

Abstract
Increasingly intensive strategies to maintain biodiversity and ecosystem function are being deployed in response to global anthropogenic threats, including intentionally introducing and eradicating species via assisted migration, rewilding, biological control, invasive species eradications, and gene drives. These actions are highly contentious because of their potential for unintended consequences. We conducted a global literature review of these conservation actions to quantify how often unintended outcomes occur and to elucidate their underlying causes. To evaluate conservation outcomes, we developed a community assessment framework for systematically mapping the range of possible interaction types for 111 case studies. Applying this tool, we quantified the number of interaction types considered in each study and documented the nature and strength of intended and unintended outcomes. Intended outcomes were reported in 51% of cases, a combination of intended outcomes and unintended outcomes in 26%, and strictly unintended outcomes in 10%. Hence, unintended outcomes were reported in 36% of all cases evaluated. In evaluating overall conservations outcomes (weighing intended vs. unintended effects), some unintended effects were fairly innocuous relative to the conservation objective, whereas others resulted in serious unintended consequences in recipient communities. Studies that assessed a greater number of community interactions with the target species reported unintended outcomes more often, suggesting that unintended consequences may be underreported due to insufficient vetting. Most reported unintended outcomes arose from direct effects (68%) or simple density‐mediated or indirect effects (25%) linked to the target species. Only a few documented cases arose from more complex interaction pathways (7%). Therefore, most unintended outcomes involved simple interactions that could be predicted and mitigated through more formal vetting. Our community assessment framework provides a tool for screening future conservation actions by mapping the recipient community interaction web to identify and mitigate unintended outcomes from intentional species introductions and eradications for conservation.
Keywords: assisted migration, biological control, biological invasion, gene drive, invasive species eradication, nontarget effects, rewilding, species introduction, control biológico, efectos no considerados, erradicación de especies invasoras, genética dirigida, introducción de especies, invasión biológica, migración asistida, retorno a la vida silvestre
Short abstract
Article impact statement: A global literature review reveals that many unintended outcomes of species introductions and eradications for conservation can be avoided.
Abstract
Evaluación de las Consecuencias Involuntarias de las Introducciones y Erradicaciones Intencionales de Especies para el Manejo Mejorado de la Conservación
Resumen
Actualmente se despliegan estrategias cada vez más intensas para mantener la biodiversidad y la función del ecosistema como respuesta a las amenazas antropogénicas mundiales, incluyendo la introducción y erradicación intencionales de especies por medio de la migración asistida, el retorno a la vida silvestre, el control biológico, la erradicación de especies invasoras y la genética dirigida. Estas acciones son muy polémicas por el potencial que tienen para generar consecuencias involuntarias. Realizamos una revisión de la literatura mundial sobre estas acciones de conservación para cuantificar cuán seguido ocurren las consecuencias involuntarias y cuáles son sus causas subyacentes. Para evaluar los resultados de conservación, desarrollamos un marco de trabajo de evaluación comunitaria para mapear sistemáticamente el rango de posibles interacciones para 111 estudios de caso. Con la aplicación de esta herramienta cuantificamos el número de tipos de interacción consideradas en cada estudio y documentamos la naturaleza y la fuerza de los resultados involuntarios. Se reportaron los resultados voluntarios en 51% de los casos, una combinación de resultados voluntarios e involuntarios en 26% de los casos y estrictamente los resultados involuntarios en el 10% de los casos. Por lo tanto, los resultados involuntarios fueron reportados en el 36% de todos los casos evaluados. En la evaluación general de los resultados de conservación (sopesando los efectos voluntarios y. los involuntarios), algunos efectos involuntarios fueron bastante inocuos en relación con el objetivo de conservación, mientras que otros resultaron en consecuencias involuntarias severas para las comunidades receptoras. Los estudios que evaluaron un mayor número de interacciones comunitarias con la especie objetivo reportaron resultados involuntarios con mayor frecuencia, lo que sugiere que las consecuencias involuntarias pueden estar subvaloradas debido al escrutinio insuficiente. La mayoría de los resultados involuntarios reportados surgieron de los efectos directos (68%) o de los efectos indirectos o mediados por la densidad (25%) vinculados con la especie diana. Solamente unos cuantos casos documentados surgieron de interacciones más complejas (7%). Por lo tanto, la mayoría de los resultados involuntarios involucran interacciones simples que podrían ser pronosticadas y mitigadas por medio de un escrutinio más formal. Nuestro marco de trabajo de evaluación comunitaria proporciona una herramienta para la revisión de las acciones de conservación en el futuro mediante el mapeo de la red de interacciones entre comunidades receptoras y para la mitigación de los resultados involuntarios surgidos de las introducciones y erradicaciones intencionales de especies a favor de la conservación.
INTRODUCTION
Management of natural systems in the Anthropocene is becoming increasingly intensive. Threats to biodiversity, such as climate change, species extinctions, and biological invasions, are being met with intentional species introductions in the name of assisted migration (Hoegh‐Guldberg et al., 2008), rewilding (Corlett, 2016), and biological control (Hoddle, 2004). In other cases, species are intentionally eradicated, fully or functionally, over large areas, where they act as introduced (Glen et al., 2013; Zavaleta et al., 2001) or native pests (Ripple et al., 2014). Moreover, new technologies, such as gene drives, promise even more powerful eradication tools (Webber et al., 2015). Intentional species introductions and eradications successfully mitigate some very substantive conservation threats such as extirpating problematic invasive species (Brooke et al., 2018; Gurr & Wratten, 2000; Jones et al., 2016; Prior et al., 2018; Zavaleta et al., 2001). However, these approaches have also caused unintended consequences to nontarget species, with negative, sometimes severe, effects permeating entire ecosystems (Bergstrom et al., 2009; Courchamp et al., 2003; Doak et al., 2008; Pearson & Callaway, 2006; Prior et al., 2018; Simberloff & Stiling, 1996; Zavaleta et al., 2001). Such negative outcomes arising from introductions and eradications have generated extensive controversy over the application of each of these conservation strategies (Hoddle, 2004; Lorimer et al., 2015; McLachlan et al., 2007; Ricciardi & Simberloff, 2009; Rubenstein, & Rubenstein, 2016; Simberloff & Stiling, 1996; Webber et al., 2015; Zavaleta et al., 2001). Despite such warnings and documented impacts, these conservation actions continue, driven by the substantial conservation risks associated with taking no action (e.g., Hoddle, 2004; Hoegh‐Guldberg et al., 2008; Marvier & Kareiva, 2020).
Many unintended outcomes of intentional species introductions and eradications arise from relatively simple two‐ and three‐species interactions that could be predicted from ecological theory. For example, introduced cats (Felis catus) were eradicated from Macquarie Island to alleviate cat predation on native seabirds. However, cat eradication generated an unintended trophic cascade by releasing introduced rabbit (Oryctolagus cuniculus) populations from cat predation, allowing rabbit herbivory to devastate native plant communities (Bergstrom et al., 2009; but see Dowding et al., 2009). These herbivore‐driven declines in native plants then increased exotic plant populations via apparent competition. In another case, the introduction of red squirrels (Tamiasciurus hudsonicus) to Newfoundland, an island, an action intended to bolster American marten (Martes americana) populations via food subsidies, was linked to a serious decline of the endemic Newfoundland red crossbill (Loxia curvirostra percna) due to marten‐crossbill competition for black spruce (Picea mariana) seeds (Benkman, 2010). Given that these basic interactions represent well‐understood community interactions, such as resource competition (Gause, 1934), apparent competition (Holt, 1977), and trophic cascades (Paine, 1980), an important question is why have these core concepts from community ecology not been better integrated to avoid such unintended outcomes in conservation?
Although many unintended conservation outcomes may result from simple interactions, others can arise from more cryptic and complex pathways associated with longer interaction chains or trait‐mediated indirect interactions (see Courchamp et al., 2003; McGregor et al., 2020; Pearson & Callaway, 2003; Simberloff & Stiling, 1996). Hence, effectively screening for such unintended outcomes requires assessment of multispecies assemblages participating in a variety of direct and indirect interactions (Wootton, 2002). Yet, most ecological theory has been built from basic community interactions focusing on only a few species at a time (e.g., Gause, 1934; Holt, 1977; Paine, 1980; Tilman, 1980). Although pairwise Lotka–Volterra type equations representing such interactions can be integrated into quantitative community‐level models (Godoy et al., 2018), they carry high data demands, requiring precise information on species abundances and interaction coefficients for all key components within a web (but see Adams et al. [2020] for caveats). Hence, ecological theory holds great potential to inform management and conservation decisions, but traditional modeling approaches are commonly too data hungry to satisfy. Meanwhile, imminent anthropogenic threats are forcing intensive, real‐time management actions to be taken with incomplete information. In short, better tools are needed to inform and guide complex conservation actions.
In an effort to better understand and address the challenge of unintended consequences of conservation management, we first drew from theoretical and empirical work in community ecology to develop a community assessment framework to examine how species introductions and eradications can influence recipient communities. Next, we conducted a global literature review of large‐scale studies to quantify unintended outcomes arising from intentional species introductions (assisted migration, rewilding, and biological control) and eradications (invasive species removal and gene drives). Although these conservation strategies represent a range of disparate conservation objectives, they share a critical commonality: by manipulating an entire species (whether adding or removing them) each strategy serves as a community‐level perturbation with the potential to profoundly affect other organisms via similar community interaction pathways. Hence, community ecology holds potential to inform and improve each of these practices similarly. Collating the results from the literature review allowed us to assess the rate of unintended outcomes arising from these conservation actions and identify the types of interaction pathways commonly associated with unintended outcomes to inform conservation. We applied the community assessment framework to 111 case studies from aquatic and terrestrial ecosystems around the globe to illustrate its potential as a tool for systematically considering and mapping the relevant recipient community interaction web to screen for species most susceptible to unintended consequences and mitigate such outcomes.
METHODS
Framework for assessing community‐level effects of species manipulations
In community ecology and network theory (Godoy et al., 2018; May, 1972; Wootton, 1994), four basic components make up a community interaction web: species in the web (i.e., network nodes); linkages connecting interacting species or nodes; type (e.g., predation, mutualism) and hence directionality of the linkages (i.e., positive or negative); and strength of the interactions (May, 1972; Wootton, 1994). With information on each of these components, it is theoretically possible to model community interactions with a standard community interaction matrix to determine the effects of changing the abundance (or presence) of one community member on the remaining community components (May, 1972; Ramsey & Veltman, 2005). Although abundance information can be important for modeling, it is not essential for determining community outcomes, so we focused on these four elements. Hence, these are the basic elements required for understanding a community's response to perturbations, such as species introductions and eradications. Building from these basic elements, we determined which of the multitude of species and interactions in the recipient community were most likely to be affected by introducing or removing a species targeted for management (hereafter, target species). Fortunately, empirical studies in community ecology demonstrate that most communities are composed of many weak and few strong interactions (Neutel et al., 2002; Paine, 1992). Hence, most interactions and species can be ignored, but determining which ones should be included to best capture substantive changes within the community is a challenging and potentially subjective process.
To address this problem, we drew from the wealth of research in community ecology to construct a community assessment framework for systematically considering the types of interaction linkages most relevant to the species targeted for management (Figure 1). Although there are a multitude of specific types of community interactions, most biotic interactions fall into the broad categories of resource uptake (consumption or intake of foods, nutrients, energy), competition (resource and interference competition), consumption (including herbivory and predation), parasitism (pathogen–host interactions), and mutualism (Wootton, 1994). In addition, ecosystem engineering is a powerful interaction by which engineers may strongly interact with many community members (Jones et al., 1997). Of course, anthropogenic factors, including management, can greatly influence organisms and their interactions and should be considered. Finally, all organisms have physical requirements that explicitly define their fundamental niches and determine their realized niches as a function of biotic interactions that are often environmentally conditioned (Hutchinson, 1957). By outlining the basic components of a community interaction web, this framework provides a tool for systematically considering the range of interactions directly linking a target species to other community members in order to identify the key nodes of the community interaction web and determine the nature and strength of their linkages—the essential components for understanding community outcomes. Further, assuming that strong interactions are those most likely to perpetuate indirect effects (e.g., Paine, 1980), this same process can be repeated for any organisms presumed to be strongly directly linked to the target species in order to incorporate potentially important indirect interactions. In sum, the community assessment framework provides a tool for understanding community interactions and for systematically constructing a community interaction web relevant to the management action that can help to elucidate past outcomes and vet proposed actions.
FIGURE 1.
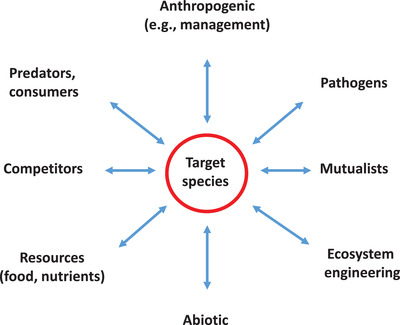
Community assessment framework for generating a community interaction web to understand how a target species intentionally introduced or eradicated for conservation purposes may influence community outcomes. Interactions included may link the target species to other network nodes (species or system components) to identify the species most likely to be affected by the action, the nature of each interaction linkage (i.e., positive or negative), and the strength of each interaction. There can be multiple sets of interactions in one interaction type (e.g., multiple competitors of the target species) that may need to be considered. The initial community assessment focuses on immediate linkages to the target species likely to be strong enough to substantively alter the abundance or function of other system components. If this assessment indicates the target species is likely to have strong effects on particular community members, then the same process should be applied to the affected species or node to extend the web and include indirect effects, under the assumption that most strong indirect effects derive from strong direct effects. Hence, this approach systematically identifies and follows out strong linkages until they become weak, thereby delineating the relevant community of concern
Literature review
To quantify the prevalence of unintended outcomes across a range of management actions and better understand the community interaction pathways by which they arise, we conducted a global literature search to generate a sample of peer‐reviewed publications addressing proposed or executed conservation actions involving intentional species introductions (assisted migration, rewilding, and biocontrol) or eradications (invader removal or gene drive). We used the Web of Science All Databases search engine to generate a preliminary list of papers on our five focal categories related to intentional species introductions (assisted migration, rewilding, and biocontrol) or eradications (invader removal or gene drive). Our search was conducted on April 11, 2019. For all four searches, we included a subject search term to focus on environmental sciences and ecology (SU [subject], Environmental Sciences & Ecology) plus additional search terms to screen titles and topics for each of our four focal categories. For assisted migration and rewilding, we included the following terms: TI [title search] (rewilding OR “assisted migration” OR “assisted colonization”) AND TS [topic search] (propos* OR outcome OR result OR consequence). This generated a list of 131 papers. For biocontrol introduction, we included the following terms: TI = (biocontrol OR “biological control”) AND TS = (intro*) AND TS = (plant OR insect OR invertebrate) AND TS = (propos* OR outcome OR result OR consequence) NOT TS = (crop OR agri* OR pathogen OR lab* OR greenhouse). This generated a list of 179 papers. For invader removals, we included the following terms: TI = (remov*) AND TI = (inva* OR exotic OR introduced) AND TS = (propos* OR outcome OR result OR consequence). This generated a list of 156 papers. For gene drives used to eradicate pest species, we included the following search terms: TI = (gene drive) AND TS = (control). This generated a list of 87 papers.
We further screened the preliminary list of 553 papers from the above search results to include in our literature review. We screened each paper to ensure that each study either proposed or executed a species introduction or eradication and that it was conducted at a large enough scale to be considered a realistic community‐level management action. We included studies that were conducted on whole ecosystems (e.g., ponds, islands) or a minimum of 1 ha. Studies were not included, for example, if an invasive species was experimentally removed from small plots (e.g., 1‐m2 plots) or potential biocontrol insects were screened on host plants in a lab. In addition, executed actions were restricted to those that effectively established the introduced species (e.g., for biological control, the control agent established) or effected some reduction of the species targeted for eradication to ensure that we only included studies that achieved the minimal objective with the target organism. A small number of articles included multiple cases, which we included as independent observations. In total, this resulted in 172 cases in 140 articles of our original list of 553 potential articles. For articles that met the above criteria, we classified each case based on management action (introduction or eradication), management status (proposed or executed), and management category (assisted migration, biocontrol, gene drive, invasive species removal, or rewilding).
We recognize that the published literature may not represent a comprehensive assessment of all the factors and interactions considered prior to each management action because not all conservation actions are published in peer‐reviewed journals (e.g., Wainwright et al., 2018). Many conservation actions around the world are documented in part or whole in the gray literature of government outlets or may go undocumented. However, because the rules and regulations governing management differ by local and national governments, as do the processes for documenting such efforts, it is logistically infeasible to acquire a globally representative, comprehensive accounting of all such actions. This problem has been noted by others who have gone on to demonstrate the value of reviewing the published literature for advancing conservation, despite these limitations (e.g., Wainwright et al., 2018). Accordingly, our inferences are based on the assumption that our survey of the peer‐reviewed scientific literature established from our targeted keyword search of this topic provides an index representative of the underlying efforts and the types of outcomes that occur, rather than an exhaustive evaluation of the underlying case studies.
Quantifying outcomes from the literature review
To understand the extent to which each management action resulted in intended versus unintended outcomes, we scored the outcomes of the management action based on the objectives of the effort and assessments of the authors as follows. We categorized an outcome as intended when the management action successfully accomplished the management goal (e.g., an exotic pest was successfully extirpated from an island). We categorized an outcome as unintended when the authors reported an outcome occurred that was not part of the management objective (i.e., there was a change in a nontarget species abundance or other community component). When the authors reported both intended and unintended outcomes of the management action, we categorized the outcome as “mixed,” and when no detectable effect of the action was reported, we scored the outcome as “neutral.” If an unintended or mixed outcome was reported, we also recorded the nature of the unintended effects (density mediated or trait mediated) and indicated whether it was a direct or indirect effect, including the number of nontarget species involved (e.g., a direct density‐mediated effect [D1], an indirect trait‐mediated effect [T2]) whenever possible.
In our assessment, we defined the terms unintended effects and unintended outcomes (used synonymously) in an ecological context relative to the intended management action and the reported response of nontarget species or system components. We considered an unintended effect negative or positive as measured by the response of the nontarget organism or system component (e.g., if a nontarget species declined, the effect was negative; if it increased, it was positive). Hence, unintended effects were objectively, ecologically defined relative to the intended management goals. In contrast, interpreting whether the overall conservation outcome was successful or deleterious (i.e., evaluating the sum of the intended and unintended effects) was far more subjective. For example, the Macquarie Island case (Bergstrom et al., 2009) discussed in the INTRODUCTION might be seen as a success by those focused on mitigating impacts on seabirds, but those focused on the native plant response might consider this a case of deleterious unintended consequences. Because of this subjectivity, we did not attempt to judge the overall conservation outcome for each case study. Rather, we highlight in the DISCUSSION the general types of overall outcomes observed, reserving the phrase unintended consequences for deleterious overall outcomes as distinct from specific unintended effects within a case study.
To determine the extent to which community‐level interactions may have been considered for each management action, as reflected in the published literature, we also evaluated the degree to which each study addressed the critical elements of a basic community interaction web (Figure 1). We examined each article and case to determine the number of types of interactions with the target organism that were addressed (0–8) as well as the number of associated interaction strengths that were at least qualitatively considered (0–8) as identified in the community assessment framework (Figure 1). Because the direction of the interaction was almost always defined in the context of its linkage to the target species (e.g., predation is negative for the prey), we did not separately track this element. Finally, because the quantification of community interactions ranged from none to highly quantitative, we assigned scores to account for the sophistication of the analysis of the community components as follows: 0, no model; 1, qualitative model (e.g., verbal model or path diagram); 2, quantitative model (e.g., linear model); and 3, mechanistic model (e.g., Lotka–Volterra models.). Summing the scores across these categories provided a scale for evaluating results from the literature review across management actions and management categories (range 0–19: 0–8 interaction types, 0–8 interaction strengths, and 0–3 analytical score). Obviously, there can be many species and linkages represented in a single interaction type (e.g., a target species could have many competitors). However, we did not account for multiple interactions within a single interaction type because our primary objective was to determine the degree to which the range of key elements of a basic community interaction web were addressed to account for potential unintended outcomes. Accounting for only one interaction type, no matter how many interactions within that type were addressed, would not sufficiently account for the breadth of unintended outcomes that might arise from a species‐level community perturbation.
Data analyses
We carried out analyses in R (R Core Team, 2018). To understand the relationship between the extent to which different community components were addressed and the likelihood and nature of unintended outcomes reported, we modeled the unintended impacts as categorical variables, which described the nature and complexity of unintended effects (see above), as a function of the community assessment score. To do so, we used multinomial log‐linear models with the nnet R package (Venables & Ripley, 2002). To control for confounding differences between each study in our statistical analyses, we also included the variables management status and category, taxa (e.g., plant vs. invertebrate vs. mammal), habitat, and region of the target organism. We did not include management action in this analysis because of collinearity with management category (e.g., rewilding involves intentional introductions, not eradications). Starting with a model containing all candidate explanatory variables and interactions, we simplified models with backward selection until we minimized the AIC, retaining only the top model (Burnham & Anderson, 2003). Additionally, we generated summary statistics (mean [SE]) for management action and management categories based on the scores for each community element.
RESULTS
Our screening of papers from the literature search generated 28 assisted migration, 15 rewilding, 63 biocontrol, 13 gene drive, and 53 invasive species removal cases (Appendix S1). These cases were widely, although not evenly, distributed globally (North America 91, Australia 22, Islands including New Zealand 15, Europe 13, Asia 9, South America 8, Africa 7, Antarctica 1, arctic regions of both Europe and North America 6). Overall, when outcomes from these management actions were documented, 51% (57 of 111 cases that documented management outcomes) were classified as strictly intended effects, 10% as strictly unintended effects, 26% as mixed, and 6% as neutral (i.e., biocontrol introductions neither reduced the target nor affected nontarget species or had unknown outcomes) (Figure 2a). Hence, 36% of cases reported some unintended outcomes. For those unintended effects reported, most arose from simple density‐mediated direct effects of the manipulated species on a community member (68%), followed by simple density‐mediated indirect effects (25%) and a few more complex trait‐mediated indirect interactions (7%) (Figure 2b). However, most documented unintended outcomes arose from invader removal and biocontrol cases. This result is linked to the fact that these management actions have been executed far more often than the newer management actions, such as assisted migration, rewilding, and gene drives. Notably, higher community assessment scores correlated with increased probabilities of unintended outcomes (Appendices S2 and S3). This result that was unaffected by potentially confounding factors, such as taxa or geographic region (Appendix S2), suggesting that our results are conservative and more comprehensive screening would identify additional unintended outcomes. Regarding the breadth of interaction types reported on, 51% of studies addressed more than two interaction types, 39% mentioned only one type of interaction, and 10% of studies did not address any interactions between nontarget species and recipient community members (Figure 3a). Fewer studies yet discussed interaction strengths, even qualitatively; 42% of studies made no mention of interaction strength (Figure 3b). Finally, few studies offered quantitative evaluation of community outcomes (Figure 3c). Overall, eradication efforts touched on more components than introduction efforts (sum of the scores for total number of interaction types, interaction strengths, and modeling scores was x̅ [SE] = 4.7 [0.3] vs. 3.2 [0.3], respectively), whereas proposed actions addressed fewer components than executed actions (2.7 [0.3] vs. 4.5 [0.2], respectively). These results provide little evidence that newly proposed actions involve greater consideration of unintended outcomes. The published literature suggested that the extent to which community components were considered was fairly limited across all conservation action categories (mean [SE]: 3.8 [0.2]; range, 0–13) (Figure 3c). Longer‐standing fields, such as invasive species removal and biological control, tended to address more components (invader removal = 5.0 [0.3], biological control = 4.3 [0.3], gene drive = 3.5 [0.5], rewilding = 2.9 [0.8], assisted migration = 1.1 [0.3]).
FIGURE 2.
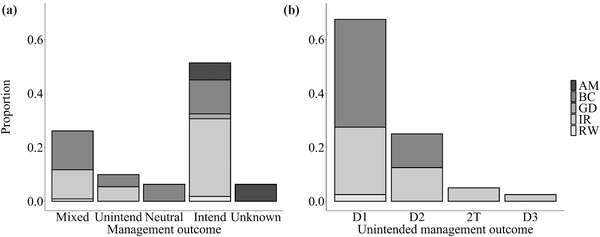
Proportion of studies identified from a review of the global literature on executions of species introductions or eradications for conservation purposes for which outcomes were separated by management category (AM, assisted migration; BC, biocontrol; GD, gene drive; IR, invader removal; RW, rewilding) for (a) management outcome (mixed, intended and unintended outcomes; unintend, unintended outcomes only; neutral, no changes among nontarget species [all cases where a biocontrol agent established but neither controlled the target nor affected nontarget species]; intend, strictly intended outcomes; unknown, outcome unclear) and (b) interaction web distance to unintended outcome (D1, density‐mediated direct effects; D2, density‐mediated indirect effect; 2T, trait‐mediated indirect effect)
FIGURE 3.
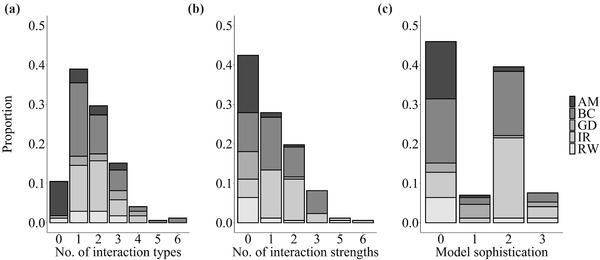
Results from a global literature review of studies proposing or executing species introductions or eradications for conservation purposes. Case studies are broken out by (a) interaction type (e.g., competition vs. predation), (b) interaction strength, and (c) level of model sophistication (0, no model; 1, qualitative model; 2, quantitative model; 3, mechanistic model) to show the proportion of cases addressing each category (e.g., the proportion of studies addressing zero, one, … or six interaction types) by management category (AM, assisted migration; BC, biocontrol; GD, gene drive; IR, invader removal; RW, rewilding)
DISCUSSION
Management actions proposing intentional species introductions and eradications for conservation purposes are controversial due to risks to nontarget species (Doak et al., 2008; Hoegh‐Guldberg et al., 2008; Ricciardi & Simberloff, 2009; Rubenstein, & Rubenstein, 2016; Webber et al., 2015). However, these debates are largely founded in anecdotes and inference about such risks because outcomes are not well quantified. In our literature review, we found that collectively these conservation actions generated intended outcomes most of the time, but unintended outcomes were common side effects of success and sometimes the sole outcome of well‐intended efforts. Understanding whether unintended effects result in deleterious unintended consequences in the context of the overall conservation outcome requires consideration of the relative strength of intended and unintended effects. Although many unintended effects are minor and can be written off as acceptable collateral damage relative to overall benefits (Ferrero et al., 2013; Johnson & Cushman, 2007; Leege & Kilgore, 2014; Lindenmayer et al., 2017), others may result in serious unintended consequences, including substantial declines in native species populations (Bateman et al., 2015; Bergstrom et al., 2009; Darrah & van Riper, 2018), secondary invasion following invader removal (Dickie et al., 2014; reviewed in Pearson et al., 2016), and increased disease risk to humans (Pearson & Callaway, 2006). Such strong unintended effects are particularly concerning if the intended outcome is not achieved or is fully offset by the unintended effects (Pearson et al., 2016). Although some have suggested that unintended consequences are visages of past conservation failures from the 1960s and 1970s (Marvier & Kareiva, 2020), most of the cases we reviewed took place since that time (up through 2019). Moreover, we found the likelihood of unintended outcomes being reported increased as studies addressed more interaction types, suggesting that unintended outcomes may be underreported because they are undervetted. Most importantly, our finding that most unintended outcomes arose from simple direct effects (68%), instead of more complex interactions, suggests that many unintended effects could potentially be identified by more formal screening.
The finding that 51% (87 of 172) of the cases we reviewed addressed two or more types of interactions with the target species (with multiple sets of interactions within an interaction type commonly considered) suggests that managers are attempting to account for unintended outcomes much of the time. However, 10% of the articles mentioned no interactions with nontarget species and 39% addressed only a single type of interaction with the target organism. If one assumes that most organisms targeted for conservation management experience minimal resource requirements, competitors, and top‐down interactions (e.g., a consumer, predator, or parasite), then one might expect most organisms to have at minimum three substantive types of interactions within the affected community that should be considered. Our finding that studies reporting on more interaction types with the target species also reported more unintended outcomes suggests that more formal vetting of these conservation actions prior to their enactment may highlight these unintended consequences and help managers plan for them.
Our review of the published scientific literature provides evidence that managers are attempting to address nontarget species and unintended outcomes. However, our results also indicate that potentially important nontarget species (e.g., an important predator or competitor of the target species) and system components are commonly not considered. Lacking is a systematic means for both vetting potential unintended outcomes and documenting this process in a transparent manner. For example, even in the most comprehensive and well‐quantified assessments of intentional species eradications, where multiple potential direct and indirect interactions are identified and outcomes are predictively modeled (e.g., Dexter et al., 2012; Raymond et al., 2011), it is often unclear what process was used to identify the system components that were assessed, which ones were evaluated and disregarded as irrelevant, and which were not considered due to oversight. Even in classical biological control of exotic plants, a field with arguably the most rigorous and well‐documented prescreening testing of any of the conservation actions we assessed (e.g., Briese, 2005), only one type of interaction is normally considered—the direct attack or consumption of nontarget species by the target (but see Todd et al., 2020). Yet, many unintended outcomes of these biological control introductions have arisen through other interaction pathways (Carvalheiro et al., 2008; Pearson & Callaway, 2003; Veldtman et al., 2011). Finally, certain types of interactions that can be among the most important may not be recognized or considered for their potential to cause strong unintended outcomes. For example, introducing artificial water sources to facilitate the rewilding of tortoises on desert islands (Falcón & Hansen, 2018) could strongly affect other endemic and non‐native species in such water‐limited systems. A framework for systematically and objectively considering the range of biotic, abiotic, and anthropogenic factors that may strongly link a species targeted for management to other community members could provide a simple tool for identifying potential unintended outcomes for further consideration before introductions as well as provide a means for consistently documenting this process.
The community assessment framework we developed for elucidating how species introductions and eradications may affect recipient communities (Figure 1) provides a formal assessment tool that could be applied to more holistically consider how an organism proposed for conservation action interacts with other species in the community and how these interactions may permeate through the ecosystem. Because the framework is built from foundational ecological theory reinforced by decades of empirical work, it provides a generic tool for systematically evaluating the basic components common to all communities to identify which components may be most relevant to a specific conservation action. In conducting our review and applying this framework to a variety of systems from saltwater to freshwater to terrestrial ecosystems, ranging from deserts to tropical forests, we found it to be widely applicable. This tool can be readily implemented by using natural history information, available literature, and, if available, empirical data from the system to systematically assess the range of possible interactions that might strongly link the target organism to other community members. This process can be used to quickly generate a community interaction web that highlights strong direct and indirect interactions linking the target organism to other community members, thereby identifying potential unintended outcome pathways (Figure 1).
Although this method is more systematic and objective than the various approaches we found in the literature, it is still a subjective process to decide which interactions are strong enough to consider. This sort of subjectivity can be further addressed by gathering managers, subject experts, and stakeholders as appropriate to assimilate the relevant information and define the community interaction in a consensus approach (e.g., Özesmi & Özesmi, 2004). Once the interaction web is completed, the risks associated with potential unintended outcomes can then be weighed against the potential benefits of taking the conservation action (Marvier & Kareiva, 2020) through consensus decision or additional research can be conducted to explicitly assess the identified risks prior to action to determine whether the proposed action is warranted. Although this strategy does not completely remove subjectivity, it establishes a process for broader input that can help overcome subjective bias, and it establishes a formal, systematic, and transparent process that can be clearly defined and defended on the grounds that important factors are considered and documented and stakeholders have opportunity for input.
Another limitation of the tool as presented above is that it provides a purely qualitative back‐of‐the‐envelope approach to map and identify possible community outcomes. Although our literature review suggested that simply formalizing this mapping approach could help reduce many fairly obvious unintended outcomes in conservation management (68% were simple direct effects), qualitative modeling tools have been developed that can incorporate the basic information derived from applying the framework in Figure 1 to formally evaluate the potential for the various interactions to play out (Baker & Bode, 2020; Baker et al., 2018; Geary et al., 2020; Hobbs et al., 2002; Ramsey & Veltman, 2005; Ramsey et al., 2012; Ramsey & Norbury, 2009; Raymond et al., 2011). These approaches greatly add objectivity to the process and are highly recommended. However, as noted above, identifying the community interaction web to be applied in these models remains fairly arbitrary; the types of interactions evaluated, species selected, and the depth to which the web is extended into the community are not always clearly justified or systematically applied. For example, in one of the more rigorous qualitative modeling attempts to vet unintended effects of a conservation action, Raymond et al. (2011) state, “Our particular situation is further complicated by another form of ambiguity: model structure uncertainty (there are a number of interactions that could potentially be included or excluded from the model).” They addressed this problem by “considering a large number of model structures, encompassing all possible combinations of unknown interactions.” We propose that our community assessment framework provides an ecologically based and systematic means for delineating the community interaction web that is prerequisite to such modeling approaches. Hence, whether the community assessment is used to map the relevant community interaction web for purely qualitative evaluation or as the basis for applying modeling approaches, this tool provides a mechanism for systematically assessing the community of interest in a manner that is more ecologically grounded, systematic, and reproducible than current methods.
Our literature review revealed that unintended outcomes commonly arise from intentional species introductions and eradications. Although many unintended outcomes may be relatively minor, some are serious. Importantly, most documented cases of unintended outcomes arise from basic direct and simple indirect interactions that could potentially be identified and managed through formal vetting of proposed management actions prior to execution. Toward this end, we drew from community ecology theory to develop a community assessment framework that provides a generic tool for systematically defining the community interaction web most strongly linked to the target organism in order to highlight potential unintended effects on nontarget community members. We propose that applying this simple tool in conservation planning is not overly burdensome, and doing so could greatly reduce unintended outcomes. The tool also provides increased transparency and defensibility for executing these conservation actions. Finally, this tool provides a systematic approach for developing the information necessary for more formal modeling of both intended and unintended outcomes, thereby delineating a pathway toward more advanced conservation to reduce unintended consequences.
Supporting information
Table S1
Appendix 1. Database of case studies and their assigned community assessment scores will be provided as an uploaded .csv file.
Appendix 2. Model selection for predictor variables which might influence the relationship between the sum of the scores for community elements and model complexity and the probability of and complexity of unintended management outcomes being reported, for executed species introductions or eradications for conservation purposes. Model selection was carried out by ΔAIC using backward selection.
Appendix 3. Parameters estimated by the model reported in Appendix S2.
ACKNOWLEDGMENTS
We thank Y. K. Ortega, P. Ramsey, M. K. Schwartz, and anonymous reviewers for input on previous drafts. D.E.P. was supported by the Rocky Mountain Research Station, USDA Forest Service and MPG Ranch. T.J.C. was supported by the National Science Foundation (NSF) Graduate Research Fellowship under grant 366280. P.G.H. was supported by funding from MPG Ranch and NSF DEB‐1901552.
Pearson DE., Clark TJ & Hahn PG. (2022). Evaluating unintended consequences of intentional species introductions and eradications for improved conservation management. Conservation Biology, 36, e13734. 10.1002/cobi.13734
Article impact statement: A global literature review reveals that many unintended outcomes of species introductions and eradications for conservation can be avoided.
LITERATURE CITED
- Adams, M. P. , Sisson, S. A. , Helmstedt, K. J. , Baker, C. M. , Holden, M. H. , Plein, M. , Holloway, J. , Mengersen, K. L. , & McDonald‐Madden, E. (2020). Informing management decisions for ecological networks, using dynamic models calibrated to noisy time‐series data. Ecology Letters, 23, 607–619. [DOI] [PubMed] [Google Scholar]
- Baker, C. M. , & Bode, M . (2020). Recent advances of quantitative modeling to support invasive species eradication on islands. Conservation Science and Practice, 3(2), e246. [Google Scholar]
- Baker, C. M. , Holden, M. H. , Plein, M. , McCarthy, M. A. , & Possingham, H. P . (2018). Informing network management using fuzzy cognitive maps. Biological Conservation, 224, 122–128. [Google Scholar]
- Bateman, H. L. , Merritt, D. M. , Glenn, E. P. , & Nagler, P. L . (2015). Indirect effects of biocontrol of an invasive riparian plant (Tamarix) alters habitat and reduces herpetofauna abundance. Biological Invasions, 17(1), 87–97. [Google Scholar]
- Benkman, C. W . (2010). Diversifying coevolution between crossbills and conifers. Evolution: Education and Outreach, 3, 47–53. [Google Scholar]
- Bergstrom, D. M. , Lucieer, A. , Kiefer, K. , Wasley, J. , Belbin, L. , Pedersen, T. K. , & Chown, S. L . (2009). Indirect effects of invasive species removal devastate World Heritage Island. Journal of Applied Ecology, 46, 73–81. [Google Scholar]
- Briese, D. T . (2005). Translating host‐specificity test results into the real world: The need to harmonize the yin and yang of current testing procedures. Biological Control, 35, 208–214. [Google Scholar]
- Brooke, M. D. L. , Bonnaud, E. , Dilley, B. J. , Flint, E. N. , Holmes, N. D. , Jones, H. P. , Provost, P. , Rocamora, G. , Ryan, P. G. , Surman, C. , & Buxton, R. T . (2018). Seabird population changes following mammal eradications on islands. Animal Conservation, 21, 3–12. [Google Scholar]
- Burnham, K. P. , & Anderson, D . (2003). Model selection and multimodel inference: A practical information‐theoretic approach. Spring Science & Business Media. [Google Scholar]
- Carvalheiro, L. G. , Buckley, Y. M. , Ventim, R. , Fowler, S. V. , & Memmott, J . (2008). Apparent competition can compromise the safety of highly specific biocontrol agents. Ecology Letters, 11, 690–700. [DOI] [PubMed] [Google Scholar]
- Corlett, R. T . (2016). Restoration, reintroduction, and rewilding in a changing world. Trends in Ecology and Evolution, 31, 453–462. [DOI] [PubMed] [Google Scholar]
- Courchamp, F. , Woodroffe, R. , & Roemer, G . (2003). Removing protected populations to save endangered species. Science, 302, 1532–1532. [DOI] [PubMed] [Google Scholar]
- Darrah, A. J. , & van Riper, C . (2018). Riparian bird density decline in response to biocontrol of Tamarix from riparian ecosystems along the Dolores River in SW Colorado, USA. Biological Invasions, 20, 709–720. [Google Scholar]
- Dexter, N. , Ramsey, D. S. L. , MacGregor, C. , & Lindenmayer, D . (2012). Predicting ecosystem wide impacts of wallaby management using a fuzzy cognitive map. Ecosystems, 15, 1363–1379. [Google Scholar]
- Dickie, I. A. , St John, M. G. , Yeates, G. W. , Morse, C. W. , Bonner, K. I. , Orwin, K. , & Peltzer, D. A . (2014). Belowground legacies of Pinus contorta invasion and removal result in multiple mechanisms of invasional meltdown. AoB Plants, 6, plu056. 10.1093/aobpla/plu056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doak, D. , Estes, J. , Halpern, B. , & Ute, J . (2008). Understanding and predicting ecological dynamics: Are major surprises inevitable? Ecology, 89(4), 952–961. [DOI] [PubMed] [Google Scholar]
- Dowding, J. E. , Murphy, E. C. , Springer, K. , Peacock, A. J. , & Krebs, C. J. (2009). Cats, rabbits, Myxoma virus, and vegetation on Macquarie Island: A comment on Bergstrom et al.(2009). Journal of Applied Ecology, 46, 1129–1132. [Google Scholar]
- Falcón, W. , & Hansen, D. M . (2018). Island rewilding with giant tortoises in an era of climate change. Philosophical Transactions of the Royal Society B: Biological Sciences, 373, 20170442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrero, V. , Castro, S. , Costa, J. , Acuña, P. , Navarro, L. , & Loureiro, J . (2013). Effect of invader removal: Pollinators stay but some native plants miss their new friend. Biological Invasions, 15, 2347–2358. [Google Scholar]
- Gause, G. F . (1934). Experimental analysis of Vito Volterra's mathematical theory of the struggle for existence. Science, 79, 16–17. [DOI] [PubMed] [Google Scholar]
- Geary, W. , Bode, M. , Doherty, T. , Fulton, E. , Nimmo, D. , Tulloch, A. , Tulloch, V. , & Ritchie, E . (2020). A guide to ecosystem models and their environmental applications. Nature Ecology & Evolution, 4(11), 1459–1471. [DOI] [PubMed] [Google Scholar]
- Glen, A. S. , Atkinson, R. , Campbell, K. J. , Hagen, E. , Holmes, N. D. , Keitt, B. S. , Parkes, J. P. , Saunders, A. , Sawyer, J. , & Torres, H . (2013). Eradicating multiple invasive species on inhabited islands: The next big step in island restoration ? Biological Invasions, 15, 2589–2603. [Google Scholar]
- Godoy, O. , Bartomeus, I. , Rohr, R. P. , & Saavedra, S . (2018). Towards the integration of niche and network theories. Trends in Ecology & Evolution, 33, 287–300. [DOI] [PubMed] [Google Scholar]
- Gurr, G. , & Wratten, S . (2000). Biological control: Measures of success. Kluwer Academic Publishers. [Google Scholar]
- Hobbs, B. F. , Ludsin, S. A. , Knight, R. L. , Ryan, P. A. , & Ciborowski, J. J. H . (2002). Fuzzy cognitive mapping as a tool to define management. Ecological Applications, 12, 1548–1565. [Google Scholar]
- Hoddle, M. S . (2004). Restoring balance: Using exotic species to control invasive exotic species. Conservation Biology, 18, 38–49. [Google Scholar]
- Hoegh‐Guldberg, O. , Hughes, L. , Mcintyre, S. , Lindenmayer, D. B. , Parmesan, C. , Possingham, H. P. , & Thomas, C. D . (2008). Assisted colonization and rapid climate change. Science, 321, 345–346. [DOI] [PubMed] [Google Scholar]
- Holt, R . (1977). Predation, apparent competition, and the structure of prey communities. Theoretical Population Biology, 12, 197–229. [DOI] [PubMed] [Google Scholar]
- Hutchinson, G . (1957). Concluding remarks. Cold Spring Harbour Symposium on Quantitative Biology, 22, 415–427. [Google Scholar]
- Johnson, B. E. , & Cushman, J. H . (2007). Influence of a large herbivore reintroduction on plant invasions and community composition in a California grassland. Conservation Biology, 21, 515–526. [DOI] [PubMed] [Google Scholar]
- Jones, C. G. , Lawton, J. H. , & Shachak, M . (1997). Positive and negative effects of organisms as physical ecosystem engineers. Ecology, 78, 1946–1957. [Google Scholar]
- Jones, H. P. , Holmes, N. D. , Butchart, S. H. , Tershy, B. R. , Kappes, P. J. , Corkery, I. , Aguirre‐Muñoz, A. , Armstrong, D. P. , Bonnaud, E. , Burbidge, A. A. , Campbell, K. , Courchamp, F. , Cowan, P. E. , Cuthbert, R. J. , Ebbert, S. , Genovesi, P. , Howald, G. R. , Keitt, B. S. , Kress, S. W. , … Croll, D. A. (2016). Invasive mammal eradication on islands results in substantial conservation gains. Proceedings of the National Academy of Sciences of the United States of America, 113, 4033–4038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leege, L. M. , & Kilgore, J. S . (2014). Recovery of foredune and blowout habitats in a freshwater dune following removal of invasive Austrian pine (Pinus nigra). Restoration Ecology, 22, 641–648. [Google Scholar]
- Lindenmayer, D. B. , Wood, J. , MacGregor, C. , Hobbs, R. J. , & Catford, J. A . (2017). Non‐target impacts of weed control on birds, mammals, and reptiles. Ecosphere, 8, e01804. [Google Scholar]
- Lorimer, J. , Sandom, C. , Jepson, P. , Doughty, C. E. , Barua, M. , & Kirby, K . (2015). Rewilding: Science, practice, and politics. Annual Review of Environmental Resources, 40, 39–62. [Google Scholar]
- Marvier, M. , & Kareiva, P . (2020). It is time to rebalance the risk equation. Frontiers in Ecology and the Environment, 18(8), 423. 10.1002/fee.2256 [DOI] [Google Scholar]
- May, R . (1972). Will a large complex system be stable? Nature, 238, 413‐414. [DOI] [PubMed] [Google Scholar]
- McGregor, H. , Moseby, K. , Johnson, C. N. , & Legge, S . (2020). The short‐term response of feral cats to rabbit population decline: Are alternative native prey more at risk? Biological Invasions, 22, 799–811. [Google Scholar]
- McLachlan, J. S. , Hellmann, J. J. , & Schwartz, M. W . (2007). A framework for debate of assisted migration in an era of climate change. Conservation Biology, 21, 297–302. [DOI] [PubMed] [Google Scholar]
- Neutel, A.‐M. , Heesterbeek, J. A. P. , & De Ruiter, P. C. (2002). Stability in real food webs: Weak links in long loops. Science, 296, 1120–1123. [DOI] [PubMed] [Google Scholar]
- Özesmi, U. , & Özesmi, S . (2004). Ecological models based on people's knowledge: A multi‐step fuzzy cognitive mapping approach. Ecological Modelling, 176, 47–59. [Google Scholar]
- Paine, R . (1992). Food‐web analysis through field measurement of per capita interaction strength. Nature, 355, 73–75. [Google Scholar]
- Paine, R. T . (1980). Food webs: Linkage, interaction strength and community infrastructure. Journal of Animal Ecology, 49, 666–685. [Google Scholar]
- Pearson, D. E. , & Callaway, R. M . (2003). Indirect effects of host‐specific biological control agents. Trends in Ecology & Evolution, 18, 456–461. [Google Scholar]
- Pearson, D. E. , & Callaway, R. M . (2006). Biological control agents elevate hantavirus by subsidizing deer mouse populations. Ecology Letters, 9, 443–450. [DOI] [PubMed] [Google Scholar]
- Pearson, D. E. , Ortega, Y. K. , Runyon, J. B. , & Butler, J. L . (2016). Secondary invasion: The bane of weed management. Biological Conservation, 197, 8–17. [Google Scholar]
- Prior, K. M. , Adams, D. C. , Klepzig, K. D. , & Hulcr, J . (2018). When does invasive species removal lead to ecological recovery? Implications for management success. Biological Invasions, 20, 267–283. [Google Scholar]
- Ramsey, D. , & Veltman, C . (2005). Predicting the effects of perturbations on ecological communities: What can qualitative models offer? Journal of Animal Ecology, 74, 905–916. [Google Scholar]
- Ramsey, D. S. L. , Forsyth, D. M. , Veltman, C. J. , Nicol, S. J. , Todd, C. R. , Allen, R. B. , Allen, W. J. , Bellingham, P. J. , Richardson, S. J. , Jacobson, C. L. , & Barker, R. J . (2012). An approximate Bayesian algorithm for training fuzzy cognitive map models of forest responses to deer control in a New Zealand adaptive management experiment. Ecological Modelling, 240, 93–104. [Google Scholar]
- Ramsey, D. S. L. , & Norbury, G. L . (2009). Predicting the unexpected: Using a qualitative model of a New Zealand dryland ecosystem to anticipate pest management outcomes. Austral Ecology, 34, 409–421. [Google Scholar]
- Raymond, B. , McInnes, J. , Dambacher, J. M. , Way, S. , & Bergstrom, D. M . (2011). Qualitative modelling of invasive species eradication on subantarctic Macquarie Island. Journal of Applied Ecology, 48, 181–191. [Google Scholar]
- R Core Team . (2018). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. [Google Scholar]
- Ricciardi, A. , & Simberloff, D . (2009). Assisted colonization is not a viable conservation strategy. Trends in Ecology and Evolution, 24, 248–253. [DOI] [PubMed] [Google Scholar]
- Ripple, W. J. , Estes, J. A. , Beschta, R. L. , Wilmers, C. C. , Ritchie, E. G. , Hebblewhite, M. , Berger, J. , Elmhagen, B. , Letnic, M. , Nelson, M. P. , Schmitz, O. J. , Smith, D. W. , Wallach, A. D. , & Wirsing A. J. (2014). Status and ecological effects of the world's largest carnivores. Science, 343(6167), 1241484. [DOI] [PubMed] [Google Scholar]
- Rubenstein, D. R. , & Rubenstein, D. I . (2016). From Pleistocene to trophic rewilding: A wolf in sheep's clothing. Proceedings of the National Academy of Sciences of the United States of America, 113, E1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simberloff, D. , & Stiling, P . (1996). How risky is biological control? Ecology, 77, 1965–1974. [Google Scholar]
- Tilman, D . (1980). A graphical‐mechanistic approach to competition and predation. The American Naturalist, 116, 362–393. [Google Scholar]
- Todd, J. H. , Pearce, B. M. , & Barratt, B. I. P . (2020). Using qualitative food webs to predict species at risk of indirect effects from a proposed biological control agent. BioControl, 66, 45–58. [Google Scholar]
- Veldtman, R. , Lado, T. F. , Botes, A. , Procheş, Ş. , Timm, A. E. , Geertsema, H. , & Chown, S. L . (2011). Creating novel food webs on introduced Australian acacias: Indirect effects of galling biological control agents. Diversity and Distributions, 17, 958–967. [Google Scholar]
- Venables, W. , & Ripley, B . (2002). Modern applied statistics with S (4th ed.). Springer. [Google Scholar]
- Wainwright, C. E. , Staples, T. L. , Charles, L. S. , Flanagan, T. C. , Lai, H. R. , Loy, X. , Reynolds, V. A. , & Mayfield, M. M. (2018). Links between community ecology theory and ecological restoration are on the rise. Journal of Applied Ecology, 55, 570–581. [Google Scholar]
- Webber, B. L. , Raghu, S. , & Edwards, O. R . (2015). Opinion: Is CRISPR‐based gene drive a biocontrol silver bullet or global conservation threat? Proceedings of the National Academy of Sciences of the United States of America, 112, 10565–10567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wootton, J. T . (1994). The nature and consequences of indirect effects in ecological communities. Annual Review of Ecology and Systematics, 25, 443–466. [Google Scholar]
- Wootton, J. T . (2002). Indirect effects in complex ecosystems: Recent progress and future challenges. Journal of Sea Research, 48, 157–172. [Google Scholar]
- Zavaleta, E. S. , Hobbs, R. J. , Mooney, H. A. , Zavaleta, E. , & Mooney, H. A . (2001). Viewing invasive species removal in a whole‐ecosystem context. Trends in Ecology & Evolution, 16, 454–459. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1
Appendix 1. Database of case studies and their assigned community assessment scores will be provided as an uploaded .csv file.
Appendix 2. Model selection for predictor variables which might influence the relationship between the sum of the scores for community elements and model complexity and the probability of and complexity of unintended management outcomes being reported, for executed species introductions or eradications for conservation purposes. Model selection was carried out by ΔAIC using backward selection.
Appendix 3. Parameters estimated by the model reported in Appendix S2.