

ABSTRACT
The Middle to Upper Palaeolithic transition, between 50 000 and 40 000 years ago, is a period of important ecological and cultural changes. In this framework, the Rock Shelter of Uluzzo C (Apulia, southern Italy) represents an important site due to Late Mousterian and Uluzzian evidence preserved in its stratigraphic sequence. Here, we present the results of a multidisciplinary analysis performed on the materials collected between 2016 and 2018 from the Uluzzian stratigraphic units (SUs) 3, 15 and 17. The analysis involved lithic technology, use‐wear, zooarchaeology, ancient DNA of sediments and palaeoproteomics, completed by quartz single‐grain optically stimulated luminescence dating of the cave sediments. The lithic assemblage is characterized by a volumetric production and a debitage with no or little management of the convexities (by using the bipolar technique), with the objective to produce bladelets and flakelets. The zooarchaeological study found evidence of butchery activity and of the possible exploitation of marine resources, while drawing a picture of a patchy landscape, composed of open forests and dry open environments surrounding the shelter. Ancient mitochondrial DNA from two mammalian taxa were recovered from the sediments. Preliminary zooarchaeology by mass spectrometry results are consistent with ancient DNA and zooarchaeological taxonomic information, while further palaeoproteomics investigations are ongoing. Our new data from the re‐discovery of the Uluzzo C Rock Shelter represent an important contribution to better understand the meaning of the Uluzzian in the context of the Middle/Upper Palaeolithic transition in south‐eastern Italy.
Keywords: ancient DNA of sediment, lithic technology, Uluzzian, zooarchaeology, ZooMS
Abbreviations
- DC
dimensional class
- FA
formic acid
- HPA
high‐power approach
- LP
Lower Palaeolithic
- LPA
low‐power approach
- MH
Modern Human
- MNI
minimum‐number of individuals
- MPN
Museo Della Preistoria di Nardò (Lecce, Apulia, Italy)
- NISP
number of identified specimens
- SU
stratigraphic unit
- UP
Upper Palaeolithic
- WoRMS
World Register of Marine Species
Introduction
The Middle to Upper Palaeolithic (MP/UP) transition corresponds to the period between around 50 000 and 40 000 years ago and is a key period of change in the prehistory of the Old World. This timespan covers the last millennia of Neanderthal presence in the fossil record, together with the appearance of Modern Human (MH) populations in Europe (Benazzi et al., 2011a; Douka et al., 2014; Higham et al., 2014).
Mediterranean Europe, and in particular Italy, plays an essential role in the study of this period due to: (i) its geographical position and ecological variability; (ii) the presence of key archaeological sites dating back to the MP/UP transition; and (iii) the evidence of different techno‐complexes – such as Late Mousterian, Uluzzian and Protoaurignacian – associated with human fossil remains. In the Palaeolithic context of Mediterranean Europe, Neanderthals are associated with Mousterian assemblages, while, according to Benazzi et al. (2011b, 2015) MHs are associated with Uluzzian and Protoaurignacian assemblages.
The Uluzzian is one of the first lithic assemblages related to the arrival of MHs in Europe. It is characterized by its own originality and coherence, observable in lithic technology (Riel‐Salvatore, 2009, 2010; De Stefani et al., 2012; Moroni et al., 2013, 2018; Ronchitelli et al., 2018; Villa et al., 2018; Peresani et al., 2019; Arrighi et al., 2020a; Collina et al., 2020; Marciani et al., 2020a), manufacturing of formal bone tools (d'Errico et al., 2012; Peresani et al., 2016; Villa et al., 2018; Peresani et al., 2019; Arrighi et al., 2020a) and ornaments (Arrighi et al., 2020b, 2020c), which point towards the sharing of common behaviours among its social groups and group members.
From a technological point of view, the Uluzzian is in fact characterized by the following. (i) A specific conceptualization of production that consists in the application of a straightforward debitage method, which implies the collection of raw blocks presenting convexities, angles and guide ribs appropriate for knapping; the striking platform being either opened by a single or few removals, or not opened at all by using a natural or cortical plan; the debitage surfaces are roughly managed, and the production of blanks follows unidirectional, bidirectional or orthogonal directions. (ii) The deliberate selection of the bipolar technique (aiming at the production of small blades/bladelets and small flakes/flakelets). (iii) The idea of a ‘simple’ production for complex tools (the use of composite tools) (Riel‐Salvatore, 2009, 2010; Moroni et al., 2013, 2018; Sano et al., 2019; Collina et al., 2020; Marciani et al., 2020a).
Among retouched tools we note the production of lunates (tools characterized by a curved backed side opposite to a rectilinear cutting edge), which at Grotta del Cavallo were used as armatures (Sano et al., 2019), and the systematic production of end‐scrapers (Palma di Cesnola, 1964, 1993, 2004; Gambassini, 1997).
Zooarchaeological data suggest that human adaptive strategies changed over time to cope with variations in the local topographic and ecological conditions, as well as to deal with the uncertainty of resource availability (Romandini et al., 2020). However, while in southern Italy more substantial data on the distribution of ungulate limb elements might point to a marked change in ungulate exploitation between the Late Mousterian and the Early Upper Palaeolithic (Boscato and Crezzini, 2012; Romandini et al., 2020), in northern sites a higher variety of processing techniques were already present at the beginning of the Late Mousterian (Romandini et al., 2020). Accordingly, our understanding of the Uluzzian is far from exhaustive, and additional evidence is required to empirically test inferences regarding Late Mousterian, Uluzzian and Protoaurignacian contexts.
The Uluzzo C Rock Shelter provides another piece of evidence towards a better understanding of the role of the Uluzzian within the MP/UP transition in south‐eastern Italy. The site is located in the Bay of Uluzzo, where numerous Uluzzian sites have already been reported (Moroni et al., 2013, 2018; Marciani et al., 2020a) – including Grotta del Cavallo – and where the Uluzzian was first discovered and described (Palma di Cesnola, 1964). Moreover, the site of Uluzzo C includes a long stratigraphic sequence composed of several levels, including Romanellian, Uluzzian and a long and rich Mousterian sequence. Here we report the results from our multidisciplinary investigation on the sediments from the cave entrance and the archaeological materials from Uluzzo C Rock Shelter, which were collected during previous and new excavations (i.e. in the 1960s and between 2016 and 2018). Our approach includes the analysis of lithic technology and use‐wear, zooarchaeological remains, sedimentary DNA and palaeoproteomics to provide further insights into the lithic behaviour (as far as reduction sequences, goals and knapping techniques are concerned) and the hunting strategies of the human groups that inhabited Uluzzo C during the MP/UP transition. Moreover, we present palaeoecological evidence on the composition of the landscape surrounding the rock shelter during this period.
Archaeological and chronological context
The Uluzzo C Rock Shelter is located in the Parco Naturale di Porto Selvaggio (Nardò, Lecce, southern Italy), on the western side of the Apulian Coast facing the Ionian Sea (40°9′27.84″N, 17°57′35.34″E) (Fig. 1). The Uluzzo C site was discovered during archaeological investigations carried out in the area in the 1960s by the Italian Institute of Prehistory and Protohistory and led by Borzatti von Löwenstern (Borzatti von Lowerstern, 1965; Borzatti von Löwenstern and Magaldi, 1966). During this first exploration, the stratified deposit of the cave yielded significant lithic assemblages that spanned from the Mousterian to the Bronze Age, including the Uluzzian techno‐complex.
Figure 1.
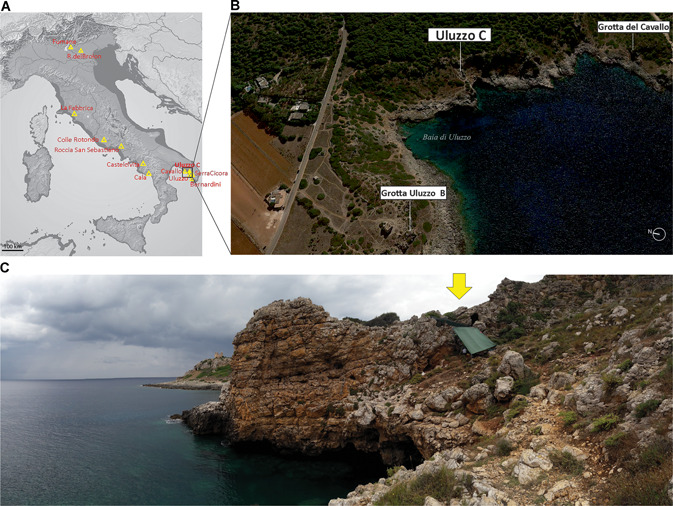
A. Uluzzian sites in Italy. B. Location of Uluzzo C in the bay of Uluzzo (Apulia, southern Italy) from Google Earth. C. General view of the site. [Color figure can be viewed at wileyonlinelibrary.com].
Today, the site is formed by a central hall with a smaller cavity on its right, whereas the stratigraphic sequence consists of 10 different layers (Borzatti von Lowerstern, 1965). From a cultural point of view, the stratigraphic sequence of Uluzzo C includes: a Romanellian layer (A); a sterile layer (B); two Uluzzian layers (C, D); a layer composed of a mixture of Upper Palaeolithic and Mousterian deposits (E); and a large sequence of Mousterian layers (F–L) (Borzatti von Lowerstern, 1965; Borzatti von Löwenstern and Magaldi, 1966; Spinapolice, 2018) (Fig. 2A). This sequence was confirmed by recent excavations, as well as by sedimentological and micromorphological analyses (for a detailed description see Spinapolice et al. in this special issue), which further detailed the sequence by identifying 22 stratigraphic units (SUs) within the layers originally identified by Borzatti.
Figure 2.
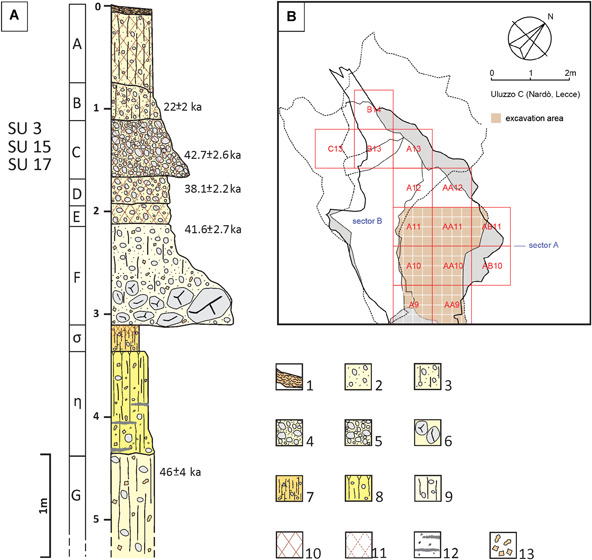
A. Log of the stratigraphic sequence of the deposit of the Uluzzo C Rock Shelter. For a full interpretation of the log the reader is referred to Spinapolice et al. (in this special issue). OSL dating results are indicated. Key: (1) flowstone; (2) sandy unit with rock fragments and bioturbation; (3) sandy unit with rock fragments and bioturbation; (4) clast‐supported breccia displaying weak oblique lamination (sandy matrix); (5) matrix‐supported breccia (sandy matrix); (6) large blocks due to roof collapse; (7) clay‐rich deposit with scarce rock fragments; (8) slightly weathered silty deposit with scarce rock fragments; (9) silty deposit with scarce rock fragments; (10) CaCO3‐cemented deposit; (11) weakly CaCO3‐cemented deposit; (12) charcoal fragments and ash‐rich lenses; (13) CaCO3 nodules and/or concretions. B. New excavation in sector A. [Color figure can be viewed at wileyonlinelibrary.com].
In the new excavation, the site of Uluzzo C was divided into three sectors: A, B and C. Sectors A and B are located inside the rock shelter and correspond to the surface of the actual deposit (sector A) and to the bottom of Borzatti's trench (sector B) (Fig. 2B), whereas sector C is outside the rock shelter and has been explored to identify possible archaeological deposits on the terrace (Fiorini et al., 2018, 2019). The recent investigations (2016–2018) focused on the area inside the rock shelter (sector A) and involved the stratigraphic excavation of the Uluzzian occupation (Borzatti's layer C), which in the new excavations corresponds to SUs 3, 15 and 17. Samples for sedimentological analysis and optically stimulated luminescence (OSL) dating, however, were not taken only from this layer, but from the entire stratigraphic succession accessible in sector A (i.e. Borzatti's layers B, C–L) (Fig. 2).
We performed specific investigations on sediments. The technical details of the sedimentological and micromorphological analyses and of the OSL dating campaign, as well as the individual OSL ages, are reported in Spinapolice et al. (this issue). Based on this single‐grain OSL chronology, the grand weighted mean age for the Uluzzian occupations – layers C, D and E (OSL samples ULOC 3, 4 and 5) is 40.6 ± 1.4 ka (Fig. 2A). These OSL time constraints generated for the Uluzzian layers at Uluzzo C match the chronology of the main Uluzzian sequences of the region (e.g. Grotta del Cavallo). The Uluzzian occupation of Grotta del Cavallo (which probably corresponds to layers D and E at Uluzzo C) is constrained to between 45.5 ± 1.0 ka (Layer Fa/Y‐6) and 39.85 ± 0.14 ka (CI) by Zanchetta et al. (2018). At Uluzzo C the Mousterian layer that underlies the Uluzzian complex (Layer G) has been OSL dated as well (i.e. 46 ± 4.0 ka, Layer G – OSL sample ULOC 1; Fig. 2A) (Spinapolice et al. in this special issue). Hence, despite the stratigraphic diversity and difficulties in matching the Uluzzian layers between Grotta del Cavallo and Uluzzo C, the chronological constraints for the end of the Mousterian appear to be congruent in the two caves: 45.5 ± 1.0 ka at Grotta del Cavallo (Zanchetta et al., 2018) and OSL dated at 46 ± 4.0 ka at Uluzzo C (Layer G – ULOC 1) (Spinapolice et al. in this special issue).
Materials and methods
This paper considers the archaeological finds (i.e. lithics and faunal remains) and sediment samples from the SUs 3, 15 and 17 of the new excavations (2016–2018), which correspond to Borzatti's layer C. In addition, it includes the lithics from layers C, D and E from the excavations carried out by Borzatti during the 1960s (Tables S1–S2). SUs 3, 15 and 17 are three components of the same level of occupation (Layer C) (Fig. 3).
Figure 3.
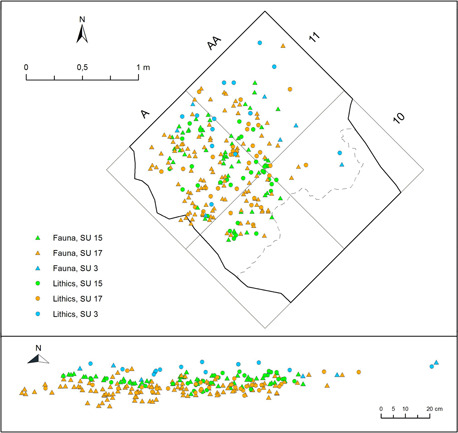
Vertical and horizontal distribution of lithic and faunal remains of SUs 3, 15 and 17 from Uluzzo C Rock Shelter. [Color figure can be viewed at wileyonlinelibrary.com].
The whole lithic assemblage was analysed by using a technological approach. Furthermore, traceological analysis, with both low‐ and high‐power approaches, was performed on the six retouched items.
Taxonomic and taphonomic evaluations were performed on the macromammal, micromammal, avifaunal and malacofaunal remains. Moreover, DNA analysis of sediments and palaeoproteomics were used to corroborate the zooarchaeological studies.
Lithic technological analysis
The technological analysis was carried out on the lithic materials coming from layers C, D and E of Borzatti's excavations and on the materials coming from SUs 3, 15, 17 of the new excavations.
We decided to include all the layers excavated by Borzatti because of the limited number of the items (i.e. layer C, corresponding to the SUs 3, 15 and 17 included only five lithic items).
Borzatti von Lowerstern and Magaldi (1966) report retrieving 145 lithic items from layers C, D and E. Currently, only 61 of these pieces are stored at the Museo della Preistoria di Nardò (MPN) (Table S1). A total of 339 pieces from SUs 3, 15 and 17 of the new excavations were here analysed (Table S2).
This lithic assemblage was analysed by using a technological approach: Geneste (1991) was used as a fundamental conceptual text on how to approach the reduction sequences; Inizan et al. (1999) was used for the definition of technological categories, whereas Boëda (2013) was used to approach lithic technology and, more precisely, regarding the description of the cores. All the archaeological material was sorted according to the lithology and texture of the raw material (chert, jasper, siliceous limestone, limestone and quartz sandstone), the colour of the cortex, the colour of the inner portion of each item, and the presence and type of post‐depositional alteration (chemical, mechanical or thermal). Subsequently, items were divided into five dimensional classes (DC) (DC1: 0–50 mm2, DC2: 50–100 mm2, DC3: 100–150 mm2, DC4: 150–200 mm2, DC5: >200 mm2) on the basis of the area covered by each specimen (Marciani et al., 2020b; Spagnolo et al., 2020). Complete items that were larger than the first DC were additionally measured according to both their technological and morphological axes.
As the material recorded at Uluzzo C is few in number, and the Uluzzian techno‐complex is characterized by an absence of standardization of the categories, in particular regarding production (i.e. Collina et al., 2020), we decided to define the categories of blank based on metric attributes.
Thus, we considered the following technological categories: flakes and blades based on the ratio between length and width, i.e. flakes (ratio length/width ratio < 2) and blades (length/width ratio ≥ 2) (Laplace, 1966). This definition of flakes and blades was used for the Uluzzian layer EIII of Grotta del Cavallo (Moroni et al., 2018). Moreover, flakes whose length is <2.5 cm are named flakelets (<2.5 cm), whereas blades whose length is <2.5 cm are named bladelets (<2.5 cm). The other technological classes are cores, pebbles, debris (items smaller than DC1) and indeterminates (fragmented pieces bigger than DC3, altered pieces, non‐orientatable pieces).
For complete flakes, flakelets, blades and bladelets, we registered the localization and extension of the cortex. We identified the concept and methods of debitage and considered: volumetric aspects (morphology, symmetry, profile and section shape); the number and orientation of dorsal scars; the type of butt and bulb; and the position of the impact point (if present). Lastly, the occurrence, type and localization of the retouch or use‐wear were also noted.
For each core we observed: the nature and morphology of the raw block; the volumetric conception of the exploitation; the hierarchy of surfaces; the type, location and preparation of the striking platform; the number and direction of the negatives on the surface of the debitage; the level of exhaustion and the possible reason for its abandonment.
Use of the bipolar technique on an anvil was evaluated on the basis of specific traits that characterized this kind of technique: rectilinear longitudinal profile of the ventral face, similar ventral and dorsal faces, pronounced ripple marks, shattered point‐form or linear butts, diffused impact points, sheared bulbs of percussion, and the presence of a parasite scar (e.g. Guyodo and Marchand, 2005; Bietti et al., 2010; Soriano et al., 2010; Duke and Pargeter, 2015; de la Peña, 2015; Collina et al., 2020).
Lithic use wear analysis
To evaluate the functional potential of the lithic assemblage from Uluzzo C, a preliminary use‐wear analysis was carried out on the retouched items recovered during the 2016–2018 excavations (six specimens). Traceological analysis was undertaken following both the low‐power approach (LPA) (Tringham et al., 1974; Odell and Odell‐Vereecken, 1980; Odell, 1981) and the high‐power approach (HPA) (Keeley, 1980; Plisson, 1985; Van Gijn, 2010). Traces were observed by means of a Hirox KH‐7700 3D digital microscope using an MX‐G 5040Z body equipped with an AD‐5040 Lows and an AD‐5040HS lens working at low magnification (20–50×) to observe the macro‐traces (fractures, edge damage, diagnostic impact fractures), and an MXG‐10C body and an OL‐140II lens (140–480×) used to analyse micro use‐wear (polishes, abrasions and striations). Before examination with the microscope, the artefacts were washed in fresh water and subsequently cleaned with pure acetone to remove traces of soil and finger grease.
Macromammal remains and zooarchaeological analysis
Identifications of both the skeletal element and the taxa were based on the reference collection stored at the Laboratory of Osteoarchaeology and Paleoanthropology (BONES Lab) of the Department of Cultural Heritage of the University of Bologna (Ravenna, Italy). Microscopic analysis of the bone surfaces was carried out using a Leica stereomicroscope. To identify the nature of the surface alterations on the bones, and to discriminate human from animal traces, trampling abrasion and modern mechanical modifications produced by excavation tools, a well‐established taphonomic literature was used as a reference (Binford, 1981; Potts and Shipman, 1981; Shipman, 1981; Brain, 1983; Shipman and Rose, 1984; Blumenschine and Selvaggio, 1988; Capaldo and Blumenschine, 1994; Lyman, 1994; Blumenschine, 1995; Fisher, 1995). The degree of combustion was evaluated according to Stiner et al. (1995). Sex and age at death were estimated to reconstruct the exploitation strategies of the different species (Aitken, 1974; Mariezcurrena and Altuna, 1983; Vigal and Machordom, 1985; D'Errico and Vanhaeren, 2002; Fiore and Tagliacozzo, 2006). To evaluate species abundance, the number of identified specimens (NISP) was considered (Grayson, 1984).
Micromammal sorting and palaeontological and taphonomic study
The few recovered micromammal fossil remains analysed in this study were disarticulated mandible fragments and isolated teeth (which were collected by water screening the sediments from the excavation of 2018 with a 0.5‐mm mesh screen). The fragments were identified following the general criteria given by Galán‐García (2019) for bats and rodents (Berto, 2013). The specific attribution of this material is based principally on the most diagnostic elements: mandible for the genus Myotis; and mandible and isolated teeth for the subfamily Arvicolinae and genus Apodemus. Moreover, the identified remains were grouped by using the minimum number of individuals (MNI) method, by which we determined the sample by counting the most represented diagnostic elements. Finally, a preliminary taphonomic study was performed to investigate the alterations caused by digestion, which were present in the first lower molars of the arvicoline rodent species (according to Andrews, 1990; Fernández‐Jalvo et al., 2016a), ultimately attesting to the action of predation.
Avifaunal remains
The fossil avifauna from Uluzzo C analysed in this contribution consists of 12 bone remains. The bird remains were recovered after dry/wet sieving of the sediment, numbered with the acronym UC and subjected to taxonomic and taphonomic analyses. Bone identification was possible thanks to the comparison with modern skeletal specimens from two comparative collections: the Marco Pavia Osteological Collection at the Department of Earth Sciences of the University of Torino and the collection at the Department of Humanities of the University of Ferrara. For identification of the remains, we also used osteology handbooks on particular bird families and orders (Janossy, 1983; Tomek and Bochenski, 2000). Taphonomic analysis was carried out with a 30× lens and a Leica S6D Verde Ough 0.75–70× stereomicroscope, available at the Laboratory of Archaeozoology and Taphonomy of the Prehistoric and Anthropological Sciences Section of the University of Ferrara.
Malacofaunal remains
The classification and nomenclature used for taxonomic analysis of the malacofaunal remains were based on the updated datasets available online on the World Register of Marine Species (WoRMS). The NISP was used to define the number of specimens in the assemblage. The taphonomic study focused on three main groups of alterations (Claassen, 1998): pre‐depositional alterations (e.g. marine abrasion, predation by other molluscs and bioerosion); intentional/unintentional anthropogenic alterations (e.g. thermic alterations, anthropic damage caused by consumption); and post‐depositional transformations (e.g. fragmentation, abrasions, root marks, decalcification and excavation damage).
ZooMS
We selected 12 unidentified bone samples (>1 cm in size) for zooarchaeology by mass spectrometry (ZooMS) analysis. We tested the protocol designed by Van Doorn et al. (2011) and used a warm (65 °C) ammonium bicarbonate buffer (50 mm) to leach bone collagen without acid digestion. Then, trypsin digestion was carried out for 18 h at 37 °C using 0.5 μL of sequencing‐grade trypsin (Sigma). Enzymatic digestion was ended using 5 μL of 5% formic acid (FA), then the tryptic digests were purified and concentrated using C18 SpinTips (Thermo Scientific). Peptide elution was performed with 15 μL of 50% acetonitrile (ACN)/0.1% FA (v/v). Samples were dried overnight under a class 100 laminar flow hood. After re‐suspension, each sample (1 μL) was spotted on a target steel plate, and mixed with 1 μL α‐cyano‐4‐hydroxycinnamic acid (CHCA; Sigma) as matrix. The samples were then analysed in duplicate with MALDI‐ToF (Bruker) over a mass‐to‐charge range of 700–3500 m/z. Spectra were manually inspected and averaged using mMass (Strohalm et al., 2010), after setting a signal‐to‐noise ratio equal to 4. Taxonomic identification was performed comparing identified peptides with a database of peptide markers for all European, Pleistocene medium to large size mammals (Welker et al., 2016).
Ancient DNA analysis
A total of 14 sediment samples, which were collected from the site on two different occasions, were tested for the preservation of ancient faunal or hominin DNA. The first set consisted of 11 samples from squares A11 and AA11 in SU 15. The second set was composed of three samples collected from square A11 in SU 17 (Table S3). Subsamples of each sample, ranging between 42 and 100 mg, were used as input for DNA extraction using a silica‐based protocol (Dabney et al., 2013), which was performed either manually or using a liquid handling platform (Bravo NGS workstation, Agilent Technologies) as described by Rohland et al. (2018).
All subsequent laboratory procedures were performed on the liquid handling platform as described by Slon et al. (2017). A single‐stranded DNA library was prepared from each DNA extract using the procedure described by Gansauge et al. (2017, 2020), the efficiency of which was assessed by quantitative PCR. The number of library molecules generated from sediment samples was higher than those prepared from associated negative controls (Table S3), which demonstrates that the library preparation procedures were successful (Gansauge and Meyer, 2013).
The number of molecules generated from a control oligonucleotide was similar between the sediment libraries and the controls, thus indicating that potential inhibitory substances that may have been co‐extracted with the sediment were not interfering with the library preparation procedure to an appreciable extent (Glocke and Meyer, 2017).
Each library was then amplified to PCR plateau and tagged with two indices (Kircher et al., 2012). Libraries originating from sample set 1 were enriched by hybridization capture for mammalian mitochondrial DNA (mtDNA) (Slon et al., 2016), as well as specifically for human mtDNA (Maricic et al., 2010; Fu et al., 2013), while libraries from set 2 were only enriched for human mtDNA. Paired‐end sequencing was performed on an Illumina MiSeq platform using 76 cycles. We note that all sample libraries were sequenced to an adequate depth, as indicated by the duplication rate observed (Table S3). Negative controls for the DNA extraction and library preparation procedures were carried along with the samples.
The processing of sequencing data was performed as described in detail by Slon et al. (2017). After removing PCR duplicates and fragments shorter than 35 bases, taxonomic identification at the family level was carried out for each sequenced DNA fragment by comparing it to a database of reference mammalian mtDNA genomes (Altschul et al., 1990; Huson et al., 2007). At least 10 fragments and at least 1% of identified fragments were required to be assigned to a family for it to be deemed present in a sample. For each identified family, we then tested whether the DNA fragments assigned to it displayed elevated frequencies (i.e. significantly higher than 10%, tested using an exact binomial test) of terminal cytosine (C) to thymine (T) nucleotide substitutions compared to a reference genome. These substitutions are a typical feature of ancient DNA and allow us to determine whether ancient DNA fragments are present in a sample (Briggs et al., 2007). None of the negative controls tested positive for the preservation of ancient DNA (Table S3).
Results
Lithic technology
Technological analysis: Borzatti's excavation
The 61 lithic artefacts from Borzatti's excavation in 1964 (five items from layer C, six items from layer D, 50 items from layer E), which are stored at the Museo della Preistoria di Nardò (Lecce, Apulia), all have fresh margins, except for nine items with blunted edges. Moreover, 11 pieces show traces of a yellow or white patina, and traces of combustion are visible on four items. Generally, their state of preservation is coherent with the lithics coming from the current excavations. In all the layers there is a predominance of fine‐grained chert, followed by siliceous limestone. Occasional limestone and jasper are attested to in layer E (Table 1). Most of the pieces, corresponding to 57.4% of the collection, pertain to DC 5 (corresponding to pieces >200 mm2) (Tables 2 and 3). No lithics pertain to DC 1 (0–50 mm2) and only seven pertain to DC 2 (50–100 mm2) (Table 2). After comparison with the lithics from the more recent excavation (see below), it is evident that a selection bias must have affected the retrieval of the lithic artefacts in the 1960s, probably because researchers were more interested in large diagnostic items, such as retouched pieces. Given the biased composition of the collection and the high degree of the retouch, it was not possible to determine from which reduction sequence these items had come from.
Table 1.
Raw material (Excavation Borzatti)
| Raw material | Layer C | Layer D | Layer E | Total | % |
|---|---|---|---|---|---|
| Chert | 3 | 4 | 38 | 45 | 73.8 |
| Siliceous limestone | 2 | 2 | 7 | 11 | 18.0 |
| Limestone | 0 | 0 | 4 | 4 | 6.6 |
| Jasper | 0 | 0 | 1 | 1 | 1.6 |
| Total | 5 | 6 | 50 | 61 | 100 |
Table 2.
Dimensional classes (Excavation Borzatti)
| Dimensional classes | Layer C | Layer D | Layer E | Total | % | |||
|---|---|---|---|---|---|---|---|---|
| DC 2 | 0 | 0 | 7 | 7 | 11.5 | |||
| DC 3 | 0 | 1 | 7 | 8 | 13.1 | |||
| DC 4 | 1 | 0 | 10 | 11 | 18.0 | |||
| DC 5 | 4 | 5 | 26 | 35 | 57.4 | |||
| Total | 5 | 6 | 50 | 61 | 100 | |||
Table 3.
Technological categories: R indicates the retouched pieces, B indicates the pieces produced by the bipolar technique (Excavation Borzatti)
| Layer C | Layer D | Layer E | Total | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Technological category | Total | R | B | Total | R | B | Total | R | B | Total | R | B |
| Cortical flakes | 2 | 2 | ||||||||||
| Flakes | 1 | 1R | 2 | 1R | 8 | 3R | 1B | 11 | 5R | 1B | ||
| Flakelets <2.5 cm | 16 | 1R | 6B | 16 | 1R | 6B | ||||||
| Flakes fragmented | 2 | 2R | 7 | 2R | 2B | 9 | 4R | 2B | ||||
| Blades | 5 | 3R | 4B | 5 | 3R | 4B | ||||||
| Bladelets <2.5 cm | 1 | 1 | 1R | 1B | 2 | 1R | 1B | |||||
| Cores | 2 | 1R | 2B | 1 | 1R | 3 | 2R | 2B | ||||
| Indeterminate fragment >DC 3 | 1 | 1 | 1R | 1B | 7 | 3R | 1B | 9 | 4R | 2B | ||
| Debris DC 1–2 | 4 | 4 | ||||||||||
| Total | 5 | 2R | 2B | 6 | 5R | 2B | 50 | 13R | 14B | 61 | 20R | 18B |
Layer C is represented by only five items, and noteworthy is the presence of two cores that were reduced by the bipolar technique, one of which was then retouched (Fig. 4C). Layer D presents six items, among which there are two flakes and one bladelet. Layer E is the one with the most items (50), including one core, 16 flakelets, eight flakes and five blades. Complete cortex coverage is present in only two flakes, whereas three flakes present between 50 and 75%; all the other items present between 25% and 0 cortex coverage. The bipolar technique is attested to on 18 items in almost all the categories (Table 3).
Figure 4.
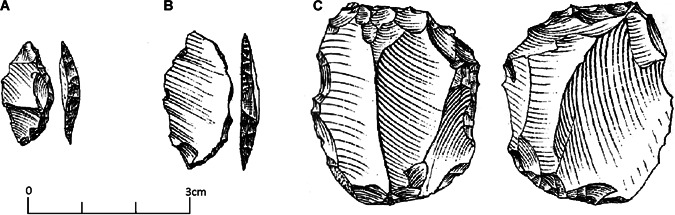
A. Lunates from layer E. B. Lunates from layer C. C. Splintered piece/core from layer C (modified from Borzatti, 1965).
The assemblage includes 20 retouched pieces made mainly on flakes, flake fragments and indeterminate fragments. Most of the retouched tools occurred on pieces with a length >3 cm. Typologically, there is a prevalence of scrapers (n = 11). Noteworthy is the presence of one end scraper and two lunates (Fig. 4A <B). The latter is the most representative retouched tool of the Uluzzian (Table 4).
Table 4.
Retouched tools (Excavation Borzatti)
| Retouched tools | Layer C | Layer D | Layer E | Total |
|---|---|---|---|---|
| Unilateral scraper | 1 | 2 | 7 | 10 |
| Bilateral scraper | 1 | 1 | ||
| End scraper | 1 | 1 | ||
| Lunate | 1 | 1 | 2 | |
| Limace | 1 | 1 | ||
| Point | 1 | 2 | 3 | |
| Indeterminate retouched | 2 | 2 | ||
| Total | 2 | 5 | 13 | 20 |
From a diachronic point of view, chert is the prevailing raw material in all technological classes in the three layers. A greater variety of raw material and greater quantity of pieces is found in layer E. Layers C and D present very few entire pieces, in both cases the majority being retouched tools. Layer E displays a set of large retouched tools and a smaller component of flakelets (Fig. 5).
Figure 5.
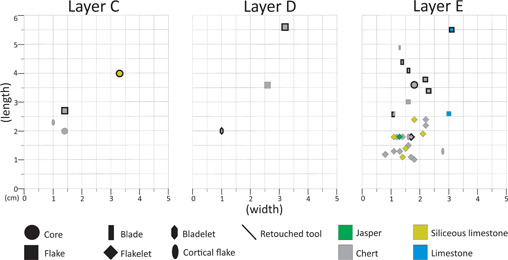
Scatter plot of the lithic items coming from Borzatti's excavation. The technological categories are shown with different symbols; the raw materials are shown with different colours. The items marked with a dark black line are the retouched tools. As the plot is based on length/width, only entire items were considered. [Color figure can be viewed at wileyonlinelibrary.com].
Technological analysis: 2016–2018 excavation
The industry is not particularly affected by post‐depositional alterations and there are no pieces presenting double patinas. All lithics have fresh margins, except for two pieces that present blunt edges, and only six pieces show traces of combustion.
Regarding the raw materials, we observe a selection of fine‐grained raw materials of different nature: nine pieces were extracted from pebbles, four pieces from slabs of local siliceous limestone and 11 from local chert lists. Even without evident traces of cortex, it was still possible to associate the siliceous limestone with the local outcrops, used in the nearby sites throughout the Mousterian and the Uluzzian (Spinapolice, 2012, 2018; Moroni et al., 2013; Spinapolice et al., 2018). The chert, by contrast, is partly local (white and blue chert), coming from the same outcrops where the siliceous limestone was found, and partly (beige or very glassy grey chert) similar to the materials of the Apennines or of the Gargano area. We also noted the sporadic presence of jasper and quartzite (Table 5), probably coming from the Bradano River basin (Basilicata, southern Italy).
Table 5.
Raw material (Excavation 2016–2018)
| SU 3 | SU 15 | SU 17 | ||||||
|---|---|---|---|---|---|---|---|---|
| Raw material | N | % | N | % | N | % | Total | % |
| Chert | 25 | 78.1 | 56 | 74.7 | 170 | 73.3 | 251 | 74.0 |
| Siliceous limestone | 3 | 9.4 | 14 | 18.7 | 46 | 19.8 | 63 | 18.6 |
| Jasper | 1 | 3.1 | 4 | 5.3 | 4 | 1.7 | 9 | 2.7 |
| Quartz sandstone | 1 | 1.3 | 3 | 1.3 | 4 | 1.2 | ||
| Indeterminate | 3 | 9.4 | 9 | 3.9 | 12 | 3.5 | ||
| Total | 32 | 100 | 75 | 100 | 232 | 100 | 339 | 100 |
Most of the lithics, corresponding to 80.7% of the collection, are included in the first (71.4%) and second (11.5%) dimensional classes (Table 6). Most of them (250 items) are debris, which is the waste of the debitage and indicates that flaking activities were performed in situ. For these fragments, it is not possible to detect the concept of debitage. Among these artefacts, 26.2% (corresponding to 89 items) are the more diagnostic pieces, which are useful to infer information on the reduction sequences performed at the site. The initialization of the reduction sequence is attested to by only two semi‐cortical flakes, and hence the first stage of debitage is not documented at the site (or at least in the portion of the site under investigation).
Table 6.
Dimensional classes (Excavation 2016‐2018)
| SU 3 | SU 15 | SU 17 | ||||||
|---|---|---|---|---|---|---|---|---|
| Dimensional class | N | % | N | % | N | % | Total | % |
| DC 1 | 19 | 59.4 | 39 | 52.0 | 184 | 79.3 | 242 | 71.4 |
| DC 2 | 4 | 12.5 | 10 | 13.3 | 25 | 10.8 | 39 | 11.5 |
| DC 3 | 1 | 3.1 | 7 | 9.3 | 11 | 4.7 | 19 | 5.6 |
| DC 4 | 2 | 6.3 | 9 | 12.0 | 5 | 2.2 | 16 | 4.7 |
| DC 5 | 6 | 18.8 | 10 | 13.3 | 7 | 3.0 | 23 | 6.8 |
| Total | 32 | 100 | 75 | 100 | 232 | 100 | 339 | 100 |
The phases of full debitage are mainly attested to by the production of flakelets (Fig. 6), blades and bladelets (Fig. 7) (Table 7). Also, a significant number of fragmented flakes and fragmented blade‐bladelets are present (Table 7). The blanks show predominantly unidirectional scar patterns (n = 43). Some blanks show convergent (n = 8), orthogonal (n = 8) or bidirectional (n = 5) scar patterns. Most pieces exhibit a rectilinear profile (n = 58), but there are some that present a convex (n = 7), wavy (n = 4) or concave (n = 2) profile. The butts are mainly flat (n = 15), point‐form (n = 15) or linear (n = 14). The impact point is mainly central or diffused. The percussion bulb is mostly not prominent. The flat or linear butt, the non‐prominent bulb, the diffused point of percussion and the linear profile, as well as the considerable variability in the debitage‐objective's morphology (Figs. 6 and 7), are typical traits of the bipolar technique. This technique is found on 32 items, mainly flakelets, flake fragments, bladelets and the four cores.
Figure 6.

Flakelets (A–G, L) and fragmented flakes (H–K). Note the small size and the absence of a standardization in the shapes. Blanks produced by the bipolar technique on an anvil (B, D, E, F, H, I, J). Note that J has very similar ventral and dorsal face of the flake; E, F and H have ripple marks, and E, F and J have flat or smashed bulbs. [Color figure can be viewed at wileyonlinelibrary.com].
Figure 7.

Bladelets (A, B, D, E, F, G), blade (C, K), fragmented blade‐bladelets (H, I, J, L, M). Blanks produced by the bipolar technique on an anvil (A–D); blanks produced by direct percussion technique (E–M). Note that the raw material is fine‐grained chert. [Color figure can be viewed at wileyonlinelibrary.com].
Table 7.
Technological categories: R indicates the retouched pieces, B indicates the pieces produced by the bipolar technique (Excavation 2016–2018)
| SU 3 | SU 15 | SU 17 | Total | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Technological category | N | % | R | B | N | % | R | B | N | % | R | B | N | % | R | B |
| Flakes | 2 | 2.7 | 2 | 0.6 | ||||||||||||
| Flakelets <2.5 cm | 4 | 12.5 | 1B | 10 | 13.3 | 2R | 5B | 13 | 5.6 | 5B | 27 | 8 | 2R | 11B | ||
| Flakes fragmented | 2 | 6.3 | 1B | 6 | 8.0 | 1R | 1B | 17 | 7.3 | 2R | 6B | 25 | 7.4 | 3R | 8B | |
| Blades | 4 | 5.3 | 1B | 4 | 1.2 | 1B | ||||||||||
| Bladelets <2.5 cm | 4 | 5.3 | 1B | 3 | 1.3 | 2B | 7 | 2.1 | 3B | |||||||
| Blade–bladelets fragmented | 1 | 3.1 | 5 | 6.7 | 1B | 6 | 2.6 | 1B | 12 | 3.5 | 2B | |||||
| Cores | 1 | 3.1 | 1B | 3 | 1.3 | 3B | 4 | 1.2 | 4B | |||||||
| Indeterminate fragments >DC 3 | 2 | 6.3 | 1 | 1.3 | 1R | 5 | 2.2 | 1B | 8 | 2.4 | 1R | 1B | ||||
| Debris DC 1–2 | 22 | 68.8 | 43 | 57.3 | 2B | 185 | 79.7 | 250 | 73.7 | 2B | ||||||
| Total | 32 | 100 | 0R | 3B | 75 | 100 | 4R | 11B | 232 | 100 | 2R | 18B | 339 | 100 | 6R | 32B |
| Total (without debris) | 10 | 32 | 47 | 89 | ||||||||||||
The assemblage counts six retouched pieces made on flakelets, flake fragments and indeterminate fragments (Table 7). Typologically, there are four side scrapers and two transversal scrapers. Four are made from chert, one from jasper and one from siliceous limestone. These items pertain to DCs 4 and 5.
The four cores are characterized by: (i) an advanced state of exploitation and (ii) the use of the bipolar technique on an anvil (Fig. 8). Specifically, the raw blocks chosen to be flaked are flakes or fragments made of chert. The dimension of the natural supports is relatively small (length 22–16 mm; width 15–12 mm; thickness 11–6 mm). The striking platforms were used without any preparation and are orthogonal to the debitage surface. The scars on the surface of the debitage have mainly a bidirectional pattern. Due to the small dimensions of the cores, the extracted blanks are also very small: <2 cm. This size is compatible with the objective of the debitage.
Figure 8.

Bipolar cores from SU 17 (A–C). [Color figure can be viewed at wileyonlinelibrary.com].
To summarize: SU 3 is characterized by very few items, mainly made of chert; SU 15 is characterized by the most scattered distribution of pieces based on size and presents the most significant variety of raw material, i.e. mainly chert, but there is also a flake made in jasper and three pieces made in siliceous limestone. SU 17 is the one that has the most even distribution of items based on their dimensions, most of its blanks are flakelets and it is where the cores were found (Fig. 9). These three SUs represent an occupation palimpsest made up by at least two types of activities: (i) the small blanks made of chert are the result of in situ flaking; while (ii) the bigger items, the retouched tools and the blanks made of a raw material other than chert, come from reduction sequences that are not attested to in the excavated portion of the site.
Figure 9.
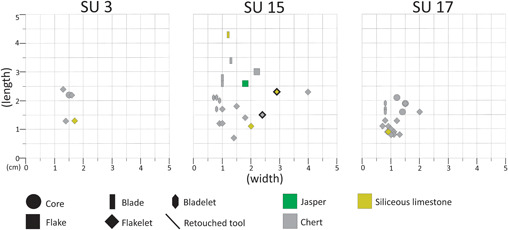
Scatter plot of the lithic items coming from the new excavation. The technological categories are shown with different symbols; the raw materials are shown with different colours. The items marked with a dark black line are the retouched tools. As the plot is based on length/width, only entire items were considered. [Color figure can be viewed at wileyonlinelibrary.com].
Lithic use‐wear analysis
Within the sample of six retouched specimens, diagnostic use‐wear was identified on only one transversal scraper (R76 – Fig. 10A). It exhibits a developed edge rounding and a weak polish with rough texture on the retouched portion (Fig. 10). The traces suggest the tool was used for hide‐working activities by using a transversal motion (scraping). A few bright spots were also detected on the proximal edge which could indicate that the tool had been hafted.
Figure 10.

Retouched tools. The pointed line indicates the presence of use‐wear. A, B: transverse scrapers, C: lateral scraper. [Color figure can be viewed at wileyonlinelibrary.com].
The other transversal scraper (R54 – Fig. 10B) shows unclear traces on the retouched edge. The remaining four pieces did not reveal any traces at all.
Zooarchaeological analysis
Macromammal bone assemblage: Borzatti's excavation
The composition of the faunal assemblage for all the stratigraphic sequence was analysed by Borzatti following his excavation (1964). The bone remains appeared fragmented and scarce, limiting the zooarchaeological considerations. However, in spite of this, he tried to infer information on the palaeoclimatic and palaeoenvironmental conditions at the Uluzzo C Rock Shelter. The faunal composition was characterized by the presence of Equus caballus, Equus asinus hydruntinus, Cervus elaphus, Dama dama, Bos sp., Sus scrofa, Vulpes vulpes, Canis lupus, Lepus sp., Merula merula, Falco tinnunculus, Turdus musicus and Columba palumbus.
In the stratigraphic sequence, considering the transition from Borzatti's layer D to C, it is important to highlight that the increase in the presence of equid remains (dry open environments) corresponds to the decrease of cervids (forest) and bovids (open humid landscape). Borzatti noted the change of a pivotal element in the stratigraphic sequence: the presence of humidity in different phases. For layer C, he observed an abundant presence of Equus, which indicates grassland habitats, and minor evidence regarding the presence of Cervidae and Bovidae compared to layer D (Borzatti von Lowerstern, 1965). He suggested that a fluctuation in the climatic conditions may have affected the extension or reduction of the forests and grasslands surrounding the site.
Macromammal bone assemblage: 2016–2018 excavation
Only 119 out of 2304 bone remains from the 2016–2018 excavations (5.1% of the faunal assemblage) were identifiable to a taxonomic level (Table 8). Eleven genera and nine species belonging to four different orders of mammals (lagomorphs, carnivores, perissodactyls and artiodactyls) were recognized. Some of these mammals are represented by very few specimens (Mustela nivalis, Martes sp., Meles meles, Capreolus capreolus, Rupicapra sp.), while others are represented by a fair quantity of bone elements (Vulpes vulpes, Cervus elaphus). Among the layers that were considered, differences in faunal composition are minimal, with Vulpes vulpes and Cervus elaphus always present, while Equus ferus, S. scrofa and Caprinae present only in SUs 17 and 15 (Fig. 11).
Table 8.
Mammal NISP (number of identified specimens) for SUs 3, 15 and 17
| SU 3 | SU15 | SU 17 | ||||
|---|---|---|---|---|---|---|
| Uluzzo C – Sector A | NISP | NISP | NISP | Total | % | |
| Lepus sp. | 1 | 5 | 6 | 5.0 | ||
| Total Rod. –Lagomorpha | 1 | 5 | 6 | 5.0 | ||
| Vulpes vulpes | 1 | 3 | 6 | 10 | 8.4 | |
| Vulpes sp. | 2 | 1 | 3 | 2.5 | ||
| Mustela nivalis | 1 | 1 | 0.8 | |||
| Meles meles | 1 | 1 | 0.8 | |||
| Martes sp. | 1 | 1 | 0.8 | |||
| Carnivora indet. | 2 | 4 | 6 | 5.0 | ||
| Total Carnivora | 1 | 7 | 14 | 22 | 18.4 | |
| Equus ferus | 5 | 5 | 10 | 8.4 | ||
| Sus scrofa | 4 | 3 | 7 | 5.8 | ||
| Cervus elaphus | 1 | 2 | 10 | 13 | 10.9 | |
| Capreolus capreolus | 1 | 1 | 0.8 | |||
| Cervidae | 1 | 4 | 5 | 4.2 | ||
| Bos/Bison | 5 | 5 | 4.2 | |||
| Rupicapra sp. | 1 | 1 | 0.8 | |||
| Caprinae | 2 | 3 | 5 | 4.2 | ||
| Ungulata | 1 | 5 | 38 | 44 | 36.9 | |
| Total Ungulata | 2 | 19 | 70 | 91 | 76.4 | |
| Total NISP | 4 | 26 | 89 | 119 | 100 | |
| Small mammals | 1 | 1 | 12 | 14 | 0.64 | |
| Small to medium‐sized mammals | 1 | 6 | 7 | 0.32 | ||
| Medium‐sized mammals | 1 | 4 | 8 | 13 | 0.59 | |
| Medium‐large sized mammals | 1 | 7 | 30 | 38 | 1.7 | |
| Large mammals | 3 | 7 | 24 | 34 | 1.5 | |
| Indet. sized mammals | 94 | 269 | 1716 | 2079 | 95.1 | |
| Total mammals indet. | 100 | 289 | 1796 | 2185 | 94.8 | |
| Total NR | 104 | 315 | 1885 | 2304 | 100 | |
| Fragm. burned | 15 | 24 | 108 | 147 | 96.7 | |
| Fragm. calcined | 1 | 3 | 1 | 5 | 3.2 | |
| Total burned | 16 | 27 | 109 | 152 | 6.5 |
Figure 11.

Some determined mammal remains from SUs 15 and 17. (1) Equus ferus. LM3. SU 15. (2) Equus ferus. Ilower. top SU 17. (3) Sus scrofa. Jugal tooth. SU 15 V tg. (4) Sus scrofa. RI1−2. SU 17 I tg. (5) Cervus elaphus. LI1 SU 17. (6) Cervus elaphus. RM1 SU 17 I tg. (7) Cervus elaphus. First phalanx SU 17. (8) Cervus elaphus. I3. SU 15. (9) Capreolus capreolus. RP2. SU 17 I tg. (10) Bos sp. I1−2. SU 17 I tg. (11) Bos/Bison. LI4. SU 17. (12) Rupicapra sp. RM3. SU 17. (13) Lepus sp. Left posterior proximal diaphysis of femur. SU 17 II tg. (14) Vulpes vulpes. RP4. SU 17 II tg. (15) Vulpes sp. Proximal phalanx. SU 15. (16) Meles meles. Lower incisor. SU 17 I tg. (17) Mustela nivalis. C1. SU 17. (18) Martes sp. c1. SU 17 I tg. [Color figure can be viewed at wileyonlinelibrary.com].
However, in SU 17 the presence of red deer increases compared to SUs 15 and 3. Moreover, only in SU 17 is there the presence of hare, weasel, badger, marten, auroch or bison, roe deer and chamois, which suggests a low‐temperate climate with woodland covering and the presence of open environments.
Given the small number of determined elements and the preliminary state of the analysis of the faunal remains, the MNI was not calculated. Bones are mostly fragmented and over 98.3% of the specimens are smaller than 3 cm. Among the analysed remains, only 9% [number of remains (NR) = 209] are burned: over 97.6% (NR = 204) of these show a brown or black colour (indicating a burning temperature of ca. 200–400 °C) (Stiner et al., 1995; Costamagno et al., 2005) and only 2.3% (NR = 5) are small calcinated fragments (<2 cm). Among the analysed bones, 19.2% (NR = 444) have concretions, while 15.7% (NR = 362) are characterized by a manganese coating and a few other remains present root furrows.
Evidence of butchering activity is provided by three bone remains with cutmarks that are probably linked to defleshing activities. In detail, two of the three elements are diaphyseal fragments of ribs, one from a large ungulate and one from either Bos primigenius or Bison priscus. In both these remains, the butchering traces are located on the dorsal surface and are short, closely related, and oblique to the major axis of the anatomical element (Fig. 12). The third is a left medial proximal diaphyseal fragment of Lepus sp. with evident scraping‐marks, probably associated with the removal of muscular mass and flesh (Fig. 12). Moreover, some bone fragments showed traces of acid corrosion from partial digestion.
Figure 12.

Bone remains with anthropic marks from SUs 15 and 17. (1) Bos/Bison rib with defleshing cutmarks from SU 17 II tg: a–e, close up of traces. (2) Impact flakes from SUs 17 II tg. and 17. (3) Medium‐sized to large ungulate rib with defleshing cutmarks from SU 17 II tg. Close up of single stria. (4) Lepus sp. proximal diaphysis of femur with scrape‐marks from SU 17 II tg. a and a', close up of traces. [Color figure can be viewed at wileyonlinelibrary.com].
Finally, we identified three percussion cones indicating the activity of bone fracturing for the extraction of marrow (Fig. 12).
Micromammals
The micromammal assemblage consists of 25 identified specimens, corresponding to a minimum of 17 individuals, and representing only three taxa [Myotis myotis, Microtus (Terricola) savii and Apodemus (Sylvaemus) sp.] (Fig. 13; Table 9).
Figure 13.
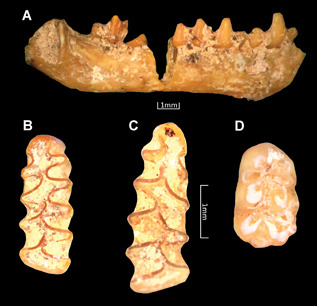
Some micromammal remains identified from Uluzzo C Rock Shelter. A (UC2018/sett. A/AA11/SU 17 tetto): right mandible of Myotis myotis in buccal view; B (UC2018/sett. A/A11/SU 15–17) and C (UC2018/sett. A/A11n/SU 17 II tg.): left and right first lower molars of Microtus (Terricola) savii in occlusal view; D (UC2018/sett. A/A11/SU 17 I tg.): right first lower molar of Apodemus (Sylvaemus) sp. Scale bars, 1 mm. [Color figure can be viewed at wileyonlinelibrary.com].
Table 9.
Minimum number of individuals (MNI) and number of identified specimens (NISP) of the identified species by stratigraphic units of Uluzzo C Rock Shelter
| SU 15 | SU 17 | Total | |
|---|---|---|---|
| Myotis myotis | 0 | 1 | 1 |
| Microtus (Terricola) savii | 3 | 12 | 15 |
| Apodemus (Sylvaemus) sp. | 0 | 1 | 1 |
| MNI total | 3 | 14 | 17 |
| NISP total | 4 | 21 | 25 |
A total of 24 lower first molars of Savi's pine vole [Microtus (Terricola) savii] from the totality of the stratigraphic units were studied. All the observed teeth presented some degree of digestion (Table 10), indicating that the micromammal accumulation was caused by the action of predation. According to Andrews (1990) and Fernández‐Jalvo et al. (2016a), predators from Categories 1 to 4 – which include a broad spectrum of nocturnal and diurnal birds of prey – could be responsible for such an accumulation of micromammal remains.
Table 10.
Percentages of Microtus (Terricola) savii lower first molars from Uluzzo C Rock Shelter showing the different degrees of digestion. NR, number of remains used for the taphonomic analysis
| NR | % | |
|---|---|---|
| Light | 16 | 64 |
| Moderate | 3 | 12 |
| Heavy | 5 | 20 |
| Total | 25 | 100 |
Avifauna
Of 12 bird bone remains, 11 have been identified either at the species level or at the supraspecific level. The poor state of preservation of the remains, due to their fragmentary nature and the presence of concretions that concealed the bones’ diagnostic features, allowed us to identify at a specific level only two remains, which luckily provided precise palaeoenvironmental indications. Melanocorypha calandra suggests the presence, in the surroundings of the cave, of open environments such as grasslands and steppes, with bare terrains and low vegetation, whereas Pyrrhocorax pyrrhocorax points to the presence of rocky cliffs and crags with grassland areas (Cramp, 1998).
The taphonomic analysis detected the presence of some natural modifications. The most abundant ones are manganese dioxide staining and sediment concretions, but we also have identified traces of root etching and trampling (Fernández‐Jalvo and Andrews, 2016b). No modifications by carnivores, nocturnal raptors or humans have been found. The finding of Corvidae and Falco sp. remains, together with the P. pyrrhocorax remains, could be due to the natural death of birds living close to the rock shelter, which could have served as a nesting place for these species (Cramp, 1998).
Malacofauna
The malacological assemblage includes 20 specimens belonging to Patella sp. and Phorcus turbinatus (Table 11). Moreover, a sea urchin fragment was discovered in SU 17. Six small fragments are the only invertebrate remains found in SU 15 and just one was identified as Patella sp. Moving down the stratigraphic sequence (SU 17), the amount of invertebrate remains becomes more abundant and two whole Ph. turbinatus were detected (Fig. 14). These species live in a littoral environment, i.e. intertidal rocky shores. The malacological assemblage does not show pre‐depositional alterations (Table 12) – generally connected to post‐mortem damage caused by shore abrasion, bioerosion, carnivorous gastropods or other predators (such as birds or crabs) – ultimately suggesting that the shells were gathered alive and carried to the site. Nonetheless, we do not exclude the possibility at some of the damages might have been caused by post‐depositional processes. Furthermore, 5% of the remains are characterized by cracks that are consistent with exposure to high temperatures, as also suggested by the decalcification index (total layers: 60%), since heat exposure (above 300 °C according to Milano et al., 2016) modifies the shell structure and leads to dissolution and fragmentation. Thin sections of small fragments of unidentified mollusc shells collected from most of the SUs of the deposit were observed (Spinapolice et al., this special issue).
Table 11.
Mollusc taxa found at Uluzzo C with relative abundances (as NISP) and their habitat
| Taxon | Authority | Habitat | SU 15 | SU 17 | Total |
|---|---|---|---|---|---|
| Gasteropoda indet. | 5 | 2 | 7 | ||
| Patella sp. | Linnaeus 1758 | Interdital rocky shore | 1 | 5 | 6 |
| Phorcus turbinatus | (Born, 1778) | Interdital rocky shore | – | 7 | 7 |
| Total | 6 | 14 | 20 |
Figure 14.

Two whole individuals of Phorcus turbinatus discovered in Uluzzo C site from SU 17 (A–B). [Color figure can be viewed at wileyonlinelibrary.com].
Table 12.
List of taphonomic processes by identified marine taxa
| Layers | SU 15 | SU 17 | Total | % | |
|---|---|---|---|---|---|
| Nisp | 6 | 14 | 20 | 100 | |
| Pre‐dep. Alt. | – | – | – | – | |
| Anthrop. Alt. | TAlt. | – | 1 | 1 | 5 |
| Post‐dep. Alt. | Decal. | 1 | 11 | 12 | 60 |
| Fragm. | 2 | 10 | 12 | 60 | |
| EscD. | – | 1 | 1 | 5 | |
Abbreviations: pre‐dep. Alt. – pre‐depositional alterations; Antrop. Alt. – anthropogenic alterations; TAlt – thermo‐alteration damage; Post‐dep. Alt. – post‐depositional alterations; Decal – decalcification; Fragm. – fragmentation; EscD. – excavation damage. Note: some specimens could have been exposed to more than one type of taphonomic alteration.
ZooMS
Twelve bone fragments coming from SUs 15 and 17 were analysed to test the collagen preservation at Uluzzo C Rock Shelter. From this preliminary assessment, we observed that collagen is poorly preserved. Of the 12 samples here considered, only five provided taxonomic information, using standard peptide markers (Buckley et al., 2009; Welker et al., 2016). In all the samples, P1105 is highly deamidated, suggesting that these bones probably yielded antique endogenous collagen. We identified R116 as an Equidae, UC81 as a Bos/Bison and UC92 as a Cervidae. R35 and UC91 could belong either to Cervidae or Equidae, due to the lack of distinctive diagnostic peptides (Table 13).
Table 13.
Diagnostic peptides detected for the different samples and taxonomic identification based on CO1 peptide markers (see Welker et al., 2016)
| Peptide markers | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample | P1 | A | A′ | B | C | P2 | D | E | F | F′ | G | G′ | Taxonomic identification |
| R35 | 1105.6 | 1427.7 | 1550.8 | 2883.4 | Cervidae/Equus | ||||||||
| R116 | 1105.6 | 1427.7 | 1550.8 | 2145.1 | 2883.4 | Equidae | |||||||
| UC81 | 1105.6 | 1427.7 | 1580.8 | 1648.8 | 2131.1 | 2853.4 | Bos/Bison | ||||||
| UC91 | 1105.6 | 1427.7 | 1550.8 | 2883.4 | Cervidae/Equus | ||||||||
| UC92 | 1105.6 | 1427.7 | 1550.8 | 1648.8 | 2131.1 | Cervidae | |||||||
All the other tested fragments did not yield sufficient collagen/peptides for ZooMS taxonomic identification.
Ancient DNA analysis
Of the 14 sediment samples tested from SUs 3, 15 and 17, three contained traces of ancient mammalian mtDNA. All three positive samples were collected in SU 15. They contained ancient mtDNA fragments assigned to Equidae and/or Hyaenidae (Table 14). Given the low number of mtDNA fragments representing these mammalian families, no attempts were made to identify the taxa at the genus or species levels. None of the tested samples contained evidence for the preservation of ancient hominin mtDNA (Table S3λ).
Table 14.
Ancient mammalian taxa identified in sediment samples using ancient DNA analyses. Only the three samples positive for the preservation of ancient DNA and only the taxa identified as ancient in each sample are shown. The number of mtDNA fragments assigned to each family is noted on the left column, and the percentage of fragments carrying a cytosine to thymine (C → T) substitution to the relevant reference genome at their 5′‐ and 3′‐ends [with the lower bound of a one‐sided 95% confidence interval (CI) in parentheses] are noted on the middle and right columns, respectively. Detailed results for all samples and negative controls are shown in Table S3
| Equidae | Hyaenidae | ||||||
|---|---|---|---|---|---|---|---|
| Sample | Provenance | No. of fragments | % 5 ′ C → T (low 95% CI) | % 3 ′ C → T (low 95% CI) | No. of fragments | % 5 ′ C → T (low 95% CI) | % 3 ′ C → T (low 95% CI) |
| C147 | SU 15; Square A11 | 21 | 50.0 | 75.0 | 68 | 50.0 | 50.0 |
| (22.2) | (24.9) | (24.5) | (27.9) | ||||
| C170 | SU 15; Square A11 | 20 | 42.9 | 75.0 | – | – | – |
| (12.9) | (24.9) | ||||||
| C173 | SU 15; Square AA11 | – | – | – | 23 | 50.0 | 75.0 |
| (15.3) | (24.9) | ||||||
Discussion
Lithic behaviour
Despite the small number of available pieces, we can draw some general conclusions on the production and use of lithic resources at Uluzzo C, based on analysis of the lithic assemblage pertaining to both the old and new excavations. Layer C and SUs 3, 15 and 17 represent a palimpsest of different partial reduction sequences. Due to the presence of small fraction debris (DC 1–2), part of the knapping activities was apparently performed directly on site, whereas the initialization of the block possibly occurred outside the shelter (or in an area that has not yet been excavated). Blocks were introduced to the site at a medium stage of exploitation (as suggested by the low amount of attested cortical pieces). We highlight the presence of two main debitage objectives: bladelets (Fig. 7) and flakelets (Fig. 6). At the current state of research, these objectives seem to have been achieved by: (i) unidirectional debitage using the bipolar technique with no or little management of the convexities – which was observed on the basis of the four bipolar cores and the bipolar debitage products; and (ii) a unidirectional production characterized by the direct percussion technique with the lateral and distal convexities being managed – based on the aligned negatives on the blanks. At this stage of the research, it is not possible to determine whether these two components are part of the same reduction sequence – which would imply the use of both direct and bipolar techniques in different moments of the reduction (e.g. such as at Roccia San Sebastiano – Collina et al., 2020)‐ or if they are two different and independent reduction sequences. In both cases, the result is a low degree of standardization for both shape and edge delineation of the products, but a consistency in blank dimensions can be observed.
Among the retouched tools, two lunates were found in levels C and D of Borzatti's excavation. However, their use as projectiles – as has been attested to at Grotta del Cavallo (Sano et al., 2019) – has not yet been tested.
Based on the technical evidence collected so far, we can consider the lithic assemblage of Uluzzo C as falling within the framework of the Uluzzian, since it presents several of the main characteristics of this techno‐complex: principally local raw materials; additional concepts of debitage (unidirectional reduction sequence); use of the bipolar technique on an anvil; production of flakes and bladelets with several morphologies; low degree of standardization of the products; presence of lunates and end‐scrapers and the absence of integrated concepts (i.e. Levallois) (Moroni et al., 2018; Collina et al., 2020; Marciani et al., 2020a).
A point that deserves more attention is the production of blade‐bladelets. These items are usually related to UP industries, even if their production has also been attested to in the Uluzzian (for a review see Marciani et al., 2020a) and before, during the Mousterian (Ranaldo et al., 2017; for a review see Carmignani, 2017). Even though the end‐products are similar, the method of their production and the role that these objects may have played within these techno‐complexes are different. Blade/bladelet reduction systems in the UP come from an integrated production where both the procedure and the traits of the obtained object are standardized (Marciani et al., 2020a). For example, the Protoaurignacian is a bladelet‐dominated industry with a significant technical investment in the production phase and standardized products that are obtained by a laminar–lamellar debitage on unidirectional and prismatic cores (e.g. Falcucci, 2018; Negrino and Riel‐Salvatore, 2018). In contrast, the degree of standardization of the Uluzzian bladelets is undoubtedly lower. Furthermore, and perhaps of even greater importance, the conceptualization of the manufacture procedure is different, as the Uluzzian uses an additional unidirectional and orthogonal debitage. Moreover, the bipolar technique is used as a deliberate choice and is not dictated by the raw material (i.e. Moroni et al., 2018; Collina et al., 2020; Marciani et al., 2020a). The presence of a possibly more standardized blade–bladelet production at Uluzzo C needs to be assessed in further detail, preferably with the addition of the materials from the other layers.
Hunting behaviour
At Uluzzo C, the lithic industry is associated with bones from different taxa. Despite the fragmentary state of the macromammal remains, there is evidence for the defleshing of large ungulates, as in other areas of Italy in this period (for a complete review, see Romandini et al., 2020). The analysis of the faunal assemblage from the more recent excavations agrees with the results obtained by Borzatti, both for the identified taxa (Lepus sp., Vulpes vulpes, Equus ferus, Sus scrofa, Cervus elaphus, Bos/Bison) and the relatively large quantity of Equus remains that were recognized (Borzatti von Lowerstern,1965).
As for the Uluzzian levels of the nearby Grotta del Cavallo (i.e. EIII5), the most represented taxa in Uluzzo C correspond to Equus and Cervus elaphus. The taxonomic information obtained from the analysis of the faunal assemblage is coherent with the preliminary results of ZooMS and ancient DNA analyses.
Moreover, there are some common qualitative elements between these two sites: the predominant distribution of cutmarks on ungulate ribs and the presence of traces related to breakage of long bones, such as percussion cones or fractures and impact traces on diaphyseal long bones which are related to marrow extraction activities (Boscato and Crezzini, 2012; Romandini et al., 2020). Unlike at Grotta del Cavallo, within Uluzzo C small to medium‐sized mammals such as Vulpes vulpes and Lepus sp. are abundantly present.
Furthermore, this site is interesting for the exploitation of marine resources. Despite the limited amount of malacological remains, we can speculate on human consumption of this resource: collected gastropods are indeed commonly associated with dietary purposes during the MP and UP (Stiner, 1999, 2009; Colonese and Wilkens, 2005; Zilhão et al., 2010; Colonese et al., 2011; Hunt et al., 2011; Bosch et al., 2015; Hill et al., 2015; Milano et al., 2016; Ramos‐Muñoz et al., 2016). Today, the site of Uluzzo C is a few metres above the sea, but at the time it was a few kilometres beyond the emerged shelf, so it is unlikely that animals or other agents could have been responsible for the transportation of the molluscs. Additionally, a few fragments of Ph. turbinatus present evidence of thermal alterations that might be consistent with intentional heating for consumption purposes. From these preliminary findings, the presence of humans and rocky shore intertidal molluscs appears to be correlated, but more data are required to validate this hypothesis.
Climatic and environmental reconstruction
Together, the information gathered through micromammals, macromammals and bird remains has disentangled the palaeoclimatic and palaeoenvironmental conditions related to the Uluzzian occupation. The micromammal assemblage of Uluzzo C is relatively dominated by the presence of Microtus (Terricola) savii (25 lower first molars), a Mediterranean species that inhabits open meadows and avoids dense forest areas (Amori, 2016). All the observed teeth presented some degree of digestion (Table 10), meaning that the micromammal accumulation must have been the result of predation (see Andrews, 1990; Fernández‐Jalvo et al., 2016a). As pointed out by Berto et al. (2017), Savi's pine vole is a dominant species in both cold and warm phases during the late Pleistocene of southern Apulia in Grotta del Cavallo (Dalla Valle, 2008), Grotta dei Cervi di Porto Badisco (Cason, 2012) and Grotta delle Cipolliane (Bon and Menon, 2000). This means that it is not a good climatic indicator in this area. However, the relative predominance of Microtus (Terricola) savii, together with the presence of Apodemus (Sylvaemus) sp. and Myotis myotis, indicates a patchy landscape composed of open forest and grassland habitats, and compatible with the overall macrofaunal composition.
In accordance with other south‐eastern Ionian sites, an increasing frequency of Equus ferus is registered in the Uluzzian sequence, suggesting the occurrence of sparse woodland and steppic environments, as is the presence of Cervidae, which are typical of Mediterranean evergreen forests (Romandini et al., 2020). The presence of Equidae was attested to by both the DNA analysis of sediment samples and from palaeoproteomics. In this context, in accordance with the archaeozoological record, sample R116, identified by ZooMS as an Equidae, can be attributed to Equus ferus.
In addition, the ancient DNA data shows the presence of Hyaenidae at the site. However, the presence of hyena was not registered by morphological analysis on the osteological remains. This could be caused by the high degree of bone fragmentation of the faunal assemblage from Uluzzo C, which may have hampered the identification of this species. Notably, some bone fragments appear as corroded and rounded after being digested by carnivores.
Concerning avifaunal remains, two species provided precise palaeoenvironmental indications, coherent with the other investigated proxies. In particular, Melanocorypha calandra suggests the presence of grasslands and steppes, while Pyrrhocorax pyrrhocorax indicates the presence of rocky cliffs and crags with grassland areas (Cramp, 1998).
Faunal‐based reconstruction of environmental settings at the time of the Uluzzian occupation of the Uluzzo C Rock Shelter fits well with the palaeoenvironmental reconstruction based on the study of sediments from the stratigraphic sequences reported in Spinapolice et al. (this special issue). The opening of the environment, the increase of aridity and the wind strength are registered in these units by an increase of the silt fraction, corresponding to loess. Dismantling of the roof and walls of the rock shelter and the accumulation of wind‐blown sediments are two sedimentary processes compatible with the cold and arid environmental conditions that occurred during Marine Isotope Stages 4 to 2. Moreover, the occurrence of a frost‐related breccia in layer C, probably formed under severe cold conditions, suggests decreased temperatures and cooler environmental conditions, probably during a stadial event (see Spinapolice et al. in this special issue).
Conclusions
The reopening of the excavation at Uluzzo C Rock Shelter, the reviewing of old material and the study of the artefacts coming from the new excavations allowed us to better bring better into focus the evidence from this site in the context of the Bay of Uluzzo and, more generally, in the framework of the Uluzzian occupation in Italy. The Bay of Uluzzo is a crucial area for understanding of the Uluzzian – not only it is the place where the Uluzzian was first identified and described (Palma di Cesnola, 1964), but it is also where the entire development of the Uluzzian techno‐complex was defined (Palma di Cesnola, 1993). Several sites with Uluzzian occupations are clustered in this area within a few kilometres from each other: Grotta del Cavallo, Grotta di Uluzzo, Uluzzo C Rock Shelter (in the Bay) and Grotta di Serra Cicora (a few kilometres in the hinterland). Palma di Cesnola described here the chrono‐cultural development of this techno‐complex based on typological traits, frequencies of retouched tools and the raw materials that were used. He defined the archaic Uluzzian based on layer EIII of Grotta del Cavallo; the evolved Uluzzian based on layers EII‐I of Grotta del Cavallo; and the late Uluzzian based on layer D of Grotta del Cavallo, layer N of Uluzzo B, and layers D and C of Uluzzo C. The last phase of the Uluzzian (final Uluzzian) is absent at Grotta del Cavallo, and it has only been found at Serra Cicora in horizon D of layer B. The Uluzzian cycle is closed by a phase called the Uluzzo‐Aurignacian, found at Serra Cicora in horizons A, B and C of layer B (Palma di Cesnola, 1993). Within the Bay of Uluzzo, the only other assemblage that was recently studied with a technological approach is the one from layer EIII at Grotta del Cavallo (Moroni et al., 2018).
The acquired evidence suggests that modern human Uluzzian groups settled at Uluzzo C Rock Shelter between 42 and 40 ka (Spinapolice et al., in this issue).
The lithic production of that layer is characterized by a significant component of mostly un‐retouched small blades/bladelets derived mainly from bipolar reduction. Among formal tools, end‐scrapers and backed elements (mainly lunates) have a key role. The lithic assemblage from layer EIII consists of larger tools (e.g. end‐scrapers and side‐scrapers), and smaller tools (that include backed pieces that were presumably used in composite devices) (Moroni et al., 2018).
The lithic materials from Uluzzo C show: (i) the production of bladelets and flakelets; (ii) the presence of the bipolar technique on an anvil; and (iii) the presence of two lunates among the retouched tools. Recognizing the layer C/SUs 3, 15 and 17 as Uluzzian confirms and enriches with further information Borzatti's interpretation, which was mainly based on typological observations. Moreover, this recognition is in accordance with: (i) the reconstructed stratigraphy of the site, where the Uluzzian occupation follows the Mousterian occupation; (ii) the site's location, since the Bay of Uluzzo is the key area for the Uluzzian; and (iii) the chronology, since the grand weighted mean age for the Uluzzian occupations is 40.6 ± 1.4 ka, whereas for the end of the Mousterian it is 46 ± 4.0 ka (Spinapolice et al., in this issue). This chronology is in accordance with the Uluzzian occupation of Grotta del Cavallo dated between 45.5 ± 1.0 and 39.85 ± 0.14 ka (Zanchetta et al., 2018).
Data obtained from the macrofaunal composition of Uluzzo C are consistent with the Uluzzian levels of the nearby Grotta del Cavallo (i.e. EIII5), where Cervidae and Equidae were also identified as the most abundant taxa. The only difference is in the exploitation of small‐ to medium‐sized mammals, such as Vulpes vulpes and Lepus sp., towards which Uluzzo C hunters seem to have been more inclined compared to those of Grotta del Cavallo. Except for the hyena, ZooMS, ancient DNA and archaeozoological analyses are consistent in identifying taxa within this context. Further analyses on the bone fragments recovered during the last excavation season in 2019 from the same levels are in progress. The aim is to improve our knowledge regarding the faunal composition through the application of palaeoproteomics.
In general, the analysis of macromammal, micromammal and bird remains agrees with the data obtained from sediments and from the stratigraphic sequence in indicating a patchy landscape composed of forests, grassland and rocky habitats in a cold and arid environment.
They butchered and consumed their food through the use of fire, hunted – probably near the site – medium‐sized and large ungulates, as well as Leporidae, and collected marine gastropods (Ph. turbinatus) – probably for sustenance purposes – within a few kilometres from the rock shelter.
The application of a multidisciplinary methodological protocol – including lithic technology, use‐wear, zooarchaeology, DNA from sediments and palaeoproteomics – shows the potential of integrated studies in understanding the technical and hunting behaviour of humans in relation to their environment. Finally, this study represents a new fundamental piece in the composition and understanding of the meaning of the complex Uluzzian cultural and ecological mosaic in Italy during the MP/UP transition.
Author contribution
ES, SB are the research and excavation coordinators and conceptualized the paper; SS, GM, MR wrote the first draft of the paper; SS, MR analysed the macrofaunal remains; GM, FR performed the lithic technological analysis; SA performed the use‐wear analysis of the lithic items; LC carried out the analysis of the avifaunal remains; JMLG analysed the small mammal remains; FL, SS performed the ZooMS analysis; VS, MM analysed the DNA from sediments; LT analysed the malacofaunal remains; AF, EB made the topographic mapping; AC described the stratigraphic sequence; AZ was responsible for the geoarchaeological investigation on the stratigraphic sequence; MCM was responsible for the OSL dating campaign; OAH reviewed and edited the language and the manuscript; ES, SB, SS, MR, AF, EB, GO participated in the archaeological excavation of the site. All the authors collaborated in writing, reviewing and editing the final version of the paper.
Supporting information
This article includes online‐only Supplemental Data.
Table S1. Lithic items in each layer of Borzatti excavation, according to Borzatti (1965) and items currently stored at Museo della Preistoria di Nardò MPN.
Table S2. Lithic items in SUs 3, 15 and 17 (excavation 2016–2018).
Table S3. Sequencing results for all tested sediment samples and relevant negative controls. For the mammalian mitochondrial capture, results are only shown for Hominidae and for the two taxa identified as ancient in at least one of the samples. L – length; MQ – mapping quality; C – cytosine; T – thymine; CI – confidence interval.
Acknowledgements
This study received funding from the European Research Council (ERC) under the European Union's Horizon 2020 research and innovation programme (grant agreement No. 724046 – SUCCESS, www.erc-success.eu). Research at Uluzzo C is supported by Leakey Foundation, Spring 2015 Grant awarded to Marco Peresani. The authors are grateful to MiBACT (now MIC) that released the permits to excavate the site and analyse archaeological materials, as well as to the Museo della Preistoria di Nardò, the Soprintendenza Archeologia, Belle Arti e Paesaggio per le province di Brindisi, Lecce e Taranto, and the Gruppo Speleologico Neretino. JMLG was supported by a Ramon y Cajal contract (RYC‐2016‐19386), with financial sponsorship of the Spanish Ministry of Science, Innovation and Universities. This is publication No. 354 of the Museo di Geologia e Paleontologia collections at the Università degli Studi di Torino. The ancient DNA analysis was funded by the Max Planck Society and the European Research Council (grant agreement No. 694707 to Svante Pääbo). We thank E. Essel, F. Müller, B. Nickel, J. Richter, B. Schellbach and A. Weihmann for the preparation of the sediment samples for sequencing. Sequencing data were deposited in the European Nucleotide Archive (accession PRJEB33875). Open access funding provided by Universita degli Studi di Bologna within the CRUI‐CARE Agreement.
Data availability statement
The data that support the findings of this study are available on request from the corresponding author. The data are not publicly available due to privacy or ethical restrictions.
References
- Amori G. 2016. Microtus savii. The IUCN Red List of Threatened Species 2016: e.T13491A513760.
- Aitken RJ. 1974. Delayed implantation in roe deer (Capreolus capreolus). Journal of Reproduction and Fertility 39: 225–233. [DOI] [PubMed] [Google Scholar]
- Altschul SF, Gish W, Miller W et al. 1990. Basic local alignment search tool. Journal of Molecular Biology 215: 403–410. [DOI] [PubMed] [Google Scholar]
- Andrews P editor. 1990. Fossils. Predation, Preservation and Accumulation of Small Mammal Bones in Caves, with an Analysis of the Pleistocene Cave Faunas from Westbury‐Sub‐Mendip. University of Chicago Press: Chicago. [Google Scholar]
- Arrighi S, Bortolini E, Tassoni L et al. 2020c. Backdating systematic shell ornament making in Europe to 45,000 years ago. Archaeological and Anthropological Sciences 12. https://link.springer.com/article/10.1007/s12520-019-00985-3 [Google Scholar]
- Arrighi S, Marciani G, Rossini M et al. 2020a. Between the hammerstone and the anvil: bipolar knapping and other percussive activities in the Late Mousterian and the Uluzzian of Grotta di Castelcivita (Italy). Archaeological and Anthropological Sciences 12. https://link.springer.com/article/10.1007%2Fs12520-020-01216-w [Google Scholar]
- Arrighi S, Moroni A, Tassoni L et al. 2020b. Bone tools, ornaments and other unusual objects during the Middle to Upper Palaeolithic transition in Italy. Quaternary International 551: 169–187. [Google Scholar]
- Benazzi S, Douka K, Fornai C et al. 2011a. Early dispersal of modern humans in Europe and implications for Neanderthal behaviour. Nature 479: 525–528. [DOI] [PubMed] [Google Scholar]
- Benazzi S, Slon V, Talamo S et al. 2015. Archaeology. The makers of the Protoaurignacian and implications for Neandertal extinction. Science 348: 793–796. [DOI] [PubMed] [Google Scholar]
- Benazzi S, Viola B, Kullmer O et al. 2011b. A reassessment of the Neanderthal teeth from Taddeo cave (southern Italy). Journal of Human Evolution 61: 377–387. [DOI] [PubMed] [Google Scholar]
- Berto C (ed). 2013. Distribuzioneevoluzione delle associazioni a piccoli mammiferi nella penisola italiana durante il Pleistocene superiore. ed PhD Thesis, Università degli Studi di Ferrara.
- Berto C, Boscato P, Boschin F et al. 2017. Paleoenvironmental and paleoclimatic context during the Upper Palaeolithic (late Upper Pleistocene) in the Italian Peninsula. The small mammal record from Grotta Paglicci (Rignano Garganico, Foggia, Southern Italy). Quaternary Science Reviews 168: 30–41. [Google Scholar]
- Bietti A, Cancellieri E, Corinaldesi C et al. 2010. La percussion sur enclume en Italie centrale Tyrrhénienne [The bipolar‐on‐anvil percussion in central Thyrrenian Italy]. Paléo (spécial) 143–180. [Google Scholar]
- Binford L. 1981. Bones, Ancient Men and Modern Myths. Academic Press: New York. [Google Scholar]
- Blumenschine RJ. 1995. Percussion marks, tooth marks, and experimental determinations of the timing of hominid and carnivore access to long bones at FLK Zinjanthropus, Olduvai Gorge, Tanzania. Journal of Human Evolution 29: 21–51. [Google Scholar]
- Blumenschine RJ, Selvaggio MM. 1988. Percussion marks on bone surfaces as a new diagnostic of hominid behaviour. Nature 333: 763–765. [Google Scholar]
- Boëda E. 2013. Techno‐logique & technologie. Une Paléo‐histoire des objets lithiques tranchants. @ rchéo‐éditions. com: Paris.
- Bon M, Menon F. 2000. I micromammiferi della grotta delle Cipolliane. Atti. Società per la preistoria e protostoria della regione Friuli Venezia Giulia 8: 37–44. [Google Scholar]
- von Borzatti Lowerstern E. 1965. La Grotta‐riparo di Uluzzo C (campagna di scavi 1964). Rivista di Scienze Preistoriche XX: 1–31. [Google Scholar]
- Borzatti von Lowerstern E, Magaldi D. 1966. Risultati conclusivi dello studio paletnologico e sedimentologico della Grotta di Uluzzo C (Nardò‐Lecce). Rivista di Scienze Preistoriche 21: 16–64. [Google Scholar]
- Boscato P, Crezzini J. 2012. Middle‐Upper Palaeolithic transition in Southern Italy: Uluzzian macromammals from Grotta del Cavallo (Apulia). Quaternary International 252: 90–98. [Google Scholar]
- Bosch MD, Wesselingh FP, Mannino MA. 2015. The Ksâr'Akil (Lebanon) mollusc assemblage: zooarchaeological and taphonomic investigations. Quaternary International 390: 85–101. [Google Scholar]
- Brain CK. 1983. The Hunters or the Hunted?: an Introduction to African Cave Taphonomy. University of Chicago Press: Chicago. [Google Scholar]
- Briggs AW, Stenzel U, Johnson PLF et al. 2007. Patterns of damage in genomic DNA sequences from a Neandertal. Proceedings of the National Academy of Sciences of the United States of America 104: 14616–14621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckley M, Collins M, Thomas‐Oates J et al. 2009. Species identification by analysis of bone collagen using matrix‐assisted laser desorption/ionisation time‐of‐flight mass spectrometry. Rapid Communications in Mass Spectrometry 23: 3843–3854. [DOI] [PubMed] [Google Scholar]
- Capaldo SD, Blumenschine RJ. 1994. A quantitative diagnosis of notches made by hammerstone percussion and carnivore gnawing on bovid long bones. American Antiquity 59: 724–748. [Google Scholar]
- Carmignani L. 2017. From the flake to the blade: the technological evolution of the Middle Paleolithic blade phenomenon. PhD Thesis, Universitat Rovira i Virgili.
- Cason M. 2012. I piccoli mammiferi della fine dell'ultimo Glaciale della Grotta dei cervi a Porto Badisco (Lecce). PhD Thesi, Tesi di Laurea Magistrale in Quaternario, Preistoria e Archeologia.
- Claassen C. 1998. Shells. Cambridge University Press: Cambridge. [Google Scholar]
- Collina C, Marciani G, Martini I et al. 2020. Refining the Uluzzian through a new lithic assemblage from Roccia San Sebastiano (Mondragone, southern Italy). Quaternary International 551: 150–168. [Google Scholar]
- Colonese AC, Mannino MA, Bar‐Yosef Mayer DE et al. 2011. Marine mollusc exploitation in Mediterranean prehistory: an overview. Quaternary International 239: 86–103. [Google Scholar]
- Colonese AC, Wilkens B. 2005. The Malacofauna of the Upper Paleolithic Levels at Grotta Della Serratura (Salerno, Southern Italy). Preliminary Data. Archaeomalacology: Molluscs in Former Environments of Human Behaviour. Oxford Books: Oxford; 63–70. [Google Scholar]
- Costamagno S, Théry‐Parisot I, Brugal JP et al. 2005. Taphonomic consequences of the use of bones as fuel. Experimental data and archaeological applications. In Biosphere to Lithosphere: New Studies Invertebrate Taphonomy (Procedings of 9th ICAZ Conference, Durham 2002), O'Connor T (ed.). Oxbow Books: Oxford; 51–62.
- Cramp S. 1998. The Complete Birds of the Western Palaearctic on CD‐ROM. Optimedia. Oxford University Press: Oxford. [Google Scholar]
- d'Errico F, Borgia V, Ronchitelli A. 2012. Uluzzian bone technology and its implications for the origin of behavioural modernity. Quaternary International 259: 59–71. [Google Scholar]
- d'Errico F, Vanhaeren M. 2002. Criteria for identifying red deer (Cervus elaphus) age and sex from their canines. Application to the study of Upper Palaeolithic and Mesolithic ornaments. Journal of Archaeological Science 29: 211–232. [Google Scholar]
- Dabney J, Knapp M, Glocke I et al. 2013. Complete mitochondrial genome sequence of a Middle Pleistocene cave bear reconstructed from ultrashort DNA fragments. Proceedings of the National Academy of Sciences of the United States of America 110: 15758–15763. [PubMed: 24019490]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dalla Valle C. 2008. I micromammiferi dei livelli musteriani e romanelliani della Grotta del Cavallo (Lecce). Thesis, Università “Cà Foscari” di Venezia.
- De La Peña P. 2015. A qualitative guide to recognize bipolar knapping for flint and quartz. Lithic Technology 40: 316–331. [Google Scholar]
- De Stefani M, Dini M, Klempererova H et al. 2012. Continuity and replacement in flake production across the Middle‐Upper Palaeolithic transition: a view over the Italian Peninsula. Palaeolithic Eur Mettmann 135–151. [Google Scholar]
- Douka K, Higham TFG, Wood R et al. 2014. On the chronology of the Uluzzian. Journal of Human Evolution 68: 1–13. [DOI] [PubMed] [Google Scholar]
- Duke H, Pargeter J. 2015. Weaving simple solutions to complex problems: an experimental study of skill in bipolar cobble‐splitting. Lithic Technology 40: 349–365. [Google Scholar]
- Falcucci A. 2018. Towards a renewed definition of the Protoaurignacian. Mitteilungen der Gesellschaft für Urgeschichte 27: 87–130. [Google Scholar]
- Fernández‐Jalvo Y, Andrews P. 2016b. Atlas of Taphonomic Identifications: 1001 + Images of Fossil and Recent Mammal Bone Modification. Springer: Dordrecht. [Google Scholar]
- Fernández‐Jalvo Y, Andrews P, Denys C et al. 2016a. Taphonomy for taxonomists: implications of predation in small mammal studies. Quaternary Science Reviews 139: 138–157. [Google Scholar]
- Fiore I, Tagliacozzo A. 2006. Lo sfruttamento dello stambecco nel Tardiglaciale di Riparo Dalmeri (TN): il livello 26c. Archaeozoological Studies in Honour of Alfredo Riedel 59–76. [Google Scholar]
- Fiorini A, Curci A, Benazzi S et al. 2018. Il sistema di documentazione digitale dello scavo archeologico nel sito di Uluzzo C (Nardò, LE). Sezione di Museologia Scientifica e Naturalistica 13: 68–70. [Google Scholar]
- Fiorini A, Curci A, Spinapolice EE et al. 2019. Grotta di Uluzzo C (Nardò‐Lecce): risultati preliminari, strumenti e metodi dell'indagine archeologica. Fasti Online Documents and Research 440: 1–18. [Google Scholar]
- Fisher JW. 1995. Bone surface modifications in zooarchaeology. Journal of Archaeological Method and Theory 2: 7–68. [Google Scholar]
- Fu Q, Meyer M, Gao X et al. 2013. DNA analysis of an early modern human from Tianyuan Cave, China. Proceedings of the National Academy of Sciences of the United States of America 110: 2223–2227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galán‐García J. 2019. Estudio de los Quirópteros (Chiroptera, Mammalia) del Cuaternario de Aragón y el norte de la Península Ibérica PhD Thesis, Universidad de Zaragoza, Zaragoza.
- Gambassini P. Electa, 1997. Il Paleolitico di Castelcivita: culture e Ambiente.
- Gansauge MT, Aximu‐Petri A, Nagel S et al. 2020. Manual and automated preparation of single‐stranded DNA libraries for the sequencing of DNA from ancient biological remains and other sources of highly degraded DNA. Nature Protocols 15: 2279–2300. [DOI] [PubMed] [Google Scholar]
- Gansauge MT, Gerber T, Glocke I et al. 2017. Single‐stranded DNA library preparation from highly degraded DNA using T4 DNA ligase. Nucleic Acids Research 45: e79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gansauge MT, Meyer M. 2013. Single‐stranded DNA library preparation for the sequencing of ancient or damaged DNA. Nature Protocols 8: 737–748. [DOI] [PubMed] [Google Scholar]
- Geneste JM. 1991. Systèmes techniques de production lithique: variations techno – économiques dans les processus de réalisation des outillages paléolithiques. Techniques and Culture 17–18: 3–35. [Google Scholar]
- Glocke I, Meyer M. 2017. Extending the spectrum of DNA sequences retrieved from ancient bones and teeth. Genome Research 27: 1230–1237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grayson DK. 1984. Quantitative Zooarchaeology: Topics in the Quantification of Archaeofaunas. Academic Press: Orlando. [Google Scholar]
- Guyodo JN, Marchand G. 2005. The bipolar percussion on an anvil in Western France from the end of the Paleolithic up to the Chalcolithic: an economic and social lecture. Bulletin de la Société Préhistorique Française 102: 539–549. [Google Scholar]
- Higham T, Douka K, Wood R et al. 2014. The timing and spatiotemporal patterning of Neanderthal disappearance. Nature 512: 306–309. [DOI] [PubMed] [Google Scholar]
- Hill EA, Hunt CO, Lucarini G et al. 2015. Land gastropod piercing during the Late Pleistocene and Early Holocene in the Haua Fteah. Libya. Journal of Archaeological Science: Reports 4: 320–325. [Google Scholar]
- Hunt CO, Reynolds TG, El‐Rishi HA et al. 2011. Resource pressure and environmental change on the North African littoral: Epipalaeolithic to Roman gastropods from Cyrenaica, Libya. Quaternary International 244: 15–26. [Google Scholar]
- Huson DH, Auch AF, Qi J et al. 2007. MEGAN analysis of metagenomic data. Genome Research 17: 377–386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inizan ML, Roche H, Tixier J et al. 1999. Technology and terminology of knapped stone. Crep: Nanterre.
- Janossy D. 1983. Humeri of central European smaller Passeriformes. Fragmenta Mineralogica et Palaeontologica 11: 85–112. [Google Scholar]
- Keeley LH editor. 1980. Experimental Determination of Stone Tool Uses: a Microwear Analysis. University of Chicago Press: London. [Google Scholar]
- Kircher M, Sawyer S, Meyer M. 2012. Double indexing overcomes inaccuracies in multiplex sequencing on the Illumina platform. Nucleic Acids Research 40: e3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laplace G. 1966. Recherches sur l'origine et l'evolution des complexes leptolithiques. Collection de l'École française de Rome 4.
- Lyman RL. 1994. Vertebrate Taphonomy. Cambridge University Press: Cambridge. [Google Scholar]
- Marciani G, Ronchitelli A, Arrighi S et al. 2020a. Lithic techno‐complexes in Italy from 50 to 39 thousand years BP: an overview of lithic technological changes across the Middle‐Upper Palaeolithic boundary. Quaternary International 551: 123–149. [Google Scholar]
- Marciani G, Spagnolo V, Martini I et al. 2020b. Neanderthal occupation during the tephra fall‐out: technical and hunting behaviours, sedimentology and settlement patterns in SU 14 of Oscurusciuto rock shelter (Ginosa, southern Italy). Archaeological and Anthropological Sciences 12. [Google Scholar]
- Maricic T, Whitten M, Pääbo S. 2010. Multiplexed DNA sequence capture of mitochondrial genomes using PCR products. PLoS ONE 5 e14004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mariezcurrena K, Altuna J. 1983. Contribución al conocimiento del desarollo de la dentición y el esqueleto poscraneal de cervus elaphus. Sociedad de Ciencias Aranzadi.
- Milano S, Prendergast AL, Schöne BR. 2016. Effects of cooking on mollusk shell structure and chemistry: implications for archeology and paleoenvironmental reconstruction. Journal of Archaeological Science: Reports 7: 14–26. [Google Scholar]
- Moroni A, Boscato P, Ronchitelli A. 2013. What roots for the Uluzzian? Modern behaviour in Central‐Southern Italy and hypotheses on AMH dispersal routes. Quaternary International 1316: 27–44. [Google Scholar]
- Moroni A, Ronchitelli A, Arrighi S et al. 2018. Grotta del cavallo [Apulia–Southern Italy]. the uluzzian in the mirror. Journal of Anthropological Sciences 96: 125–160. [DOI] [PubMed] [Google Scholar]
- Negrino F, Riel‐Salvatore J. 2018. From Neanderthals to Anatomically Modern Humans in Liguria (Italy): the current state of knowledge. In: Palaeolihic Italy. Advanced studies on early human adaptations in the Apennine peninsula; 161–181.
- Odell GH. 1981. The mechanics of use‐breakage of stone tools: some testable hypotheses. Journal of Field Archaeology 8: 197–209. [Google Scholar]
- Odell GH, Odell‐Vereecken F. 1980. Verifying the reliability of lithic use‐wear assessments by ‘blind tests’: the low‐power approach. Journal of Field Archaeology 7: 87–120. [Google Scholar]
- Palma di Cesnola A. 1964. Prima campagna di scavi nella Grotta del Cavallo presso Santa Caterina (Lecce) – Dialnet. Rivista di Scienze Preistoriche.
- Palma di Cesnola A. 1993. Il Paleolitico superiore in Italia. Introduzione allo studio.
- Palma di Cesnola A. 2004. Paglicci. L'Aurignaziano e il Gravettiano antico, Grenzi C (ed.). Foggia.
- Peresani M, Bertola S, Delpiano D et al. 2019. The Uluzzian in the north of Italy: insights around the new evidence at Riparo Broion. Archaeological and Anthropological Sciences 11: 3503–3536. [Google Scholar]
- Peresani M, Cristiani E, Romandini M. 2016. The Uluzzian technology of Grotta di Fumane and its implication for reconstructing cultural dynamics in the Middle‐Upper Palaeolithic transition of western Eurasia. Journal of Human Evolution 91: 36–56. [DOI] [PubMed] [Google Scholar]
- Plisson H. 1985. Etude fonctionnelle d'outillages lithiques préhistoriques par l'analyse des micro‐usures: recherche méthodologique et archéologique. Université de Paris I.
- Potts R, Shipman P. 1981. Cutmarks made by stone tools on bones from Olduvai Gorge, Tanzania. Nature 291: 577–580. [Google Scholar]
- Ramos‐Muñoz J, Cantillo‐Duarte JJ, Bernal‐Casasola D et al. 2016. Early use of marine resources by Middle/Upper Pleistocene human societies: the case of Benzú rockshelter (northern Africa). Quaternary International 407: 6–15. [Google Scholar]
- Ranaldo F, Boscato P, Moroni A et al. 2017. Riparo dell'Oscurusciuto (Ginosa – TA): la chiusura del ciclo Levallois alla fine del Paleolitico medio. In Preistoria e Protostoria Della Puglia, Radina F (ed.). Firenze; 169–174.
- Riel‐Salvatore J. 2009. What Is a ‘Transitional' Industry? The Uluzzian of Southern Italy as a Case Study, in: Sourcebook of Paleolithic Transitions. Springer: New York; 377–396. [Google Scholar]
- Riel‐Salvatore J. 2010. A niche construction perspective on the middle‐Upper Paleolithic transition in Italy. Journal of Archaeological Method and Theory 17: 323–355. [Google Scholar]
- Rohland N, Glocke I, Aximu‐Petri A et al. 2018. Extraction of highly degraded DNA from ancient bones, teeth and sediments for high‐throughput sequencing. Nature Protocols 13: 2447–2461. [DOI] [PubMed] [Google Scholar]
- Romandini M, Crezzini J, Bortolini E et al. 2020. Macromammal and bird assemblages across the late Middle to Upper Palaeolithic transition in Italy: an extended zooarchaeological review. Quaternary International 551: 188–223. [Google Scholar]
- Ronchitelli A, Moroni A, Boscato P et al. 2018. The Uluzzian 50 years later. In: Multas per Gentes et Multa per Saecula, Valde‐Nowak KSP (ed.), (a cura di). Alter Radosław Palonka (Publisher): Krakow; 71–76.
- Sano K, Arrighi S, Stani C et al. 2019. The earliest evidence for mechanically delivered projectile weapons in Europe. Nature Ecology and Evolution 3: 1409–1414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shipman P. 1981. Life History of a Fossil and Introduction to Taphonomy and Paleoecology. Harvard University Press, Harvard.
- Shipman P, Rose JJ. 1984. Cutmark mimics on modern and fossil bovid bones. Current Anthropology 25: 116–117. [Google Scholar]
- Slon V, Glocke I, Barkai R et al. 2016. Mammalian mitochondrial capture, a tool for rapid screening of DNA preservation in faunal and undiagnostic remains, and its application to Middle Pleistocene specimens from Qesem Cave (Israel). Quaternary International 398: 210–218. [Google Scholar]
- Slon V, Hopfe C, Weiß CL et al. 2017. Neandertal and Denisovan DNA from Pleistocene sediments. Science 356: 605–608. [DOI] [PubMed] [Google Scholar]
- Soriano S, Robert A, Huysecom É. 2010. Percussion bipolaire sur enclume: choix ou contrainte? L'exemple du Paléolithique d'Ounjougou (Pays dogon, Mali). Paléo (spécial) 123–132. [Google Scholar]
- Spagnolo V, Crezzini J, Marciani G et al. 2020. Neandertal camps and hyena dens. Living floor 150A at Grotta dei Santi (Monte Argentario, Tuscany, Italy). Journal of Archaeological Science: Reports 30.
- Spinapolice EE. 2012. Raw material economy in Salento (Apulia, Italy): new perspectives on Neanderthal mobility patterns. Journal of Archaeological Science 39: 680–689. [Google Scholar]
- Spinapolice EE. 2018. Les Néandertaliens du talon Technologie lithique et mobilité au Paléolithique moyen dans le Salento (Pouilles, Italie méridionale). Archaeopress: Oxford. [Google Scholar]
- Spinapolice EE, Borgia V, Cristiani E. 2018. Neanderthal Mobility Pattern and Technological Organization in the Salento (Apulia, Italy). Palaeolithic Italy Advanced Studies on Early Human Adaptations in the Apennine Peninsula. Sidestone Press Acadmics: Leiden; 95–124. [Google Scholar]
- Stiner MC. 1999. Palaeolithic mollusc exploitation at Riparo Mochi (Balzi Rossi, Italy): food and ornaments from the Aurignacian through Epigravettian. Antiquity 73: 735–754. [Google Scholar]
- Stiner MC. 2009. Prey choice, site occupation intensity & economic diversity in the Middle – early Upper Palaeolithic at the Üçağizli Caves, Turkey. Before Farming 3: 1–20. [Google Scholar]
- Stiner MC, Kuhn SL, Weiner S et al. 1995. Differential burning, recrystallization, and fragmentation of archaeological bone. Journal of Archaeological Science 22: 223–237. [Google Scholar]
- Strohalm M, Kavan D, Novák P et al. 2010. mMass 3: a cross‐platform software environment for precise analysis of mass spectrometric data. Analytical Chemistry 82: 4648–4651. [DOI] [PubMed] [Google Scholar]
- Tomek T, Bochenski ZM. 2000. The Comparative Osteology of European Corvids (Aves: Corvidae), with a Key to the Identification of Their Skeletal Elements. Institute of Systematics and Evolution of Animals, Polish Academy of Sciences: Krakow. [Google Scholar]
- Tringham R, Cooper G, Odell G et al. 1974. Experimentation in the formation of edge damage: a new approach to lithic analysis. Journal of Field Archaeology 1: 171–196. [Google Scholar]
- Van Doorn NL, Hollund H, Collins MJ. 2011. A novel and non‐destructive approach for ZooMS analysis: ammonium bicarbonate buffer extraction. Archaeological and Anthropological Sciences 3: 281–289. [Google Scholar]
- Van Gijn A. 2010. Chert in Focus: Lithic Biographies in the Neolithic and Bronze Age. Sidestone Press: Leiden. [Google Scholar]
- Vigal CR, Machordom A. 1985. Tooth eruption and replacement in the Spanish wild goat. Acta theriologica 30: 305–320. plates 4. [Google Scholar]
- Villa P, Pollarolo L, Conforti J et al. 2018. From Neandertals to modern humans: new data on the Uluzzian. PLoS ONE 13 e0196786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welker F, Hajdinjak M, Talamo S et al. 2016. Palaeoproteomic evidence identifies archaic hominins associated with the Châtelperronian at the Grotte du Renne. Proceedings of the National Academy of Sciences of the United States of America 113: 11162–11167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zanchetta G, Giaccio B, Bini M et al. 2018. Tephrostratigraphy of Grotta del Cavallo, Southern Italy: insights on the chronology of Middle to Upper Palaeolithic transition in the Mediterranean. Quaternary Science Reviews 182: 65–77. [Google Scholar]
- Zilhão J, Angelucci DE, Badal‐García E et al. 2010. Symbolic use of marine shells and mineral pigments by Iberian Neandertals. Proceedings of the National Academy of Sciences of the United States of America 107: 1023–1028. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
This article includes online‐only Supplemental Data.
Table S1. Lithic items in each layer of Borzatti excavation, according to Borzatti (1965) and items currently stored at Museo della Preistoria di Nardò MPN.
Table S2. Lithic items in SUs 3, 15 and 17 (excavation 2016–2018).
Table S3. Sequencing results for all tested sediment samples and relevant negative controls. For the mammalian mitochondrial capture, results are only shown for Hominidae and for the two taxa identified as ancient in at least one of the samples. L – length; MQ – mapping quality; C – cytosine; T – thymine; CI – confidence interval.
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding author. The data are not publicly available due to privacy or ethical restrictions.