

Abstract
Cryptosporidium parvum, which is resistant to chlorine concentrations typically used in water treatment, is recognized as a significant waterborne pathogen. Recent studies have demonstrated that chlorine dioxide is a more efficient disinfectant than free chlorine against Cryptosporidium oocysts. It is not known, however, if oocysts from different suppliers are equally sensitive to chlorine dioxide. This study used both a most-probable-number–cell culture infectivity assay and in vitro excystation to evaluate chlorine dioxide inactivation kinetics in laboratory water at pH 8 and 21°C. The two viability methods produced significantly different results (P < 0.05). Products of disinfectant concentration and contact time (Ct values) of 1,000 mg · min/liter were needed to inactivate approximately 0.5 log10 and 2.0 log10 units (99% inactivation) of C. parvum as measured by in vitro excystation and cell infectivity, respectively, suggesting that excystation is not an adequate viability assay. Purified oocysts originating from three different suppliers were evaluated and showed marked differences with respect to their resistance to inactivation when using chlorine dioxide. Ct values of 75, 550, and 1,000 mg · min/liter were required to achieve approximately 2.0 log10 units of inactivation with oocysts from different sources. Finally, the study compared the relationship between easily measured indicators, including Bacillus subtilis (aerobic) spores and Clostridium sporogenes (anaerobic) spores, and C. parvum oocysts. The bacterial spores were found to be more sensitive to chlorine dioxide than C. parvum oocysts and therefore could not be used as direct indicators of C. parvum inactivation for this disinfectant. In conclusion, it is suggested that future studies address issues such as oocyst purification protocols and the genetic diversity of C. parvum, since these factors might affect oocyst disinfection sensitivity.
Many treatment plants which employ free chlorine as a primary disinfectant are unable to achieve desired microbial inactivation levels without forming disinfection by-products that may exceed regulatory levels. In addition, pathogens of concern to the water industry, such as Giardia lamblia and especially Cryptosporidium parvum, are known to be resistant to chlorine at concentrations typically applied for water treatment (5, 16, 25). Consequently, alternative disinfectants such as chlorine dioxide (ClO2) may be considered. Few studies have been published concerning the inactivation of protozoan parasites in water when chlorine dioxide is used; however, enough data have been collected so far to suggest that chlorine dioxide is a stronger oxidant than free chlorine. Chlorine dioxide does not form halogenated by-products typically associated with chlorine, including trihalomethanes and haloacetic acids; it does, however react to form chlorite and chlorate, which may be toxic at high concentrations (8, 20, 21, 23, 29, 33).
Disparity exists in the chlorine dioxide inactivation data reported for Cryptosporidium (11, 13, 19, 25), and some investigators remain cautious about the possibility of using this disinfectant to inactivate protozoal agents (14). Other researchers have stated that chlorine dioxide is an effective disinfectant for Cryptosporidium, capable of inactivating 2.1 log10 units (Ct value [product of disinfectant concentration and contact time] of 120 mg · min/liter) at 22°C (13). In part, the disparity may be attributed to the different analytical methods which have been used for measuring both parasite viability and chlorine dioxide concentrations, and it suggests that more research is warranted.
Current enumeration techniques to measure C. parvum concentrations in drinking water are cumbersome, expensive, and time-consuming and are not appropriate for routine monitoring (7, 28). Microbial indicators (or surrogates) of these pathogens could potentially be used to evaluate the efficacy of disinfection during water treatment. Aerobic spores of the bacterial genus Bacillus have been proposed as potential candidates for such an evaluation (22, 28). Comparative studies are needed in order to relate the chlorine dioxide inactivation of microbial indicators, such as Bacillus spores, to that of G. lamblia and C. parvum.
The objectives of this study were (i) to test the inactivation by chlorine dioxide of C. parvum oocysts purchased from three different commercial suppliers and (ii) to evaluate Bacillus subtilis spores and Clostridium sporogenes spores as potential microbial indicators of the chlorine dioxide inactivation of C. parvum.
MATERIALS AND METHODS
Organisms.
Purified C. parvum oocysts (Iowa bovine isolate) were purchased from three different suppliers (Table 1). All three isolates originated from the Harley Moon Collection (National Disease Center, Ames, Iowa). The original isolate was obtained from the feces of a naturally infected calf in the 1980s, and the parasite was maintained by infecting newborn calves. Purified oocysts were purchased from (i) the Pleasant Hill Farm, Troy, Idaho; (ii) the Sterling Parasitology Research Laboratory, University of Arizona, Tucson.; and (iii) the University of Alberta, Edmonton, Alberta, Canada (Table 1). Prior to shipment the Pleasant Hill Farm oocysts were purified by ethyl ether extraction followed by centrifugation and a one-step sucrose gradient. Oocysts from the Sterling Parasitology Research Laboratory were purified by using discontinuous sucrose gradients followed by cesium chloride gradients. The University of Alberta oocysts were similarly purified using sucrose flotation and cesium chlorine gradient ultracentrifugation. In all cases, purified oocysts were suspended in an antibiotic solution and shipped on ice. The oocysts used for chlorine dioxide inactivation trials were always used within 2 months of shedding. Upon reception of the cultures, they were maintained at 4°C in the antibiotic solution in which they were shipped. Prior to commencement of an experiment, the oocysts were washed by centrifugation (10,000 × g, 10 min) and resuspended in the experimental water matrix.
TABLE 1.
Characteristics of the purified C. parvum oocysts purchased from three different suppliers
| Supplier | Strain | Supplier's lot no. | Purification method | 50% Infective dose in mice (supplier's data) | Antibiotic solution used for storage | % Infectivity in cell culture (n)a |
|---|---|---|---|---|---|---|
| Pleasant Hill Farm | Iowa | 99-23, 00-3, 00-11 | Ethyl ether extraction, centrifugation, and a one-step sucrose gradient | NRb | PBS–penicillin (20 U/ml)–streptomycin (20 μg/ml) | 1.48 ± 1.18 (7) |
| Sterling Parasitology Laboratory | Iowa | 00619-25 | Discontinuous sucrose gradients followed by cesium chloride gradients | 121 | Deionized water–penicillin (100 U/ml)–gentamicin (100 μg/ml)–0.01% Tween 20 | 2.31 ± 1.94 (2) |
| University of Alberta | Iowa | CP-020900-1 | Sucrose flotation and cesium chloride gradient ultracentrifugation | 103 | Deionized water–penicillin (100 U/ml)–gentamicin (100 μg/ml)–streptomycin (100 μg/ml)–0.1% Tween 20 | 7.49 ± 2.11 (2) |
Results are means and standard deviations.
NR, not reported.
The in vitro production of B. subtilis (ATCC 19659) spores was accomplished by cultivating the bacteria at 37°C in 1/10 strength Columbia broth (Difco Laboratories) supplemented with 0.1 mM MnSO4 · 4H2O for 96 h (S. Sattar [University of Ottawa, Ottawa, Canada], personal communication). This procedure yielded up to 5.0 × 108 spores/ml. The spores were further isolated from vegetative cells by heating at 80°C for 12 min. The spores were then plated out for enumeration using a spread plate method on nutrient agar medium containing 0.015 g of trypan blue per liter and incubated at 37°C for 24 h. The spores were stored in sterile deionized distilled water (type I reagent water). Variability in the enumeration of spores was approximately 10%. Replicate samples were averaged for a particular time point. The in vitro production of Clostridium sporogenes (ATCC 319) spores was done by cultivating the bacteria under anaerobic conditions at 37°C in Duncan-Strong sporulation medium (2) for 7 to 10 days. The spores were enumerated under anaerobic conditions using nutrient agar plates. This procedure typically yielded 106 to 107 spores/ml.
Chlorine dioxide generation.
Chlorine dioxide was generated using a modified version of Standard Method 4500 (1). A 25% (wt/vol) solution of NaClO2 was introduced by pumping it at a feed rate of 2 to 3 ml/min into a gas-generating bottle containing 12 N H2SO4. This bottle was connected to a chlorine scrubber bottle containing a 10% (wt/vol) solution of NaClO2. The scrubber was connected to a chlorine dioxide collection bottle filled with deionized distilled water (maintained on ice). At the end of the series, an additional chlorine dioxide trap bottle with 10% (wt/vol) KI was present to trap any remaining chlorine dioxide. Overall, the stock chlorine dioxide solution purity averaged 99% (range of 97 to 100%), and the solution was essentially free from chlorite, chlorate, and chlorine contamination. The stock chlorine dioxide solution was usually diluted to obtain a concentration of about 1 g/liter in order to facilitate the addition of low chlorine dioxide concentrations to water samples. Diluted chlorine dioxide stock solutions were stored in headspace-free 40-ml amber vials at 4°C and in the dark.
Chlorine dioxide residual measurement.
Chlorine dioxide residual concentrations were measured using the lissamine green spectrophotometric method. This method has been shown to be free from interferences by chlorine, chlorite, and chlorate (17). Chlorine dioxide standards were prepared and tested on a regular basis.
Inactivation experiments.
Microbial inactivation experiments were conducted in sterile 500-ml demand-free polyethylene bottles containing sterile deionized distilled water adjusted to pH 8.0 (alkalinity of 7.0 mg/liter as CaCO3; chlorine dioxide demand of 0.15 mg/liter). Approximately 108 B. subtilis spores, Clostridium sporogenes spores, or C. parvum oocysts were added to each bottle. Chlorine dioxide was then added to all vials (except to control vials) at a desired disinfectant dose. The samples were mixed on a shaker at 150 rpm. Microbial samples (1 to 10 ml) were collected at selected time intervals and placed in sterile microtubes containing sterile sodium thiosulfate to quench any residual disinfectant. Samples (1 ml) for chlorine dioxide residual measurements were collected at each sampling time and immediately analyzed. All experiments were conducted in duplicate at 21 ± 1oC in a temperature-controlled incubator. For each batch of oocysts, duplicate control bottles (oocysts without chlorine dioxide) were prepared, incubated, and processed exactly as the chlorine dioxide-treated bottles were. No reduction in oocyst concentration and infectivity over time (up to 120 min, which was the longest incubation time period) was ever recorded for control bottles (data not shown).
Cell culture.
Oocyst viability was determined by a most-probable-number (MPN)–cell culture infectivity assay modified from that of Slifko et al. (32). The modifications included in our study consisted of using a different cell culture (Madin-Darby canine kidney [MDCK] cells [ATCC CCL-34]) from the one used by Slifko et al., avoiding the bleach pretreatment of the oocysts, and incorporating a direct immunodetection assay. MDCK cells were maintained in 25-cm2 flasks and passaged every 3 to 4 days. The growth medium was RPMI 1640 (Cellgro; Fisher Scientific, Pittsburgh, Pa.) with 25 mM HEPES buffer with 300 mg of l-glutamine per liter and supplemented with 5% (vol/vol) fetal bovine serum (Cellect; ICN Biomedicals, Inc., Aurora, Ohio).
MPN-cell culture infectivity assay.
In the present study, the MPN-cell culture infectivity assay was performed by seeding eight-well chamber slides (Lab-Tek Brand Products; Nalge Nunc International, Naperville, Ill.) with MDCK cells and growing them (in the medium described above) at 37°C for 24 h to a confluence of approximately 80%. After incubation, the wells were washed with phosphate-buffered saline (PBS), and 150 μl of medium was added to each well. Oocyst suspensions were diluted in 10-fold increments, and 50-μl aliquots from each appropriate dilution were inoculated into five replicate wells. Typically, four or five 10-fold dilutions were inoculated (undiluted, 10−1, 10−2, 10−3, and 10−4), with five replicate wells per dilution. The slides were then placed into a CO2-enriched environment (Bio-Bag Environmental Chamber Type C; Becton Dickinson and Company, Cockeysville, Md.) at 37°C for 48 h. After this period, the overlying medium was removed, and each well was washed four times with PBS supplemented with 0.03% (vol/vol) Tween 20 (Fisher Scientific) to remove any unexcysted oocysts. The cells were fixed with 100 μl of methanol for 10 min and rehydrated in PBS for 30 min. After rehydration, horse serum (1% [vol/vol]; ICN Biomedicals) was added to each well as a blocking agent and left for 1 h. The cells were then washed with PBS and directly stained with a fluorescein isothiocyanate-labeled polyclonal antibody (Sporo-Glo; Waterborne, Inc., New Orleans, La.) reactive toward the intracellular reproductive stages of this organism. Staining was done at 20°C for 30 min with gentle rocking. Finally, a coverslip was placed on each slide and it was sealed with clear nail polish.
Detection of infection foci and MPN calculations.
For the MPN calculations, five replicate wells from at least three consecutive dilutions were scored. Each well was examined by epifluorescence microscopy (excitation at 450 to 490 nm) at a magnification of ×200 or ×400 using an Olympus BX-60 microscope. A well was scored as positive when an infection focus, representing secondary infections and numerous stages, was observed (Fig. 1). No infection foci were observed with heat-treated (80°C, 15 min) oocytes (data not shown). Sporo-Glo can cross-react with oocysts, which occasionally remained bound to the cell culture after washing. Care was taken to recognize oocysts by differentiating them from other developmental stages (meronts or microgamonts, etc.) using both epifluorescence and bright-field differential interference contrast microscopy (staining patterns, shape, and size). Oocyst MPN values with the Salama correction were calculated using the MPN Calculator Software version 4.04, which was downloaded from the U.S. Environmental Protection Agency website (http://www.epa.gov/nerlcwww/other.htm). The concentration of organisms at each sampling time or in each treatment (MPNt) was normalized by expressing it as a percentage of the original concentration (MPN0) in control samples. Therefore, inactivation, when measured by the MPN-cell culture assay, was expressed as log10 MPNt/MPN0. Percent infectivity was calculated using the equation percent infectivity = (MPN/ml)/(number of oocysts/ml) × 100. The number of oocysts per milliliter was determined by four replicate hemacytometer counts.
FIG. 1.
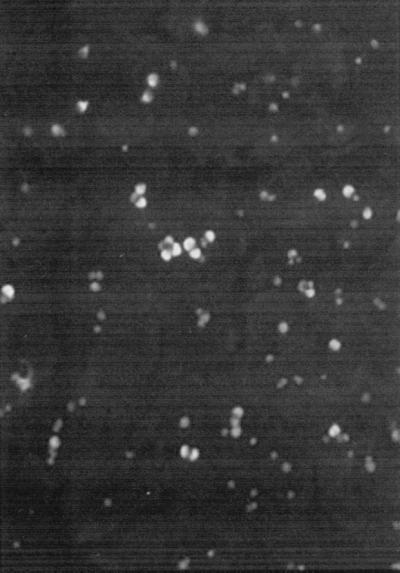
Epifluorescence photograph showing an example of an infection focus with numerous intracellular stages of C. parvum infecting MDCK cells. Magnification, ×400.
In vitro excystation.
The relative viability of the oocysts was also assessed by an in vitro excystation procedure (6). In vitro excystation was done by exposing the organism to an acidified (pH 2.0) Hanks' balanced salt solution (Gibco Laboratories) for 60 min at 37°C, followed by incubation (120 min, 37°C) in PBS containing 0.005 g of trypsin per ml and 0.015 g of taurocholic acid per ml (Sigma Chemical Co., St. Louis, Mo.). After incubation, at least 100 oocysts per slide were observed by differential interference contrast microscopy (Olympus BX-60 microscope), and the numbers of full, partially empty, and empty oocysts were determined. Motile sporozoites were also enumerated. The percent excystation (percentage of viable oocysts) was given by the following equation:
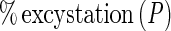 |
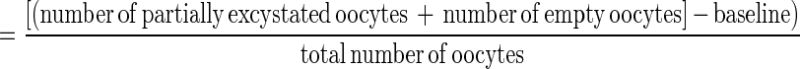 |
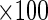 |
A baseline was established by determining the ratio of empty to full oocysts prior to performing in vitro excystation. The concentration of organisms at each sampling time or in each treatment (Pt) was normalized by expressing it as a percentage of the original concentration (P0) in control samples. Therefore, inactivation, when measured by the in vitro excystation assay, was expressed as log10 Pt/P0.
Ct values.
Ct values were calculated by integration of the disinfectant residual concentration (C) up to the given sampling time (t). For each time point, the Ct value was calculated by multiplying the measured chlorine dioxide concentration by the time period since the previous ClO2 measurement. This value was then added to the Ct value calculated at the previous time point to give the overall Ct value for a desired sampling time. Therefore, for a sample n and a sampling time tn with a chlorine dioxide concentration Cn, the (Ct)n is calculated as follows: (Ct)n = Cn · (tn − tn − 1) + (Ct)n − 1.
Physical and chemical parameters.
The pH of the water matrix was recorded at the beginning of each experiment (pH meter model 6071; Jenco Instruments Inc., San Diego, Calif.). Alkalinity was measured using Standard Method 2320B, and UV254 measurement used Standard Method 5910B (1). Total organic carbon analysis was done using an OI Corp. (College Station, Tex.) analytical model 1010 total organic carbon analyzer with autosampler and was based on Standard Method 5310D (1).
Statistical analyses.
Generalized linear models were used to compare inactivation data. These models were constructed using SAS (version 8.0; SAS Institute, Cary, N.C.) with the logarithm of inactivation as the dependent measure and Ct values (milligrams · minute/liter) as independent variables. Model adequacy checks were performed for each of these models and included residual plots, q-q plots, normality tests, and box plots. When significant effects were found to be present, pairwise comparisons were performed using Bonferroni's t test on adjusted mean inactivation levels generated by generalized linear models.
RESULTS
C. parvum inactivation by chlorine dioxide was assessed using (i) an MPN-cell culture infectivity assay with MDCK cells and immunofluorescence detection and (ii) in vitro excystation. The percent infectivities of various batches of C. parvum oocysts were tested over time (up to 75 days) and were found to vary from 0.24 to 7.49% (average of 2.29%; standard deviation of 1.97%; n = 14) (Fig. 2). Oocysts used in chlorine dioxide inactivation experiments, however, were always used within 2 months of shedding.
FIG. 2.

Percent infective C. parvum oocysts purchased from Pleasant Hill Farm (PHF), Sterling Parasitology Laboratory, and the University of Alberta. Infectivity was measured by the MPN-cell infectivity assay.
Bench scale experiments were conducted with C. parvum oocysts (at a concentration of 2.0 × 105 oocysts/ml) using deionized distilled water and demonstrated that cell culture infectivity was more sensitive than in vitro excystation when measuring inactivation (Table 2). For example, an approximate Ct value of 1,000 mg · min/liter yielded 0.5 and 2.0 log10 units of inactivation as measured by in vitro excystation and cell infectivity, respectively (Table 2). Pairwise comparisons performed on the adjusted mean inactivation levels showed that differences between the MPN-cell culture infectivity assay and in vitro excystation results were significant (P < 0.05).
TABLE 2.
Summary of selected data for the inactivation of C. parvum oocysts by chlorine dioxide and comparison to results of the present study
| Expt no. | Experimental summarya | Ct (mg · min/liter) | Log10 inactivationc | Viability or infectivity measure | Referenced |
|---|---|---|---|---|---|
| 1 | C. parvum, room temp, 0.43 mg of ClO2/liter at start, 0.22 mg of ClO2/liter at end, contact time of 30 min | 12.9b | 1.24 (1.0) | Mouse infectivity | 25 |
| 2 | C. parvum, 0.01 M phosphate-buffered pH 7 water, 25°C 1.3 mg of ClO2/liter at start, as low as 0.4 mg of ClO2/liter at end, contact time of 60 min | 78b | 1.0 (2.0) | Mouse infectivity | 18 |
| 3 | C. parvum, oxidant demand-free 0.05 M phosphate-buffered pH 8 water, 22 ± 1°C, 2.0 mg of ClO2/liter at start, contact time of 30 min | 60b | 0.99 | Mouse infectivity | 19 |
| 4 | Same as for expt 3 but with a contact time of 61 min | 122b | 1.57 | Mouse infectivity | 19 |
| 5 | Same as for expt 3 but with 3.3 mg of ClO2/liter at start and a contact time of 116 min | 383b | >3.22 | Mouse infectivity | 19 |
| 6 | C. parvum, 0.05 M phosphate-buffered pH 6 water, 22°C, two trials | 120 | 1.8 and 2.1 | Mouse infectivity | 13 |
| 7 | C. parvum (Iowa isolate, genotype 2 [C], from a calf at the University of Arizona), 0.01 M phosphate-buffered pH 8 ddH2O, 20°C | 150 | 2.0e | Modified in vitro excystation | 31 |
| 8 | C. parvum (Iowa strain from PHF), ddH2O adjusted to pH 8, 21°C | 1,000 | 2.0e | MPN-cell culture | This study |
| 9 | C. parvum (Iowa strain from PHF), ddH2O adjusted to pH 8, 21°C | 1,000 | 0.5e | In vitro excystation | This study |
| 10 | C. parvum (Iowa strain from Sterling), ddH2O adjusted to pH 8, 21°C | 550 | 2.0e | MPN-cell culture | This study |
| 11 | C. parvum (Iowa strain, from University of Alberta), ddH2O adjusted to pH 8, 21°C | 75 | 2.0e | MPN-cell culture | This study |
ddH2O, deionized distilled water, PHF, Pleasant Hill Farm.
The Ct value is the theoretical maximum Ct, calculated by multiplying the initial disinfectant dose by the exposure time. Note that Liyanage et al. (19) provided data for the final and initial chlorine dioxide residuals from their C. parvum oocyst inactivation experiments, and in all cases the final residual was 60% or less of the initial value.
Values in parentheses are recalculations by Finch et al. (11).
Only selected data from each reference are presented.
Approximate value.
Three different lots of C. parvum oocysts from Pleasant Hill Farm (lots 99-23, 00-3, and 00-11) were utilized over the course of this study. Lot 99-23 was used in preliminary experiments with control oocysts. The other two lots were used to perform chlorine dioxide inactivation experiments. Lot 00-3 was shed on 18 January 2000, whereas lot 00-11 was shed on 8 June 2000. In order to assess any possible lot-to-lot variations with respect to chlorine dioxide sensitivity, the inactivations of these two lots of oocysts from Pleasant Hill Farm were compared (Fig. 3). Chlorine dioxide inactivation was measured by the MPN-cell infectivity method and was performed at 21°C in deionized distilled water at pH 8.0. Pairwise comparison done on the adjusted mean inactivation levels showed that there was no significant difference (P < 0.05) in sensitivity to chlorine dioxide observed between the two lots tested, suggesting no significant variability between the two lots and good reproducibility of the method used.
FIG. 3.

Linear regression comparison of the inactivation by chlorine dioxide of two different lots of C. parvum oocysts from Pleasant Hill Farm (PHF). Lot 00-3 (solid line) was shed on 18 January 2000, whereas lot 00-11(dashed line) was shed on 8 June 2000. Inactivation was measured by the MPN-cell infectivity method and was performed at 21°C in deionized distilled water at pH 8.0.
In addition to possible lot-to-lot variations, another source of variation in disinfection studies with C. parvum oocysts may be the origin of the parasite tested. To evaluate these effects, purified oocysts from three different suppliers (Pleasant Hill Farm, Sterling Parasitology Laboratory, and the University of Alberta) were purchased and tested with chlorine dioxide. The results indicated different levels of resistance to chlorine dioxide (Fig. 4). For example, for approximately 2.0 log10 units of inactivation, Ct values of 1,000, 550, and 75 mg · min/liter were required for the Pleasant Hill Farm oocysts, Sterling Parasitology Laboratory oocysts, and University of Alberta oocysts, respectively (Fig. 4 and Table 2). Pairwise comparisons of the adjusted mean inactivation levels showed that these differences were significant for each supplier (P < 0.05). Table 2 summarizes the results from various research groups on C. parvum inactivation by chlorine dioxide. In the present study, inactivation data obtained using the University of Alberta oocysts were comparable to the data previously published by the University of Alberta group (13, 19), suggesting that when the same oocysts are used, inactivation data with mouse infectivity and the MPN-cell infectivity assay are similar (Table 2). So far, no published study on chlorine dioxide inactivation has been performed using Pleasant Hill Farm oocysts, making direct comparisons to other studies difficult because they were carried out with different oocysts.
FIG. 4.

Linear regression comparison of the chlorine dioxide inactivation of C. parvum from three different suppliers (Pleasant Hill Farm, Sterling Parasitology Laboratory, and the University of Alberta). Inactivation was performed in deionized distilled water (21°C, pH 8.0) and measured by the MPN-cell infectivity assay (A) and by in vitro excystation (B). For comparison, chlorine dioxide inactivation (under the same conditions) of B. subtilis spores and Clostridium sporogenes spores is shown in panel A.
The inactivation data were fitted by first-order linear regressions (Table 3). All of the data sets measured by the MPN-cell culture infectivity assay had r2 values of greater than 0.75, and the y-intercept values were minimal, suggesting that the Chick-Watson model may be a good approximation. The r2 values of the in vitro excystation data sets were less than 0.50 for two of the three data sets evaluated, suggesting that in vitro excystation is not an adequate measurement of inactivation and is prone to variations.
TABLE 3.
Linear regressions of C. parvum inactivation results
| Oocyst supplier | In vitro excystation
|
MPN-cell culture assay
|
||||
|---|---|---|---|---|---|---|
| r2 | k (liters/mg · min)a | y intercept | r2 | k (liters/mg · min) | y intercept | |
| Pleasant Hill Farm | 0.475 | −0.0007 | −0.0179 | 0.902 | −0.00233 | 0.338 |
| Sterling Parasitology Laboratory | 0.3697 | −0.0018 | −0.398 | 0.752 | −0.0035 | −0.080 |
| University of Alberta | 0.9622 | −0.0058 | −0.092 | 0.986 | −0.0270 | −0.160 |
k is the pseudo-first-order reaction rate constant, taken as the slope of the linear regression of the data.
A final objective of this study was to evaluate the usefulness of two bacterial spore indicators: B. subtilis and Clostridium sporogenes. Both spore types were cultured in the laboratory and suspended in deionized distilled water for inactivation experiments with chlorine dioxide. Both types of spores showed the same susceptibility to chlorine dioxide, and both were significantly (pairwise comparison on the adjusted mean inactivation levels, P < 0.05) more susceptible to chlorine dioxide inactivation than the C. parvum oocysts from both Pleasant Hill Farm and Sterling Parasitology Laboratory. For example, in deionized distilled water at pH 8.0 and 21°C, a Ct of 200 mg · min/liter provided less than 0.5 log10 unit of C. parvum (Pleasant Hill Farm isolate) inactivation, whereas spore inactivation exceeded 5.0 log10 units for the same conditions (Fig. 4). On the other hand, the spores showed sensitivity to chlorine dioxide inactivation similar to that of the University of Alberta Cryptosporidium (Fig. 4).
DISCUSSION
In the past decade, as C. parvum became recognized as a ubiquitous waterborne pathogen, research has focused on finding alternative disinfectants or disinfection methods that will provide more efficient inactivation of this protozoan parasite during water treatment, since it is known that oocysts of this parasite can survive in chlorinated drinking water (26). One of these alternative disinfectants is chlorine dioxide, which is a stronger oxidant than free chlorine. When using chlorine, Ct values of at least 7,000 mg · min/liter are required to inactivate 2 log10 units of C. parvum (18). As in other studies (13, 18, 31), the results of this work confirm that much lower Ct values are required when using chlorine dioxide to obtain similar levels of inactivation. Among the other advantages attributed to chlorine dioxide is the fact that it does not form many halogenated, and potentially carcinogenic, disinfection by-products typically associated with free chlorine. Chlorine dioxide does, however, react to form chlorite and chlorate (8, 20, 21, 23, 29, 33). The U.S. Environmental Protection Agency maximum contaminant level for chlorite is 1.0 mg/liter.
C. parvum inactivation was measured both by in vitro excystation and by an MPN-cell culture infectivity assay. Oocysts were suspended in water at concentrations of 2.0 × 105 oocysts/ml. These two assays were used in order to compare the present data with published data obtained with infectivity and excystation procedures. The cell culture assay chosen was based on a recently developed method (32), but it incorporated a few significant changes such as using a different cell culture (MDCK cells) and eliminating the oocyst pretreatment in a bleach solution. In our experience, the bleach pretreatment did not improve infectivity rates in MDCK cells. In addition, pretreating with a bleach solution introduces another oxidant to the experimental protocol, which is not desirable in inactivation studies. The use of MPN to quantify infectivity made the assay very sensitive to low levels of infection. In addition, the microscopist does not need to count infection foci but simply needs to recognize them, making the assay much less tedious than one that would incorporate the enumeration of foci. In the present study, MDCK cells were utilized as host cells. C. parvum was shown to produce very efficient infection in MDCK cells (9), and these cells are easily cultured and passaged in the laboratory. Our results indicate that the infectivities of different lots of control oocysts were consistent in MDCK cells: in the present study, the infectivities of control oocysts from three suppliers, as measured by the MPN-cell culture infectivity assay, were similar and ranged from 0.24 to 7.49%. Moreover, when chlorine dioxide inactivation was performed on the University of Alberta oocysts under conditions previously tested by Finch and coworkers (13, 19) (laboratory water, pH 8, 21 to 22°C), our results were comparable to those obtained by the Alberta researchers, who measured inactivation using mouse infectivity. As mentioned above, in vitro excystation was used for comparison to the results in the literature; however, excystation data were significantly different from infectivity data. As reported in other studies (4, 10), our results clearly demonstrate that in vitro excystation consistently underestimates Cryptosporidium inactivation compared to infectivity assays. Oocysts capable of in vitro excystation may not necessarily be infectious, and oocysts incapable of in vitro excystation are almost certainly incapable of causing an infection. In addition, the data obtained by the cell culture were fitted by linear regression. The Chick-Watson model (log10 N/N0 = −k′Cnt) (16) was shown to be a good approximation of these data but not of the excystation data. Overall, all of these factors suggest that the MPN-cell culture infectivity assay utilized in this work adequately measured oocyst viability.
The initial experiments in this study were performed using oocysts purchased from the Pleasant Hill Farm. It rapidly became evident that inactivation data with these oocysts were very different from published data. It was suspected that this difference could be due to the C. parvum isolate used. Comparison of chlorine dioxide inactivation data from different studies is difficult because, without a practical in vitro cultivation system capable of producing oocysts in the concentrations used in bench scale studies, no standard reference strains exist (15). Oocysts are, therefore, produced in animals such as newborn calves and purified from the feces of the infected animals by using various purification protocols. When animals are used for oocyst production, there is always the possibility that contamination from a naturally occurring infection takes place. In addition, oocysts maintained by different facilities may experience evolutionary divergence over time. To investigate this issue, oocysts from two additional suppliers were tested, and all three isolates showed marked differences with respect to resistance to disinfection. Oocysts from all three suppliers originate from the Harley Moon Collection in Ames, Iowa. The chlorine dioxide inactivation measured with the Sterling Parasitology Laboratory oocysts approached the levels observed by Ruffell et al. (31), who also used the Sterling oocysts, but those researchers measured inactivation using a modified in vitro excystation assay. Differences observed in this study may be due to evolutionary divergence of the oocysts, possible contamination from a naturally occurring infection, or differences in isolation and purification of the oocysts from fecal material. The Sterling Parasitology Laboratory oocysts used by Ruffell et al. (31) and in the present study were purified using discontinuous sucrose and cesium chloride centrifugation gradients. The oocysts were prepared using a cesium chloride microcentrifugation procedure consisting of overlaying 1 ml of cesium chloride gradient with 0.5 ml of secondary oocyst suspension. The suspension was centrifuged at 16,000 × g for 3 min. The oocyst-containing layer was then washed twice in a saline solution (22,000 × g for 3 min) (Marilyn Marshall [Sterling Parasitology Laboratory], personal communication). The Alberta oocysts were prepared by sucrose flotation followed by cesium chloride ultracentrifugation. The cesium chloride step involved overlaying three layers of cesium chloride gradient (27 ml) with approximately 3 ml of secondary oocysts. The gradient was then centrifuged (16,000 × g for 60 min). After centrifugation, the oocyst fraction was washed twice with Milli-Q water containing Tween 20 (14,500 × g for 10 min) (12). Cesium chloride centrifugation gradients produce oocysts that appear to be free from organic and fecal debris and, thus, more sensitive to chlorine dioxide disinfection than oocysts purified by a different method (5). For the Pleasant Hill Farm oocysts, ethyl ether extraction was used to remove fat and fecal debris and was followed by centrifugation to remove residual ether and bacteria. Further purification was performed by a one-step sucrose gradient and repeated washings. With the information available, it seems reasonable to suggest that the difference in inactivation kinetics observed could be at least partially explained by different oocyst preparation protocols, which may have selectively concentrated oocysts (for example, a stock consisting of a higher percentage of fully intact and viable oocysts) and/or altered the sensitivities of the oocysts to chlorine dioxide. Different protocols for oocyst purification from feces may therefore have been a factor contributing to the differences in chlorine dioxide inactivation kinetics. These different purification methods and their effect on disinfection sensitivity should be more thoroughly evaluated.
In addition to different sensitivities to chlorine dioxide between isolates from different suppliers, some studies have shown that different lots of the same isolate may respond differently to the same disinfection process. Ruffell et al. (31) used two different lots of oocysts from the same supplier and reported that one lot was more resistant to chlorine dioxide than the other. Slifko et al. (32) described significant lot-to-lot variability in their measurements of infectivity (by the MPN-cell culture infectivity assay) of C. parvum oocysts obtained from the Pleasant Hill Farm, with infectivity ranging from 3.1 to 63.5% in eight lots tested. This variation in C. parvum oocyst infectivity from different lots has been observed by others (3). However, this lot-to-lot variability was not observed in the present study, as with another study which also noted that two different lots of C. parvum oocysts (Iowa strain) used in experiments 6 months apart showed similar sensitivities to (ozone) disinfection (27). Two different lots of C. parvum oocysts (shed approximately 5 months apart) from the Pleasant Hill Farm were used for inactivation experiments in deionized distilled water adjusted to pH 8. There was no statistical difference in the inactivation rate between the two lots when using the MPN-cell culture assay or in vitro excystation. Lot-to-lot variability, when observed, may result from pathological differences upon infection of the host or differences in subsequent oocyst processing (30, 31, 32). In addition, measurements of viability of oocysts isolated from environmental samples may reflect not only the effects of the environment and/or treatment but also the effects of sample collection and processing (15). This rationale may also apply to inactivation studies on C. parvum oocysts collected and purified from animal feces: oocyst sensitivity may be affected by the purification protocol.
Although bench scale inactivation experiments with C. parvum are logistically feasible, they are expensive to conduct because of the cost associated with both producing the oocysts and measuring their infectivity. Pilot scale experiments are also very difficult to perform, again because of high costs (associated with the large number of parasites required) and the potential biohazards related to using large numbers of parasites in a pilot plant. For the latter experiments, it would be desirable to have adequate and nonpathogenic microbial indicators for C. parvum and to use these indicators in inactivation experiments as surrogates for the parasites. Previous studies have suggested that bacterial spores may serve as indicators (22, 24). Bacterial spores of B. subtilis and Clostridium sporogenes were therefore tested in parallel experiments to evaluate their usefulness as indicators of oocyst inactivation by chlorine dioxide. Both spore cultures were similarly sensitive to chlorine dioxide, but their inactivation by chlorine dioxide was similar only to that of the University of Alberta oocysts. Consequently, additional studies are needed to evaluate Bacillus and Clostridium spores (and especially environmental isolates) as possible surrogates or indicators for C. parvum inactivation when using chlorine dioxide. It is possible that environmental isolates may be more resistant than the laboratory cultures tested. However, the results of this study suggest that since there may be a wide range of disinfection resistance among C. parvum isolates, no single spore indicator may be suitable to adequately model C. parvum inactivation by chlorine dioxide.
In conclusion, the MPN-cell culture infectivity method has been shown to be an excellent method for assessing inactivation of C. parvum by chlorine dioxide. Using this method, it was demonstrated that oocysts purchased from different suppliers were significantly different with respect to their resistance to chlorine dioxide, suggesting that future studies must address issues such as oocyst purification protocols and the genetic diversity of C. parvum. Finally, the two spore cultures tested in this study were shown to be inadequate indicators of C. parvum inactivation. Ongoing work will serve to evaluate the chlorine dioxide resistance of environmental spore-forming isolates.
ACKNOWLEDGMENTS
We acknowledge the Chlorine Chemistry Council, Sterling Pulp Chemicals Ltd., the Canadian Chlorine Coordinating Committee, the Canadian Chemical Producers Association, and the Natural Sciences and Engineering Research Council of Canada for their financial support.
The technical help of Nancy Hartman with cell cultures is acknowledged.
REFERENCES
- 1.American Public Health Association. Standard methods for the examination of water and wastewater. 20th ed. Washington, D.C.: American Public Health Association; 1998. [Google Scholar]
- 2.Atlas R. Handbook of microbiological media. Boca Raton, Fla: CRC Press, Inc.; 1997. [Google Scholar]
- 3.Belosevic M, Guy R A, Taghi-Kilani R, Neumann N F, Gyurek L L, Lyanage L R J, Millards P J, Finch G R. Nucleic acid stains as indicators of Cryptosporidium parvum oocyst viability. Int J Parasitol. 1997;27:787–798. doi: 10.1016/s0020-7519(97)00033-7. [DOI] [PubMed] [Google Scholar]
- 4.Bukhari Z, Hargy T M, Bolton J R, Dussert B, Clancy J L. Medium-pressure UV for oocyst inactivation. J Am Water Works Assoc. 1998;91:86–94. [Google Scholar]
- 5.Carpenter C, Fayer R, Trout J, Beach M J. Chlorine disinfection of recreational water for Cryptosporidium parvum. Emerg Infect Dis. 1999;5:579–584. doi: 10.3201/eid0504.990425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chauret C, Nolan K, Chen P, Springthorpe S, Sattar S. Aging of Cryptosporidium parvum oocysts in river water and their susceptibility to disinfection by chlorine and monochloramine. Can J Microbiol. 1998;44:1154–1160. doi: 10.1139/cjm-44-12-1154. [DOI] [PubMed] [Google Scholar]
- 7.Clancy J L, Hollnitz W D, Tabib Z. Commercial labs: how accurate are they? J Am Water Works Assoc. 1994;86:89–94. [Google Scholar]
- 8.Colclough C A, Johnson J D, Christman R F, Millington D S. Water chlorination: environmental impact and health effects. 4, book 1. Ann Arbor, Mich: Ann Arbor Science Publishers; 1983. Organic reaction products of chlorine dioxide and natural aquatic fulvic acids; pp. 219–229. [Google Scholar]
- 9.Fayer R. Cryptosporidium and cryptosporidiosis. Boca Raton, Fla: CRC Press; 1997. [Google Scholar]
- 10.Finch G R, Black E K, Gyurek L G, Belosevic M. Ozone inactivation of Cryptosporidium parvum in demand-free phosphate buffer determined by in vitro excystation and animal infectivity. Appl Environ Microbiol. 1993;59:4203–4210. doi: 10.1128/aem.59.12.4203-4210.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Finch G R, Lyanage L R J, Belosevic M. Symposium Proceedings of Chlorine Dioxide: Drinking Water, Process Water, and Wastewater Issues. Denver, Colo: American Water Works Association; 1995. Effect of chlorine dioxide on Cryptosporidium and Giardia; pp. 23–32. [Google Scholar]
- 12.Finch G R, Gyurek L L, Liyanage L R J, Belosevic M. Effects of various disinfection methods on the inactivation of Cryptosporidium. Denver, Colo.: American Water Works Association; 1997. [Google Scholar]
- 13.Finch G R, Li H. Inactivation of Cryptosporidium at 1°C using ozone or chlorine dioxide. Ozone Sci Eng. 1999;21:477–486. [Google Scholar]
- 14.Fricker C R, Crabb J H. Water-borne cryptosporidiosis: detection methods and treatment options. Adv Parasitol. 1998;40:241–278. doi: 10.1016/s0065-308x(08)60123-2. [DOI] [PubMed] [Google Scholar]
- 15.Gasser R B, O'Donoghue P. Isolation, propagation and characterisation of Cryptosporidium. Int J Parasitol. 1999;29:1379–1413. doi: 10.1016/s0020-7519(99)00113-7. [DOI] [PubMed] [Google Scholar]
- 16.Gyurek L L, Finch G R. Modeling water treatment chemical disinfection kinetics. J Environ Eng. 1998;124:783–793. [Google Scholar]
- 17.Hofmann R, Andrews R C, Ye Q. Comparison of spectrophotometric methods for measuring chlorine dioxide in drinking water treatment. Environ Technol. 1998;19:761–773. [Google Scholar]
- 18.Korich D G, Mead J R, Madore M S, Sinclair N A, Sterling C R. Effects of ozone, chlorine dioxide, chlorine and monochloramine on Cryptosporidium parvum oocyst viability. Appl Environ Microbiol. 1990;56:1423–1428. doi: 10.1128/aem.56.5.1423-1428.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Liyanage L R J, Finch G R, Belosevic M. Sequential disinfection of Cryptosporidium parvum by ozone and chlorine dioxide. Ozone Sci Eng. 1997;19:409–423. [Google Scholar]
- 20.Lykins B W, Koffskey W E, Patterson K S K S. Alternative disinfectants for drinking water treatment. J Environ Eng. 1994;120:745–758. [Google Scholar]
- 21.Muttamara S, Sales C I, Gazali Z. The formation of trihalomethane from chemical disinfectants and humic substances in drinking water. Water Supply. 1995;13:105–117. [Google Scholar]
- 22.Nieminski E C, Bellamy W D, Moss L R. Using surrogates to improve plant performance. J Am Water Works Assoc. 2000;92:67–78. [Google Scholar]
- 23.Oehler K E, Köhler A, Schüttler A. Formation of chlorite during raw water treatment with chlorine dioxide and the removal of chlorite by water treatment. Water Supply. 1986;4:127–139. [Google Scholar]
- 24.Payment P, Franco E. Clostridium perfringens and somatic coliphages as indicators of the efficiency of drinking water treatment for viruses and protozoan cysts. Appl Environ Microbiol. 1993;59:2418–2424. doi: 10.1128/aem.59.8.2418-2424.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Peeters J E, Ares Mazás E, Masschelein W J, Villacorta Martinez de Maturana I, Debacker E. Effect of disinfection of drinking water with ozone or chlorine dioxide on survival of Cryptosporidium parvum oocysts. Appl Environ Microbiol. 1989;55:1519–1522. doi: 10.1128/aem.55.6.1519-1522.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Quinn C M, Betts W B. Longer-term viability status of chlorine-treated Cryptosporidium oocysts in tap water. Biomed Lett. 1993;48:315–318. [Google Scholar]
- 27.Rennecker J L, Mariñas B J, Owens J H, Rice E W. Inactivation of Cryptosporidium parvum oocysts with ozone. Water Res. 1999;33:2481–2488. [Google Scholar]
- 28.Rice E W, Fox K R, Miltner R J, Lytle D A, Johnson C H. Evaluating plant performance with endospores. J Am Water Works Assoc. 1996;88:122–136. [Google Scholar]
- 29.Rice R G, Gomez-Taylor M. Occurrence of by-products of strong oxidants reacting with drinking water contaminants—scope of the problem. Environ Health Persp. 1986;69:31–44. doi: 10.1289/ehp.866931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Robertson L J, Campbell A T, Smith H V. Survival of Cryptosporidium parvum oocysts under various environmental pressures. Appl Environ Microbiol. 1992;58:3494–3500. doi: 10.1128/aem.58.11.3494-3500.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ruffell K M, Rennecker J L, Mariñas B J. Inactivation of Cryptosporidium parvum oocysts with chlorine dioxide. Water Res. 2000;34:868–876. doi: 10.1016/s0043-1354(01)00245-7. [DOI] [PubMed] [Google Scholar]
- 32.Slifko T R, Huffman D E, Rose J B. A most-probable-number assay for enumeration of infectious Cryptosporidium parvum oocysts. Appl Environ Microbiol. 1999;65:3936–3941. doi: 10.1128/aem.65.9.3936-3941.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Werdehoff K S, Singer P C. Chlorine dioxide effects on THMFP, TOXFP, and the formation of inorganic by-products. J Am Water Works Assoc. 1987;79:107–113. [Google Scholar]