

Abstract
Skeletal muscles and bones, the largest tissues in the body of a non-obese person, comprise the musculoskeletal system, which allows mobility and protects internal organs. Although muscles and bones are closely related throughout life, observations during development and aging and in human and animal disuse models have revealed the synchronization of tissue mass such that muscle phenotype changes precede alterations in bone mineral density and strength. This review discussed that mechanical forces, which have been the traditional research focus, are not the only mechanism by which muscle-derived signals may affect bone metabolism and emphasized the significance of skeletal muscles as an endocrine organ that secretes bone-regulatory factors. Consequently, both mechanical and biochemical aspects should be considered to fully understand muscle–bone crosstalk. This review also suggested that specific myokines could be ideal therapeutic targets for osteoporosis to both increase bone formation and reduce bone resorption; moreover, these myokines could also be potential circulating biomarkers to predict musculoskeletal health.
Keywords: Biomarkers, Skeletal muscle, Bone, Fractures
INTRODUCTION
Fragility fractures, which occur owing to low-energy trauma such as a fall from a standing height or lower, are a major public health and economic concern worldwide, with an incidence estimated to outnumber those of stroke, heart attack, and breast cancer combined.1,2) With aging populations, fragility fractures are predicted to become a burden on healthcare systems. Although fragility fractures occur at various sites, including the wrist, humerus, and spine, commonly causing pain and disability and reducing quality of life, hip fractures present the most serious consequences. A recent epidemiological study using a nationwide database revealed that 14% and 21% of women and men, respectively, died within 1 year of hip fractures.3) Even if they recover, more than six out of ten older adults with hip fractures require assistance to carry out their activities of daily living.4,5) Therefore, preventing these low-trauma fractures is critical for independent living in older age.
Osteoporosis is a metabolic skeletal disorder characterized by low bone mass and poor bone quality, which result in an increased risk of fragility fractures. The prevalence of osteoporosis increases with age in both men and women in Korea, reaching nearly 70% in women aged >70 years.3) Based on the underlying mechanisms of osteoporosis, many effective drugs, such as anti-resorptives or bone-forming agents, have been developed and used in clinical practice to improve bone strength.6) However, despite these efforts, decreases in the incidence of osteoporosis and resultant fractures have been stagnant, thereby forming the major cause of increased morbidity and mortality in older adults. Given the multifaceted pathogenesis of fragility fractures, a systemic approach to various risk factors besides deteriorated bone health is essential for fracture prevention.
Sarcopenia corresponds to a progressive loss of skeletal muscle mass and function due to an imbalance between protein synthesis and breakdown.7) Several lines of evidence indicate that sarcopenia is closely associated with higher mortality and adverse health outcomes, such as metabolic and degenerative diseases, disability, comorbidities, and institutionalization in older adults.8-10) As a result, this syndrome has received attention globally in the last decade and is now considered a “geriatric giant” in super-aged societies.11) In particular, sarcopenia directly enhances fragility fractures by increasing the risk of falling due to aberrant balance and poor physical performance.12,13) Consequently, to effectively reduce fragility fractures, not only the metabolism of each bone and muscle but also the mechanisms underlying their interactions should be elucidated. The present review discusses muscle–bone communication, with a focus on the impact of muscles on bone health.
CLINICAL EVIDENCE SUPPORTING THE HIGHLY INTEGRATED NATURE OF MUSCLES AND BONES
Skeletal muscles and bones represent the largest tissues in the body of a non-obese person and comprise the musculoskeletal system, the structure of which allows mobility and protects the internal organs.14) In the law of the jungle, the coordination between muscles and bones is critical. For example, even if an animal has strong bones, weak muscles make it difficult to avoid predators. Conversely, in cases of weak bones but strong muscles, abrupt muscle-derived stress on the neighboring bones might increase fracture risk. Therefore, muscular and bone changes are likely matched throughout life.
Epidemiological studies have revealed that muscle and bone losses frequently occur simultaneously, especially in older individuals. In a nationwide population-based study of Asians aged >65 years, lower height-adjusted appendicular skeletal muscle was associated with lower bone mineral density (BMD) after considering potential confounders.15) Other studies, including a systematic review and meta-analysis, have consistently suggested that middle-aged and older adult men and women with sarcopenia are more likely to have a higher risk of developing osteoporosis than those without sarcopenia.16-18) More importantly, skeletal muscles affect not only bone mass but also bone quality and strength. Poor structural parameters of the femoral neck and distal radius, including thinner cortices, reduced cortical area, deteriorated microarchitecture, and lower section modulus, are more common in men with lower muscle mass and weaker handgrip strength.19,20) In a representative cohort of the general Korean population aged ≥50 years, women with sarcopenia had markedly decreased femoral neck composite strength indices for compression, bending, and impact.21) These findings provide clinical evidence of the strong relationship between muscles and bones in aging.
To describe the concurrent development of osteoporosis and sarcopenia, “osteosarcopenia,” a unique geriatric syndrome, was initially proposed by Hirschfeld et al.22) Although common hormonal, nutritional, lifestyle, and genetic determinants may explain at least some of the highly integrated nature of muscles and bones,23-26) the pathogenesis of osteosarcopenia is multifactorial and remains actively investigated. From the perspective of physicians investigating the musculoskeletal system, the clinical implication of osteosarcopenia is that treatment strategies that target either osteoporosis or sarcopenia separately may not be sufficient for fracture prevention; thus, novel approaches to simultaneously improve both tissues are needed. Several comprehensive reviews on osteosarcopenia have been published recently.27-29)
MUSCLE CHANGES OCCUR FIRST, FOLLOWED BY BONE ALTERATIONS
Although the crosstalk between muscles and bones is bidirectional, some clinical and preclinical observations indicate the dominant role of muscles over bones in synchronizing the mass and quality of these two tissues. For example, a longitudinal study of 138 boys and girls during pubertal development revealed that the rate of muscle force accrual was the highest a few months before the peak gain in bone strength.30) Similarly, other studies including adolescents and young adults showed that lean mass changes preceded alterations in whole-body BMD and bone strength.31,32) Antecedent muscle atrophy also affected bone loss in an animal model.33) Furthermore, when astronauts return to normal gravity, their muscle loss is recovered six times faster than the bone loss.34) Consequently, these data suggest that muscle changes occur first, followed by bone alterations, in the sequence of muscle–bone communication.
MUSCLE FORCE-GENERATED MECHANICAL SIGNALS TO THE BONE
The impact of muscles on bones has traditionally been addressed from a mechanical standpoint. The skeletal muscles attach to bones along the motion axis, transforming skeletal segments into a lever system that requires significant muscle force to generate the torque necessary for movement.35) The muscle force-generated strain transducing anabolic activity in nearby bones has been theoretically supported by the “mechanostat” theory.36,37) Osteocytes, the most mechanosensitive bone cells, respond to fluid shear stress and convert mechanical strain into biochemical signals to recruit osteoclasts or osteoblasts.38,39) Therefore, osteocytes are key players in mediating the loading effects on bone strength. The model of bone as a biomechanical tissue is further supported by the results of murine experiments showing the beneficial effects of low-magnitude mechanical stimuli on bone mass and structure.40,41)
Patients with spinal cord injury (SCI) are a useful human model to better understand the importance of muscle atrophy in bone health.42) This devastating condition refers to temporary or permanent spinal cord damage, mainly resulting from trauma, such as sports injuries, falls, or vehicle accidents, and mimics neurectomy by surgical procedures to induce deteriorated musculoskeletal phenotypes. Paralyzed patients enter an extreme catabolic state and undergo dramatic contractile and morphological muscular changes below the injury level, leading to a substantial loss of muscle mass, function, and endurance.43-45) Osteoporosis is a common consequence of SCI, with bone loss occurring at a rate of 1% per week during the first 6–12 months after SCI, that is, 5–20 times faster than that observed with aging, prolonged bed rest, or microgravity.43,46-48) Interestingly, bone loss is observed mainly in the paralyzed limbs and not in the non-affected areas.46) Other studies have shown that electrical stimulation of paralyzed muscles markedly reduces deleterious post-SCI bone adaptations and even reverses, at least partially, bone loss after SCI.44,49,50) These human data point to the elimination of internal loading via muscle contractions and regular gravity loading via ground reaction forces as the primary causes of SCI-related weak bone strength.
Consistent with findings showing concomitant muscle atrophy and bone loss in rodent disuse models of hindlimb unloading and botulinum toxin injection,33,42,51,52) muscle-induced mechanical impulses are the main factors with positive effects on bone metabolism.
BEYOND MECHANICAL: THE ROLE OF MYOKINES ON THE MUSCLE–BONE CROSSTALK
To describe cytokines or other peptides that are expressed, synthesized, and secreted from the skeletal muscles and exert biological activity in the human body, Pedersen et al.53) coined the term “myokine,” from the Greek words for “muscle” and “motion.” Recognizing muscles as an endocrine organ marks a watershed moment in our knowledge of how muscles communicate with other organs, such as the liver, brain, and adipose tissues, and establishes the notion of the critical need to maintain muscle health to reduce clinical disorders. With the development of modern technologies, including quantitative mass spectrometry-based proteomics, several secretome analyses have led to the identification of novel muscle-derived factors and various candidates from the supernatants of mouse C2C12 cells, human skeletal muscle cells, and L6 rat myotubes.54-58) Determining their biological roles in human health is currently a hot topic in this field.
Because skeletal muscles and bones are in close anatomical proximity, myokines likely biochemically influence bone homeostasis in a paracrine manner, a possibility strongly supported by long-standing observations in humans and rodents. Extensive muscle damage in open fractures is well known to impede fracture healing, with more negative consequences if the muscles are lost rather than crushed.35,59-61) Quantitative peripheral computed tomography, mechanical testing, and histomorphometry analysis in a mouse model indicated that covering open tibial fractures stripped from the periosteum with muscle flaps increased bone regeneration quality and rate compared to those obtained by covering them with similarly vascularized fasciocutaneous tissues.62,63) Furthermore, early soft tissue cover with a vascularized muscle flap markedly improved the healing of severe open tibial fractures after trauma in humans.64) These findings supported the idea that muscle production of local growth factors may induce bone formation, irrespective of physical load, and that biochemical and mechanical stimuli work together for muscle–bone crosstalk.65)
Several research groups have experimentally demonstrated the direct effect of myokines on bone metabolism. Conditioned media (CM) collected from C2C12 myotubes exerted protective effects against glucocorticoid-induced osteoblast and osteocyte apoptosis through β-catenin activation,66) and the primary myoblast CM of exercised mice significantly enhanced in vitro osteoblastogenesis.67) We first demonstrated that myotube CM suppressed in vitro bone resorption by inhibiting osteoclastogenesis and the resorptive activity of individual osteoclasts, whereas the same CM increased osteoblast viability and migration, thereby stimulating calvaria bone formation (Fig. 1).68) Furthermore, systemic treatment with myotube CM through the tail vein in ovariectomized mice increased bone mass by 30.7% compared to that of the non-CM.68) Although skeletal muscles can secrete complex factors, including positive or negative regulators, in terms of bone metabolism, our results showed the osteoprotective in vitro and in vivo net effects of various myokines on bone metabolism.
Fig. 1.
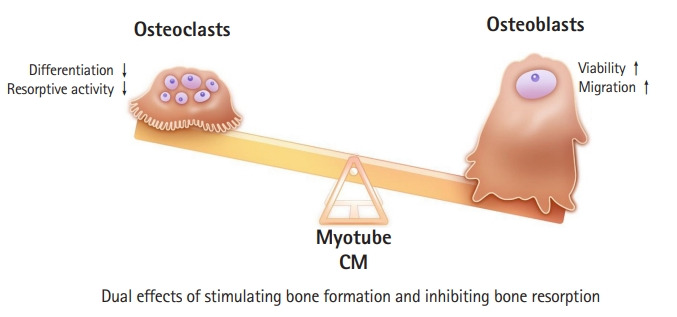
Myotube-conditioned media (CM) show dual osteoprotective effects of simultaneously stimulating bone formation and inhibiting bone resorption.
The finding that CM from skeletal muscle cells can promote bone formation while inhibiting bone resorption has crucial therapeutic implications in the management of osteoporosis. Although anti-resorptive agents, such as bisphosphonate and denosumab, have been most frequently used to treat osteoporosis worldwide, these drugs also concomitantly suppress bone formation due to a coupling phenomenon, raising questions about their long-term side effects and efficacy.69,70) Bone-forming medications such as teriparatide or abaloparatide have been suggested as alternatives. However, these drugs also sequentially stimulate bone resorption, rendering their anabolic action ineffective after 2 years of use.71) Therefore, dissociation of bone resorption from bone formation is essential for effective osteoporosis treatment. In this regard, muscle-secreted factors with opposing effects on osteoblasts and osteoclasts might be ideal candidates as potential therapeutic targets against metabolic bone diseases.
MUSCLE-DERIVED FACTORS INFLUENCING BONE METABOLISM
Specific myokines affecting bone homeostasis are being continuously discovered. Among these factors, myostatin and irisin have attracted the most attention in muscle–bone interactions. Myostatin, also known as growth differentiation factor 8, is a protein released by myocytes.72) Its levels are increased in catabolic situations, causing muscle atrophy, such as inflammation, microgravity, and immobilization.73) Although myostatin has been primarily evaluated as a negative regulator of muscles,74-76) increasing data indicate its direct function in bone remodeling. Myostatin strongly accelerates receptor activator of nuclear factor-κB ligand (RANKL)-mediated osteoclastogenesis,77) whereas osteogenic differentiation of bone marrow-derived mesenchymal stem cells increases in a load-dependent manner in myostatin-deficient mice, resulting in improved bone mass and strength.78,79) Consequently, myostatin plays a detrimental role in both bones and muscles. Irisin is a myokine that regulates energy metabolism and is activated by physical activity. In bone metabolism, irisin stimulated in vitro osteoblastogenesis and in vivo bone formation and prevented bone loss in hindlimb-suspended mice,67,80,81) while inhibiting bone resorption by directly suppressing osteoclast differentiation or indirectly downregulating RANKL expression in osteoblasts and osteocytes.82,83) Therefore, exercise-induced irisin is a pro-osteogenic factor explaining the parallel muscle and bone changes.
Our group proposed lumican, a small leucine-rich repeat proteoglycan, as a muscle-derived osteoprotective factor.84) Norheim et al.85) identified lumican in CM collected from human myotubes based on the results of proteomic analyses using database searches and reported significantly upregulated lumican expression in human skeletal muscles following strength training. These results matched our findings of its strong production and secretion in both cell lysates and the CM of myotubes.84) Importantly, lumican knockdown markedly reduced the known beneficial effects of myotube CM on the bones, while adding lumican to these CM restored the reduced osteoblast viability caused by lumican silencing.84) Additional in vitro and animal experiments revealed that lumican not only increased bone formation by stimulating osteoblast viability and differentiation but also suppressed osteoclastogenesis and in vitro bone resorption.84,86) These findings indicate that lumican may be a myokine involved in bone anabolism.
β-Aminoisobutyric acid (BAIBA), a metabolite released during muscle contraction, is involved in various metabolic processes such as improved insulin resistance and white adipose tissue browning.87,88) BAIBA has also recently demonstrated bone-protective activities such as enhancing osteocyte survival under oxidative stress and reducing bone loss with hindlimb unloading.89,90) Brain-derived neurotrophic factors,91) follistatin,92) leptin,93) interleukin (IL)-6,94) and IL-795) have been suggested as myokines that link muscle activity with skeletal health. In contrast, biochemical muscle–bone communication is a complicated process involving various beneficial or detrimental mediators to preserve musculoskeletal homeostasis. Therefore, in addition to the important roles of individual myokines, future studies should focus on identifying how combinations of these muscle-derived factors precisely regulate bone metabolism, especially in vivo.
CONCLUSION
Although muscles and bones have a close relationship throughout life, observations during development and aging and in both human and animal disuse models revealed that the synchronization of tissue mass occurs in such a way that changes in muscle phenotypes precede BMD and bone strength alterations.30-34) The present review discussed evidence that mechanical forces, which have been the traditional focus, are not the only mechanisms by which muscle-derived signals may affect bone metabolism and emphasized the significance of skeletal muscles as an endocrine organ that secretes bone-regulatory factors. Consequently, both mechanical and biochemical aspects should be considered to fully understand muscle–bone crosstalk. Moreover, muscle-secreted factors could be ideal therapeutic targets for osteoporosis, with dual effects of increasing bone formation and reducing bone resorption.
A particularly intriguing characteristic of myokines is that they can affect muscle metabolism in an autocrine manner. In addition to myostatin and irisin, which have well-established functions in muscles,96-98) apelin rejuvenated behavioral and circadian phenotypes and prevented muscle weakness and poor physical activity in aged mice.99,100) These results suggested that muscle-derived factors may be promising pharmacological candidates against osteosarcopenia and can simultaneously control bone and muscle losses beyond those due to osteoporosis (Fig. 2).
Fig. 2.
Muscle-secreted factors as promising pharmacological candidates against osteosarcopenia that can prevent both bone and muscle loss.
The ultimate objective of musculoskeletal research is to prevent fragility fractures and their consequent morbidity and mortality. To achieve this objective, the early detection of high-risk populations who are vulnerable to these low-trauma fractures is critical, in addition to effective treatment for osteoporosis and sarcopenia. In this regard, the levels of muscle-released factors can be easily measured in the blood; thus, myokines, whose actions on bone and/or muscle metabolism have been verified, could be useful as potential circulating biomarkers to predict musculoskeletal health. However, despite tremendous efforts to uncover the pivotal role of myokines in muscle and bone metabolism, human evidence of the clinical applicability of these muscle-derived factors as therapeutic targets and blood-based biomarkers for musculoskeletal diseases is lacking. Future efforts are expected to provide a solution to these limitations, allowing older individuals to live healthy lives through independent daily activities.
Footnotes
CONFLICT OF INTEREST
The researcher claims no conflicts of interest.
FUNDING
This study was supported by a grant from the National Research Foundation of Korea (NRF) funded by the South Korean government (MSIT) (No. 2019R1A2C2006527).
REFERENCES
- 1.Burge R, Dawson-Hughes B, Solomon DH, Wong JB, King A, Tosteson A. Incidence and economic burden of osteoporosis-related fractures in the United States, 2005-2025. J Bone Miner Res. 2007;22:465–75. doi: 10.1359/jbmr.061113. [DOI] [PubMed] [Google Scholar]
- 2.Leslie WD, O’Donnell S, Lagace C, Walsh P, Bancej C, Jean S, et al. Population-based Canadian hip fracture rates with international comparisons. Osteoporos Int. 2010;21:1317–22. doi: 10.1007/s00198-009-1080-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ahn SH, Park SM, Park SY, Yoo JI, Jung HS, Nho JH, et al. Osteoporosis and osteoporotic fracture fact sheet in Korea. J Bone Metab. 2020;27:281–90. doi: 10.11005/jbm.2020.27.4.281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Miller CW. Survival and ambulation following hip fracture. J Bone Joint Surg Am. 1978;60:930–4. [PubMed] [Google Scholar]
- 5.Amarilla-Donoso FJ, Lopez-Espuela F, Roncero-Martin R, Leal-Hernandez O, Puerto-Parejo LM, Aliaga-Vera I, et al. Quality of life in elderly people after a hip fracture: a prospective study. Health Qual Life Outcomes. 2020;18:71. doi: 10.1186/s12955-020-01314-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Brown JP. Long-term treatment of postmenopausal osteoporosis. Endocrinol Metab (Seoul) 2021;36:544–52. doi: 10.3803/EnM.2021.301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Dao T, Green AE, Kim YA, Bae SJ, Ha KT, Gariani K, et al. Sarcopenia and muscle aging: a brief overview. Endocrinol Metab (Seoul) 2020;35:716–32. doi: 10.3803/EnM.2020.405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Baek JY, Lee E, Jung HW, Jang IY. Geriatrics fact sheet in Korea 2021. Ann Geriatr Med Res. 2021;25:65–71. doi: 10.4235/agmr.21.0063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Baek JY, Lee E, Oh G, Park YR, Lee H, Lim J, et al. The Aging Study of Pyeongchang Rural Area (ASPRA): findings and perspectives for human aging, frailty, and disability. Ann Geriatr Med Res. 2021;25:160–9. doi: 10.4235/agmr.21.0100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cruz-Jentoft AJ, Bahat G, Bauer J, Boirie Y, Bruyere O, Cederholm T, et al. Sarcopenia: revised European consensus on definition and diagnosis. Age Ageing. 2019;48:16–31. doi: 10.1093/ageing/afy169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lee SY. Sarcopenia: a geriatric giant facing a huge transition. Ann Geriatr Med Res. 2021;25:1–3. doi: 10.4235/agmr.21.0023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Choi KA, Heu E, Nam HC, Park Y, Kim D, Ha YC. Relationship between low muscle strength, and protein intake: a preliminary study of elderly patients with hip fracture. J Bone Metab. 2022;29:17–21. doi: 10.11005/jbm.2022.29.1.17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wong RM, Wong H, Zhang N, Chow SK, Chau WW, Wang J, et al. The relationship between sarcopenia and fragility fracture-a systematic review. Osteoporos Int. 2019;30:541–53. doi: 10.1007/s00198-018-04828-0. [DOI] [PubMed] [Google Scholar]
- 14.Tagliaferri C, Wittrant Y, Davicco MJ, Walrand S, Coxam V. Muscle and bone, two interconnected tissues. Ageing Res Rev. 2015;21:55–70. doi: 10.1016/j.arr.2015.03.002. [DOI] [PubMed] [Google Scholar]
- 15.Kim S, Won CW, Kim BS, Choi HR, Moon MY. The association between the low muscle mass and osteoporosis in elderly Korean people. J Korean Med Sci. 2014;29:995–1000. doi: 10.3346/jkms.2014.29.7.995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Verschueren S, Gielen E, O’Neill TW, Pye SR, Adams JE, Ward KA, et al. Sarcopenia and its relationship with bone mineral density in middle-aged and elderly European men. Osteoporos Int. 2013;24:87–98. doi: 10.1007/s00198-012-2057-z. [DOI] [PubMed] [Google Scholar]
- 17.Lima RM, de Oliveira RJ, Raposo R, Neri SG, Gadelha AB. Stages of sarcopenia, bone mineral density, and the prevalence of osteoporosis in older women. Arch Osteoporos. 2019;14:38. doi: 10.1007/s11657-019-0591-4. [DOI] [PubMed] [Google Scholar]
- 18.Nielsen BR, Abdulla J, Andersen HE, Schwarz P, Suetta C. Sarcopenia and osteoporosis in older people: a systematic review and meta-analysis. Eur Geriatr Med. 2018;9:419–34. doi: 10.1007/s41999-018-0079-6. [DOI] [PubMed] [Google Scholar]
- 19.Szulc P, Blaizot S, Boutroy S, Vilayphiou N, Boonen S, Chapurlat R. Impaired bone microarchitecture at the distal radius in older men with low muscle mass and grip strength: the STRAMBO study. J Bone Miner Res. 2013;28:169–78. doi: 10.1002/jbmr.1726. [DOI] [PubMed] [Google Scholar]
- 20.Szulc P, Beck TJ, Marchand F, Delmas PD. Low skeletal muscle mass is associated with poor structural parameters of bone and impaired balance in elderly men: the MINOS study. J Bone Miner Res. 2005;20:721–9. doi: 10.1359/JBMR.041230. [DOI] [PubMed] [Google Scholar]
- 21.Kim BJ, Ahn SH, Kim HM, Lee SH, Koh JM. Low skeletal muscle mass associates with low femoral neck strength, especially in older Korean women: the Fourth Korea National Health and Nutrition Examination Survey (KNHANES IV) Osteoporos Int. 2015;26:737–47. doi: 10.1007/s00198-014-2959-z. [DOI] [PubMed] [Google Scholar]
- 22.Hirschfeld HP, Kinsella R, Duque G. Osteosarcopenia: where bone, muscle, and fat collide. Osteoporos Int. 2017;28:2781–90. doi: 10.1007/s00198-017-4151-8. [DOI] [PubMed] [Google Scholar]
- 23.Seeman E, Hopper JL, Young NR, Formica C, Goss P, Tsalamandris C. Do genetic factors explain associations between muscle strength, lean mass, and bone density?: a twin study. Am J Physiol. 1996;270(2 Pt 1):E320–7. doi: 10.1152/ajpendo.1996.270.2.E320. [DOI] [PubMed] [Google Scholar]
- 24.Zofkova I. Hormonal aspects of the muscle-bone unit. Physiol Res. 2008;57 Suppl 1:S159–69. doi: 10.33549/physiolres.931501. [DOI] [PubMed] [Google Scholar]
- 25.Kaji H. Interaction between muscle and bone. J Bone Metab. 2014;21:29–40. doi: 10.11005/jbm.2014.21.1.29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yoo JI, Lee KH, Choi Y, Lee J, Park YG. Poor dietary protein intake in elderly population with sarcopenia and osteosarcopenia: a nationwide population-based study. J Bone Metab. 2020;27:301–10. doi: 10.11005/jbm.2020.27.4.301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kirk B, Zanker J, Duque G. Osteosarcopenia: epidemiology, diagnosis, and treatment-facts and numbers. J Cachexia Sarcopenia Muscle. 2020;11:609–18. doi: 10.1002/jcsm.12567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sepulveda-Loyola W, Phu S, Bani Hassan E, Brennan-Olsen SL, Zanker J, Vogrin S, et al. the joint occurrence of osteoporosis and sarcopenia (osteosarcopenia): definitions and characteristics. J Am Med Dir Assoc. 2020;21:220–5. doi: 10.1016/j.jamda.2019.09.005. [DOI] [PubMed] [Google Scholar]
- 29.Cedeno-Veloz B, Lopez-Doriga Bonnardeauxa P, Duque G. Osteosarcopenia: a narrative review. Rev Esp Geriatr Gerontol. 2019;54:103–8. doi: 10.1016/j.regg.2018.09.010. [DOI] [PubMed] [Google Scholar]
- 30.Rauch F, Bailey DA, Baxter-Jones A, Mirwald R, Faulkner R. The ‘muscle-bone unit’ during the pubertal growth spurt. Bone. 2004;34:771–5. doi: 10.1016/j.bone.2004.01.022. [DOI] [PubMed] [Google Scholar]
- 31.Jackowski SA, Lanovaz JL, Van Oort C, Baxter-Jones AD. Does lean tissue mass accrual during adolescence influence bone structural strength at the proximal femur in young adulthood? Osteoporos Int. 2014;25:1297–304. doi: 10.1007/s00198-013-2592-2. [DOI] [PubMed] [Google Scholar]
- 32.Bogl LH, Latvala A, Kaprio J, Sovijarvi O, Rissanen A, Pietilainen KH. An investigation into the relationship between soft tissue body composition and bone mineral density in a young adult twin sample. J Bone Miner Res. 2011;26:79–87. doi: 10.1002/jbmr.192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lloyd SA, Lang CH, Zhang Y, Paul EM, Laufenberg LJ, Lewis GS, et al. Interdependence of muscle atrophy and bone loss induced by mechanical unloading. J Bone Miner Res. 2014;29:1118–30. doi: 10.1002/jbmr.2113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Keyak JH, Koyama AK, LeBlanc A, Lu Y, Lang TF. Reduction in proximal femoral strength due to long-duration spaceflight. Bone. 2009;44:449–53. doi: 10.1016/j.bone.2008.11.014. [DOI] [PubMed] [Google Scholar]
- 35.Cianferotti L, Brandi ML. Muscle-bone interactions: basic and clinical aspects. Endocrine. 2014;45:165–77. doi: 10.1007/s12020-013-0026-8. [DOI] [PubMed] [Google Scholar]
- 36.Frost HM. Bone’s mechanostat: a 2003 update. Anat Rec A Discov Mol Cell Evol Biol. 2003;275:1081–101. doi: 10.1002/ar.a.10119. [DOI] [PubMed] [Google Scholar]
- 37.Frost HM, Schonau E. The “muscle-bone unit” in children and adolescents: a 2000 overview. J Pediatr Endocrinol Metab. 2000;13:571–90. doi: 10.1515/jpem.2000.13.6.571. [DOI] [PubMed] [Google Scholar]
- 38.Bonewald LF. The amazing osteocyte. J Bone Miner Res. 2011;26:229–38. doi: 10.1002/jbmr.320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Klein-Nulend J, Bacabac RG, Bakker AD. Mechanical loading and how it affects bone cells: the role of the osteocyte cytoskeleton in maintaining our skeleton. Eur Cell Mater. 2012;24:278–91. doi: 10.22203/ecm.v024a20. [DOI] [PubMed] [Google Scholar]
- 40.Chan ME, Adler BJ, Green DE, Rubin CT. Bone structure and B-cell populations, crippled by obesity, are partially rescued by brief daily exposure to low-magnitude mechanical signals. FASEB J. 2012;26:4855–63. doi: 10.1096/fj.12-209841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lam H, Qin YX. The effects of frequency-dependent dynamic muscle stimulation on inhibition of trabecular bone loss in a disuse model. Bone. 2008;43:1093–100. doi: 10.1016/j.bone.2008.07.253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bettis T, Kim BJ, Hamrick MW. Impact of muscle atrophy on bone metabolism and bone strength: implications for muscle-bone crosstalk with aging and disuse. Osteoporos Int. 2018;29:1713–20. doi: 10.1007/s00198-018-4570-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Qin W, Bauman WA, Cardozo C. Bone and muscle loss after spinal cord injury: organ interactions. Ann N Y Acad Sci. 2010;1211:66–84. doi: 10.1111/j.1749-6632.2010.05806.x. [DOI] [PubMed] [Google Scholar]
- 44.Dudley-Javoroski S, Shields RK. Muscle and bone plasticity after spinal cord injury: review of adaptations to disuse and to electrical muscle stimulation. J Rehabil Res Dev. 2008;45:283–96. doi: 10.1682/jrrd.2007.02.0031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Biering-Sorensen B, Kristensen IB, Kjaer M, Biering-Sorensen F. Muscle after spinal cord injury. Muscle Nerve. 2009;40:499–519. doi: 10.1002/mus.21391. [DOI] [PubMed] [Google Scholar]
- 46.Jiang SD, Dai LY, Jiang LS. Osteoporosis after spinal cord injury. Osteoporos Int. 2006;17:180–92. doi: 10.1007/s00198-005-2028-8. [DOI] [PubMed] [Google Scholar]
- 47.Warden SJ, Bennell KL, Matthews B, Brown DJ, McMeeken JM, Wark JD. Quantitative ultrasound assessment of acute bone loss following spinal cord injury: a longitudinal pilot study. Osteoporos Int. 2002;13:586–92. doi: 10.1007/s001980200077. [DOI] [PubMed] [Google Scholar]
- 48.Garland DE, Adkins RH, Kushwaha V, Stewart C. Risk factors for osteoporosis at the knee in the spinal cord injury population. J Spinal Cord Med. 2004;27:202–6. doi: 10.1080/10790268.2004.11753748. [DOI] [PubMed] [Google Scholar]
- 49.Shields RK, Dudley-Javoroski S. Musculoskeletal plasticity after acute spinal cord injury: effects of long-term neuromuscular electrical stimulation training. J Neurophysiol. 2006;95:2380–90. doi: 10.1152/jn.01181.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Shields RK, Dudley-Javoroski S, Law LA. Electrically induced muscle contractions influence bone density decline after spinal cord injury. Spine (Phila Pa 1976) 2006;31:548–53. doi: 10.1097/01.brs.0000201303.49308.a8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Hanson AM, Harrison BC, Young MH, Stodieck LS, Ferguson VL. Longitudinal characterization of functional, morphologic, and biochemical adaptations in mouse skeletal muscle with hindlimb suspension. Muscle Nerve. 2013;48:393–402. doi: 10.1002/mus.23753. [DOI] [PubMed] [Google Scholar]
- 52.Lloyd SA, Lewis GS, Zhang Y, Paul EM, Donahue HJ. Connexin 43 deficiency attenuates loss of trabecular bone and prevents suppression of cortical bone formation during unloading. J Bone Miner Res. 2012;27:2359–72. doi: 10.1002/jbmr.1687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Pedersen BK, Steensberg A, Fischer C, Keller C, Keller P, Plomgaard P, et al. Searching for the exercise factor: is IL-6 a candidate? J Muscle Res Cell Motil. 2003;24:113–9. doi: 10.1023/a:1026070911202. [DOI] [PubMed] [Google Scholar]
- 54.Yoon JH, Song P, Jang JH, Kim DK, Choi S, Kim J, et al. Proteomic analysis of tumor necrosis factor-alpha (TNF-α)-induced L6 myotube secretome reveals novel TNF-α-dependent myokines in diabetic skeletal muscle. J Proteome Res. 2011;10:5315–25. doi: 10.1021/pr200573b. [DOI] [PubMed] [Google Scholar]
- 55.Hartwig S, Raschke S, Knebel B, Scheler M, Irmler M, Passlack W, et al. Secretome profiling of primary human skeletal muscle cells. Biochim Biophys Acta. 2014;1844:1011–7. doi: 10.1016/j.bbapap.2013.08.004. [DOI] [PubMed] [Google Scholar]
- 56.Henningsen J, Rigbolt KT, Blagoev B, Pedersen BK, Kratchmarova I. Dynamics of the skeletal muscle secretome during myoblast differentiation. Mol Cell Proteomics. 2010;9:2482–96. doi: 10.1074/mcp.M110.002113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Deshmukh AS, Cox J, Jensen LJ, Meissner F, Mann M. Secretome analysis of lipid-induced insulin resistance in skeletal muscle cells by a combined experimental and bioinformatics workflow. J Proteome Res. 2015;14:4885–95. doi: 10.1021/acs.jproteome.5b00720. [DOI] [PubMed] [Google Scholar]
- 58.Grube L, Dellen R, Kruse F, Schwender H, Stuhler K, Poschmann G. Mining the secretome of C2C12 muscle cells: data dependent experimental approach to analyze protein secretion using label-free quantification and peptide based analysis. J Proteome Res. 2018;17:879–90. doi: 10.1021/acs.jproteome.7b00684. [DOI] [PubMed] [Google Scholar]
- 59.Utvag SE, Iversen KB, Grundnes O, Reikeras O. Poor muscle coverage delays fracture healing in rats. Acta Orthop Scand. 2002;73:471–4. doi: 10.1080/00016470216315. [DOI] [PubMed] [Google Scholar]
- 60.Stein H, Perren SM, Cordey J, Kenwright J, Mosheiff R, Francis MJ. The muscle bed: a crucial factor for fracture healing: a physiological concept. Orthopedics. 2002;25:1379–83. doi: 10.3928/0147-7447-20021201-16. [DOI] [PubMed] [Google Scholar]
- 61.Landry PS, Marino AA, Sadasivan KK, Albright JA. Effect of soft-tissue trauma on the early periosteal response of bone to injury. J Trauma. 2000;48:479–83. doi: 10.1097/00005373-200003000-00018. [DOI] [PubMed] [Google Scholar]
- 62.Harry LE, Sandison A, Paleolog EM, Hansen U, Pearse MF, Nanchahal J. Comparison of the healing of open tibial fractures covered with either muscle or fasciocutaneous tissue in a murine model. J Orthop Res. 2008;26:1238–44. doi: 10.1002/jor.20649. [DOI] [PubMed] [Google Scholar]
- 63.Harry LE, Sandison A, Pearse MF, Paleolog EM, Nanchahal J. Comparison of the vascularity of fasciocutaneous tissue and muscle for coverage of open tibial fractures. Plast Reconstr Surg. 2009;124:1211–9. doi: 10.1097/PRS.0b013e3181b5a308. [DOI] [PubMed] [Google Scholar]
- 64.Gopal S, Majumder S, Batchelor AG, Knight SL, De Boer P, Smith RM. Fix and flap: the radical orthopaedic and plastic treatment of severe open fractures of the tibia. J Bone Joint Surg Br. 2000;82:959–66. doi: 10.1302/0301-620x.82b7.10482. [DOI] [PubMed] [Google Scholar]
- 65.Hamrick MW, McNeil PL, Patterson SL. Role of muscle-derived growth factors in bone formation. J Musculoskelet Neuronal Interact. 2010;10:64–70. [PMC free article] [PubMed] [Google Scholar]
- 66.Jahn K, Lara-Castillo N, Brotto L, Mo CL, Johnson ML, Brotto M, et al. Skeletal muscle secreted factors prevent glucocorticoid-induced osteocyte apoptosis through activation of β-catenin. Eur Cell Mater. 2012;24:197–210. doi: 10.22203/ecm.v024a14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Colaianni G, Cuscito C, Mongelli T, Oranger A, Mori G, Brunetti G, et al. Irisin enhances osteoblast differentiation in vitro. Int J Endocrinol. 2014;2014:902186. doi: 10.1155/2014/902186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Lee JY, Park SJ, Han SA, Lee SH, Koh JM, Hamrick MW, et al. The effects of myokines on osteoclasts and osteoblasts. Biochem Biophys Res Commun. 2019;517:749–54. doi: 10.1016/j.bbrc.2019.07.127. [DOI] [PubMed] [Google Scholar]
- 69.Kim BJ, Koh JM. Coupling factors involved in preserving bone balance. Cell Mol Life Sci. 2019;76:1243–53. doi: 10.1007/s00018-018-2981-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Reyes C, Hitz M, Prieto-Alhambra D, Abrahamsen B. Risks and Benefits of bisphosphonate therapies. J Cell Biochem. 2016;117:20–8. doi: 10.1002/jcb.25266. [DOI] [PubMed] [Google Scholar]
- 71.Canalis E, Giustina A, Bilezikian JP. Mechanisms of anabolic therapies for osteoporosis. N Engl J Med. 2007;357:905–16. doi: 10.1056/NEJMra067395. [DOI] [PubMed] [Google Scholar]
- 72.Gonzalez-Cadavid NF, Taylor WE, Yarasheski K, Sinha-Hikim I, Ma K, Ezzat S, et al. Organization of the human myostatin gene and expression in healthy men and HIV-infected men with muscle wasting. Proc Natl Acad Sci U S A. 1998;95:14938–43. doi: 10.1073/pnas.95.25.14938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Hamrick MW. A role for myokines in muscle-bone interactions. Exerc Sport Sci Rev. 2011;39:43–7. doi: 10.1097/JES.0b013e318201f601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Saunders MA, Good JM, Lawrence EC, Ferrell RE, Li WH, Nachman MW. Human adaptive evolution at Myostatin (GDF8), a regulator of muscle growth. Am J Hum Genet. 2006;79:1089–97. doi: 10.1086/509707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Giannesini B, Vilmen C, Amthor H, Bernard M, Bendahan D. Lack of myostatin impairs mechanical performance and ATP cost of contraction in exercising mouse gastrocnemius muscle in vivo. Am J Physiol Endocrinol Metab. 2013;305:E33–40. doi: 10.1152/ajpendo.00651.2012. [DOI] [PubMed] [Google Scholar]
- 76.Suh J, Lee YS. Myostatin inhibitors: panacea or predicament for musculoskeletal disorders? J Bone Metab. 2020;27:151–65. doi: 10.11005/jbm.2020.27.3.151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Dankbar B, Fennen M, Brunert D, Hayer S, Frank S, Wehmeyer C, et al. Myostatin is a direct regulator of osteoclast differentiation and its inhibition reduces inflammatory joint destruction in mice. Nat Med. 2015;21:1085–90. doi: 10.1038/nm.3917. [DOI] [PubMed] [Google Scholar]
- 78.Hamrick MW, Shi X, Zhang W, Pennington C, Thakore H, Haque M, et al. Loss of myostatin (GDF8) function increases osteogenic differentiation of bone marrow-derived mesenchymal stem cells but the osteogenic effect is ablated with unloading. Bone. 2007;40:1544–53. doi: 10.1016/j.bone.2007.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Hamrick MW, Samaddar T, Pennington C, McCormick J. Increased muscle mass with myostatin deficiency improves gains in bone strength with exercise. J Bone Miner Res. 2006;21:477–83. doi: 10.1359/JBMR.051203. [DOI] [PubMed] [Google Scholar]
- 80.Colaianni G, Cuscito C, Mongelli T, Pignataro P, Buccoliero C, Liu P, et al. The myokine irisin increases cortical bone mass. Proc Natl Acad Sci U S A. 2015;112:12157–62. doi: 10.1073/pnas.1516622112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Colaianni G, Mongelli T, Cuscito C, Pignataro P, Lippo L, Spiro G, et al. Irisin prevents and restores bone loss and muscle atrophy in hind-limb suspended mice. Sci Rep. 2017;7:2811. doi: 10.1038/s41598-017-02557-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Kawao N, Moritake A, Tatsumi K, Kaji H. Roles of irisin in the linkage from muscle to bone during mechanical unloading in mice. Calcif Tissue Int. 2018;103:24–34. doi: 10.1007/s00223-018-0387-3. [DOI] [PubMed] [Google Scholar]
- 83.Colaianni G, Cinti S, Colucci S, Grano M. Irisin and musculoskeletal health. Ann N Y Acad Sci. 2017;1402:5–9. doi: 10.1111/nyas.13345. [DOI] [PubMed] [Google Scholar]
- 84.Lee JY, Park SJ, Kim DA, Lee SH, Koh JM, Kim BJ. Muscle-derived lumican stimulates bone formation via integrin α2β1 and the downstream ERK signal. Front Cell Dev Biol. 2020;8:565826. doi: 10.3389/fcell.2020.565826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Norheim F, Raastad T, Thiede B, Rustan AC, Drevon CA, Haugen F. Proteomic identification of secreted proteins from human skeletal muscle cells and expression in response to strength training. Am J Physiol Endocrinol Metab. 2011;301:E1013–21. doi: 10.1152/ajpendo.00326.2011. [DOI] [PubMed] [Google Scholar]
- 86.Lee JY, Kim DA, Kim EY, Chang EJ, Park SJ, Kim BJ. Lumican inhibits osteoclastogenesis and bone resorption by suppressing Akt activity. Int J Mol Sci. 2021;22:4717. doi: 10.3390/ijms22094717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Jung TW, Hwang HJ, Hong HC, Yoo HJ, Baik SH, Choi KM. BAIBA attenuates insulin resistance and inflammation induced by palmitate or a high fat diet via an AMPK-PPARδ-dependent pathway in mice. Diabetologia. 2015;58:2096–105. doi: 10.1007/s00125-015-3663-z. [DOI] [PubMed] [Google Scholar]
- 88.Roberts LD, Bostrom P, O’Sullivan JF, Schinzel RT, Lewis GD, Dejam A, et al. β-Aminoisobutyric acid induces browning of white fat and hepatic β-oxidation and is inversely correlated with cardiometabolic risk factors. Cell Metab. 2014;19:96–108. doi: 10.1016/j.cmet.2013.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Hamrick MW, McGee-Lawrence ME. Blocking bone loss with l-BAIBA. Trends Endocrinol Metab. 2018;29:284–6. doi: 10.1016/j.tem.2018.03.005. [DOI] [PubMed] [Google Scholar]
- 90.Kitase Y, Vallejo JA, Gutheil W, Vemula H, Jahn K, Yi J, et al. β-aminoisobutyric acid, l-BAIBA, is a muscle-derived osteocyte survival factor. Cell Rep. 2018;22:1531–44. doi: 10.1016/j.celrep.2018.01.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Zhang Z, Zhang Y, Zhou Z, Shi H, Qiu X, Xiong J, et al. BDNF regulates the expression and secretion of VEGF from osteoblasts via the TrkB/ERK1/2 signaling pathway during fracture healing. Mol Med Rep. 2017;15:1362–7. doi: 10.3892/mmr.2017.6110. [DOI] [PubMed] [Google Scholar]
- 92.Kawao N, Morita H, Obata K, Tatsumi K, Kaji H. Role of follistatin in muscle and bone alterations induced by gravity change in mice. J Cell Physiol. 2018;233:1191–201. doi: 10.1002/jcp.25986. [DOI] [PubMed] [Google Scholar]
- 93.Baek K, Bloomfield SA. Beta-adrenergic blockade and leptin replacement effectively mitigate disuse bone loss. J Bone Miner Res. 2009;24:792–9. doi: 10.1359/jbmr.081241. [DOI] [PubMed] [Google Scholar]
- 94.Kurihara N, Bertolini D, Suda T, Akiyama Y, Roodman GD. IL-6 stimulates osteoclast-like multinucleated cell formation in long term human marrow cultures by inducing IL-1 release. J Immunol. 1990;144:4226–30. [PubMed] [Google Scholar]
- 95.Weitzmann MN, Cenci S, Rifas L, Brown C, Pacifici R. Interleukin-7 stimulates osteoclast formation by up-regulating the T-cell production of soluble osteoclastogenic cytokines. Blood. 2000;96:1873–8. [PubMed] [Google Scholar]
- 96.Reza MM, Subramaniyam N, Sim CM, Ge X, Sathiakumar D, McFarlane C, et al. Irisin is a pro-myogenic factor that induces skeletal muscle hypertrophy and rescues denervation-induced atrophy. Nat Commun. 2017;8:1104. doi: 10.1038/s41467-017-01131-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Chang JS, Kong ID. Irisin prevents dexamethasone-induced atrophy in C2C12 myotubes. Pflugers Arch. 2020;472:495–502. doi: 10.1007/s00424-020-02367-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Tsuchida K. Targeting myostatin for therapies against muscle-wasting disorders. Curr Opin Drug Discov Devel. 2008;11:487–94. [PubMed] [Google Scholar]
- 99.Vinel C, Lukjanenko L, Batut A, Deleruyelle S, Pradere JP, Le Gonidec S, et al. The exerkine apelin reverses age-associated sarcopenia. Nat Med. 2018;24:1360–71. doi: 10.1038/s41591-018-0131-6. [DOI] [PubMed] [Google Scholar]
- 100.Rai R, Ghosh AK, Eren M, Mackie AR, Levine DC, Kim SY, et al. Downregulation of the apelinergic axis accelerates aging, whereas its systemic restoration improves the mammalian healthspan. Cell Rep. 2017;21:1471–80. doi: 10.1016/j.celrep.2017.10.057. [DOI] [PMC free article] [PubMed] [Google Scholar]