed using density gradient centrifugation. In the eukaryotic extracellular vesicle field, the use of SEC to separate vesicle populations and to remove contaminants is increasing in importance, as it is simpler, faster, and less expensive than DGC29. In addition, SEC has the advantage of being possible to automate, unlike DGC29. Thus, while DGC remains the "gold standard" of vesicle isolation in the bacterial OMV field, we propose that the numerous advantages of SEC make it an extremely useful, if not better, method of OMV purification than DGC. In this work, we have demonstrated that a 1.5 x 50 cm column of Sephacryl S-1000 is capable of separating two subpopulations of OMVs. We have also observed that the approach is capable of removing nucleic acids and free proteins from the OMV solution. Previous reports have found SEC to be able to remove free LPS from OMV preparations, as well28.
In conclusion, we propose that SEC holds much promise in the purification of bacterial vesicles. While we have demonstrated the ability of the technique to separate subpopulations of OMVs produced by a specific bacterium (A. actinomycetemcomitans), we anticipate that the technique will be found to be extremely valuable in analyzing other bacterial vesicle samples, as it sees additional use. In particular, as the biotechnological applications of OMVs increase, the need for consistent and pure vesicle preparations will also increase; SEC is a promising method for these applications.
Materials
| Name | Company | Catalog Number | Comments |
|---|---|---|---|
| 1-Step Ultra TMB-ELISA | Thermo Scientific | 34028 | |
| Amicon 50 kDa filters | Millipore Sigma | UFC905024 | |
| Bovine Serum Albumin (BSA) | Fisher Scientific | BP9704-100 | |
| ELISA Immuno plates | Thermo Scientific | 442404 | |
| FM 4-64 | Thermo Scientific | T13320 | 1.5 x 50 cm |
| Glass Econo-Column | BioRad | 7371552 | |
| Infinite 200 Pro plate Reader | Tecan | ||
| Potassium Chloride (KCl) | Amresco (VWR) | 0395-500G | |
| Potassium Phosphate Monobasic Anhydrous (KH2PO4) | Amresco (VWR) | 0781-500G | |
| Sephacryl S-1000 Superfine | GE Healthcare | 17-0476-01 | |
| Sodium Chloride (NaCl) | Fisher Chemical | S271-3 | |
| Sodium Phosphate Dibasic Anhydrous (Na2HPO4) | Amresco (VWR) | 0404-500G | |
| Tris Base | VWR | 0497-1KG | |
| Tween(R) 20 | Acros Organics | 23336-2500 |
Acknowledgments
This work was funded by the National Science Foundation (1554417) and National Institutes of Health (DE027769).
Footnotes
A complete version of this article that includes the video component is available at http://dx.doi.org/10.3791/62429.
Disclosures
The authors have no conflicts of interest to report.
References
- 1.Kuehn MJ, Kesty NC Bacterial outer membrane vesicles and the host-pathogen interaction. Genes and Development. 19, 2645–2655 (2005). [DOI] [PubMed] [Google Scholar]
- 2.Kulp A, Kuehn MJ Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annual Reviews Microbiology. 64, 163–184 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kato S, Kowashi Y, Demuth DR Outer membrane-like vesicles secreted by Actinobacillus actinomycetemcomitans are enriched in leukotoxin. Microbial Pathogenesis. 32 (1), 1–13 (2002). [DOI] [PubMed] [Google Scholar]
- 4.Nice JB et al. Aggregatibacter actinomycetemcomitans leukotoxin is delivered to host cells in an LFA-1-independent manner when associated with outer membrane vesicles. Toxins. 10 (10), 414 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Haurat MF et al. Selective sorting of cargo proteins into bacterial membrane vesicles. Journal of Biological Chemistry. 286 (2), 1269–1276 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Horstman AL, Kuehn MJ Enterotoxigenic Escherichia coli secretes active heat-labile enterotoxin via outer membrane vesicles. The Journal of Biological Chemistry. 275 (17), 12489–12496 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wai SN et al. Vesicle-mediated export and assembly of pore-forming oligomers of the Enterobacterial ClyA cytotoxin. Cell. 115, 25–35 (2003). [DOI] [PubMed] [Google Scholar]
- 8.Balsalobre C et al. Release of the Type I secreted α-haemolysin via outer membrane vesicles from Escherichia coli. Molecular Microbiology. 59 (1), 99–112 (2006). [DOI] [PubMed] [Google Scholar]
- 9.Donato GM et al. Delivery of Bordetella pertussis adenylate cyclase toxin to target cells via outer membrane vesicles. FEBS Letters. 586, 459–465 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kim YR et al. Outer membrane vesicles of Vibrio vulnificus deliver cytolysin-hemolysin VvhA into epithelial cells to induce cytotoxicity. Biochemical and Biophysical Research Communications. 399, 607–612 (2010). [DOI] [PubMed] [Google Scholar]
- 11.Maldonado R, Wei R, Kachlany SC, Kazi M, Balashova NV Cytotoxic effects of Kingella kingae outer membrane vesicles on human cells. Microbial Pathogenesis. 51 (1-2), 22–30 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Turner L et al. Helicobacter pylori outer membrane vesicle size determines their mechanisms of host cell entry and protein content. Frontiers in Immunology. 9, 1466 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zavan L, Bitto NJ, Johnston EL, Greening DW, Kaparakis-Liaskos M Helicobacter pylori growth stage determines the size, protein composition, and preferential cargo packaging of outer membrane vesicles. Proteomics. 19 (1-2), e1800209 (2019). [DOI] [PubMed] [Google Scholar]
- 14.Rompikuntal PK et al. Perinuclear localization of internalized outer membrane vesicles carrying active cytolethal distending toxin from Aggregatibacter actinomycetemcomitans. Infections and Immunity. 80 (1), 31–42 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Nice JB et al. Aggregatibacter actinomycetemcomitans leukotoxin is delivered to host cells in an LFA-1-independent manner when associated with outer membrane vesicles. Toxins. 10 (10), 414 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Stevenson TC et al. Immunization with outer membrane vesicles displaying conserved surface polysaccharide antigen elicits broadly antimicrobial antibodies. Proceedings of the National Academy of Sciences. 115 (14), E3106–E3115 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gujrati V et al. Bioengineered bacterial outer membrane vesicles as cell-specific drug-delivery vehicles for cancer therapy. ACS Nano. 8 (2), 1525–1537 (2014). [DOI] [PubMed] [Google Scholar]
- 18.Huang W et al. Development of novel nanoantibiotics using an outer membrane vesicle-based drug efflux mechanism. Journal of Controlled Release. 317, 1–22 (2020). [DOI] [PubMed] [Google Scholar]
- 19.Chen DJ et al. Delivery of foreign antigens by engineered outer membrane vesicle vaccines. Proceedings of the National Academy of Sciences. 107 (7), 3099–3104 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Chen L et al. Outer membrane vesicles displaying engineered glycotopes elicit protective antibodies. Proceedings of the National Academy of Sciences. 113 (26), E3609–E3618 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Singorenko PD et al. Isolation of membrane vesicles from prokaryotes: A technical and biological comparison reveals heterogeneity. Journal of Extracellular Vesicles. 6 (1), 1324731 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zeringer E, Barta T, Li M, Vlassov AV Strategies for isolation of exosomes. Cold Spring Harbor Protocols. 2015 (4), 319–323 (2015). [DOI] [PubMed] [Google Scholar]
- 23.Benedikter BJ et al. Ultrafiltration combined with size exclusion chromatography efficiently isolates extracellular vesicles from cell culture media for compositional and functional studies. Science Reports. 7 (1), 15297 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mol EA, Goumans MJ, Doevendans PA, Sluijter JPG, Vader P Higher functionality of extracellular vesicles isolated using size-exclusion chromatography compared to ultracentrifugation. Nanomedicine. 13 (6), 2061–2065 (2017). [DOI] [PubMed] [Google Scholar]
- 25.Lally ET, Golub EE, Kieba IR Identification and immunological characterization of the domain of Actinobacillus actinomycetemcomitans leukotoxin that determines its specificity for human target cells. Journal of Biological Chemistry. 269 (49), 31289–31295 (1994). [PubMed] [Google Scholar]
- 26.Chang EH, Giaquinto P, Huang J, Balashova NV, Brown AC Epigallocatechin gallate inhibits leukotoxin release by Aggregatibacter actinomycetemcomitans by promoting association with the bacterial membrane. Molecular Oral Microbiology. 35 (1), 29–39 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Klimentová J, Stulík J Methods of isolation and purification of outer membrane vesicles from gram-negative bacteria. Microbiological Research. 170, 1–9 (2015). [DOI] [PubMed] [Google Scholar]
- 28.Dauros Singorenko P et al. Isolation of membrane vesicles from prokaryotes: a technical and biological comparison reveals heterogeneity. Journal of Extracellular Vesicles. 6 (1), 1324731 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Monguió-Tortajada M, Gálvez-Montón C, Bayes-Genis A, Roura S, Borràs FE Extracellular vesicle isolation methods: rising impact of size-exclusion chromatography. Cellular and Molecular Life Sciences. 76 (12), 2369–2382 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]