

Abstract
Calreticulin (CRT) is located in the endoplasmic reticulum (ER), it helps proteins fold correctly inside the ER, and acts as a modulator of Ca2+ homeostasis. Aberrant expression of CRT is implicated in several cancer types, qualifying CRT as a potential therapeutic target. However, it remains unclear how CRT affects specific oncogenic pathways. In this study, we used histone deacetylase inhibitors (HDACis) to establish drug-resistant liver cancer cells and further analyzed the molecular mechanism of development of drug resistance in those cells. The 2D gel electrophoresis and RT-PCR data showed that CRT was downregulated in HDACis-resistant cells by comparing with HA22T parental cells. We previously elucidated the development of drug-resistance in HCC cells via activation of PP1-eIF2α pathway, but not via ER stress pathway. Here, we show that thapsigargin induced ER stress through mechanism other than ER stress downstream protein GRP78-PERK to regulate CRT expression in HDACis-R cells. Moreover, the expression level of CRT was not the main cause of apoptosis in HDACis-resistant cells. Mechanistic studies identified the apoptosis factors in the nucleus—the HDACis-mediated overexpression of CRT, CRT translocation to the cell nucleus, and reduced CaM/CaMKII/CREB pathway—that led to chemosensitivity in HDACis-R HCC cells.
Keywords: calreticulin, hepatocellular carcinoma, HDACis-resistant cells, chemosensitivity
INTRODUCTION
Several chemotherapeutic drugs are developed and approved by the Food and Drug Administration (FDA) in recent years against liver cancer, such as cabozantinib-s-malate [1], ramucirumab [2], pembrolizumab [3], lenvatinib mesylate [3], sorafenib tosylate [4], nivolumab [5], regorafenib [6], etc. Despite advances chemotherapeutic agents for treatment of liver cancer, development of drug resistance has remained the major impediment in the treatment of patients.
Most of the chemotherapeutic drugs approved for the treatment of malignant liver tumors inhibit specific tyrosine kinases, such as vascular endothelial growth factor receptors (VEGFR) [4, 7], Raf kinase [4, 7], platelet-derived growth factor (PDGF) [7], and PD-1 receptor [6]. However, epigenetic mechanisms are found to play a key role in cancer development [8]. Acetylation of lysine residues is one of the epigenetic histone modification events. Histone acetyltransferase (HATs)-mediated histone acetylation leads to chromatin decondensation and binding of transcriptional machinery to the DNA [9]. In contrast, histone deacetylation by histone deacetylases (HDACs) favors a more condensed, transcriptionally inactive chromatin structure [9]. These alterations affect both gene transcription and DNA repair. Thus, the development of epigenetic therapeutics is a potential and attractive domain in HCC.
In our previous studies, we used HDAC inhibitors (HDACis) to treat liver cancer cell. Moreover, we generated HDACis-resistant cells (HDACis-R cells) by treating HA22T cell line with apicidin, a novel HDAC-inhibitor, and investigated the molecular mechanisms of developing resistance in liver cancer cells [10–12]. The expression of phosphatase and tensin homolog (PTEN) [11], a tumor-suppressor gene, was decreased, while that of members of phosphoinositide 3-kinase (PI3K)-Akt pathway (a pro-survival protein) [10, 11] and EMT-like signaling (as a marker of metastatic) [12] was increased in HDACis-R cells than that in HDAC-sensitive cells. Therefore, these studies showed that HDACis act as potential therapeutic agents in HCC and elucidated the development of resistance.
Here, we aimed to further investigate the expression of calreticulin (CRT) that was differential between HA22T and HDACis-R cells [13]. CRT is a Ca2+-binding chaperone that helps proteins fold correctly in the lumen of endoplasmic reticulum (ER) and also functions as a multi-functional protein in non-ER activities, such as calcium homeostasis [14], cell adhesion [15], cell migration [16], transcriptional activities [17], and resistance to anoikis [18]. Several studies suggest CRT to be involved in the development of different cancer types via tumor generation and progression. For example, CRT affects cancer metastasis by interacting with integrins [19, 20], regulating cell proliferation via upregulation of proangiogenic VEGF in various cancer cells [21–23], and suppressing cell proliferation and enhancing cell differentiation in neuroblastoma cells [24]. However, CRT expressed (ecto-CRT) on the cell surface serves as a phagocytic signal for immunogenic cell death (ICD) mediated by dendritic cells (DCs) and cytotoxic T-cell activation [25–27].
Moreover, cytosolic CRT functions as a receptor for nuclear export of glucocorticoid receptor [28, 29] to suppress transcriptional activity of steroid hormone receptors in the development of hormone-dependent tumors [30]. However, how cytosolic CRT plays a role in the development of drug-resistant cancer remains unclear.
Calmodulin (CaM) is an intermediate for Ca2+-stimulated signaling cascades in many cell types. The Ca2+/CaM complex may activate a variety of enzymes and CaM Kinase (CaMK) family that control many cancer-related functions in several tumor types. Additionally, the CaMKII is a cancer marker that is frequently overexpressed and regulates cell proliferation, survival, invasion, and migration in various cancer types [31]. Our previous study demonstrated critical roles of CaM/CaMKII-γ pathway in the apicidin-persistent HCC prosurvival capability via ERK1/2/CREB/c-fos signaling activation [32]. These findings suggest that CaMKII-CREB is linked with differentiation, proliferation, apoptosis, and tumorigenicity; however, its role in the regulation of CRT remains unclear. Thus, we hypothesized that CRT may control drug-resistance in cancer cells by regulating the ER stress pathway or stimulating CRT translocation pathway. In the present study, CRT was overexpressed, which led to its translocation to the nucleus causing apoptosis in HCC cells treated with HDACis. The data showed that CRT was not regulated via ER stress pathway in HDACis-resistant HCC cells. Moreover, translocation of CRT, rather than its accumulation, led to apoptosis in HCC cells. These results indicate that CRT may serve as a target for developing anticancer therapeutics and preventive strategies by stimulating CRT translocation to the nucleus.
RESULTS
Analysis of the drug sensitivity and differential protein expression between parental and resistance HCC cells
First, we assessed the effect of HDACis on cell viability of HA22T and HDACis-R (apicidin-R and SAHA-R) cells using 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium-bromide (MTT) assay. Different concentrations of HDACis (apicidin and SAHA) were used to evaluate cell viability after incubating for 48 h. The analysis indicated that HDACis failed to reduce cell proliferation in HDACis-R cells; however, HDACis significantly attenuated the proliferation of HA22T parental cells in a dose-dependent manner (Figure 1A, 1B).
Figure 1.
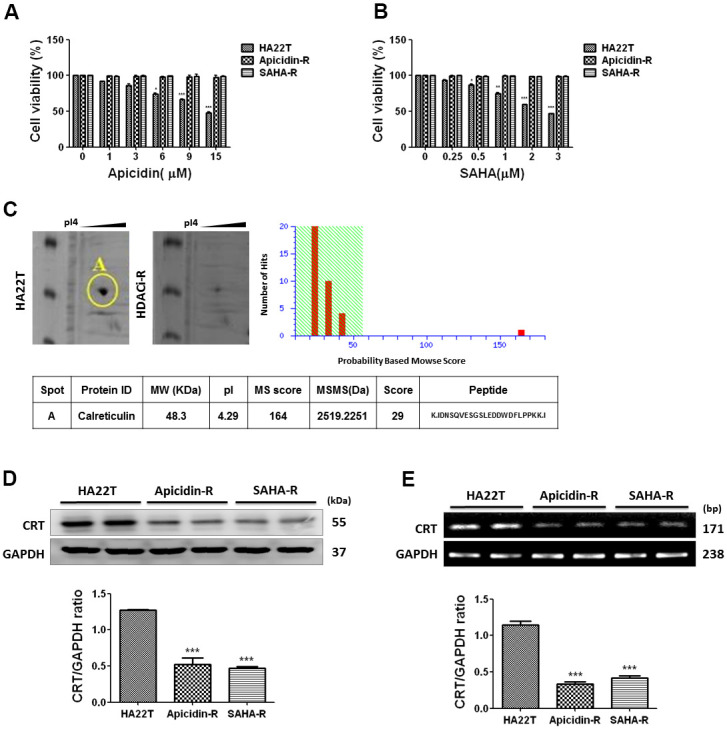
HCC cells resistance to HDACis correlated with downregulation of CRT. (A) Liver cancer cells are incubated with apicidin at various concentrations for 48 h and proliferation measured by MTT assay. (B) Liver cancer cells are incubated with SAHA at various concentrations for 48 h and proliferation measured by MTT assay. Data are expressed as percentage of the control and presented as mean ± S.D. of three independent experiments. (C) Two-dimensional gel electrophoresis analysis of proteins extracted from liver cancer cells, and further identification of candidate protein, calreticulin (CRT), by tandem mass spectrometer. (D, E) Western blotting and reverse transcription PCR analysis indicate decreased expression of CRT in HDACis-R cells than that in HA22T parental cells at protein and mRNA levels. Protein and mRNA expression are normalized to expression of GAPDH. *p <0.05, **p <0.01, ***p <0.001 compared with parental cell group.
Following, we used two-dimensional gel electrophoresis (2-DE) to identify differentially expressed proteins, and selected a protein spot differential between HA22T and HDACis-R cells. The protein spot was analyzed by mass spectrometry and identified as calreticulin; it was significantly downregulated in the HDACis-R cells than in HA22T parental cells (Figure 1C). Furthermore, western blotting and qPCR analysis confirmed the downregulation of CRT in HDACis-R cells than in HA22T parental cells at protein and mRNA levels, respectively (Figure 1D, 1E). Therefore, we hypothesized that resistance to HDACis in HA22T cells correlated with downregulation of CRT.
Overexpression of CRT enhances chemosensitivity to HDACis-mediated HCC cytotoxicity
Recent studies have shown that CRT plays a crucial role during tumor development. Therefore, we hypothesized that HDACis-R in HA22T cells was developed either by upregulation or downregulation of CRT. We performed short hairpin RNA (shRNA)-mediated knockdown and plasmid-based overexpression of CRT to examine its role in HA22T and HDACis-R cells. The analysis showed that knockdown of CRT had no significant effect on HA22T cells. The HDACis suppressed p-Akt levels, but the expression of cleaved caspase 3, cleaved PARP, and CRT increased in HA22T cells. Moreover, knockdown of CRT rescued HDACis-induced apoptosis by reducing levels of cleaved caspase 3 and cleaved PARP, and increased p-Akt levels in HA22T cells (Figure 2A, 2B). In contrast, transient transfection with CRT-plasmid showed no effect on the levels of p-Akt, cleaved caspase 3, and cleaved PARP. However, overexpression of CRT enhanced HDACis-induced apoptosis between HA22T and HDACis-R cells via decreased levels of p-Akt, along with increased expression of cleaved caspase 3 or cleaved PARP (Figure 2C–2F). Additionally, studies have demonstrated that CRT could be modified [33], degraded [34], and translocated to the nucleus [35]. Therefore, we investigated that whether the posttranslational modifications in CRT could enhance chemosensitivity in HCC cells. First, we assessed the effect of MG132 on the viability of HA22T and HDACis-R cells using MTT assay. The analysis indicated that MG132 (10μM) significantly attenuated viability of liver cancer cells in 24–48 h (Supplementary Figure 1A). We treated liver cancer cells with MG132 (10μM) for 8 h that induced the accumulation of CRT and posttranslational modifications in CRT via polyubiquitination between HA22T and HDACis-R cells (Supplementary Figure 1B, 1C). Moreover, CRT ubiquitination increased in the HDACis-R cells than that in HA22T parental cells after treatment with MG132 (10 μM) for 8 h and subsequent immunoprecipitation (IP)/immunoblotting (IB) analysis (Supplementary Figure 1B, 1C). Nevertheless, after incubation with HDACis (10 μM of Apicidin or 3 μM SAHA) for 24 h and addition of MG132 at the 16th hour enhanced chemosensitivity in HA22T parental cells, especially in the HDACis-R cells (Supplementary Figure 1D, 1E). The results indicate that stress stimulates translocation of CRT that is upregulated and further induces cell apoptosis.
Figure 2.
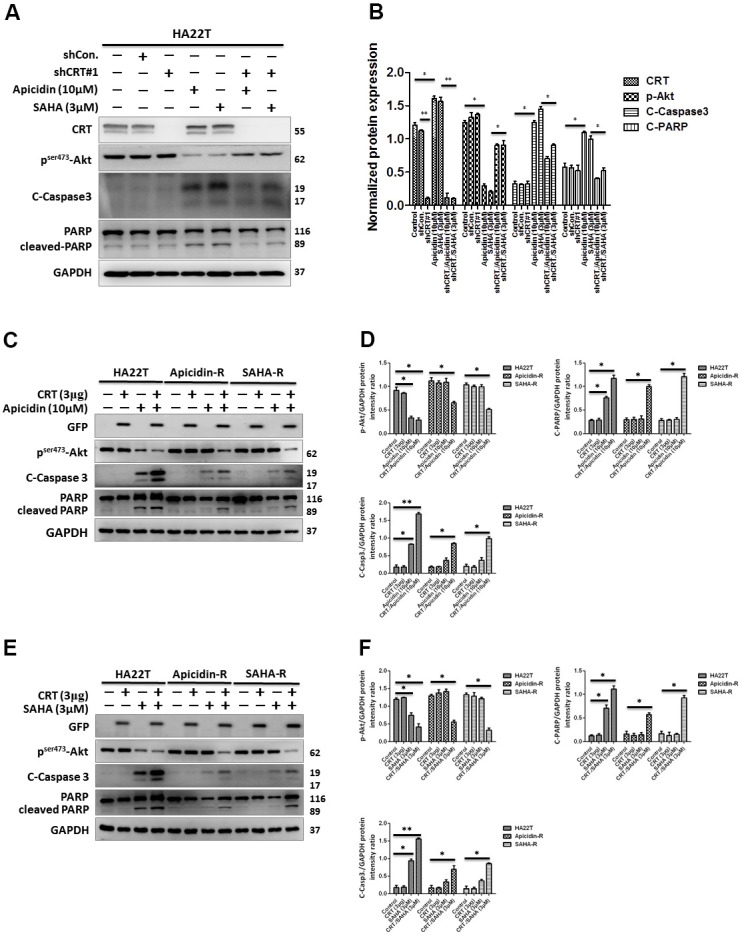
Effect of overexpression and downregulation of CRT overexpression on cell apoptosis in HCC cells treated with HDAC inhibitors. (A, B) shRNA-mediated knockdown of CRT enhances drug-resistance in HDAC inhibitor-treated cells than in HA22T parental cells. (C–F) Overexpression of CRT overexpression enhances HDAC inhibitor-induced chemosensitivity in HDAC inhibitor-resistant cells than in parental cells. All protein samples are analyzed using western blotting. Protein expression is normalized to expression of GAPDH. *p <0.05, **p <0.01, ***p<0.001 compared with control group.
Expression of CRT is not regulated by ER stress pathway in HDACis-R cells
Calreticulin resides mainly in the endoplasmic reticulum (ER), where it functions as a chaperone [36] and calcium homeostasis regulator [37]. To explore the molecular mechanism by CRT, we examined the protein expression level of the traditional ER stress pathways. The analysis indicated that expression of the ER stress signaling proteins (GRP78, p-PERK, p-eIF2, ERp57, and CRT) in the HDACis-R cells was downregulated than that in parental cells under normal conditions (Figure 3A, 3B). This indicated that the CRT expression was regulated by the ER stress pathway in HDACis-R cells.
Figure 3.

ER stress-dependent pathway was not significantly involved in HDAC inhibitors-resistant HA22T cells. (A, B) Protein expression levels of CRT-regulated ER stress pathway proteins (GRP78, p-PERK, p-eIF2, ERp57 and CRT) are measured by western blotting analysis. (C) Effect of thapsigargin (TG)-induced ER stress on the cell viability of liver cancer cells. Liver cancer cells are incubated with different concentration of TG for 24 h and cell viability is tested by MTT assay. (D, E) Expression of CRT-regulated ER stress pathway proteins (GRP78, p-PERK, p-eIF2, ERp57 and CRT) are measured using western blotting. All protein samples are analyzed by western blotting. Protein expression is normalized to expression of GAPDH. *p<0.05, **p<0.01, ***p<0.001 compared with control group.
To explore the regulation of CRT expression by the ER stress pathway activity, we treated the HCC cells with ER stress activator, thapsigargin (TG) [38] for 24 h. The analysis showed that TG could significantly attenuate HA22T parental cell viability in a dose-dependent manner, but not in apicidin-R cells (Figure 3C). TG initiated apoptosis via activation of p-PERK, a protein functioning upstream to the ER stress in HA22T parental cells. Moreover, expression of GRP78, p-PERK, and p-eIF2α was upregulated in HA22T parental cells than that in HDACis-R (Figure 3D, 3E), consistent with our previous observations with fisetin-induced chemosensitivity in HCC cells [39].
Additionally, expression of CRT was upregulated by TG between HA22T and HDACis-R cells. These results suggest that expression of CRT was not regulated via activation of the ER stress pathway—PERK-eIF2α—in HDACis-R cells.
Nuclear translocation of calreticulin increases cell apoptosis in HDACis-R cells
To demonstrate that whether HDACis led translocation of CRT to the nucleus and further induced HCC apoptosis, we treated HCC cells with HDACis (10 μM of Apicidin or 3 μM of SAHA) and measured protein levels of the members of CaM/CaMKII/CREB signaling pathway in both the cytosolic and nuclear fractions. The results showed that HDACis significantly induced translocation of CRT to the nucleus and further led to apoptosis via attenuation of the CaM/CaMKII/CREB signaling pathway in the nuclear fraction of HA22T parental cells, but not in the HDACis-R cells (Figure 4A, 4B). Next, we transiently transfected a CRT plasmid (3 μg) in HCC cells for 24 h before treating them with HDACis to investigate whether overexpression of CRT could enhance HDACis-induced chemosensitivity in HDACis-R cells. The analysis showed that overexpression of CRT was not significantly affected between HA22T and HDACis-R cells. Moreover, HDACis significantly induced apoptosis by initiating translocation of CRT to the nucleus and further attenuating CaM/CaMKII/CREB signaling pathway between CRT-transfection in HA22T and HDACis-R cells in nuclear fractions (Figure 4C, 4D). Further, using TUNEL assay, we found that HDACis only induced cell apoptosis in HA22T cells than in control cells. Overexpression of CRT and treatment with HDACis significantly increased the TUNEL-positive cells than with HDACis group between HA22T and HDACis-R cells (Figure 4E–4H). Additionally, abnormal calcium signaling is related to occurrence of various cancer types and therapy resistance [40, 41]. However, it remains unclear how CRT affects calcium regulation in the nucleus. To determine whether CRT overexpression affected nuclear calcium levels in HDACi-mediated HCC cytotoxicity, we measured calcium levels in the HCC cells by using calcium detection reagents. The analysis demonstrated that HDACis significantly induced calcium accumulation in HA22T parental cells. Moreover, HDACis significantly enhanced translocation of CRT to the nucleus and led to increased calcium accumulation in CRT-transfected HCC cells than that in untransfected cells (Figure 4I–4K). Taken together, these findings indicate that HDACis induce translocation of CRT to the nucleus and further caused cell apoptosis via CaM/CaMKII/CREB signaling activation, and also enhance nuclear calcium accumulation in CRT-transfected HCC cells.
Figure 4.

HDAC inhibitors trigger CRT translocation to the nucleus in liver cancer cells overexpressing CRT. (A, B) HDAC inhibitors activate CRT nuclear translocation and inhibit CaM/CaMKII/CREB signaling pathway in the nuclear isolate of the HA22T parental cells. (C, D) HDAC inhibitors activate CRT nuclear translocation and inhibit CaM/CaMKII/CREB signaling pathway in the nuclear isolate of HCC cells overexpressing CRT.
Figure 4.

HDAC inhibitors trigger CRT translocation to the nucleus in liver cancer cells overexpressing CRT. (E–G) HCC cells treated with vehicle, transfected with CRT-plasmid, or HDAC inhibitors alone or in combination exhibit DNA damage, as determined by TUNEL assays. Scale bar = 100 μm. Upper panel: Green fluorescence indicates CRT-transfected HCC cells. HCC cells are stained with fluorescein-labeled dUTP according to the protocols reported in the Methods section. Red fluorescence indicates TUNEL-positive cells. Lower panel: HCC cells in the upper panel are stained with 4',6-diamidino-2-phenylindole (DAPI) to identify cell nuclei. (H) Rate of apoptosis is calculated as the percentage of TUNEL-positive cells among the total number of counted parental and resistant cells (mean ± SD, n = 3). (I–K) Cells are seeded in an 8-well chamber and treated with vehicle, CRT-plasmid, or HDAC inhibitors alone or in combination. HDAC inhibitors trigger CRT translocation to the nucleus and stimulate calcium accumulation, analyzed by confocal microscopy. Scale bar = 10 μm. Mean values are significantly different than that in control group. *p< 0.05, **p< 0.01, ***p< 0.001.
DISCUSSION
In this study, we selected CRT was found to be a differentially expressed protein between the HA22T parental cells and HDACis-resistant cells previous studies. Many studies showed CRT to be highly expressed in several cancer types [42], such as high level microsatellite instability colorectal adenocarcinomas [43], esophageal squamous cell carcinoma [16], oral squamous cell carcinoma [44], breast cancer [45], and acute myeloid leukemia [46]. In contrast, some studies observed downregulation of CRT in laryngeal squamous cell carcinoma lesions [47], endometrial cancer [48], vaginal and cervical carcinoma [49], and non-small-cell lung cancer [50]; expression of CRT is especially low in advanced human prostate cancer and ovarian serous carcinoma than that in benign tissue [51, 52]. Therefore, expression of CRT may correlate with carcinogenesis and cancer progression. The present study showed the expression of CRT to be lower in HDACis-R cells than that in HA22T parental cells at the protein and mRNA levels, suggesting it to be a new therapeutic target in HCC. The function of CRT is to bind to URP and induces ER stress in the UPR pathway via IRE-1α, XBP-1 and ATF-6. We detected the IRE-1, XBP-1 and ATF-6 expression after treated with thapsigargin in HCC cells. The results indicated that expression of IRE-1, XBP-1 and CHOP were higher in HA22T cells than in HDACi-R cells (Supplementary Figure 2A, 2B). Additionally, IRE-1 and CHOP expression upregulated after treated with TG in HCC cells but XBP-1 and ATF-6 were not. These results suggest that CHOP-induced apoptosis in HDACis-R cells were not mediated through PERK, XBP-1, ATF-6 signaling pathway.
Overexpression of CRT can significantly suppress cell viability and proliferation in MCF-7 [53] and neuroblastoma cells [54]. Here, overexpression of CRT-alone showed no effect on cell apoptosis. Interestingly, overexpression of CRT enhanced chemosensitivity in HDACis-mediated HCC cytotoxicity. Furthermore, CRT can be localized to the plasma membrane and nucleus either as native or post-translationally modified isoforms, performing diverse and important non-ER functions [55]. For instance, CRT is counterbalanced by CD47 for the adaptive immune response on the cell surface [56]. We also explored the role of UB-CRT in HCC apoptosis. The studies indicated that the levels of posttranslational modifications in CRT were higher in HDACi-R cells; also UB-CRT may lead to increased apoptosis in HDACis-mediated HCC cells; but UB-CRT-alone failed to affect the expression level of these proteins. CRT was stabilized by integrin-ligand binding on the extracellular matrix (ECM) and activated focal adhesion kinase (FAK) for regulating cell-adhesion [57, 58]. To understand how CRT regulates cell apoptosis, we evaluated the effect of CRT expression on ER stress pathway. Recent studies found that CRT is an ER-binding protein regulated by ERP57 [59–62]. Here, we observed that in HDACis-R cells, the expression of GRP78, p-PERK, p-eIF2α, ERp57, and CRT was significantly lower than that in HA22T parental cells. Therefore, we used TG (one of the most popular inducers of ER stress) in HA22T parental cells and HDACis-resistant cells. TG is a specific inhibitor of the sarcoplasmic/endoplasmic reticulum Ca2+-ATPase (SERCA) that causes ER stress through depletion of Ca2+ store in the ER [63–66]. CRT acts as a chaperone and affects Ca2+ homeostasis [67], which influences gene expression and cell adhesion [68]. CRT activated the Wnt and CaM/CaMK II pathways by regulating Ser/Thr phosphatase in regulating cellular adhesion, growth, migration, and differentiation [69–71]. Previous studies indicated that decreased CRT expression levels were paralleled by an increase in CaM abundance, with concomitant decrease in cell spreading [69]. Notably, inhibition of CRT expression could activate c-src expression compared to the overexpressing cells. Activation of c-src could increasing the affinity of calmodulin for CaMK II [72, 73]. However, Tyrosine phosphorylated calmodulin effectively activates CaMK II and this tyrosine phosphorylation event is inhibited by high calcium concentration [74, 75]. Slight differences in Ca2+ signaling mechanisms can disrupt cell function, such as using too much/ little Ca2+ at the wrong time/place can lead to rapid cell death by necrosis, or inducing cell apoptosis. Following this suggestion, we check the expression of Ca channel-related markers in HCC cells and obtained that treated with HDAC inhibitors up-regulation of Ca channel-related markers in the HA22T cells but no effect in HDACis-R cells (Supplementary Figure 3A–3D). Interestingly, treated with HDAC inhibitors in overexpression of CRT HCC cells up-regulated expression of L-type Ca++ CP α1C, SERCA2 and IP3R in HCC cells. These results hint that HCC cells resistance to HDACi through CRT expression but not related with Ca channel. In the current study, overexpressed CRT translocated to the nucleus and activated the CaM/CaMKII/CREB pathway in HDACi-mediated HCC cytotoxicity. Additionally, calcium accumulation activated cell apoptosis signaling by CRT in the nucleus. The study findings can aid in clinical treatment: measurement of CRT levels and determination of sensitivity to HDACis before chemotherapy would help in better planning the therapeutic scheme for effective treatment of HCC.
In conclusion, the study suggests that HDACis induce cell death by increasing cytoplasmic localization of CRT in the nucleus and further enhance calcium accumulation and CaM/CaMKII/CREB pathway suppression in HA22T parental cells. In contrast, low cytoplasmic levels of CRT prevent its nuclear translocation by HDACis, leading to chemoresistance and suppression of apoptosis in HDACis-R cells (Figure 5). Moreover, we identified a new drug resistance mechanism mediated by suppression of CRT translocation to the nucleus. In response to treatment with HDACis, reduced expression of CRT suppresses its translocation to the nucleus, inhibiting calcium accumulation and activation of CaM/CaMKII/CREB pathway. These results suggest that stimulating CRT translocation to the nucleus can be a novel target for treating drug resistance in liver cancer.
Figure 5.
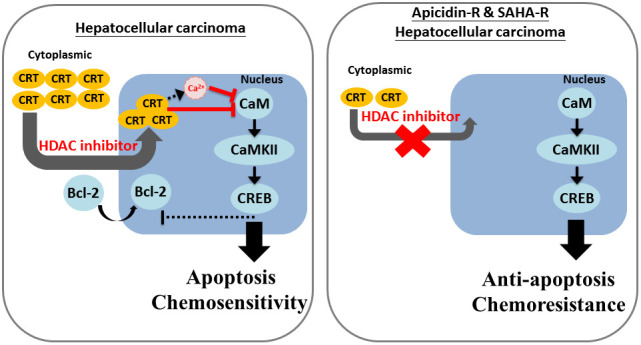
Mechanism of CRT-mediated chemosensitivity in HCC cells via nucleus translocation and reduction in CaM/CaMKII/CREB signaling. We revealed that decreased expression of CRT lead to down-regulated the chemosensitivity of HDACi in HCC cells. Moreover, we also obtain that CRT translocation into the nucleus is an important mechanism of chemoresistance and HDACi-induced cell apoptosis in HCC cells.
MATERIALS AND METHODS
Cell culture and generation of HDACis-resistant cells
HA22T (BCRC, Taiwan; Cat. No. 60168) or HDACis-resistant (HDACis-R; apicidin-R and SAHA-R) cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM; Sigma, St. Louis, MO, USA, Cat. No. D5523) supplemented with 10% FBS (characterized fetal bovine serum, Millipore, Billerica, MA, USA) and 1% penicillin (Corning, NY, USA, Cat. No. 30-002-CI) in humidified air with 5% CO2 at 37° C. The HDACis-R cells were established from HA22T cells by following the protocol elaborated in our previous report [10, 11, 39, 76].
Cell viability assay
Cells (1×104/well) were plated in 24-well plates and treated with increasing concentrations of apicidin, SAHA and thapsigargin for 48 h, 48 h and 24 h, respectively. Furthermore, a 10 μM concentration of MG132 (carbobenzoxy-Leu-Leu-leucinal, Sigma-Aldrich Inc., St. Louis, MO, USA) was used to treat liver cancer cells at different time points. A 0.5 mg/mL concentration of yellow colored 3-(4, 5-dimethylthiazolyl-2)-2, 5-diphenyltetrazolium bromide (MTT; Sigma-Aldrich Inc., St. Louis, MO, USA) was added to each well for 3 h in humidified air with 5% CO2 at 37° C, until conversion into formazan crystals via mitochondrial dehydrogenase enzymes [77]. The purple MTT-formazan crystals were dissolved in 200 μL of DMSO and the formazan level was measured as absorbance at 570 nm using an automated microplate reader (Thermo Fisher Scientific Inc., Grand Island, NY, USA).
Reverse transcription PCR
Total RNA was isolated and purified from cells using Quick-RNA™ MiniPrep kit (Zymo Research, Irvine, CA, USA), according to the manufacturer’s protocol. A total of 1 μg of RNA from each sample was reverse transcribed with a TaqMan™ reverse transcription kit (part no. N808-0234; Applied Biosystems), following the manufacturer’s protocol; and placed the PCR tubes on a thermal cycler (Thermo Fisher Scientific Inc., Grand Island, NY, USA) and incubated at 42° C for 65 min, 90° C for 5 min, and then at 4° C. The synthesized cDNAs were further amplified by RT-PCR using CRT primers (Forward: 5’-CGTCTACTTCAAGGAGCAGT-3’, Reverse: 5’-AGAGCATAAAAGCGTGCATC-3’, 171 bp), GAPDH primers (Forward: 5’-AAG GCTGTGGGCAAGG-3’, Reverse: 5’-TGGAGGAGTGGGTGTCG-3’, 238 bp), with the ProTaq System using ProTaq Master Mix (Protech Technology Enterprise CO., Ltd., Taiwan). One μL of the cDNA was amplified in a total volume of 25 μL reaction mixture containing 10× Reaction buffer (2.5 μL), 25 mM MgCl2 (1.5 μL), 10 mM dNTP mixture (0.5 μL), and 10 μM of forward and reverse primers. The RT-PCR cycles were estimated in a preliminary study and optimized in the PCR exponential phase. The conditions were as follows: initiation, 95° C for 5 min; followed by 28 cycles of denaturation at 95° C for 30 sec, annealing at 54° C for 30 sec, and extension at 72° C for 50 sec; final extension, 72° C for 5 min; stop reaction, 4° C. PCR product was analyzed by gel electrophoresis on a 1.5% agarose gel. Gene expression was normalized to expression of GAPDH in each sample.
Two-dimensional (2D)-gel electrophoresis
Total protein was dissolved in urea lysis buffer containing 1% protease inhibitor cocktail, sonicated, and centrifuged at 17,000 ×g for 30 min. We used an 18-cm Immobiline DryStrip (GE Healthcare Life Sciences Inc., Pittsburgh, PA, USA) and IPGphor isoelectric focusing unit (GE Healthcare Life Sciences Inc.) at 62000 Vh to perform isoelectric focusing of protein (100 μg) obtained from gradient-purified total protein. A 12.5% sodium lauryl sulfate-polyacrylamide gel electrophoresis was used for second-dimension separation and used silver staining to detect the protein. Targeted protein spots were excised from the gel and subjected to in-gel digestion with trypsin to recover the peptide. The peptides were identified and analyzed by electrospray ionization mass spectrometry (MS/MS) from Proteomics Core Laboratory (Office of Research and Development, China Medical University, Taiwan).
Antibodies and drug formulations
The following antibodies were used in the study: anti-GRP78 (sc-1050, Santa Cruz, CA, USA), anti-p-PERK Thr981 (sc-32577, Santa Cruz), anti-eIf2α (sc-133132, Santa Cruz), anti-CRT (sc-166837, Santa Cruz), anti-ERp57 (sc-28823, Santa Cruz), anti-Ub (sc-8017, Santa Cruz), anti-p-Akt1/2/3 (sc-7985, Santa Cruz), anti-GAPDH (sc-32233, Santa Cruz), anti-GFP (sc-8334, Santa Cruz), anti-Lamin B1 (sc-374015, Santa Cruz), anti-CaM (sc-137079, Santa Cruz), anti-CaMKIIγ (sc-1541, Santa Cruz), CREB-1 (sc-186, Santa Cruz), anti-p-CaMKII (ab182647, abcam, Waltham, MA, USA), anti-Bcl-2 (610539, BD Biosciences, Franklin Lakes, NJ, USA), anti-cleaved-caspase 3 (9664, Cell Signaling Technology, Danvers, MA, USA), anti-p-CREB Ser133 (9198, Cell Signaling Technology), anti-p-eIF2α Ser51 (9721, Cell Signaling Technology), and anti-PARP (9542, Cell Signaling Technology). All secondary antibodies (HRP-conjugated anti-rabbit, anti-mouse, and anti-goat) were purchased from Sigma (Sigma-Aldrich Inc., St. Louis, MO, USA). All the drug formulations were purchased from Sigma (Sigma-Aldrich Inc.).
Western blotting analysis
Cell samples were extracted with lysis buffer (20 mM Tris base, 1% NP-40, 150 mM NaCl, 1 mM EDTA, 0.1% phosphatase inhibitor, and proteinase K inhibitor). Cell pellets were lysed for 30 min on ice and clarified by centrifugation at 12,000 rpm for 20 min, and protein concentration was measured using Bradford protein assay (Bio-Rad, Hercules, CA, USA). Further, an aliquot of each sample extraction equivalent to 20 μg of protein was boiled after adding an appropriate amount of 5-fold protein sample buffer (40 % glycerol, 10% SDS, 5% β-mercaptoethanol, 0.02% bromophenol blue, and 0.5 M Tris-HCl pH 6.8). The proteins were separated using 8–12% SDS-polyacrylamide gels, and transferred to PVDF membranes (Millipore, Belford, MA, USA) using Bio-Rad electrotransfer system (Bio-Rad Laboratories Inc., Hercules, CA, USA). The membranes were blocked with blocking buffer [78] (5 % non-fat milk, 20 mM Tris-Base, pH 7.4, 150 mM NaCl, and 0.1% Tween-20) at room temperature for 1–2 h and probed with specific primary antibodies at 4° C overnight. Furthermore, the protein bands were measured with HRP-conjugated secondary antibodies (Sigma-Aldrich, Inc.) at room temperature for 1 h and using custom-made ECL (Millipore, Billerica, MA, USA) detection reagents to detect the target protein on immunoblot PVDF membranes by the ImageQuant LAS 4000 digital imaging system (Digital Imaging System, Commerce, CA, USA).
Immunoprecipitation assay
The same procedure, as described for western blotting, was used to obtain the total protein. Further, 50 μL of the Protein G beads slurry was transferred to an Eppendorf tube and washed three times with TBST buffer (20 mM Tris-Base, 150 mM NaCl, 0.1 % Tween-20, and pH 7.4). Next, 2 μg of primary antibody was added in 250–500 μL of TBST buffer and transferred to an Eppendorf tube containing the Protein G beads slurry and subjected it to IP rotor at 4° C for 6 h. Furthermore, the contents were washed three times with TBST buffer and >500 μg of protein sample was added in an Eppendorf tube containing the Protein G beads slurry and subjected the samples to IP rotor at 4° C overnight. Finally, the supernatant was completely and carefully removed and the beads were washed 3–5 times with TBST buffer, and 50 μL of 1-fold sample buffer was boiled at 100° C for 10 min, followed by western blotting to separate the proteins.
Plasmid and gene transfection
Plasmid (pCMV3-CALR-GFPSpark) encoding calreticulin was purchased from Asia Bioscience Co., Ltd., Taiwan. An siRNA against CRT was purchased from Sigma. The cells were 60–70 % confluent at the time of transfection. The plasmid or siRNA of CRT were transfected in the cells using jetPRIME® (Polyplus Transfection Inc, Illkirch, France) transfection reagent, according to the manufacturer’s protocol. The cells were incubated in humidified air with 5 % CO2 at 37° C for 24 h.
Cytoplasmic and nuclear extraction
Cells were grown in 100 mm dishes, and at 60–70 % confluent growth, were transfected with 3 μg of plasmid for 24 h, and treated with 10 μM of apicidin or 3 μM of SAHA for 24 h. Cytoplasmic and nuclear fractions were obtained using the Nuclear/Cytosol Fractionation Kit (BioVision, Milpitas, CA, USA), following the manufacturer’s protocol. The extracted proteins were stored at -80° C for further western blotting assay.
TUNEL assay
Cells were grown in 8-well chamber slides (MatTek; Ashland, MA, USA) and transfected with pCMV3-CALR-GFPSpark (Asia Bioscience Co. Ltd. HG13539-ACG) for 24 h, and treated with HDACis (10 μM apicidin or 3 μM SAHA) for 24 h. We measured apoptosis/ DNA damage in HA22T cells or HDACis-resistant cells using In Situ Cell Death Detection Kit and TMR red (Roche, Basel, Switzerland), according to the manufacturer’s instructions [79]. Fluorescence was visualized using a fluorescence microscope coupled to an image analysis system. Four microscopic fields were randomly-selected, and average counts of TUNEL-positive cells were calculated.
Intracellular calcium staining
Cells were seeded and transfected with pCMV3-CALR-GFPSpark (Asia Bioscience Co. Ltd. HG13539-ACG) for 24 h in chamber slides, and treated with HDACis (10 μM apicidin or 3 μM SAHA) for 24 h. Cells were stained with 5 μM of Cal-630™ AM kit (AAT Bioquest Inc., Sunnyvale, CA, USA), and fixed with 4 % paraformaldehyde for 30 min at room temperature. Finally, the samples were washed by PBS for three times and stained with Fluoroshield™ with DAPI (blue) (Sigma-Aldrich, Inc.) as nuclear counterstain. The samples were analyzed under a fluorescence microscope with an excitation wavelength of 475 nm (green) to detect CRT translocation, an excitation wavelength of 600/640 nm to detect calcium, and UV light microscope to detect cell nucleus (blue).
Statistical analysis
Statistical analysis was performed by one-way analysis of variance (One-way ANOVA) and SigmaPlot 10.0 software (Systat Software Inc., San Jose, CA, USA), along with GraphPad Prism 8. In all tests, p values < 0.05 were considered as statistically significant. * p < 0.05, **p < 0.01, and ***p < 0.001.
Supplementary Material
Footnotes
AUTHOR CONTRIBUTIONS: Conceptualization: CYH, PHL, YSL; Funding Acquisition: YSL, CYH; Investigation: YCC; Methodology: YCC, PHL, TSC, YML; Project Administration: WWK; Resources: CYH, YSL; Supervision: WWK, MCC, TFW; Validation: WWK; Writing -Original Draft Preparation: YSL; Writing and Editing: YCC, PHL, CYH; English editing: CCL. PHL* and CYH* contributed equally to this work.
CONFLICTS OF INTEREST: The authors declare no conflicts of interest.
FUNDING: This study was supported by the Kaohsiung Armed Forces General Hospital, China Medical University: CMU107-TC-05, China Medical University Hospital: DMR-108-143 and Ministry of Science and Technology grants from Taiwan MOST 106-2320-B-303 -004 -MY3.
REFERENCES
- 1.Verslype C, Cohn AL, Kelley RK, Yang TS, Su WC, Ramies DA, Lee Y, Shen X, Van Cutsem E. Activity of cabozantinib (XL184) in hepatocellular carcinoma: results from a phase II randomized discontinuation trial (RDT). J Clin Oncol. 2012; 30:4007. 10.1200/jco.2012.30.15_suppl.4007 [DOI] [Google Scholar]
- 2.Zhu AX, Baron AD, Malfertheiner P, Kudo M, Kawazoe S, Pezet D, Weissinger F, Brandi G, Barone CA, Okusaka T, Wada Y, Park JO, Ryoo BY, et al. Ramucirumab as Second-Line Treatment in Patients With Advanced Hepatocellular Carcinoma: Analysis of REACH Trial Results by Child-Pugh Score. JAMA Oncol. 2017; 3:235–43. 10.1001/jamaoncol.2016.4115 [DOI] [PubMed] [Google Scholar]
- 3.Rinninella E, Cerrito L, Spinelli I, Cintoni M, Mele MC, Pompili M, Gasbarrini A. Chemotherapy for Hepatocellular Carcinoma: Current Evidence and Future Perspectives. J Clin Transl Hepatol. 2017; 5:235–48. 10.14218/JCTH.2017.00002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Daher S, Massarwa M, Benson AA, Khoury T. Current and Future Treatment of Hepatocellular Carcinoma: An Updated Comprehensive Review. J Clin Transl Hepatol. 2018; 6:69–78. 10.14218/JCTH.2017.00031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.El-Khoueiry AB, Sangro B, Yau T, Crocenzi TS, Kudo M, Hsu C, Kim TY, Choo SP, Trojan J, Welling TH 3rd, Meyer T, Kang YK, Yeo W, et al. Nivolumab in patients with advanced hepatocellular carcinoma (CheckMate 040): an open-label, non-comparative, phase 1/2 dose escalation and expansion trial. Lancet. 2017; 389:2492–502. 10.1016/S0140-6736(17)31046-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bruix J, Qin S, Merle P, Granito A, Huang YH, Bodoky G, Pracht M, Yokosuka O, Rosmorduc O, Breder V, Gerolami R, Masi G, Ross PJ, et al. , and RESORCE Investigators. Regorafenib for patients with hepatocellular carcinoma who progressed on sorafenib treatment (RESORCE): a randomised, double-blind, placebo-controlled, phase 3 trial. Lancet. 2017; 389:56–66. 10.1016/S0140-6736(16)32453-9 [DOI] [PubMed] [Google Scholar]
- 7.Wilhelm SM, Adnane L, Newell P, Villanueva A, Llovet JM, Lynch M. Preclinical overview of sorafenib, a multikinase inhibitor that targets both Raf and VEGF and PDGF receptor tyrosine kinase signaling. Mol Cancer Ther. 2008; 7:3129–40. 10.1158/1535-7163.MCT-08-0013 [DOI] [PubMed] [Google Scholar]
- 8.Fardi M, Solali S, Farshdousti Hagh M. Epigenetic mechanisms as a new approach in cancer treatment: An updated review. Genes Dis. 2018; 5:304–11. 10.1016/j.gendis.2018.06.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sawan C, Herceg Z. Histone modifications and cancer. Adv Genet. 2010; 70:57–85. 10.1016/B978-0-12-380866-0.60003-4 [DOI] [PubMed] [Google Scholar]
- 10.Hsieh CH, Cheng LH, Hsu HH, Ho TJ, Tu CC, Lin YM, Chen MC, Tsai FJ, Hsieh YL, Huang CY. Apicidin-resistant HA22T hepatocellular carcinoma cells strongly activated the Wnt/β-catenin signaling pathway and MMP-2 expression via the IGF-IR/PI3K/Akt signaling pathway enhancing cell metastatic effect. Biosci Biotechnol Biochem. 2013; 77:2397–404. 10.1271/bbb.130503 [DOI] [PubMed] [Google Scholar]
- 11.Hsu HH, Cheng LH, Ho TJ, Kuo WW, Lin YM, Chen MC, Lee NH, Tsai FJ, Tsai KH, Huang CY. Apicidin-resistant HA22T hepatocellular carcinoma cells massively promote pro-survival capability via IGF-IR/PI3K/Akt signaling pathway activation. Tumour Biol. 2014; 35:303–13. 10.1007/s13277-013-1041-3 [DOI] [PubMed] [Google Scholar]
- 12.Tu CC, Cheng LH, Hsu HH, Chen LM, Lin YM, Chen MC, Lee NH, Tsai FJ, Huang CY, Wu WJ. Activation of snail and EMT-like signaling via the IKKαβ/NF-κB pathway in Apicidin-resistant HA22T hepatocellular carcinoma cells. Chin J Physiol. 2013; 56:326–33. 10.4077/CJP.2013.BAB158 [DOI] [PubMed] [Google Scholar]
- 13.Liao PH, Hsu HH, Chen TS, Chen MC, Day CH, Tu CC, Lin YM, Tsai FJ, Kuo WW, Huang CY. Phosphorylation of cofilin-1 by ERK confers HDAC inhibitor resistance in hepatocellular carcinoma cells via decreased ROS-mediated mitochondria injury. Oncogene. 2017; 36:1978–90. 10.1038/onc.2016.357 [DOI] [PubMed] [Google Scholar]
- 14.Michalak M, Robert Parker JM, Opas M. Ca2+ signaling and calcium binding chaperones of the endoplasmic reticulum. Cell Calcium. 2002; 32:269–78. 10.1016/s0143416002001884 [DOI] [PubMed] [Google Scholar]
- 15.Papp S, Fadel MP, Opas M. ER-to-cell surface signalling: calreticulin and cell adhesion. J Appl Biomed. 2004; 2:1–14. 10.32725/jab.2004.001 [DOI] [Google Scholar]
- 16.Shi F, Shang L, Pan BQ, Wang XM, Jiang YY, Hao JJ, Zhang Y, Cai Y, Xu X, Zhan QM, Wang MR. Calreticulin promotes migration and invasion of esophageal cancer cells by upregulating neuropilin-1 expression via STAT5A. Clin Cancer Res. 2014; 20:6153–62. 10.1158/1078-0432.CCR-14-0583 [DOI] [PubMed] [Google Scholar]
- 17.Yokoyama M, Hirata K. New function of calreticulin: calreticulin-dependent mRNA destabilization. Circ Res. 2005; 97:961–3. 10.1161/01.RES.0000193564.46466.2a [DOI] [PubMed] [Google Scholar]
- 18.Pallero MA, Elzie CA, Chen J, Mosher DF, Murphy-Ullrich JE. Thrombospondin 1 binding to calreticulin-LRP1 signals resistance to anoikis. FASEB J. 2008; 22:3968–79. 10.1096/fj.07-104802 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Dedhar S. Novel functions for calreticulin: interaction with integrins and modulation of gene expression? Trends Biochem Sci. 1994; 19:269–71. 10.1016/0968-0004(94)90001-9 [DOI] [PubMed] [Google Scholar]
- 20.Coppolino MG, Dedhar S. Ligand-specific, transient interaction between integrins and calreticulin during cell adhesion to extracellular matrix proteins is dependent upon phosphorylation/dephosphorylation events. Biochem J. 1999; 340:41–50. 10.1042/bj3400041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Weng WC, Lin KH, Wu PY, Lu YC, Weng YC, Wang BJ, Liao YF, Hsu WM, Lee WT, Lee H. Calreticulin Regulates VEGF-A in Neuroblastoma Cells. Mol Neurobiol. 2015; 52:758–70. 10.1007/s12035-014-8901-8 [DOI] [PubMed] [Google Scholar]
- 22.Chen CN, Chang CC, Su TE, Hsu WM, Jeng YM, Ho MC, Hsieh FJ, Lee PH, Kuo ML, Lee H, Chang KJ. Identification of calreticulin as a prognosis marker and angiogenic regulator in human gastric cancer. Ann Surg Oncol. 2009; 16:524–33. 10.1245/s10434-008-0243-1 [DOI] [PubMed] [Google Scholar]
- 23.Roberts E, Cossigny DA, Quan GM. The role of vascular endothelial growth factor in metastatic prostate cancer to the skeleton. Prostate Cancer. 2013; 2013:418340. 10.1155/2013/418340 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lin KH, Weng WC, Wu PY, Lu YC, Wang BJ, Liao YF, Hsu WM, Lee H. Calreticulin Up-Regulates VEGFs Expression in Neuroblastoma Cell Lines. FASEB J. 2015; 29:974–7. 10.1096/fasebj.29.1_supplement.974.7 [DOI] [PubMed] [Google Scholar]
- 25.Garg AD, Dudek AM, Agostinis P. Calreticulin surface exposure is abrogated in cells lacking, chaperone-mediated autophagy-essential gene, LAMP2A. Cell Death Dis. 2013; 4:e826. 10.1038/cddis.2013.372 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Wang HT, Lee HI, Guo JH, Chen SH, Liao ZK, Huang KW, Torng PL, Hwang LH. Calreticulin promotes tumor lymphocyte infiltration and enhances the antitumor effects of immunotherapy by up-regulating the endothelial expression of adhesion molecules. Int J Cancer. 2012; 130:2892–902. 10.1002/ijc.26339 [DOI] [PubMed] [Google Scholar]
- 27.Gardai SJ, McPhillips KA, Frasch SC, Janssen WJ, Starefeldt A, Murphy-Ullrich JE, Bratton DL, Oldenborg PA, Michalak M, Henson PM. Cell-surface calreticulin initiates clearance of viable or apoptotic cells through trans-activation of LRP on the phagocyte. Cell. 2005; 123:321–34. 10.1016/j.cell.2005.08.032 [DOI] [PubMed] [Google Scholar]
- 28.Holaska JM, Black BE, Rastinejad F, Paschal BM. Ca2+-dependent nuclear export mediated by calreticulin. Mol Cell Biol. 2002; 22:6286–97. 10.1128/MCB.22.17.6286-6297.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Roderick HL, Campbell AK, Llewellyn DH. Nuclear localisation of calreticulin in vivo is enhanced by its interaction with glucocorticoid receptors. FEBS Lett. 1997; 405:181–5. 10.1016/s0014-5793(97)00183-x [DOI] [PubMed] [Google Scholar]
- 30.Zheng Y, Murphy LC. Regulation of steroid hormone receptors and coregulators during the cell cycle highlights potential novel function in addition to roles as transcription factors. Nucl Recept Signal. 2016; 14:e001. 10.1621/nrs.14001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Brzozowski JS, Skelding KA. The Multi-Functional Calcium/Calmodulin Stimulated Protein Kinase (CaMK) Family: Emerging Targets for Anti-Cancer Therapeutic Intervention. Pharmaceuticals (Basel). 2019; 12:8. 10.3390/ph12010008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hsu WC, Le HN, Lin YJ, Chen MC, Wang TF, Li CC, Kuo WW, Mahalakshmi B, Singh CH, Chen MC, Huang CY. Calmodulin/CaMKII-γ mediates prosurvival capability in apicidin-persistent hepatocellular carcinoma cells via ERK1/2/CREB/c-fos signaling pathway. J Cell Biochem. 2021; 122:612–25. 10.1002/jcb.29892 [DOI] [PubMed] [Google Scholar]
- 33.Denning GM, Leidal KG, Holst VA, Iyer SS, Pearson DW, Clark JR, Nauseef WM, Clark RA. Calreticulin biosynthesis and processing in human myeloid cells: demonstration of signal peptide cleavage and N-glycosylation. Blood. 1997; 90:372–81. [PubMed] [Google Scholar]
- 34.Goitea VE, Hallak ME. Calreticulin and Arginylated Calreticulin Have Different Susceptibilities to Proteasomal Degradation. J Biol Chem. 2015; 290:16403–14. 10.1074/jbc.M114.626127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Black BE, Paschal BM. Calreticulin-Mediated Nuclear Protein Export. Calreticulin: Springer. 2003; 75–84. 10.1007/978-1-4419-9258-1_8 [DOI] [Google Scholar]
- 36.Owusu BY, Zimmerman KA, Murphy-Ullrich JE. The role of the endoplasmic reticulum protein calreticulin in mediating TGF-β-stimulated extracellular matrix production in fibrotic disease. J Cell Commun Signal. 2018; 12:289–99. 10.1007/s12079-017-0426-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Smith MJ, Koch GL. Multiple zones in the sequence of calreticulin (CRP55, calregulin, HACBP), a major calcium binding ER/SR protein. EMBO J. 1989; 8:3581–6. 10.1002/j.1460-2075.1989.tb08530.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kamiya T, Hara H, Adachi T. Effect of endoplasmic reticulum (ER) stress inducer thapsigargin on the expression of extracellular-superoxide dismutase in mouse 3T3-L1 adipocytes. J Clin Biochem Nutr. 2013; 52:101–5. 10.3164/jcbn.12-46 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Liu YS, Kuo WW, Chen MC, Hsu HH, Tu CC, Yeh YL, Viswanadha VP, Liao PH, Huang CY, Chang YC. Inhibition of protein phosphatase 1 stimulates noncanonical ER stress eIF2α activation to enhance fisetin-induced chemosensitivity in HDAC inhibitor-resistant hepatocellular carcinoma cells. Cancers (Basel). 2019; 11:918. 10.3390/cancers11070918 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Varghese E, Samuel SM, Sadiq Z, Kubatka P, Liskova A, Benacka J, Pazinka P, Kruzliak P, Büsselberg D. Anti-Cancer Agents in Proliferation and Cell Death: The Calcium Connection. Int J Mol Sci. 2019; 20:3017. 10.3390/ijms20123017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Florea AM, Büsselberg D. Anti-cancer drugs interfere with intracellular calcium signaling. Neurotoxicology. 2009; 30:803–10. 10.1016/j.neuro.2009.04.014 [DOI] [PubMed] [Google Scholar]
- 42.Zamanian M, Veerakumarasivam A, Abdullah S, Rosli R. Calreticulin and cancer. Pathol Oncol Res. 2013; 19:149–54. 10.1007/s12253-012-9600-2 [DOI] [PubMed] [Google Scholar]
- 43.Vougas K, Gaitanarou E, Marinos E, Kittas C, Voloudakis-Baltatzis IE. Two-dimensional electrophoresis and immunohistochemical study of calreticulin in colorectal adenocarcinoma and mirror biopsies. J BUON. 2008; 13:101–7. [PubMed] [Google Scholar]
- 44.Harada K, Takenawa T, Ferdous T, Kuramitsu Y, Ueyama Y. Calreticulin is a novel independent prognostic factor for oral squamous cell carcinoma. Oncol Lett. 2017; 13:4857–62. 10.3892/ol.2017.6062 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Lwin ZM, Guo C, Salim A, Yip GW, Chew FT, Nan J, Thike AA, Tan PH, Bay BH. Clinicopathological significance of calreticulin in breast invasive ductal carcinoma. Mod Pathol. 2010; 23:1559–66. 10.1038/modpathol.2010.173 [DOI] [PubMed] [Google Scholar]
- 46.Helbling D, Mueller BU, Timchenko NA, Schardt J, Eyer M, Betts DR, Jotterand M, Meyer-Monard S, Fey MF, Pabst T. CBFB-SMMHC is correlated with increased calreticulin expression and suppresses the granulocytic differentiation factor CEBPA in AML with inv(16). Blood. 2005; 106:1369–75. 10.1182/blood-2004-11-4392 [DOI] [PubMed] [Google Scholar]
- 47.Ogino T, Shigyo H, Ishii H, Katayama A, Miyokawa N, Harabuchi Y, Ferrone S. HLA class I antigen down-regulation in primary laryngeal squamous cell carcinoma lesions as a poor prognostic marker. Cancer Res. 2006; 66:9281–9. 10.1158/0008-5472.CAN-06-0488 [DOI] [PubMed] [Google Scholar]
- 48.Xu Q, Chen C, Chen G, Chen W, Zhou D, Xie Y. Significance of calreticulin as a prognostic factor in endometrial cancer. Oncol Lett. 2018; 15:8999–9008. 10.3892/ol.2018.8495 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Hellman K, Alaiya AA, Becker S, Lomnytska M, Schedvins K, Steinberg W, Hellström AC, Andersson S, Hellman U, Auer G. Differential tissue-specific protein markers of vaginal carcinoma. Br J Cancer. 2009; 100:1303–14. 10.1038/sj.bjc.6604975 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Fucikova J, Becht E, Iribarren K, Goc J, Remark R, Damotte D, Alifano M, Devi P, Biton J, Germain C, Lupo A, Fridman WH, Dieu-Nosjean MC, et al. Calreticulin Expression in Human Non-Small Cell Lung Cancers Correlates with Increased Accumulation of Antitumor Immune Cells and Favorable Prognosis. Cancer Res. 2016; 76:1746–56. 10.1158/0008-5472.CAN-15-1142 [DOI] [PubMed] [Google Scholar]
- 51.Alur M, Nguyen MM, Eggener SE, Jiang F, Dadras SS, Stern J, Kimm S, Roehl K, Kozlowski J, Pins M, Michalak M, Dhir R, Wang Z. Suppressive roles of calreticulin in prostate cancer growth and metastasis. Am J Pathol. 2009; 175:882–90. 10.2353/ajpath.2009.080417 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Vaksman O, Davidson B, Tropé C, Reich R. Calreticulin expression is reduced in high-grade ovarian serous carcinoma effusions compared with primary tumors and solid metastases. Hum Pathol. 2013; 44:2677–83. 10.1016/j.humpath.2013.07.009 [DOI] [PubMed] [Google Scholar]
- 53.Prathyuman S, Sellappa S, Joseph S, Keyan KS. Enhanced calreticulin expression triggers apoptosis in the MCF-7 cell line. Asian Pac J Cancer Prev. 2010; 11:1133–6. [PubMed] [Google Scholar]
- 54.Hsu WM, Hsieh FJ, Jeng YM, Kuo ML, Chen CN, Lai DM, Hsieh LJ, Wang BT, Tsao PN, Lee H, Lin MT, Lai HS, Chen WJ. Calreticulin expression in neuroblastoma--a novel independent prognostic factor. Ann Oncol. 2005; 16:314–21. 10.1093/annonc/mdi062 [DOI] [PubMed] [Google Scholar]
- 55.Wiersma VR, Michalak M, Abdullah TM, Bremer E, Eggleton P. Mechanisms of Translocation of ER Chaperones to the Cell Surface and Immunomodulatory Roles in Cancer and Autoimmunity. Front Oncol. 2015; 5:7. 10.3389/fonc.2015.00007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Chao MP, Jaiswal S, Weissman-Tsukamoto R, Alizadeh AA, Gentles AJ, Volkmer J, Weiskopf K, Willingham SB, Raveh T, Park CY, Majeti R, Weissman IL. Calreticulin is the dominant pro-phagocytic signal on multiple human cancers and is counterbalanced by CD47. Sci Transl Med. 2010; 2:63ra94. 10.1126/scitranslmed.3001375 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Coppolino MG, Woodside MJ, Demaurex N, Grinstein S, St-Arnaud R, Dedhar S. Calreticulin is essential for integrin-mediated calcium signalling and cell adhesion. Nature. 1997; 386:843–7. 10.1038/386843a0 [DOI] [PubMed] [Google Scholar]
- 58.Coppolino M, Leung-Hagesteijn C, Dedhar S, Wilkins J. Inducible interaction of integrin alpha 2 beta 1 with calreticulin. Dependence on the activation state of the integrin. J Biol Chem. 1995; 270:23132–8. 10.1074/jbc.270.39.23132 [DOI] [PubMed] [Google Scholar]
- 59.Xu Q, Chen C, Lin A, Xie Y. Endoplasmic reticulum stress-mediated membrane expression of CRT/ERp57 induces immunogenic apoptosis in drug-resistant endometrial cancer cells. Oncotarget. 2017; 8:58754–64. 10.18632/oncotarget.17678 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Panaretakis T, Joza N, Modjtahedi N, Tesniere A, Vitale I, Durchschlag M, Fimia GM, Kepp O, Piacentini M, Froehlich KU, van Endert P, Zitvogel L, Madeo F, Kroemer G. The co-translocation of ERp57 and calreticulin determines the immunogenicity of cell death. Cell Death Differ. 2008; 15:1499–509. 10.1038/cdd.2008.67 [DOI] [PubMed] [Google Scholar]
- 61.Obeid M. ERP57 membrane translocation dictates the immunogenicity of tumor cell death by controlling the membrane translocation of calreticulin. J Immunol. 2008; 181:2533–43. 10.4049/jimmunol.181.4.2533 [DOI] [PubMed] [Google Scholar]
- 62.Liu CC, Leclair P, Pedari F, Vieira H, Monajemi M, Sly LM, Reid GS, Lim CJ. Integrins and ERp57 Coordinate to Regulate Cell Surface Calreticulin in Immunogenic Cell Death. Front Oncol. 2019; 9:411. 10.3389/fonc.2019.00411 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Schmeits PC, Katika MR, Peijnenburg AA, van Loveren H, Hendriksen PJ. DON shares a similar mode of action as the ribotoxic stress inducer anisomycin while TBTO shares ER stress patterns with the ER stress inducer thapsigargin based on comparative gene expression profiling in Jurkat T cells. Toxicol Lett. 2014; 224:395–406. 10.1016/j.toxlet.2013.11.005 [DOI] [PubMed] [Google Scholar]
- 64.Gu Y, Cheng S, Chen G, Shen Y, Li X, Jiang Q, Li J, Cao Y. The effects of endoplasmic reticulum stress inducer thapsigargin on the toxicity of ZnO or TiO2 nanoparticles to human endothelial cells. Toxicol Mech Methods. 2017; 27:191–200. 10.1080/15376516.2016.1273429 [DOI] [PubMed] [Google Scholar]
- 65.Treiman M, Caspersen C, Christensen SB. A tool coming of age: thapsigargin as an inhibitor of sarco-endoplasmic reticulum Ca(2+)-ATPases. Trends Pharmacol Sci. 1998; 19:131–5. 10.1016/s0165-6147(98)01184-5 [DOI] [PubMed] [Google Scholar]
- 66.Kitamura M, Hiramatsu N. Real-time monitoring of ER stress in living cells and animals using ESTRAP assay. Methods Enzymol. 2011; 490:93–106. 10.1016/B978-0-12-385114-7.00005-2 [DOI] [PubMed] [Google Scholar]
- 67.Michalak M, Groenendyk J, Szabo E, Gold LI, Opas M. Calreticulin, a multi-process calcium-buffering chaperone of the endoplasmic reticulum. Biochem J. 2009; 417:651–66. 10.1042/BJ20081847 [DOI] [PubMed] [Google Scholar]
- 68.Czarnowski A, Papp S, Szaraz P, Opas M. Calreticulin affects cell adhesiveness through differential phosphorylation of insulin receptor substrate-1. Cell Mol Biol Lett. 2014; 19:77–97. 10.2478/s11658-014-0181-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Szabo E, Papp S, Opas M. Differential calreticulin expression affects focal contacts via the calmodulin/CaMK II pathway. J Cell Physiol. 2007; 213:269–77. 10.1002/jcp.21122 [DOI] [PubMed] [Google Scholar]
- 70.Parsons SJ, Parsons JT. Src family kinases, key regulators of signal transduction. Oncogene. 2004; 23:7906–9. 10.1038/sj.onc.1208160 [DOI] [PubMed] [Google Scholar]
- 71.Alvandi Z, Al-Mansoori LJ, Opas M. Calreticulin regulates Src kinase in osteogenic differentiation from embryonic stem cells. Stem Cell Res. 2020; 48:101972. 10.1016/j.scr.2020.101972 [DOI] [PubMed] [Google Scholar]
- 72.Abdel-Ghany M, el-Gendy K, Zhang S, Racker E. Control of src kinase activity by activators, inhibitors, and substrate chaperones. Proc Natl Acad Sci USA. 1990; 87:7061–5. 10.1073/pnas.87.18.7061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Benaim G, Villalobo A. Phosphorylation of calmodulin. Functional implications. Eur J Biochem. 2002; 269:3619–31. 10.1046/j.1432-1033.2002.03038.x [DOI] [PubMed] [Google Scholar]
- 74.Fukami Y, Nakamura T, Nakayama A, Kanehisa T. Phosphorylation of tyrosine residues of calmodulin in Rous sarcoma virus-transformed cells. Proc Natl Acad Sci USA. 1986; 83:4190–3. 10.1073/pnas.83.12.4190 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Corti C, Leclerc L’Hostis E, Quadroni M, Schmid H, Durussel I, Cox J, Dainese Hatt P, James P, Carafoli E. Tyrosine phosphorylation modulates the interaction of calmodulin with its target proteins. Eur J Biochem. 1999; 262:790–802. 10.1046/j.1432-1327.1999.00441.x [DOI] [PubMed] [Google Scholar]
- 76.Huang TY, Peng SF, Huang YP, Tsai CH, Tsai FJ, Huang CY, Tang CH, Yang JS, Hsu YM, Yin MC, Huang WW, Chung JG. Combinational treatment of all-trans retinoic acid (ATRA) and bisdemethoxycurcumin (BDMC)-induced apoptosis in liver cancer Hep3B cells. J Food Biochem. 2020; 44:e13122. 10.1111/jfbc.13122 [DOI] [PubMed] [Google Scholar]
- 77.Chang WS, Tsai CW, Yang JS, Hsu YM, Shih LC, Chiu HY, Bau DT, Tsai FJ. Resveratrol inhibited the metastatic behaviors of cisplatin-resistant human oral cancer cells via phosphorylation of ERK/p-38 and suppression of MMP-2/9. J Food Biochem. 2021; 45:e13666. 10.1111/jfbc.13666 [DOI] [PubMed] [Google Scholar]
- 78.Liu SP, Shibu MA, Tsai FJ, Hsu YM, Tsai CH, Chung JG, Yang JS, Tang CH, Wang S, Li Q, Huang CY. Tetramethylpyrazine reverses high-glucose induced hypoxic effects by negatively regulating HIF-1α induced BNIP3 expression to ameliorate H9c2 cardiomyoblast apoptosis. Nutr Metab (Lond). 2020; 17:12. 10.1186/s12986-020-0432-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Hsu HH, Kuo WW, Shih HN, Cheng SF, Yang CK, Chen MC, Tu CC, Viswanadha VP, Liao PH, Huang CY. FOXC1 Regulation of miR-31-5p Confers Oxaliplatin Resistance by Targeting LATS2 in Colorectal Cancer. Cancers (Basel). 2019; 11:1576. 10.3390/cancers11101576 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.