

Abstract
Background and Purpose
Cannabis has been used to treat epilepsy for millennia, with such use validated by regulatory approval of cannabidiol (CBD) for Dravet syndrome. Unregulated artisanal cannabis‐based products used to treat children with intractable epilepsies often contain relatively low doses of CBD but are enriched in other phytocannabinoids. This raises the possibility that other cannabis constituents might have anticonvulsant properties.
Experimental Approach
We used the Scn1a +/− mouse model of Dravet syndrome to investigate the cannabis plant for phytocannabinoids with anticonvulsant effects against hyperthermia‐induced seizures. The most promising, cannabigerolic acid (CBGA), was further examined against spontaneous seizures and survival in Scn1a +/− mice and in electroshock seizure models. Pharmacological effects of CBGA were surveyed across multiple drug targets.
Key Results
The initial screen identified three phytocannabinoids with novel anticonvulsant properties: CBGA, cannabidivarinic acid (CBDVA) and cannabigerovarinic acid (CBGVA). CBGA was most potent and potentiated the anticonvulsant effects of clobazam against hyperthermia‐induced and spontaneous seizures, and was anticonvulsant in the MES threshold test. However, CBGA was proconvulsant in the 6‐Hz threshold test and a high dose increased spontaneous seizure frequency in Scn1a +/− mice. CBGA was found to interact with numerous epilepsy‐relevant targets including GPR55, TRPV1 channels and GABAA receptors.
Conclusion and Implications
These results suggest that CBGA, CBDVA and CBGVA may contribute to the effects of cannabis‐based products in childhood epilepsy. Although these phytocannabinoids have anticonvulsant potential and could be lead compounds for drug development programmes, several liabilities would need to be overcome before CBD is superseded by another in this class.
Keywords: cannabinoids, CBGA, Dravet syndrome, epilepsy
Abbreviations
- CBD
cannabidiol
- CBDVA
cannabidivarinic acid
- CBG
cannabigerol
- CBGA
cannabigerolic acid
- CBGV
cannabigerivarin
- CBGVA
cannabigerovarinic acid
- CBDV
cannabidivarin
- GTCS
generalized tonic–clonic seizure
- Δ9‐THCV
tetrahydrocannabivarin
- MES
maximal electroshock
What is already known
Cannabis has been used since ancient times to treat epilepsy.
Some phytocannabinoids derived from cannabis, including cannabidiol (CBD), reduce seizures or epileptiform activity.
What does this study add
Three phytocannabinoids were anticonvulsant: cannabigerolic acid (CBGA), cannabidivarinic acid (CBDVA) and cannabigerovarinic acid (CBGVA).
CBGA had both anticonvulsant and proconvulsant effects across seizure models and affected epilepsy‐relevant molecular targets.
What is the clinical significance
Phytocannabinoid acids may contribute to the effects of cannabis‐based products used to treat intractable epilepsies.
1. INTRODUCTION
Cannabis has been used to treat epilepsy for millennia and prior to prohibition was routinely prescribed by physicians for this purpose. Recent years have seen widespread legalization and utilization of medicinal cannabis worldwide, partly driven by stories of artisanal cannabis products having remarkable therapeutic effects in children with severe, intractable epilepsy. This phenomenon has intensified efforts to unlock the medicinal potential of this ancient herb.
Cannabis is a complex mixture of bioactive molecules including >140 terpenophenolic compounds known as phytocannabinoids. The main psychoactive phytocannabinoid, Δ9‐tetrahydrocannabinol (Δ9‐THC) and the principal non‐intoxicating component, cannabidiol (CBD), have well‐characterized pharmacological effects. Although Δ9‐THC has shown mixed proconvulsant and anticonvulsant effects, CBD has exhibited anticonvulsant properties across multiple preclinical models (Anderson, Absalom, et al., 2019; Anderson et al., 2020; Chiu et al., 1979; Kaplan et al., 2017; Karler et al., 1973; Karler & Turkanis, 1979; Socała et al., 2019). CBD has now received regulatory approval, in the purified formulation Epidiolex®, for the treatment of Dravet syndrome and Lennox–Gastaut syndrome following a series of successful randomized controlled trials (Devinsky et al., 2018, 2017; Thiele et al., 2018).
However, many childhood epilepsy patients continue to be treated with unregulated artisanal products extracted from the cannabis plant, often due to high expense and limited availability of Epidiolex® and, in some cases, a preference for extracts containing a ‘full spectrum’ of multiple phytocannabinoids and terpenoids rather than isolated CBD. Artisanal cannabis‐based products have diverse phytocannabinoid profiles and may often deliver low CBD doses, suggesting that other phytocannabinoids may contribute to the anticonvulsant properties of cannabis (Huntsman et al., 2019; Pamplona et al., 2018; Suraev et al., 2018; Tzadok et al., 2016). Indeed, preclinical research has uncovered a suite of phytocannabinoids, which reduce epileptiform activity and seizures in animal models, including Δ9‐tetrahydrocannabidivarin (Δ9‐THCV), cannabidivarin (CBDV) and cannabidiolic acid (CBDA) (Amada et al., 2013; Anderson et al., 2020, 2021; Anderson, Low, et al., 2019; Benson et al., 2020; Chiu et al., 1979; Hill et al., 2012, 2010, 2013; Huntsman et al., 2019; Karler et al., 1973; Karler & Turkanis, 1979).
Further exploration of the cannabis plant may therefore uncover additional lead molecules with efficacy against drug‐resistant epilepsies such as Dravet syndrome. Loss‐of‐function mutations in SCN1A, the gene that encodes voltage‐gated sodium channel α1 subunit (Nav1.1), are present in approximately 80% of children with Dravet syndrome (Marini et al., 2011). Heterozygous deletion of Scn1a (Scn1a +/−) in mice reproduces the key features of Dravet syndrome, thereby providing a screening platform for potential Dravet syndrome therapies (Hawkins et al., 2017; Kaplan et al., 2017; Yamakawa, 2009). Here, we used the Scn1a +/− mouse model of Dravet syndrome to assess the anticonvulsant activity of seven different phytocannabinoids, including cannabigerolic acid (CBGA), the acid form of cannabigerol (CBG), a molecule that serves as the biosynthetic precursor to both Δ9‐THC and CBD in the cannabis plant.
2. METHODS
2.1. Animals
All animal care and experimental procedures were approved by the University of Sydney Animal Ethics Committee in accordance with the Australian Code of Practice for the Care and Use of Animals for Scientific Purposes. Animal studies are reported in compliance with the ARRIVE guidelines (Percie du Sert et al., 2020) and with the recommendations made by the British Journal of Pharmacology (Lilley et al., 2020). Scn1a +/− mice were purchased from The Jackson Laboratory (Stock 37107‐JAX; Bar Harbor, USA) and maintained as a congenic line on the 129S6/SvEvTac background (129.Scn1a +/−). For experiments, F1 mice were generated by breeding heterozygous 129.Scn1a +/− mice with wild‐type C57BL/6J mice (Jackson Laboratory Stock 000664; Animal Resources Centre, Canning Vale, Australia). The Scn1a genotype was determined as previously described (Hawkins et al., 2017; Miller et al., 2014). Swiss outbred mice were purchased from Australian BioResources (Stock SwissTacAusb; Moss Vale, Australia). Mice were group housed (up to six mice per cage) in individually ventilated cages with corncob bedding in a specific pathogen‐free facility that operated under a 12‐h light/12‐h dark cycle with ad libitum access to food and water. At the end of the study, mice were killed humanely using isoflurane followed by decapitation, unless otherwise specified.
2.2. Pharmacokinetic analysis
A pharmacokinetic study of cannabigerivarin (CBGV) to determine the target experimental time point for the hyperthermia‐induced seizure protocol was performed as previously described (Anderson, Low, et al., 2019). Briefly, male and female wild‐type mice received a single intraperitoneal injection of 1‐mg·kg−1 CBGV. At selected time points, mice were anaesthetized with isoflurane and whole blood was collected via cardiac puncture, with n = 4–5 per time point. Plasma was isolated by centrifugation and stored at −80°C until assayed. CBGV concentrations in plasma at each time point were averaged (Figure S1).
2.3. Hyperthermia‐induced seizures
Hyperthermia‐induced seizure experiments were conducted on male and female Scn1a +/− mice as previously described (Anderson, Absalom, et al., 2019; Hawkins et al., 2017). Briefly, mice aged postnatal days 14–16 (P14–16) were pretreated with vehicle or cannabinoid by a single intraperitoneal injection, with n = 12–18 per treatment. Fifteen minutes prior to the target experimental time point for each drug, the rectal probe was inserted, mice were acclimated for 5 min, and the hyperthermia protocol was initiated. Mouse core body temperature was elevated 0.5°C every 2 min until the onset of the first clonic convulsion with loss of posture or until 42.5°C was reached. Mice that reached 42.5°C were held at temperature for 3 min and were considered seizure free if no seizure occurred during the hold. In this assay, we and others have shown that CBD is anticonvulsant against hyperthermia‐induced seizures at 100 mg·kg−1 (Anderson, Absalom, et al., 2019; Anderson et al., 2020; Kaplan et al., 2017). The doses of the seven cannabinoids tested were chosen to provide a point of direct comparison with CBD at this dose. CBG and CBDV were not tested at the 100‐mg·kg−1 dose due to poor aqueous solubility and Δ9‐THCV was tested at doses known to antagonize the cannabinoid CB1 receptor in vivo in mice (3 and 10 mg·kg−1) (Pertwee et al., 2007). Following the hyperthermia‐induced seizure protocol, cohorts of mice (n = 5–10 per treatment) were anaesthetized with isoflurane and whole blood was collected via cardiac puncture. Plasma was isolated and samples were stored at −80°C until assayed. Seizure threshold temperatures were compared using Mantel–Cox log‐rank test in GraphPad Prism 8.2.1 (GraphPad Software, San Diego, USA) and P < 0.05 was considered statistically significant.
Experimental time points were based on previously determined time‐to‐peak plasma and brain concentrations or effect from the literature or our pharmacokinetic study (Figure S1) (Anderson, Low, et al., 2019; Deiana et al., 2012). Experimental time points were as follows: 120 min—CBG; 45 min—CBGA; 30 min—CBDV, CBGV, Δ9‐THCV and clobazam; and 15 min—CBDVA and CBGVA, which were administered after the 5 min acclimation period. Matched vehicle controls were administered at each experimental time point.
2.4. Spontaneous seizures and survival
Male and female Scn1a +/− mice were exposed to a single hyperthermia‐induced seizure event as described previously (Anderson et al., 2020; Hawkins et al., 2017). Briefly, at P18 mice were exposed to a thermally induced seizure as described above. At the onset of clonic convulsion with loss of posture, mice were immediately placed on a cooling pad to terminate the seizure. Mice were cooled to a core body temperature of 37°C, with a mortality rate as a result of the febrile seizure priming <2%. Mice were then randomly assigned to drug treatment groups, with n = 15–19 per treatment. Drugs were administered orally through supplementation in chow formulated in‐house using R&M Standard Diet (Specialty Feeds, Glen Forrest, Australia) irradiated powder with the following formulations: 1400‐mg CBGA·kg−1 chow, 2500‐mg CBGA·kg−1 chow, 80‐mg clobazam·kg−1 chow and 80‐mg clobazam plus 1400‐mg CBGA·kg−1 chow. Continuous video recording was captured using a day/night camera with an IR lens (Samsung SCB5003, Tamron 13F604IRSQ) and the number of spontaneous generalized tonic–clonic seizures (GTCS) in the 60‐h recording session was quantified offline by an observer blinded to treatment. Mice continued drug treatment to P30 to monitor survival. Plasma samples were isolated as described above on P31 within 30 min of lights on and stored at −80°C until assayed.
Statistical comparisons were made with GraphPad Prism using Fisher's exact test (proportion of mice seizure free), one‐way ANOVA followed by Dunnett's post hoc (seizure frequency) and P < 0.05 was considered statistically significant. Post hoc analyses were conducted only when F in ANOVA achieved P < 0.05 and there was no significant variance in homogeneity. Sample sizes subjected to statistical analysis had at least five samples per group (n = 5), where n is equal to the number or independent values. Statistical comparisons of survival were made with GraphPad Prism using Mantel–Cox log‐rank test and P < 0.05 was considered statistically significant. The data and statistical analysis comply with the recommendations of the British Journal of Pharmacology on experimental design and analysis in pharmacology (Curtis et al., 2018).
2.5. Maximal electroshock and 6‐Hz threshold tests
Maximal electroshock (MES) and 6‐Hz threshold tests were conducted in Swiss male mice using a rodent Electro‐Convulsive‐Therapy unit (Model 57800; Ugo Basile, Gemonio, Italy) as previously described (Benson et al., 2020). Briefly, mice were pretreated with vehicle or CBGA and returned to their home cage for 45 min. Positive control mice were treated with valproic acid (300 mg·kg−1) 15 min prior to seizure induction. For MES threshold test experiments, corneal electroshocks (60 Hz, 0.4 s, 0.5‐ms rectangular pulse width) were administered starting at 50 mA, with n = 12 per treatment. Mice were scored by an observer blinded to treatment for the presence or absence of GTCS with full hindlimb extension (hindlimbs at a 180° angle to the torso). Corneal electroshocks (6 Hz, 3 s, 0.2‐ms rectangular pulse width) starting at 20 mA were used for 6‐Hz threshold test experiments, with n = 12 per treatment. Mice were scored by an observer blinded to treatment for the presence or absence of a psychomotor seizure occurring within 30 s of the shock delivery. Seizure response was characterized by the presence of rhythmic jaw, forelimb clonus, immobility and/or Straub tail. The critical current (mA) at which 50% of mice seized (CC50) was determined using the ‘up‐and‐down’ method described by Kimball et al. (1957). Statistical comparisons of CC50 values between treatment groups were made using one‐way ANOVA followed by Dunnett's post hoc and P < 0.05 was considered statistically significant. Post hoc analyses were conducted only when F in ANOVA achieved P < 0.05 and there was no significant variance in homogeneity.
2.6. Plasma analytical chemistry
Phytocannabinoid and clobazam concentrations is plasma were assayed by LC–MS/MS as previously described (Anderson, Absalom, et al., 2019; Anderson, Low, et al., 2019). Briefly, plasma samples were prepared using solid–liquid extraction with methyl tert‐butyl ether and assayed by LC–MS/MS using a Shimadzu Nexera ultra‐high‐performance liquid chromatograph coupled to a Shimadzu 8030 triple quadrupole mass spectrometer (Shimadzu Corp., Kyoto, Japan). The mass spectrometer was operated in positive electrospray ionization mode with multiple reaction monitoring with mass transition pairs (Table S1). Quantification was achieved by comparing experimental samples to standards prepared with known amounts of drug. Cannabinoid concentrations were measured numerical values, which were not influenced by observer bias so blinding was not considered necessary. Statistical comparisons of plasma concentrations were made with GraphPad Prism using unpaired Student's t‐test and P < 0.05 was considered statistically significant. Sample sizes subjected to statistical analysis had at least five samples per group (n = 5), where n is equal to the number of independent values.
2.7. GPCR screen
Agonist and antagonist activity of CBGA (3 μM) at GPCRs was performed by Eurofins DiscoverX (Freemont, USA). The PathHunter® β‐arrestin functional cell‐based panel was used to investigate CBGA agonist and antagonist activity at 168 GPCRs and 74 orphan GPCRs (Table S2).
2.8. Cell culture
HEK 293 cells were used for all in vitro inhibition assays (CLS Cat# 300192/p777_HEK293, RRID:CVCL_0045). Cells were grown in DMEM supplemented with 10% FBS, 100‐U·ml−1 penicillin and 100‐μg·ml−1 streptomycin (P/S) in a humidified 5% CO2 atmosphere at 37°C. Cells stably expressing human GPR55 were generously provided by Dr Julia Kargl (University of Graz, Austria) and grown in supplemented culture media with added G418 (400 μg·ml−1) (Kargl et al., 2012). Cells stably expressing human TRPV1 were generated as previously described and grown in supplemented culture media with added zeocin (100 μg·ml−1) and blasticidin S (15 μg·ml−1) (Heblinski et al., 2020). All in vitro experiments were performed over a minimum of five independent experiments performed in duplicate. Data were fit to a three‐parameter logistic equation to determine IC50 values.
2.9. GPR55 ERK phosphorylation assay
ERK phosphorylation (pERK) was detected using the AlphaScreen® SureFire® p‐ERK ½ (Thr202/Tyr204) assay kit (PerkinElmer, Waltham, USA) according to the manufacturer's instructions. Briefly, 20–24 h prior to the assay, cells were resuspended in Leibovitz's L‐15 media supplemented with 1% FBS, 15‐mM glucose and P/S. Cells (110,000 per well) were plated into a Corning CellBIND (Corning Inc., Corning, USA) clear, flat‐bottomed 96‐well plate and incubated overnight at 37°C in ambient CO2. Media were aspirated and phenol red‐free DMEM/F‐12 (50 μl) was added to each well and incubated for 2–3 h at 37°C in a humidified 5% CO2 atmosphere and then drugs were added and incubated for 20 min. Media were then aspirated, lysis buffer was added and cells were placed on an orbital shaker for 10 min (500 r.p.m., room temperature). Lysates (5 μl) were mixed with reaction mixture (7 μl) and incubated for 3.5 h at room temperature. A CLARIOstar plate reader (BMG Labtech, Offenburg, Germany) was used to take fluorescence readings over 740 ms (excitation 680 nm and emission 520–620 nm). Baseline fluorescence as measured from an untreated cell condition was subtracted and fluorescence readings were normalized to those of 100‐nM phorbol 12‐myrisate 13‐acetate (PMA), a pERK stimulant. Final drug concentrations had ≤0.2% DMSO. Comparisons of curve fits for concentration–response curves were calculated using the extra‐sum‐of‐squares F test using GraphPad Prism.
2.10. TRPV1 intracellular calcium assay
Intracellular calcium was measured using the fluorescence imaging plate reader (FLIPR) Calcium 5 assay kit (Molecular Devices). Cells were grown to 90% confluency and 24 h prior to the assay, cells were detached from the flask and resuspended in Leibovitz's L‐15 media supplemented with 1% FBS, P/S and 15‐mM glucose. Cells (80,000–110,000 cells per well) were plated into a black, clear‐bottomed, poly‐d‐lysine‐coated 96‐well plate and incubated overnight at 37°C in ambient CO2. L‐15 media supplemented with tetracycline (10 μg·ml−1) were added to each well 4.5 h prior to the assay. Dye was prepared with high‐potassium HBSS assay buffer with (in mM) 13.8 NaCl, 2.2 HEPES, 0.0338 Na2HPO4, 0.4117 NaHCO3, 0.0441 KH2PO4, 0.0407 MgSO4, 0.0493 MgCl2, 0.533 KCl, 0.126 CaCl2, 250 probenecid and 1‐mg·ml−1 glucose with pH 7.4. Dye solution was added to each well and incubated for 90 min at 37°C in ambient CO2. A FlexStation 3 Microplate reader was used to take fluorescence readings every 2 s (excitation 485 nm and emission 565 nm) for 2 min prior to vehicle or CBGA addition. Three minutes following the addition of vehicle or CBGA, 10‐μM capsaicin was added and fluorescence was recorded for an additional 5 min. Baseline recordings were taken as the average fluorescence over 20 s prior to each drug addition. Changes in fluorescence were measured as the maximal increase in fluorescence from baseline following drug addition and normalized to the response of 10‐μM protease activated receptor 1 peptide (AusPep; Tullamarine, Australia). Final drug concentrations had ≤0.2% DMSO.
2.11. Two‐electrode voltage clamp
Cell currents were recorded from Xenopus laevis oocytes expressing the concatenated human γ2–β2–α1–β2–α1 GABA receptor using two‐electrode voltage clamp as previously described (Anderson, Absalom, et al., 2019; Liao et al., 2019). Briefly, GABA (15 μM) was applied three times and currents were normalized to the mean current of the last two GABA applications for modulation experiments. A washout of 7–12 min was performed between GABA applications and GABA was preincubated for 60 s. Per cent modulation by CBGA was derived by the equation described previously (Absalom et al., 2019). Data were fit to the Hill equation to determine I max and EC50 values using GraphPad Prism.
2.12. Desensitization assay
To quantify the level of desensitization induced by CBGA, dead volume was reduced and two successive pulses of 1‐mM GABA were applied to the oocyte. CBGA (10 μM) was preapplied to oocytes for 30, 60 and 180 s prior to coapplications of CBGA and GABA. Peak currents were measured and compared with applications of 1‐mM GABA alone under the same protocol. Statistical comparisons were made in GraphPad Prism using an unpaired Student's t‐test and P < 0.05 was considered statistically significant.
2.13. Materials
CBDV, CBG and Δ9‐THCV were purchased from THC Pharm GmbH (Frankfurt, Germany). CBGA was purchased from THC Pharm GmbH and Epichem (Bentley, Australia). CBGV was purchased from Toronto Research Chemicals Inc. (Ontario, Canada). CBDVA and CBGVA were synthesized by Professor Michael Kassiou at the University of Sydney (Australia). Clobazam and valproic acid were purchased from Sigma‐Aldrich Co. (St. Louis, USA). When available, analytical standards were purchased from Novachem Pty Ltd (Heidelberg West, Australia). For in vitro experiments, drugs were prepared as stocks in DMSO. For acute administration, drugs were prepared fresh on the day of the experiment. CBDV, CBG, CBGV and Δ9‐THCV were prepared in ethanol:Tween 80:saline (1:1:18). CBGV (100 mg·kg−1) was a suspension, with CBDV (60 mg·kg−1) and CBG (30 mg·kg−1) near solubility limits. CBDVA, CBGA, CBGVA and clobazam were prepared as solutions in vegetable oil. Drugs were administered as an intraperitoneal injection in a volume of 10 ml·kg−1. ML‐186 was purchased from Enamine (Kyiv, Ukraine). CID16020046 and lysophosphatidylinositol were purchased from Tocris Biosciences (Bristol, United Kingdom) and Sigma‐Aldrich (St. Louis, USA).
2.14. Nomenclature of targets and ligands
Key protein targets and ligands in this article are hyperlinked to corresponding entries in the IUPHAR/BPS Guide to PHARMACOLOGY http://www.guidetopharmacology.org and are permanently archived in the Concise Guide to PHARMACOLOGY 2019/20 (Alexander et al., 2019).
3. RESULTS
3.1. Novel anticonvulsant activity of phytocannabinoids against thermally induced seizures in Scn1a +/− mice
We assessed phytocannabinoids for anticonvulsant properties against hyperthermia‐induced seizures in Scn1a +/− mice, a model of the febrile seizures that occur in children with Dravet syndrome. Between P14 and P16, Scn1a +/− mice were treated with a single intraperitoneal injection of an individual phytocannabinoid or vehicle and then subjected to hyperthermia to assess the effect of the phytocannabinoids on the temperature threshold for a generalized tonic–clonic seizure (GTCS). Cannabidivarin (CBDV), cannabidivarinic acid (CBDVA), CBGA and cannabigerovarinic acid (CBGVA) displayed anticonvulsant properties, significantly elevating the temperature threshold for GTCS onset (Figures 1 and S2). CBDV exhibited an inverted U‐shaped dose effect, with an increase in GTCS temperature threshold at 30 mg·kg−1 relative to vehicle but not 60 mg·kg−1. CBDVA (100 mg·kg−1) and CBGVA (100 mg·kg−1) both significantly elevated the temperature threshold for GTCS compared with vehicle. CBGA dose‐dependently increased the GTCS temperature threshold compared with vehicle. Neither cannabigerol (CBG) nor cannabigerivarin (CBGV) affected thresholds for hyperthermia‐induced seizures. In contrast, Δ9‐THCV (3 mg·kg−1) treatment was proconvulsant, with a lower threshold temperature for GTCS. Plasma phytocannabinoid levels were measured following completion of the assay (Table S3).
FIGURE 1.
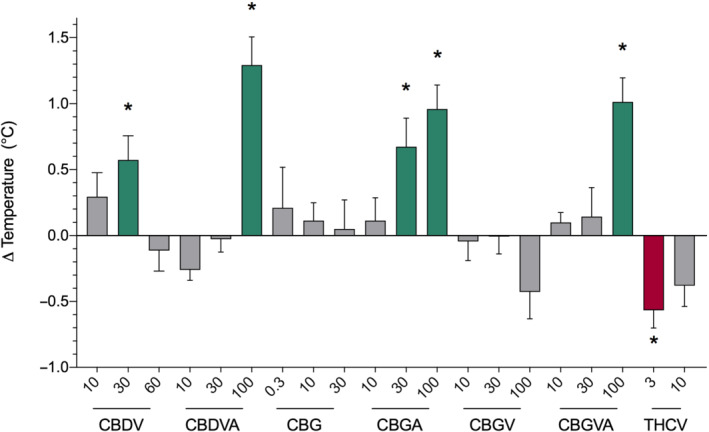
Phytocannabinoid screen against thermally induced seizures in Scn1a +/− mice yields compounds with novel anticonvulsant activity. Change in threshold temperature for generalized tonic–clonic seizure (GTCS) induced by hyperthermia compared with vehicle‐treated controls following acute intraperitoneal (mg·kg−1) administration of varying doses of compounds. Cannabidivarin (CBDV; 30 mg·kg−1), cannabidivarinic acid (CBDVA; 100 mg·kg−1), cannabigerolic acid (CBGA; 30 and 100 mg·kg−1) and cannabigerovarinic acid (CBGVA; 100 mg·kg−1) treatments were anticonvulsant resulting in a significantly improved response to thermal seizure induction (green bars), whereas Δ9‐tetrahydrocannabivarin (Δ9‐THCV) (3 mg·kg−1) was proconvulsant and significantly lowered the thermal threshold (red bar). Error bars represent SEM, with n = 12–18 per group (* P < 0.05; log‐rank Mantel–Cox). Treatments where mice were protected from GTCS were as follows: ‐ 0.3‐mg·kg−1 cannabigerol (CBG), four mice seizure free; 10‐mg·kg−1 CBG, one mouse seizure free. All vehicle‐treated mice had seizures. Temperatures of GTCS induction for individual mice, including vehicle‐treated mice, are found in Figure S2
3.2. Cannabigerolic acid (CBGA) increases the frequency of spontaneous seizures in Scn1a +/− mice
CBGA was the most potent of the seven phytocannabinoids against hyperthermia‐induced seizures, so it was further assessed against spontaneous seizures and survival of Scn1a +/− mice. Dravet syndrome often presents with febrile seizures that progress to other seizure types, including spontaneous afebrile GTCS. We modelled this seizure progression by priming Scn1a +/− mice with a single hyperthermia‐induced seizure at P18 and measuring subsequent spontaneous GTCS frequency. The average temperature of seizure onset was 40.2 ± 0.1°C (range: 37.9°C to 41.3°C). Mice received subchronic treatment of CBGA via supplementation in chow (1400‐ and 2500‐mg·kg−1 chow) following the thermally induced GTCS. These doses resulted in steady‐state plasma concentrations similar to those achieved following acute intraperitoneal treatment with 30‐ and 100‐mg·kg−1 CBGA, respectively (Figure 2a,b). Neither dose of CBGA affected the proportion of mice experiencing GTCS during the observation period. However, treatment with the higher dose of CBGA (2500‐mg·kg−1 chow) was proconvulsant and significantly increased GTCS frequency.
FIGURE 2.
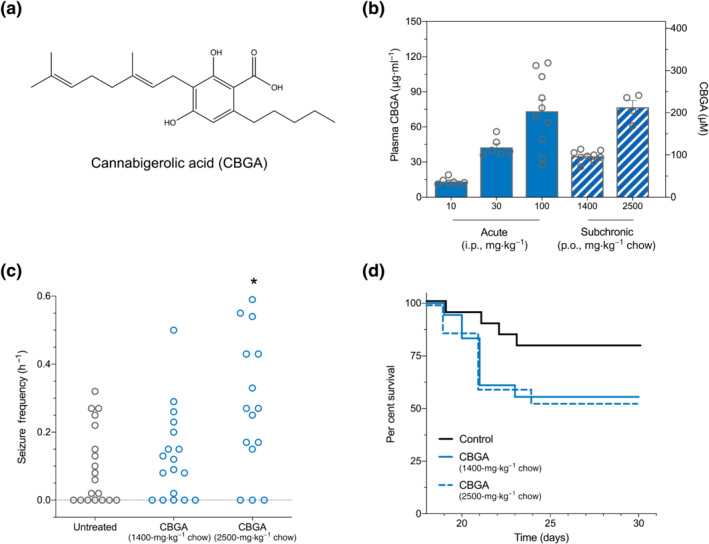
Cannabigerolic acid (CBGA) worsens spontaneous seizure frequency but not survival in Scn1a +/− mice. (a) Chemical structure of CBGA. (b) Average CBGA plasma concentrations in experimental Scn1a +/− mice treated with acute (solid bars) or subchronic (hatched bars) CBGA. Acute treatment of CBGA was administered as a single intraperitoneal injection for hyperthermia‐induced seizure experiments. Subchronic CBGA treatment was administered orally through supplementation in chow for spontaneous seizure and survival experiments. Error bars represent SEM, with n = 4–8 per group. CBGA concentrations are depicted as both mass concentrations (left Y axis) and molar concentrations (right Y axis). (c) Spontaneous generalized tonic–clonic seizure (GTCS) frequency of individual untreated and CBGA‐treated mice. Treatment began after the induction of a single hyperthermia‐induced seizure and spontaneous GTCS frequency was subsequently quantified over a 60‐h recording period. Treatment with the higher dose of CBGA (2500‐mg·kg−1 chow) significantly increased seizure frequency, with n = 15–19 per treatment (* P < 0.05; one‐way ANOVA followed by Dunnett's post hoc). (d) Survival curves comparing untreated and CBGA‐treated mice. Treatment with CBGA had no effect on survival of Scn1a +/− mice, with n = 15–19 per treatment (log‐rank Mantel–Cox, P > 0.05)
Dravet syndrome patients have a reduced lifespan, which is replicated in Scn1a +/− mice. CBGA treatment was continued until P30 to monitor effects on survival. Despite increasing GTCS frequency, the higher dose of CBGA did not affect survival of Scn1a +/− mice (Figure 2d). Survival did not differ with either CBGA dose relative to controls.
3.3. CBGA increases the anticonvulsant efficacy of clobazam against thermally induced seizures
CBD is approved as an adjunctive therapy in childhood epilepsy and is typically used in combination with clobazam (Devinsky et al., 2017). Indeed, CBD augments the anticonvulsant efficacy of clobazam (Anderson, Absalom, et al., 2019). Here, we assessed whether CBGA could similarly potentiate the anticonvulsant effects of clobazam in Scn1a +/− mice. The anticonvulsant effects of each drug and their combination were first assessed against hyperthermia‐induced seizures. Clobazam (1 mg·kg−1) and CBGA (30 mg·kg−1) administered individually significantly elevated the temperature threshold for GTCS compared with vehicle‐treated mice (Figure 3a). When administered in combination, the thermal threshold was elevated compared with either drug administered alone. A pharmacokinetic interaction was also observed whereby clobazam significantly increased the plasma concentration of CBGA during acute cotreatment (Figure 3b). CBGA co‐administration did not affect plasma clobazam concentrations but reduced plasma concentrations of its active metabolite, N‐desmethylclobazam (N‐CLB) (Figure S3).
FIGURE 3.
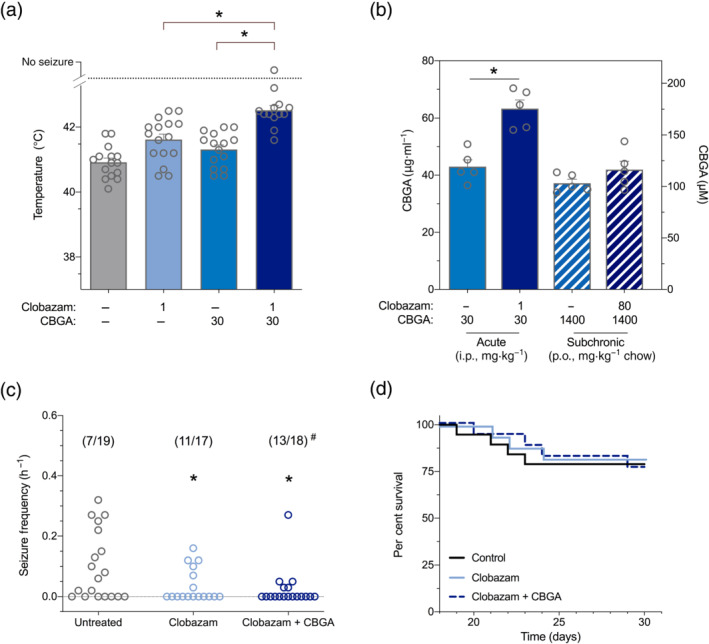
Cannabigerolic acid (CBGA) enhanced the anticonvulsant effect of clobazam against hyperthermia‐induced seizure in Scn1a +/− mice. (a) Temperature threshold for generalized tonic–clonic seizure (GTCS) induced by hyperthermia in individual mice following acute intraperitoneal treatment with CBGA (30 mg·kg−1, blue bar) or clobazam (1 mg·kg−1, light blue bar) administered individually or in combination (dark blue bar). All treatments resulted in a significantly improved response to thermal seizure induction compared with vehicle (P < 0.05; log‐rank Mantel–Cox). Combination CBGA and clobazam treatment was significantly more effective than either treatment alone, with n = 13–16 per group (* P < 0.05; log‐rank Mantel–Cox). (b) Average CBGA plasma concentrations in Scn1a +/− mice from hyperthermia‐induced seizure and spontaneous seizure experiments. CBGA was administered acutely (solid bars) or subchronically (hatched bars) in the presence (blue) and absence (dark blue) of clobazam, with n = 5 per treatment. Significantly higher CBGA plasma levels were observed following acute combination treatment with clobazam (* P < 0.05; Student's t‐test). CBGA concentrations are depicted as both mass concentrations (left Y axis) and molar concentrations (right Y axis). (c) Spontaneous GTCS frequency of individual untreated and drug‐treated Scn1a +/− mice. Treatments were administered orally via supplementation in chow and began at P18 following a hyperthermia‐induced seizure. Clobazam in both the absence and presence of CBGA significantly reduced the frequency of GTCS, with n = 17–19 per group (* P < 0.05; one‐way ANOVA followed by Dunnett's post hoc). Combination clobazam and CBGA treatment also significantly reduced the proportion of mice experiencing GTCS (# P < 0.05, Fisher's exact test). Untreated data are replotted from Figure 3c for clarity. (d) Survival curves comparing untreated and drug‐treated mice. Clobazam treatment alone or in combination with CBGA had no effect on survival, with n = 17–19 per group (log‐rank Mantel–Cox). Untreated data are replotted from Figure 2d for clarity. Additional data presented in Figure S3
We then sought to determine the effects of CBGA and clobazam combination treatment on spontaneous seizures and survival in Scn1a +/− mice. A CBGA dose (1400‐mg·kg−1 chow) was used which achieved steady‐state plasma concentrations equivalent to those found following a single 30 mg·kg−1 i.p. dose. As expected, clobazam treatment was anticonvulsant against spontaneous seizures, significantly reducing GTCS frequency (Figure 3c). Combination CBGA and clobazam treatment resulted in a lower mean GTCS frequency than controls and also significantly reduced the proportion (28%) of mice displaying GTCS compared with controls (63%). Clobazam treatment alone did not reduce the proportion of mice experiencing a GTCS (35%) compared with vehicle, although this group also did not differ from combination‐treated mice.
Despite clobazam treatment alone or in combination with CBGA being anticonvulsant against spontaneous seizures, neither treatment had any effect on survival (Figure 3d). In contrast to acute observations, subchronic cotreatment with clobazam had no effect on plasma CBGA concentrations (Figure 3b). However, plasma concentrations of N‐desmethylclobazam (N‐CLB) were significantly lower when clobazam was administered in combination with CBGA (Figure S3). In conclusion, CBGA enhanced the anticonvulsant effects of clobazam in the Scn1a +/− mouse model of Dravet syndrome.
3.4. CBGA exhibits divergent effects in conventional seizure models
To further investigate the anticonvulsant potential of CBGA, we evaluated its effects in conventional seizure models using wild‐type Swiss mice: the maximal electroshock (MES) threshold test model of GTCS and the 6‐Hz threshold test of focal, psychomotor seizures (Benson et al., 2020). Threshold tests were used where the CC50 (critical current at which 50% of mice exhibit a seizure) was determined. CBGA was anticonvulsant in the MES threshold test, where CBGA (30 mg·kg−1) significantly increased the CC50 compared with vehicle‐treated mice (Figure 4a). In contrast, CBGA was proconvulsant in the 6‐Hz threshold test (Figure 4b). The CC50 values for 10‐ and 30‐mg·kg−1 CBGA were significantly lower than that of vehicle‐treated controls, but 100‐mg·kg−1 CBGA did not differ from vehicle. By comparison, 300‐mg·kg−1 valproic acid yielded 100% protection against seizure activity induced by both MES and 6‐Hz threshold tests (n = 12 for each seizure model, data not graphed). The divergent properties of CBGA suggest that it may be useful against GTCS but perhaps contraindicated against focal seizures.
FIGURE 4.
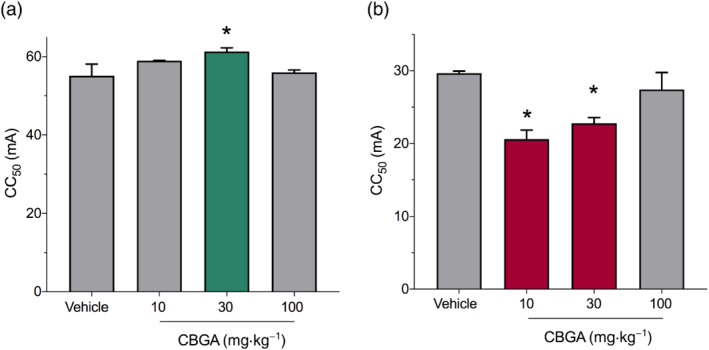
Cannabigerolic acid (CBGA) has divergent anticonvulsant and proconvulsant effects in conventional seizure models. (a) Dose–response of CBGA in the maximal electroshock (MES) acute seizure model. CBGA (30 mg·kg−1) was anticonvulsant and significantly increased the critical current (CC50) at which 50% of the mice exhibit a seizure with maximal hindlimb extension (green bar). Data are expressed as mean ± SEM, with n = 12 per treatment (* P < 0.05; one‐way ANOVA followed by Dunnett's post hoc). By comparison, 100% of mice treated with the positive control, valproic acid, were protected from hindlimb extension, with n = 12. (b) Dose–response of CBGA in the 6‐Hz seizure model. CBGA was proconvulsant with 10‐ and 30‐mg·kg−1 doses significantly reducing the CC50 for mice to have a psychomotor seizure (red bars). Data are expressed as mean + SEM, with n = 12 per treatment (* P < 0.05; one‐way ANOVA followed by Dunnett's post hoc). By comparison, 100% of mice treated with valproic acid were protected from psychomotor seizures, with n = 12
3.5. Molecular targets of CBGA
The major phytocannabinoids Δ9‐THC and CBD have been characterized as pharmacologically promiscuous, exerting effects on numerous targets and rendering mechanistic understanding of their therapeutic effects extremely challenging. Here, a Eurofins DiscoverX β‐arrestin signalling assay was used to screen the pharmacological actions of CBGA at GPCRs. No agonist or antagonist activity was identified for 3 μM CBGA at 242 GPCRs, which includes 74 orphan GPCRs. Of note, CBGA did not affect cannabinoid CB1 or CB2 receptors included in the panel, consistent with a prior report (Navarro et al., 2020).
The anticonvulsant mechanism of CBD is thought to reflect activity at numerous therapeutic targets, potentially including the GPCR 55 (GPR55), transient receptor potential cation channel V1 (TRPV1) and GABA (GABAA) receptors (Anderson, Absalom, et al., 2019; Chen et al., 2013; Gray & Whalley, 2020; Jia et al., 2015; Kaplan et al., 2017; Vilela et al., 2017). Therefore, the molecular actions of CBGA at these epilepsy‐relevant drug targets were explored.
In the Eurofins DiscoverX screen, CBGA was not an agonist at GPR55. However, no antagonist activity could be determined because GPR55 is classified as an orphan receptor with no endogenous agonist by Eurofins. We used HEK293 cells expressing human GPR55 to determine whether CBGA inhibits activation of GPR55 by the putative endogenous ligand lysophosphatidylinositol (LPI) and the selective agonist ML‐186. Concentration–response curves were generated for EC80 concentrations of ML‐186 (0.45 μM) and LPI (4 μM) in the presence of varying concentrations of CBGA (Figures 5a and S4A). CBGA inhibited ML‐186 and LPI‐mediated GPR55 activity with IC50 values of 4.8 and 5.7 μM, respectively. By comparison, IC50 values for inhibition of GPR55 activation by ML‐186 and LPI by the established GPR55 antagonist, CID16020046, were 238 and 303 nM, respectively (Figure S4B). The antagonist activity of CBGA was further characterized by generating concentration–response curves for ML‐186 in the presence of varying concentrations of CBGA (Figure 5b). CBGA appeared to be a non‐competitive antagonist of GPR55, with no effect on the potency of ML‐186 but significantly reducing its efficacy (Table S4).
FIGURE 5.
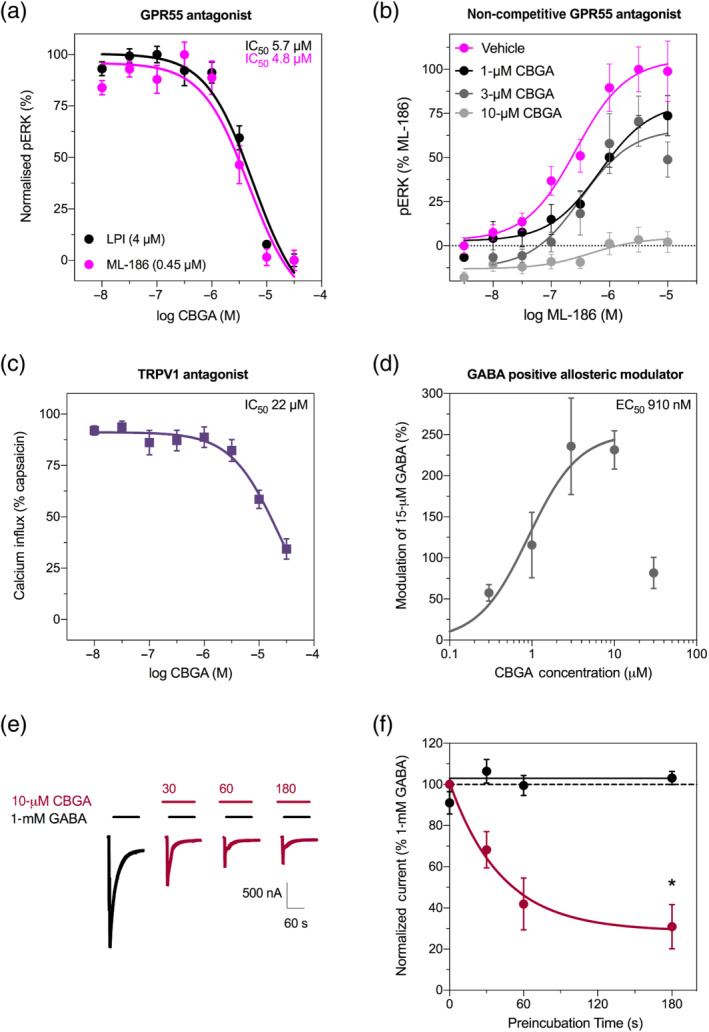
Cannabigerolic acid (CBGA) affects various epilepsy‐relevant drug targets in vitro. (a) Concentration–response for CBGA inhibition of GPR55 activation measured by ERK phosphorylation (pERK). GPR55 activation was measured following treatment with either LPI (black symbols) or ML‐186 (magenta symbols) in the presence of varying concentrations of CBGA and expressed as a percentage of responses to ligand alone. Data are expressed as mean ± SEM, with n = 6–7 per group. Curves represent fit to a three‐parameter log function and IC50 values are listed. (b) Concentration–response curves for activation of GPR55 by ML‐186 across varying concentrations of CBGA. Data are expressed as mean ± SEM, with n = 6 per group. Curve represents fit to the Hill equation. (c) Concentration–response for CBGA inhibition of TRPV1 determined by a FLIPR calcium influx assay to measure channel activity in cells expressing human TRPV1 channels. TRPV1 activity was measured following treatment with capsaicin (10 μM) in the presence of varying concentrations of CBGA and expressed as a percentage of capsaicin response. Data are expressed as mean ± SEM, with n = 4–8 per group. Curve represents fit to a three‐parameter log function. (d) Concentration–response curve for CBGA at GABAA receptors expressed in oocytes. CBGA modulates currents evoked by 15 μM GABA. Data are shown as mean ± SEM fit to the Hill equation, with n = 5 per group. (e) Representative trace currents evoked by GABA (1 mM) with (red) and without (black) preincubation of CBGA (10 μM) with varying preincubation times. (f) Time course of CBGA preincubation on GABAA receptor activity. Currents evoked by 1 mM GABA were measured following varying preincubation times with 10 μM CBGA (red symbols) and without (black symbols) and expressed as a percentage of GABA response. Dotted line represents no change in current. Data are shown as mean ± SEM, with n = 5 per group (* P < 0.05, Student's t‐test), and curve represents fit to a one‐phase decay. Additional data presented in Figures S4 and S5
Next, we examined the effect of CBGA on TRPV1 channel function. HEK293 cells expressing human TRPV1 channels were used to evaluate effects of CBGA on TRPV1 channel function. CBGA did not display agonist activity, but a non‐specific increase in intracellular calcium was observed at 30 and 100 μM of CBGA (Figure S5). Increasing concentrations of CBGA inhibited TRPV1 channel function with an IC50 value of 22 μM (9–123 μM, 95% confidence interval [CI]; Figure 5c).
CBD is a positive allosteric modulator (PAM) of GABAA receptors, so we explored whether CBGA exhibited similar activity (Anderson, Absalom, et al., 2019; Bakas et al., 2017). Using Xenopus oocytes expressing concatenated human GABAA receptors (γ2–β2–α1–β2–α1), a concentration–response curve of CBGA modulation of GABA‐evoked currents was generated (Figure 5d). CBGA increased the peak currents induced by GABA with a maximum modulation of 271% (E max 271 ± 40%) and a half maximal effective concentration (EC50) of 910 nM (log EC50 = −6.09 ± 0.14); however, the modulation was substantially reduced with 30 μM CBGA, so the Hill equation could not be fit when incorporating this high CBGA concentration. Some positive allosteric modulators of GABAA receptors also induce desensitization (Rosso et al., 2015). To determine whether CBGA induced desensitization, we preincubated the receptor with CBGA for 30–180 s before coapplication with GABA (Figure 5e,f). Peak GABA current was significantly reduced following 180‐s preincubation with CBGA (31 ± 11% compared with 103 ± 3% without preincubation; P < 0.05). Collectively, these data demonstrate that CBGA is an unconventional modulator of inhibitory GABAA receptors. Although CBGA enhances receptor activation at low GABA concentrations, prolonged exposure to CBGA at a higher concentration increased the desensitizing properties of the GABAA receptor. This suggests a potential liability of extended exposure to CBGA.
4. DISCUSSION
New therapies are urgently needed for treatment‐resistant epilepsies such as Dravet syndrome. The regulatory approval of CBD as an adjunctive treatment for Dravet syndrome has offered a new therapeutic avenue to patients, representing a modern continuation of the ancient practice of using hemp extracts to treat epilepsy (Friedman & Sirven, 2017). CBD is one of many possible anticonvulsant compounds in the cannabis plant and the current study continued the search for these constituents. Three phytocannabinoids, CBGA, CBDVA and CBGVA, had anticonvulsant effects on hyperthermia‐induced seizures in Scn1a +/− mice. The anticonvulsant activity of CBDV was also further confirmed (Amada et al., 2013; Hill et al., 2012, 2013). Interestingly, the three novel anticonvulsant phytocannabinoids uncovered here are all acid precursor molecules that are usually present in raw cannabis.
Commercially available cannabis‐based products rarely contain these phytocannabinoid acid precursors because manufacturing processes usually heat cannabis to decarboxylate the acidic compounds into neutral phytocannabinoids (e.g. CBGA to CBG). However, the artisanal cannabis oils used to treat children with intractable epilepsies are often cold extracted, which allows preservation of these phytocannabinoid acids (Huntsman et al., 2019; Suraev et al., 2018; Vincent et al., 2020). The current results suggest that the presence of CBGA, CBDVA and CBGVA, in addition to those identified previously CBDA, cannabichromenic acid (CBCA), cannabichromevarinic acid (CBCVA) and Δ9‐THCA, might contribute to the reported efficacy of such cold‐extracted artisanal preparations (Anderson et al., 2021; Anderson, Low, et al., 2019; Benson et al., 2020).
An extensive literature has explored the therapeutic potential of neutral phytocannabinoids Δ9‐THC and CBD. However, corresponding research on acidic precursor phytocannabinoids is in its infancy (Amin & Ali, 2019; Devinsky et al., 2014; Kis et al., 2019; Pisanti et al., 2017). CBDA and Δ9‐THCA are reported to have anti‐emetic, anxiolytic and anticonvulsant effects in animal models (Anderson, Low, et al., 2019; Assareh et al., 2020; Benson et al., 2020; Bolognini et al., 2013; Pertwee et al., 2018; Rock et al., 2017; Rock et al., 2018). CBCA and CBCVA displayed anticonvulsant effects against hyperthermia‐induced seizures in Scn1a +/− mice in our recent report (Anderson et al., 2021). The present findings of the anticonvulsant properties of CBGA, CBDVA and CBGA further support the consideration of phytocannabinoids in the discovery and development of novel cannabis‐based therapeutics. However, as phytocannabinoid acids tend to be relatively unstable and often exhibit poor brain penetration, bioisosteric replacement of the unstable acid moiety may provide one pathway for the development of these molecules towards the clinic.
The potency of CBDVA and CBGVA against hyperthermia‐induced seizures in the Scn1a +/− mouse model was similar to that previously reported with CBD, with effective doses of 100 mg·kg−1 (Anderson, Absalom, et al., 2019; Anderson et al., 2021; Kaplan et al., 2017). In contrast, CBGA achieved an anticonvulsant effect in this model at 30 mg·kg−1, suggesting greater potency than CBD and warranting further characterization of its anticonvulsant potential. This characterization provided a contrasting set of outcomes. CBGA showed further anticonvulsant properties in the MES threshold test and potentiated the anticonvulsant effects of clobazam against thermally induced seizures in Scn1a +/− mice. CBGA also appeared to modestly improve the effect of clobazam against spontaneous seizures in Scn1a +/− mice. However, CBGA as a monotherapy had proconvulsant effects on spontaneous seizures in Scn1a +/− mice and psychomotor seizures in the 6‐Hz threshold test. CBGA had no effect on survival of Scn1a +/− mice. The 78% survival of the untreated controls here is somewhat higher than what is usually observed for Scn1a +/− mice (Anderson, Absalom, et al., 2019; Anderson et al., 2020; Benson et al., 2020). Survival of Scn1a +/− mice is typically ~50% but has been known to vary between different laboratories and even between cohorts within the same laboratory under identical experimental conditions (Anderson, Absalom, et al., 2019; Anderson et al., 2020; Benson et al., 2020; Han et al., 2020; Hawkins et al., 2017; Miller et al., 2014; Mistry et al., 2014; Patra et al., 2020; Yu et al., 2006).
Divergent profiles across different preclinical seizure models are not unusual and underscore the importance of model choice when designing experiments to evaluate novel therapeutics for different epilepsy types. Lamotrigine, commonly prescribed for tonic–clonic and focal seizures, is anticonvulsant against MES‐induced seizures yet is proconvulsant in Scn1a +/− mice (Hawkins et al., 2017; Luszczki et al., 2003; Miller et al., 1986). Similarly, Δ9‐THCV was proconvulsant against hyperthermia‐induced seizures in the present study but showed anticonvulsant effects against pentylenetetrazole (PTZ)‐induced seizures previously (Hill et al., 2010). Additionally, co‐administration of CBD and Δ9‐THC is anticonvulsant against hyperthermia‐induced seizures but proconvulsant against spontaneous seizures in the Scn1a +/− mice (Anderson et al., 2020). These conflicting properties are not surprising as thermally induced seizures and spontaneous seizures have distinct neurochemical influences (Cheah et al., 2012; Kalume et al., 2007; Rubinstein et al., 2015; Stein et al., 2019). Further, in conventional seizure models, CBGA was anticonvulsant against MES‐induced seizures yet proconvulsant in the 6‐Hz seizure model.
Collectively, this indicates that CBGA may not possess the anticonvulsant versatility of CBD, which has shown consistent anticonvulsant activity across various animal models. In addition to CBD being anticonvulsant in Scn1a +/− mice, it is anticonvulsant in corneal kindled models and in the induced seizure models MES, 6‐Hz and PTZ (Anderson et al., 2020; Kaplan et al., 2017; Klein et al., 2017; Patra et al., 2020, 2019; Socała et al., 2019). Here, CBGA did not affect survival of Scn1a +/− mice similar to what we have observed previously with CBD (Anderson, Absalom, et al., 2019; Anderson et al., 2020). A different study did show survival of Scn1a +/− mice significantly improved with CBD treatment, however, a higher dose of CBD was administered (Patra et al., 2020). Overall, it appears that CBGA may not share the same versatility as CBD as an anticonvulsant despite having superior potency against hyperthermia‐induced seizures.
Although the pharmacological actions of the major phytocannabinoids CBD and Δ9‐THC are well understood, the pharmacological actions of the minor cannabinoids are poorly characterized. Here, we provide extensive characterization of the molecular targets of CBGA using both a commercially available screening platform (Eurofins DiscoverX) and several in‐house assays. We found CBGA to be relatively inactive, as it exhibited no agonist or antagonist activity at the 242 GPCRs screened. Although further assays interrogating epilepsy‐relevant targets for CBGA showed inhibition at GPR55 and TRPV1 channels, as well as an atypical positive allosteric modulator‐like profile on GABAA receptors. It is important to consider the concentrations of CBGA achieved in vivo when interpreting the relevance of the in vitro pharmacological action of CBGA at these molecular targets. Plasma concentrations of CBGA in the experimental animals used here were 36–202 μM (13–73 μg·ml−1). CBGA has been shown to have low brain penetrance with a brain–plasma ratio of 0.02 (Anderson, Low, et al., 2019). Based on this value of 2% brain penetration, the estimated concentrations of CBGA in the brains of the experimental mice would be 720 nM to 4 μM. CBGA inhibited GPR55 activation with an IC50 value of roughly 5 μM, so GPR55 inhibition could be contributing to the effects observed with the higher doses of CBGA. Although CBGA does inhibit TRPV1, the high IC50 value (22 μM) suggests that this target is likely less relevant to its anticonvulsant mechanism of action. The atypical positive allosteric modulator‐like profile of CBGA at GABAA receptors, however, could be quite relevant as the EC50 value of 910 nM is well within the therapeutic range of CBGA.
Whether these targets are relevant to the anticonvulsant and proconvulsant effects of CBGA requires further experiments. Additionally, the effects exerted by CBGA could result from pharmacological action of its metabolites rather than a direct effect of CBGA itself. Although the metabolic pathways of CBGA are unknown, CBD and Δ9‐THC are extensively metabolized by CYP450 metabolic enzymes and their metabolites appear to exert pharmacological effects including having anticonvulsant properties (dos Santos et al., 2015; Jiang et al., 2011; Nahas & Paton, 1979; Patilea‐Vrana & Unadkat, 2019; Stout & Cimino, 2014). Future studies could explore the metabolic profile of CBGA and characterize the pharmacological activity of its metabolites.
The effects of CBGA at this array of molecular targets are also observed for CBD, suggesting a possible overlap in mechanism of action between these two cannabinoids (Anderson, Absalom, et al., 2019; Bakas et al., 2017; Gray & Whalley, 2020; Kaplan et al., 2017; Vilela et al., 2017). CBD has been suggested to exert its anticonvulsant effects via inhibition of GPR55 (Gray & Whalley, 2020; Kaplan et al., 2017; Vilela et al., 2017). TRPV1 antagonists have anticonvulsant effects in animal models and the anticonvulsant effect of CBD against MES‐induced seizures is mediated by TRPV1 (Chen et al., 2013; Gonzalez‐Reyes et al., 2013; Gray et al., 2020; Iannotti et al., 2014; Jia et al., 2015; Vilela et al., 2017). Although our recent study in Scn1a +/− mice suggests that Trpv1 may not be disease modifying in Dravet syndrome (Satpute Janve et al., 2021).
Lastly, the divergent activity of CBGA on GABA modulation of GABAA receptors was striking. The actions of CBGA as a GABAA receptor positive allosteric modulator may contribute to the anticonvulsant effects of CBGA on hyperthermia‐induced seizures in Scn1a +/− mice and in the MES threshold test. Positive allosteric modulation at GABAA receptors could also account for the increased anticonvulsant effect observed with CBGA and clobazam co‐administration, similar to that previously observed between CBD and clobazam (Anderson, Absalom, et al., 2019). However, similar to the tolerance and GABAA receptor desensitization that is observed with chronic benzodiazepine treatment, prolonged exposure to CBGA resulted in GABAA receptor desensitization, which presents a potential liability with repeated CBGA treatment (Brown et al., 2002; Gallager et al., 1985; Mierlak & Farb, 1988; Nicholson et al., 2018). A previous study has shown that venoms from coral snakes, which are GABAA receptor positive allosteric modulators, also desensitized GABAA receptors in Xenopus oocytes (Rosso et al., 2015). The toxin‐induced desensitization of GABAA receptors correlated with increased neuronal activity and seizures in mice. We expect that GABAA receptor desensitization observed with prolonged CBGA exposure would similarly translate to mice in vivo. The proconvulsant effect of CBGA in the 6‐Hz threshold test and the increased spontaneous seizure frequency with subchronic CBGA treatment could be the result of GABAA receptor desensitization.
5. CONCLUSION
The current study shows for the first time that three phytocannabinoid acid constituents of the cannabis plant— cannabigerolic acid (CBGA), cannabidivarinic acid (CBDVA) and cannabigerovarinic acid (CBGVA)—have anticonvulsant effects against hyperthermia‐induced seizures in the Scn1a +/− mouse model of Dravet syndrome. This preclinical study confirms that the therapeutic actions of cannabis in epilepsy do not begin and end with CBD and that other phytocannabinoids might be considered in future drug discovery and development programmes. CBGA exhibited multimodal activity affecting several epilepsy‐relevant drug targets. Although CBGA shows promise, some proconvulsant effects were observed. Such liabilities would need to be overcome before another supersedes CBD in the new phytocannabinoid class of anticonvulsant drugs.
AUTHOR CONTRIBUTIONS
L.L.A., I.S.M., J.A.K. and J.C.A. contributed to the conception and design of the study. L.L.A., M.J.B., D.B., P.T.D., N.A.H., J.A.K. and J.C.A. contributed to the acquisition and analyses of the in vivo pharmacokinetic and anticonvulsant experiments. M.B., I.S.M. and J.C.A. designed the Eurofins DiscoverX screen. N.L.A., F.Z. and M.C. contributed to the acquisition and analyses of the GABAA receptor experiments. M.H., L.L.A. and J.C.A. contributed to the acquisition and analyses of the in vitro GPR55 experiments. All authors contributed to drafting the manuscript or figures.
CONFLICT OF INTEREST
J.C.A. has served as an expert witness in various medicolegal cases involving cannabis and cannabinoids and served as a temporary advisor to the World Health Organization (WHO) on their review of cannabis and the cannabinoids. I.S.M. is involved in an NHMRC‐funded clinical trial using the cannabis extract, nabiximols (Sativex®). He has served as an expert witness in various medicolegal cases involving cannabis and cannabinoids. J.C.A., L.L.A. and I.S.M. hold patents on cannabinoid therapies (PCT/AU2018/05089 and PCT/AU2019/050554). The remaining authors have no conflicts of interest.
DECLARATION OF TRANSPARENCY AND SCIENTIFIC RIGOUR
This Declaration acknowledges that this paper adheres to the principles for transparent reporting and scientific rigour of preclinical research as stated in the BJP guidelines for Design & Analysis and Animal Experimentation and as recommended by funding agencies, publishers and other organizations engaged with supporting research.
Supporting information
Figure S1. Pharmacokinetic analysis of CBGV. Concentration‐time curve for CBGV in mouse plasma following a 1 mg·kg‐1 i.p. injection. Data are expressed as means ± SEM, with n = 4–5 per time point.
Figure S2. Hyperthermia‐induced seizure screen. Temperature threshold for GTCS in individual mice treated acutely with (A) CBDV or THCV, (B) CBDVA, (C) CBG, (D) CBGA, (E) CBGV or (F) CBGVA. CBDV (30 mg·kg‐1), CBDVA (100 mg·kg‐1), CBGA (30 and 100 mg·kg‐1) and CBGVA (100 mg·kg‐1) treatments were anticonvulsant resulting in a significantly improved response to thermal seizure induction (blue‐green bars); whereas, THCV (3 mg·kg‐1) was proconvulsant and significantly lowered the thermal threshold (red bar). Error bars represent SEM, with n = 12–20 per group (*p < 0.05, **p < 0.01, ****p < 0.0001; logrank Mantel‐Cox).
Figure S3. Pharmacokinetic interaction between CBGA and N‐CLB, the active metabolite of clobazam. Average plasma concentrations of (a) clobazam and (b) N‐CLB in Scn1a +/− mice from hyperthermia‐induced seizure and spontaneous seizure experiments. Clobazam was administered acutely (solid bars) or subchronically (hatched bars) in the presence (dark green bars) and absence of CBGA (light green bars), with n = 5 per treatment. Significantly lower plasma N‐CLB levels were observed following combination treatment of CBGA and clobazam (*p < 0.05, **p < 0.01; Student's ttest).
Figure S4. In vitro characterization of GPR55 activity. HEK293 cells expressing human GPR55 were used to characterize inhibitory and agonist activity of compounds. ERK phosphorylation (pERK) was used as a measure of GPR55 activation. (a) Concentration‐response curves for activation of GPR55 by ML‐186 (blue symbols) and LPI (gray symbols). Data are expressed as mean ± SEM normalized to 100 nM phorbol 12‐myrisate 13‐acetate (PMA) and fit to the Hill equation, with n = 3–4 per concentration. (b) Concentration‐response for CID16020046 inhibition of GPR55 activation measured by ERK phosphorylation (pERK). GPR55 activation was measured following treatment with either LPI (gray symbols) or ML‐186 (blue symbols) in the presence of varying concentrations of CID16020046 and expressed as a percentage of responses to ligand alone. Data are expressed as mean ± SEM, with n = 6 per group. Curves represent fit to a three‐parameter log function and IC50 values are listed.
Figure S5. In vitro characterization of TRPV1 agonists. HEK293 cells expressing human
TRPV1 were used to characterize agonist activity of capsaicin and CBGA. Changes in intracellular calcium were used as a measure of TRPV1 activation. Concentration‐response curves for capsaicin (red) and CBGA (purple) in (a) wildtype and (b) TRPV1‐expressing cells. Data are expressed as mean ± SEM normalized to PAR‐1 and fit to the Hill equation, with n = 4–5 per group. The EC50 value of capsaicin for TRPV1 was 3.3 nM (95% Confidence Interval, 1.9–5.8 nM). Non‐specific calcium influx was observed with CBGA treatment (30 and 100 μM).
Table S1. Parameters for LC–MS/MS Detection of Phytocannabinoids
Table S2. Eurofins DiscoverX GPCR Screen
Table S3. Plasma cannabinoid concentrations in experimental Scn1a+/− mice
Table S4. Effect on GPR55 activity
ACKNOWLEDGEMENTS
The authors gratefully acknowledge Barry and Joy Lambert for their continued support of the Lambert Initiative for Cannabinoid Therapeutics. J.A.K. acknowledges the US National Institutes of Health (NIH) (R01 NS084959). M.C., J.C.A., J.A.K. and I.S.M. acknowledge the Australian National Health and Medical Research Council (NHMRC) (GNT 1161571 and 1185122). In addition, we thank Katelyn Lambert for inspiring our work on novel cannabinoid therapies for childhood epilepsy. We also thank Ivan Low, Charlotte Fletcher and Declan Everett‐Morgan for technical assistance.
Anderson, L. L. , Heblinski, M. , Absalom, N. L. , Hawkins, N. A. , Bowen, M. T. , Benson, M. J. , Zhang, F. , Bahceci, D. , Doohan, P. T. , Chebib, M. , McGregor, I. S. , Kearney, J. A. , & Arnold, J. C. (2021). Cannabigerolic acid, a major biosynthetic precursor molecule in cannabis, exhibits divergent effects on seizures in mouse models of epilepsy. British Journal of Pharmacology, 178(24), 4826–4841. 10.1111/bph.15661
Funding information National Health and Medical Research Council, Grant/Award Numbers: 1161571, 1185122; National Institutes of Health, Grant/Award Number: R01 NS084959; Lambert Initiative for Cannabinoid Therapeutics
DATA AVAILABILITY STATEMENT
The data that support the findings of this study are available from the corresponding author upon reasonable request.
REFERENCES
- Absalom, N. L. , Ahring, P. K. , Liao, V. W. , Balle, T. , Jiang, T. , Anderson, L. L. , Arnold, J. C. , McGregor, I. S. , Bowen, M. T. , & Chebib, M. (2019). Functional genomics of epilepsy‐associated mutations in the GABAA receptor subunits reveal that one mutation impairs function and two are catastrophic. The Journal of Biological Chemistry, 294, 6157–6171. 10.1074/jbc.RA118.005697 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander, S. P. H. , Christopoulos, A. , Davenport, A. P. , Kelly, E. , Mathie, A. , Peters, J. A. , Veale, E. L. , Armstrong, J. F. , Faccenda, E. , Harding, S. D. , Pawson, A. J. , Sharman, J. L. , Southan, C. , Davies, J. A. , Abbracchio, M. P. , Alexander, W. , Al‐hosaini, K. , Bäck, M. , Beaulieu, J. , … Yao, C. (2019). The Concise Guide to PHARMACOLOGY 2019/20: G protein‐coupled receptors. British Journal of Pharmacology, 176, S21–S141. 10.1111/bph.14748 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amada, N. , Yamasaki, Y. , Williams, C. M. , & Whalley, B. J. (2013). Cannabidivarin (CBDV) suppresses pentylenetetrazole (PTZ)‐induced increases in epilepsy‐related gene expression. PeerJ, 1, e214. 10.7717/peerj.214 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amin, M. R. , & Ali, D. W. (2019). Pharmacology of medical cannabis. In Advances in experimental medicine and biology. Adv Exp Med Biol. (pp. 151–165). 10.1007/978-3-030-21737-2_8 [DOI] [PubMed] [Google Scholar]
- Anderson, L. L. , Absalom, N. L. , Abelev, S. V. , Low, I. K. , Doohan, P. T. , Martin, L. J. , Chebib, M. , McGregor, I. S. , & Arnold, J. C. (2019). Coadministered cannabidiol and clobazam: Preclinical evidence for both pharmacodynamic and pharmacokinetic interactions. Epilepsia, 60, 2224–2234. 10.1111/epi.16355 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson, L. L. , Ametovski, A. , Lin Luo, J. , Everett‐Morgan, D. , McGregor, I. S. , Banister, S. D. , & Arnold, J. C. (2021). Cannabichromene, related phytocannabinoids, and 5‐fluoro‐cannabichromene have anticonvulsant properties in a mouse model of Dravet syndrome. ACS Chemical Neuroscience, 12, 330–339. 10.1021/acschemneuro.0c00677 [DOI] [PubMed] [Google Scholar]
- Anderson, L. L. , Low, I. K. , Banister, S. D. , McGregor, I. S. , & Arnold, J. C. (2019). Pharmacokinetics of phytocannabinoid acids and anticonvulsant effect of cannabidiolic acid in a mouse model of Dravet syndrome. Journal of Natural Products, 82, 3047–3055. 10.1021/acs.jnatprod.9b00600 [DOI] [PubMed] [Google Scholar]
- Anderson, L. L. , Low, I. K. , McGregor, I. S. , & Arnold, J. C. (2020). Interactions between cannabidiol and Δ9‐tetrahydrocannabinol in modulating seizure susceptibility and survival in a mouse model of Dravet syndrome. British Journal of Pharmacology, 177, 4261–4274. 10.1111/bph.15181 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Assareh, N. , Gururajan, A. , Zhou, C. , Luo, J. L. , Kevin, R. C. , & Arnold, J. C. (2020). Cannabidiol disrupts conditioned fear expression and cannabidiolic acid reduces trauma‐induced anxiety‐related behaviour in mice. Behavioural Pharmacology, 31, 591–596. 10.1097/FBP.0000000000000565 [DOI] [PubMed] [Google Scholar]
- Bakas, T. , van Nieuwenhuijzen, P. S. , Devenish, S. O. , McGregor, I. S. , Arnold, J. C. , & Chebib, M. (2017). The direct actions of cannabidiol and 2‐arachidonoyl glycerol at GABAA receptors. Pharmacological Research, 119, 358–370. 10.1016/j.phrs.2017.02.022 [DOI] [PubMed] [Google Scholar]
- Benson, M. J. , Anderson, L. L. , Low, I. K. , Luo, J. L. , Kevin, R. C. , Zhou, C. , McGregor, I. S. , & Arnold, J. C. (2020). Evaluation of the possible anticonvulsant effect of Δ9‐tetrahydrocannabinolic acid in murine seizure models. Cannabis Cannabinoid Res, can.2020.0073. 10.1089/can.2020.0073 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolognini, D. , Rock, E. , Cluny, N. , Cascio, M. , Limebeer, C. , Duncan, M. , Stott, C. , Javid, F. , Parker, L. , & Pertwee, R. (2013). Cannabidiolic acid prevents vomiting in Suncus murinus and nausea‐induced behaviour in rats by enhancing 5‐HT1A receptor activation. British Journal of Pharmacology, 168, 1456–1470. 10.1111/bph.12043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown, M. J. , Wood, M. D. , Coldwell, M. C. , & Bristow, D. R. (2002). γ‐Aminobutyric acidA receptor function is desensitised in rat cultured cerebellar granule cells following chronic flunitrazepam treatment. Journal of Neurochemistry, 71, 1232–1240. 10.1046/j.1471-4159.1998.71031232.x [DOI] [PubMed] [Google Scholar]
- Cheah, C. S. , Yu, F. H. , Westenbroek, R. E. , Kalume, F. K. , Oakley, J. C. , Potter, G. B. , Rubenstein, J. L. , & Catterall, W. A. (2012). Specific deletion of NaV1.1 sodium channels in inhibitory interneurons causes seizures and premature death in a mouse model of Dravet syndrome. Proceedings of the National Academy of Sciences, 109, 14646–14651. 10.1073/pnas.1211591109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, C.‐Y. , Li, W. , Qu, K.‐P. , & Chen, C.‐R. (2013). Piperine exerts anti‐seizure effects via the TRPV1 receptor in mice. European Journal of Pharmacology, 714, 288–294. 10.1016/j.ejphar.2013.07.041 [DOI] [PubMed] [Google Scholar]
- Chiu, P. , Olsen, D. M. , Borys, H. K. , Karler, R. , & Turkanis, S. A. (1979). The influence of cannabidiol and Δ9‐tetrahydrocannabinol on cobalt epilepsy in rats. Epilepsia, 20, 365–375. 10.1111/j.1528-1157.1979.tb04816.x [DOI] [PubMed] [Google Scholar]
- Curtis, M. J. , Alexander, S. , Cirino, G. , Docherty, J. R. , George, C. H. , Giembycz, M. A. , Hoyer, D. , Insel, P. A. , Izzo, A. A. , Ji, Y. , MacEwan, D. J. , Sobey, C. G. , Stanford, S. C. , Teixeira, M. M. , Wonnacott, S. , & Ahluwalia, A. (2018). Experimental design and analysis and their reporting II: Updated and simplified guidance for authors and peer reviewers. British Journal of Pharmacology, 175(7), 987–993. 10.1111/bph.14153 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deiana, S. , Watanabe, A. , Yamasaki, Y. , Amada, N. , Arthur, M. , Fleming, S. , Woodcock, H. , Dorward, P. , Pigliacampo, B. , Close, S. , Platt, B. , & Riedel, G. (2012). Plasma and brain pharmacokinetic profile of cannabidiol (CBD), cannabidivarine (CBDV), Δ9‐tetrahydrocannabivarin (THCV) and cannabigerol (CBG) in rats and mice following oral and intraperitoneal administration and CBD action on obsessive–compulsive behavi. Psychopharmacology, 219, 859–873. 10.1007/s00213-011-2415-0 [DOI] [PubMed] [Google Scholar]
- Devinsky, O. , Cilio, M. R. , Cross, H. , Fernandez‐Ruiz, J. , French, J. , Hill, C. , Katz, R. , Di Marzo, V. , Jutras‐Aswad, D. , Notcutt, W. G. , Martinez‐Orgado, J. , Robson, P. J. , Rohrback, B. G. , Thiele, E. , Whalley, B. , & Friedman, D. (2014). Cannabidiol: Pharmacology and potential therapeutic role in epilepsy and other neuropsychiatric disorders. Epilepsia, 55, 791–802. 10.1111/epi.12631 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devinsky, O. , Cross, J. H. , Laux, L. , Marsh, E. , Miller, I. , Nabbout, R. , Scheffer, I. E. , Thiele, E. A. , Wright, S. , & Cannabidiol in Dravet Syndrome Study Group . (2017). Trial of cannabidiol for drug‐resistant seizures in the Dravet syndrome. The New England Journal of Medicine, 376, 2011–2020. 10.1056/NEJMoa1611618 [DOI] [PubMed] [Google Scholar]
- Devinsky, O. , Patel, A. D. , Cross, J. H. , Villanueva, V. , Wirrell, E. C. , Privitera, M. , Greenwood, S. M. , Roberts, C. , Checketts, D. , VanLandingham, K. E. , & Zuberi, S. M. (2018). Effect of cannabidiol on drop seizures in the Lennox–Gastaut syndrome. The New England Journal of Medicine, 378, 1888–1897. 10.1056/NEJMoa1714631 [DOI] [PubMed] [Google Scholar]
- dos Santos, R. G. , Hallak, J. E. C. , Leite, J. P. , Zuardi, A. W. , & Crippa, J. A. S. (2015). Phytocannabinoids and epilepsy. Journal of Clinical Pharmacy and Therapeutics, 40, 135–143. 10.1111/jcpt.12235 [DOI] [PubMed] [Google Scholar]
- Friedman, D. , & Sirven, J. I. (2017). Historical perspective on the medical use of cannabis for epilepsy: Ancient times to the 1980s. Epilepsy & Behavior, 70, 298–301. 10.1016/j.yebeh.2016.11.033 [DOI] [PubMed] [Google Scholar]
- Gallager, D. W. , Malcolm, A. B. , Anderson, S. A. , & Gonsalvez, S. F. (1985). Continuous release of diazepam: Electrophysiological, biochemical and behavioral consequences. Brain Research, 342, 26–36. 10.1016/0006-8993(85)91349-6 [DOI] [PubMed] [Google Scholar]
- Gonzalez‐Reyes, L. E. , Ladas, T. P. , Chiang, C.‐C. , & Durand, D. M. (2013). TRPV1 antagonist capsazepine suppresses 4‐AP‐induced epileptiform activity in vitro and electrographic seizures in vivo. Experimental Neurology, 250, 321–332. 10.1016/j.expneurol.2013.10.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray, R. A. , Stott, C. G. , Jones, N. A. , Di Marzo, V. , & Whalley, B. J. (2020). Anticonvulsive properties of cannabidiol in a model of generalized seizure are transient receptor potential vanilloid 1 dependent. Cannabis and Cannabinoid Research, 5, 145–149. 10.1089/can.2019.0028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray, R. A. , & Whalley, B. J. (2020). The proposed mechanisms of action of CBD in epilepsy. Epileptic Disorders, 22, S10–S15. 10.1684/epd.2020.1135 [DOI] [PubMed] [Google Scholar]
- Han, Z. , Chen, C. , Christiansen, A. , Ji, S. , Lin, Q. , Anumonwo, C. , Liu, C. , Leiser, S. C. , Aznarez, I. , Liau, G. , & Isom, L. L. (2020). Antisense oligonucleotides increase Scn1a expression and reduce seizures and SUDEP incidence in a mouse model of Dravet syndrome. Science Translational Medicine, 12, eaaz6100. 10.1126/scitranslmed.aaz6100 [DOI] [PubMed] [Google Scholar]
- Hawkins, N. A. , Anderson, L. L. , Gertler, T. S. , Laux, L. , George, A. L. , & Kearney, J. A. (2017). Screening of conventional anticonvulsants in a genetic mouse model of epilepsy. Annals of Clinical Translational Neurology, 4, 326–339. 10.1002/acn3.413 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heblinski, M. , Santiago, M. , Fletcher, C. , Stuart, J. , Connor, M. , McGregor, I. S. , & Arnold, J. C. (2020). Terpenoids commonly found in Cannabis sativa do not modulate the actions of phytocannabinoids or endocannabinoids on TRPA1 and TRPV1 channels. Cannabis and Cannabinoid Research, 5, 305–317. 10.1089/can.2019.0099 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill, A. , Mercier, M. , Hill, T. , Glyn, S. , Jones, N. , Yamasaki, Y. , Futamura, T. , Duncan, M. , Stott, C. , Stephens, G. , Williams, C. , & Whalley, B. (2012). Cannabidivarin is anticonvulsant in mouse and rat. British Journal of Pharmacology, 167, 1629–1642. 10.1111/j.1476-5381.2012.02207.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill, A. J. , Weston, S. E. , Jones, N. A. , Smith, I. , Bevan, S. A. , Williamson, E. M. , Stephens, G. J. , Williams, C. M. , & Whalley, B. J. (2010). Δ9‐Tetrahydrocannabivarin suppresses in vitro epileptiform and in vivo seizure activity in adult rats. Epilepsia, 51, 1522–1532. 10.1111/j.1528-1167.2010.02523.x [DOI] [PubMed] [Google Scholar]
- Hill, T. D. M. , Cascio, M. G. , Romano, B. , Duncan, M. , Pertwee, R. G. , Williams, C. M. , Whalley, B. J. , & Hill, A. J. (2013). Cannabidivarin‐rich cannabis extracts are anticonvulsant in mouse and rat via a CB1 receptor‐independent mechanism. British Journal of Pharmacology, 170, 679–692. 10.1111/bph.12321 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huntsman, R. J. , Tang‐Wai, R. , Alcorn, J. , Vuong, S. , Acton, B. , Corley, S. , Laprairie, R. , Lyon, A. W. , Meier, S. , Mousseau, D. D. , Newmeyer, D. , Prosser‐Loose, E. , Seifert, B. , Tellez‐Zenteno, J. , Huh, L. , Leung, E. , & Major, P. (2019). Dosage related efficacy and tolerability of cannabidiol in children with treatment‐resistant epileptic encephalopathy: Preliminary results of the CARE‐E study. Frontiers in Neurology, 10, 716. 10.3389/fneur.2019.00716 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iannotti, F. A. , Hill, C. L. , Leo, A. , Alhusaini, A. , Soubrane, C. , Mazzarella, E. , Russo, E. , Whalley, B. J. , Di Marzo, V. , & Stephens, G. J. (2014). Nonpsychotropic plant cannabinoids, cannabidivarin (CBDV) and cannabidiol (CBD), activate and desensitize transient receptor potential vanilloid 1 (TRPV1) channels in vitro: Potential for the treatment of neuronal hyperexcitability. ACS Chemical Neuroscience, 5, 1131–1141. 10.1021/cn5000524 [DOI] [PubMed] [Google Scholar]
- Jia, Y.‐F. , Li, Y.‐C. , Tang, Y.‐P. , Cao, J. , Wang, L.‐P. , Yang, Y.‐X. , Xu, L. , & Mao, R.‐R. (2015). Interference of TRPV1 function altered the susceptibility of PTZ‐induced seizures. Frontiers in Cellular Neuroscience, 9. 10.3389/fncel.2015.00020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang, R. , Yamaori, S. , Takeda, S. , Yamamoto, I. , & Watanabe, K. (2011). Identification of cytochrome P450 enzymes responsible for metabolism of cannabidiol by human liver microsomes. Life Sciences, 89, 165–170. 10.1016/j.lfs.2011.05.018 [DOI] [PubMed] [Google Scholar]
- Kalume, F. , Yu, F. H. , Westenbroek, R. E. , Scheuer, T. , & Catterall, W. A. (2007). Reduced sodium current in Purkinje neurons from NaV1.1 mutant mice: Implications for ataxia in severe myoclonic epilepsy in infancy. The Journal of Neuroscience, 27, 11065–11074. 10.1523/JNEUROSCI.2162-07.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaplan, J. S. , Stella, N. , Catterall, W. A. , & Westenbroek, R. E. (2017). Cannabidiol attenuates seizures and social deficits in a mouse model of Dravet syndrome. Proceedings of the National Academy of Sciences, 114, 11229–11234. 10.1073/pnas.1711351114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kargl, J. , Balenga, N. , Parzmair, G. P. , Brown, A. J. , Heinemann, A. , & Waldhoer, M. (2012). The cannabinoid receptor CB1 modulates the signaling properties of the lysophosphatidylinositol receptor GPR55. The Journal of Biological Chemistry, 287, 44234–44248. 10.1074/jbc.M112.364109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karler, R. , Cely, W. , & Turkanis, S. A. (1973). The anticonvulsant activity of cannabidiol and cannabinol. Life Sciences, 13, 1527–1531. 10.1016/0024-3205(73)90141-0 [DOI] [PubMed] [Google Scholar]
- Karler, R. , & Turkanis, S. (1979). Cannabis and epilepsy. In Marihuana biological effects (pp. 619–641). Elsevier. 10.1016/B978-0-08-023759-6.50052-4 [DOI] [PubMed] [Google Scholar]
- Kimball, A. W. , Burnett, W. T. , & Doherty, D. G. (1957). Chemical protection against ionizing radiation: I. Sampling methods for screening compounds in radiation protection studies with mice. Radiation Research, 7, 1–12. 10.2307/3570549 [DOI] [PubMed] [Google Scholar]
- Kis, B. , Ifrim, F. C. , Buda, V. , Avram, S. , Pavel, I. Z. , Antal, D. , Paunescu, V. , Dehelean, C. A. , Ardelean, F. , Diaconeasa, Z. , Soica, C. , & Danciu, C. (2019). Cannabidiol—From plant to human body: A promising bioactive molecule with multi‐target effects in cancer. International Journal of Molecular Sciences, 20, 5905. 10.3390/ijms20235905 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klein, B. D. , Jacobson, C. A. , Metcalf, C. S. , Smith, M. D. , Wilcox, K. S. , Hampson, A. J. , & Kehne, J. H. (2017). Evaluation of cannabidiol in animal seizure models by the Epilepsy Therapy Screening Program (ETSP). Neurochemical Research, 42, 1939–1948. 10.1007/s11064-017-2287-8 [DOI] [PubMed] [Google Scholar]
- Liao, V. W. Y. , Chua, H. C. , Kowal, N. M. , Chebib, M. , Balle, T. , & Ahring, P. K. (2019). Concatenated γ‐aminobutyric acid type A receptors revisited: Finding order in chaos. The Journal of General Physiology, 151, 798–819. 10.1085/jgp.201812133 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lilley, E. , Stanford, S. C. , Kendall, D. E. , Alexander, S. P. , Cirino, G. , Docherty, J. R. , George, C. H. , Insel, P. A. , Izzo, A. A. , Ji, Y. , Panettieri, R. A. , Sobey, C. G. , Stefanska, B. , Stephens, G. , Teixeira, M. , & Ahluwalia, A. (2020). ARRIVE 2.0 and the British Journal of Pharmacology: Updated guidance for 2020. British Journal of Pharmacology, 177(16), 3611–3616. 10.1111/bph.15178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luszczki, J. J. , Czuczwar, M. , Kis, J. , Krysa, J. , Pasztelan, I. , Swiader, M. , & Czuczwar, S. J. (2003). Interactions of lamotrigine with topiramate and first‐generation antiepileptic drugs in the maximal electroshock test in mice: An isobolographic analysis. Epilepsia, 44, 1003–1013. 10.1046/j.1528-1157.2003.10003.x [DOI] [PubMed] [Google Scholar]
- Marini, C. , Scheffer, I. E. , Nabbout, R. , Suls, A. , De Jonghe, P. , Zara, F. , & Guerrini, R. (2011). The genetics of Dravet syndrome. Epilepsia, 52, 24–29. 10.1111/j.1528-1167.2011.02997.x [DOI] [PubMed] [Google Scholar]
- Mierlak, D. , & Farb, D. (1988). Modulation of neurotransmitter receptor desensitization: Chlordiazepoxide stimulates fading of the GABA response. The Journal of Neuroscience, 8, 814–820. 10.1523/JNEUROSCI.08-03-00814.1988 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller, A. A. , Wheatley, P. , Sawyer, D. A. , Baxter, M. G. , & Roth, B. (1986). Pharmacological studies on lamotrigine, a novel potential antiepileptic drug. Epilepsia, 27, 483–489. 10.1111/j.1528-1157.1986.tb03572.x [DOI] [PubMed] [Google Scholar]
- Miller, A. R. , Hawkins, N. A. , McCollom, C. E. , & Kearney, J. A. (2014). Mapping genetic modifiers of survival in a mouse model of Dravet syndrome. Genes, Brain, and Behavior, 13, 163–172. 10.1111/gbb.12099 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mistry, A. M. , Thompson, C. H. , Miller, A. R. , Vanoye, C. G. , George, A. L. , & Kearney, J. A. (2014). Strain‐ and age‐dependent hippocampal neuron sodium currents correlate with epilepsy severity in Dravet syndrome mice. Neurobiology of Disease, 65, 1–11. 10.1016/j.nbd.2014.01.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nahas, G. G. , & Paton, W. D. M. (1979). International Congress of Pharmacology (7th: 1978: Paris, France). In Marihuana biological effects: Analysis, metabolism, cellular responses, reproduction and brain. Pergamon Press. [PubMed] [Google Scholar]
- Navarro, G. , Varani, K. , Lillo, A. , Vincenzi, F. , Rivas‐Santisteban, R. , Raïch, I. , Reyes‐Resina, I. , Ferreiro‐Vera, C. , Borea, P. A. , Sánchez de Medina, V. , Nadal, X. , & Franco, R. (2020). Pharmacological data of cannabidiol‐ and cannabigerol‐type phytocannabinoids acting on cannabinoid CB1, CB2 and CB1/CB2 heteromer receptors. Pharmacological Research, 159, 104940. 10.1016/j.phrs.2020.104940 [DOI] [PubMed] [Google Scholar]
- Nicholson, M. W. , Sweeney, A. , Pekle, E. , Alam, S. , Ali, A. B. , Duchen, M. , & Jovanovic, J. N. (2018). Diazepam‐induced loss of inhibitory synapses mediated by PLCδ/Ca2+/calcineurin signalling downstream of GABAA receptors. Molecular Psychiatry, 23, 1851–1867. 10.1038/s41380-018-0100-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pamplona, F. A. , da Silva, L. R. , & Coan, A. C. (2018). Potential clinical benefits of CBD‐rich cannabis extracts over purified CBD in treatment‐resistant epilepsy: Observational data meta‐analysis. Frontiers in Neurology, 9. 10.3389/fneur.2018.00759 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patilea‐Vrana, G. I. , & Unadkat, J. D. (2019). Quantifying hepatic enzyme kinetics of (−)‐∆9‐tetrahydrocannabinol (THC) and its psychoactive metabolite, 11‐OH‐THC, through in vitro modeling. Drug Metabolism and Disposition, 47, 743–752. 10.1124/dmd.119.086470 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patra, P. H. , Barker‐Haliski, M. , White, H. S. , Whalley, B. J. , Glyn, S. , Sandhu, H. , Jones, N. , Bazelot, M. , Williams, C. M. , & McNeish, A. J. (2019). Cannabidiol reduces seizures and associated behavioral comorbidities in a range of animal seizure and epilepsy models. Epilepsia, 60, 303–314. 10.1111/epi.14629 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patra, P. H. , Serafeimidou‐Pouliou, E. , Bazelot, M. , Whalley, B. J. , Williams, C. M. , & McNeish, A. J. (2020). Cannabidiol improves survival and behavioural co‐morbidities of Dravet syndrome in mice. British Journal of Pharmacology, 177, 2779–2792. 10.1111/bph.15003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Percie du Sert, N. , Hurst, V. , Ahluwalia, A. , Alam, S. , Avey, M. T. , Baker, M. , Browne, W. J. , Clark, A. , Cuthill, I. C. , Dirnagl, U. , Emerson, M. , Garner, P. , Holgate, S. T. , Howells, D. W. , Karp, N. A. , Lazic, S. E. , Lidster, K. , MacCallum, C. J. , Macleod, M. , … Würbel, H. (2020). The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biology, 18(7), e3000410. 10.1371/journal.pbio.3000410 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pertwee, R. G. , Rock, E. M. , Guenther, K. , Limebeer, C. L. , Stevenson, L. A. , Haj, C. , Smoum, R. , Parker, L. A. , & Mechoulam, R. (2018). Cannabidiolic acid methyl ester, a stable synthetic analogue of cannabidiolic acid, can produce 5‐HT1A receptor‐mediated suppression of nausea and anxiety in rats. British Journal of Pharmacology, 175, 100–112. 10.1111/bph.14073 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pertwee, R. G. , Thomas, A. , Stevenson, L. A. , Ross, R. A. , Varvel, S. A. , Lichtman, A. H. , Martin, B. R. , & Razdan, R. K. (2007). The psychoactive plant cannabinoid, Δ9‐tetrahydrocannabinol, is antagonized by Δ8‐ and Δ9‐tetrahydrocannabivarin in mice in vivo. British Journal of Pharmacology, 150, 586–594. 10.1038/sj.bjp.0707124 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pisanti, S. , Malfitano, A. M. , Ciaglia, E. , Lamberti, A. , Ranieri, R. , Cuomo, G. , Abate, M. , Faggiana, G. , Proto, M. C. , Fiore, D. , Laezza, C. , & Bifulco, M. (2017). Cannabidiol: State of the art and new challenges for therapeutic applications. Pharmacology & Therapeutics, 175, 133–150. 10.1016/j.pharmthera.2017.02.041 [DOI] [PubMed] [Google Scholar]
- Rock, E. M. , Limebeer, C. L. , & Parker, L. A. (2018). Effect of cannabidiolic acid and ∆9‐tetrahydrocannabinol on carrageenan‐induced hyperalgesia and edema in a rodent model of inflammatory pain. Psychopharmacology, 235, 3259–3271. 10.1007/s00213-018-5034-1 [DOI] [PubMed] [Google Scholar]
- Rock, E. M. , Limebeer, C. L. , Petrie, G. N. , Williams, L. A. , Mechoulam, R. , & Parker, L. A. (2017). Effect of prior foot shock stress and Δ9‐tetrahydrocannabinol, cannabidiolic acid, and cannabidiol on anxiety‐like responding in the light‐dark emergence test in rats. Psychopharmacology, 234, 2207–2217. 10.1007/s00213-017-4626-5 [DOI] [PubMed] [Google Scholar]
- Rosso, J. P. , Schwarz, J. R. , Diaz‐Bustamante, M. , Céard, B. , Gutiérrez, J. M. , Kneussel, M. , Pongs, O. , Bosmans, F. , & Bougis, P. E. (2015). MmTX1 and MmTX2 from coral snake venom potently modulate GABAA receptor activity. Proceedings of the National Academy of Sciences of the United States of America, 112, E891–E900. 10.1073/pnas.1415488112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rubinstein, M. , Han, S. , Tai, C. , Westenbroek, R. E. , Hunker, A. , Scheuer, T. , & Catterall, W. A. (2015). Dissecting the phenotypes of Dravet syndrome by gene deletion. Brain, 138, 2219–2233. 10.1093/brain/awv142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satpute Janve, V. , Anderson, L. L. , Bahceci, D. , Hawkins, N. A. , Kearney, J. A. , & Arnold, J. C. (2021). The heat sensing Trpv1 receptor is not a viable anticonvulsant drug target in the Scn1a +/− mouse model of Dravet syndrome. Frontiers in Pharmacology, 12. 10.3389/fphar.2021.675128 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Socała, K. , Wyska, E. , Szafarz, M. , Nieoczym, D. , & Wlaź, P. (2019). Acute effect of cannabidiol on the activity of various novel antiepileptic drugs in the maximal electroshock‐ and 6 Hz‐induced seizures in mice: Pharmacodynamic and pharmacokinetic studies. Neuropharmacology, 158, 107733. 10.1016/j.neuropharm.2019.107733 [DOI] [PubMed] [Google Scholar]
- Stein, R. E. , Kaplan, J. S. , Li, J. , & Catterall, W. A. (2019). Hippocampal deletion of NaV1.1 channels in mice causes thermal seizures and cognitive deficit characteristic of Dravet syndrome. Proceedings of the National Academy of Sciences, 116, 16571–16576. 10.1073/pnas.1906833116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stout, S. M. , & Cimino, N. M. (2014). Exogenous cannabinoids as substrates, inhibitors, and inducers of human drug metabolizing enzymes: A systematic review. Drug Metabolism Reviews, 46, 86–95. 10.3109/03602532.2013.849268 [DOI] [PubMed] [Google Scholar]
- Suraev, A. , Lintzeris, N. , Stuart, J. , Kevin, R. C. , Blackburn, R. , Richards, E. , Arnold, J. C. , Ireland, C. , Todd, L. , Allsop, D. J. , & McGregor, I. S. (2018). Composition and use of cannabis extracts for childhood epilepsy in the Australian community. Scientific Reports, 8, 10154. 10.1038/s41598-018-28127-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thiele, E. A. , Marsh, E. D. , French, J. A. , Mazurkiewicz‐Beldzinska, M. , Benbadis, S. R. , Joshi, C. , Lyons, P. D. , Taylor, A. , Roberts, C. , Sommerville, K. , Gunning, B. , Gawlowicz, J. , Lisewski, P. , Mazurkiewicz Beldzinska, M. , Mitosek Szewczyk, K. , Steinborn, B. , Zolnowska, M. , Hughes, E. , McLellan, A. , … Wilfong, A. (2018). Cannabidiol in patients with seizures associated with Lennox‐Gastaut syndrome (GWPCARE4): A randomised, double‐blind, placebo‐controlled phase 3 trial. Lancet, 391, 1085–1096. 10.1016/S0140-6736(18)30136-3 [DOI] [PubMed] [Google Scholar]
- Tzadok, M. , Uliel‐Siboni, S. , Linder, I. , Kramer, U. , Epstein, O. , Menascu, S. , Nissenkorn, A. , Yosef, O. B. , Hyman, E. , Granot, D. , Dor, M. , Lerman‐Sagie, T. , & Ben‐Zeev, B. (2016). CBD‐enriched medical cannabis for intractable pediatric epilepsy. Seizure, 35, 41–44. 10.1016/j.seizure.2016.01.004 [DOI] [PubMed] [Google Scholar]
- Vilela, L. R. , Lima, I. V. , Kunsch, É. B. , Pinto, H. P. P. , de Miranda, A. S. , Vieira, É. L. M. , de Oliveira, A. C. P. , Moraes, M. F. D. , Teixeira, A. L. , & Moreira, F. A. (2017). Anticonvulsant effect of cannabidiol in the pentylenetetrazole model: Pharmacological mechanisms, electroencephalographic profile, and brain cytokine levels. Epilepsy & Behavior, 75, 29–35. 10.1016/j.yebeh.2017.07.014 [DOI] [PubMed] [Google Scholar]
- Vincent, D. , Savin, K. , Rochfort, S. , & Spangenberg, G. (2020). The power of three in cannabis shotgun proteomics: Proteases, databases and search engines. Proteomes, 8, 13. 10.3390/proteomes8020013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamakawa, K. (2009). Molecular basis of severe myoclonic epilepsy in infancy. Brain & Development, 31, 401–404. 10.1016/j.braindev.2008.11.015 [DOI] [PubMed] [Google Scholar]
- Yu, F. H. , Mantegazza, M. , Westenbroek, R. E. , Robbins, C. A. , Kalume, F. , Burton, K. A. , Spain, W. J. , McKnight, G. S. , Scheuer, T. , & Catterall, W. A. (2006). Reduced sodium current in GABAergic interneurons in a mouse model of severe myoclonic epilepsy in infancy. Nature Neuroscience, 9, 1142–1149. 10.1038/nn1754 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. Pharmacokinetic analysis of CBGV. Concentration‐time curve for CBGV in mouse plasma following a 1 mg·kg‐1 i.p. injection. Data are expressed as means ± SEM, with n = 4–5 per time point.
Figure S2. Hyperthermia‐induced seizure screen. Temperature threshold for GTCS in individual mice treated acutely with (A) CBDV or THCV, (B) CBDVA, (C) CBG, (D) CBGA, (E) CBGV or (F) CBGVA. CBDV (30 mg·kg‐1), CBDVA (100 mg·kg‐1), CBGA (30 and 100 mg·kg‐1) and CBGVA (100 mg·kg‐1) treatments were anticonvulsant resulting in a significantly improved response to thermal seizure induction (blue‐green bars); whereas, THCV (3 mg·kg‐1) was proconvulsant and significantly lowered the thermal threshold (red bar). Error bars represent SEM, with n = 12–20 per group (*p < 0.05, **p < 0.01, ****p < 0.0001; logrank Mantel‐Cox).
Figure S3. Pharmacokinetic interaction between CBGA and N‐CLB, the active metabolite of clobazam. Average plasma concentrations of (a) clobazam and (b) N‐CLB in Scn1a +/− mice from hyperthermia‐induced seizure and spontaneous seizure experiments. Clobazam was administered acutely (solid bars) or subchronically (hatched bars) in the presence (dark green bars) and absence of CBGA (light green bars), with n = 5 per treatment. Significantly lower plasma N‐CLB levels were observed following combination treatment of CBGA and clobazam (*p < 0.05, **p < 0.01; Student's ttest).
Figure S4. In vitro characterization of GPR55 activity. HEK293 cells expressing human GPR55 were used to characterize inhibitory and agonist activity of compounds. ERK phosphorylation (pERK) was used as a measure of GPR55 activation. (a) Concentration‐response curves for activation of GPR55 by ML‐186 (blue symbols) and LPI (gray symbols). Data are expressed as mean ± SEM normalized to 100 nM phorbol 12‐myrisate 13‐acetate (PMA) and fit to the Hill equation, with n = 3–4 per concentration. (b) Concentration‐response for CID16020046 inhibition of GPR55 activation measured by ERK phosphorylation (pERK). GPR55 activation was measured following treatment with either LPI (gray symbols) or ML‐186 (blue symbols) in the presence of varying concentrations of CID16020046 and expressed as a percentage of responses to ligand alone. Data are expressed as mean ± SEM, with n = 6 per group. Curves represent fit to a three‐parameter log function and IC50 values are listed.
Figure S5. In vitro characterization of TRPV1 agonists. HEK293 cells expressing human
TRPV1 were used to characterize agonist activity of capsaicin and CBGA. Changes in intracellular calcium were used as a measure of TRPV1 activation. Concentration‐response curves for capsaicin (red) and CBGA (purple) in (a) wildtype and (b) TRPV1‐expressing cells. Data are expressed as mean ± SEM normalized to PAR‐1 and fit to the Hill equation, with n = 4–5 per group. The EC50 value of capsaicin for TRPV1 was 3.3 nM (95% Confidence Interval, 1.9–5.8 nM). Non‐specific calcium influx was observed with CBGA treatment (30 and 100 μM).
Table S1. Parameters for LC–MS/MS Detection of Phytocannabinoids
Table S2. Eurofins DiscoverX GPCR Screen
Table S3. Plasma cannabinoid concentrations in experimental Scn1a+/− mice
Table S4. Effect on GPR55 activity
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.