

Abstract
A theoretical analysis of the detachment of bacteria adhering to substratum surfaces upon the passage of an air-liquid interface is given, together with experimental results for bacterial detachment in the absence and presence of a conditioning film on different substratum surfaces. Bacteria (Streptococcus sobrinus HG1025, Streptococcus oralis J22, Actinomyces naeslundii T14V-J1, Bacteroides fragilis 793E, and Pseudomonas aeruginosa 974K) were first allowed to adhere to hydrophilic glass and hydrophobic dimethyldichlorosilane (DDS)-coated glass in a parallel-plate flow chamber until a density of 4 × 106 cells cm−2 was reached. For S. sobrinus HG1025, S. oralis J22, and A. naeslundii T14V-J1, the conditioning film consisted of adsorbed salivary components, while for B. fragilis 793E and P. aeruginosa 974K, the film consisted of adsorbed human plasma components. Subsequently, air bubbles were passed through the flow chamber and the bacterial detachment percentages were measured. For some experimental conditions, like with P. aeruginosa 974K adhering to DDS-coated glass and an air bubble moving at high velocity (i.e., 13.6 mm s−1), no bacteria detached upon passage of an air-liquid interface, while for others, detachment percentages between 80 and 90% were observed. The detachment percentage increased when the velocity of the passing air bubble decreased, regardless of the bacterial strain and substratum surface hydrophobicity involved. However, the variation in percentages of detachment by a passing air bubble depended greatly upon the strain and substratum surface involved. At low air bubble velocities the hydrophobicity of the substratum had no influence on the detachment, but at high air bubble velocities all bacterial strains were more efficiently detached from hydrophilic glass substrata. Furthermore, the presence of a conditioning film could either inhibit or stimulate detachment. The shape of the bacterial cell played a major role in detachment at high air bubble velocities, and spherical strains (i.e., streptococci) detached more efficiently than rod-shaped organisms. The present results demonstrate that methodologies to study bacterial adhesion which include contact with a moving air-liquid interface (i.e., rinsing and dipping) yield detachment of an unpredictable number of adhering microorganisms. Hence, results of studies based on such methodologies should be referred as “bacterial retention” rather than “bacterial adhesion”.
Bacterial adhesion forms the basis for several diverse problems in medicine (5, 10, 21, 38), industry (3, 15, 29, 32), and environmental areas (8, 33). The most detrimental effects are encountered in medicine, where failure of implanted devices may result from surface-associated bacterial infections. Different environmental, biological, and substratum-related factors have been proposed to influence bacterial adhesion to surfaces at the onset of the formation of a mature biofilm. Bacterial adhesion in natural and industrial environments is preceded by the formation of conditioning films, consisting of adsorbed organic components. These films may inhibit or promote bacterial adhesion (35, 37), and the final properties of the conditioning film have been suggested to be under the control of the physicochemical properties of the underlying substratum surface, such as its hydrophobicity and charge (40).
Adhesion of microorganisms to substratum surfaces is difficult to measure, and a variety of static and dynamic (25, 36) systems have been developed by different research groups, currently often with the aid of image analysis. Sometimes mass transport considerations are included in the design (9, 39), but frequently no distinction is made between kinetic and stationary or equilibrium effects (36). The lack of a ubiquitously accepted methodology and way of reporting bacterial adhesion data impedes a comparison of results from different laboratories and is unlike the standardization seen in many other fields of science.
Contributing to this unfortunate situation are the so-called “slight rinsing” and “dipping” procedural artifacts to remove loosely adhering bacteria. These experimental procedures are still used quite often by many groups (16, 30, 44) but neglect observations made over the past decade that a passing air-liquid interface has the ability to displace and detach micron-sized particles, including dust particles (28) and adhering bacteria (34), from surfaces. By consequence, upon close inspection, many papers on bacterial adhesion do not deal with adhesion but rather with the ability of adhering bacteria to withstand detachment, i.e., bacterial retention. Bos et al. (1) recently demonstrated, using micropatterned surfaces, that bacterial adhesion is less influenced by substratum surface hydrophobicity than by bacterial retention. In fact, for many natural processes bacterial retention has been described to be more influential on the final development of a biofilm than adhesion. In the oral cavity, for instance, adhering bacteria have to withstand detachment forces due to eating, speaking, drinking, and swallowing in order to form a biofilm (6). On the eye and on contact lenses, blinking of the eye lid creates a detachment force on adhering bacteria (27), similar to the action of waves on rocks and ship hulls in the marine environment (8, 12). Adhering organisms, responsible for biodeterioration of monumental buildings, have to adhere strongly enough to withstand rain (41).
Several processes occur in rapid sequence when an air-liquid interface passes over an adhering micron-sized particle. These processes have only recently been fully understood and need to be introduced into microbiology in order to stimulate methodological advances in the field. Detachment of adhering polystyrene particles by the passage of air bubbles has been demonstrated to be more efficient when (i) the velocity of the passing air bubble decreases (19, 20), (ii) the air-liquid interfacial tension increases (19, 20), (iii) particle size decreases (17), (iv) the substratum surface is more hydrophobic (19), and (v) the substratum surface is negatively charged (18).
The aim of the present paper is to provide a theoretical background of the processes occurring during the passage of an air-liquid interface over bacteria adhering to a substratum surface and to report on the air bubble-induced detachment of different bacterial strains from hydrophilic and hydrophobic substratum surfaces in the absence and presence of a conditioning film.
THEORETICAL BACKGROUND
Detachment of an adhering bacterium from substratum surfaces by a passing air bubble is controlled by a sequence of processes that includes the approach of the air bubble to the adhering bacterium; interception of the bacterium by the air bubble; deformation of the air-liquid interface, yielding a thin liquid film separating the bacterium and the air bubble; formation of a three-phase contact between an adhering bacterium, air, and liquid; and subsequent transport of the bacterium-bubble aggregate away from the substratum surface (19). Hence, the efficiency of the bacterial detachment (E) by a passing air-liquid interface can be expressed as E = EC × EA × ES, where EC is the air bubble-bacterium collision efficiency, EA is the bubble-bacterium attachment efficiency, and ES is the stability of the bubble-bacterium aggregate.
The direction and velocity of the air bubble with respect to the adhering bacterium determine the collision efficiency (EC) (31). In a parallel-plate flow chamber, for an air bubble fully spanning the width of the chamber, the collision efficiency is 1. However, an air bubble moving in a narrow channel is surrounded by a liquid film (4), the thickness of which increases with increasing air bubble velocity. As this liquid film thickness decreases, the collision efficiency increases, as outlined in Fig. 1.
FIG. 1.

Schematic presentation of the different degrees of immersion of a bacterium upon the passage of an air bubble. The air-liquid interface velocity decreased from panel a to panel c, yielding to different degrees of immersion. d represents the distance between the bacterium and collector surface, φ is the angle giving the position of the particle with respect to the air-liquid interface, θ is the contact angle between the surface of the bacterium and the air-liquid interface, b is the liquid film thickness, γlv is the air-liquid interfacial tension, and Fγ is the surface tension result of the air-liquid interfacial tension exerted by the bend meniscus on the bacterium.
The interaction between a passing air bubble and an adhering bacterium in an aqueous medium includes, apart from hydrodynamic forces, Lifshitz-van der Waals, electrostatic, and hydrophobic (acid-base) forces (22). Hence, the bubble-bacterium attachment efficiency will increase with bacterial cell surface hydrophobicity (14). Note that an effective three-phase contact between the bubble and an adhering bacterium (Fig. 1) can form only when the bacterium-bubble contact time is long enough to thin the liquid film and form a three-phase boundary (23). Hence, the bubble-bacterium attachment efficiency will also increase with decreasing air bubble velocity. During thinning of the liquid film, a viscous force opposes the attachment of the bacteria to the air bubble. This viscous drag force increases with the size of the bacterium and the air bubble velocity (17).
Once and only if an effective three-phase contact between the bubble and an adhering bacterium has formed, a balance between the surface tension force (Fγ), the bacterial adhesion force (FA), and the viscous drag force determines whether detachment will occur. Evidently (Fig. 1), the surface tension force opposes the adhesion and viscous drag forces depending on the position of the three-phase contact and can be expressed by the equation 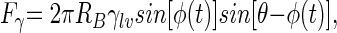 in which RB is the radius of the bacterium, γlv is the air-liquid interfacial tension, φ represents the angle determining the position of the bacterium with respect to the air-liquid interface, t is time (in seconds), and θ is the contact angle between the surface of the bacterium and the air-liquid interface. Figure 2 presents the surface tension detachment force as a function of φ for different bacterial contact angles or cell surface hydrophobicities, θ. As can be seen, for each bacterial cell surface hydrophobicity, the surface tension detachment force passes through a maximum value at a different degree of immersion of the adhering bacteria (see also Fig. 1).
in which RB is the radius of the bacterium, γlv is the air-liquid interfacial tension, φ represents the angle determining the position of the bacterium with respect to the air-liquid interface, t is time (in seconds), and θ is the contact angle between the surface of the bacterium and the air-liquid interface. Figure 2 presents the surface tension detachment force as a function of φ for different bacterial contact angles or cell surface hydrophobicities, θ. As can be seen, for each bacterial cell surface hydrophobicity, the surface tension detachment force passes through a maximum value at a different degree of immersion of the adhering bacteria (see also Fig. 1).
FIG. 2.

Theoretically calculated surface tension detachment forces from the equation 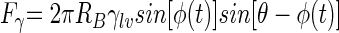 for different values of φ and θ and for a bacterium with an RB of 800 nm and a γlv of 72 mJ m−2. Positive forces are directed away from the substratum surface.
for different values of φ and θ and for a bacterium with an RB of 800 nm and a γlv of 72 mJ m−2. Positive forces are directed away from the substratum surface.
The adhesion force can be calculated theoretically according to the extended DLVO (Derjaguin-Landau-Verney-Overbeek) approach as a combination of Lifshitz-van der Waals, electrostatic, and acid-base forces (42) as follows: 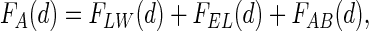 where FLW(d) and FEL(d) denote the decay with the distance (d) to the collector surface of the Lifshitz-van der Waals and electrostatic interaction forces, respectively, and FAB denotes acid-base interaction forces, but theoretical adhesion forces that match experimentally reported adhesion forces have not yet been calculated. The most reliable estimates of bacterial adhesion forces have become available from atomic-force microscopy and range from 7 × 10−9 to 4 × 10−9 N between sulfate-reducing bacteria and Si3N4 or mica surfaces (13). Such adhesion forces are orders of magnitude smaller than the surface tension detachment forces induced by a passing air-liquid interface, ranging up to 2 × 10−7 N, as can be seen in Fig. 2.
where FLW(d) and FEL(d) denote the decay with the distance (d) to the collector surface of the Lifshitz-van der Waals and electrostatic interaction forces, respectively, and FAB denotes acid-base interaction forces, but theoretical adhesion forces that match experimentally reported adhesion forces have not yet been calculated. The most reliable estimates of bacterial adhesion forces have become available from atomic-force microscopy and range from 7 × 10−9 to 4 × 10−9 N between sulfate-reducing bacteria and Si3N4 or mica surfaces (13). Such adhesion forces are orders of magnitude smaller than the surface tension detachment forces induced by a passing air-liquid interface, ranging up to 2 × 10−7 N, as can be seen in Fig. 2.
Once a bacterium is detached from a substratum surface, the balance between the surface tension forces acting on the bacterium as opposed to external forces in the flow (i.e., gravitational force, buoyancy force, hydrostatic pressure, and viscous drag force opposing the separation of the bacteria from the collector surfaces) determines the bacterium-bubble aggregate stability (38). For the dimensions considered in the parallel-plate flow chamber (i.e., d/RB ≪ 1 and RB ≪ 10 μm), the surface tension force dominates (28).
MATERIALS AND METHODS
Microorganisms and culture conditions.
Five bacterial strains were used in this study. S. oralis J22 was cultured aerobically at 37°C in Todd-Hewitt broth (Oxoid, Basingstoke, United Kingdom), while S. sobrinus HG1025 and P. aeruginosa 974K were cultured under similar conditions in tryptone soya broth (Oxoid). A. naeslundii T14V-J1 was cultured in Schaedler's broth supplemented with 0.01 g of hemin liter−1 and B. fragilis 793E was cultured in chopped-meat medium (in one liter: 0.30% beef extract [Difco Laboratories, Detroit, Mich.], 30 g of peptone, 5 g of yeast extract, 5 g of K2HPO4, and 0.56 g of l-cysteine HCl · H2O [pH 7.0]) both in an anaerobic cabinet (Don Whitley Scientific, West Yorkshire, United Kingdom) and in an atmosphere of 10% H2, 85% N2, and 5% CO2 at 37°C. For each experiment, strains were inoculated from blood agar plates in batch culture for 24 h. This culture was used to inoculate a second culture, which was grown for 16 h prior to harvesting.
Bacteria were harvested in stationary phase by centrifugation (5 min at 5,500 × g) and washed twice with demineralized water, and to break bacterial chains and aggregates, cells were sonicated for 30 s at 30 W (Vibra Cell model 375; Sonics and Materials, Inc., Danbury, Conn.). Sonication was performed intermittently while cells were cooling in an ice-water bath. Finally, bacteria were suspended in a buffer (1 mM calcium chloride, 2 mM potassium phosphate, and 50 mM potassium chloride [pH 6.8]) to a concentration of 6 × 108 ml−1 for adhesion experiments.
Substratum surfaces and conditioning films.
Glass microscope slides (76 by 25 by 1.5 mm) were used as substratum surfaces. Glass surfaces were cleaned with 2% RBS 35 detergent (Omnilabo International B.V., Breda, The Netherlands) and alternately rinsed in water, methanol, water again, and finally Millipore-Q water. Cleaned glass plates were used as a negatively charged, hydrophilic substratum surface, while hydrophobic, negatively charged surfaces were prepared from cleaned glass first by drying the plates in an oven at 80°C and then by silanization in 0.05% (wt/vol) dimethyldichlorosilane (DDS) in trichloroethylene. After silanization for 15 min, the surfaces were rinsed with trichloroethylene, methanol, and Millipore-Q water.
Conditioning films were formed on the substratum surfaces by exposure of the surfaces to saliva (for S. sobrinus HG1025, S. oralis J22, and A. naeslundii T14V-J1), or human plasma (for P. aeruginosa 974K and B. fragilis 793E). Whole saliva samples from approximately 20 healthy human volunteers of both sexes were collected in ice-chilled beakers after stimulation by chewing Parafilm (American National Can, Neenah, Wis.). Thereafter, the saliva was pooled and centrifuged at 10,000 × g for 5 min at 4°C, while phenylmethylsulfonyl fluoride (0.1 M; Merck, Darmstadt, Germany) was added to a final concentration of 1 mM as a protease inhibitor. The solution was again centrifuged, dialyzed (molecular weight cutoff, 6,000 to 8,000; Spectra/por; Spectrum Medical Industries, Inc., Los Angeles, Calif.) overnight at 4°C against demineralized water, and freeze-dried for storage. A 10% dilution of pooled human plasma anticoagulated with sodium citrate and 1.5 g of freeze-dried saliva reconstituted in adhesion buffer liter−1 were used for conditioning-film formation on the substrata.
Surface characterization.
Water contact angles on lawns of bacteria were determined using the sessile drop technique (7, 42). Briefly, bacteria were layered from Millipore-Q water onto 0.45-μm-pore-sized filters (Millipore) using negative pressure. The filters were left to air dry in ambient air until so-called plateau contact angles could be measured. Contact angles of the substratum surfaces were measured directly on the substratum surfaces, prepared as described above.
Electrophoretic mobilities of the bacteria in adhesion buffer were measured with a Lazer Zee Meter 501 (PenKem, Bedford Hills, N.Y.), which uses scattering of incident lazer light to enable detection of the bacteria at relatively low magnifications. Aliquots of the bacterial suspensions were added to a density of approximately 108 cells ml−1 in adhesion buffer. The voltage difference across the electrophoresis chamber was set to 150 V, and the electrophoretic mobilities measured were converted to apparent zeta potentials through the use of the Smoluchowski equation (24). All zeta potentials reported are the mean values of measurements on three different cultures. Zeta potentials of the glass or DDS-coated glass collector surfaces in the adhesion buffer were measured by streaming potentials in a parallel-plate flow chamber, as described by Van Wagenen and Andrade (43).
Parallel-plate flow chamber and image analysis.
The experiments were carried out in a parallel-plate flow chamber (20, 39) containing three basic modules: the flow loop, the syringe pump, and the image analysis components. The chamber is made of poly(methyl methacrylate) with channel dimensions (76 by 5 by 0.6 mm, length by width by height) enabling control of the passage and velocity of the air bubbles introduced. The top plate is made of glass and separated from the bottom plate by two Teflon spacers. The bottom plate is made of poly(methyl methacrylate) with a rectangular section cut out in the center (76 by 25 by 1.5 mm) in which the substrata could be inserted. A pulse-free liquid flow was achieved by hydrostatic pressure, and a roller pump recirculated the liquid.
For enumeration of the number of bacteria adhering to the bottom plate before and after the passage of an air bubble, the entire flow chamber was mounted on the stage of a phase-contrast microscope (Olympus BH-2) equipped with a 40× ultralong working-distance objective (Olympus ULWD-CD Plan 40 PL). A charge-coupled device camera (model CCD-MXR/5010; Adimec Advanced Image Systems B.V., Eindhoven, The Netherlands) was mounted on the phase-contrast microscope and connected to a PCVision+ video frame grabber interface card (Imaging Technology Incorporated, Woburn, Mass.) to digitize the video signal. A cell finder slide (76 by 26 mm) attached to the outside of the bottom plate was focused in order to obtain an accurate repositioning (±2 μm) of the flow chamber in the enumeration process.
Experimental protocol.
All experiments were performed at room temperature in triplicate with separate bacterial suspensions and collector surfaces. Before each experiment, all tubes, the flow chamber, and the syringe pump were filled with buffer and care was taken to remove all air bubbles from the system. The buffer solution was first perfused through the chamber for 30 min. At this stage and when appropriate, the flow was switched to adhesion buffer supplemented with 1.5 g of lyophilized saliva liter−1 or with 10% pooled plasma for 2 h to create a conditioning film, after which the flow was switched for 20 min to buffer to remove all remnants of saliva or plasma from the tubing and the flow chamber. Thereafter, bacteria were allowed to adhere until a density of 4 × 106 cm−2 was reached, as enumerated on 10 arbitrarily chosen areas of 0.017 mm2 distributed over the length of the substratum surface. The volumetric flow was 0.025 ml s−1 (wall shear rate, 10 s−1), which yields a laminar flow (Reynolds number 0.6). Finally, the flow was switched again for 30 min to the buffer solution in order to remove all nonadhering bacteria from the flow chamber and the tubing system.
In order to introduce air bubbles in the flow system, a syringe pump (model SC512; Terumo Tokyo, Japan) filled with the buffer solution was connected to the flow chamber by a three-way valve to control the velocity of the air bubble. Air bubbles (0.1 ± 0.02 ml) were introduced through injection in the tubing by a 1-ml syringe. The approximate projected dimensions of the air bubbles were 25 by 5 mm (length by width), i.e., they fully spanned the width of the flow chamber. Subsequently, the bacteria that remained adhering were enumerated, after the positions used before were retrieved with the aid of the cell finder.
Data analysis.
Detachment data were analyzed by a one-way analysis of variance. Analysis of variance tests the ratios of the mean squares between groups and within groups, and provides an F ratio (i.e., Fisher distribution) and its corresponding P value. If the null hypothesis is true, the F ratio should be approximately 1 since both the mean squares between and within groups should be about the same. If the ratio is much larger than 1, the null hypothesis is false. The shape of the F distribution depends on the degrees of freedom within and between groups. The lower the degree of freedom, the larger the value of F needed to be significant. A P value of less than 5% was considered to indicate a significant difference between groups. The subscript of F indicates the value of F needed for a hypothesis to be significant at the 5% level. When differences among the means were determined, post hoc Dunnett's range tests and pairwise multiple comparisons were employed to establish differences between means.
RESULTS
Table 1 presents the water contact angles on the different bacterial strains after drying to plateau state and on the substratum surfaces, together with their zeta potentials in adhesion buffer. A. naeslundii T14V-J1 was the most hydrophobic strain involved in this study, while S. oralis J22 was the most hydrophilic, as judged from their water contact angles. At this stage it must be noted, however, that the initial water contact angle on a bacterial lawn prior to drying was essentially 0, i.e., all bacterial strains are likely to be fully wettable during the passage of an air bubble in the parallel-plate flow chamber. The glass surface had a water contact angle of 33 degrees, while after the plate was coated with DDS, the water contact angle increased to 97 degrees. All bacterial and substratum surfaces were negatively charged, and zeta potentials ranged from −12 mV for A. naeslundii T14V-J1 to −26 mV for P. aeruginosa 974K.
TABLE 1.
Water contact angles on the different bacterial strains and substratum surfaces involved in this study, together with their zeta potentials in adhesion buffera
| Bacterial strain or substratum surface | θw (°) | ζ (mV) |
|---|---|---|
| Bacterial strain | ||
| S. sobrinus HG1025 | 29 | −14 |
| S. oralis J22 | 24 | −16 |
| A. naeslundii T14V-J1 | 64 | −12 |
| B. fragilis 793E | 35 | −17 |
| P. aeruginosa 974K | 43 | −26 |
| Substratum surface | ||
| Glass | 33 | −17 |
| DDS-coated glass | 97 | −11 |
Adhesion buffer contained 1 mM calcium chloride, 2 mM potassium phosphate, and 50 mM potassium chloride (pH 6.8). All data are the mean results of three experiments with separately grown bacterial strains and substratum surfaces, yielding an average standard deviation of around 5 degrees for the contact angles (θw) and 3 mV for the zeta potentials (ζ).
Table 2 shows the percentages of detachment of the different bacterial strains from hydrophilic glass and hydrophobic DDS-coated glass by a passing air bubble in the absence and presence of a conditioning film. Regardless of the bacterial strain involved and substratum surface hydrophobicity, the percentage of detachment was highest at the lowest velocity of the passing air bubble (F4.2 = 43.2, P < 0.000007). However, the percentage of detachment by a passing air bubble depended greatly upon the strain and substratum surface involved, as well as on the presence or absence of a conditioning film on the substratum surface. At the lowest air bubble velocity (i.e., 2.36 mm s−1), all bacterial strains presented similar detachment percentages when they adhered to hydrophilic glass substrata (F3.48 = 1.2, P = 0.37) but differed significantly when they adhered to hydrophobic DDS-coated substrata (F3.48 = 18.21, P = 0.0001). At the highest air bubble velocity (i.e., 13.6 mm s−1), all bacterial strains detached more easily from hydrophilic glass than from hydrophobic DDS-coated glass substrata (F4.20 = 22.0, P = 0.00006). Both streptococcal strains (S. oralis J22 and S. sobrinus HG1025) were more effectively detached than rod-shaped organisms (A. naeslundii T14V-J1, B. fragilis 793E, and P. aeruginosa 974K) when the air bubble moved at the highest air bubble velocity (F4.96 = 19.3, P = 0.0009).
TABLE 2.
Percentages of bacterial detachment by a passing air bubble moving at different velocities from glass and DDS-coated glass in the presence and absence of a conditioning filmc
| Bacterial strain | Bacterial detachment in the absence of a conditioning film (%) by an air bubble moving at:
|
Bacterial detachment in the presence of a conditioning film (%) by an air bubble moving at:
|
||||||
|---|---|---|---|---|---|---|---|---|
| 2.36 mm s−1
|
13.6 mm s−1
|
2.36 mm s−1
|
13.6 mm s−1
|
|||||
| Glass | DDS-coated glass | Glass | DDS-coated glass | Glass | DDS-coated glass | Glass | DDS-coated glass | |
| S. sobrinus HG1025a | 52 ± 15 | 53 ± 13 | 39 ± 17 | 26 ± 18 | 80 ± 15 | 91 ± 5 | 41 ± 16 | 27 ± 11 |
| S. oralis J22a | 72 ± 15 | 52 ± 20 | 60 ± 15 | 46 ± 22 | 65 ± 27 | 79 ± 17 | 44 ± 21 | 24 ± 18 |
| A. naeslundii T14V-J1a | 68 ± 18 | 41 ± 22 | 23 ± 13 | 17 ± 10 | 67 ± 16 | 61 ± 16 | 29 ± 19 | 22 ± 6 |
| B. fragilis 793Eb | 57 ± 14 | 60 ± 21 | 34 ± 15 | 33 ± 19 | 58 ± 20 | 60 ± 10 | 29 ± 12 | 14 ± 9 |
| P. aeruginosa 974Kb | 58 ± 22 | 90 ± 7 | 28 ± 14 | 0 ± 76 | 72 ± 12 | 45 ± 19 | 39 ± 18 | 12 ± 18 |
Conditioning film consisted of saliva.
Conditioning film consisted of adsorbed plasma proteins.
Note that prior to the passage of an air bubble, the number of adhering bacteria was set at 4 × 106 cm−2, as enumerated by the image analysis system. All data are means ± standard deviations based on the results of three experiments with separately grown bacterial strains and substratum surfaces.
The presence of a conditioning film had various effects on detachment of adhering bacteria. While its presence did not affect the detachment of B. fragilis 793E, A. naeslundii T14V-J1, and P. aeruginosa 974K (F4.30 = 0.3, P = 0.64), it stimulated the detachment of the streptococcal strains at the lowest air bubble velocity regardless of the hydrophobicities of the substrata (F4.30 = 7.4, P = 0.012).
Redeposition of bacteria once they were detached or displacement of adhering bacteria by a passing air bubble was not observed, except with P. aeruginosa 974K after its detachment from hydrophobic DDS-coated glass in the absence of a conditioning film. While 90% of the P. aeruginosa 974K organisms were effectively detached without redeposition or displacement by air bubbles at a velocity of 2.36 mm s−1 (Fig. 3), no bacterial detachment was observed at an air bubble velocity of 13.6 mm s−1 (Fig. 4). Instead, areas on the substratum located near the sides of the flow chamber did not show any adhering bacteria after the passage of an air bubble (Fig. 4a) but areas near the center showed large bacterial agglomerates (Fig. 4b) from displacement and redeposition of adhering bacteria.
FIG. 3.

Example of the effect of a passing air bubble through the parallel-plate flow chamber with P. aeruginosa 974K adhering to DDS-coated glass in the absence of a conditioning film. Shown are bacteria prior to (a) and after (b) the passage of an air bubble moving at a velocity of 2.36 mm s−1. Bars, 20 μm.
FIG. 4.

Example of the effect of a passing air bubble through the parallel-plate flow chamber with P. aeruginosa 974K adhering to DDS-coated glass in the absence of a conditioning film. Shown are bacteria prior to (a) and after (b) the passage of an air bubble moving at a velocity of 13.6 mm s−1. Bars, 20 μm.
DISCUSSION
In this study a theoretical analysis of the detachment of bacteria adhering to substratum surfaces upon the passage of air-liquid interfaces is presented, together with experimental results for bacterial detachment in the absence and presence of a conditioning film on different substratum surfaces. Greater bacterial detachment was achieved by decreasing the air bubble velocity, regardless of the hydrophobicities of the substrata or bacterial strains or the presence of a conditioning film. These results are in line with previous observations on detachment of polystyrene particles from different substrata where a linear decrease in detachment was observed with an increasing air bubble velocity (17–20). Presumably, at high air bubble velocities, the bacterium-bubble attachment efficiency decreases due to a lower degree of immersion of the bacterium in the air-liquid interface (Fig. 1) and, hence, the detachment force 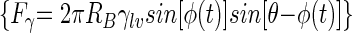 induced by a passing air-liquid interface decreases (19).
induced by a passing air-liquid interface decreases (19).
Nevertheless, bacterial cell and substratum hydrophobicity play a role in the detachment process. At low air bubble velocities, the detachment efficiencies were similar for all bacterial strains when they adhered to a hydrophilic substratum but differed when they adhered to a hydrophobic substratum. Presumably, bacterial surface hydrophobicity plays a more important role when bacteria adhere to a hydrophobic substratum, as the detachment of hydrophobic polystyrene particles (i.e., water contact angle of 90 degrees) stimulated by a passing air bubble was not influenced by substratum hydrophobicity at low air bubble velocities (19). At high air bubble velocities, strains detached more efficiently from a hydrophilic substratum surface. In this respect it must be realized that at low air bubble velocities, the detachment force is about 2 orders of magnitude greater than the bacterium-substratum adhesion force (Fig. 2) due to a high degree of immersion of the bacterium (Fig. 1c). Hence, substratum properties are not decisive in the detachment process. Conversely, at high velocities, the detachment force induced by the passing air bubble is lower (i.e., due to a lower degree of immersion [Fig. 1b]) and becomes comparable to the bacterium-substratum adhesion force, making substratum properties decisive in bacterial detachment.
The present study reveals two new aspects of air-liquid interface passage-stimulated detachment of bacteria which have not been observed for polystyrene particles. For P. aeruginosa 974K detaching from DDS-coated glass, the detachment efficiency was almost 0 and redeposition and displacement were seen. These findings likely indicate that a combination of lateral capillary forces between bacterial cells and adhesion forces counteracts the surface tension detachment force to form aggregates, as was also suggested for proteins and latex spheres (11, 26). In addition, the present study reveals an effect of particle shape on detachment, since it appears that coccal cells detach more readily upon the passage of an air-liquid interface than rod-shaped organisms, like P. aeruginosa 974K, B. fragilis 793E, and A. naeslundii T14V-J1.
Conditioning films have been described to promote or hinder bacterial adhesion (4, 37), and in this study conditioning films of adsorbed salivary components or plasma proteins are described to have various effects on detachment of different strains after the passage of an air-liquid interface. In an earlier inventory it was suggested that the presence of a conditioning film yielded higher detachment than when bacteria adhered to a bare substratum surface (2). However, in the present study the number of adhering bacteria prior to the passage of an air-liquid interface was set for all combinations of strains and substratum surfaces to 4 × 106 cm−2, whereas in an inventory by Bos et al. (2), the number of adhering bacteria varied strongly prior to detachment.
This study yields several important conclusions with respect to the study of bacterial adhesion to surfaces. First, since an unknown number of adhering bacteria is detached by passages of an air-liquid interface, as during slight rinsing and dipping, in situ enumeration methods should be preferred for the study of bacterial adhesion. In case in situ enumeration methods are not available, the terminology should be adjusted to emphasize that bacterial retention was measured rather than adhesion. In the latter case, a description of the detachment forces caused by slight rinsing and dipping that is as detailed as possible should be given. Second, it is important to realize that bacterial detachment by a passing air-liquid interface is smallest when the interface moves at a higher velocity, as this allows no time for a three-phase contact to form.
REFERENCES
- 1.Bos R, van der Mei H C, Gold J, Busscher H J. Retention of bacteria on a substratum surface with micro-patterned hydrophobicity. FEMS Microbiol Lett. 2000;189:311–315. doi: 10.1111/j.1574-6968.2000.tb09249.x. [DOI] [PubMed] [Google Scholar]
- 2.Bos R, van der Mei H C, Busscher H J. Physico-chemistry of initial microbial adhesive interactions—its mechanisms and methods for study. FEMS Microbiol Rev. 1999;23:179–230. doi: 10.1111/j.1574-6976.1999.tb00396.x. [DOI] [PubMed] [Google Scholar]
- 3.Bouman S, Lund D B, Driessen F M, Schmidt D G. Growth of thermoresistant streptococci and deposition of milk constituents on plates of heat exchangers during long operation times. J Food Prot. 1982;45:806–812. doi: 10.4315/0362-028X-45.9.806. [DOI] [PubMed] [Google Scholar]
- 4.Bretherton F P. The motion of long bubbles in tubes. J Fluid Mech. 1961;10:166–188. [Google Scholar]
- 5.Busscher H J, Geertsema-Doornbusch G I, van der Mei H C. Adhesion to silicone rubber of yeasts and bacteria isolated from voice prostheses: influence of salivary conditioning films. J Biomed Mat Res. 1997;34:201–210. doi: 10.1002/(sici)1097-4636(199702)34:2<201::aid-jbm9>3.0.co;2-u. [DOI] [PubMed] [Google Scholar]
- 6.Busscher H J, Cowan M M, van der Mei H C. On the relative importance of specific and non-specific approaches to oral microbial adhesion. FEMS Microbiol Lett. 1992;88:199–210. doi: 10.1111/j.1574-6968.1992.tb04988.x. [DOI] [PubMed] [Google Scholar]
- 7.Busscher H J, Weerkamp A H, van der Mei H C, van Pelt A W J, de Jong H P, Arends J. Measurements of the surface free energy of bacterial cell surfaces and its relevance for adhesion. Appl Environ Microbiol. 1984;48:980–983. doi: 10.1128/aem.48.5.980-983.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cooksey K E, Wigglesworth-Cooksey B. Adhesion of bacteria and diatoms surfaces in the sea: a review. Aquat Microbiol Ecol. 1995;9:87–96. [Google Scholar]
- 9.Dabros T, van de Ven T G M. Deposition of latex particles on glass surfaces in an impinging jet. Phys Chem Hydrodyn. 1987;8:161–172. [Google Scholar]
- 10.Dankert J, Hogt A H, Feijen J. Biomedical polymers, bacterial adhesion, colonization and infection. Crit Rev Biocompat. 1986;2:219–301. [Google Scholar]
- 11.Denkov N D, Velev O D, Kralchevsky P A, Ivanov I B, Yoshimurea H, Nagayama K. Mechanism of formation of two-dimensional crystals from latex particles on substrates. Langmuir. 1992;8:3183–3190. [Google Scholar]
- 12.Doig F, Lollar B S, Ferris F G. Microbial communities in deep Canadian shield ground waters. An in situ biofilm experiment. Geomicrobiol J. 1995;13:91–102. [Google Scholar]
- 13.Fang H H P, Chan K-Y, Xu L-C. Quantification of bacterial adhesion forces using atomic force microscopy (AFM) J Microbiol Methods. 2000;40:89–97. doi: 10.1016/s0167-7012(99)00137-2. [DOI] [PubMed] [Google Scholar]
- 14.Fielden M, Hayes R A, Ralston J. Surface and capillary forces affecting air bubble-particle interactions in aqueous electrolytes. Langmuir. 1996;12:3721–3727. [Google Scholar]
- 15.Flint S H, Bremer P J, Brooks J D. Biofilms in dairy manufacturing plant — description, current concerns and methods of control. Biofouling. 1997;11:81–97. [Google Scholar]
- 16.Foster D B, Philpott D, Abul-Milh M, Huesca M, Sherman P M, Lingwood C A. Phosphatidylethanolamine recognition promotes enteropathogenic E. coli and enterohemorrhagic E. coli host cell attachment. Microb Pathog. 1999;27:289–301. doi: 10.1006/mpat.1999.0305. [DOI] [PubMed] [Google Scholar]
- 17.Gómez-Suárez, C., H. C. van der Mei, and H. J. Busscher. Air bubble induced detachment of polystyrene particles with different size from collector surfaces in parallel plate flow chamber. Colloids Surf. A, in press.
- 18.Gómez-Suárez C, van der Mei H C, Busscher H J. Air bubble-induced detachment of positively and negatively charged polystyrene particles from collector surfaces in a parallel-plate flow chamber. J Adhes Sci Technol. 2000;14:1527–1538. [Google Scholar]
- 19.Gómez-Suárez C, Noordmans J, van der Mei H C, Busscher H J. Detachment of colloidal particles from collector surfaces with different electrostatic charge and hydrophobicity by attachment to air bubbles in a parallel plate flow chamber. Phys Chem Chem Phys. 1999;1:4423–4427. [Google Scholar]
- 20.Gómez-Suárez C, Noordmans J, van der Mei H C, Busscher H J. Removal of colloidal particles from quartz collector surfaces as stimulated by the passage of air-liquid interfaces. Langmuir. 1999;15:5123–5127. [Google Scholar]
- 21.Gristina A G, Naylor P, Myrvik Q. Infections from biomaterials and implants: a race for the surface. Med Prog Technol. 1988;14:205–224. [PubMed] [Google Scholar]
- 22.Hewitt D, Fornasiero D, Ralston J. Bubble-particle attachment. J Chem Soc Faraday Trans. 1995;94:1997–2001. [Google Scholar]
- 23.Hewitt D, Fornasiero D, Ralston J, Fisher L R. Aqueous film drainage at the quartz/water/air interface. J Chem Soc Faraday Trans. 1998;89:817–822. [Google Scholar]
- 24.Hiemenz P C. Electrophoresis and other electrokinetic phenomena. In: Lagowski J J, editor. Principles of colloid and surface chemistry. New York, N.Y: Marcel Dekker; 1977. pp. 452–487. [Google Scholar]
- 25.Kojima N, Shiota M, Sadahira Y, Handa K, Hakomori S. Cell adhesion in a dynamic flow system as compared to static system. J Biol Chem. 1992;267:1264–1270. [PubMed] [Google Scholar]
- 26.Krachesvsky P A, Nagayama K. Capillary forces between colloidal particles. Langmuir. 1994;10:23–26. [Google Scholar]
- 27.Landa A S, van de Mei H C, van Rij G, Busscher H J. Efficacy of ophthalmic solutions to detach adhering Pseudomonas aeruginosa from contact lenses. Cornea. 1998;17:293–300. [PubMed] [Google Scholar]
- 28.Leenaars A F M, O'Brien S B G. A new approach to the removal of sub-micron particles from solid (silicon) substrates. In: Mittal K L, editor. Particles on surfaces: detection, adhesion and removal. New York, N.Y: Plenum Press; 1989. pp. 361–372. [Google Scholar]
- 29.Lewis S J, Gilmour A. Microflora associated with the internal surfaces of rubber and stainless steel milk transfer pipeline. J Appl Bacteriol. 1987;62:327–333. doi: 10.1111/j.1365-2672.1987.tb04928.x. [DOI] [PubMed] [Google Scholar]
- 30.Liu Y. Adhesion kinetics of nitrifying bacteria on various thermoplastic supports. Colloids Surf B. 1995;5:316–329. [Google Scholar]
- 31.Nguyen-Van A. The collision of fine particles and single air-bubbles in flotation. J Colloid Interface Sci. 1994;162:123–128. [Google Scholar]
- 32.Notermans S, Dormans J A M A, Mead G C. Contribution of surface attachment to the establishment of microorganisms in food processing plants: a review. Biofouling. 1991;5:21–36. [Google Scholar]
- 33.Percival S L, Knapp J S, Wales D S, Edyvean R G J. Biofilm development in potable quality water. Biofouling. 1999;13:259–277. [Google Scholar]
- 34.Pitt W G, McBride M O, Barton A J, Sagers R D. Air-water interfaces displace adsorbed bacteria. Biomaterials. 1993;14:605–608. doi: 10.1016/0142-9612(93)90179-6. [DOI] [PubMed] [Google Scholar]
- 35.Reid G, Bruce A W, McGroarty J A, Cheng M C, Costerton J W. Is there a role for lactobacilli in prevention of urogenital and intestinal infections? Clin Microbiol Rev. 1990;3:335–344. doi: 10.1128/cmr.3.4.335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Rijnaarts H H M, Norde W, Bouwer E J, Lyklema J, Zehnder A J B. Bacterial adhesion under static and dynamic conditions. Appl Environ Microbiol. 1993;59:3255–3265. doi: 10.1128/aem.59.10.3255-3265.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Schneider R P. Conditioning film-induced modification of substratum physicochemistry. Analysis by contact angles. J Colloid Interface Sci. 1996;182:204–213. [Google Scholar]
- 38.Schulze H J, Wahl B, Gottschalk G. Determination of adhesive strength of particles with the liquid/gas interface in flotation by means of a centrifugal method. J Colloid Interface Sci. 1989;128:57–65. [Google Scholar]
- 39.Sjollema J, Busscher H J, Weerkamp A H. Experimental approaches for studying adhesion of microorganisms to solid substrata: applications and mass transport. J Microbiol Methods. 1989;9:79–90. [Google Scholar]
- 40.Taylor G T, Zheng D, Lee M, Troy P J, Gyananath G, Sharma S K. Influence of surface properties on accumulation of conditioning films and marine bacteria on substrata exposed to oligotrophic waters. Biofouling. 1997;11:31–57. [Google Scholar]
- 41.Urzi C, Lisi S, Criseo G, Pernice A. Adhesion and degradation of marble by Micrococcus strain isolated from it. Geomicrobiol J. 1991;9:81–90. [Google Scholar]
- 42.Van Oss C J. Interfacial forces in aqueous media. New York, N.Y: Marcel Dekker; 1994. Polar or Lewis acid-base interactions; pp. 18–46. [Google Scholar]
- 43.Van Wagenen R J, Andrade J D. Flat plate streaming potential investigations: hydrodynamics and electrokinetic equivalency. J Colloid Interface Sci. 1980;76:305–314. [Google Scholar]
- 44.Wirtanen G, Husmark U, Mattila-Sandholm T. Microbial evaluation of the biotransfer potential from surfaces with Bacillus biofilms after rinsing and cleaning procedures in closed food-processing systems. J Food Prot. 1996;59:727–733. doi: 10.4315/0362-028X-59.7.727. [DOI] [PubMed] [Google Scholar]