

Abstract
Non-alcoholic fatty liver disease (NAFLD) is a clinically progressive illness that can advance from simple fatty liver to non-alcoholic hepatitis and liver fibrosis. Cirrhosis and hepatocellular carcinoma are two of the most common diseases caused by NAFLD. As there are no early disease biomarkers and no US Food and Drug Administration-approved medications, treatment for NAFLD is still focused on altering lifestyle and dietary habits, which makes it difficult to treat effectively. As a result, a novel treatment is urgently needed to prevent NAFLD progression. Calcium (Ca2+) channels regulate intracellular Ca2+ homeostasis via the mediation of Ca2+ flow. Previous studies have reported that Ca2+ channel expression varies throughout the development and progression of NAFLD, which results in the dysregulation of intracellular Ca2+ homeostasis, endoplasmic reticulum stress, mitochondrial dysfunction and autophagy suppression, all of which contribute to NAFLD progression. Several types of Ca2+ channels (including two-pore segment channel 2, transient receptor potential, inositol triphosphate receptor, voltage-dependent anion channel 1, store-operated Ca2+ entry, purinergic receptor X7 and potassium Ca2+-activated channel subfamily N member 4) have been identified as potential targets for preventing NAFLD development and controlling intracellular Ca2+ homeostasis. To achieve this, these channels can be blocked or activated, which exerts anti-steatotic, anti-inflammatory, anti-fibrotic and other effects, which ultimately prevents the development of NAFLD. In the present review NAFLD therapeutics and the treatments that target Ca2+ channels that are currently being developed were examined.
Keywords: non-alcoholic fatty liver disease, calcium channels, endoplasmic reticulum stress, mitochondrial dysfunction, therapeutic strategies
1. Introduction
Non-alcoholic fatty liver disease (NAFLD) is the excessive accumulation of lipids in hepatocytes and is a result of a combination of circulating free fatty acids, de novo adipogenesis and dietary fat intake. NAFLD is diagnosed following the exclusion of other causes of hepatic steatosis, such as excessive alcohol consumption or other diseases (1,2). NAFLD is one of the most common chronic liver diseases worldwide and the incidence of NAFLD in Europe and America is >20% (3). According to the pathological status of the liver, NAFLD can be divided into simple fatty liver and non-alcoholic steatohepatitis (NASH), which will eventually develop into liver fibrosis, cirrhosis and hepatocellular carcinoma (HCC) (1,4). Extrahepatic problems, including cardiovascular disease, obesity, type 2 diabetes, dyslipidemia and neurodegenerative conditions, are frequently associated with NAFLD. It has been reported that as the incidence of obesity and diabetes rise the prevalence of NAFLD also increases (5,6).
The pathophysiology of NAFLD is complex and involves free fatty acid accumulation, liver inflammation, oxidative stress, lipid peroxidation and hepatocyte damage (7). The double-hit model was previously considered to be the most widely accepted and popular NAFLD theory. In this model the first strike is insulin resistance (IR), which causes fat to build up in the liver, and the second blow is lipotoxicity, which causes mitochondrial dysfunction, oxidative stress and endoplasmic reticulum (ER) stress, as a result of fat accumulation. As a result, the vulnerability of the liver to inflammatory necrosis and fibrosis increases and inflammatory damage leads to NASH, liver fibrosis, cirrhosis and HCC, which hastens NAFLD progression (8). It is now widely accepted that multiple factors are involved in the development of NAFLD (9,10). As there is presently no satisfactory therapeutic approach for NAFLD due to its uncertain pathophysiology, NAFLD therapeutics are still dependent on improving the lifestyle and changing the eating habits of individuals (11,12). Overall, the prevalence of NAFLD has increased over the past few decades, which imposes an increasingly severe economic burden (13). It has also been reported that NAFLD will become the leading indication for liver transplantation within a decade (14). Therefore, it is important to understand in detail the regulatory mechanism of lipid metabolism in hepatocytes and to develop novel therapeutic strategies for the control of NAFLD.
Ion channels are polymeric proteins that generate ion selective holes in the plasma or organelle membrane in response to certain stimuli, such as membrane potential or ligand binding (15). Calcium (Ca2+) channels are highly selective for Ca2+ permeability and mainly mediate Ca2+ flow, which thereby regulates Ca2+ homeostasis inside and outside the cell and therefore it serves an important role in specific physiological functions. Ca2+ channels are mainly divided into the following two types: i) Voltage-gated Ca2+ channels, including T, L, N, P/Q and R types; and ii) non-voltage-gated Ca2+ channels, including transient receptor potential (TRP) and Orai calcium release-activated calcium modulator (Orai) families and their partners, stromal interaction molecules (STIMs) (16). Ca2+ channels are distributed in the cell membranes and organelles of various organs and regulate numerous physiological functions of the body (Table I). Increasing evidence has demonstrated that Ca2+ channels serve a key role in the pathological process of liver diseases, including NAFLD. These channels mediate pathophysiological changes via the upregulation or downregulation of expression levels, such as those of diagnostic markers of early liver cancer and drug treatment targets (17,18).
Table I.
Localization and function of Ca2+ channels.
| Name | Location | Function |
|---|---|---|
| TPC2 | Lysosome | Ca2+ release from the lysosome |
| TRPV1 | Plasma membrane | Ca2+ entry from the extracellular space to the cytoplasm |
| TRPV4 | Plasma membrane | Ca2+ entry from the extracellular space to the cytoplasm |
| TRPM2 | Plasma membrane | Ca2+ entry from the extracellular space to the cytoplasm |
| IP3R1 | ER membrane | Ca2+ release from the ER to the cytoplasm |
| IP3R2 | ER membrane | Ca2+ release from the ER to the cytoplasm |
| VDAC1 | Mitochondrial outer membrane | Entry of Ca2+ and other metabolites |
| Orai | Plasma membrane | Ca2+ entry from the extracellular space to the cytoplasm |
| P2X7R | Plasma membrane | Ca2+ and Na+ entry from the extracellular space to the cytoplasm, and K+ outflow from the cytoplasm to the extracellular space |
| KCa3.1 | Plasma membrane | Ca2+ entry from the extracellular space to the cytoplasm, and K+ outflow from the cytoplasm to the extracellular space |
Ca2+, calcium; ER, endoplasmic reticulum; Orai, Orai calcium release-activated calcium modulator; P2X7R, purinergic receptor X7; K+, potassium.
The present review aimed to summarize the regulatory mechanism of Ca2+ channels and discuss its role in the development and progression of NAFLD. Furthermore, the latest research progress in drugs that target Ca2+ channels to treat NAFLD was examined. Moreover, the present review aimed to propose novel potential therapeutic strategies involving Ca2+ channels for the treatment of NAFLD and to highlight current issues and the direction of future research in this field. The present review provided new opportunities for the prevention and treatment of NAFLD.
The databases used in the present review included PubMed, Web of Science and Embase. The key words used included 'non-alcoholic fatty liver disease', 'calcium channels', 'endoplasmic reticulum stress', 'mitochondrial dysfunction', and 'therapeutic strategies'. The references included studies performed from 1998-2021.
2. Physiological role of Ca2+ in the liver
The liver is a complex organ composed of a variety of different types of cells, which serves a crucial role as the metabolic center of the body, including in the metabolism of lipids, carbohydrates, various drugs and toxins (19). Ca2+ is a ubiquitous secondary messenger that regulates a variety of liver functions, including lipid and carbohydrate metabolism as well as bile secretion and cholestasis. Furthermore, the balance of Ca2+ signals between the nucleus, cytoplasm and mitochondria controls liver regeneration following injury and is also related to Ca2+ channel expression and distribution in different organelles (20,21). Changes in intracellular Ca2+ concentration are affected by the influx of extracellular Ca2+ and the release of Ca2+ from intracellular Ca2+ stores, such as the ER, mitochondria and lysosomes. When cells are excited by external stimuli, various Ca2+ channels are opened or closed, which results in the dysregulation of Ca2+ homeostasis (22,23). Previous studies have reported that the Ca2+ signaling pathway is a key regulator of nutrient uptake, metabolism and utilization (24,25). Therefore, if Ca2+ homeostasis in liver cells is disrupted, a series of different metabolic damage and liver regeneration disorders will be caused, including NAFLD.
3. Dysregulation of Ca2+ channel expression and the disturbance of Ca2+ homeostasis in NAFLD
Ca2+ maintains a dynamic balance between internal and external cells and organelles and regulates normal physiological functions (Fig. 1). However, Ca2+ channels partially regulate Ca2+ levels and once channel expression levels are impaired, including the upregulation, downregulation and even channel defects, a series of pathophysiological changes occur. In NAFLD the imbalance of Ca2+ in the cytoplasm, organelles, including the mitochondria and ER, and the nucleus is one of the contributing factors that promotes the development of hepatic steatosis (26). The ER and mitochondria are important organelles for lipid synthesis and glycolipid metabolism in the liver and therefore serve an important role in controlling lipid synthesis and metabolism in the liver (27). The ER is the main Ca2+ reservoir that releases Ca2+ under the stimulation of various hormones. Matrix interaction molecules, STIM1 and STIM2, are Ca2+ sensors of the ER and detect the reduction of ER Ca2+ (28). When ER Ca2+ is depleted, STIMs rapidly connect the ER and the plasma membrane, causing the plasma membrane to recruit and activate Orai Ca2+ channels, which result in Ca2+ release and activation of the Ca2+ release-activated channel. Ca2+ flows from extracellular space via the Orai channel, which results in increased cytoplasmic Ca2+ levels and downstream signaling. This process is called storage operation Ca2+ entry (SOCE). The excess Ca2+ is subsequently pumped back into the ER to replenish the deficiency and this therefore maintains a dynamic Ca2+ equilibrium (29). Liver hyperlipidemia leads to an imbalance between ER membrane lipids and phosphatidylethanolamine, which impairs the sarco/ER Ca2+ ATPase (SERCA) function of the ER Ca2+ uptake pump and decreases the opening of the inositol triphosphate receptor (InsP3R) Ca2+ channel. These disturbances lead to increased cytoplasmic Ca2+ levels and decreased ER Ca2+ levels (30-32). Furthermore, in the ER certain Ca2+-dependent molecular chaperones, including calin and calreticulin, control the folding of secretory and membrane proteins, such as insulin receptors. Therefore, low Ca2+ levels in the ER can lead to an excess of misfolded proteins and this therefore results in ER stress. Furthermore, in response to ER stress, hepatocytes activate the unfolded protein response (UPR) that causes IR via different signaling pathways (24).
Figure 1.

Schematic of intracellular Ca2+ flow. When GPCRs are activated, the action of PLCs leads to the formation of InsP3. Binding to the InsP3R triggers the release of Ca2+ from the ER to the cytoplasm. Subsequent isolation of Ca2+ from the STIM protein leads to its interaction with Orai, which allows Ca2+ to enter the cell from the extracellular space. Furthermore, cytoplasmic Ca2+ is absorbed into the ER via SERCA activity. The ER and mitochondria function via MAMs, whereby the chaperone molecule GRP75 connects InsP3R and VDAC, which allows Ca2+ to be transported from the ER to the mitochondria. Tethering proteins Mfn1/2 regulate the stability of MAM. Ca2+ enters the mitochondrial matrix via the MCU complex of the inner mitochondrial membrane. Ca2+, calcium; GPCR, G-protein-coupled receptor; PLC, phospholipase C; InsP3, inositol triphosphate; InsP3R, InsP3 receptor; ER, endoplasmic reticulum; STIM, stromal interaction molecule; Orai, Orai calcium release-activated calcium modulator; SERCA, sarco/ER Ca2+ ATPase; MAM, mitochondria-associated membrane; GRP75, glucose-regulated protein 75; VDAC, voltage-dependent anion channel; Mfn1/2, mitofusin 1/2; MCU, mitochondrial Ca2+ uniporter.
Moreover, the concentration of Ca2+ in the mitochondria is equivalent to that in the cytoplasm under normal physiological conditions. The close proximity between the ER and mitochondria generates Ca2+ microdomains that allow the rapid transport of Ca2+ to the mitochondria when needed (33). Ca2+ absorption is achieved by a voltage-dependent anion channel (VDAC) in the outer membrane of mitochondria that is connected with the ER InsP3R1 via a molecular partner, glucose-regulated protein 75 (34). Mitochondrial Ca2+ concentration is controlled via the coordinated interaction between the mitochondrial Ca2+ uniporter (MCU) complex and the sodium or hydrogen (H+)/Ca2+ reverse transporters, solute carrier family 8 member B1 and leucine zipper and EF-hand containing transmembrane protein 1. The fine-regulation of mitochondrial Ca2+ levels according to cellular needs contributes to the regulation of numerous liver functions, including lipid and carbohydrate metabolism, proliferation and apoptosis (24). In compensatory diseases with chronic elevated cytoplasmic Ca2+ levels, Ca2+ uptake via the MCU increases and the exit via the Ca2+ transporter becomes saturated. The overload of mitochondrial Ca2+ buffer capacity leads to an increase in mitochondrial Ca2+ levels, which can lead to mitochondrial dysfunction and increase reactive oxygen species levels. Oxidative stress promotes hepatocyte apoptosis via the consumption of ATP, NAD and glutathione, as well as via the damage of DNA, lipids and proteins (24,35).
Lysosomes are also important for intracellular Ca2+ storage and H+-ATPase establishes a H+ gradient on the lysosome membrane that provides energy for Ca2+ entry via the Ca2+/H+ exchanger (36). Lysosomal Ca2+ release is triggered via numerous Ca2+ mobilization messengers, including nicotinic acid adenine dinucleotide phosphate (NAADP) and phosphatidylinositol 3,5-diphosphate, via Ca2+ permeable channels, including two-pore channels (TPCs) and TRP mucilipids (TRPMLs) (26). Furthermore, lysosomal Ca2+ is a key regulator of autophagy and the process of lipophagy decomposition of intracellular lipid drops is named lipid phagocytosis (37). Autophagy is important in NASH and it has previously been reported that drugs enhancing autophagy have therapeutic potential for the treatment of NASH (38). Therefore, targeting Ca2+ channels on lysosomes to regulate Ca2+ levels may also be a potential therapeutic approach for the treatment of NAFLD.
Lipid accumulation in liver cells induces changes in Ca2+ signaling, mainly via the decreased concentration of ER Ca2+ and the increased concentration of Ca2+ in the cytoplasmic matrix [(Ca2+)cyt] and mitochondrial matrix (39). It can also lead to the inhibition of lipid autophagy (40), ER stress (41), mitochondrial dysfunction (2) and oxidative stress, which results in increased liver fat accumulation, steatosis and the progression of NASH (Fig. 2) (27,42). These aforementioned studies have therefore indicated that Ca2+ signaling serves an important role in NAFLD. By directly targeting Ca2+ channels that regulate Ca2+ levels, novel NAFLD therapeutics may be developed to prevent steatosis and the transformation of inflammation into fibrosis.
Figure 2.
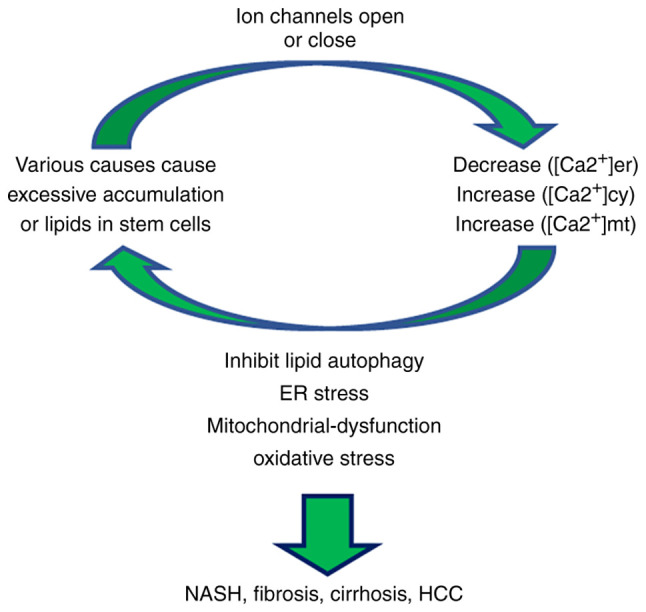
Ca2+ homeostasis. Lipid accumulation dysregulates Ca2+ homeostasis in the intracellular matrix, mitochondria and endoplasmic reticulum via affecting ion channel permeability. This leads to further lipid accumulation, which ultimately promotes the further development of NAFLD. Ca2+, calcium; NAFLD, non-alcoholic fatty liver disease.
4. Ca2+ channels in the development and progression of NAFLD and as an NAFLD therapeutic target
Ca2+ channels serve an important role in the development and progression of NAFLD, including via their overexpression and low expression. Therefore, Ca2+ channels may be used as a potential target for the diagnosis, prevention and treatment of NAFLD. The following is a summary of the mechanism of action and/or treatment strategies of several types of Ca2+ channels in NAFLD (Table II) (43-62).
Table II.
Role of Ca2+ channels in the development of NAFLD.
| Name | Role and therapeutic target | Therapeutic strategy | (Refs.) |
|---|---|---|---|
| TPC2 | Downregulated expression; promotes the progression | Inhibition | (43) |
| TRPV1 | After activation inhibits the progression | Activation | (44,45) |
| TRPV4 | Upregulated expression; promotes the progression | Inhibition | (46,47) |
| TRPM2 | Upregulated expression; promotes the progression | Inhibition | (48,49) |
| IP3R1 | Upregulated expression; promotes the progression | Inhibition | (50,51) |
| IP3R2 | Downregulated expression; promotes the progression | Activation | (52) |
| VACD1 | Upregulated expression; promotes the progression | Inhibition | (53,54) |
| SOCE | Downregulated expression; promotes the progression | Activation | (55,56) |
| Orai1 | Upregulated expression; promotes the progression | Inhibition | (57,58) |
| P2X7R | Upregulated expression; promotes the progression | Inhibition | (59,60) |
| KCa3.1 | Upregulated expression; promotes the progression | Inhibition | (61,62) |
Ca2+, calcium; NAFLD, non-alcoholic fatty liver disease; SOCE, storage operation Ca2+ entry.
TPC2 defects contribute to NAFLD progression
TPCs are endolysomal ion channels. There are three TPC subtypes. TPC1 and TPC2 are ubiquitous, whereas TPC3 is not present in numerous animal genomes (63). TPCs are mainly located in acidic organelles, such as lysosomes, with TPC2 expression being predominantly lysosomal and TPC1 having a wider distribution within the endolysomal system. TPC1 is found in lysosomes and early and recycling endosomes (43,63). Previous studies have reported that TPCs mediate the release of Ca2+ in lysosomes following activation via NAADP, which regulates Ca2+ signal stabilization to control a series of Ca2+-dependent events (64). In NAFLD, the opening of the TPC2 channel initiates the final fusion between late endosomes and lysosomes, which mediates lipid degradation transport. However, TPC2 deficiency leads to impaired transport that promotes hepatic cholesterol accumulation and hyperlipoproteinemia, which thereby significantly contributes to the development of NASH (43). Furthermore, in a previous study it was demonstrated that in the liver cells of TPC2-knockout embryonic mice, substances and receptors transported along the degradation pathway, such as low-density lipoprotein or epidermal growth factor (EGF)/EGFR, accumulate rapidly in intracellular vesicles. This resulted in increased cholesterol accumulation and liver transaminase, which suggested that the TPC2-deficient mice exhibited cholesterol overload and liver damage consistent with NAFLD (43). Furthermore, high Ca2+ can cause lysosome damage, which leads to impaired lipid degradation, liver cholesterol accumulation and hyperlipidemia, which further progresses NAFLD into NASH (17,43). Therefore, it can be hypothesized that regulating the high expression of TPC2 may prevent NAFLD development.
TRP channels
TRP channels are a superfamily of univalent and bivalent cation osmotic ion channels. More than 30 channels have been identified, including TRP channel, TRP vanilloid (TRPV), TRP melastatin (TRPM), TRP subfamily A, TRP polycystic, TRPML and TRP no mechanoreceptor potential C (65). TRP channels mainly exist on the plasma membrane of metabolically active tissues, including the liver, gastrointestinal tract, brain, pancreas and adipose tissue (66). They serve an important role in sensory function (sight, hearing, taste, smell, pain, mechanical and thermal sensations), homeostasis (absorption and reabsorption of ions and fluid flow), as well as cell survival (67,68). TRP channels have the following three important characteristics: i) Ca2+ permeability; ii) thermal sensation; and iii) mechanical sense. Moreover, TPC channels have a high permeability for Ca2+ ions, regulate the intracellular Ca2+ steady-state and affect the energy intake, storage and consumption of the body (65,66). It has previously been reported that the following TRP channels induce the development and progression of NAFLD via the mediation of intracellular Ca2+ homeostasis, which could help identify new treatment strategies for NAFLD.
Following activation TRPV1 upregulates the expression of uncoupling protein 2 (UCP2) and decreases peroxisome proliferator activated receptor δ (PPARδ) and autophagy
TRPV1 is found in mouse preadipocytes and adipocytes, as well as in human and mouse visceral adipose tissue. It mediates the elevation of Ca2+ in fat cells and therefore significantly inhibits adipogenesis and reduces the accumulation of triglycerides (69). Capsaicin, the main irritant in pepper, is an effective agonist of TRPV1 (70) that blocks adipogenesis and obesity in anterior fat cells in vivo and in vitro via the activation of TRPV1 channels (71). Previous studies have reported that NAFLD is often associated with obesity, which is specifically reflected in autophagy disorders during obesity that contribute to the pathological progression of fatty liver disease, such as steatohepatitis and HCC (71-73). In an in vitro model of NAFLD induced via a high-fat diet (HFD), capsaicin treatment was demonstrated to reduce liver steatosis and inflammation, but not in TRPV1-knockout mice (44,45). In an in vitro model of NAFLD induced by HFD, UCP2 expression was upregulated in the liver of wild-type mice following TRPV1 activation, but not in TRPV1-knockout mice. UCP2 promotes liver β-oxidation and lipid metabolism (74). These aforementioned results suggested that the activation of TRPV1 prevents the development of fatty liver and may be associated with increased hepatic β-oxidation, mediated via UCP2 (45,75). Another study reported that a long-term HFD significantly reduced the expression of liver PPARδ and autophagy molecules in wild-type mouse liver tissue (44). Autophagy regulates intracellular lipid levels by removing lipid droplets, which consequently enhances autophagy, liver steatosis and IR, and leads to improvements in HFD-treated mice (76). It was also demonstrated that TRPV1 activation via capsaicin treatment promoted lipolysis and enhanced autophagy, whereas dietary capsaicin did not have this effect in TRPV1-deficient mice. Therefore, capsaicin also inhibits NAFLD by increasing the expression levels of PPARδ, which causes autophagy enhancement (44). In conclusion, the activation of TRPV1 potentially has clinical value in the treatment of the development and progression of NAFLD.
TRPV4 inhibits the action of cytochrome P450 family 2 subfamily E member 1 (CYP2E1) via the activation of Kupffer cell nitric oxide (NO) synthase 3 (NOS3) and NO release via the paracrine system
TRPV4 is a Ca2+ channel that is widely distributed in the kidneys, lungs, brain, bladder, fat, liver, heart, skin, airway smooth muscle cells, vascular endothelial cells, chondrocytes, osteoclasts and other tissues. TRPV4 is activated by numerous physical and chemical stimuli, including heat, mechanical stress, arachidonic acid and its derivatives (77). Certain studies have reported that TRPV4 has a pro-inflammatory effect on adipose tissue. Its inhibition by drugs will lead to an increase in the thermogenic gene program and a decrease in adipose tissue inflammation. This indicates that TRPV4 has therapeutic benefits for obesity and metabolic diseases (78). TRPV4 can also increase autophagy and inhibit stellate cell apoptosis (79). Furthermore, TRPV4 expression is significantly increased in hepatic fibrosis tissues (80). CYP2E1 is a cytochrome P450 enzyme, which can lead to lipid peroxidation following its activation and serves a pro-inflammatory role in NAFLD and enhances the progression of NAFLD (46). In progressive NAFLD, the promoter region of the TRPV4 gene is methylated, which inhibits TRPV4 expression and therefore reduces the levels of channel proteins associated with the progression of NAFLD (47). In a previous study, the NAFLD model was established in TRPV4-knockout mice and the deterioration of NAFLD was observed 1 week following CYP2E1 activation. This was exhibited via necrosis, liver cell swelling, the upregulation of inflammatory factor expression levels, Kupffer cell activation and an increase in IL-1β levels. Furthermore, the mRNA and protein expression levels of the damage-related nuclear DNA binding protein, high mobility group box 1 (HMGB1), were also increased, which promoted the further development of NAFLD. This aforementioned study therefore concluded that TRPV4 serves a protective role in the liver (47). This protective effect of TRPV4 may block the role of CYP2E1 via the activation of Kupffer cell NOS3 and the release of NO via the paracrine system (47). Therefore, as an endogenous defense molecule, TRPV4 has the potential to treat NAFLD.
Salidroside prevents the further development of NAFLD via inhibition of the TRPM2/Ca2+/Ca2+/calmodulin-dependent protein kinase II (CaMKII) signaling pathway
TRPM2 is a non-selective cationic channel that mediates Ca2+ transmission and is expressed in the skin, brain, pancreas, spleen, kidney and immune cells (65). However, its presence in the liver has only been confirmed at the mRNA expression level (81). TRPM2 is activated via temperature, oxidative stress and intracellular endogenous ligands, of which oxidative stress is the most important (48,65). It also regulates Ca2+ signal transduction in tissues and mediates various pathophysiological processes. TRPM2-mediated Ca2+ entry has been reported to contribute to drug-induced hepatotoxicity and the progression of NAFLD into cirrhosis, fibrosis and HCC (49). In a previous study, following the establishment of NAFLD models in vitro with palmitic acid (PA), the expression of TRPM2 was increased and cytoplasmic Ca2+ levels were significantly increased. This subsequently resulted in the downstream phosphorylation of CaMKII. Furthermore, treatment with salidroside reduced the accumulation of fat droplets in liver L02 cells of PA-treated mice in a dose-dependent manner, which was potentially a result of the inhibition of TRPM2 channel activation in NAFLD liver cells. Moreover, this reduced the progression of NAFLD-related disease symptoms via reducing Ca2+ influx-induced phosphorylated-CaMKII downregulation. These results suggested that the TRPM2/Ca2+/CaMKII signaling pathway further leads to lipid accumulation, mitochondrial damage and ER stress, which may exacerbate the progression of NAFLD. This can lead to NASH and progression to cirrhosis and HCC (48). Therefore, TRPM2 inhibitors may be useful therapeutic targets for NAFLD treatment (48,49,82).
1,4,5-Trisphosphate receptor (IP3R)1 deletion inhibits the development of NAFLD, whereas IP3R2 deletion can promote the development of NAFLD
The IP3R is a type of important intracellular Ca2+ release channel that mediates a series of pathophysiological processes by participating in the regulation of intracellular Ca2+ levels (83). There are three main types, named IP3R1, IP3R2 and IP3R3. Moreover, IP3R1 and IP3R2 are both expressed in liver cells, whereas IP3R3 is physiologically deficient. IP3R1 is mainly distributed in the ER, which controls mitochondrial Ca2+ signaling and lipid metabolism via connecting with VDAC on the outer membrane of the mitochondria. IP3R2 is concentrated near the apical membrane and regulates the bile solute secretion of liver cells (50). Therefore, IP3R1 and IP3R2 serve different roles in the progression of NAFLD. It has previously been reported that IP3R1 expression is increased in the mitochondria-bound ER of HFD mice, which leads to mitochondrial dysfunction and the disruption of metabolic homeostasis (84). Furthermore, the results of this previous study suggested that IP3R1 may promote the development of NAFLD. A HFD diet was used to induce fatty liver in mice. When compared with the control group, IP3R1-knockout mice exhibited reduced triglycerides in the serum and liver and lipid droplet accumulation. Moreover, liver histological examinations demonstrated that IP3R1-knockout mice were free from steatosis (51). Further investigations also detected a significant decrease in Ca2+ signaling in the mitochondria of IP3R1-deficient mice (51). Therefore, this aforementioned study concluded that IP3R1-knockout mice have reduced mitochondrial Ca2+ signal impairment, reduced hepatic triglycerides and reduced lipid droplet formation and are resistant to the development of fatty liver disease (51). In a study, hepatocytes were treated with PA to establish an NAFLD model in vitro. The increased expression of IP3R1, mitochondrial dysfunction and apoptosis were detected. The results also demonstrated that the inhibition of IP3R1 expression alleviated the decrease in mitochondrial membrane potential and of mitochondrial superoxide accumulation in hepatocytes following PA treatment. This may have been due to the increased phosphorylation of Tyr353 in IP3R1 via the tyrosine kinase signaling pathway that led to the increased stability of the IP3R1 protein. Therefore, mitochondrial Ca2+ overload may induce mitochondrial dysfunction in hepatocytes (50). In summary, the specific inhibition of IP3R1 channels to reduce mitochondrial dysfunction in hepatocytes may be a potential novel clinical strategy for the treatment of NAFLD and the prevention of its further development.
In a previous study, liver cell models of NAFLD were constructed using a HFD. c-Jun expression was reported to be elevated, whereas IP3R2 expression was significantly decreased (52). Moreover, liver biopsies from patients with simple steatosis and NASH were also analyzed and the results were consistent with the in vitro models (52). Compared with normal cells, IP3R2-deficient cells displayed a significantly reduced nuclear Ca2+ signal amplitude, which suggested that nuclear Ca2+ signaling may be dependent on inositol 1,4,5-trisphosphate receptor type 2 channel activation (52). [Ca2+]cyt is known to be essential for cell proliferation (85). Further research has reported significant diffuse steatosis and impaired hepatocyte regeneration in IP3R2-deficient livers following a hepatectomy (52). These aforementioned results suggested that increased c-Jun expression levels may potentially negatively regulate IP3R2 expression in the livers of patients with NAFLD. This may therefore affect nuclear Ca2+ and lead to impaired liver regeneration and cell proliferation, which eventually causes the progression of NAFLD (52). Although the specific underlying mechanism remains to be studied further, it can be hypothesized that the factors regulating IP3R2 expression have potential clinical significance in the treatment of NAFLD.
VDAC1 expression is upregulated in NAFLD-associated HCC
VDAC1 is an anionic channel that is located on the outer membrane of the mitochondria and can form hydrophilic channels to regulate the passage of anions, cations, ATP and other metabolites into and out of the mitochondria. It also interacts with numerous proteins that are also involved in lipid metabolism and cholesterol transport. VDAC1 serves an important role in regulating cell metabolism, maintaining intracellular Ca2+ homeostasis, apoptosis and necrosis (86,87). Cardiolipin (CL), the signature phospholipid of the inner mitochondrial membrane (IMM), serves an important role in maintaining the structure and function of mitochondria (88). CL deletion occurs in various metabolic disorders, including NAFLD. Certain studies have analyzed patients with NAFLD-driven HCC, and have demonstrated that VDAC1 is dysregulated in NAFLD-driven HCC. Furthermore, immunohistochemical analysis has previously demonstrated that compared with non-tumor tissues, VDAC1 expression levels in HCC tissues are upregulated and gradually increase with the progression of the tumor (53,54). Furthermore, high levels of VDAC1 expression are associated with poor clinical outcomes, possibly because VDAC1 is significantly positively correlated with protein tyrosine phosphatase mitochondrial 1 and negatively correlated with the CL remodeling enzyme tafazzin. This results in changes to CL composition and damages the stability of the IMM structure, which ultimately leads to mitochondrial dysfunction. VDAC1 therefore promotes the development of NAFLD to HCC (54). Although the specific underlying mechanism remains unclear, the targeted inhibition of VDAC1 may be a potential therapeutic approach to prevent the progression of NAFLD to HCC.
Inhibition of SOCE and SERCA2b leads to the development of NAFLD, whereas inhibition of Orai1 inhibits this effect
SOCE, composed of STIM1, is located on the ER and the Orai1 protein is located on the cell membrane and is an important channel that mediates the entry of extracellular Ca2+ into cells (89). As a Ca2+ sensor, when STIM1 senses a decrease in ER Ca2+ concentration it will undergo rapid activation reactions, such as translocation and polymerization, and will subsequently combine with Orai1 on the plasma membrane, leading to the activation of the Orai1 pore, which results in the entry of intracellular Ca2+ (89,90). SOCE signals control a variety of functions, including gene expression, migration, proliferation and apoptosis. Dysregulation of STIM and Orai1 protein expression levels can lead to SOCE dysfunction and consequently induce pathophysiological changes (90). Ca2+ entry via the SOCE signaling pathway induces cAMP production, protein kinase C (PKC) activation, hormone-sensitive lipase activation and enhanced lipolysis (39). The accumulation of cholesterol during steatosis is associated with ER Ca2+ depletion, which leads to the induction of UPR and apoptosis (91) and participates in the regulation of lipid metabolism (92). Previous studies have reported that lipid accumulation induces the inhibition of SOCE and SERCA2b, the major isoform of SERCA protein in the liver, and reduces the entry of Ca2+ via the activation of PKC-mediated phosphorylation of the Orai1 protein. As the ER stress response is related to Ca2+ homeostasis, a decrease of Ca2+ concentration in the ER induces the ER stress response and leads to a decrease in protein folding, as well as protein chaperone activity and glucose tolerance. This therefore stimulates the synthesis of diacylglycerol and triacylglycerol (55,56). This process ultimately intensifies lipid accumulation and leads to IR, which promotes NAFLD progression to NASH, liver fibrosis and even HCC. Furthermore, the inhibition of SOCE and subsequent increase in intracellular lipid accumulation are associated with increased lipid autophagy (4,55,56). Previous research on the Orai1 protein has reported that Orai1 expression regulates de novo fat formation (57). Hepatic BRL-3A cells were previously treated with high concentrations of nesfatins to construct a NAFLD cell model. The expression levels of Orai1 and NF-κB p65 were significantly upregulated, which promoted ER and oxidative stress and led to the inflammatory response of hepatic lipid deposition. Treatment with Orai1 inhibitor 2-aminoethoxydiphenyl borate and NF-κB inhibitor wogonin reduced these effects (57,58). Therefore, it can be hypothesized that the activation of SOCE or the reduction of SOCE inhibition increases Ca2+ in the ER, which may reduce ER stress and decrease lipid accumulation. However, lipid deposition may also be reduced via the regulation of Orai1 protein expression. The aforementioned strategies may serve as potential therapeutic approaches to prevent NAFLD progression.
Deletion of the purinergic receptor X7 (P2X7R) against fatty liver injury and fibrosis
P2X7R, a member of the purinergic receptor family, is an ion-type ligand-gated ion channel that is expressed on the membrane of hepatic cells and is activated via ATP to regulate liver metabolic processes, such as the insulin response, glycogen and lipid metabolism and bile secretion (93). P2X7R has been reported to increase NADPH oxidase activity, increase the expression of major histocompatibility complex II in Kupffer cells and participate in the regulation of inflammation (59). Previous studies have demonstrated that P2X7R expression was increased in hepatocytes, Kupffer cells and hepatic sinusoidal endothelial cells via the construction of an in vitro NASH model (60,94). P2X7R is associated with autophagy, disruption of normal lysosomal function, as well as autophagosome release into the extracellular matrix of microglia, which ultimately leads to inflammation (95). Autophagy is one of the key characteristics of NASH and drugs that enhance autophagy have been reported to have therapeutic potential for the treatment of NASH (38). Moreover, P2X7R is a key regulator of autophagy in NASH (96). LC3B depletion, lysosome-associated membrane protein 2 and heat shock cognate protein 70 increasing, which act as autophagy markers, have been reported in mice with the P2X7R gene deletion. Furthermore, LI-1, IFN-α and HMGB1 release was demonstrated to be subsequently reduced, which resulted in enhanced autophagy and reduced inflammation (60). Other studies have reported that liver cells treated with a HFD and carbon tetrachloride (CCL4) exhibited improved apoptosis, as well as reduced inflammation and fibrosis in mice with a P2X7R deletion (59,60). Furthermore, P2X7R activated Kupffer cells and increased the production of TNF-α and monocyte chemotactic protein-2 in HFD mice treated with CCL4 (59). In summary, P2X7R deletion mice were protected from the damage of steatosis and fibrosis. However, the exact molecular underlying mechanism of P2X7R is unclear. It can be hypothesized that enhancing lipid autophagy via the inhibition of P2X7R may be a novel therapeutic approach for the treatment of NASH.
Senicapoc inhibits the KCa3.1 signaling pathway and serves anti-inflammatory and antisteatotic roles
Medium conductance Ca2+-activated potassium (K+) channels (KCa3.1) are widely expressed throughout the body, mainly in hematopoietic cells, the gastrointestinal tract, lungs, endocrine glands, exocrine glands, vascular endothelial cells, fibroblasts and proliferating neointimal vascular smooth muscle cells (97). Via the promotion of K+ outflow from hyperpolarized cell membranes, Ca2+ entry is promoted, which affects cell proliferation, migration and vascular resistance (61). Triarylmethane-34, an inhibitor of KCa3.1, can reduce the proliferation of stellate cells via the induction of cell cycle arrest and reduces the gene expression of TGF-β1-induced collagen, α-smooth muscle actin and TGF-β1 to serve an anti-fibrotic role (98). Moreover, in a previous study, to further confirm the role of KCa3.1 in NAFLD-related liver fibrosis, rat liver fibrosis models were induced using thioacetamide and HFD. Furthermore, the pharmacodynamic effects of senicapoc were determined via detecting biomarkers of apoptosis, inflammation, steatosis and fibrosis (62). The upregulation of the KCa3.1 signaling pathway has previously been observed in three different pathological models. However, in the aforementioned previous study senicapoc-induced inhibition of the KCa3.1 signaling pathway reduced hepatic triglyceride content and served an anti-inflammatory and anti-steatotic role (62). Therefore, drug inhibition of the KCa3.1 channel may serve as a potential therapeutic target for liver fibrosis induced via NAFLD.
5. Conclusion and future perspectives
NAFLD is one of the most common causes of cirrhosis and HCC, however to date, there are no effective treatments (1). Therefore, improving the quality of human life with respect to NAFLD is a challenge. In conclusion, NAFLD involves increased hepatic fatty acid synthesis, decreased fatty acid catabolism and cellular stress, including oxidative stress, the inflammatory response and ER stress (57). Dysregulation of Ca2+ channel expression leads to the disruption of Ca2+ signal homeostasis, which results in the aforementioned changes discussed in the present review. This suggests that ion channels serve an important role in the occurrence and development of NAFLD. The inhibition or activation of these channel proteins may be a potential therapeutic approach to prevent NAFLD progression to NASH, fibrosis, cirrhosis and HCC. Furthermore, it is unclear if Ca2+ channels serve a role in all stages of NAFLD, including simple steatosis, chronic inflammation, fibrosis or cirrhosis. Therefore, further research is required in this area of research. At present, there are relatively few studies concerning NAFLD and most of these are still at the laboratory stage. The clinical feasibility and effectiveness of NAFLD Ca2+ channel-targeted therapeutics need to be studied. Therefore, more systematic studies are needed in the future.
Acknowledgments
We thank Dr Hai Jin (Department of Gastroenterology, Digestive Disease Hospital, Affiliated Hospital of Zunyi Medical University, Zunyi, Guizhou) for providing suggestions for the review.
Funding Statement
The present study was supported by a grant from the National Natural Science Foundation of China (grant no. 82073087), the National Natural Science Foundation of China (grant no. 81960507) and the Science and Technology Bureau fund of Zunyi city [grant no. ZUN SHI KE HE HZ ZI(2019)93-HAO] Collaborative Innovation Center of Chinese Ministry of Education (2020-39).
Availability of data and materials
Not applicable.
Authors' contributions
XC made substantial contributions to the conception design of the study as well as wrote the manuscript and performed the literature search. LZha, LZhe and BT were involved in revising the manuscript critically for important intellectual content. Data authentication is not applicable. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Brunt EM, Wong VW, Nobili V, Day CP, Sookoian S, Maher JJ, Bugianesi E, Sirlin CB, Neuschwander-Tetri BA, Rinella ME. Nonalcoholic fatty liver disease. Nat Rev Dis Primers. 2015;1:15080. doi: 10.1038/nrdp.2015.80. [DOI] [PubMed] [Google Scholar]
- 2.Gusdon AM, Song KX, Qu S. Nonalcoholic fatty liver disease: Pathogenesis and therapeutics from a mitochondria-centric perspective. Oxid Med Cell Longev. 2014;2014:637027. doi: 10.1155/2014/637027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zhu JZ, Dai YN, Wang YM, Zhou QY, Yu CH, Li YM. Prevalence of nonalcoholic fatty liver disease and economy. Dig Dis Sci 2015. 2015 Nov;60:3194–3202. doi: 10.1007/s10620-015-3728-3. [DOI] [PubMed] [Google Scholar]
- 4.Neuschwander-Tetri BA. Non-alcoholic fatty liver disease. BMC Med. 2017;15:45. doi: 10.1186/s12916-017-0806-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Stefan N, Häring H, Cusi K. Non-alcoholic fatty liver disease: Causes, diagnosis, cardiometabolic consequences, and treatment strategies. Lancet Diabetes Endocrinol. 2019;7:313–324. doi: 10.1016/S2213-8587(18)30154-2. [DOI] [PubMed] [Google Scholar]
- 6.Manne V, Handa P, Kowdley KV. Pathophysiology of nonalcoholic fatty liver disease/nonalcoholic steatohepatitis. Clin Liver Dis. 2018;22:23–37. doi: 10.1016/j.cld.2017.08.007. [DOI] [PubMed] [Google Scholar]
- 7.Varela-Rey M, Embade N, Ariz U, Lu SC, Mato JM, Martínez-Chantar ML. Non-alcoholic steatohepatitis and animal models: Understanding the human disease. Int J Biochem Cell Biol. 2009;41:969–976. doi: 10.1016/j.biocel.2008.10.027. [DOI] [PubMed] [Google Scholar]
- 8.Day CP, James OF. Steatohepatitis: A tale of two 'hits'? Gastroenterology. 1998;114:842–845. doi: 10.1016/S0016-5085(98)70599-2. [DOI] [PubMed] [Google Scholar]
- 9.Friedman SL, Neuschwander-Tetri BA, Rinella M, Sanyal AJ. Mechanisms of NAFLD development and therapeutic strategies. Nat Med. 2018;24:908–922. doi: 10.1038/s41591-018-0104-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Buzzetti E, Pinzani M, Tsochatzis E. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD) Metabolism. 2016;65:1038–1048. doi: 10.1016/j.metabol.2015.12.012. [DOI] [PubMed] [Google Scholar]
- 11.Schuppan D, Schattenberg JM. Non-alcoholic steatohepatitis: Pathogenesis and novel therapeutic approaches. J Gastroenterol Hepatol. 2013;28(Suppl 1):S68–S76. doi: 10.1111/jgh.12212. [DOI] [PubMed] [Google Scholar]
- 12.Cortez-Pinto H, de Moura MC, Day CP. Non-alcoholic steatohepatitis: From cell biology to clinical practice. J Hepatol. 2006;44:197–208. doi: 10.1016/j.jhep.2005.09.002. [DOI] [PubMed] [Google Scholar]
- 13.Younossi ZM, Blissett D, Blissett R, Henry L, Stepanova M, Younossi Y, Racila A, Hunt S, Beckerman R. The economic and clinical burden of nonalcoholic fatty liver disease in the United States and Europe. Hepatology. 2016;64:1577–1586. doi: 10.1002/hep.28785. [DOI] [PubMed] [Google Scholar]
- 14.Wong RJ, Aguilar M, Cheung R, Perumpail RB, Harrison SA, Younossi ZM, Ahmed A. Nonalcoholic steatohepatitis is the second leading etiology of liver disease among adults awaiting liver transplantation in the United States. Gastroenterology. 2015;148:547–555. doi: 10.1053/j.gastro.2014.11.039. [DOI] [PubMed] [Google Scholar]
- 15.Kiselyov K, Muallem S. ROS and intracellular ion channels. Cell Calcium. 2016;60:108–114. doi: 10.1016/j.ceca.2016.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Guéguinou M, Chantôme A, Fromont G, Bougnoux P, Vandier C, Potier-Cartereau M. KCa and Ca(2+) channels: The complex thought. Biochim Biophys Acta. 2014;1843:2322–2333. doi: 10.1016/j.bbamcr.2014.02.019. [DOI] [PubMed] [Google Scholar]
- 17.Ramírez A, Vázquez-Sánchez AY, Carrión-Robalino N, Camacho J. Ion channels and oxidative stress as a potential link for the diagnosis or treatment of liver diseases. Oxid Med Cell Longev. 2016;2016:3928714. doi: 10.1155/2016/3928714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ali ES, Rychkov GY, Barritt GJ. Targeting Ca2+ signaling in the initiation, promotion and progression of hepatocellular carcinoma. Cancers (Basel) 2020;12:2755. doi: 10.3390/cancers12102755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ben-Moshe S, Itzkovitz S. Spatial heterogeneity in the mammalian liver. Nat Rev Gastroenterol Hepatol. 2019;16:395–410. doi: 10.1038/s41575-019-0134-x. [DOI] [PubMed] [Google Scholar]
- 20.Garcin I, Tordjmann T. Calcium signalling and liver regeneration. Int J Hepatol. 2012;2012:630670. doi: 10.1155/2012/630670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Taira Z, Ueda Y, Monmasu H, Yamase D, Miyake S, Shiraishi M. Characteristics of intracellular Ca2+ signals consisting of two successive peaks in hepatocytes during liver regeneration after 70% partial hepatectomy in rats. J Exp Pharmacol. 2016;8:21–33. doi: 10.2147/JEP.S106084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Berridge MJ, Bootman MD, Roderick HL. Calcium signalling: Dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol. 2003;4:517–529. doi: 10.1038/nrm1155. [DOI] [PubMed] [Google Scholar]
- 23.Wu L, Lian W, Zhao L. Calcium signaling in cancer progression and therapy. FEBS J. 2021;288:6187–6205. doi: 10.1111/febs.16133. [DOI] [PubMed] [Google Scholar]
- 24.Oliva-Vilarnau N, Hankeova S, Vorrink SU, Mkrtchian S, Andersson ER, Lauschke VM. Calcium signaling in liver injury and regeneration. Front Med (Lausanne) 2018;5:192. doi: 10.3389/fmed.2018.00192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bartlett P, Gaspers L, Pierobon N, Thomas A. Calcium-dependent regulation of glucose homeostasis in the liver. Cell Calcium. 2014;55:306–316. doi: 10.1016/j.ceca.2014.02.007. [DOI] [PubMed] [Google Scholar]
- 26.Chen CC, Hsu LW, Chen KD, Chiu KW, Chen CL, Huang KT. Emerging roles of calcium signaling in the development of non-alcoholic fatty liver disease. Int J Mol Sci. 2021;23:256. doi: 10.3390/ijms23010256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wang J, He W, Tsai PJ, Chen PH, Ye M, Guo J, Su Z. Mutual interaction between endoplasmic reticulum and mitochondria in nonalcoholic fatty liver disease. Lipids Health Dis. 2020;19:72. doi: 10.1186/s12944-020-01210-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Arruda A, Pers B, Parlakgul G, Güney E, Goh T, Cagampan E, Lee GY, Goncalves RL, Hotamisligil GS. Defective STIM-mediated store operated Ca2+ entry in hepatocytes leads to metabolic dysfunction in obesity. Elife. 2017;6:e29968. doi: 10.7554/eLife.29968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhang SL, Yu Y, Roos J, Kozak JA, Deerinck TJ, Ellisman MH, Stauderman KA, Cahalan MD. STIM1 is a Ca2+ sensor that activates CRAC channels and migrates from the Ca2+ store to the plasma membrane. Nature. 2005;437:902–905. doi: 10.1038/nature04147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Park SW, Zhou Y, Lee J, Lee J, Ozcan U. Sarco(endo)plasmic reticulum Ca2+-ATPase 2b is a major regulator of endoplasmic reticulum stress and glucose homeostasis in obesity. Proc Natl Acad Sci USA. 2010;107:19320–19325. doi: 10.1073/pnas.1012044107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Egnatchik RA, Leamy AK, Jacobson DA, Shiota M, Young JD. ER calcium release promotes mitochondrial dysfunction and hepatic cell lipotoxicity in response to palmitate overload. Mol Metab. 2014;3:544–553. doi: 10.1016/j.molmet.2014.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Fu S, Yang L, Li P, Hofmann O, Dicker L, Hide W, Lin X, Watkins SM, Ivanov AR, Hotamisligil GS. Aberrant lipid metabolism disrupts calcium homeostasis causing liver endoplasmic reticulum stress in obesity. Nature. 2011;473:528–531. doi: 10.1038/nature09968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Amaya M, Nathanson M. Calcium signaling in the liver. Compr Physiol. 2013;3:515–539. doi: 10.1002/cphy.c120013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Szabadkai G, Bianchi K, Várnai P, De Stefani D, Wieckowski MR, Cavagna D, Nagy AI, Balla T, Rizzuto R. Chaperone-mediated coupling of endoplasmic reticulum and mitochondrial Ca2+ channels. J Cell Biol. 2006;175:901–911. doi: 10.1083/jcb.200608073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Masarone M, Rosato V, Dallio M, Gravina AG, Aglitti A, Loguercio C, Federico A, Persico M. Role of oxidative stress in pathophysiology of nonalcoholic fatty liver disease. Oxid Med Cell Longev. 2018;2018:9547613. doi: 10.1155/2018/9547613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Luzio JP, Hackmann Y, Dieckmann NM, Griffiths GM. The biogenesis of lysosomes and lysosome-related organelles. Cold Spring Harb Perspect Biol. 2014;6:a016840. doi: 10.1101/cshperspect.a016840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Singh R, Kaushik S, Wang Y, Xiang Y, Novak I, Komatsu M, Tanaka K, Cuervo AM, Czaja MJ. Autophagy regulates lipid metabolism. Nature. 2009;458:1131–1135. doi: 10.1038/nature07976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lin CW, Zhang H, Li M, Xiong X, Chen X, Chen X, Dong XC, Yin XM. Pharmacological promotion of autophagy alleviates steatosis and injury in alcoholic and non-alcoholic fatty liver conditions in mice. J Hepatol. 2013;58:993–999. doi: 10.1016/j.jhep.2013.01.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ali ES, Rychkov GY, Barritt GJ. Deranged hepatocyte intracellular Ca2+ homeostasis and the progression of non-alcoholic fatty liver disease to hepatocellular carcinoma. Cell Calcium. 2019;82:102057. doi: 10.1016/j.ceca.2019.102057. [DOI] [PubMed] [Google Scholar]
- 40.Miyagawa K, Oe S, Honma Y, Izumi H, Baba R, Harada M. Lipid-induced endoplasmic reticulum stress impairs selective autophagy at the step of autophagosome-lysosome fusion in hepatocytes. Am J Pathol. 2016;186:1861–1873. doi: 10.1016/j.ajpath.2016.03.003. [DOI] [PubMed] [Google Scholar]
- 41.Koo JH, Han CY. Signaling nodes associated with endoplasmic reticulum stress during NAFLD progression. Biomolecules. 2021;11:242. doi: 10.3390/biom11020242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ali ES, Petrovsky N. Calcium signaling as a therapeutic target for liver steatosis. Trends Endocrinol Metab. 2019;30:270–281. doi: 10.1016/j.tem.2019.02.005. [DOI] [PubMed] [Google Scholar]
- 43.Grimm C, Holdt LM, Chen CC, Hassan S, Müller C, Jörs S, Cuny H, Kissing S, Schröder B, Butz E, et al. High susceptibility to fatty liver disease in two-pore channel 2-deficient mice. Nat Commun. 2014;5:4699. doi: 10.1038/ncomms5699. [DOI] [PubMed] [Google Scholar]
- 44.Li Q, Li L, Wang F, Chen J, Zhao Y, Wang P, Nilius B, Liu D, Zhu Z. Dietary capsaicin prevents nonalcoholic fatty liver disease through transient receptor potential vanilloid 1-mediated peroxisome proliferator-activate receptor delta activation. Pflugers Arch. 2013;465:1303–1316. doi: 10.1007/s00424-013-1274-4. [DOI] [PubMed] [Google Scholar]
- 45.Li L, Chen J, Ni Y, Feng X, Zhao Z, Wang P, Sun J, Yu H, Yan Z, Liu D, et al. TRPV1 activation prevents nonalcoholic fatty liver through UCP2 upregulation in mice. Pflugers Arch. 2012;463:727–732. doi: 10.1007/s00424-012-1078-y. [DOI] [PubMed] [Google Scholar]
- 46.Wang K, Tan W, Liu X, Deng L, Huang L, Wang X, Gao X. New insight and potential therapy for NAFLD: CYP2E1 and flavonoids. Biomed Pharmacother. 2021;137:111326. doi: 10.1016/j.biopha.2021.111326. [DOI] [PubMed] [Google Scholar]
- 47.Seth RK, Das S, Dattaroy D, Chandrashekaran V, Alhasson F, Michelotti G, Nagarkatti M, Nagarkatti P, Diehl AM, Bell PD, et al. TRPV4 activation of endothelial nitric oxide synthase resists nonalcoholic fatty liver disease by blocking CYP2E1-mediated redox toxicity. Free Radic Biol Med. 2017;102:260–273. doi: 10.1016/j.freeradbiomed.2016.11.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Feng Q, Liu C, Gao W, Geng XL, Dai N. Salidroside-mitigated inflammatory injury of hepatocytes with non-alcoholic fatty liver disease via inhibition TRPM2 ion channel activation. Diabetes Metab Syndr Obes. 2019;12:2755–2763. doi: 10.2147/DMSO.S210764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Ali ES, Rychkov GY, Barritt GJ. TRPM2 non-selective cation channels in liver injury mediated by reactive oxygen species. Antioxidants (Basel) 2021;10:1243. doi: 10.3390/antiox10081243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Yu T, Zheng E, Li Y, Li Y, Xia J, Ding Q, Hou Z, Ruan XZ, Zhao L, Chen Y. Src-mediated Tyr353 phosphorylation of IP3R1 promotes its stability and causes apoptosis in palmitic acid-treated hepatocytes. Exp Cell Res. 2021;399:112438. doi: 10.1016/j.yexcr.2020.112438. [DOI] [PubMed] [Google Scholar]
- 51.Feriod CN, Oliveira AG, Guerra MT, Nguyen L, Richards KM, Jurczak MJ, Ruan HB, Camporez JP, Yang X, Shulman GI, et al. Hepatic inositol 1,4,5 trisphosphate receptor type 1 mediates fatty liver. Hepatol Commun. 2017;1:23–35. doi: 10.1002/hep4.1012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Khamphaya T, Chukijrungroat N, Saengsirisuwan V, Mitchell-Richards KA, Robert ME, Mennone A, Ananthanarayanan M, Nathanson MH, Weerachayaphorn J. Nonalcoholic fatty liver disease impairs expression of the type II inositol 1,4,5-trisphosphate receptor. Hepatology. 2018;67:560–574. doi: 10.1002/hep.29588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Smedlund K, Dube P, Vazquez G. Early steatohepatitis in hyperlipidemic mice with endothelial-specific gain of TRPC3 function precedes changes in aortic atherosclerosis. Physiol Genomics. 2016;48:644–649. doi: 10.1152/physiolgenomics.00067.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Zhu Y, Zhang C, Xu F, Zhao M, Bergquist J, Yang C, Liu X, Tan Y, Wang X, Li S, et al. System biology analysis reveals the role of voltage-dependent anion channel in mitochondrial dysfunction during non-alcoholic fatty liver disease progression into hepatocellular carcinoma. Cancer Sci. 2020;111:4288–4302. doi: 10.1111/cas.14651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ali ES, Rychkov GY, Barritt GJ. Metabolic disorders and cancer: Hepatocyte store-operated Ca2+ channels in nonalcoholic fatty liver disease. Adv Exp Med Biol. 2017;993:595–621. doi: 10.1007/978-3-319-57732-6_30. [DOI] [PubMed] [Google Scholar]
- 56.Wilson CH, Ali ES, Scrimgeour N, Martin AM, Hua J, Tallis GA, Rychkov GY, Barritt GJ. Steatosis inhibits liver cell store-operated Ca2+ entry and reduces ER Ca2+ through a protein kinase C-dependent mechanism. Biochem J. 2015;466:379–390. doi: 10.1042/BJ20140881. [DOI] [PubMed] [Google Scholar]
- 57.Zhang B, Yang W, Zou Y, Li M, Guo H, Zhang H, Xia C, Xu C. NEFA-sensitive Orai1 expression in regulation of de novo lipogenesis. Cell Physiol Biochem. 2018;47:1310–1317. doi: 10.1159/000490226. [DOI] [PubMed] [Google Scholar]
- 58.Zhang B, Li M, Zou Y, Guo H, Zhang B, Xia C, Zhang H, Yang W, Xu C. NFκB/Orai1 facilitates endoplasmic reticulum stress by oxidative stress in the pathogenesis of non-alcoholic fatty liver disease. Front Cell Dev Biol. 2019;7:202. doi: 10.3389/fcell.2019.00202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Chatterjee S, Rana R, Corbett J, Kadiiska MB, Goldstein J, Mason RP. P2X7 receptor-NADPH oxidase axis mediates protein radical formation and Kupffer cell activation in carbon tetrachloride-mediated steatohepatitis in obese mice. Free Radic Biol Med. 2012;52:1666–1679. doi: 10.1016/j.freeradbiomed.2012.02.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Das S, Seth RK, Kumar A, Kadiiska MB, Michelotti G, Diehl AM, Chatterjee S. Purinergic receptor X7 is a key modulator of metabolic oxidative stress-mediated autophagy and inflammation in experimental nonalcoholic steatohepatitis. Am J Physiol Gastrointest Liver Physiol. 2013;305:G950–G963. doi: 10.1152/ajpgi.00235.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Freise C, Heldwein S, Erben U, Hoyer J, Köhler R, Jöhrens K, Patsenker E, Ruehl M, Seehofer D, Stickel F, Somasundaram R. K+-channel inhibition reduces portal perfusion pressure in fibrotic rats and fibrosis associated characteristics of hepatic stellate cells. Liver Int. 2015;35:1244–1252. doi: 10.1111/liv.12681. [DOI] [PubMed] [Google Scholar]
- 62.Paka L, Smith DE, Jung D, McCormack S, Zhou P, Duan B, Li JS, Shi J, Hao YJ, Jiang K, et al. Anti-steatotic and anti-fibrotic effects of the KCa3.1 channel inhibitor, senicapoc, in non-alcoholic liver disease. World J Gastroenterol. 2017;23:4181–4190. doi: 10.3748/wjg.v23.i23.4181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Morgan AJ, Galione A. Two-pore channels (TPCs): Current controversies. Bioessays. 2014;36:173–183. doi: 10.1002/bies.201300118. [DOI] [PubMed] [Google Scholar]
- 64.Patel S, Kilpatrick BS. Two-pore channels and disease. Biochim Biophys Acta Mol Cell Res. 2018;1865:1678–1686. doi: 10.1016/j.bbamcr.2018.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Bishnoi M, Khare P, Brown L, Panchal SK. Transient receptor potential (TRP) channels: A metabolic TR(i)P to obesity prevention and therapy. Obes Rev. 2018;19:1269–1292. doi: 10.1111/obr.12703. [DOI] [PubMed] [Google Scholar]
- 66.Zhu Z, Luo Z, Ma S, Liu D. TRP channels and their implications in metabolic diseases. Pflugers Arch. 2011;461:211–223. doi: 10.1007/s00424-010-0902-5. [DOI] [PubMed] [Google Scholar]
- 67.Nilius B, Owsianik G. The transient receptor potential family of ion channels. Genome Biol. 2011;12:218. doi: 10.1186/gb-2011-12-3-218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Venkatachalam K, Montell C. TRP channels. Annu Rev Biochem. 2007;76:387–417. doi: 10.1146/annurev.biochem.75.103004.142819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Zhang LL, Yan Liu D, Ma LQ, Luo ZD, Cao TB, Zhong J, Yan ZC, Wang LJ, Zhao ZG, Zhu SJ, et al. Activation of transient receptor potential vanilloid type-1 channel prevents adipogenesis and obesity. Circ Res. 2007;100:1063–1070. doi: 10.1161/01.RES.0000262653.84850.8b. [DOI] [PubMed] [Google Scholar]
- 70.Uchida K, Dezaki K, Yoneshiro T, Watanabe T, Yamazaki J, Saito M, Yada T, Tominaga M, Iwasaki Y. Involvement of thermosensitive TRP channels in energy metabolism. J Physiol Sci. 2017;67:549–560. doi: 10.1007/s12576-017-0552-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Park HW, Lee JH. Calcium channel blockers as potential therapeutics for obesity-associated autophagy defects and fatty liver pathologies. Autophagy. 2014;10:2385–2386. doi: 10.4161/15548627.2014.984268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Polyzos SA, Kountouras J, Mantzoros CS. Obesity and nonalcoholic fatty liver disease: From pathophysiology to therapeutics. Metabolism. 2019;92:82–97. doi: 10.1016/j.metabol.2018.11.014. [DOI] [PubMed] [Google Scholar]
- 73.Li J, Li X, Liu D, Zhang S, Tan N, Yokota H, Zhang P. Phosphorylation of eIF2α signaling pathway attenuates obesity-induced non-alcoholic fatty liver disease in an ER stress and autophagy-dependent manner. Cell Death Dis. 2020;11:1069. doi: 10.1038/s41419-020-03264-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Baffy G. Uncoupling protein-2 and non-alcoholic fatty liver disease. Front Biosci. 2005;10:2082–2096. doi: 10.2741/1683. [DOI] [PubMed] [Google Scholar]
- 75.Panchal SK, Bliss E, Brown L. Capsaicin in metabolic syndrome. Nutrients. 2018;10:630. doi: 10.3390/nu10050630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Yang L, Li P, Fu S, Calay E, Hotamisligil GS. Defective hepatic autophagy in obesity promotes ER stress and causes insulin resistance. Cell Metab. 2010;11:467–478. doi: 10.1016/j.cmet.2010.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Everaerts W, Nilius B, Owsianik G. The vanilloid transient receptor potential channel TRPV4: From structure to disease. Prog Biophys Mol Biol. 2010;103:2–17. doi: 10.1016/j.pbiomolbio.2009.10.002. [DOI] [PubMed] [Google Scholar]
- 78.Ye L, Kleiner S, Wu J, Sah R, Gupta RK, Banks AS, Cohen P, Khandekar MJ, Boström P, Mepani RJ, et al. TRPV4 is a regulator of adipose oxidative metabolism, inflammation, and energy homeostasis. Cell. 2012;151:96–110. doi: 10.1016/j.cell.2012.08.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Zhan L, Yang Y, Ma TT, Huang C, Meng XM, Zhang L, Li J. Transient receptor potential vanilloid 4 inhibits rat HSC-T6 apoptosis through induction of autophagy. Mol Cell Biochem. 2015;402:9–22. doi: 10.1007/s11010-014-2298-6. [DOI] [PubMed] [Google Scholar]
- 80.Song Y, Zhan L, Yu M, Huang C, Meng X, Ma T, Zhang L, Li J. TRPV4 channel inhibits TGF-β1-induced proliferation of hepatic stellate cells. PLoS One. 2014;9:e101179. doi: 10.1371/journal.pone.0101179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Fonfria E, Murdock PR, Cusdin FS, Benham CD, Kelsell RE, McNulty S. Tissue distribution profiles of the human TRPM cation channel family. J Recept Signal Transduct Res. 2006;26:159–178. doi: 10.1080/10799890600637506. [DOI] [PubMed] [Google Scholar]
- 82.Kheradpezhouh E, Ma L, Morphett A, Barritt GJ, Rychkov GY. TRPM2 channels mediate acetaminophen-induced liver damage. Proc Natl Acad Sci USA. 2014;111:3176–3181. doi: 10.1073/pnas.1322657111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Vanderheyden V, Devogelaere B, Missiaen L, De Smedt H, Bultynck G, Parys JB. Regulation of inositol 1,4,5-trisphosphate-induced Ca2+ release by reversible phosphorylation and dephosphorylation. Biochim Biophys Acta. 2009;1793:959–970. doi: 10.1016/j.bbamcr.2008.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Arruda AP, Pers BM, Parlakgül G, Güney E, Inouye K, Hotamisligil GS. Chronic enrichment of hepatic endoplasmic reticulum-mitochondria contact leads to mitochondrial dysfunction in obesity. Nat Med. 2014;20:1427–1435. doi: 10.1038/nm.3735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Rodrigues MA, Gomes DA, Leite MF, Grant W, Zhang L, Lam W, Cheng YC, Bennett AM, Nathanson MH. Nucleoplasmic calcium is required for cell proliferation. J Biol Chem. 2007;282:17061–17068. doi: 10.1074/jbc.M700490200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Lemasters JJ, Holmuhamedov E. Voltage-dependent anion channel (VDAC) as mitochondrial governator-thinking outside the box. Biochim Biophys Acta. 2006;1762:181–190. doi: 10.1016/j.bbadis.2005.10.006. [DOI] [PubMed] [Google Scholar]
- 87.Shoshan-Barmatz V, De Pinto V, Zweckstetter M, Raviv Z, Keinan N, Arbel N. VDAC, a multi-functional mitochondrial protein regulating cell life and death. Mol Aspects Med. 2010;31:227–285. doi: 10.1016/j.mam.2010.03.002. [DOI] [PubMed] [Google Scholar]
- 88.Pittala S, Krelin Y, Kuperman Y, Shoshan-Barmatz V. A mitochondrial VDAC1-based peptide greatly suppresses steatosis and NASH-associated pathologies in a mouse model. Mol Ther. 2019;27:1848–1862. doi: 10.1016/j.ymthe.2019.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Prakriya M, Lewis RS. Store-operated calcium channels. Physiol Rev. 2015;95:1383–1436. doi: 10.1152/physrev.00020.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kappel S, Borgström A, Stoklosa P, Dörr K, Peinelt C. Store-operated calcium entry in disease: Beyond STIM/Orai expression levels. Semin Cell Dev Biol. 2019;94:66–73. doi: 10.1016/j.semcdb.2019.01.003. [DOI] [PubMed] [Google Scholar]
- 91.Li Y, Ge M, Ciani L, Kuriakose G, Westover EJ, Dura M, Covey DF, Freed JH, Maxfield FR, Lytton J, Tabas I. Enrichment of endoplasmic reticulum with cholesterol inhibits sarcoplasmic-endoplasmic reticulum calcium ATPase-2b activity in parallel with increased order of membrane lipids: Implications for depletion of endoplasmic reticulum calcium stores and apoptosis in cholesterol-loaded macrophages. J Biol Chem. 2004;279:37030–37039. doi: 10.1074/jbc.M405195200. [DOI] [PubMed] [Google Scholar]
- 92.Maus M, Cuk M, Patel B, Lian J, Ouimet M, Kaufmann U, Yang J, Horvath R, Hornig-Do HT, Chrzanowska-Lightowlers ZM, et al. Store-operated Ca2+ entry controls induction of lipolysis and the transcriptional reprogramming to lipid metabolism. Cell Metab. 2017;25:698–712. doi: 10.1016/j.cmet.2016.12.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Jain S, Jacobson KA. Purinergic signaling in liver pathophysiology. Front Endocrinol (Lausanne) 2021;12:718429. doi: 10.3389/fendo.2021.718429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Jiang M, Cui BW, Wu YL, Zhang Y, Shang Y, Liu J, Yang HX, Qiao CY, Zhan ZY, Ye H, et al. P2X7R orchestrates the progression of murine hepatic fibrosis by making a feedback loop from macrophage to hepatic stellate cells. Toxicol Lett. 2020;333:22–32. doi: 10.1016/j.toxlet.2020.07.023. [DOI] [PubMed] [Google Scholar]
- 95.Takenouchi T, Nakai M, Iwamaru Y, Sugama S, Tsukimoto M, Fujita M, Wei J, Sekigawa A, Sato M, Kojima S, et al. The activation of P2X7 receptor impairs lysosomal functions and stimulates the release of autophagolysosomes in microglial cells. J Immunol. 2009;182:2051–2062. doi: 10.4049/jimmunol.0802577. [DOI] [PubMed] [Google Scholar]
- 96.Chatterjee S, Das S. P2X7 receptor as a key player in oxidative stress-driven cell fate in nonalcoholic steatohepatitis. Oxid Med Cell Longev. 2015;2015:172493. doi: 10.1155/2015/172493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Wulff H, Castle NA. Therapeutic potential of KCa3.1 blockers: Recent advances and promising trends. Expert Rev Clin Pharmacol. 2010;3:385–396. doi: 10.1586/ecp.10.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Alkhouri N, Dixon LJ, Feldstein AE. Lipotoxicity in nonalcoholic fatty liver disease: Not all lipids are created equal. Expert Rev Gastroenterol Hepatol. 2009;3:445–451. doi: 10.1586/egh.09.32. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.