

ABSTRACT
There are several competing hypotheses on tooth origins, with discussions eventually settling in favour of an ‘outside‐in’ scenario, in which internal odontodes (teeth) derived from external odontodes (skin denticles) in jawless vertebrates. The evolution of oral teeth from skin denticles can be intuitively understood from their location at the mouth entrance. However, the basal condition for jawed vertebrates is arguably to possess teeth distributed throughout the oropharynx (i.e. oral and pharyngeal teeth). As skin denticle development requires the presence of ectoderm‐derived epithelium and of mesenchyme, it remains to be answered how odontode‐forming skin epithelium, or its competence, were ‘transferred' deep into the endoderm‐covered oropharynx. The ‘modified outside‐in’ hypothesis for tooth origins proposed that this transfer was accomplished through displacement of odontogenic epithelium, that is ectoderm, not only through the mouth, but also via any opening (e.g. gill slits) that connects the ectoderm to the epithelial lining of the pharynx (endoderm). This review explores from an evolutionary and from a developmental perspective whether ectoderm plays a role in (pharyngeal) tooth and denticle formation. Historic and recent studies on tooth development show that the odontogenic epithelium (enamel organ) of oral or pharyngeal teeth can be of ectodermal, endodermal, or of mixed ecto–endodermal origin. Comprehensive data are, however, only available for a few taxa. Interestingly, in these taxa, the enamel organ always develops from the basal layer of a stratified epithelium that is at least bilayered. In zebrafish, a miniaturised teleost that only retains pharyngeal teeth, an epithelial surface layer with ectoderm‐like characters is required to initiate the formation of an enamel organ from the basal, endodermal epithelium. In urodele amphibians, the bilayered epithelium is endodermal, but the surface layer acquires ectodermal characters, here termed ‘epidermalised endoderm’. Furthermore, ectoderm–endoderm contacts at pouch–cleft boundaries (i.e. the prospective gill slits) are important for pharyngeal tooth initiation, even if the influx of ectoderm via these routes is limited. A balance between sonic hedgehog and retinoic acid signalling could operate to assign tooth‐initiating competence to the endoderm at the level of any particular pouch. In summary, three characters are identified as being required for pharyngeal tooth formation: (i) pouch–cleft contact, (ii) a stratified epithelium, of which (iii) the apical layer adopts ectodermal features. These characters delimit the area in which teeth can form, yet cannot alone explain the distribution of teeth over the different pharyngeal arches. The review concludes with a hypothetical evolutionary scenario regarding the persisting influence of ectoderm on pharyngeal tooth formation. Studies on basal osteichthyans with less‐specialised types of early embryonic development will provide a crucial test for the potential role of ectoderm in pharyngeal tooth formation and for the ‘modified outside‐in’ hypothesis of tooth origins.
Keywords: oropharynx, endoderm, ectoderm, periderm, pharyngeal pouches, tooth development, dermal denticles, pharyngeal denticles
I. INTRODUCTION: GERM LAYERS AND THE EVOLUTIONARY ORIGIN OF TEETH
Teeth play a pivotal role in tracing the evolutionary history of vertebrates (e.g. Janvier, 1996; Ungar, 2010). Not surprisingly, the evolutionary origin of teeth has itself been at the centre of evo‐devo research. Based on their composition and development, it was recognised in the 19th century that teeth derive from skin denticles, elements of the ancient vertebrate dermal skeleton (Hertwig, 1874). Both teeth and skin denticles qualify as ‘odontodes’ – isolated structures that consist of a dentine cone, are covered by a hypermineralised layer and have a bony base which serves as an attachment (Reif, 1982; see Table 1). The canonical view that skin denticles invaded the mouth cavity and gave rise to teeth at the dawn of gnathostome evolution, in conjunction with the origin of jaws, dominated our thinking for well over a century (e.g. Romer, 1949; Jollie, 1968; Peyer, 1968; Ørvig, 1977).
Table 1.
Glossary. In its original concept, the term ‘odontode’ was used to refer to ‘dermal teeth’ – hard tissue units in the corium, corresponding very closely to teeth, composed principally of dentine or dentinous tissue, with or without an outer covering of enameloid, and forming ontogenetically in a single undivided mesenchymal dental papilla (Ørvig, 1967, 1977). Reif (1982) slightly modified the definition of Ørvig (1977) and, importantly, expanded it to encompass both ‘dermal denticles’ and ‘teeth’. In Reif's view, only those organ systems that are formed by a dental lamina qualify as dentitions, and its components referred to as teeth. Reif's (1982) definition of an odontode has gained wide acceptance, but the dependence on a dental lamina for teeth is now no longer considered valid: the dental lamina has been seriously challenged as a patterning device for the dentition in early jawed vertebrates (Qu et al., 2013), or is absent altogether in developing teeth in many extant species (see text). Donoghue & Rücklin (2016) found an elegant way to circumvent this problem by referring to external and internal odontodes. While we support this distinction, it does not do justice to the diversity of internal odontodes. Thus, in the current review, we will adopt the terminology shown in this table
| Odontode | An isolated superficial structure of the dermal skeleton which consists of a dentinous tissue and a hypermineralised cap of enamel or enameloid either present or absent, and which rests on a base of acellular or cellular bone functioning as an attachment tissue. Developmentally, odontodes form in a single undivided mesenchymal dental papilla, bounded at its surface by an epithelial dental (or enamel) organ (Reif, 1982). Skin (or dermal) denticles, teeth, pharyngeal denticles, and placoid scales all qualify as odontodes. |
| External odontodes | Odontodes located in the skin, synonymous with skin (or dermal) denticles. Chondrichthyans have external odontodes that are usually called placoid scales. These can be non‐growing and replaced, or growing and retained (Reif, 1978b ). |
| Internal odontodes | Odontodes located within the oropharynx, with their patterning and replacement highly coordinated compared to external odontodes (Doeland et al., 2019). Two categories of internal odontodes can be distinguished – teeth, and (pharyngeal) denticles – although differences are somewhat blurred as a result of their conserved structure and development. |
| Teeth | Internal odontodes with an important function in food uptake, transport and other types of food manipulation. Usually large units, and in low numbers. Teeth can be associated with the jaw margins and palate (oral teeth) but also with any of the post‐mandibular arches (pharyngeal teeth). In a number of extant teleosts, tooth structures appear in the skin but are phylogenetically discontinuous with skin denticles, and represent the outcome of a tooth developmental program outside the mouth. These have been called ‘extra‐oral teeth’ (Sire & Allizard, 2001). |
| Pharyngeal denticles | Internal odontodes that are topologically related to teeth (i.e. located inside the oropharynx), but that functionally resemble external odontodes, for example in providing the oropharyngeal lining with a rough surface. Usually small units, and in high numbers. Many extant chondrichthyans have internal odontodes that, like external odontodes, are often called ‘placoid scales’ (Nelson, 1970), or ‘mucous membrane denticles’ (Peyer, 1968). In tetrapodomorph fishes and early tetrapods the denticles, as well as the region where they are found, are called ‘branchial’ rather than ‘pharyngeal’ (e.g. Schoch, 2002). |
| Dental lamina | An epithelial thickening invaginating into the underlying mesenchyme, yielding one or multiple anlagen of enamel organs, and thus tooth germs (Reif, 1978a , 1982). |
In the 1990s, an alternative hypothesis about tooth origins was proposed, based on a revised analysis of palaeontological and neontological data (Smith & Coates, 1998, 2000, 2001; Smith, 2003; Johanson & Smith, 2005). It became known as the ‘inside‐out’ hypothesis, to distinguish it from the classical hypothesis that became known as the ‘outside‐in’ hypothesis. The arguments were: (i) conodonts were reinterpreted as jawless vertebrates and mineralised conodont structures were accepted as teeth. Since conodonts have no dermal skeleton and the conodont elements were believed to be located inside the animals' pharynx, it was concluded that not skin denticles but endoderm‐derived pharyngeal structures gave rise to teeth (Smith & Coates, 1998). (ii) The discovery of thelodonts with pharyngeal denticles was argued to provide evidence for an origin of teeth independent of jaws (Smith & Coates, 1998, 2000, 2001). (iii) Patterned denticle arrays were identified on the posterior wall of the pharyngeal cavity, the postbranchial lamina, in placoderms (early jawed vertebrates) (Johanson & Smith, 2005). (iv) It was claimed that transitory stages between teeth and skin denticles cannot be found (Smith & Coates, 1998, 2000, 2001). (v) It was suggested that teeth must always develop from, and be patterned by, a specialised epithelial structure, the dental lamina (Smith & Coates, 1998). That skin denticles develop directly from the interaction between the epithelium and the mesenchyme, without the formation of a dental lamina, was used as an argument against their homology with teeth. The conclusion was that teeth evolved first in the pharynx from the interaction between endoderm and mesenchyme, and not, like skin denticles, from ectoderm and mesenchyme (Smith & Coates, 1998, 2000, 2001; Smith, 2003; Johanson & Smith, 2005; Fraser et al., 2009). This view was further fostered as a result of comparing the patterning of shark skin denticles with that of dentitions (Fraser & Smith, 2011).
The debate extended for over a decade before the pendulum eventually swung back to the earlier, canonical view and the recognition that internal odontodes (oral or pharyngeal teeth or denticles) in jawed vertebrates are evolutionarily derived from the expansion of odontogenic epithelium from the skin into the oropharynx (Witten, Sire & Huysseune, 2014). Key elements that led to this insight were: (i) that tooth‐like elements such as found in derived conodont taxa are an innovation peculiar to that lineage (Murdock et al., 2013); (ii) that the internal scales in thelodonts fail the test of phylogenetic continuity with teeth of jawed vertebrates (Rücklin et al., 2011); (iii) that the tooth‐like structures on the postbranchial lamina in placoderms are in fact focal developments of bone (Rücklin et al., 2012); (iv) the recognition of intermediate stages between external odontodes and teeth (Blais, MacKenzie & Wilson, 2011; King, Marone & Rücklin, 2021), and (v) a reappraisal of the significance of the dental lamina (Huysseune, Sire & Witten, 2009, 2010; Qu et al., 2013; Vandenplas, De Clercq & Huysseune, 2014). The different arguments have been well covered in a series of papers, that also demonstrate that teeth are genuine elements of the dermal skeleton (Sire & Huysseune, 2003; Reif, 2006; Harris et al., 2008; Huysseune et al., 2009, 2010; Turner et al., 2010; Blais et al., 2011; Debiais‐Thibaud et al., 2011; Rücklin et al., 2011, 2012; Murdock et al., 2013; Qu et al., 2013; Donoghue & Rücklin, 2016; Haridy et al., 2019; Chen et al., 2020).
Huysseune et al. (2009, 2010) proposed a slightly modified version of the canonical view, designated as the ‘modified outside‐in’ hypothesis. This proposal followed the recognition that odontodes in the oral and pharyngeal cavity are serial homologues and to a large extent share genetic developmental pathways (Stock, 2001; Wise & Stock, 2006; Debiais‐Thibaud et al., 2007, 2008). Thus, the ‘modified outside‐in’ hypothesis was introduced to explain the presence of teeth throughout the mouth and pharynx (collectively called the oropharynx), that is, not just teeth associated with the mandibular arch and the palatal bones (oral teeth), but also teeth developing in association with the more posterior pharyngeal arches, that is the hyoid and the branchial arches sensu stricto [pharyngeal teeth (Nelson, 1969; Berkovitz & Shellis, 2016)]. In its original version, the ‘modified outside‐in’ hypothesis attached a great deal of importance to open oral and pharyngeal orifices. It suggested that epithelium with odontode‐forming competence may have entered the oropharynx via these orifices, in particular the gill slits. It was furthermore suggested that an invasion of tooth‐competent epithelium into the oropharynx may have occurred independent from (and perhaps even prior to) the presence of jaws (Huysseune et al., 2009, 2010). In this respect, it is interesting to note that several authors have emphasised the disparate evolutionary history of teeth and jaws (McCollum & Sharpe, 2001; Rücklin et al., 2012; Rücklin & Donoghue, 2015).
That the origin of teeth has been placed either in the pharynx or in the skin hints at the crucial role that germ layers have played in the discussions summarised above. Vertebrates, like all bilaterian animals, are built on a three‐layered plan, the three layers of embryonic tissue, or germ layers: ectoderm, endoderm and mesoderm. Ectoderm gives rise to the epithelium of the skin, the epidermis, while endoderm forms the epithelial lining of the digestive tube and associated organs. In addition, the ectoderm is at the origin of the neural crest, a cell population that produces diverse tissues and is thus often considered as the fourth germ layer (Hall, 1998, 2000). Vertebrate teeth, as a paradigmatic example of organogenesis, develop through reciprocal interactions between an epithelial component (derived from ectoderm or endoderm, and forming the enamel organ) and the underlying mesenchyme (neural crest‐derived, and forming the dental papilla) (Kollar & Baird, 1969; Balic & Thesleff, 2015; Balic, 2019; Yu & Klein, 2020) (Fig. 1A). The enamel organ is responsible, at least in part, for the production of the hypermineralised tooth cap; the dental papilla for the production of dentine.
Fig 1.
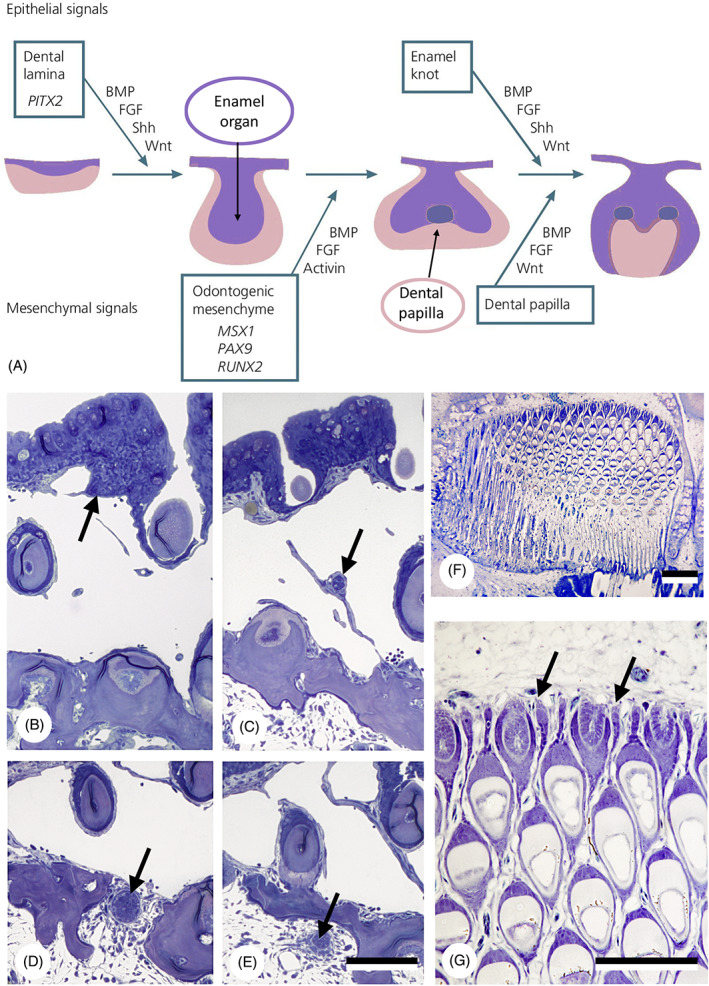
Epithelial primacy in tooth formation. (A) Schematic representation of tooth formation in mammals and transcription factors and signalling molecules involved (modified from Koch, Thesleff & Kreiborg, 2017); purple: epithelium/enamel organ; pink: mesenchyme/dental papilla. BMP, bone morphogenetic protein; FGF, fibroblast growth factor; Shh, sonic hedgehog. (B–E) Intramedullary tooth formation in the lower pharyngeal jaw of the cichlid Hemichromis bimaculatus, 28 mm standard length (SL). Successive cross sections through the epithelial strand (arrow) giving rise to the enamel organ of the replacement tooth, showing its origin (B), its course across the large vascular cavity (C), its penetration through the jaw bone (D) and its distal end within the medullary cavity of the jaw bone (E). Only few mesenchymal cells have condensed around the distal end. Total length of the epithelial strand is approximately 300 μm. Scale bar for B–E = 100 μm. (F, G) Section through the left premaxilla of a scraping loricariid catfish, Ancistrus cf. triradiatus, of 33.5 mm SL. Note virtual absence of mesenchymal cells at the distal end of each successional lamina (arrows). Scale bars: F = 100 μm; G = 50 μm.
In the ‘inside‐out’ model, the germ layer origin of the epithelium (ectoderm for external odontodes, endoderm for teeth) was considered to impart real differences to the two systems (Smith & Coates, 2001; Smith & Johanson, 2015). With the experimental evidence that the enamel organ can be of ectodermal, endodermal or mixed origin (Soukup et al., 2008), the focus in discussions shifted to the pivotal role of the mesenchyme, and it was proposed that the germ layer origin of the epithelial enamel organ does not matter (Fraser et al., 2010; Calamari, Kuang‐Hsien Hu & Klein, 2018). Fraser et al. (2010) argued that teeth develop wherever specific signalling networks drive interactions between neural crest‐derived mesenchyme and competent epithelium, independent from the germ layer origin of the epithelial layer. This suggestion, labelled as the ‘inside and out’ model, moved the focus away from arguments about primacy of location and/or cell type (Fraser et al., 2010; Fraser & Smith, 2011). In this model the expansion of odontogenic competence from external to internal epithelia is attributed to the primacy of neural crest‐derived ectomesenchyme, rather than to the origin of the epithelia (Soukup et al., 2008; Jheon et al., 2012; Donoghue & Rücklin, 2016).
There are nevertheless several arguments to reject the primacy of the ectomesenchyme in odontode induction. From an evolutionary viewpoint, it has been argued that enamel‐like (i.e. epithelium‐derived) substances may well have been the most primitive vertebrate hard tissues (Maisey, 1988), although the early diversity of vertebrate mineralised tissues makes this a contentious issue (Donoghue & Sansom, 2002; Donoghue, Sansom & Downs, 2006; Kawasaki et al., 2020). From a developmental viewpoint, epithelial primacy in tooth initiation seems well underscored by reports detailing the first molecular evidence of mammalian tooth initiation in the epithelium (Peters et al., 1998; Peters & Balling, 1999; Ohazama et al., 2010; Prochazka et al., 2015; Calamari et al., 2018; Yuan & Chai, 2019) (Fig. 1A). Epithelial primacy appears also evident in the case of intramedullary (also called intraosseous) tooth formation (Trapani, 2001), as observed in advanced teleost fish [e.g. Trachinotus teraia (Françillon‐Vieillot et al., 1994); Hemichromis bimaculatus (Huysseune & Thesleff, 2004)]. Here, replacement tooth formation starts with the production of an epithelial strand (the successional dental lamina) branching off from the outer dental epithelium. This strand grows over a large distance, piercing through bone, to reach the medullary cavity. Only then, within the medullary cavity, does mesenchyme start to condense around the tip of the strand, suggesting a later role in tooth development (Fig. 1B–G). The crucial role of the epithelium in initiating tooth formation justifies our focus on the odontogenic epithelium. The germ layer origin of the odontogenic mesenchyme (i.e. neural crest‐derived ectomesenchyme) is taken for granted here (see e.g. Raven, 1935; de Beer, 1947; Chibon, 1966, 1967; Chai et al., 2000; Hall & Gillis, 2013).
This review intends to assess, both from an evolutionary and developmental perspective, whether ectoderm plays a role in tooth formation, as proposed in the ‘modified outside‐in’ hypothesis, and whether this can explain the presence and distribution of teeth in the oropharynx. The emphasis is on pharyngeal teeth and denticles, as these present a greater challenge to the classical ‘outside‐in’ view than oral teeth, which are located close to the mouth entrance.
II. GERM LAYERS CONTRIBUTING TO TEETH AND THEIR ROLE IN TOOTH INDUCTION
Crucial in the discussion about the homology of external odontodes (skin denticles) with internal odontodes (teeth and pharyngeal denticles) is the extent to which the ectoderm penetrates into the oropharynx. According to embryology textbooks, the distribution of ectoderm is limited to the stomodaeal area, whereas the post‐mandibular pharyngeal arches are lined by endoderm (Balinsky, 1975; Hildebrand, 1995). Yet, recent studies show that the oral cavity can be covered by endoderm, by ectoderm, or can have a mixed lining, depending on the process of primary mouth formation (Soukup, Horacek & Cerny, 2013; Chen et al., 2017; Minarik et al., 2017) (Fig. 2).
Fig 2.
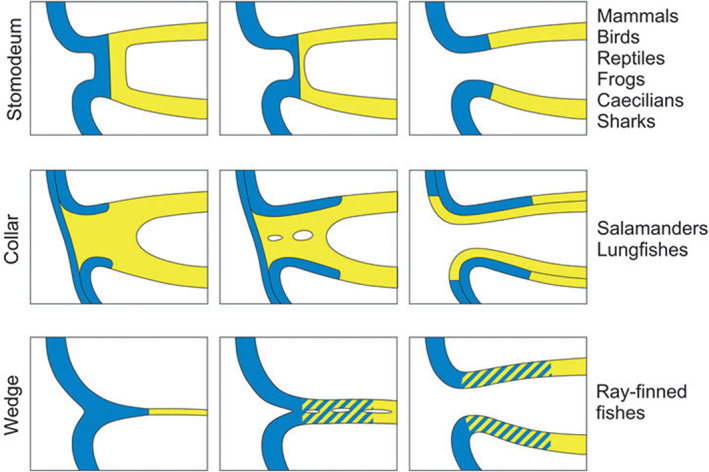
Schematic representation of primary mouth formation in vertebrates [from Soukup et al., 2013, with permission from the authors and publisher]. Yellow, endoderm; blue, ectoderm.
(1). Chondrichthyans
Little attention has been paid to the germ layer origin of chondrichthyan oral teeth. The ectodermal nature of the enamel organ is usually taken for granted, likely because of the close proximity of the dental lamina to the mouth entrance (Hertwig, 1874; Rasch et al., 2016; Fig. 3A, B). At the time the first oral teeth form, the epidermis is clearly stratified, and the basal epithelial layer appears to have a prominent role in early morphogenesis of the enamel organ [illustrated by Rasch et al., 2016 for the spotted catshark (Scyliorhinus stellaris and S. canicula)].
Fig 3.
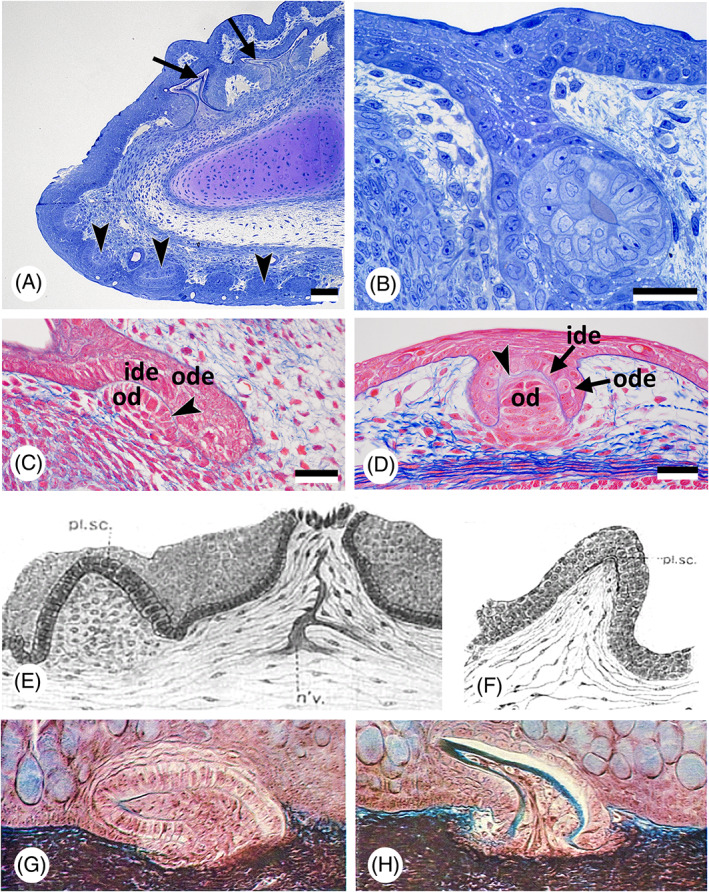
Oropharyngeal epithelium, teeth, and dermal denticles in chondrichthyans. (A) Scyliorhinus canicula; sagittal section of the lower jaw of a 85 mm embryo, showing skin (= dermal) denticles (arrowheads), and the dental lamina with developing teeth (arrows). Scale bar = 100 μm. (B) Scyliorhinus canicula; larger magnification of the early dental lamina with a developing tooth in a 85 mm embryo. Scale bar = 20 μm. (C, D) Scyliorhinus canicula; sagittal section of a tooth anlage in the lower jaw with the invading dental lamina (C) compared to the anlage of a dermal denticle (D) in the same 50 mm embryo. Outer (ode) and inner dental epithelium (ide), the basal lamina of the enamel organ (arrowhead) and the mesenchymal condensation that will give rise to the odontoblasts (od) characterise the anlagen of teeth and dermal denticles. Scale bars = 25 μm. (E, F) Squalus acanthias; two stages of development of a ‘placoid scale’ (from Cook & Neal, 1921). n'v. = nerve; pl. sc. = placoid scale. (G, H) Mustelus sp.; two stages of development of a ‘mucous membrane denticle’ (from Peyer, 1968).
Few studies have been conducted on the development of pharyngeal denticles in sharks, let alone on their germ layer origin (Fig. 3C, D). Cook & Neal (1921) searched for embryological evidence for the origin of the pharyngeal denticles (that they called “pharyngeal placoid scales”) scattered over the surface of the pharynx of the common spiny dogfish (Squalus acanthias). Based on histological sections, they concluded that the entire pharyngeal cavity is of endodermal origin. They observed little or no inward migration of ectoderm into the pharynx except in the region of the upper and lower jaws and thus considered pharyngeal denticles to be endodermal (Fig. 3E, F). Interestingly, the early stage of pharyngeal denticle formation is drawn with a multilayered epithelium, an observation whose significance will be discussed later (see Sections II.4 and III). Peyer (1968) illustrated the formation of pharyngeal denticles in the smooth‐hound shark (Mustelus sp.), insisting on homology with skin denticles. He showed that they form from a stratified epithelium, but did not assign the epithelium to a particular germ layer (Fig. 3G, H). Regrettably the species which, over the past years, has become a popular chondrichthyan model for developmental studies, the lesser spotted dogfish (Scyliorhinus canicula), possesses no pharyngeal denticles.
(2). Osteichthyans: actinopterygians
Actinopterygians possess teeth whose distribution can extend deep into the alimentary canal. Teeth can be found associated with the hyoid or any of the branchial arches, even down into the oesophagus (Gilchrist, 1922; Isokawa et al., 1965). Thus, actinopterygians represent a particularly interesting group to investigate the germ layer origin of the teeth (see e.g. Stock, 2001; Berkovitz & Shellis, 2016).
(a). Non‐teleost actinopterygians
While the development of the teeth in basal (non‐teleost) actinopterygians has been studied in some detail [e.g. bichir, Polypterus senegalus (Sire et al., 2002); bowfin, Amia calva (Degener, 1924); gar, Lepisosteus oculatus (Sasagawa et al., 2019); Siberian sturgeon, Acipenser baeri (Gisbert et al., 1998)], the germ layer origin of the enamel organ has usually not been addressed. Still, endoderm forms substantial parts of the orofacial epithelia in these taxa, and that includes contributions to the teeth (Minarik et al., 2017). It can therefore be expected that the enamel organs of many (if not all) teeth are endoderm‐derived. Interestingly, at least in bichir and sturgeon, the odontogenic pharyngeal epithelium is clearly stratified (Fig. 4A, B).
Fig 4.
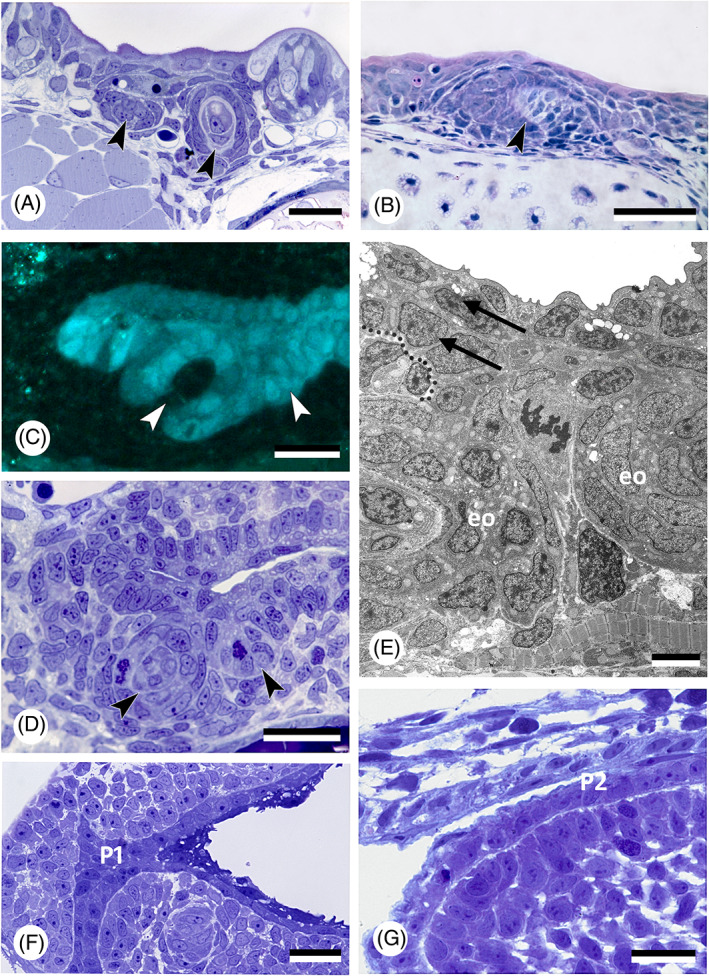
Oropharyngeal epithelium and teeth in actinopterygians. (A) Polypterus senegalus; pharyngeal epithelium and forming tooth germs (arrowheads) in an 18 mm larva. Scale bar = 20 μm. (B) Acipenser ruthenus; tooth germ (arrowhead) in the buccopharyngeal epithelium in a 15 mm larva. Scale bar = 50 μm. (C, D) Danio rerio, cross section of a 56 h post‐fertilisation (hpf) embryo at the level of the first forming teeth (arrowheads), shown in a transgenic (sox17:egfp) line (C), and corresponding semithin, toluidine blue‐stained section of a wildtype fish (D). Scale bars = 20 μm. (E) Danio rerio, transmission electron microscopy (TEM) image of the bilayered pharyngeal epithelium (two arrows, delimited by dotted line) of a 72 hpf embryo at the level of the first two teeth (eo, enamel organ). Scale bar = 5 μm. (F) Hemichromis bimaculatus, cross section of the oropharynx of a 1 day post‐hatching (dph) (4.0 mm total length) specimen at the level of pouch 1 (P1). Scale bar = 20 μm. (G) Oncorhynchus tshawytscha, sagittal section of the pharyngeal epithelium in an unhatched embryo 17 days post‐fertilisation (dpf), at the level of the (open) pouch 2 (P2). Scale bar = 20 μm.
(b). Teleosts
The first detailed study on germ layer participation in teleost teeth comes from Edwards (1929), working on common carp (Cyprinus carpio). The carp is a close relative of the zebrafish (Danio rerio), today a common experimental model for vertebrate development and genetics. As is typical for cyprinids, both species lack oral teeth. Their pharyngeal teeth are restricted to the last (seventh) pharyngeal arch, or fifth branchial (gill) arch. Edwards (1929) drew the following conclusions: (i) the superficial layer of the epidermis invades the gill slits and spreads out over the entire pharynx. Edwards illustrated this for the first gill slit but inferred it to be similar for the other gill slits. As a consequence, (ii) the pharyngeal epithelium is composed of two layers: a deep, columnar layer, of endodermal origin, and a superficial flattened layer, of ectodermal origin. (iii) The enamel organs of the teeth derive from the deep layer and are therefore of endodermal origin. Nearly a century later, Rosa et al. (2019), and Oralová et al. (2020), using transgenic zebrafish lines, largely confirmed Edwards' results on carp. The pharyngeal epithelium in zebrafish is bilayered and the enamel organs are derived from the basal layer only, that is endoderm (Fig. 4C–E), observations also fitting with those of Waterman & Kao (1982). Rosa et al. (2019) also confirmed the invasion of the pharyngeal pouches (i.e. the structures in which the gill slits form, see Section III.1) by the superficial epidermal layer, or periderm (for a discussion on the germ layer origin of the periderm, see Section III.3). In contrast to Edwards' results on carp, however, they established that periderm invasion in zebrafish is arrested halfway into each pouch (Fig. 5A–C). Instead of spreading throughout the oropharynx, the periderm joins another, endogenous population of cells covering the midline endoderm. The latter cells, designated as ‘periderm‐like’ cells, are highly similar to the invading periderm cells and cannot be distinguished from these once both populations have joined (Fig. 5A–E). Apart from their similar morphology, both periderm and periderm‐like cells (i) express periderm markers (krt4, and zc1044), (ii) develop microridges that otherwise characterise periderm cells, and (iii) both cell populations merge imperceptibly, suggesting compatible adhesion; there is no further mixing (Rosa et al., 2019; Fig. 5A–C). The periderm‐like cells furthermore display ultrastructural features that are distinctive from the endodermal cells on which they rest. Apart from size and shape, they notably differ from the endodermal cells by their electrondense cytoplasm (Rosa et al., 2019). The layer of periderm‐like cells expands posteriorly along the midline until the oesophageal–intestinal border, where it ends abruptly. Along its way it joins with periderm invading each of the pouches. Like Edwards (1929), Rosa et al. (2019) observed that the presence of periderm or periderm‐like cells precedes lumenisation of the pouches and of the pharynx, respectively. Whether zebrafish, as a miniaturised species (Witten et al., 2017), truly differs from carp in that periderm invasion is arrested, and the periderm instead joins a periderm‐like layer that expands along the midline, still needs to be established. Significantly, Oralová et al. (2020) showed that zebrafish teeth form from the basal, that is endodermal, layer, but only when this is covered by the periderm‐like layer. Thus, a layer is required for pharyngeal tooth formation that bears similarities to the periderm, or outer epidermal layer (Fig. 5F–H). Whether the periderm‐like layer possesses a signalling function remains unclear (Oralová et al., 2020). A mechanical role should also be considered. In mice, the suprabasal layer of the odontogenic epithelium forms a tensile canopy required for the basal layer of the tooth placode to invaginate (Panousopoulou & Green, 2016). Eventually, elucidating the precise role of the periderm‐like cells in tooth induction will only be possible through ablation experiments, using, for example, genetically engineered lines (Chen et al., 2011; Eisenhoffer et al., 2017).
Fig 5.
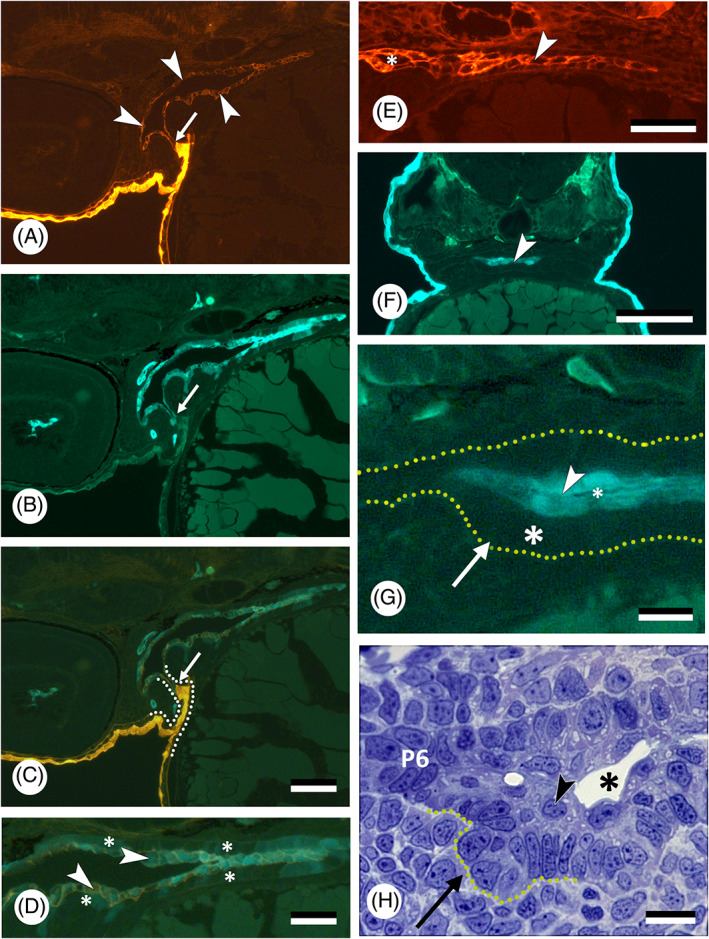
Germ layer contribution to oropharyngeal epithelium and teeth in zebrafish, Danio rerio. (A–C) Sagittal section of a double‐transgenic (krt4:tomatoCAAX;sox17:egfp) 55 h post‐fertilisation (hpf) embryo showing periderm having partially invaded pouch 2 (intense orange), and connecting to periderm‐like cells investing the oropharynx (red, indicated by white arrowheads, shown also in E); boundary indicated by a small white arrow; both populations do not show further mixing. Micrographs are shown in the red (A) and green channel (B), and as overlay (C). Dotted line in C marks the outline of pouch 2. Scale bar for A–C = 20 μm. (D) Magnification of the pharynx lining shown in C; periderm‐like cells (arrowheads) express both krt4 and sox17 and cover endoderm, in which sox17 is downregulated (asterisks). Scale bar = 20 μm. (E) Sagittal section of a transgenic (krt4:tomatoCAAX) 55 hpf embryo showing periderm‐like layer (arrowhead). A lumen has appeared rostrally between the two periderm‐like layers (asterisk). Scale bar = 20 μm. (F) Cross section of a 60 hpf Tg(krt4:gfp) embryo at the level of the forming pharyngeal dentition; periderm‐like layer indicated by an arrowhead. Scale bar = 100 μm. (G) Magnification of F, showing the placode of the first pharyngeal tooth (arrow). This develops from the unlabelled basal (endodermal) layer (large asterisk), while being covered by a krt4‐positive superficial layer (arrowhead). A dotted line delimits the pharyngeal epithelium; a lumen starts to appear between the two layers of periderm‐like cells (small asterisk). Scale bar = 10 μm. (H) Toluidine blue‐stained semithin cross section, showing placode stage of first pharyngeal tooth formation corresponding to G (tooth placode indicated by a dotted outline). Note superficial periderm‐like layer (arrowhead) with forming pharyngeal lumen (asterisk). Laterally, the pouch (pouch 6, P6) is still bilayered. Scale bar = 10 μm.
Several other teleost species (both basal and derived) have been shown also to possess a bilayered pharyngeal epithelium early in development, with superficial flattened cells covering basal cuboidal cells (Gonzalez, Blanquez & Rojo, 1996; Rosa et al., 2019) (Fig. 4F, G), suggesting that the bilayered condition could be a common teleost character.
(3). Osteichthyans: sarcopterygians
(a). Dipnoans
Kerr (1903) examined the formation of the mouth cavity in the South American lungfish Lepidosiren paradoxa, comparing it with mouth formation in Ambystoma mexicanum (Fig. 6A, B). He described that the superficial layer of endoderm transformed into a cell layer with ectoderm‐like characters, along with breakdown of its yolk content. In this way, a layer of definite epithelium arose, continuous with the external epiblast anteriorly, and with the yolk‐laden endoderm posteriorly. Kerr (1903, p. 423) states:
Fig 6.
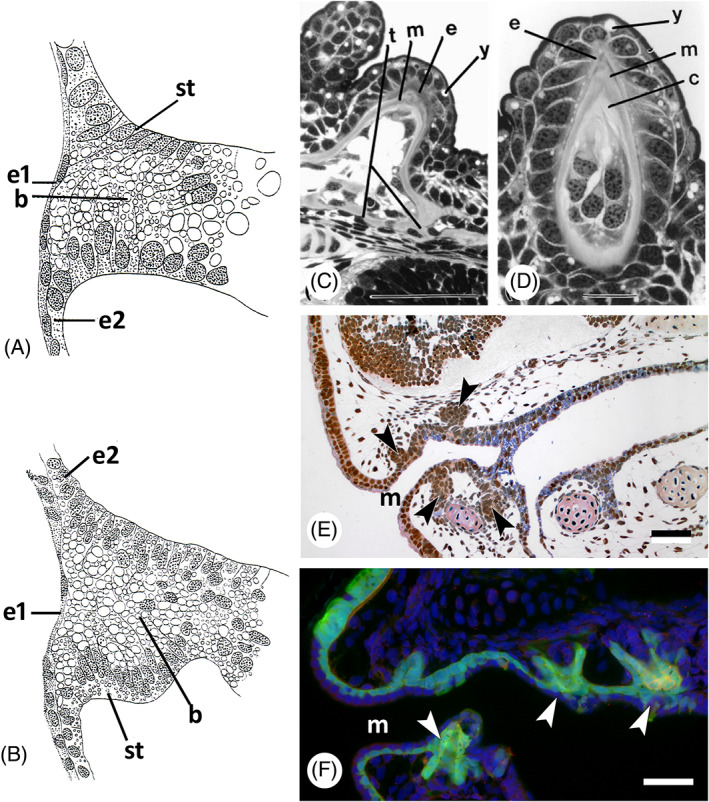
Germ layer contribution to oropharyngeal epithelium and teeth in lungfish and amphibians. (A) Lepidosiren paradoxa; drawing of a sagittal section of a stage 30 specimen (after Kerr, 1903); drawing mirrored to show anterior to the left. b, yolk‐laden cells in the buccal cavity; e1, superficial ectodermal layer; e2, basal ectodermal layer; st, lining epithelium of buccal cavity arising in situ. (B) Ambystoma mexicanum; drawing of a similar section to A, embryo of 7.5 mm length (after Kerr, 1903; drawing mirrored to show anterior to the left). (C, D) Neoceratodus forsteri, cusp of pterygopalatine tooth plate [from Kemp, 2002a , with permission from the author and publisher]. c, circumdenteonal dentine; e, enamel; m, mantle dentine; t, trabeculae; y, yolk remnants. Scale bars: C = 100 μm; D = 20 μm. (E) Ambystoma mexicanum; sagittal section of a stage 43 embryo; compare with F. Tooth buds indicated by arrowheads. m, mouth opening. Scale bar = 50 μm. (F) Ambystoma mexicanum; transgenic embryo of stage 43 with green fluorescent protein (GFP) labelling in the ectoderm [modified from Soukup et al., 2008, courtesy of the authors]. Arrowheads: tooth buds arising in ectoderm; m, mouth opening. Scale bar = 50 μm.
“It is, as it were, as if an influence were spreading inwards from the external epiblast, gradually transforming the original “endoderm” yolk‐laden cells into ectoderm like itself. I find no evidence of an actual bodily involution of ectoderm such as is ordinarily associated with the term stomodaeum.”
He concluded (p. 452) that there is
“no invagination of ectoderm to form a true stomodaeum. The epithelium of the buccal cavity is developed in situ from the outer layer of the solid anterior portion of the yolk‐laden enteric rudiment. … The tooth germs appear while the mouth region is still without a lumen.”
Development in the Australian lungfish, Neoceratodus forsteri, resembles that in Lepidosiren: an endodermal plug makes contact with the stomodaeal plate, upon which the ectoderm disappears. Once a lumen is formed, the lining of the oral cavity is made up of endoderm (Kemp, 2002a ). Up to this point, this resembles the situation in the axolotl (see Section II.3b). Interestingly, as described for Lepidosiren (Kerr, 1903), the endodermal oral epithelium in Neoceratodus differentiates into two layers. The cells of the superficial layer adopt ectodermal characters, like a brush border, or cilia, while their yolk disappears. Only the basal (proximal) layer of the oral epithelium produces the enamel organs (Kemp, 2002a , 2002b ) (Fig. 6C, D).
(b). Lissamphibians
The majority of studies that have attempted to identify germ layer contributions to vertebrate teeth used amphibians, in particular urodeles. Anurans start to develop teeth only at metamorphosis, making them less attractive for experimentation (reviewed in Davit‐Béal et al., 2007). The mouth region in urodeles is formed by an endodermal plug surrounded by an ectodermal sleeve, the so‐called stomodaeal collar (Adams, 1924). This observation was confirmed with the use of transgenic axolotls (Soukup et al., 2008) (Fig. 6E, F).
In a very detailed account, Adams (1924) described the succession of tooth germs developing on the jaws of the axolotl, Ambystoma (then called Amblystoma) punctatum, all of which appear between stages 37 and 39 (her staging system partly derived from Harrison, 1918). Adams concluded that palatine and splenial (= coronoid) teeth possess endodermal enamel organs, while maxillary (= premaxillary), vomerine, and dentary teeth, with one exception, arise entirely within the ectodermal collar. The median pair of dentary teeth has enamel organs of mixed ectodermal–endodermal composition. Germ layer contributions were identified based on histology, in particular the presence of large yolk granules in the endoderm. Adams (1924) furthermore saw a similarity in the sequence of development of the teeth in the axolotl (first the endodermal, then the ectodermal teeth) and in dipnoans. She attributed this to the presence, of “a thick endodermal component in the oral plate” in both groups (p. 320). Based on the results of extirpation and transplantation experiments, Adams suggested that the formation of an ectodermal collar around the endodermal plug is essential for the development of maxillary, vomerine, and dentary (i.e. ectodermal) teeth. The endodermal splenial and palatine tooth germs were constant in their time of appearance in the stomodaeal regeneration experiments, despite variable appearance of the ectodermal collar (Fig. 7A, B). According to Adams, no tooth germs formed in stomodaeal ectoderm transplanted to the gill region. However, homoplastic ventral body wall ectoderm transplanted to the stomodaeal region could apparently initiate tooth germ development. The conclusion that one can draw from Adams' studies is that normal ectoderm–endoderm contact (as in a one‐layered ectodermal sleeve around an endodermal plug) is required for ectodermal tooth formation. This requirement appears to be less stringent for endodermal teeth, which appeared to develop more autonomously (Fig. 7B). Adams (1924) nevertheless also reported the presence of ‘maxillary bulbs’, consisting of ectoderm alone, and which contained tooth germs. To her, this indicated some potentiality for independent invagination without endodermal contact (Fig. 7C). In a follow‐up study, Adams (1931) removed endoderm from axolotl embryos with closing or closed neural folds (i.e. prospective neural crest mesenchyme). Teeth with endoderm‐derived enamel organs were reduced. Their number nevertheless suggested that a re‐established ecto–endodermal contact may have promoted the development of such teeth. Teeth with ectoderm‐derived enamel organs were also reduced (although no ectoderm was removed), leading Adams to conclude that these teeth need an interaction with the endoderm.
Fig 7.
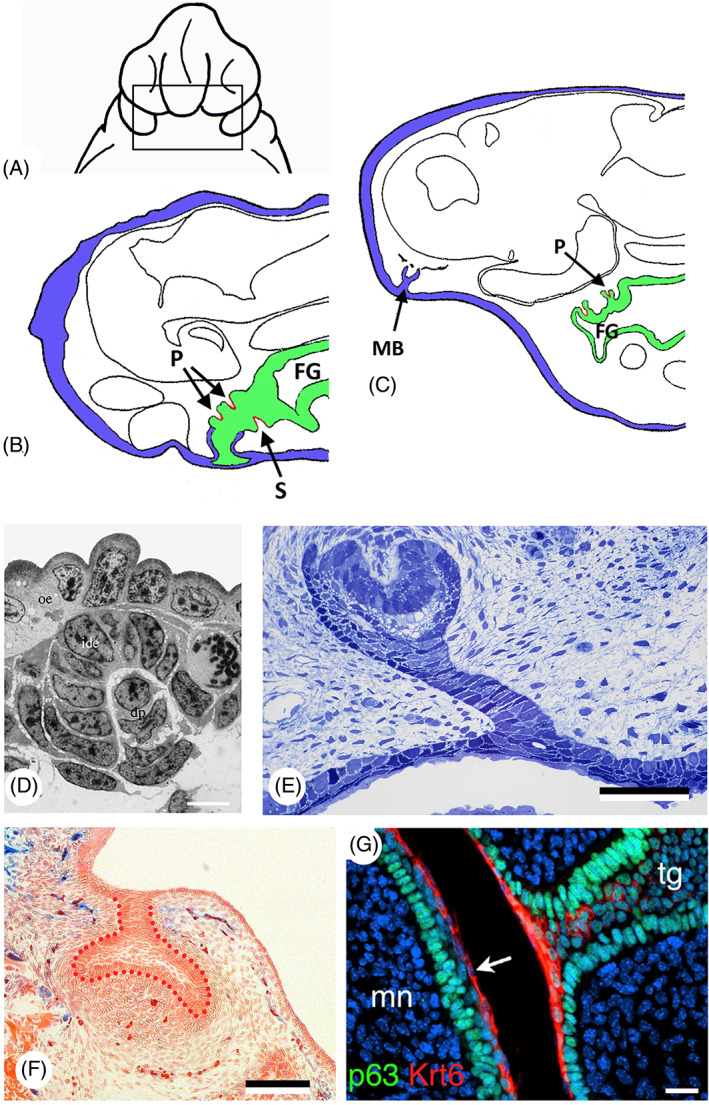
Germ layer contribution to oropharyngeal epithelium and teeth in amphibians and amniotes. (A) Ambystoma punctatum, extirpation of the ectoderm of a stage 24 embryo, ventral view (redrawn after Adams, 1924). (B) Ambystoma punctatum; sagittal section of a stage 39 embryo after extirpation of the ectoderm; teeth in the ectoderm (blue) are reduced or absent, while endoderm (green) has given rise to splenial (S) and palatine (P) tooth germs (red); FG, foregut (adapted after Adams, 1924). (C) Ambystoma punctatum; sagittal section of a stage 45 embryo showing a maxillary bulb (MB) separated from any connection with the foregut; no contact between ectoderm (blue) and endoderm (green), yet a palatine tooth (P) is present (red); FG, foregut (adapted after Adams, 1924). (D) Pleurodeles waltl, first‐generation tooth, stage 36. dp, dental papilla; ide, inner dental epithelium; oe, oral epithelium (from Davit‐Béal et al., 2007, with permission from the publisher). Scale bar = 10 μm. (E) Chalcides viridanus; cross section of the dental lamina on the upper jaw of a 30 mm embryo. Scale bar = 50 μm. (F) Mus musculus; frontal section through a cap‐stage molar tooth germ of a Sox17‐2A‐iCre/R26R mouse, with the epithelium outlined with red dots; there is no contribution of the endoderm (blue) [modified from Rothova et al., 2012, courtesy of the authors ]. Scale bar = 100 μm. (G) Mus musculus; immunostaining for keratin 6 (Krt6, red) and tumor protein p63 (p63, green) in a E15.5 embryo. Krt6 is expressed uniformly in the periderm superficial to the tooth germ (arrow). mn, mandible; tg, tooth germ (from Peyrard‐Janvid et al., 2014, with permission from the publisher). Scale bar = 20 μm.
In the years that followed, several authors critically assessed Adams' results, drawing from their own studies on axolotl. Based on extensive series of experiments, both Ströer (1933) and Holtfreter (1935) concluded that endoderm does not participate in tooth formation, but is nevertheless required. Likewise, Sellman (1946) concluded that all the axolotl enamel organs are most probably ectodermal, a finding we know now to be incorrect (Soukup et al., 2021). More importantly, however, his studies revealed that three tissue layers are required for tooth formation: neural crest, mouth ectoderm and stomodaeal endoderm. Sellman did not rule out the possibility of endoderm‐derived enamel organs but insisted that ectoderm would be necessary for endodermal teeth to form (p. 112). Sellman (1946) agreed with Adams (1924) in that extirpation of mouth ectoderm causes tooth reduction, especially of anterior teeth. Still, he contested Adams' interpretation that the unaffected posterior teeth were endodermal in origin. Sellman also noticed an abnormal formation of the visceral skeleton in these experiments and concluded that tooth reduction was related to disturbance of the neural crest‐derived ectomesenchyme. In contrast to Sellman (1946), but in agreement with Adams (1924), de Beer (1947) concluded that enamel organs can be endodermal or ectodermal.
Rather than using transplantation experiments, Wilde (1955) removed parts of the cranial neuroepithelium in Ambystoma maculatum prior to the migration of neural crest cells. The epithelium was explanted into a nutrient medium, alone or in combination with stomodaeal ectoderm, foregut endoderm or endomesoderm. He observed teeth only when both stomodaeal ectoderm and foregut endoderm were present in the explants, further supporting the idea that all three tissue layers are required for teeth to be initiated.
From his studies on a urodele, the Spanish newt (Pleurodeles waltlii), Chibon (1966) concluded that the neural crest‐derived odontoblasts exert an inductive action upon the epithelium, which can be either ectodermal (in normal conditions) or endodermal. A few years later, Chibon (1970) transplanted the stomodaeal ectoderm of 3H‐thymidine‐labelled neurula‐stage embryos of Pleurodeles waltlii onto non‐labelled siblings. Based on 22 embryos that developed normally, he observed that anterior tooth germs are derived from ectoderm, posterior ones from endoderm, while some contained both labelled and non‐labelled ameloblasts, which he considered indicative of a mixed origin.
Cassin & Capuron (1979) examined the formation of buccal structures both by intrablastocelic transplantation and by in vitro culture, using stage 15 and 18 neurulae of the urodele Pleurodeles waltlii. They obtained teeth when neural fold was associated with stomodaeal endoderm, and claimed that stomodaeal ectoderm is incapable on its own of forming teeth, even when associated with neural fold. There are, however, a number of problems associated with this paper, both at the experimental and analytical level. For example, teeth were reported to develop in transplants that had no neural fold (transplants with stomodaeum alone, that is ectoderm and endoderm, with or without tissues underlying the median neural fold). At best, these results are in line with those of Adams (1924), suggesting that endoderm can form teeth alone with neural fold, while ectoderm needs an interaction with endoderm.
To avoid any bias in their study of neural crest potential, Graveson, Smith & Hall (1997) explanted neural fold segments of Ambystoma mexicanum along with both stomodaeal ectoderm and pharyngeal endoderm as “inductive epithelia” to elicit tooth formation. In a study primarily aimed at investigating taste buds, Barlow & Northcutt (1995) labelled presumptive cephalic endoderm in Ambystoma sp. with the carbocyanine dye, DiI, shortly after the onset of gastrulation and found teeth in areas of labelled cells, suggesting an endodermal origin of the enamel organs.
Soukup et al. (2008) re‐investigated germ layer origin of enamel organs in the axolotl by means of ectodermal transplantations, taking advantage of transgenic axolotls expressing GFP ubiquitously. The experiments demonstrated conclusively that the enamel organs can be of ectodermal, endodermal, or mixed origin. This confirmed the observations of Adams (1924) and de Beer (1947) based on histology, and of Chibon (1970), based on isotope labelling. Whether these results also apply to other urodele species is currently not known.
In conclusion, while it is now firmly established that the enamel organ in urodeles can be of ectodermal, endodermal, or mixed origin, most studies also indicate that both ectoderm and endoderm are required in conjunction with neural crest‐derived mesenchyme to make teeth. Moreover, it has been recently revealed that the entire oral dentition in the axolotl, comprising both the outer and inner tooth arcades, develop from the common competent zone alongside the ectoderm–endodermal boundary (Soukup et al., 2021). Yet, two reported experiments on axolotl suggest that endoderm can form teeth in the absence of ectoderm. First, the extirpation of ectoderm did not affect the formation of endodermal teeth (Adams, 1924), and second, transplants or cultures of stomodaeal endoderm alone, together with neural fold, could generate teeth – ectoderm was not required (Cassin & Capuron, 1979). At first sight, this violates the need for ectoderm as stipulated in the ‘modified outside‐in’ hypothesis. We will return to these findings in Section VI. Importantly, the urodele enamel organ develops from the basal layer of a stratified epithelium (Fig. 7D).
(c). Sauropsids
Data on sauropsids are scarce, given that reptiles are not easily accessible for experimental manipulation, and given that all modern birds lack teeth. In the absence of studies using labelling techniques, enamel organs in the different lineages have been assigned to the ectoderm (e.g. snakes; Buchtová et al., 2008), likely because teeth in reptiles are restricted to the oral region (Richman & Handrigan, 2011). Pythons, however, have two distinct types of dental development that appear to be dependent on the degree to which sonic hedgehog (Shh) expression is maintained in the oral epithelium (Buchtová et al., 2008). Whether this reflects differences in germ layer origin of these teeth is unknown. Data on the possible stratification of the odontogenic epithelia are even more scarce. At least in the scincid lizard Chalcides viridanus, the first tooth placode derives solely from the basal layer of the stratified buccal epithelium (Delgado et al., 2005) (Fig. 7E). Kollar & Mina (1991) transplanted oral epithelium from mice, which is of ectodermal origin (see Section II.3d), onto the mandibular mesenchyme of chickens and generated teeth. Perhaps this experiment showed the undisputed odontogenic potential of oral epithelium of mice rather than a hidden odontogenic potential of chicken. However, Mitsiadis, Caton & Cobourne (2006) transplanted mouse neural folds into chicken and obtained tooth anlagen which suggests a potential of the oral epithelium of chicken to generate an enamel organ.
(d). Mammals
It has long been thought that the posterior part of the mammalian mouth is covered with endodermal epithelium, and the anterior part with ectoderm. Thus, incisor and molar teeth could have a different germ layer origin. Imai, Osumi & Eto (1998) used an endodermal cell‐tracing system in the rat with a recombinant adenovirus, but could only demonstrate that tooth germs form in ectoderm adjacent to labelled endodermal cells (foregut endoderm); there were no functional studies involved. Ohazama et al. (2010) showed that the expression of five endodermally expressed genes extends anterior to the rostral extent of pharyngeal endoderm cells, into what is considered to be oral ectoderm. They concluded that molar (but not incisor) teeth develop from epithelium that shares molecular characteristics with pharyngeal endoderm. The issue was finally settled by Rothova et al. (2012). Using Sox17‐2A‐iCre/Rosa26 reporter mice, their study clearly demonstrated that endoderm does not contribute at any stage to tooth development, including not to molar development. The odontogenic epithelium was identified unequivocally as ectodermal (Rothova et al., 2012) (Fig. 7F). Interestingly, during mouse embryonic development, an initially single‐layered ectoderm stratifies to produce an outer layer of flattened periderm cells (Richardson et al., 2014). Whether the periderm has a function in tooth development has not been investigated, but its appearance in mice from E10.5 onwards (Richardson et al., 2014), that is just before the onset of tooth formation, warrants such a study (Fig. 7G).
(4). Conclusion
The general conclusion drawn from the studies reviewed above is that, depending on taxon and position of the teeth, enamel organs can derive from ectoderm, endoderm, or both. Thus, several authors have put the discussion about germ layer origin of the enamel organs to rest (Chibon, 1970; Fraser et al., 2010; Donoghue & Rücklin, 2016; Calamari et al., 2018). Must we then accept that specificity of the germ layers is not a stringent criterion for homology as is sometimes assumed? Hall (1998) argues that homologous structures need not, and often do not, arise from the same germ layer. Coincidentally, teleost scales and fin rays, considered odontode derivatives (Huysseune & Sire, 1998) and, like teeth, components of the dermal skeleton, form a prime example. Elements that cover the postcranial region derive from mesodermal mesenchyme, elements in the head region from neural crest‐derived mesenchyme (Lee, Thiery & Carney, 2013; Mongera & Nüsslein‐Volhard, 2013). Common to scales, fin rays and odontodes, nevertheless, is their intimate relationship with ectodermal epithelium (Sire & Huysseune, 2003), the latter serving as an important signalling partner for the underlying mesenchyme (e.g. Aman, Fulbright & Parichy, 2018). Thus, it may well be possible that ectoderm is also required in some (hidden) form for the development of internal odontodes, as suggested by the ‘modified outside‐in’ hypothesis.
The experimental studies discussed above are rather inconclusive as to whether endoderm can develop autonomously into enamel organs without any ectodermal influence. Yet, a far more significant outcome in our view is the observation that enamel organs always develop from stratified epithelia. In several teleosts, a cell layer with peridermal characters overlies the endoderm prior to the formation of pharyngeal enamel organs. In urodeles, anteriorly positioned teeth develop from the basal (i.e. ectodermal) layer of a double‐layered epithelium, with endoderm forming the superficial layer. Even teeth that develop in an exclusive ‘endodermal territory’ develop from a stratified epithelium, with a flattened cell layer that covers the basal epithelial layer [Reisinger, 1933 and references therein; Davit‐Béal, Allizard & Sire, 2006; Davit‐Béal et al., 2007]. The superficial layer has distinct staining properties and shows apical differentiation. This is not just the case for urodele endodermal teeth but also for dentary teeth in the lungfish Neoceratodus forsteri (Kemp, 2002b ). Because teeth always develop from a stratified epithelium, one must consider the possibility that this epithelium has a composite nature. Whether the superficial layer is ectoderm or an ectoderm equivalent is discussed in the next section.
III. ECTODERMAL INFLUX OR CO‐OPTION?
There are various ways in which ectoderm could be part of the composite nature of the oropharyngeal epithelium and, in this way, continue (in evolutionary terms) to exert an influence on tooth formation. One possibility is that odontode‐competent ectodermal epithelium effectively invades the oropharynx during development and becomes associated with endoderm. This was originally proposed in the ‘modified outside‐in’ hypothesis (Huysseune et al., 2009, 2010). An obvious place for the influx of ectoderm into the oropharynx is where the ectoderm contacts the endoderm, and where an orifice is eventually created, that is at the mouth opening and the gill slits. The ectoderm may also reflect inwards around endoderm without the need for a physical opening, as in the urodele stomodaeal collar. Finally, intraepithelial migration should not be excluded. For example, Prochazka et al. (2015) showed that a fibroblast growth factor 8 (Fgf8)‐expressing epithelial cell population migrates toward a Shh‐expressing region in the mandible, the tooth placode initiation site, and that this is required for tooth development. This cell behaviour is not the only type of intraepithelial migration; diverse examples of intraepithelial cell motility are currently being uncovered (reviewed in Kapsimali, 2017). Motile epithelial cells may ultimately participate in tooth formation, by directly building the enamel organ, as a signalling partner, or by providing the necessary mechanical stimuli.
An alternative hypothesis for the invasion of odontode‐competent ectoderm is the co‐option of ectodermal characters by endoderm, facilitating the development of odontodes inside the pharynx. Such a co‐option event would most likely have been initiated at the ecto–endoderm contact zone, that is in the mouth, and at the pharyngeal openings, or gill slits. Given our emphasis on pharyngeal teeth, we focus on the latter.
(1). Ectoderm meets endoderm at pouch–cleft contacts
The endodermal epithelium of the developing pharynx produces a series of bilateral outpocketings, the pharyngeal pouches. These extend from the midline endoderm and eventually contact the ectoderm at corresponding clefts (Graham, Okabe & Quinlan, 2005; Grevellec & Tucker, 2010; Graham & Richardson, 2012; Frisdal & Trainor, 2014; Shone & Graham, 2014). [Note that the term ‘cleft’ is used here although there may not necessarily be a visible skin depression]. In chondrichthyans and non‐amniote osteichthyans, most pouch–cleft contacts eventually break through to create orifices, called gill slits (Goodrich, 1930; Bjerring, 1977; Kimmel et al., 1995; Shone & Graham, 2014). The pouches, and later gill slits, separate clusters of mesenchymal cells that give rise to, amongst others, cartilaginous pharyngeal arches. Choe & Crump (2015) provide an overview on epithelial morphogenesis at the pouch–cleft contacts, and its molecular control.
Counting the slits behind the first, or hyomandibular, pouch (i.e. pouch 1, P1), there are five in selachians. The exceptions are the sixgill shark Hexanchus (six pouches), and the sevengill shark Heptranchias (seven pouches), which however do not represent a basal condition. The persistent notion that elasmobranchs primitively had more than five gill slits is unsubstantiated by fossil evidence (Maisey, 2004). Other vertebrates have four or five gill slits beyond the first one (Goodrich, 1930; Brachet, 1935; Graham et al., 2005, 2019). In actinopterygian fishes, commonly six pharyngeal pouches form from anterior to posterior (Fig. 8A). The first pouch (P1) separates the mandibular from the hyoid arch, and is homologous to the first pouch in chondrichthyans, the spiraculum. Basal actinopterygians (e.g. Polypterus) and the lobe‐finned fish Eusthenopteron have spiracular canals, as do anuran tadpoles (Bartsch, Gemballa & Piotrowski, 1997; Haas, 2003; Brazeau & Ahlberg, 2006). Dipnoans on the other hand lack the canal and among the neopterygians, Lepisosteus retains only a rudimentary canal (Balfour & Parker, 1882; Goodrich, 1930). Initially the spiraculum may have been used for gas exchange. The stapes of Devonian stem tetrapods traverses a deep and expansive spiracular space homologous with the crown tetrapod middle ear space (Gardiner & Schaeffer, 1989; Brazeau & Ahlberg, 2005). An open spiracular canal is, however, not always present in adults; it became obliterated many times in evolution (Brachet, 1935). As in chondrichthyans, it still gives rise to an opening in species at chondrostean level (e.g. Acipenser) and in Polypterus (Brachet, 1935; Bertin, 1958; Kunz, Luer & Kapoor, 2009; Hilton, Grande & Bemis, 2011). By contrast, the spiraculum does not open any longer in neopterygians (e.g. Lepisosteus) and in teleosts (Goette, 1901; Goodrich, 1930; Brachet, 1935; Arambourg & Bertin, 1958; Bertin, 1958; Gardiner & Schaeffer, 1989).
Fig 8.
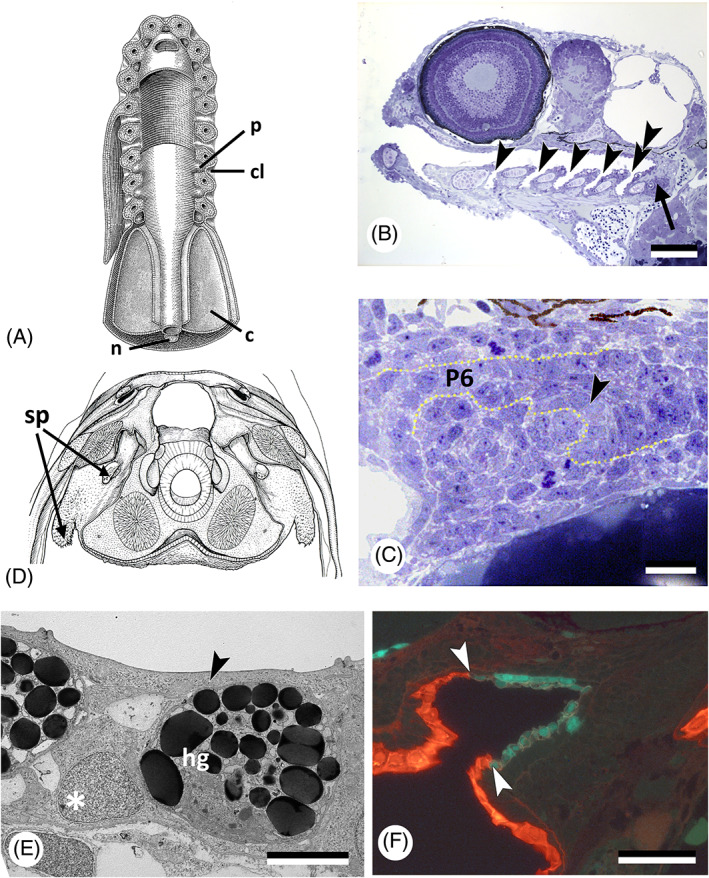
Pouch–cleft contacts and tooth formation. (A) Diagrammatic ventral view of dissected branchial region of an ‘idealised' primitive craniate. c, coelom; cl, cleft; n, notochord; p, pouch [after Bjerring, 1977, with permission from the author and publisher]. (B) Danio rerio; sagittal section showing the open pouches (gill slits) in a 96 h post‐fertilisation (hpf) specimen (arrowheads); pharyngeal teeth (arrow) are associated with the last pouch (pouch 6, double arrowhead). Scale bar = 100 μm. (C) Danio rerio; cross section at the level of the forming pharyngeal dentition in a 50 hpf specimen. Note connection of the tooth‐forming epithelium (arrowhead) with that lining pouch 6 (P6). Dotted line follows basal lamina, delimiting the pharyngeal epithelium. Scale bar = 10 μm. (D) Latimeria chalumnae; teeth formed in the spiracular pouch (sp); based on a photograph of a transverse section at the level of the intracranial articulation (modified after Millot & Anthony, 1958, with permission from the publisher). (E) Oryzias latipes; transmission electron microscopy (TEM) picture of a cross section of the pharyngeal epithelium in a 7 days post‐fertilisation (dpf) embryo, showing that a flattened cell layer (arrowhead) covers the hatching gland cells (hg) and basal endodermal layer (asterisk) of the pharyngeal epithelium. Scale bar = 5 μm. (F) Danio rerio; sagittal section through the mouth entrance of a 55 hpf Tg(krt4:tomatoCAAX;sox17:egfp) embryo, periderm (red) has partially invaded the mouth (boundary indicated by arrowheads); endoderm is green. Scale bar = 50 μm.
The second pouch (P2) separates the hyoid arch from the third pharyngeal arch (also called first branchial or gill arch), pouch 3 (P3) separates pharyngeal arch 3 from 4, and so on. The last pouch (P6) opens anterior to the seventh pharyngeal arch [see Holzschuh et al., 2005, Kopinke et al., 2006, Choe et al., 2013, Rosa et al., 2019 and Oralová et al., 2020 for timing of development of the pouches in zebrafish; Okada et al., 2016 for the Japanese medaka, Oryzias latipes; Le Pabic, Stellwag & Scemama, 2009 for Nile tilapia, Oreochromis niloticus]. The epithelium from pharyngeal arches 3–6 differentiates into gills; the seventh arch is non‐gill bearing. Once the gill slits are open, respiratory water flows over the gills.
How the pouch–cleft contacts eventually perforate varies considerably across vertebrates, although few studies have been devoted to this topic. In zebrafish, separate microscopically small lumina appear within a pouch after invasion of periderm, and subsequently coalesce. Formation of the opening is possibly aided by the secretion of hatching gland cells, at least in rainbow trout, Oncorhynchus mykiss (Gonzalez et al., 1996). Alternatively, cell interdigitations of ectoderm and endoderm produce a single cell layer that eventually ruptures, a situation encountered in the branchial membranes of the chick (reviewed in Soukup et al., 2013; Shone & Graham, 2014).
(2). Pharyngeal teeth form at ectoderm–endoderm contacts
If pouch–cleft contacts are the place where ectoderm can invade the pharynx, co‐localisation of gill slit openings and internal odontodes can be expected. Explicit statements on their co‐localisation are, however, rare. According to Smith & Coates (2001, p. 237), oropharyngeal denticles in the agnathan thelodont Loganellia occur “associated with gill pouches and slits” [see also Van der Brugghen & Janvier, 1993; Rücklin et al., 2011]. Jollie (1968) refers to gnathostome teeth that extend dorsally in each pharyngeal pouch. In Polypterus senegalus, the position of small tooth plates (with a single tooth each) that cover the inner surfaces of ceratobranchials 1–3 and the outer surfaces of all ceratobranchials corresponds with the position of the gill slits (Wacker, Bartsch & Clemen, 2001). In zebrafish, tooth buds develop from midline endoderm just posterior to where the last (sixth) pouch connects to the ectoderm (Oralová et al., 2020) (Fig. 8B, C). Several actinopterygian fish species possess oesophageal teeth (Isokawa et al., 1965). Gilchrist (1922), who studied oesophageal teeth in members of the deep‐sea fish family Stomateidae, noted that these teeth are found in two large saccular outgrowths of the alimentary tract just behind the branchial region, “suggestive of a pair of closed gill‐slits” (p. 249). By contrast, we know of no gnathostome species with teeth in the stomach or beyond. An explanation for the occurrence of oesophageal teeth is offered in Section VI.
Among sarcopterygians, the osteolepiform Eusthenopteron possessed a spiracular pouch paved with teeth (Bjerring, 1998; Brazeau & Ahlberg, 2006). Spiracular teeth are also found in the extant coelacanth Latimeria chalumnae (Fig. 8D). In this species, the absence of well‐defined rows of tooth plates associated with the dorsal parts of the gill arches has been related in part to the virtual absence of the dorsal parts of the gill slits (Nelson, 1969). In temnospondyl amphibians, one of the largest late Palaeozoic tetrapod clades, branchial denticles were common and are considered by palaeontologists as evidence for the presence of open gill slits and an aquatic lifestyle (Schoch, 2001, 2002). Branchial denticles disappeared at metamorphosis (e.g. Boy, 1988). The questionable identification of branchial denticles in extant amphibians, and the distinction between denticles and gill rakers, is discussed in Section IV.1. In amniotes, pouch–cleft contacts may well break through, but pharyngeal teeth are never present.
The observations summarised above suggest that pharyngeal teeth or denticles indeed are closely associated with pharyngeal pouches. If so, does ectoderm use pouch–cleft contact areas (or the orifices formed in them) as passageways to invade the oropharynx and participate in tooth formation? Possibly but not necessarily. The two vertebrate lineages for which details are available to address this question are teleosts and amphibians.
(a). Teleosts
Edwards (1929), studying tooth formation in the common carp, showed that the endodermal pharyngeal folds first come into contact with the basal ectodermal layer (i.e. the layer resting on the basal lamina), which responds by sending inward a wedge‐shaped plug of cells. The latter ruptures the apex of the pharyngeal fold which then becomes continuous with the basal ectodermal layer. The surface layer of the ectoderm then grows inwards as rapidly as the cleft‐like lumen in the endodermal cells proceeds inward. This results in a lining of the lumen of the pharyngeal cleft and the pharyngeal cavity composed of two cell layers: an apical layer made from flattened (i.e. ectodermal) cells, and a basal (i.e. endodermal) layer of columnar cells. The endodermal layer gives rise to the enamel organs. Rosa et al. (2019) demonstrated that in zebrafish the surface layer of the ectoderm (i.e. the periderm) indeed invades each pouch, even prior to orifice formation, but is arrested halfway into the pouch. Moreover, the invasion into pouch 6 (the pouch just anterior to the tooth‐forming region) occurs only after the first tooth germ has been initiated (recall that, instead, a periderm‐like layer overlies the midline endoderm from which teeth form) (Fig. 5F–H). Pouch–cleft contact is nevertheless required before teeth are initiated (Oralová et al., 2020). This conclusion was based on observations on vgo mutants, which carry a mutation in the transcription factor tbx1 (Kochilas et al., 2003; Piotrowski et al., 2003). These mutants display defective pouch formation and therefore disturbed pouch–cleft contact. The disturbance is often asymmetric, while the medial, unsegmented, pharyngeal endoderm (that is, where teeth normally develop) is less affected (Piotrowski & Nüsslein‐Volhard, 2000). Oralová et al. (2020) established a significant correlation between the absence of teeth in these mutants and failure of endodermal pouch 6 to contact the surface ectoderm. Teeth were present if this contact was established. A coincidental, non‐causal relationship between pouch–cleft contact and tooth initiation was excluded by the observation of unilateral teeth (i.e. teeth present only on the left or on the right side of the animal) in vgo mutants, associated with a normal pouch–cleft contact at that side only. Similar data were obtained on embryos with natural or induced pericardial oedema, which mechanically prevents contact of pouch endoderm with ectoderm. These data strongly suggest that contact of pouch endoderm with skin ectoderm is required for tooth formation. However, since teeth in zebrafish are initiated prior to any invasion of the corresponding pouch–cleft contact by cells from the body surface, a direct participation of the ectoderm in the epithelial enamel organ, via this route, appears excluded. Likewise, Atukorala et al. (2011) could not see an ectodermal influx into the pouches in Japanese medaka. Unlike Oralová et al. (2020), this led them to conclude that the rostral endoderm possesses an intrinsic odontogenic competence. Yet, as in zebrafish, the pharyngeal epithelium in medaka has a superficial layer of flattened cells, similar to the periderm‐like layer in zebrafish (Yamamoto, Iuchi & Yamagami, 1979; Fig. 8E). Interestingly, in mice, loss of the transcription factor Pax9 (Peters et al., 1998) leads not only to pouch defects, but also to defective tooth development, tooth germs being arrested at the bud stage. In zebrafish, pax9 is expressed in pharyngeal arch mesenchyme and both medial and lateral to the tooth germ in pharyngeal epithelium, but not in the tooth germ itself (Jackman, Draper & Stock, 2004). Morpholino knockdown of pax9 does not prevent the formation of teeth in zebrafish (Swartz et al., 2011), albeit teeth are reduced in number.
It is unknown whether the pouch–cleft (i.e. endoderm–ectoderm) contact that is needed for pharyngeal tooth development reflects the need for chemical (signalling) factors or for mechanical stimulation. The need for a contact is, however, also apparent from studies on other organs. Balinsky (1975) noted (without referring to a particular taxon) that ectodermal clefts are induced by endodermal pouches when they touch the epidermis but are absent if the endodermal pouch does not reach the epidermis. Holzschuh et al. (2005) observed a close correlation between enlarged or reduced contacts made by pouches with the ectoderm and epibranchial defects in vgo‐mutant zebrafish, and proposed a contact‐dependent signal from the endoderm. Specification and patterning of the pharyngeal endoderm is required for ectodermal expression of gcm2, a gene necessary for gill filament budding (Hogan et al., 2004). This expression requires Fgf3 and Fgf receptor‐like 1a (Fgfrl1a) in the pouch endoderm (Hall et al., 2006). Thymic dysgenesis in nude mice that is caused by the failure of third pouch endoderm to contact the ectoderm (Cordier & Haumont, 1980), is reminiscent of the above‐described pouch formation defects.
Oralová et al. (2020) clearly indicated that a pouch–cleft contact is necessary for tooth initiation, in addition to the presence of a layer of periderm‐like cells covering the odontogenic endodermal epithelium. The situation may be different for the oral teeth in teleosts. In cichlid fishes, the start of oral tooth formation consistently follows mouth opening (Huysseune, 1990). This sequence of events suggests that invasion of the oral cavity by ectoderm or periderm could occur prior to the start of tooth formation. Rosa et al. (2019) showed that periderm partially invades the zebrafish mouth cavity (Fig. 8F). However, given the absence of oral teeth in zebrafish (and other cyprinids), nothing can be concluded on the role of periderm or ectoderm in oral tooth formation in this species. It will be interesting to investigate whether the processes observed during zebrafish tooth development represent the general teleost, or even actinopterygian, situation – whether for oral or pharyngeal teeth. Indeed, unlike non‐teleost actinopterygians and amphibians, teleosts have a highly derived early development with meroblastic cleavage, and unique patterns of gastrulation, endoderm formation and pouch development (Collazo, Bolker & Keller, 1994). Egg type, cleavage and gastrulation have been proposed to impact the formation of the primary mouth, the other obvious place where ectoderm meets endoderm (Soukup et al., 2013). Clearly, studies on primary mouth and gill slit formation in basal, non‐teleost, actinopterygians with holoblastic cleavage are needed to acquire a broader insight into the composition of the oropharyngeal epithelium.
(b). Lissamphibians
Extant amphibians, exemplified by urodeles, have teeth along the jaw margins but no pharyngeal teeth. The oral teeth are ectoderm‐derived anteriorly, and endoderm‐derived posteriorly, some being of mixed origin (see Section II.3b; Fig. 6F). The stomodaeal region is covered by an epithelium consisting of flattened cells on top of cuboidal cells, as described for the urodele Ambystoma mexicanum (de Beer, 1947). The epithelial composition in A. mexicanum is remarkably similar to the two‐layered pharyngeal epithelium in zebrafish. Yet, different from teleosts, the flattened cells are yolk‐filled, pigment‐free and have been identified as endoderm, while the basal cuboidal cells are yolk‐free, pigment‐containing cells and represent the ectodermal stomodaeal collar. This was confirmed by Soukup et al. (2008) using transgenic axolotls, and by Takahama, Sasaki & Watanabe (1988) for the Tokyo salamander, Hynobius tokyoensis. Interestingly, in the latter species, the apical cells of the oral epithelium (and thus the endoderm) contain secretory granules as in the epidermis (Takahama et al., 1988).
Pharyngeal teeth were lost during amphibian evolution, despite the presence of pouch–cleft contacts. Larval and perennibranchiate amphibians retain an open operculum and gill clefts supported by the branchial skeleton (Coates & Clack, 1991). In Section IV.2, we will argue that the absence of teeth despite pouch–cleft contacts does not falsify the hypothesised need for ectoderm–endoderm contact to initiate tooth development.
(3). The possible co‐option of endoderm
In zebrafish, the invasion of periderm into the pharyngeal pouches is arrested halfway into the pouches and limited in extent in the mouth (see Section III.2a). The germ layer origin of the periderm‐like layer that expands along the midline of the oropharynx to the end of the oesophagus, is not known (Rosa et al., 2019). Yet, odontogenesis starts only when these periderm‐like cells have covered the endoderm‐derived prospective odontogenic epithelium. Careful observation suggests that these periderm‐like cells in zebrafish may represent a population of endodermal cells that behaves different from the original endoderm. The cells express periderm markers, raising the possibility that these cells have been co‐opted to function as a periderm. We use the term ‘co‐option’ in the sense of True & Carroll (2002, p. 66) in that “novel expression of just one or a few regulatory genes may result in the redeployment of entire sets of genes in novel contexts”. We propose that conserved ectoderm/periderm gene functions required for tooth development have been redeployed in a subset of endoderm. Thus, the periderm‐like cells express the transitory endodermal marker SRY‐box transcription factor 17 (sox17) but also keratin 4 (krt4), an intermediate filament marker expressed in the periderm (Oralová et al., 2020). Redeployment of genes in a novel context is not exceptional. Hogan et al. (2004) suggest that a transfer of expression of the transcription factor glial cells missing 2 (gcm2) occurred from ectoderm to endoderm at the origin of tetrapods. In non‐tetrapod osteichthyans gcm2 is required for gill filament development, in tetrapods gcm2 is required for parathyroid development. Thus, the parathyroid gland in tetrapods may have evolved by co‐option of ectoderm characters to endoderm, together with the transition from an aquatic to terrestrial environment (reviewed in Graham et al., 2005; Graham & Richardson, 2012).
Why periderm invasion is arrested in zebrafish, and endoderm is possibly co‐opted into a periderm‐like fate to cover the remaining (medial) part of the oropharynx, remains puzzling. Possibly, this is related to the miniaturisation of zebrafish compared to other cyprinids. Recall that Edwards (1929) saw invasion of the superficial ectodermal layer (i.e. periderm) as deep as the midsagittal plane in the much larger carp (an observation that nevertheless needs to be reassessed). It is known that miniaturisation entails a number of morphological and developmental changes, such as reductions, and structural simplifications but also innovations (Hanken & Wake, 1993). Alternatively, it is possible that the dual origin of the superficial layer of the oropharyngeal epithelium, that is from periderm (in the pouches) and from a periderm‐like layer (along the midline), is general for teleosts and relates to their derived mode of early development, as suggested in Section III.2a. At the onset of gastrulation, the outer layer of the embryo, the enveloping layer (EVL), is said to become lineage‐restricted and to give rise to the periderm only (Kimmel, Warga & Schilling, 1990), transiently covering the ectoderm. Following this model most authors consider the periderm as extra‐embryonic, and not as ectodermal. This view needs to be revised in light of recent findings. First, contrary to what was traditionally assumed, periderm cells persist in the epidermis until at least one month of age (Fukazawa et al., 2010; Fischer et al., 2014; Lee, Asharani & Carney, 2014). Second, recent findings show that the EVL gives rise to other cell types of the organism, notably to the so‐called ‘forerunner cells’ that give rise to Kupffer's vesicle (whose cells later disperse and assume caudal notochord or muscle fates) (Warga & Kane, 2018). In another example, a study on mouse endoderm has shown that segregation of extra‐embryonic and embryonic tissues is not as strict as classically accepted. Indeed, a cell lineage previously considered exclusively extra‐embryonic, contributes to the early gut tube of the mouse embryo (Kwon, Viotti & Hadjantonakis, 2008). In contrast to teleosts, non‐teleost actinopterygians such as Polypterus, Acipenser or Amia, undergo complete cleavage. A periderm does not form (e.g. Acipenser), or is less specialised than in teleosts (as in Lepisosteus and Amia) (Dettlaff, 1993). In sturgeon, for example, in the absence of a periderm layer, the ectoderm is bilayered from early stages onwards. Thus, it is tempting to consider the teleost periderm as being derived from the outer ectodermal layer in early actinopterygians, due to the peculiar and derived type of embryonic development, a suggestion also made by Warga & Kane (2018) [but see Collazo et al., 1994 for alternative scenarios for the origin of the EVL]. Significantly, in mammals, the periderm is a direct ectodermal derivative.
The ectoderm–periderm relationship outlined above allows us to speculate about a possible evolutionary scenario. In this hypothesis, the outer ectodermal layer penetrates deep into the pharynx in non‐teleost actinopterygians. During actinopterygian evolution, substantial changes are known to have taken place in the mode of embryonic development (Collazo et al., 1994). We propose that an invasion of the outer ectodermal layer would then have become more restricted to the entrance of the pouches. Instead, endoderm would have been co‐opted to take up the function of the outer ectodermal (now called peridermal) layer. A comparison of miniaturised teleosts such as zebrafish and medaka with non‐miniaturised teleost species, and between teleosts and basal actinopterygians that develop a bilayered ectoderm, will be imperative to test these ideas. How this hypothesis then relates to the origin and evolution of internal odontodes (oral and pharyngeal teeth or denticles) is further elaborated upon in Section VI.
IV. GERM LAYERS AND THE DISTRIBUTION OF POST‐MANDIBULAR TEETH
(1). The distribution pattern of teeth in the oropharynx
In vertebrates with internal gills such as chondrichthyans, actinopterygians, and sarcopterygian fishes, the pharynx can be completely covered in odontodes. In extant sharks, these are frequently very small and often referred to as (mucous membrane) denticles (Peyer, 1968) (Fig. 9A). Species of batoids (skates and rays) also possess denticles in the pharynx (Atkinson & Collin, 2012). The pattern of distribution of pharyngeal denticles varies widely, from tightly crowded, to widely scattered, depending on the species (Nelson, 1970). The denticles can be distributed throughout the oropharynx, be restricted to its posterior region, or be missing altogether. In selachians they can reach as far back as the oesophagus (Peyer, 1968). Ratfish, members of the holocephalans (a basal clade among extant chondrichthyans) have tooth plates in the oral cavity (Didier, Stahl & Zangerl, 1994) but there is no report of pharyngeal denticles (Peyer, 1968). Nelson (1969) assumed that primitively the dermal skeleton was represented by small toothed elements distributed in a nearly uniform manner over the surfaces of the oropharyngeal cavity, a condition possibly represented by pharyngeal denticles scattered over the oropharyngeal cavity in elasmobranchs (illustrated for recent forms in Nelson, 1970). Not surprisingly, shark teeth and denticles are the most commonly collected craniate fossil material (Maisey, 2000). There is, however, a discussion as to what extent the condition in early sharks is representative for tooth development in much older early gnathostomes (Reif, 1978b ; Smith & Coates, 2001; Donoghue & Rücklin, 2016; Haridy et al., 2019; Rücklin et al., 2021). Large tooth‐bearing elements occurring in various locations on the gill arches would have arisen from smaller elements by enlargement and/or fusion. This was accompanied by reductions elsewhere in the visceral apparatus (Nelson, 1969).
Fig 9.
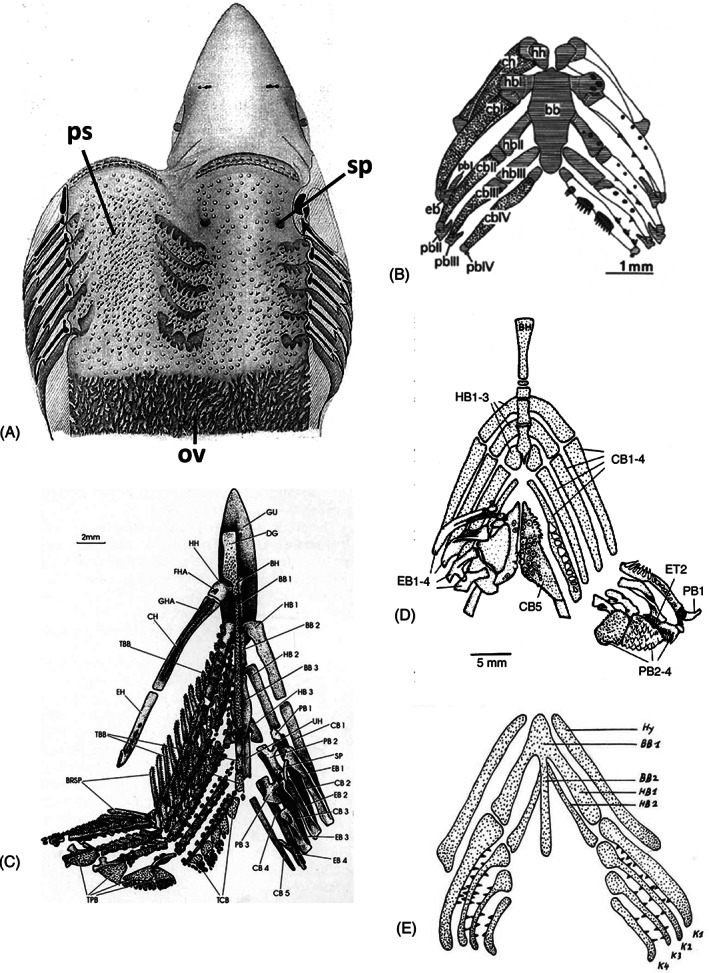
Distribution of pharyngeal teeth. (A) Squalus acanthias; adult pharynx. ov, oesophageal villi; ps, ‘placoid scales’; sp, spiracle (after Cook & Neal, 1921). (B) Polypterus senegalus; visceral skeleton and dentition of pterolarva. bb, basibranchial; cb, ceratobranchial; ch, ceratohyal; eb, epibranchial; hb, hypobranchial; hh, hypohyal; pb, pharyngobranchial (from Wacker et al., 2001, with permission from the publisher). (C) Elops lacerta; visceral skeleton and dentition. Tooth plates associated with (amongst others): BB, basibranchials; HB, hypobranchials; CB, ceratobranchials; EB, epibranchials; PB, pharyngobranchials (after Taverne, 1974). (D) Haemulon sciurus; visceral skeleton and dentition. BH, branchiohyoideus; CB, ceratobranchial; EB, epibranchial; ET, epibranchial tooth plate; HB, hypobranchial; PB, pharyngobranchial [after Wainwright, 2006, with permission from the author and publisher]. (E) Triton alpestris; visceral skeleton and dentition of stage 50, ‘Reusenzähnchen’, structures considered by the author to be true tooth germs. BB, basibranchial; HB, hypobranchial; Hy, hyoid; K, ceratobranchial [from Wagner, 1955; reproduced with permission from the Journal of Embryology and Experimental Morphology].
Extant lineages that are close to the osteichthyan key divergence into ray‐finned (actinopterygians) and lobe‐finned fishes (sarcopterygians) are represented by bichirs (Polypterus), sturgeons (Acipenser) and paddlefish (Polyodon), and gars (Lepisosteus, Atractosteus). Pharyngeal teeth in Polypterus senegalus are associated with the basibranchial, hypobranchials 1–3, ceratobranchials 1–4, and pharyngobranchials 2 and 3 (Clemen, Bartsch & Wacker, 1998) (Fig. 9B). Adult sturgeons are edentulous (Peyer, 1968; Hilton et al., 2011; Warth et al., 2017) but juvenile sturgeons possess teeth on the dentary, dermopalatine (= maxilla), palatopterygoid, elements of the hyoid arches, hypobranchial 1 and the region around infrapharyngobranchial 1 (= the parasphenoid tooth plate) (Hilton et al., 2011; Warth et al., 2017). Likewise, young American paddlefish (Polyodon spathula) carry teeth on upper (dermopalatine and palatopterygoid) and lower (dentary) jaw, as well as on the hyoid and first two branchial arches (hypobranchials, epibranchials and infrapharyngobranchials), but adults are edentulous (Smith et al., 2015). The Cuban gar (Atractosteus tristoechus) possesses pharyngeal teeth at least in the posterior pharynx (Comabella et al., 2012). Amia calva and Lepisosteus spatula have a significantly developed upper pharyngeal dentition (Nelson, 1969).
In teleosts, pharyngeal tooth distribution is extremely variable (Nelson, 1969, 1970; Vandewalle et al., 1994) (Fig. 9C, D). The upper pharyngeal dentition in teleosts consists in part of 1–5 usually paired tooth plates or groups of tooth plates (Nelson, 1969). Across the different lineages of teleosts, there was a – probably independent – loss of tooth plates. The repeated reduction, in particular of the fourth and fifth upper pharyngeal tooth plates, appears to be a general trend towards consolidation of the extensive upper pharyngeal dentition of basal teleosts such as Elops lacerta (Fig. 9C), into a single pair of toothed areas (Fig. 9D). This would be formed mainly (if not entirely) by the third infrapharyngobranchials (Nelson, 1969; Vandewalle et al., 1994). The evolution of the dentition associated with the ventral parts of the gill arches is more complex. This dentition shows secondary modifications, reductions and losses, as is extensively discussed by Nelson (1969). Again, Elops is considered to display the basal teleost condition. Based on what is known about the dermal skeleton of the gill arches, Nelson (1969) assumes that the other toothed surfaces of the buccopharyngeal cavity arose by ‘consolidation’ of tooth plates primitively supported by the visceral endoskeleton (Fig. 9C). Reduction in the pharyngeal dentition took place within the ostariophysans (Nelson, 1969; Vandewalle et al., 1994; Stock, 2007). A dentition restricted to the fifth ceratobranchials [i.e. the last (7th) pharyngeal arch] is characteristic of all families in the order Cypriniformes (including the speciose family Cyprinidae that includes zebrafish). These ventral pharyngeal teeth are the only teeth left as the oral teeth also disappeared (Huysseune, Van der heyden & Sire, 1998; Stock, 2007; Pasco‐Viel et al., 2010). Even further reduction took place in another cypriniform family, the Gyrinocheilidae, which lacks pharyngeal teeth (He, Yue & Chen, 1997). A reduction in the extent of distribution of pharyngeal teeth is a general evolutionary trend (Shkil et al., 2010), but it can also be observed within individual lineages, as for example in clupeids (Nelson, 1967).
Little is known about the distribution (and especially reduction) of teeth and denticles at the transition from lobe‐finned fishes to tetrapods. Eusthenopteron possesses well‐developed marginal tooth series, palatal denticle fields as well as large fangs on the palatines, vomers and lower jaw (Smith & Coates, 2000; Doeland et al., 2019). Like Eusthenopteron, its Australian relative Mandageria has denticulated plates on the anterior basibranchial, hypobranchials and ceratobranchials (Johanson & Ahlberg, 1997). Latimeria has an elaborate dentition on upper and lower jaws as well as in the branchial region (Millot & Anthony, 1958; Nelson, 1969). Each branchial arch is ornamented with rows of tooth plates of around 20 teeth, two rows on the inner face, two on the outer. The hyoid arch has one row of tooth plates on its inner surface, and the copula is paved with hundreds of miniature teeth. Moreover, two groups of teeth are placed in the spiracular cavity (Millot & Anthony, 1958; Fig. 8D). Acanthostega has palatal bones carrying a range of tooth‐like structures, described as shagreen, fangs, toothlets and denticles (Clack, 2002). Palatal teeth appear to have been conserved better than branchial and oropharyngeal tooth plates (Smith & Coates, 2000).
Teeth in extant amphibians can be localised to the lower jaws (dentaries), upper jaws (premaxillae, maxillae) and palate (vomers, palatines). Caecilians possess teeth on the premaxillae, maxillae, vomers, palatines, and dentaries. Salamanders retain teeth on both upper and lower jaws and on the palate, but anurans, with one exception, have lost dentary teeth and have a reduced number of palatal teeth; toads are edentulous (reviewed in Davit‐Béal et al., 2007; Berkovitz & Shellis, 2016). Metamorphosis, if present, entails major changes in the dentition. As described below, larval and fetal stages of some urodeles and caecilians have a dentition very different from that of the adult (Parker & Dunn, 1964).
Many temnospondyl amphibians possessed branchial denticles, considered to be probably homologous to palatal and pharyngeal teeth of actinopterygians (Schoch, 2002). In extant amphibians, denticles disappeared from the pharynx. Exceptionally, teeth have been reported to occur in the pharynx of urodeles. Worthington & Wake (1971, p. 358) mention the frequent presence of a “mineralized gill raker, resembling a tooth crown” on epibranchials 1 and 2 in the ambystomid salamander Rhyacotriton olympicus. It remains questionable whether these are teeth and not gill rakers. Stadtmüller (1924) made a study of the development of gill rakers in salamanders, and considered them to be homologous to teeth. Wagner (1955) reported the presence of tooth structures (“Kiemenzähnchen”) on the gill arches (but not the hyoid arch) in Triton alpestris larvae (and indeed in most urodele larvae). He considered them ‘true teeth’ (“echte Zahnanlagen”), that disappear with the closure of the gill slits (Fig. 9E). These teeth consist of a collagenous matrix only, presumably predentine. The matrix does not become mineralised and there is no enamel cap. These structures do not erupt, but eventually degenerate. By contrast, Duellman & Trueb (1986) mention only gill rakers in larval salamanders. Schoch (2001) distinguishes gill rakers from branchial ossicles, tooth‐like structures located in the throat and gill pouches. In apparent contradiction to Schoch (2002), Schoch & Witzmann (2011) consider tooth‐bearing ossicles (“pharyngeal jaws”) to be not identical and not homologous to teeth and report them to be absent in lissamphibians, but common in temnospondyls. Clearly, more data about gill rakers in actinopterygians and sarcopterygians are required for an unambiguous terminology and to assess their homology to internal odontodes. Interestingly, Schoch & Witzmann (2011) assign gill rakers to the endoderm and ossicles to the ectoderm.
In reptiles, the dentition is restricted to the oral region. There is a single row of marginal teeth on the lower jaw, but a more widely varying pattern on the upper jaw and palate (reviewed in Richman & Handrigan, 2011). The first dental elements are rudimentary papillae called denticles; they consist of dentine but little or no enamel. These teeth never become functional and are resorbed.
(2). The loss of teeth from the post‐mandibular arches
Having argued for pouch–cleft contacts (prospective gill slits) as a condition for pharyngeal tooth development, can this explain the distribution of post‐mandibular teeth? Yes and no. Yes, because teeth are found near pouch–cleft contacts and prospective orifices, as reviewed above. No, because neither the presence of pouch–cleft contacts per se, nor their sequence of formation, coincides with the diverse patterns of pharyngeal tooth distribution across taxa.
How, then, can we reconcile the observation that pouch–cleft contacts are required for tooth formation, with the fact that there is no one‐to‐one relationship between pouch–cleft contact and tooth distribution? Arguably, pharyngeal teeth in jawed vertebrates were once widely distributed over all pharyngeal arches. Because of their poor mineralisation and deep location, data on gill skeletons in fossil early gnathostomes are rare (Pradel et al., 2014; Brazeau et al., 2017), and this is even more so for pharyngeal denticles, although advanced imaging techniques may expand our knowledge in the years to come. Within the crown gnathostomes, pharyngeal teeth or denticles are limited to chondrichthyans and osteichthyans (Rücklin & Donoghue, 2019). The basal condition in recent sharks is argued to be denticles covering much or all of the buccopharyngeal surface (Nelson, 1970). In osteichthyans, inference about the evolution of tooth distribution is easier because teeth are attached in some way to the arch skeleton. The basal condition is tooth plates distributed over the different arches (Nelson, 1969; Taverne, 1974; Vandewalle et al., 1994). Thus, rather than arches gaining the capacity to form teeth, one must assume that the capacity to form teeth was lost from some arches (despite the presence of pouch–cleft contacts) but retained on other arches. Stock (2001) discussed potential mechanisms accounting for evolutionary tooth loss: developmental arrest, failure of tooth replacement, or alterations in the factors eliciting tooth initiation or later stages of tooth development. In a follow‐up study addressing oral tooth loss in cypriniforms, Jackman & Stock (2006) proposed that changes in trans‐acting regulators of the distal‐less‐like (Dlx) genes may have been responsible for loss of teeth associated with the mandibular arch. Fgf signalling was identified as one of the potential upstream regulators of oral dlx2b expression (Stock, Jackman & Trapani, 2006). An unanswered question is whether such a mechanism is also responsible for tooth loss on the other pharyngeal arches.
Still, wherever pharyngeal teeth are present, one may expect the sequence of tooth formation to follow the sequence of formation of pouch–cleft contacts. Given the diversity of pharyngeal tooth distribution, teleosts are excellent candidates to test this relationship, provided the species has teeth on more than one pharyngeal arch. In medaka, pharyngeal teeth start to appear around stage 29 on the fourth pharyngobranchial (Debiais‐Thibaud et al., 2007) while the corresponding, fifth, pouch forms at stage 27 (Okada et al., 2016) (staging according to Iwamatsu, 2004). In the Nile tilapia, pouch–cleft contacts are established between 30 and 60 h post‐fertilisation (hpf), while pharyngeal tooth buds are first reported at 96 hpf (Le Pabic et al., 2009). Obviously, more taxa need to be explored to assess the relationship between timing of pouch–cleft contact, and onset of pharyngeal tooth formation, as well as their dependence on shared genetic regulatory pathways. Should such studies reveal a significant relationship, then both events might well represent an example of co‐evolutionary patterning – analogous to what has been suggested for teeth and taste buds (Bloomquist et al., 2015). In the definition of co‐evolution adopted here, the evolution of one entity is closely affected by that of another with which it interacts (Wilkins, 2002). The concept applies to different levels of biological organisation, from molecules to ecosystems, but has rarely been applied to vertebrate organs.
Other than timing and extent of pouches and pouch–cleft contacts, one must also consider molecular regulation of anterior–posterior patterning of the endoderm in determining the sequence of pharyngeal tooth formation. The proximodistal and anteroposterior polarity of the pharyngeal arch skeleton is dictated by the endoderm (reviewed in Frisdal & Trainor, 2014; Graham & Shone, 2019). This is in line with the observation that the polarity of the branchial pouch endoderm is established in the embryo in the absence of neural crest cells (Veitch et al., 1999).
(3). A special identity for pouch 2?
In zebrafish, periderm cells (covering the basal ectodermal layer of the skin) start by invading pouch 2 (the pouch that separates the hyoid from the first branchial arch) at an early developmental stage. Invasion is soon followed by lumenisation of this pouch and opening of the pouch to the exterior. These periderm cells, characterised by krt4 expression, merge with krt4‐expressing periderm‐like cells located in the pharynx midline at the level of pouch 2. Once these two populations of krt4‐expressing cells have made contact, krt4‐expressing cells expand posteriorly towards the gut entrance, as reviewed in Section II.2b (Rosa et al., 2019). A mature pharyngeal epithelium, composed of an endodermal layer covered by a periderm‐like layer, is crucial for pharyngeal tooth development in zebrafish.
The apparent involvement of pouch 2 in these events thus raises the question whether pouch 2 plays a key role in the evolutionary origin of the pharyngeal dentition. The early invasion and opening of pouch 2 in zebrafish may be independent of the behaviour of the periderm‐like cells. It may simply be related to the sequential maturation of the pouches, given that pouch 1 does not fully open any more in most taxa. Yet, several lines of arguments indicate that this is not the (only) explanation and that pouch 2 has a more significant role. In gnathostomes, pouch 2 separates the jaw–hyoid apparatus, functioning in feeding, from the branchial arches, functioning in respiration (Starck, 1982; Stundl et al., 2019). Thus, the importance of pouch 2 in evolution can be easily understood from a functional viewpoint. Aspects of the development of the first two pharyngeal pouches (and mandibular and hyoid arches) appear to be highly conserved in all vertebrates. The first two pharyngeal pouches form first and concomitantly. The posterior pouches form consecutively along with caudal extension of the pharyngeal endoderm (Graham & Shone, 2019). Their development is controlled by distinct signalling pathways (Graham & Shone, 2019). Consequently, mutations, knockdown of genes, or pharmaceutical inhibition experiments affect mandibular and hyoid arches differently from the posterior arches and pouches [several examples and references in Okada & Takada, 2020; see also Hall et al., 2006; Birkholz et al., 2009; Schwend & Ahlgren, 2009; Vaccari et al., 2010; Lovely et al., 2016; Duncan et al., 2017]. In all vertebrates, the thyroid develops from pouch 2 whereas other pouch derivatives show larger variation (Graham et al., 2005). In further support of the evolutionary significance of pouch 2 are developmental features in amniotes that reflect the evolutionary ‘memory’ of this pouch, such as an embryonic operculum (although without dermal bones) that persists in amniotes (Wall & Hogan, 1995; Richardson et al., 2011; Graham & Richardson, 2012). In humans, the second (hyoid) arch expands caudally to cover more posterior pouch clefts (as does the operculum in teleosts; Richardson et al., 2011). Second branchial cleft lesions account for 95% of all branchial cleft malformations in humans (Ford et al., 1992; Waldhausen, 2006; Adams et al., 2016).
In conclusion, while all pouches are serial homologues, they do not all appear to be equivalent in terms of potencies. How much this impinges on pharyngeal tooth distribution is largely unexplored. It will be important to assess the relationship between pouch 2 development, its contact with ectoderm, the extent of invading periderm, and the distribution of pharyngeal teeth in selected taxa that cover key steps in vertebrate evolution.
The early opening of pouch 2 before opening of the mouth in zebrafish is remarkable. Possibly this is a general character of actinopterygians as it is also observed in the sturgeon, Acipenser sturio and A. ruthenus (Goette, 1901; A. Huysseune, personal observations), and in teleosts other than zebrafish, for example in trout, Trutta fario (Moroff, 1904), Atlantic salmon, Salmo salar (A. Huysseune and P.E. Witten, personal observations), and European seabass, Dicentrarchus labrax (Sucré et al., 2009). Early pouch 2 opening also occurs in chondrichthyans, for example in selachians such as Scyllium sp. (Brachet, 1935). Stundl et al. (2019) showed acceleration of the entire hyoid arch segment in bichirs as a function of the development of external gills, further emphasising the crucial role of this pharyngeal segment.
V. MOLECULAR CONTROL REGULATING THE DISTRIBUTION OF POST‐MANDIBULAR TEETH
(1). Genes expressed in pouches and at pouch–cleft contacts
Above, we highlighted the importance of the endodermal pouches and their contact with the ectoderm for pharyngeal tooth formation. Is it possible to identify key genes responsible for tooth development by interfering with genes that are specifically expressed in pouches or at the pouch–cleft contact? In our view, the absence of such genes or their function should compromise the odontogenic competence of the pharyngeal epithelium in species with a pharyngeal dentition. Clearly, we have data about genes that interfere with tooth development in general. However, data specifically related to genes expressed in the pouches are scarce and those available come from zebrafish. Unfortunately, while some studies report defects in the pouches upon knocking down or knocking out an endodermally expressed gene, the phenotype of the teeth is rarely studied. For example, knockdown of vgll2a results in aberrant endodermal pouch morphogenesis, and hypoplastic cranial cartilages, but the phenotype of the teeth was not reported (Johnson et al., 2011). In a rare study considering both teeth and pouches, Jackman et al. (2004) mention tooth defects along with aberrant pouch morphology when inhibiting Fgf signalling. Wiweger et al. (2012) list several zebrafish mutants based on alizarin red S‐stained whole mounts, most of which do not display a tooth phenotype, and none is reported edentulous.
Two caveats should be added when critically reviewing reported tooth phenotypes. One concerns the visualisation of the teeth in minute embryos such as of the zebrafish. First, most studies that examine the pharyngeal dentition rely on alizarin red S‐stained specimens, but these reveal only the final steps of odontogenesis. Stages of initiation, morphogenesis and early cytodifferentiation up to early mineralisation of the matrix remain unnoticed. Moreover, first‐generation teeth are extremely small, and imaging of the zebrafish dentition often requires sophisticated methods (Bruneel et al., 2015). The second caveat concerns the usefulness, in some cases, of marker genes to assess the presence of teeth. For example, pitx2 has been reported to be one of the earliest dental epithelial markers for teeth in teleosts (Fraser, Bloomquist & Streelman, 2008). However, blocking Fgf signalling by the receptor tyrosine kinase inhibitor SU5402 in zebrafish, or Shh signalling by cyclopamine A, does not abolish pitx2 expression, yet tooth germs are no longer morphologically discernable (Jackman et al., 2004; Jackman, Yoo & Stock, 2010).
Signalling between ectoderm and pouches has been shown to be necessary for gill development (Hanaoka et al., 2004; Hogan et al., 2004; Okabe & Graham, 2004), as well as epibranchial placode development in various vertebrates [zebrafish (Holzschuh et al., 2005); mouse (Rizzoti & Lovell‐Badge, 2007); reviewed in Graham et al., 2005 and Graham, 2008]. Molecular players, which include Fgfs and bone morphogenetic proteins (Bmps), have been identified. A detailed analysis of signalling at the pouch–cleft contact is nonetheless hindered by the fact that the boundary between pouch endoderm and ectoderm is not easily detected. Also, expression data are usually collected through whole‐mount in situ hybridisation, preventing precise localisation of the signal (ectoderm or endoderm).
In conclusion, in contrast to gills and epibranchial placodes, no key genes have so far been identified that could act at the pouch–cleft contact to elicit tooth formation. There is nevertheless one candidate molecule, retinoic acid, which has been proposed as a molecular explanation for the distribution of post‐mandibular teeth. This will be discussed below.
(2). Tooth distribution: the role of retinoic acid and sonic hedgehog
Tooth development depends on reciprocal epithelial–mesenchymal interactions, which are mediated by the Wnt (Wingless/integrated), BMP, FGF, Shh (Sonic hedgehog), and Eda (ectodysplasin) pathways (Balic & Thesleff, 2015). Experiments that alter Wnt signalling in teleosts have produced conflicting results. No effects on (pharyngeal) tooth number or distribution were reported in zebrafish (Huysseune, Soenens & Elderweirdt, 2014) while (oral) tooth density decreased by antagonising Wnt signalling in cichlids (Bloomquist et al., 2015). In zebrafish, supernumerary pharyngeal teeth develop when Fgf signalling is up‐regulated (Gibert et al., 2010; Jackman et al., 2013), when Bmp signalling is down‐regulated (Jackman et al., 2013) or when Eda is overexpressed (Aigler et al., 2014). Supernumerary teeth can also be generated in zebrafish by thyroid hormone level manipulation (Woltmann et al., 2018). Remarkably, in none of these cases do teeth expand over the other (i.e. more anterior) arches. To the best of our knowledge, only manipulation of signalling through retinoic acid expands the dentition to anterior arches (Seritrakul et al., 2012).
Retinoic acid (RA) is a morphogen with a key role in many tissues during several developmental stages. Importantly, RA regulates Hox genes, which play a crucial role in axial patterning (reviewed in Nolte, De Kumar & Krumlauf, 2019). Gibert et al. (2010) demonstrated that pharyngeal tooth formation in zebrafish depends on RA. Interestingly, a few years earlier, Kopinke et al. (2006) showed that RA signalling is necessary for correct morphogenesis and segmentation of the endodermal pouches. Combining these two observations, one could conceive that tooth formation is only indirectly influenced by RA, namely by depending on proper formation of pouch–cleft contacts. Remarkably, Gibert et al. (2010) reported that pharyngeal tooth induction was not compromised by RA inhibition in members of two other lineages of teleosts, the beloniform Oryzias latipes (Japanese medaka), and the characiform Astyanax mexicanus (Mexican tetra). The authors concluded that in these two species, oral and pharyngeal teeth are induced independently of RA, and proposed that RA‐dependent tooth initiation was acquired in the cypriniform lineage. However, Gibert et al. (2010) commenced the RA inhibition treatment in medaka at the end of somitogenesis. Since the pharyngeal pouches are formed well before the end of somitogenesis [fifth pouch at stage 27 (Okada et al., 2016); somite completion at stage 32 (Iwamatsu, 2004)], the appropriate signals for tooth formation may have been present prior to the inhibition experiment. Clearly, RA‐inhibition experiments, fine‐tuned against the timing of pouch–cleft contact, will be required in multiple species to assess the role of pouch–cleft contacts versus direct RA signalling. In a follow‐up experiment, Seritrakul et al. (2012) applied exogenous RA to zebrafish embryos from 24 hpf onwards and demonstrated an expansion of teeth to anterior arches. The authors concluded that changes in the levels of RA may be an evolutionary mechanism controlling tooth distribution, and suggested this could be the result of altered neural crest development. In another set of experiments, Gibert et al. (2015) applied exogenous RA to zebrafish later in development (56 hpf and older), and examined adult zebrafish heterozygous for a mutation in cyp26b1, a gene encoding an enzyme involved in RA degradation. This resulted in an additional tooth in the ventral row, anterior to the tooth in position 1V. The expansion of the dentition was accompanied by expansion of the mesenchymal markers dlx2a and lhx6 and the dental epithelium marker pitx2a. The increase in tooth number was limited to the tooth‐bearing arch only, without anterior expansion over the other arches. It was concluded that RA signalling can possibly intervene in the fine‐tuning of tooth number at a taxonomically restricted level. Given our focus on the dental epithelium, it is interesting to note that RA regulates the developmental potential of the anterior endoderm (respiratory versus pharyngeal) in two other vertebrates, Xenopus laevis and mouse (Rankin et al., 2018).
Sonic hedgehog (Shh), a signalling molecule from the Hedgehog family, is another candidate for regulating tooth patterning. In modern elasmobranchs, expression of shh marks the prospective loci of tooth initiation within the dental epithelium (Smith et al., 2009; Maisey et al., 2014). In zebrafish, the paralogue shha is widely expressed in the pharyngeal epithelium, both in tooth‐forming and adjacent, non tooth‐forming, regions (Jackman et al., 2010). Its broad expression domain contrasts with the restriction of Shh expression to the dental epithelium in the oral teeth of mice (Hardcastle et al., 1998). Tooth development in zebrafish depends on shha. However, overactivation of shha signalling yields no observable effect on tooth development (Jackman et al., 2010). On the other hand, cyclopamine A, a compound that inhibits hedgehog signalling by binding to the Smoothened receptor, blocks tooth formation on the last (i.e. tooth‐bearing) arch when applied early (Jackman et al., 2010).
Recently, El Shahawy et al. (2017) reported a highly sensitive balance between SHH and RA signalling. In the mouse tongue, SHH antagonises RA signalling by maintaining expression of two RA catabolic enzymes Cyp26a1 and Cyp26c1. RA, on the other hand, controls Shh expression, for example during zebrafish fin regeneration, or in the pharyngeal region of flounder, Paralichthys olivaceus (Laforest et al., 1998; Suzuki, Oohara & Kurokawa, 1999; Sehring & Weidinger, 2019). These findings, along with the observation that tooth development in zebrafish can be elicited on normally edentulous arches by the overactivation of RA, hints at a possible epistatic relationship between these pathways. Whether such a proposed balance between RA and SHH plays a role in the distribution of post‐mandibular teeth can be tested experimentally.
VI. TOWARDS A UNIFYING CONCEPT FOR GERM LAYER INVOLVEMENT IN TOOTH INITIATION
Odontodes are outstanding examples of vertebrate characters with highly conserved development, structure and composition. The fossil record provides ample evidence for the homology of teeth and external odontodes, including transitional forms (Blais et al., 2011; Chen et al., 2020), as well as support that internal odontodes evolved through the expansion of odontogenic competence from external to internal epithelia (Rücklin et al., 2011; Donoghue & Rücklin, 2016). External odontode development in early vertebrates undoubtedly required the interaction between ectoderm and mesenchyme. The question is, was an interaction of mesenchyme with ectoderm still required when odontodes started to appear in the oropharyngeal cavity, a tenet of the ‘modified outside‐in’ hypothesis? The fossil record is naturally silent about this question (Janvier, 2015). Furthermore, is ectoderm in some form (still) required to allow tooth formation inside the oropharynx? Ongoing studies on oropharynx and tooth development in extant vertebrates can possibly provide answers.
Some of the studies presented in the previous sections appear, at first sight, to contradict the requirement for ectoderm in the formation of internal odontodes (teeth or pharyngeal denticles). For example, the experiments of Adams (1924) and of Cassin & Capuron (1979) in urodele amphibians suggested that endoderm alone can form (oral) teeth in conjunction with neural crest. Challenging these ideas, it was recently shown that pharyngeal teeth in zebrafish develop from endodermal epithelium but only where covered by a superficial layer with periderm‐like characters (Oralová et al., 2020).
Below, we propose a hypothesis that attempts to integrate the different and often conflicting results. The observations on lungfish dentitions are inspiring in this context. Here the cells that line the oral cavity are described to be of endodermal origin. Yet, the superficial cells later metabolise their yolk and acquire characters similar to cells of the outer layer of the external epidermis, such as the appearance of microvilli and tight junctions, while the enamel organ differentiates from a basal layer that maintains its yolk as the tooth primordia begin to form (Kerr, 1903; Kemp, 2002a , 2003).
The scenario that we propose for the evolutionary developmental biology of pharyngeal teeth or denticles is described below and depicted in Fig. 10A–D'. In the primitive situation, represented by fossil taxa, skin denticles are proposed to have enamel organs derived from the basal layer of a bi‐ (possibly multi‐) layered ectodermal epithelium; endodermal pouches contact the ectoderm (Fig. 10A). At some point, perhaps even before the advent of jawed vertebrates, enamel organs are suggested to have appeared in the endoderm of the pouches, likely as the result of transfer of competence from the ectoderm. However, it is hypothesised that this transfer only happened if and where the endoderm maintained an interaction with ectoderm, resulting from the invasion of the outer ectodermal layer (the periderm) into the oropharyngeal cavity (as in Fig. 10C). The recent study of Chen et al. (2020) on the expansion of dermal denticles in the stem osteichthyan Lophosteus superbus illustrates how morphogenetic movements of cell layers may play an important role in the evolutionary origin of novel structures.
Fig 10.
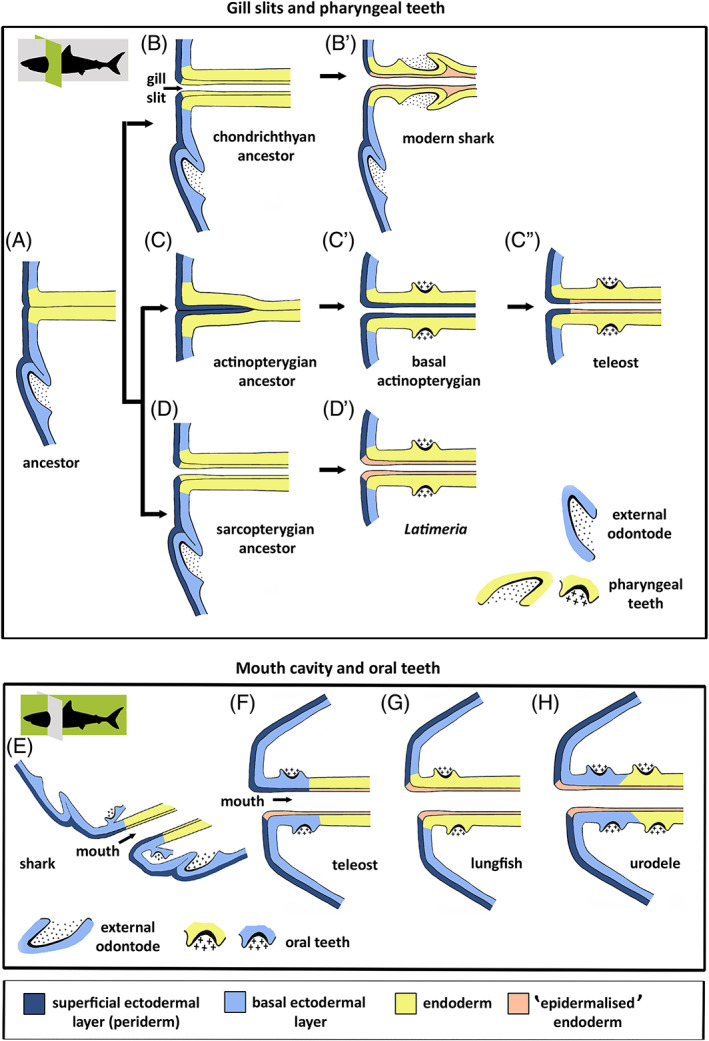
Germ layer contribution to pharyngeal and oral teeth. (A–D′) Hypothetical evolutionary scenario for the persisting influence of ectoderm on pharyngeal tooth formation. (A) Primitive situation; external odontode enamel organ derives from basal ectodermal layer; endodermal pouches contact the ectoderm and endoderm partially spreads under the periderm. (B, B′) In the chondrichthyan lineage, endoderm of the pouches is stratified and the superficial layer is ‘epidermalised'; pharyngeal teeth derive from the basal endodermal layer (modern sharks, B′). (C–C″) In the actinopterygian lineage, periderm invades the pouches, ultimately covering the entire pharyngeal endoderm (basal actinopterygians, C′); in advanced actinopterygians, periderm invasion is restricted to the distal parts of the pouches, while endoderm is ‘epidermalised' medially; pharyngeal teeth derive from the basal endodermal layer (teleosts, C″). (D, D′) In the sarcopterygian lineage, endoderm of the pouches is stratified and the superficial layer is ‘epidermalised'; pharyngeal teeth derive from the basal endodermal layer (Latimeria chalumnae, D′). It is possible that basal actinopterygians took path D, leading to the situation in teleosts (i.e. periderm invasion would be a teleost novelty rather than a regressive feature). A study of basal actinopterygians will resolve this issue. (E–H) Schematic representation of germ layer contribution to oral tooth formation in extant vertebrates, as inferred from literature data. (E) Shark; dental lamina derived purely from ectoderm. (F) Teleost; both periderm and periderm‐like layer may constitute the superficial layer of the oral epithelium; enamel organs putatively derived from ectoderm. (G) Lungfish; the oral cavity is lined by endoderm, which also gives rise to the enamel organs. However, the superficial cells have acquired characters similar to cells of the outer epidermal layer. (H) Urodele amphibian; the basal ectodermal layer invades the mouth, oral teeth derive from the basal layer (either ectoderm or endoderm) but are covered by ‘epidermalised' endoderm. Shark profiles show the plane of sectioning (in green): cross sections for gill slits and pharyngeal teeth (upper panels), sagittal sections for mouth cavity and oral teeth (lower panels).
In chondrichthyans, which can develop an extensive covering of denticles in the oropharynx, the superficial layer of endoderm is proposed to have been co‐opted into a layer with ectoderm‐like characteristics (Fig. 10B–B'). Interestingly, Cook & Neal (1921, p. 45) described the pharyngeal epithelium in Squalus acanthias (a shark with pharyngeal denticles) as follows: “the pharyngeal epidermis resembles that of the skin rather than epithelium such as that of the stomach and intestine”. Peyer (1968, p. 72) suggested that the distribution of “mucous membrane” denticles (his terminology) in sharks may be caused by “boundary displacements during embryonic development”, with which he indicated a shift of the posterior limit of the stomodaeum. A possible interpretation for chondrichthyan oral teeth is shown in Fig. 10E.
In teleosts, the pharyngeal epithelium is bilayered in taxa that represent both basal and highly evolved members of the group: members of the anguillids, salmonids, cyprinids, and cichlids (Rosa et al., 2019). Provided that zebrafish can be considered to represent the teleost situation, the basal layer – the layer that produces the enamel organ – is endodermal; it is covered in the oropharynx and oesophagus by a superficial layer with periderm‐like characters. This layer covers the endoderm prior to periderm invasion via the pouch. Again this possibly represents a case of co‐option of endoderm (Fig. 10C"). The extent of the periderm‐like layer (down to the oesophagus, but not further) delimits the area from which enamel organs can develop (thus, including the oesophagus, explaining the oesophageal teeth observed in some teleosts). Interestingly, it coincides with the demarcation between the pharyngeal endoderm anteriorly and the presumptive alimentary canal posteriorly, which develop independently from each other (Ober, Field & Stainier, 2003; Wallace & Pack, 2003; Kobayashi et al., 2006; Sucré et al., 2009). Note, in this context, that extra‐oral teeth in teleosts arise in the skin ectoderm only (Sire & Allizard, 2001), never in the gut. Whether a restricted invagination of periderm into the pouches is a teleost‐specific regressive feature (Fig. 10C–C"), or whether co‐opted endoderm (Fig. 10D–D') is rooted deep in actinopterygian or even osteichthyan phylogeny, can be tested by studying basal actinopterygians with less‐specialised early embryonic development. Unfortunately, these taxa are largely understudied in relation to the composition of ectodermal (skin) and oropharyngeal epithelia. Likewise, teleost species other than cyprinids need to be tested to assess germ layer contributions to oral teeth (Fig. 10F).
With respect to sarcopterygians, in dipnoans (which have oral teeth only), the enamel organs form from endoderm but also here the upper layer adopts features of a superficial ectodermal layer, as commented on in Section II.3a (Kerr, 1903; Kemp, 2002a , 2003) (Fig. 10G). A name that we suggest for this condition is ‘epidermalised endoderm’, similar to ‘ectodermalised endoderm’, a term that was coined by Hayashi (1972) for the events occurring at opening of the stomodaeum in the newt Triturus pyrrhogaster (“ectodermalization of endodermal cells”). In urodele amphibians, the oral teeth can form from ectoderm, from endoderm or have a mixed origin (Adams, 1924; Chibon, 1970; Soukup et al., 2008). Experimental data have shown that endodermal teeth can develop fairly independently from ectoderm, but that, by contrast, ectodermal teeth need endoderm (Adams, 1924, 1931), a situation that seems at odds with our claim of the need for ectoderm. Provided the superficial endodermal layer adopts an ectodermal phenotype (‘epidermalised endoderm’) – and histology suggests that it does (Fig. 11) – this observation is in line with the situation described for lungfish enamel organs (Fig. 10H). In the case of the ectodermal teeth, the enamel organ, forming as a folding of the basal ectodermal layer, is itself covered by a superficial layer of endoderm (or rather, ‘epidermalised endoderm’). The function of this layer is supposedly the same as for the endodermal teeth, that is it acts as a layer co‐opted into an ectodermal/peridermal fate. Moreover, in this review it is extensively argued that enamel organs never develop from a monolayer. A co‐option of the superficial endodermal layer into an ectodermal/peridermal identity would explain why endodermal teeth can develop fairly independently from the ectodermal collar, but ectodermal teeth need the endodermal plug reaching up to the body surface. This interpretation could also explain the findings of Ohazama et al. (2010). These authors reported that mouse teeth possess a dual molecular (ectoderm/endoderm) origin according to position, although we now know that the epithelial components of mouse teeth are strictly ectodermal by origin (Rothova et al., 2012).
Fig 11.
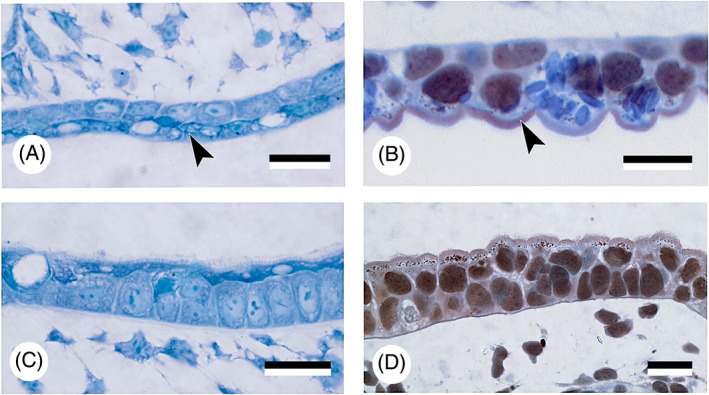
‘Epidermalised' endoderm. Detail of the ‘epidermalised' endodermal epithelium (arrowheads) in the roof of the pharyngeal cavity (A, B) and of the ectodermal epithelium of the skin (C, D) in Acipenser ruthenus (A, C) and Ambystoma mexicanum (B, D). Note similarities in differentiation of the superficial cells. Scale bars = 25 μm.
The ectoderm–endoderm boundary is often represented as a head‐to‐tail contact, as for example in the buccopharyngeal membrane, although intermingling of cells has been described [e.g. chick (Waterman & Schoenwolf, 1980); hamster (Waterman, 1977); reviewed in Soukup et al., 2013]. The presence of a periderm‐like layer covering the endoderm in zebrafish (Rosa et al., 2019; Oralová et al., 2020), or of ‘epidermalised endoderm’ in axolotls (A. Huysseune, personal observations), strongly suggests that this boundary can also take the form of an interface between two superimposed layers. Different authors have stressed the importance of an ectoderm–endoderm boundary in tooth formation. Smith & Coates (2001) and Smith (2003) suggested that the ectoderm–endoderm boundary is crucial for setting up positional information for patterning of the dentition. Recently, Soukup et al. (2021) identified the ectoderm–endoderm boundary in the axolotl as a single tooth‐competent zone giving rise to both dental arcades. In the talpid chicken mutant, the formation of teeth is coupled with alterations in the specification of the oral/aboral boundary of the jaw (Harris et al., 2006). One might speculate that this novel oral/aboral boundary reflects a shift of the ectoderm–endoderm boundary.
The scenario depicted above supports the role of ectoderm, and aligns with the ‘modified outside‐in’ hypothesis for tooth origins. Indeed, for each of the taxa listed, enamel organs develop from a stratified epithelium with distinctive characters between the basal and superficial layer, and with the superficial layer bearing ectodermal/peridermal characters. The apparent need for stratification is conceivable from a functional viewpoint. In gill‐breathing vertebrates the oropharynx is open to the surrounding environment and the superficial layer must maintain a function as a protective covering, with roles as an osmotic, immunological and/or mechanical barrier, whilst the basal layer can differentiate into a particular derivative.
A question that remains is how to explain the sustained need for pouch–cleft contact for pharyngeal tooth formation (as in zebrafish), if it is not required as a channel to allow an influx of ectoderm or periderm. An answer to this question awaits further experimental approaches and dissection of the molecular signalling taking place at these contacts, and especially, examination of both more basal and more derived taxa.
VII. CONCLUSIONS
Teeth in vertebrates can be distributed, depending on the lineage, in the mouth and on the palate (oral teeth), as well as on any of the post‐mandibular arches (pharyngeal teeth). With the evolutionary origin of internal from external odontodes being firmly re‐established, the formation of pharyngeal teeth (and denticles), continues to present a challenge to explain how odontode‐forming epithelium from the skin could have been ‘transferred' into the oropharynx to initiate tooth or denticle formation, in conjunction with neural crest‐derived mesenchyme.
The ‘modified outside‐in’ hypothesis proposed that this transfer was accomplished through displacement of competent odontogenic epithelium, that is ectoderm, not only through the mouth, but via any channel connecting the ectoderm to the epithelial lining of the pharynx (endoderm), such as the gill slits.
Historic studies based on histology, as well as recent research based on modern cell lineage‐tracing techniques, have firmly established that the epithelial component of teeth, the enamel organ – whether oral or pharyngeal – can derive from ectoderm, endoderm, or have a mixed origin. In line with the ‘modified outside‐in’ hypothesis, it is argued here that ectoderm continues to be required in some form to initiate tooth formation, even if teeth derive (partially) from endoderm.
A survey of experimental studies on pharyngeal tooth formation in teleosts and on oral tooth formation in amphibians (the two lineages for which data are available) reveals a seemingly inconclusive picture as to whether only one (either ectoderm or endoderm) or both germ layers are required to initiate tooth formation. However, upon closer inspection it appears that independent from layer origin, the prospective enamel organ is covered by a layer with characteristics reminiscent of ectoderm. In zebrafish this is a layer resembling the periderm (or outer epidermal layer), and is possibly co‐opted from endoderm through evolutionary changes in gene expression. In urodele amphibians, it is the superficial endodermal layer that adopts ectodermal characteristics, and is therefore termed ‘epidermalised endoderm’. Thus, while ectoderm may no longer invade deep into the pharynx, its role may have been taken over through co‐option.
The apparent lack in zebrafish of ectodermal invasion into the pouches prior to tooth initiation, must be considered against the highly derived type of early embryonic development displayed by teleosts, characterised by telolecithal eggs and meroblastic cleavage. There is an urgent need to study basal actinopterygians with holoblastic cleavage to establish whether a bilayered ectoderm is basal for the lineage, whether its superficial layer is homologous to the teleost periderm, whether this layer penetrates into the pharynx, and whether it participates in tooth formation.
Even though ectoderm, at least in zebrafish, does not penetrate deep into the pharynx via gill slits, pouch–cleft contacts (prospective gill slits) are nevertheless required for pharyngeal tooth initiation. A survey of tooth distribution in both actinopterygians and sarcopterygians confirms the close association of pharyngeal teeth with pouches. The nature of the signal at the pouch–cleft contact, whether molecular or mechanical, remains enigmatic.
The pouch that separates the hyoid from first branchial arch (pouch 2) may well possess a special identity. It constitutes an important borderline separating developmental programs from anterior and posterior pharyngeal arches, as it opens first in many species and serves as passageway for initial invasion of periderm in zebrafish.
While pouch–cleft contacts are demonstrably important in tooth initiation, this association cannot on its own explain the distribution of post‐mandibular teeth. Clearly, teeth are found only near pouch–cleft contacts, but neither the sequence of development of the contacts, nor their number, is indicative for how the patterning of the teeth progresses in ontogeny.
Because the likely basal condition in extant gnathostomes is to have teeth spread over all arches (Nelson, 1969, 1970), it is safe to assume that, in evolution, the capacity for tooth formation was lost on tooth‐bearing arches, rather than gained on edentulous arches. One molecule regulating tooth distribution on the different arches is retinoic acid (RA), whose role is discussed, as well as its possible intersection with sonic hedgehog (SHH) signalling. It is proposed that a balance between SHH and RA signalling may be operating to assign tooth‐initiating competence to the endoderm at any particular pouch–cleft contact.
It follows from the above description that teeth develop invariably where ectoderm meets endoderm (at pouch–cleft contacts or at the mouth), that teeth always form from the basal layer of a bi‐ (or multi‐) layered epithelium, and that the surface layer appears to adopt ectodermal characteristics (either as periderm, or as ‘epidermalised endoderm’). Ectoderm–endoderm boundaries may thus not necessarily take the shape of head‐to‐tail contacts, but can also form the interface between two superimposed cell layers, constituting a broad area with odontogenic competence.
Together, three requirements (pouch–cleft contacts, a stratified epithelium and a surface layer that is ectodermal or epidermalised) appear to limit the distribution of teeth within the oropharynx. The proposed role of ectoderm in tooth initiation, irrespective of the enamel organs' germ layer origin, is in line with the ‘modified outside‐in’ hypothesis of tooth origins.
VIII. ACKNOWLEDGMENTS
A.H. and P.E.W. are thankful to J.‐Y. Sire (Université Paris 6, Paris, France) for long‐term collaboration and many insightful discussions. In addition, the authors are very grateful for the donation of his collection of histological sections. V. Soukup and A. Pospisilova (Charles University, Prague, Czech Republic) kindly permitted access to, and use of, axolotl and sturgeon sections, respectively. A. De Clercq (Ghent University, Belgium) and A. Boutet (Station biologique de Roscoff, Sorbonne University, France) generously provided salmon and shark embryos, respectively, for histological preparation. D. Larionova (Ghent University, Belgium) kindly provided the TEM picture used in Fig. 8E. The authors gratefully acknowledge the insightful comments of three anonymous reviewers, which helped to improve the manuscript. A.H. acknowledges a grant from the Ghent University Research Fund (n° BOF24J2015001401) and grants for a sabbatical stay at the Department of Zoology, Charles University, Prague, from the Research Council of the Faculty of Sciences (Ghent University, FCWO) and from the FWO (Research Foundation Flanders ‐ n° FWOSAB2019000601).
[Correction added on 21 October 2021, after first online publication: Authors affiliations have been corrected in this version.]
REFERENCES
- Adams, A. E. (1924). An experimental study of the development of the mouth in the amphibian embryo. Journal of Experimental Zoology 40, 311–379. [Google Scholar]
- Adams, A. E. (1931). Some effects of removal of endoderm from the mouth region of early Amblystoma punctatum embryos. Journal of Experimental Zoology 58, 147–163. [Google Scholar]
- Adams, A. , Mankad, K. , Offiah, C. & Childs, L. (2016). Branchial cleft anomalies: A pictorial review of embryological development and spectrum of imaging findings. Insights Into Imaging 7, 69–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aigler, S. R. , Jandzik, D. , Hatta, K. , Uesugi, K. & Stock, D. W. (2014). Selection and constraint underlie irreversibility of tooth loss in cypriniform fishes. Proceedings of the National Academy of Sciences of the United States of America 111, 7707–7712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aman, A. J. , Fulbright, A. N. & Parichy, D. M. (2018). Wnt/β‐catenin regulates an ancient signaling network during zebrafish scale development. eLife 7, e37001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arambourg, C. & Bertin, L. (1958). Super‐Ordres des Holostéens et des Halécostomes (Holostei et Halecostomi). In Traité de Zoologie (ed. Grassé P.‐P.. Vol. 13 (3). Agnathes et Poissons. Anatomie, Ethologie, Systématique), pp. 2173–2203. Masson et Cie, Paris. [Google Scholar]
- Atkinson, C. J. L. & Collin, S. P. (2012). Structure and topographic distribution of oral denticles in elasmobranch fishes. Biological Bulletin 222, 26–34. [DOI] [PubMed] [Google Scholar]
- Atukorala, A. D. S. , Inohaya, K. , Baba, O. , Tabata, M. J. , Ratnayake, R. A. R. K. , Abduweli, D. , Kasugai, S. , Mitani, H. & Takano, Y. (2011). Scale and tooth phenotypes in medaka with a mutated ectodysplasin‐A receptor: implications for the evolutionary origin of oral and pharyngeal teeth. Archives of Histology and Cytology 73, 139–148. [DOI] [PubMed] [Google Scholar]
- Balfour, F. M. & Parker, W. N. (1882). On the structure and development of Lepidosteus . Philosophical Transactions of the Royal Society of London 173, 359–442. [Google Scholar]
- Balic, A. (2019). Concise review: cellular and molecular mechanisms regulation of tooth initiation. Stem Cells 37, 26–32. [DOI] [PubMed] [Google Scholar]
- Balic, A. & Thesleff, I. (2015). Tissue interactions regulating tooth development and renewal. Current Topics in Developmental Biology 115, 157–186. [DOI] [PubMed] [Google Scholar]
- Balinsky, B. I. (1975). An Introduction to Embryology, 4th Edition. W.B. Saunders Company, Philadelphia, 648 pp. [Google Scholar]
- Barlow, L. A. & Northcutt, R. G. (1995). Embryonic origin of amphibian taste buds. Developmental Biology 169, 273–285. [DOI] [PubMed] [Google Scholar]
- Bartsch, P. , Gemballa, S. & Piotrowski, T. (1997). The embryonic and larval development of Polypterus senegalus Cuvier, 1829: its staging with reference to external and skeletal features, behavior and locomotory habits. Acta Zoologica (Stockholm) 78, 309–328. [Google Scholar]
- Berkovitz, B. K. B. & Shellis, R. P. (2016). The Teeth of Non‐Mammalian Vertebrates. Elsevier, Amsterdam, 354 pp. [Google Scholar]
- Bertin, L. (1958). Organes de la respiration aquatique. In Traité de Zoologie (ed. Grassé P.‐P.. Vol. 13 (2). Agnathes et Poissons. Anatomie, Ethologie, Systématique), pp. 1303–1341. Masson et Cie, Paris. [Google Scholar]
- Birkholz, D. A. , Olesnicky Killian, E. C. , George, K. M. & Artinger, K. B. (2009). prdm1a is necessary for posterior pharyngeal arch development in zebrafish. Developmental Dynamics 238, 2575–2587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bjerring, H. C. (1977). A contribution to structural analysis of the head of craniate animals. Zoologica Scripta 6, 127–183. [Google Scholar]
- Bjerring, H. C. (1998). The fates of spiracular allostoses in mammals. Acta Zoologica (Stockholm) 79, 51–67. [Google Scholar]
- Blais, S. A. , MacKenzie, L. A. & Wilson, M. V. H. (2011). Tooth‐like scales in early Devonian eugnathostomes and the ‘outside‐in’ hypothesis for the origins of teeth in vertebrates. Journal of Vertebrate Paleontology 31, 1189–1199. [Google Scholar]
- Bloomquist, R. F. , Parnell, N. F. , Phillips, K. A. , Fowler, T. E. , Yu, T. Y. , Sharpe, P. T. & Streelman, J. T. (2015). Coevolutionary patterning of teeth and taste buds. Proceedings of the National Academy of Sciences of the United States of America 112, E5954–E5962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boy, J. A. (1988). On some representatives of the Eryopoidea (Amphibia: Temnospondyli) from the European Rotliegend (?uppermost Carboniferous – Permian) 1. Sclerocephalus. Paläontologische Zeitschrift 62, 107–132 [in German]. [Google Scholar]
- Brachet, A. (1935). Traité d'Embryologie des Vertébrés. 690 pp. Masson et Cie , Paris. [Google Scholar]
- Brazeau, M. & Ahlberg, P. (2005). A new look at tetrapod middle ear origins: spiracle evolution in the Tetrapodomorpha. Journal of Vertebrate Paleontology 25, 39A. [Google Scholar]
- Brazeau, M. & Ahlberg, P. (2006). Tetrapod‐like middle ear architecture in a Devonian fish. Nature 439, 318–321. [DOI] [PubMed] [Google Scholar]
- Brazeau, M. D. , Friedman, M. , Jerve, A. & Atwood, R. C. (2017). A three‐dimensional placoderm (stem‐group gnathostome) pharyngeal skeleton and its implications for primitive gnathostome pharyngeal architecture. Journal of Morphology 278, 1220–1228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruneel, B. , Mathä, M. , Paesen, R. , Ameloot, M. , Weninger, W. J. & Huysseune, A. (2015). Imaging the zebrafish dentition: from traditional approaches to emerging technologies. Zebrafish 12, 1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buchtová, M. , Handrigan, G. , Tucker, A. , Lozanoff, S. , Town, L. , Fu, K. , Diewert, V. , Wicking, C. & Richman, J. (2008). Initiation and patterning of the snake dentition are dependent on Sonic Hedgehog signaling. Developmental Biology 319, 132–145. [DOI] [PubMed] [Google Scholar]
- Calamari, Z. T. , Kuang‐Hsien Hu, J. & Klein, O. D. (2018). Tissue mechanical forces and evolutionary developmental changes act through space and time to shape tooth morphology and function. BioEssays 40(1800140), 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cassin, C. & Capuron, A. (1979). Buccal organogenesis in Pleurodeles waltlii Michah (urodele amphibian). Study by intrablastocelic transplantation and in vitro culture. Journal de Biologie Buccale 7, 61–76. [PubMed] [Google Scholar]
- Chai, Y. , Jiang, X. , Ito, Y. , Bringas, P. , Han, J. , Rowitch, D. H. , Soriano, P. , McMahon, A. P. & Sucov, H. M. (2000). Fate of the mammalian cranial neural crest during tooth and mandibular morphogenesis. Development 127, 1671–1679. [DOI] [PubMed] [Google Scholar]
- Chen, C.‐F. , Chu, C.‐Y. , Chen, T.‐H. , Lee, S.‐J. , Shen, C.‐N. & Hsiao, C.‐D. (2011). Establishment of a transgenic zebrafish line for superficial skin ablation and functional validation of apoptosis modulators in vivo. PLoS One 6, e20654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, D. , Blom, H. , Sanchez, S. , Tafforeau, P. , Märss, T. & Ahlberg, P. E. (2020). The developmental relationship between teeth and dermal odontodes in the most primitive bony fish Lophosteus . eLife 9, e60985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, J. , Jacox, L. A. , Saldanha, F. & Sive, H. (2017). Mouth development. Wiley Interdisciplinary Reviews‐Developmental Biology 6, e275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chibon, P. (1966). Analyse expérimentale de la régionalisation et des capacités morphogénétiques de la crête neurale chez l'Amphibien Urodèle Pleurodeles waltlii Michah. Mémoires de la Société Zoologique de France 36, 5–108. [Google Scholar]
- Chibon, P. (1967). Étude expérimentale par ablations, greffes, et autoradiographie de l'origine des dents chez l'Amphibien Urodèle Pleurodeles waltlii Michah. Archives of Oral Biology 12, 745–753. [DOI] [PubMed] [Google Scholar]
- Chibon, P. (1970). L'origine de l'organe adamantin des dents. Etude au moyen du marquage nucléaire de l'ectoderme stomodéal. Annales d'Embryologie et de Morphogenèse 3, 203–213. [Google Scholar]
- Choe, C. P. , Collazo, A. , Trinh, L. A. , Pan, L. , Moens, C. B. & Crump, J. G. (2013). Wnt‐dependent epithelial transitions drive pharyngeal pouch formation. Developmental Cell 24, 296–309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choe, C. P. & Crump, J. G. (2015). Dynamic epithelia of the developing vertebrate face. Current Opinion in Genetics & Development 32, 66–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clack, J. A. (2002). The dermal skull roof of Acanthostega gunnari, an early tetrapod from the Late Devonian. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 93, 17–33. [Google Scholar]
- Clemen, G. , Bartsch, P. & Wacker, K. (1998). Dentition and dentigerous bones in juveniles and adults of Polypterus senegalus (Cladistia, Actinopterygii). Annals of Anatomy 180, 211–221. [DOI] [PubMed] [Google Scholar]
- Coates, M. I. & Clack, J. A. (1991). Fish‐like gills and breathing in the earliest known tetrapod. Nature 352, 234–236. [Google Scholar]
- Collazo, A. , Bolker, J. A. & Keller, R. (1994). A phylogenetic perspective on teleost gastrulation. American Naturalist 144, 133–152. [Google Scholar]
- Comabella, Y. , Franyutti, A. H. , Hurtado, A. , Canabal, J. & García‐Galano, T. (2012). Ontogenetic development of the digestive tract in Cuban gar (Atractosteus tristoechus) larvae. Reviews in Fish Biology and Fisheries 23, 245–260. [Google Scholar]
- Cook, M. H. & Neal, H. V. (1921). Are the taste‐buds of elasmobranchs endodermal in origin? Journal of Comparative Neurology 33, 45–63. [Google Scholar]
- Cordier, A. C. & Haumont, S. M. (1980). Development of thymus, parathyroids, and ultimo‐branchial bodies in NMRI and nude mice. American Journal of Anatomy 157, 227–263. [DOI] [PubMed] [Google Scholar]
- Davit‐Béal, T. , Allizard, F. & Sire, J.‐Y. (2006). Morphological variations in a tooth family through ontogeny in Pleurodeles waltl (Lissamphibia, Caudata). Journal of Morphology 267, 1048–1065. [DOI] [PubMed] [Google Scholar]
- Davit‐Béal, T. , Chisaka, H. , Delgado, S. & Sire, J.‐Y. (2007). Amphibian teeth: current knowledge, unanswered questions, and some directions for future research. Biological Reviews 82, 49–81. [DOI] [PubMed] [Google Scholar]
- de Beer, G. R. (1947). The differentiation of neural crest cells into visceral cartilages and odontoblasts in Amblystoma, and a re‐examination of the germ‐layer theory. Proceedings of the Royal Society of London, Series B 134, 377–398. [PubMed] [Google Scholar]
- Debiais‐Thibaud, M. , Borday‐Birraux, V. , Germon, I. , Bourrat, F. , Metcalfe, C. J. , Casane, D. & Laurenti, P. (2007). Development of oral and pharyngeal teeth in the medaka (Oryzias latipes): comparison of morphology and expression of eve1 gene. Journal of Experimental Zoology (Molecular and Developmental Evolution) 308B, 693–708. [DOI] [PubMed] [Google Scholar]
- Debiais‐Thibaud, M. , Germon, I. , Laurenti, P. , Casane, D. & Borday‐Birraux, V. (2008). Low divergence in Dlx gene expression between dentitions of the medaka (Oryzias latipes) versus high level of expression shuffling in osteichtyans. Evolution & Development 10, 464–476. [DOI] [PubMed] [Google Scholar]
- Debiais‐Thibaud, M. , Oulion, S. , Bourrat, F. , Laurenti, P. , Casane, D. & Borday‐Birraux, V. (2011). The homology of odontodes in gnathostomes: insights from Dlx gene expression in the dogfish, Scyliorhinus canicula . BMC Evolutionary Biology 11, 307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Degener, L. M. (1924). The development of the dentary bone and teeth of Amia calva . Journal of Morphology 39, 113–155. [Google Scholar]
- Delgado, S. , Davit‐Béal, T. , Allizard, F. & Sire, J.‐Y. (2005). Tooth development in a scincid lizard, Chalcides viridanus (Squamata), with particular attention to enamel formation. Cell Tissue Research 319, 71–89. [DOI] [PubMed] [Google Scholar]
- Dettlaff, T. A. (1993). Evolution of the histological and functional structure of ectoderm, chordamesoderm and their derivatives in Anamnia. Roux's Archives in Developmental Biology 203, 3–9. [DOI] [PubMed] [Google Scholar]
- Didier, D. A. , Stahl, B. J. & Zangerl, R. (1994). Development and growth of compound tooth plates in Callorhinchus milii (Chondrichthyes, HoIocephaIi). Journal of Morphology 222, 73–89. [DOI] [PubMed] [Google Scholar]
- Doeland, M. , Couzens, A. M. C. , Donoghue, P. C. J. & Rücklin, M. (2019). Tooth replacement in early sarcopterygians. Royal Society Open Science 6, 191173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donoghue, P. C. J. & Rücklin, M. (2016). The ins and outs of the evolutionary origin of teeth. Evolution & Development 18, 19–30. [DOI] [PubMed] [Google Scholar]
- Donoghue, P. C. J. & Sansom, I. J. (2002). Origin and early evolution of vertebrate skeletonization. Microscopy Research and Technique 59, 352–372. [DOI] [PubMed] [Google Scholar]
- Donoghue, P. C. J. , Sansom, I. J. & Downs, J. P. (2006). Early evolution of vertebrate skeletal tissues and cellular interactions, and the canalization of skeletal development. Journal of Experimental Zoology (Molecular and Developmental Evolution) 306B, 278–294. [DOI] [PubMed] [Google Scholar]
- Duellman, W. E. & Trueb, L. (1986). Biology of the Amphibians. McGraw‐Hill Book Company, New York, 670 pp. [Google Scholar]
- Duncan, K. M. , Mukherjee, K. , Cornell, R. A. & Liao, E. C. (2017). Zebrafish models of orofacial clefts. Developmental Dynamics 246, 897–914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edwards, L. F. (1929). The origin of the pharyngeal teeth of the carp (Cyprinus carpio Linnaeus). Ohio Journal of Sciences 29, 93–130. [Google Scholar]
- Eisenhoffer, G. T. , Slattum, G. , Ruiz, O. E. , Otsuna, H. , Bryan, C. D. , Lopez, J. , Wagner, D. S. , Bonkowsky, J. L. , Chien, C.‐B. , Dorsky, R. I. & Rosenblatt, J. (2017). A toolbox to study epidermal cell types in zebrafish. Journal of Cell Science 130, 269–277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El Shahawy, M. , Reibring, C.‐G. , Neben, C. L. , Hallberg, K. , Marangoni, P. , Harfe, B. D. , Klein, O. D. , Linde, A. & Gritli‐Linde, A. (2017). Cell fate specification in the lingual epithelium is controlled by antagonistic activities of Sonic hedgehog and retinoic acid. PLoS Genetics 13, e1006914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer, B. , Metzger, M. , Richardson, R. , Knyphausen, P. , Ramezani, T. , Franzen, R. , Schmelzer, E. , Bloch, W. , Carney, T. J. & Hammerschmidt, M. (2014). p53 and TAp63 promote keratinocyte proliferation and differentiation in breeding tubercles of the zebrafish. PLoS Genetics 10, e1004048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ford, G. , Balakrishnan, A. , Evans, J. & Bailey, C. (1992). Branchial cleft and pouch anomalies. The Journal of Laryngology and Otology 106, 137–143. [DOI] [PubMed] [Google Scholar]
- Françillon‐Vieillot, H. , Trébaol, L. , Meunier, F. J. & Slembrouck, J. (1994). Histological study of odontogenesis in the pharyngeal jaws of Trachinotus teraia (Cuvier et Valenciennes, 1832) (Osteichthyes, Teleostei, Carangidae). Journal of Morphology 220, 11–24. [DOI] [PubMed] [Google Scholar]
- Fraser, G. J. , Bloomquist, R. F. & Streelman, J. T. (2008). A periodic pattern generator for dental diversity. BMC Biology 6, 32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fraser, G. J. , Cerny, R. , Soukup, V. , Bronner‐Fraser, M. & Streelman, J. T. (2010). The odontode explosion: the origin of tooth‐like structures in vertebrates. BioEssays 32, 808–817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fraser, G. J. , Hulsey, C. D. , Bloomquist, R. F. , Uyesugi, K. , Manley, N. R. & Streelman, J. T. (2009). An ancient gene network is co‐opted for teeth on old and new jaws. PLoS Biology 7, e1000031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fraser, G. J. & Smith, M. M. (2011). Evolution of developmental pattern for vertebrate dentitions: an oro‐pharyngeal specific mechanism. Journal of Experimental Zoology (Molecular and Developmental Evolution) 316, 99–112. [DOI] [PubMed] [Google Scholar]
- Frisdal, A. & Trainor, P. A. (2014). Development and evolution of the pharyngeal apparatus. Wiley Interdisciplinary Reviews‐Developmental Biology 3, 403–418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fukazawa, C. , Santiago, C. , Park, K. M. , Deery, W. J. , de la Torre Canny, S. G. , Holterhoff, C. K. & Wagner, D. S. (2010). Poky/chuk/ikk1 is required for differentiation of the zebrafish embryonic epidermis. Developmental Biology 346, 272–283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardiner, B. C. & Schaeffer, B. (1989). Interrelationships of lower actinopterygian fishes. Zoological Journal of the Linnean Society 97, 135–187. [Google Scholar]
- Gibert, Y. , Bernard, L. , Debiais‐Thibaud, M. , Bourrat, F. , Joly, J.‐S. , Pottin, K. , Meyer, A. , Retaux, S. , Stock, D. W. , Jackman, W. R. , Seritrakul, P. , Begemann, G. & Laudet, V. (2010). Formation of oral and pharyngeal dentition in teleosts depends on differential recruitment of retinoic acid signaling. FASEB Journal 24, 3298–3309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibert, Y. , Samarut, E. , Pasco‐Viel, E. , Bernard, L. , Borday‐Birraux, V. , Sadier, A. , Labbé, C. , Viriot, L. & Laudet, V. (2015). Altered retinoic acid signalling underpins dentition evolution. Proceedings of the Royal Society B 282, 20142764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilchrist, J. D. F. (1922). XXVII.—Note on the oesophageal teeth of the Stromateidæ. Annals and Magazine of Natural History 9(51), 249–255. [Google Scholar]
- Gisbert, E. , Rodriguez, A. , Castelló‐Orvay, F. & Williot, P. (1998). A histological study of the development of the digestive tract of Siberian sturgeon (Acipenser baeri) during early ontogeny. Aquaculture 167, 195–209. [Google Scholar]
- Goette, A. (1901). Uber die Kiemen der Fische. Zeitschrift für Wissenschaftliche Zoologie 69, 533–577. [Google Scholar]
- Gonzalez, M. E. , Blanquez, M. J. & Rojo, C. (1996). Early gill development in the rainbow trout, Oncorhynchus mykiss . Journal of Morphology 229, 201–217. [DOI] [PubMed] [Google Scholar]
- Goodrich, E. S. (1930). Studies on the Structure & Development of Vertebrates. MacMillan and Co, Limited, London, 837 pp. [Google Scholar]
- Graham, A. (2008). Deconstructing the pharyngeal metamere. Journal of Experimental Zoology (Molecular and Developmental Evolution) 310B, 336–344. [DOI] [PubMed] [Google Scholar]
- Graham, A. , Okabe, M. & Quinlan, R. (2005). The role of the endoderm in the development and evolution of the pharyngeal arches. Journal of Anatomy 207, 479–487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham, A. , Poopalasundaram, S. , Shone, V. & Kiecker, C. (2019). A reappraisal and revision of the numbering of the pharyngeal arches. Journal of Anatomy 235, 1019–1023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham, A. & Richardson, J. (2012). Developmental and evolutionary origins of the pharyngeal apparatus. EvoDevo 3, 24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham, A. & Shone, V. (2019). Pharyngeal remodelling in vertebrate evolution. In Evolution and Development of Fishes (eds Johanson Z., Underwood C. and Richter M.), pp. 241–251. Cambridge University Press, New York. [Google Scholar]
- Graveson, A. C. , Smith, M. M. & Hall, B. K. (1997). Neural crest potential for tooth development in a urodele amphibian: developmental and evolutionary significance. Developmental Biology 188, 34–42. [DOI] [PubMed] [Google Scholar]
- Grevellec, A. & Tucker, A. S. (2010). The pharyngeal pouches and clefts: development, evolution, structure and derivatives. Seminars in Cell and Developmental Biology 21, 325–332. [DOI] [PubMed] [Google Scholar]
- Haas, A. (2003). Phylogeny of frogs as inferred from primarily larval characters (Amphibia: Anura). Cladistics 19, 23–89. [DOI] [PubMed] [Google Scholar]
- Hall, B. K. (1998). Germ layers and the germ‐layer theory revisited. Evolutionary Biology 30, 121–186. [Google Scholar]
- Hall, B. K. (2000). The neural crest as a fourth germ layer and vertebrates as quadroblastic not triploblastic. Evolution & Development 2, 3–5. [DOI] [PubMed] [Google Scholar]
- Hall, B. K. & Gillis, J. A. (2013). Incremental evolution of the neural crest, neural crest cells and neural crest‐derived skeletal tissues. Journal of Anatomy 222, 19–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall, C. , Flores, M. V. , Murison, G. , Crosier, K. & Crosier, P. (2006). An essential role for zebrafish Fgfrl1 during gill cartilage development. Mechanisms of Development 123, 925–940. [DOI] [PubMed] [Google Scholar]
- Hanaoka, R. , Ohmori, Y. , Uyemura, K. , Hosoya, T. , Hotta, Y. , Shirao, T. & Okamoto, H. (2004). Zebrafish gcmb is required for pharyngeal cartilage formation. Mechanisms of Development 121, 1235–1247. [DOI] [PubMed] [Google Scholar]
- Hanken, J. & Wake, D. B. (1993). Miniaturization of body size: organismal consequences and evolutionary significance. Annual Review of Ecology and Systematics 24, 501–519. [Google Scholar]
- Hardcastle, Z. , Mo, R. , Hui, C.‐C. & Sharpe, P. T. (1998). The Shh signalling pathway in tooth development: defects in Gli2 and Gli3 mutants. Development 125, 2803–2811. [DOI] [PubMed] [Google Scholar]
- Haridy, Y. , Gee, B. M. , Witzmann, F. , Bevitt, J. J. & Reisz, R. R. (2019). Retention of fish‐like odontode overgrowth in Permian tetrapod dentition supports outside‐in theory of tooth origins. Biology Letters 15, 20190514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris, M. P. , Hasso, S. M. , Ferguson, M. W. J. & Fallon, J. F. (2006). The development of archosaurian first‐generation teeth in a chicken mutant. Current Biology 16, 371–377. [DOI] [PubMed] [Google Scholar]
- Harris, M. P. , Rohner, N. , Schwarz, H. , Perathoner, S. , Konstantinidis, P. & Nüsslein‐Volhard, C. (2008). Zebrafish eda and edar mutants reveal conserved and ancestral roles of ectodysplasin signaling in vertebrates. PLoS Genetics 4, 1–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison, R. G. (1918). Experiments on the development of the forelimb of Amblystoma, a self‐differentiating equipotential system. Journal of Experimental Zoology 25, 413–461. [Google Scholar]
- Hayashi, H. (1972). Opening mechanism of stomodeum cavity and transformation of yolk platelet proteins to various subcellular structures in developing newt embryos, Triturus pyrrhogaster . Journal of Electron Microscopy 21, 246 (abstract). [Google Scholar]
- He, S. , Yue, P. & Chen, Y. (1997). Comparative study on the morphology and development of the pharyngeal dentition in the families of Cypriniformes. Acta Zoologica Sinica 43, 255–262. [Google Scholar]
- Hertwig, O. (1874). Ueber Bau und Entwickelung der Placoidschuppen und der Zähne der Selachier. Jenaische Zeitschrift für Naturwissenschaft 8, 331–404. [Google Scholar]
- Hildebrand, M. (1995). Analysis of Vertebrate Structure, 4th Edition. John Wiley & Sons, New York, 657 pp. [Google Scholar]
- Hilton, E. J. , Grande, L. & Bemis, W. E. (2011). Skeletal anatomy of the shortnose sturgeon, Acipenser brevirostrum Lesueur, 1818, and the systematics of sturgeons (Acipenseriformes, Acipenseridae). Fieldiana Life Earth Sciences 3, 1–168. [Google Scholar]
- Hogan, B. M. , Hunter, M. P. , Oates, A. C. , Crowhurst, M. O. , Hall, N. E. , Heath, J. K. , Prince, V. E. & Lieschke, G. J. (2004). Zebrafish gcm2 is required for gill filament budding from pharyngeal ectoderm. Developmental Biology 276, 508–522. [DOI] [PubMed] [Google Scholar]
- Holtfreter, J. (1935). Morphologische Beeinflussung von Urodelenektoderm bei xenoplastischer Transplantation. Wilhelm Roux’ Archiv für Entwicklungsmechanik 133, 367–426. [DOI] [PubMed] [Google Scholar]
- Holzschuh, J. , Wada, N. , Wada, C. , Schaffer, A. , Javidan, Y. , Talafuss, A. , Bally‐Cuif, L. & Schilling, T. F. (2005). Requirements for endoderm and BMP signalling in sensory neurogenesis in zebrafish. Development 132, 3731–3742. [DOI] [PubMed] [Google Scholar]
- Huysseune, A. (1990). Development of the anterior part of the mandible and the mandibular dentition in two species of Cichlidae (Teleostei). Cybium 14, 327–344. [Google Scholar]
- Huysseune, A. & Sire, J.‐Y. (1998). Evolution of patterns and processes in teeth and tooth‐related tissues in non‐mammalian vertebrates. European Journal of Oral Sciences 106(Suppl. 1), 437–481. [DOI] [PubMed] [Google Scholar]
- Huysseune, A. , Sire, J.‐Y. & Witten, P. E. (2009). Evolutionary and developmental origins of the vertebrate dentition. Journal of Anatomy 214, 465–476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huysseune, A. , Sire, J.‐Y. & Witten, P. E. (2010). A revised hypothesis on the evolutionary origin of the vertebrate dentition. Journal of Applied Ichthyology 26, 152–155. [Google Scholar]
- Huysseune, A. , Soenens, M. & Elderweirdt, F. (2014). Wnt signaling during tooth replacement in zebrafish (Danio rerio): pitfalls and perspectives. Frontiers in Physiology 5, 386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huysseune, A. & Thesleff, I. (2004). Continuous tooth replacement: the possible involvement of epithelial stem cells. BioEssays 26, 665–671. [DOI] [PubMed] [Google Scholar]
- Huysseune, A. , Van der heyden, C. & Sire, J.‐Y. (1998). Early development of the zebrafish (Danio rerio) pharyngeal dentition (Teleostei, Cyprinidae). Anatomy & Embryology 198, 289–305. [DOI] [PubMed] [Google Scholar]
- Imai, H. , Osumi, N. & Eto, K. (1998). Contribution of foregut endoderm to tooth initiation of mandibular incisor in rat embryos. European Journal of Oral Sciences 106(Suppl. 1), 19–23. [DOI] [PubMed] [Google Scholar]
- Isokawa, S. , Kubota, K. , Kosakai, T. , Satomura, I. , Tsubouchi, M. & Sera, A. (1965). Some contributions to study of esophageal sacs and teeth of fishes. Journal of the Nihon University School of Dentistry 7, 103–111. [DOI] [PubMed] [Google Scholar]
- Iwamatsu, T. (2004). Stages of normal development in the medaka Oryzias latipes . Mechanisms of Development 121, 605–618. [DOI] [PubMed] [Google Scholar]
- Jackman, W. R. , Davies, S. H. , Lyons, D. B. , Stauder, C. K. , Denton‐Schneider, B. R. , Jowdry, A. , Aigler, S. R. , Vogel, S. A. & Stock, D. W. (2013). Manipulation of Fgf and Bmp signaling in teleost fishes suggests potential pathways for the evolutionary origin of multicuspid teeth. Evolution & Development 15, 107–118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackman, W. R. , Draper, B. W. & Stock, D. W. (2004). Fgf signaling is required for zebrafish tooth development. Developmental Biology 274, 139–157. [DOI] [PubMed] [Google Scholar]
- Jackman, W. R. & Stock, D. W. (2006). Transgenic analysis of Dlx regulation in fish tooth development reveals evolutionary retention of enhancer function despite organ loss. Proceedings of the National Academy of Sciences of the USA 103, 19390–19395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackman, W. R. , Yoo, J. J. & Stock, D. W. (2010). Hedgehog signaling is required at multiple stages of zebrafish tooth development. BMC Developmental Biology 10, 119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janvier, P. (1996). Early Vertebrates. Clarendon Press, Oxford, 393 pp. [Google Scholar]
- Janvier, P. (2015). Facts and fancies about early fossil chordates and vertebrates. Nature 520, 483–489. [DOI] [PubMed] [Google Scholar]
- Jheon, A. H. , Seidel, K. , Biehs, B. & Klein, O. D. (2012). From molecules to mastication: the development and evolution of teeth. Wiley Interdisciplinary Reviews‐Developmental Biology 2, 165–182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johanson, Z. & Ahlberg, P. E. (1997). A new tristichopterid (Osteolepiformes: Sarcopterygii) from the Mandagery Sandstone (Late Devonian, Famennian) near Canowindra, NSW, Australia. Transactions of the Royal Society of Edinburgh: Earth Sciences 88, 39–68. [Google Scholar]
- Johanson, Z. & Smith, M. M. (2005). Origin and evolution of gnathostome dentitions: A question of teeth and pharyngeal denticles in placoderms. Biological Reviews 80, 303–345. [DOI] [PubMed] [Google Scholar]
- Johnson, C. W. , Hernandez‐Lagunas, L. , Feng, W. , Melvin, V. S. , Williams, T. & Artinger, K. B. (2011). Vgll2a is required for neural crest cell survival during zebrafish craniofacial development. Developmental Biology 357, 269–281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jollie, M. (1968). Some implications of the acceptance of a delamination principle. In: Current Problems of Lower Vertebrate Phylogeny. Proceedings of the fourth Nobel Symposium, Stockholm, 1967 (ed. T. Ørvig). Interscience (Wiley), pp. 89–107. Almqvist and Wiksell, Stockholm, New York.
- Kapsimali, M. (2017). Epithelial cell behaviours during neurosensory organ formation. Development 144, 1926–1936. [DOI] [PubMed] [Google Scholar]
- Kawasaki, K. , Keating, J. N. , Nakatomi, M. , Welten, M. , Mikami, M. , Sasagawa, I. , Puttick, M. N. , Donoghue, P. C. J. & Ishiyama, M. (2020). Coevolution of enamel, ganoin, enameloid, and their matrix SCPP genes in osteichthyans. iScience 24, 102023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kemp, A. (2002a). Growth and hard tissue remodelling in the dentition of the Australian lungfish, Neoceratodus forsteri (Osteichthyes: Dipnoi). Journal of Zoology 257, 219–235. [Google Scholar]
- Kemp, A. (2002b). The marginal dentition of the Australian lungfish, Neoceratodus forsteri (Osteichthyes: Dipnoi). Journal of Zoology 257, 325–331. [Google Scholar]
- Kemp, A. (2003). The ultrastructure of developing tooth plates in the Australian lungfish, Neoceratodus forsteri . Tissue and Cell 35, 401–426. [DOI] [PubMed] [Google Scholar]
- Kerr, J. G. (1903). The development of Lepidosiren paradoxa. Part III. Development of the skin and its derivatives. Journal of Cell Science s2–46, 417–459. [Google Scholar]
- Kimmel, C. B. , Ballard, W. W. , Kimmel, S. R. , Ullmann, B. & Schilling, T. F. (1995). Stages of embryonic development of the zebrafish. Developmental Dynamics 203, 253–310. [DOI] [PubMed] [Google Scholar]
- Kimmel, C. B. , Warga, R. M. & Schilling, T. F. (1990). Origin and organization of the zebrafish fate map. Development 108, 581–594. [DOI] [PubMed] [Google Scholar]
- King, B. , Marone, F. & Rücklin, M. (2021). Tooth development in the Early Devonian sarcopterygian Powichthys and the evolution of the crown osteichthyan dentition. Palaeontology 64, 645–659. [Google Scholar]
- Kobayashi, D. , Jindo, T. , Naruse, K. & Takeda, H. (2006). Development of the endoderm and gut in medaka, Oryzias latipes . Development Growth and Differentiation 48, 283–295. [DOI] [PubMed] [Google Scholar]
- Koch, G. , Thesleff, I. & Kreiborg, S. (2017). Tooth development and disturbances in number and shape of teeth. Electronic file available at https://pocketdentistry.com/ Accessed 19.05.2020.
- Kochilas, L. K. , Potluri, V. , Gitler, A. , Balasubramanian, K. & Chin, A. J. (2003). Cloning and characterization of zebrafish tbx1 . Gene Expression Patterns 3, 645–651. [DOI] [PubMed] [Google Scholar]
- Kollar, E. J. & Baird, G. R. (1969). The influence of the dental papilla on the development of tooth shape in embryonic mouse tooth germs. Journal of Embryology and Experimental Morphology 21, 131–148. [PubMed] [Google Scholar]
- Kollar, E. J. & Mina, M. (1991). Role of the early epithelium in the patterning of the teeth and Meckel's cartilage. Journal of Craniofacial Genetics and Developmental Biology 11, 223–228. [PubMed] [Google Scholar]
- Kopinke, D. , Sasine, J. , Swift, J. , Stephens, W. Z. & Piotrowski, T. (2006). Retinoic acid is required for endodermal pouch morphogenesis and not for pharyngeal endoderm specification. Developmental Dynamics 235, 2695–2709. [DOI] [PubMed] [Google Scholar]
- Kunz, Y. W. , Luer, C. A. & Kapoor, B. G. (eds) (2009). Development of Non‐Teleost Fishes. Science Publishers, Enfield; 289 pp. [Google Scholar]
- Kwon, G. S. , Viotti, M. & Hadjantonakis, A. K. (2008). The endoderm of the mouse embryo arises by dynamic widespread intercalation of embryonic and extraembryonic lineages. Developmental Cell 15, 509–520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laforest, L. , Brown, C. W. , Poleo, G. , Géraudie, J. , Tada, M. , Ekker, M. & Akimenko, M.‐A. (1998). Involvement of the sonic hedgehog, patched 1 and bmp2 genes in patterning of the zebrafish dermal fin rays. Development 125, 4175–4184. [DOI] [PubMed] [Google Scholar]
- Le Pabic, P. , Stellwag, E. J. & Scemama, J.‐L. (2009). Embryonic development and skeletogenesis of the pharyngeal jaw apparatus in the cichlid Nile Tilapia (Oreochromis niloticus). The Anatomical Record 292, 1780–1800. [DOI] [PubMed] [Google Scholar]
- Lee, R. T. H. , Asharani, P. V. & Carney, T. J. (2014). Basal keratinocytes contribute to all strata of the adult zebrafish dermis. PLoS One 9, e84858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee, R. T. H. , Thiery, J. P. & Carney, T. J. (2013). Dermal fin rays and scales derive from mesoderm, not neural crest. Current Biology 23, R336–R337. [DOI] [PubMed] [Google Scholar]
- Lovely, C. B. , Swartz, M. E. , McCarthy, N. , Norrie, J. L. & Eberhart, J. K. (2016). Bmp signaling mediates endoderm pouch morphogenesis by regulating Fgf signaling in zebrafish. Development 143, 2000–2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maisey, J. G. (1988). Phylogeny of early skeletal induction and ossification patterns. Evolutionary Biology 22, 1–36. [Google Scholar]
- Maisey, J. G. (2000). Discovering Fossil Fishes. Westview Press, Boulder, 223 pp. [Google Scholar]
- Maisey, J. G. (2004). Morphology of the braincase in the broadnose sevengill shark Notorynchus (Elasmobranchii, Hexanchiformes), based on CT scanning. American Museum Novitates 3429, 1–52. [Google Scholar]
- Maisey, J. G. , Turner, S. , Naylor, G. J. P. & Miller, R. F. (2014). Dental patterning in the earliest sharks: implications for tooth evolution. Journal of Morphology 275, 586–596. [DOI] [PubMed] [Google Scholar]
- McCollum, M. & Sharpe, P. T. (2001). Evolution and development of teeth. Journal of Anatomy 199, 153–159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Millot, J. & Anthony, J. (1958). Anatomie de Latimeria chalumnae. Tome I. Squelette, muscles et formations de soutien, pp. 1–122. Editions du Centre National de la Recherche Scientifique, Paris. [Google Scholar]
- Minarik, M. , Stundl, J. , Fabian, P. , Jandzik, D. , Metscher, B. D. , Psenicka, M. , Gela, D. , Osorio‐Pérez, A. , Arias‐Rodriguez, L. , Horácek, I. & Cerny, R. (2017). Pre‐oral gut contributes to facial structures in non‐teleost fishes. Nature 547, 209–214. [DOI] [PubMed] [Google Scholar]
- Mitsiadis, T. A. , Caton, J. & Cobourne, M. (2006). Waking‐up the sleeping beauty: recovery of the ancestral bird odontogenic program. Journal of Experimental Zoology (Molecular and Developmental Evolution) 306B, 227–233. [DOI] [PubMed] [Google Scholar]
- Mongera, A. & Nüsslein‐Volhard, C. (2013). Scales of fish arise from mesoderm. Current Biology 23(9), R338–R339. [DOI] [PubMed] [Google Scholar]
- Moroff, T. (1904). Über die Entwicklung der Kiemen bei Fischen. Archiv für Mikroskopische Anatomie und Entwicklungsgeschichte 64, 189–213. [Google Scholar]
- Murdock, D. J. E. , Dong, X.‐P. , Repetski, J. E. , Marone, F. , Stampanoni, M. & Donoghue, P. C. J. (2013). The origin of conodonts and of vertebrate mineralized skeletons. Nature 502, 546–549. [DOI] [PubMed] [Google Scholar]
- Nelson, G. J. (1967). Gill arches of teleostean fishes of the family Clupeidae. Copeia 1967, 389–399. [Google Scholar]
- Nelson, G. J. (1969). Gill arches and the phylogeny of fishes, with notes on the classification of vertebrates. Bulletin of the American Museum of Natural History 141, 475–552. [Google Scholar]
- Nelson, G. J. (1970). Pharyngeal denticles (placoid scales) of sharks, with notes on the dermal skeleton of vertebrates. American Museum Novitates 2415, 1–26. [Google Scholar]
- Nolte, C. , De Kumar, B. & Krumlauf, R. (2019). Hox genes: downstream “effectors” of retinoic acid signaling in vertebrate embryogenesis. genesis 57, e23306. [DOI] [PubMed] [Google Scholar]
- Ober, E. A. , Field, H. A. & Stainier, D. Y. R. (2003). From endoderm formation to liver and pancreas development in zebrafish. Mechanisms of Development 120, 5–18. [DOI] [PubMed] [Google Scholar]
- Ohazama, A. , Haworth, K. E. , Ota, M. S. , Khonsari, R. H. & Sharpe, P. T. (2010). Ectoderm, endoderm, and the evolution of heterodont dentitions. genesis 48, 382–389. [DOI] [PubMed] [Google Scholar]
- Okabe, M. & Graham, A. (2004). The origin of the parathyroid gland. Proceedings of the National Academy of Science of the United States of America 101, 17717–17719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okada, K. , Inohaya, K. , Mise, T. , Kudo, A. , Takada, S. & Wada, H. (2016). Reiterative expression of pax1 directs pharyngeal pouch segmentation in medaka (Oryzias latipes). Development 143, 1800–1810. [DOI] [PubMed] [Google Scholar]
- Okada, K. & Takada, S. (2020). The second pharyngeal pouch is generated by dynamic remodeling of endodermal epithelium in zebrafish. Development 147, dev194738. [DOI] [PubMed] [Google Scholar]
- Oralová, V. , Rosa, J. T. , Larionova, D. , Witten, P. E. & Huysseune, A. (2020). Multiple epithelia are required to form teeth deep in the pharynx. Proceedings of the National Academy of Science of the United States of America 117, 11503–11512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ørvig, T. (1967). Phylogeny of tooth tissues: evolution of some calcified tissues in early vertebrates. In Structural and Chemical Organization of Teeth (Volume 1, ed. Miles A. E. W.), pp. 45–110. Academic Press, London. [Google Scholar]
- Ørvig, T. (1977). A survey of odontodes (‘dermal teeth’) from developmental, structural, functional, and phyletic points of view. In Problems in Vertebrate Evolution (eds Andrews S. M., Miles R. S. and Walker A. D.), pp. 53–75. Academic Press, London. [Google Scholar]
- Panousopoulou, E. & Green, J. B. A. (2016). Invagination of ectodermal placodes is driven by cell intercalation‐mediated contraction of the suprabasal tissue canopy. PLoS Biology 14, e1002405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parker, H. W. & Dunn, E. R. (1964). Dentitional metamorphosis in the Amphibia. Copeia 1964, 75–86. [Google Scholar]
- Pasco‐Viel, E. , Charles, C. , Chevret, P. , Semon, M. , Tafforeau, P. , Viriot, L. & Laudet, V. (2010). Evolutionary trends of the pharyngeal dentition in Cypriniformes (Actinopterygii: Ostariophysi). PLoS One 5, e11293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peters, H. & Balling, R. (1999). Tooth development. Trends in Genetics 15, 59–65. [DOI] [PubMed] [Google Scholar]
- Peters, H. , Neubüser, A. , Kratochwil, K. & Balling, R. (1998). Pax9‐deficient mice lack pharyngeal pouch derivatives and teeth and exhibit craniofacial and limb abnormalities. Genes & Development 12, 2735–2747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peyer, B. (1968). Comparative Odontology. The University of Chicago, Chicago & London, 347 pp. [Google Scholar]
- Peyrard‐Janvid, M. , Leslie, E. J. , Kousa, Y. A. , Smith, T. L. , Dunnwald, M. , Magnusson, M. , Lentz, B. A. , Unneberg, P. , Fransson, I. , Koillinen, H. K. , Rautio, J. , Pegelow, M. , Karsten, A. , Basel‐Vanagaite, L. , Gordon, W. , Andersen, B. , Svensson, T. , Murray, J. C. , Cornell, R. A. , Kere, J. & Schutte, B. C. (2014). Dominant mutations in GRHL3 cause Van der Woude syndrome and disrupt oral periderm development. American Journal of Human Genetics 94, 23–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piotrowski, T. , Ahn, D.‐G. , Schilling, T. F. , Nair, S. , Ruvinsky, I. , Geisler, R. , Rauch, G.‐J. , Haffter, P. , Zon, L. I. , Zhou, Y. , Foott, H. , Dawid, I. B. & Ho, R. K. (2003). The zebrafish van gogh mutation disrupts tbx1, which is involved in the DiGeorge deletion syndrome in humans. Development 130, 5043–5052. [DOI] [PubMed] [Google Scholar]
- Piotrowski, T. & Nüsslein‐Volhard, C. (2000). The endoderm plays an important role in patterning the segmented pharyngeal region in zebrafish (Danio rerio). Developmental Biology 225, 339–356. [DOI] [PubMed] [Google Scholar]
- Pradel, A. , Maisey, J. G. , Tafforeau, P. , Mapes, R. H. & Mallatt, J. (2014). A Palaeozoic shark with osteichthyan‐like branchial arches. Nature 509, 608–611. [DOI] [PubMed] [Google Scholar]
- Prochazka, J. , Prochazkova, M. , Du, W. , Spoutil, F. , Tureckova, J. , Hoch, R. , Shimogori, T. , Sedlacek, R. , Rubenstein, J. L. , Wittmann, T. & Klein, O. D. (2015). Migration of founder epithelial cells drives proper molar tooth positioning and morphogenesis. Developmental Cell 35, 713–724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qu, Q. , Sanchez, S. , Blom, H. , Tafforeau, P. & Ahlberg, P. E. (2013). Scales and tooth whorls of ancient fishes challenge distinction between external and oral ‘teeth’. PLoS One 8, e71890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rankin, S. A. , McCracken, K. W. , Luedeke, D. M. , Han, L. , Wells, J. M. , Shannon, J. M. & Zorn, A. M. (2018). Timing is everything: reiterative Wnt, BMP and RA signaling regulate developmental competence during endoderm organogenesis. Developmental Biology 434, 121–132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rasch, L. J. , Martin, K. J. , Cooper, R. L. , Metscher, B. D. , Underwood, C. J. & Fraser, G. J. (2016). An ancient dental gene set governs development and continuous regeneration of teeth in sharks. Developmental Biology 415, 347–370. [DOI] [PubMed] [Google Scholar]
- Raven, C. P. (1935). Zur Entwicklung der Ganglienleiste: IV. Untersuchugen über Zeitpunkt und Verlauf der «materiellen Determination» des präsumptiven Kopfganglienleistenmaterials der Urodelen. Wilhelm Roux' Archiv für Entwicklungsmechanik der Organismen 132, 509–575. [DOI] [PubMed] [Google Scholar]
- Reif, W. E. (1978a). Shark dentitions. Morphogenetic processes and evolution. Neues Jahrbuch für Geologie und Paläontologie‐Abhandlungen 157, 107–115. [Google Scholar]
- Reif, W. E. (1978b). Types of morphogenesis of the dermal skeleton in fossil sharks. Paläontologische Zeitschrift 52, 110–128. [Google Scholar]
- Reif, W. E. (1982). Evolution of dermal skeleton and dentition in vertebrates. The odontode regulation theory. Evolutionary Biology 15, 287–368. [Google Scholar]
- Reif, W. E. (2006). Conodonts, odontodes, stem‐groups, and the ancestry of enamel genes. Neues Jahrbuch für Geologie und Paläontologie‐Abhandlungen 241, 405–439. [Google Scholar]
- Reisinger, E. (1933). Entwicklungsgeschichtliche Untersuchungen am Amphibienvorderdarm. (Gleichzeitig ein Beitrag zur Keimblattspezifität und zur prospektiven Bedeutung des Mesektoderms). Wilhelm Roux' Archiv für Entwicklungsmechanik der Organismen 129, 445–501. [DOI] [PubMed] [Google Scholar]
- Richardson, J. , Shono, T. , Okabe, M. & Graham, A. (2011). The presence of an embryonic opercular flap in amniotes. Proceedings of the Royal Society B 279, 224–229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson, R. J. , Hammond, N. L. , Coulombe, P. A. , Saloranta, C. , Nousiainen, H. O. , Salonen, R. , Berry, A. , Hanley, N. , Headon, D. , Karikoski, R. & Dixon, M. J. (2014). Periderm prevents pathological epithelial adhesions during embryogenesis. The Journal of Clinical Investigation 124, 3891–3900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richman, J. M. & Handrigan, G. R. (2011). Reptilian tooth development. genesis 49, 247–260. [DOI] [PubMed] [Google Scholar]
- Rizzoti, K. & Lovell‐Badge, R. (2007). SOX3 activity during pharyngeal segmentation is required for craniofacial morphogenesis. Development 134, 3437–3448. [DOI] [PubMed] [Google Scholar]
- Romer, A. S. (1949). The Vertebrate Body. W.B. Saunders Company, Philadelphia & London, 643 pp. [Google Scholar]
- Rosa, J. T. , Oralová, V. , Larionova, D. , Eisenhoffer, G. T. , Witten, P. E. & Huysseune, A. (2019). Periderm invasion contributes to epithelial formation in the teleost pharynx. Scientific Reports 9, 10082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rothova, M. , Thompson, H. , Lickert, H. & Tucker, A. S. (2012). Lineage tracing of the endoderm during oral development. Developmental Dynamics 241, 1183–1191. [DOI] [PubMed] [Google Scholar]
- Rücklin, M. & Donoghue, P. C. J. (2015). Romundina and the evolutionary origin of teeth. Biology Letters 11, 20150326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rücklin, M. & Donoghue, P. C. J. (2019). Evolutionary Origin of Teeth. Chichester, John Wiley & Sons, Ltd. [Google Scholar]
- Rücklin, M. , Donoghue, P. C. J. , Johanson, Z. , Trinajstic, K. , Marone, F. & Stampanoni, M. (2012). Development of teeth and jaws in the earliest jawed vertebrates. Nature 491, 748–751. [DOI] [PubMed] [Google Scholar]
- Rücklin, M. , Giles, S. , Janvier, P. & Donoghue, P. C. J. (2011). Teeth before jaws? Comparative analysis of the structure and development of the external and internal scales in the extinct jawless vertebrate Loganellia scotica . Evolution & Development 13, 523–532. [DOI] [PubMed] [Google Scholar]
- Rücklin, M. , King, B. , Cunningham, J. A. , Johanson, Z. , Marone, F. & Donoghue, P. C. J. (2021). Acanthodian dental development and the origin of gnathostome dentitions. Nature Ecology and Evolution 5, 919–926. [DOI] [PubMed] [Google Scholar]
- Sasagawa, I. , Ishiyama, M. , Yokosuka, H. , Mikami, M. , Oka, S. , Shimokawa, H. & Uchida, T. (2019). Immunolocalization of enamel matrix protein‐like proteins in the tooth enameloid of spotted gar, Lepisosteus oculatus, an actinopterygian bony fish. Connective Tissue Research 60, 291–303. [DOI] [PubMed] [Google Scholar]
- Schoch, R. R. (2001). Can metamorphosis be recognized in Palaeozoic amphibians? Neues Jahrbuch für Geologie und Paläontologie‐Abhandlungen 220, 335–367. [Google Scholar]
- Schoch, R. R. (2002). The evolution of metamorphosis in temnospondyls. Lethaia 35, 309–327. [Google Scholar]
- Schoch, R. R. & Witzmann, F. (2011). Bystrow's paradox – gills, fossils, and the fish‐to‐tetrapod transition. Acta Zoologica (Stockholm) 92, 251–265. [Google Scholar]
- Schwend, T. & Ahlgren, S. C. (2009). Zebrafish con/disp1 reveals multiple spatiotemporal requirements for Hedgehog‐signaling in craniofacial development. BMC Developmental Biology 9, 59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sehring, I. M. & Weidinger, G. (2019). Recent advancements in understanding fin regeneration in zebrafish. Wiley Interdisciplinary Reviews‐Developmental Biology 2019, e367. [DOI] [PubMed] [Google Scholar]
- Sellman, S. (1946). Some experiments on the determination of the larval teeth in Ambystoma mexicanum . Odontologisk Tidskrift 54, 1–128. [Google Scholar]
- Seritrakul, P. , Samarut, E. , Lama, T. T. S. , Gibert, Y. , Laudet, V. & Jackman, W. R. (2012). Retinoic acid expands the evolutionarily reduced dentition of zebrafish. FASEB Journal 26, 5014–5024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shkil, F. N. , Levin, B. A. , Abdissa, B. & Smirnov, S. V. (2010). Variability in the number of tooth rows in the pharyngeal dentition of Barbus intermedius (Teleostei; Cyprinidae): genetic, hormonal and environmental factors. Journal of Applied Ichthyology 26, 315–319. [Google Scholar]
- Shone, V. & Graham, A. (2014). Endodermal/ectodermal interfaces during pharyngeal segmentation in vertebrates. Journal of Anatomy 225, 479–491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sire, J.‐Y. & Allizard, F. (2001). A fourth teleost lineage possessing extra‐oral teeth: the genus Atherion (Teleostei; Atheriniformes). European Journal of Morphology 39, 295–305. [DOI] [PubMed] [Google Scholar]
- Sire, J.‐Y. , Davit‐Béal, T. , Delgado, S. , Van der heyden, C. & Huysseune, A. (2002). First‐generation teeth in nonmammalian lineages: evidence for a conserved ancestral character? Microscopy Research and Technique 59, 408–434. [DOI] [PubMed] [Google Scholar]
- Sire, J.‐Y. & Huysseune, A. (2003). Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach. Biological Reviews 78, 219–249. [DOI] [PubMed] [Google Scholar]
- Smith, M. M. (2003). Vertebrate dentitions at the origin of jaws: when and how pattern evolved. Evolution & Development 5, 394–413. [DOI] [PubMed] [Google Scholar]
- Smith, M. M. & Coates, M. I. (1998). Evolutionary origins of the vertebrate dentition: phylogenetic patterns and developmental evolution. European Journal of Oral Sciences 106(Suppl. 1), 482–500. [DOI] [PubMed] [Google Scholar]
- Smith, M. M. & Coates, M. I. (2000). Evolutionary origins of teeth and jaws: developmental models and phylogenetic patterns. In Development, Function and Evolution of Teeth (eds Teaford M., Smith M. and Ferguson M.), pp. 133–151. Cambridge University Press, Cambridge. [Google Scholar]
- Smith, M. M. & Coates, M. I. (2001). The evolution of vertebrate dentitions: phylogenetic pattern and developmental models. In Major Events in Early Vertebrate Evolution (ed. Ahlberg P. E., Systematics Association Special Volume Series 61), pp. 223–240. CRC Press, London. [Google Scholar]
- Smith, M. M. , Fraser, G. J. , Chaplin, N. , Hobbs, C. & Graham, A. (2009). Reiterative pattern of sonic hedgehog expression in the catshark dentition reveals a phylogenetic template for jawed vertebrates. Proceedings of the Royal Society B‐Biological Sciences 276, 1225–1233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith, M. M. & Johanson, Z. (2015). Origin of the vertebrate dentition: teeth transform jaws into a biting force. In Great Transformations in Vertebrate Evolution (eds Dial K. P., Shubin N. and Brainerd E. L.), pp. 9–29. The University of Chicago Press, Chicago and London. [Google Scholar]
- Smith, M. M. , Johanson, Z. , Butts, T. , Ericsson, R. , Modrell, M. , Tulenko, F. J. , Davis, M. C. & Fraser, G. J. (2015). Making teeth to order: conserved genes reveal an ancient molecular pattern in paddlefish (Actinopterygii). Proceedings of the Royal Society B 282, 20142700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soukup, V. , Epperlein, H. H. , Horacek, I. & Cerny, R. (2008). Dual epithelial origin of vertebrate oral teeth. Nature 455, 795–799. [DOI] [PubMed] [Google Scholar]
- Soukup, V. , Horacek, I. & Cerny, R. (2013). Development and evolution of the vertebrate primary mouth. Journal of Anatomy 222, 79–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soukup, V. , Tazaki, A. , Yamazaki, Y. , Pospisilova, A. , Epperlein, H.‐H. , Tanaka, E. & Cerny, R. (2021). Oral and palatal dentition of axolotl arises from a common tooth‐competent zone along the ecto‐endodermal boundary. Frontiers in Cell and Developmental Biology 8, 622308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stadtmüller, F. (1924). Über Entwicklung und Bau der papillenförmigen Erhebungen (Filterfortsätze) auf den Branchialbogen der Salamandridenlarven. Zeitschrift für Morphologie und Anthropologie 24, 125–156. [Google Scholar]
- Starck, D. (1982). Vergleichende Anatomie der Wirbeltiere auf evolutionsbiologischer Grundlage. Bd 3. Springer Verlag, Berlin, 1110 pp. [Google Scholar]
- Stock, D. W. (2001). The genetic basis of modularity in the development and evolution of the vertebrate dentition. Philosophical Transactions of the Royal Society of London B 356, 1633–1653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stock, D. W. (2007). Zebrafish dentition in comparative context. Journal of Experimental Zoology (Molecular and Developmental Evolution) 308B, 523–549. [DOI] [PubMed] [Google Scholar]
- Stock, D. W. , Jackman, W. R. & Trapani, J. (2006). Developmental genetic mechanisms of evolutionary tooth loss in cypriniform fishes. Development 133, 3127–3137. [DOI] [PubMed] [Google Scholar]
- Ströer, W. F. H. (1933). Experimentelle Untersuchungen über die Mundentwicklung bei den Urodelen. Wilhelm Roux' Archiv für Entwicklungsmechanik der Organismen 130, 131–186. [DOI] [PubMed] [Google Scholar]
- Stundl, J. , Pospisilova, A. , Jandzik, D. , Fabian, P. , Dobiasova, B. , Minarik, M. , Metscher, B. D. , Soukup, V. & Cerny, R. (2019). Bichir external gills arise via heterochronic shift that accelerates hyoid arch development. eLife 8, e435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sucré, E. , Charmantier‐Daures, M. , Grousset, E. , Charmantier, G. & Cucchi‐Mouillot, P. (2009). Early development of the digestive tract (pharynx and gut) in the embryos and pre‐larvae of the European sea bass Dicentrarchus labrax . Journal of Fish Biology 75, 1302–1322. [DOI] [PubMed] [Google Scholar]
- Suzuki, T. , Oohara, I. & Kurokawa, T. (1999). Retinoic acid given at late embryonic stage depresses sonic hedgehog and Hoxd‐4 expression in the pharyngeal area and induces skeletal malformation in flounder (Paralichthys olivaceus) embryos. Development Growth & Differentiation 41, 143–152. [DOI] [PubMed] [Google Scholar]
- Swartz, M. E. , Sheehan‐Rooney, K. , Dixon, M. J. & Eberhart, J. K. (2011). Examination of a palatogenic gene program in zebrafish. Developmental Dynamics 240, 2204–2220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahama, H. , Sasaki, F. & Watanabe, K. (1988). Morphological changes in the oral (buccopharyngeal) membrane in urodelan embryos: development of the mouth opening. Journal of Morhology 195, 59–69. [DOI] [PubMed] [Google Scholar]
- Taverne, L. (1974). L'ostéologie d'Elops Linné, C., 1766 (Pisces, Elopiformes) et son intérêt phylogénétique. Académie Royale de Belgique . Mémoires de la Classe des Sciences 41, 6–96. [Google Scholar]
- Trapani, J. (2001). Position of developing replacement teeth in teleosts. Copeia 2001, 35–51. [Google Scholar]
- True, J. R. & Carroll, S. B. (2002). Gene co‐option in physiological and morphological evolution. Annual Review of Cell and Developmental Biology 18, 53–80. [DOI] [PubMed] [Google Scholar]
- Turner, S. , Burrow, C. J. , Schultze, H. P. , Blieck, A. , Reif, W. E. , Rexroad, C. B. , Bultynck, P. & Nowlan, G. S. (2010). False teeth: conodont‐vertebrate phylogenetic relationships revisited. Geodiversitas 32, 545–594. [Google Scholar]
- Ungar, P. S. (2010). Mammal Teeth: Origin, Evolution, and Diversity. Johns Hopkins University Press, Baltimore, 303 pp. [Google Scholar]
- Vaccari, E. , Deflorian, G. , Bernardi, E. , Pauls, S. , Tiso, N. , Bortolussi, M. & Argenton, F. (2010). prep1.2 and aldh1a2 participate to a positive loop required for branchial arches development in zebrafish. Developmental Biology 343, 94–103. [DOI] [PubMed] [Google Scholar]
- Vandenplas, S. , De Clercq, A. & Huysseune, A. (2014). Tooth replacement without a dental lamina: the search for epithelial stem cells in Polypterus senegalus . Journal of Experimental Zoology (Molecular and Developmental Evolution) 322B, 281–293. [DOI] [PubMed] [Google Scholar]
- Van der Brugghen, W. & Janvier, P. (1993). Denticles in thelodonts. Nature 364, 107.7686630 [Google Scholar]
- Vandewalle, P. , Huysseune, A. , Aerts, P. & Verraes, W. (1994). The pharyngeal apparatus in teleost feeding. In Advances in Comparative and Environmental: Biomechanics of Feeding in Vertebrates Physiology (Volume 18, eds BELS V., CHARDON M. and VANDEWALLE P.), pp. 59–92. Springer Verlag, Berlin. [Google Scholar]
- Veitch, E. , Begbie, J. , Schilling, T. F. , Smith, M. M. & Graham, A. (1999). Pharyngeal arch patterning in the absence of neural crest. Current Biology 9, 1481–1484. [DOI] [PubMed] [Google Scholar]
- Wacker, K. , Bartsch, P. & Clemen, G. (2001). The development of the tooth pattern and dentigerous bones in Polypterus senegalus (Cladistia, Actinopterygii). Annals of Anatomy 183, 37–52. [DOI] [PubMed] [Google Scholar]
- Wagner, G. (1955). Chimaerische Zahnanlagen aus Triton‐Schmelzorgan und Bombinator‐Papille. Mit Beobachtungen über die Entwicklung von Kiemenzähnchen und Mundsinnesknospen in den Triton‐Larven. Journal of Embryology and Experimental Morphology 3, 160–188. [Google Scholar]
- Wainwright, P. C. (2006). Functional morphology of the pharyngeal jaw apparatus. Fish Physiology 23, 77–101. [Google Scholar]
- Waldhausen, J. H. T. (2006). Branchial cleft and arch anomalies in children. Seminars in Pediatric Surgery 15, 64–69. [DOI] [PubMed] [Google Scholar]
- Wall, N. A. & Hogan, B. L. M. (1995). Expression of bone morphogenetic protein‐4 (BMP‐4), bone morphogenetic protein‐7 (BMP‐7), fibroblast growth factor‐8 (FGF‐8) and sonic hedgehog (SHH) during branchial arch development in the chick. Mechanisms of Development 53, 383–392. [DOI] [PubMed] [Google Scholar]
- Wallace, K. N. & Pack, M. (2003). Unique and conserved aspects of gut development in zebrafish. Developmental Biology 255, 12–29. [DOI] [PubMed] [Google Scholar]
- Warga, R. M. & Kane, D. A. (2018). Wilson cell origin for Kupffer's vesicle in the zebrafish. Developmental Dynamics 247, 1057–1069. [DOI] [PubMed] [Google Scholar]
- Warth, P. , Hilton, E. J. , Naumann, B. , Olsson, L. & Konstantinidis, P. (2017). Development of the skull and pectoral girdle in Siberian sturgeon, Acipenser baerii, and Russian sturgeon, Acipenser gueldenstaedtii (Acipenseriformes: Acipenseridae). Journal of Morphology 278, 418–442. [DOI] [PubMed] [Google Scholar]
- Waterman, R. E. (1977). Ultrastructure of oral (buccopharyngeal) membrane formation and rupture in the hamster embryo. Developmental Biology 58, 219–229. [DOI] [PubMed] [Google Scholar]
- Waterman, R. E. & Kao, R. (1982). Formation of the mouth opening in the zebrafish embryo. Scanning Electron Microscopy 1982, 1249–1257. [Google Scholar]
- Waterman, R. E. & Schoenwolf, G. C. (1980). The ultrastructure of oral (buccopharyngeal) membrane formation and rupture in the chick embryo. The Anatomical Record 197, 441–470. [DOI] [PubMed] [Google Scholar]
- Wilde, C. E. (1955). The urodele neuroepithelium. I. The differentiation in vitro of the cranial neural crest. Journal of Experimental Zoology 130, 573–591. [Google Scholar]
- Wilkins, A. S. (2002). The Evolution of Developmental Pathways. Sinauer Associates, Inc, Sunderland, 603 pp. [Google Scholar]
- Wise, S. B. & Stock, D. W. (2006). Conservation and divergence of Bmp2a, Bmp2b, and Bmp4 expression patterns within and between dentitions of teleost fishes. Evolution & Development 8, 511–523. [DOI] [PubMed] [Google Scholar]
- Witten, P. E. , Harris, M. P. , Huysseune, A. & Winkler, C. (2017). Small teleost fish provide new insights into human skeletal diseases. Methods in Cell Biology 138, 321–346. [DOI] [PubMed] [Google Scholar]
- Witten, P. E. , Sire, J.‐Y. & Huysseune, A. (2014). Old, new and new‐old concepts about the evolution of teeth. Journal of Applied Ichthyology 30, 636–642. [Google Scholar]
- Wiweger, M. I. , Zhao, Z. , van Merkesteyn, R. J. P. , Roehl, H. H. & Hogendoorn, P. C. W. (2012). HSPG‐deficient zebrafish uncovers dental aspect of multiple osteochondromas. PLoS One 7, e29734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woltmann, I. , Shkil, F. , De Clercq, A. , Huysseune, A. & Witten, P. E. (2018). Supernumerary teeth in the pharyngeal dentition of slow‐developing zebrafish (Danio rerio, Hamilton, 1822). Journal of Applied Ichthyology 34, 455–464. [Google Scholar]
- Worthington, R. D. & Wake, D. B. (1971). Larval morphology and ontogeny of the Ambystomatid salamander, Rhyacotriton olympicus . American Midland Naturalist 85, 349–365. [Google Scholar]
- Yamamoto, M. , Iuchi, I. & Yamagami, K. (1979). Ultrastructural changes of the teleostean hatching gland cell during natural and electrically induced precocious secretion. Developmental Biology 68, 162–174. [DOI] [PubMed] [Google Scholar]
- Yu, T. & Klein, O. D. (2020). Molecular and cellular mechanisms of tooth development, homeostasis and repair. Development 147, dev184754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan, Y. & Chai, Y. (2019). Regulatory mechanisms of jaw bone and tooth development. Current Topics in Developmental Biology 133, 91–118. [DOI] [PMC free article] [PubMed] [Google Scholar]