

Abstract
Populations experiencing varying levels of ionizing radiation provide an excellent opportunity to study the fundamental drivers of evolution. Radiation can cause mutations and thus supply genetic variation; it can also selectively remove individuals that are unable to cope with the physiological stresses associated with radiation exposure, or non‐selectively cull swathes of the population, reducing genetic variation. Since the nuclear power plant explosion in 1986, the Chernobyl area has experienced a spatially heterogeneous exposure to varying levels of ionizing radiation. We sampled Daphnia pulex (a freshwater crustacean) from lakes across the Chernobyl area, genotyped them at ten microsatellite loci and also calculated the current radiation dose rates. We then investigated whether the pattern of genetic diversity was positively associated with radiation dose rates, consistent with radiation‐mediated supply of de novo mutations, or negatively associated with radiation dose rates, as would be expected with strong radiation‐mediated selection. We found that measures of genetic diversity, including expected heterozygosity and mean allelic richness (an unbiased indicator of diversity), were significantly higher in lakes that experienced the highest radiation dose rates. This suggests that mutation outweighs selection as the key evolutionary force in populations exposed to high radiation dose rates. We also found significant but weak population structure, indicative of low genetic drift and clear evidence for isolation‐by‐distance between populations. This further suggests that gene flow between nearby populations is eroding population structure and that mutational input in high radiation lakes could, ultimately, supply genetic variation to lower radiation sites.
Keywords: evolution, ionizing radiation, microsatellites, population structure
We examined population genetic diversity across Chernobyl Daphnia populations. Measures of genetic diversity, including expected heterozygosity and mean allelic richness, were higher in lakes that experienced the highest radiation dose rates. This suggests that mutation outweighs selection as the key evolutionary force in populations exposed to high radiation dose rates.

1. INTRODUCTION
The explosion of the Chernobyl nuclear power plant in 1986 released approximately 1.85 × 1018 Bq of radionuclides into the atmosphere (IAEA, 2006). This radioactive material was deposited over the surrounding landscape in a very heterogeneous manner, leading to radiation dose rates that vary considerably over very small spatial scales (a few hundred metres: Goodman et al., 2019; Shestopalov, 1996). Wildlife populations in the Chernobyl area have thus experienced varying levels of radiation exposure depending on their locations (Hinton et al., 2007). This is potentially important, because high doses of ionizing radiation are known to have strong negative effects on organismal fitness (Barnthouse, 1995; IAEA, 1992; Real et al., 2004), and can cause genetic mutations that can be passed on to future generations in mammals (mice) and crustacean (Daphnia) (Adewoye et al., 2015; Parisot et al., 2015). Indeed, a recent meta‐analysis found that increased dose rates of ionizing radiation are associated with elevated mutation rates in Chernobyl bacterial, vertebrate, invertebrate and plant species (Møller & Mousseau, 2015). However, very little is known of how chronic exposure to lower doses of radiation over multiple generations affects the structure and genetic diversity across multiple populations of an individual species (c.f. Baker et al., 2017).
Nuclear accidents such as Chernobyl provide a window through which to view the net outcome of three fundamental processes in evolutionary biology: mutation, selection and genetic drift. Ionizing radiation generates mutations (Adewoye et al., 2015; Parisot et al., 2015) and can thus increase the supply of genetic variation to populations (Haldane, 1937; Kimura & Maruyama, 1966). This is important, because genetic diversity is the currency for both evolution and adaptation (Lande & Shannon, 1996). The dose rates across the Chernobyl area have considerably declined since 1986 because of radioactive decay. Nevertheless, if the current dose rates are sufficient cause mutations, there will be a greater supply of genetic variation at high radiation sites relative to their low radiation counterparts. Long‐term exposure to chronic radiation may also have exerted selection on populations. This could lead to the removal of individuals with inadequate mechanisms for protecting against radiation‐mediated cellular damage (Diehn et al., 2009; Khodarev et al., 2004; Ramana et al., 1998; Smirnov et al., 2012), thus reducing genetic variation more in populations experiencing high radiation dose rates (Schlotterer et al., 1997). Finally, the initial fallout from the accident could have caused bottlenecks across the whole area, indiscriminately depleting diversity and causing non‐selective differentiation, that is drift, among populations (Frankham et al., 2002); this would leave its mark in the form of strong population structure (Hartl & Clark, 1997), provided there was low gene flow (Gilpin, 1991; Slatkin, 1987). Indeed, these various evolutionary processes could shape populations in such a way as to reduce fitness differences among them, thus masking the negative effects of ionizing radiation (as we have previously seen in Daphnia: Goodman et al., 2019).
Here, we used 10 microsatellite loci to examine the population genetic structure of the freshwater crustacean, Daphnia pulex, in seven Daphnia‐inhabited lakes in the Chernobyl area. Five of the lakes were within the Chernobyl Exclusion Zone (CEZ), and the other two were situated outside the CEZ. Daphnia are perfectly poised to study the effects of radiation because they are known to suffer reduced fitness (Marshall, 1962) and accumulate mutations (Parisot et al., 2015) when exposed to elevated radiation dose rates. Moreover, their populations are defined by the boundary of the water body that they inhabit (i.e. they do not move great distances across landscapes), and the heterogeneous nature of radionuclide deposition across the CEZ provides variation in radiation exposures that are independent of distance between populations (Goodman et al., 2019; Shestopalov, 1996). Migration between populations is therefore limited, though still possible, and gene flow is generally restricted. As we could not explicitly test if radiation dose rate caused shifts in Daphnia population genetic diversity, we were able to evaluate whether measures of population genetic diversity across lakes experiencing differing radiation dose rates were consistent with: (1) radiation‐mediated supply of genetic variation, (2) radiation‐mediated selection or (3) genetic drift associated with a massive population bottleneck.
2. MATERIALS AND METHODS
2.1. Sample collection
Live Daphnia pulex, sediment and water samples were collected from seven of the eight Daphnia‐inhabited lakes within and immediately outside the CEZ from the 7–16 July 2016. Individual daphnids were immediately stored in 1.5‐ml Eppendorf tubes in absolute ethanol at ambient temperatures and transported back to the UK, where they were stored at −20°C prior genotyping. A total of 205 samples were collected from seven lake populations (27–38 samples per lake; see Table 2). Radiation dose rate was calculated according to the protocol detailed in our previous study (Goodman et al., 2019), where the water and sediment activity concentrations of the different radionuclides were measured for each lake and used to estimate the dose rates to Daphnia. In brief, concentrations for the dominant radionuclides in the CEZ (137Cs and 90Sr) and radionuclides that were considered representative of others present within the water column and top sediment layer (241Am and 239Pu) (IAEA, 2006) were used to estimate dose rates. Where information was available, data on radionuclide concentrations were extracted from the Ukraine atlas (Intelligence Systems GEO, 2008). Where information was unavailable, water and sediment samples were collected at each sampling site and analysed at the Ukrainian Hydrometeorological Institute (see Goodman et al., 2019).
TABLE 2.
Analysis of molecular variance (AMOVA) assessing the partitioning of genetic variation
| Source of variation | df | Sum of squares | Variance | % total | p |
|---|---|---|---|---|---|
| Between populations | 6 | 275.76 | 0.70 | 12.52 | 0.001 |
| Within populations | 197 | 1053.84 | 0.47 | 8.37 | 0.001 |
| Within samples | 204 | 900.78 | 4.42 | 79.11 | 0.001 |
| Total | 407 | 2230.38 | 5.58 | 100 |
Significant values, resulting from a randomization test of 999 samples, are highlighted in bold.
To estimate dose rates experienced by Daphnia, we used the ERICA (version 1.2) software program. ERICA calculates dose rates using an extensive database of published values for radionuclide transfer through the environment and the activity concentrations in various media (lake water, sediment, soil) and organisms, including over 24 000 data entries for freshwater organisms alone (Beresford et al., 2007; Brown et al., 2008, 2016; ICRP, 2009). The user can specify the reference taxon, media and measured radionuclide concentrations, and the tool calculates the average radiation dose rate based on the appropriate subset of data from the database. In this case, Zooplankton was selected as the reference taxon, and the contribution of sediment and water radioactive dose rates was set to 75% and 25% respectively to reflect the period of time Daphnia spend in the water column and as dormant eggs in the sediment (see Alekseev & Lampert, 2001). These percentages are conservative as the majority of radionuclides will accumulate in the surface sediment (Nazarov & Gudkov, 2009). It should be noted that radiation is not confounded with lake location; that is there is no positive relationship between pairwise distances between populations and pairwise differences in radiation dose rate (Mantel r = −0.24, p = 0.16).
2.2. DNA extraction and microsatellite genotyping
Microsatellite genotyping was used to identify differences in allele frequencies and population structure within and across lake populations following the protocol previously outlined by Auld and Brand (2017). First, genomic DNA was extracted from 205 whole Daphnia samples from the seven lake populations (see Table 2 for details) using protocols provided in NucleoSpin Tissue XS (Macherey‐Nagel). We successfully amplified eleven microsatellite markers for each Daphnia across two multiplexes (Table S1; Jansen et al., 2011), though one marker exceeded a 5% null allele rate and was thus excluded from further analysis. Multiplex PCRs consisted of 5 µl 2× Type‐it Multiplex PCR Mastermix (Qiagen), 3 µl Nuclease Free H2O, 1 µl primer mix solution and 1 µl DNA to give a total volume of 10 µl per reaction. The PCR programme was as follows: 15 min at 95°C for Taq activation, followed by 30 cycles of 30 s at 94°C for denaturation of the DNA into separate strands, 90 s at 57°C for annealing of the DNA strands to template DNA and 90 s at 72°C for extension. The final extension was performed for 30 min at 60°C. The final PCR products were analysed with an ABI 3730XL DNA Analyzer (at the Protein Phosphorylation and Ubiquitylation Unit, University of Dundee, UK) using the GeneScan‐500 LIZ size standard (Applied Biosystems). Microsatellite band scoring was completed manually using GENEIOUS software (Biomatters, version 9.1.8). The strongest peak(s) within the loci were selected to determine allele size.
2.3. Analysis
The total number of alleles, mean allelic richness (MAR), the total number of private alleles (PA), and both observed and expected heterozygosities (HO and HE, respectively) were calculated (PopGenReport package; Adamack & Gruber, 2014, adegenet package; Jombart, 2008; Jombart & Ahmed, 2011). Linear models were then used to assess the relationship between log10 of the radiation dose rate and each of MAR, HE and HO. The index of unbiased association (; Brown et al., 1980) was then calculated in order to evaluate the level of linkage disequilibrium within populations; this was done using a permutation approach that estimates the levels of recombination in order to detect association between alleles (poppr package; Kamvar et al., 2014).
The next step was to test which populations were significantly different from each other. First, an analysis of molecular variance (AMOVA) was used to partition variation within and between populations; the significance of these within‐ and among‐population variation was then estimated using 999 permutations (ade4 package; Dray & Dufour, 2007; Bougeard & Dray, 2018). We calculated fixation indices (F‐statistics; Weir & Cockerham, 1984; Wright, 1951) to quantify the extent of population structure within (F IS) and among (F ST) populations (adegenet package; Jombart, 2008; Jombart & Ahmed, 2011). Confidence intervals for the F IS values for each population were computed by bootstrapping over loci, with 999 permutations using the hierfstat package (Goudet & Jombart, 2018). A 999 permutations were used as it eliminates sufficient variation associated with resampling (Hesterberg et al., 2003).
Next, we tested whether populations in close proximity to each other were more similar than those separated by larger geographic distances (i.e. whether there was isolation‐by‐distance). This was done using a Mantel test (implemented using the ade4 package), which quantified the association between two matrices of pairwise Edward's genetic distances (Edwards, 1971) and pairwise Euclidean geographic distances between populations.
Finally, we conducted a Discriminant Analysis of Principal Components (DAPC) to allow a more comprehensive examination of population genetic structure (implemented using the adegenet package; Jombart, 2008; Jombart & Ahmed, 2011). Specifically, we specified K = 7 clusters based on our a priori knowledge of lake population identity and then used the α‐score procedure to identify the optimal number of principal components (PCs) to retain (in this case, 15). The α‐score procedure maximizes the capacity to discriminate between individuals whilst minimizing model overfitting (Jombart & Ahmed, 2011). The DAPC analysis allowed us to visualize both the clustering of individuals in multivariate genetic space and also evaluate admixture among populations. Following our initial specification of the DAPC model, we tested its performance by dividing the data into two groups: one used for model training (49 individuals) and one used for model testing (156 individuals). We specified an equivalent model with the same number of specified clusters and PCs using the training data, then used the model to assign test individuals to their appropriate populations. By comparing the plot of training and test data, we could evaluate the performance of the model.
3. RESULTS
3.1. Greater genetic diversity in highest radiation populations
We identified 204 multilocus genotypes (MLGs) among the 205 individuals (Table 2). HO ranged from 0.31 to 0.66 and expected HE ranged from 0.44 to 0.62 (Table 2). Significant linkage disequilibrium was found in Vediltsy, Yampol, Buryakovka and Krasnyansky lake populations (see values in Table 1).
TABLE 1.
Estimates of genetic diversity among seven Daphnia pulex populations at 10 microsatellite loci across Chernobyl
| Lake | Estimated lake area | Sampling date | Coord N | Coord E | Upper dose estimate | n | MLG | HE | HO | A | PA | MAR | F IS |
|
|
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Vediltsy | 0.175 | 07.06.2016 | 51.4352 | 30.8385 | 0.10 | 27 | 27 | 0.46 | 0.53 | 30 | 1 | 2.66 | −0.14 | 0.03 | |
| Smolin | 0.850 | 07.06.2016 | 51.2757 | 31.0333 | 0.12 | 28 | 28 | 0.5 | 0.45 | 36 | 2 | 3.16 | 0.1 | 0.02 | |
| Yampol | 0.020 | 11.06.2016 | 51.2095 | 30.1767 | 0.20 | 28 | 28 | 0.45 | 0.39 | 30 | 2 | 2.67 | 0.14 | 0.06 | |
| Glinka | 0.005 | 16.06.2016 | 51.2174 | 29.9371 | 1.17 | 28 | 28 | 0.44 | 0.31 | 29 | 1 | 2.62 | 0.3 | 0.06 | |
| Buryakovka | 0.350 | 11.06.2016 | 51.3978 | 29.8931 | 1.77 | 28 | 28 | 0.47 | 0.33 | 32 | 0 | 2.85 | 0.3 | 0.05 | |
| Krasnyansky | 0.066 | 13.06.2016 | 51.4429 | 30.0764 | 55.79 | 38 | 38 | 0.62 | 0.5 | 42 | 3 | 3.7 | 0.2 | 0.06 | |
| Gluboke | 0.260 | 13.06.2016 | 51.4454 | 30.0653 | 181.15 | 28 | 27 | 0.6 | 0.66 | 43 | 4 | 3.68 | −0.11 | 0.02 |
Estimate lake area is in km2 and upper dose estimate is in µGy h−1.
A, number of alleles; HE, expected heterozygosity; HO, observed heterozygosity; MAR, mean allelic richness; MLG, multilocus genotypes; n, number of individuals; PA, number of private alleles; , Index of unbiased association (linkeage disequilibrium).
There was a significant effect of radiation dose rate on both MAR (F 1,5 = 10.24, p = 0.024; Figure 1a) and HE (F 1,5 = 12.03, p = 0.018.); these patterns were clearly driven by the two populations experiencing the highest radiation dose rates (Gluboke and Krasnyansky). We tested the robustness of the relationship between dose rate and MAR by calculating the correlation coefficient (r = 0.82) and then performing a randomization test where we compared this correlation coefficient with 5000 resampled correlations where each of the MAR values was assigned a random dose rate value; we found the relationship between dose rate ad MAR to be robust (p = 0.024; Figure S1a). We repeated the randomization test for the relationship between radiation dose rate and HE, and found this correlation (r = 0.84) to also be robust (p = 0.037; Figure S1b). There was no effect of radiation dose rate on HO (F 1,5 = 1.67, p = 0.25; Table 1). There was no relationship between estimated lake area and either MAR (F 1,5 = 0.18, p = 0.69) or HE (F 1,5 = 0.008, p = 0.93).
FIGURE 1.
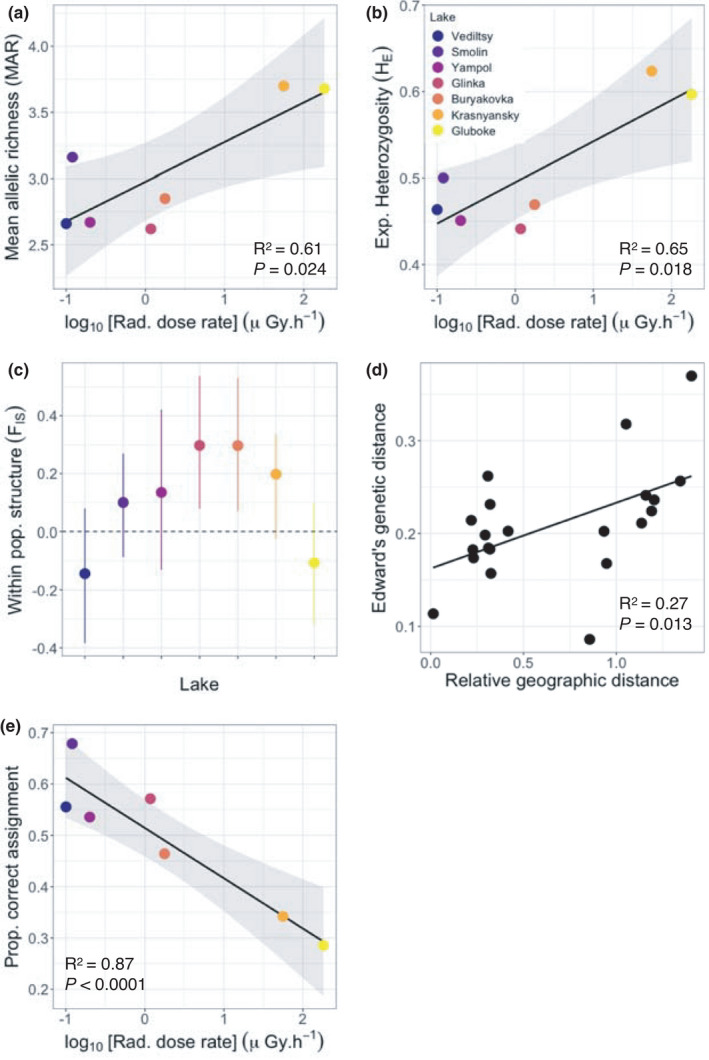
The relationships between (a) mean allelic richness (b) expected heterozygosity and log10 dose rate. Points for panels A and B show the raw data, and the shaded area shows 95% confidence intervals from linear models. The R 2 and p‐values are shown for each model fit. (c) Within population structure F IS values for each population; 95% confidence intervals were computed using Monte Carlo simulations with 999 permutations. (d) Isolation‐by‐distance plot based upon a Monte Carlo simulation using 999 permutations to test between two matrices of pairwise Edward's genetic distances and Euclidean geographic distances. (e) Likelihood of correct assignment of individuals to lake population based on DAPC analysis
3.2. Population structure and gene flow
An AMOVA revealed significant variation within samples, within populations and between populations (Table 2), confirming that there was significant population structure. The overall structure within populations (overall F IS) was 0.11, and the individual population F IS values ranged from −0.14 to 0.30 (Table 1). The lower 95% confidence intervals failed to encompass zero, as is indicative of significant within‐population structure, in two of the seven populations: Glinka and Buryakovka (Figure 1c). The structure across populations was low‐moderate: F ST was 0.14, in broad agreement with the 12.52% of molecular variance explained by population (Table 2). However, all pairwise F ST comparisons were significant (p = 0.001 in all cases), demonstrating that even though there was evidence of gene flow (see later results), each population was indeed a separate entity.
We also found a significant relationship between genetic and geographical distances, that is an isolation‐by‐distance effect (Mantel r = 0.52, p = 0.013; Figure 1d; see Table S2 for pairwise distances between lakes). When we clustered individuals using DAPC, it was clear that whereas individuals generally clustered according to lake population, there was nevertheless overlap between lake populations in terms of multivariate space, particularly between Krasnyansky and Gluboke populations (Figure 2a). Examination of population structure revealed substantial admixture (Figure 2b), with 63% of individuals being assigned to the correct lake population. Finally, the likelihood of individuals being assigned to the correct population declined with increasing population radiation dose rate (GLM: χ2 = 11.94, p < 0.001; Figure 1e).
FIGURE 2.
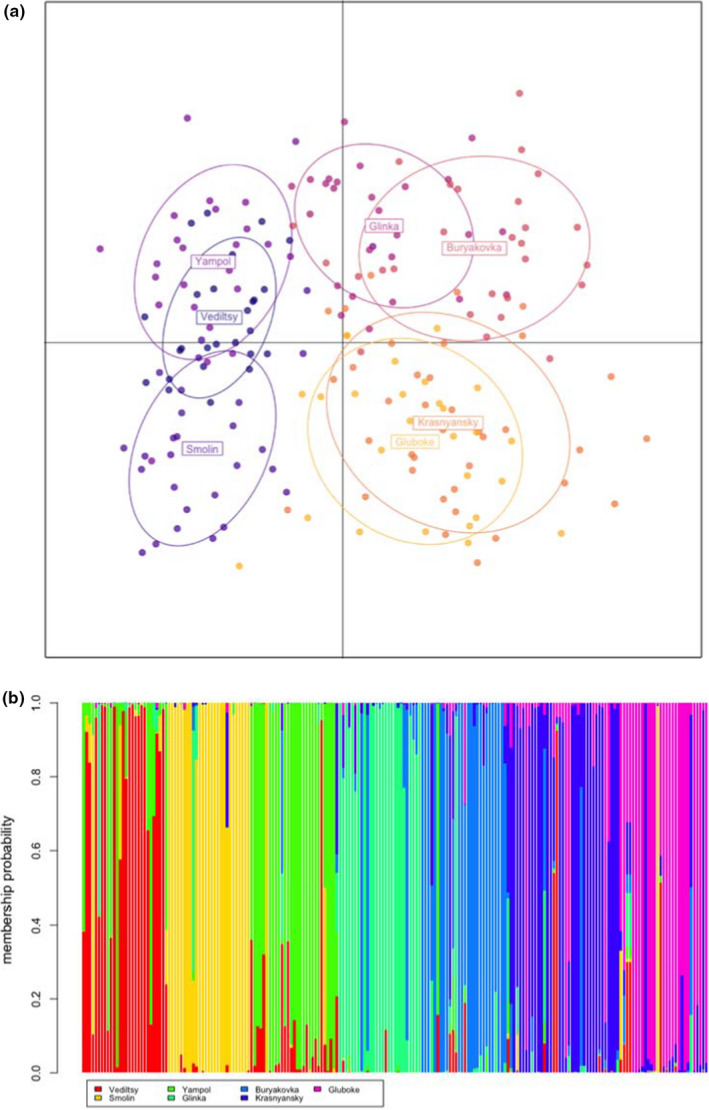
DAPC of the multilocus microsatellite genotypes of 205 Daphnia magna from Chernobyl. (a) Plot of individuals in first two discriminant functions, where colours denote population ID. (b) Structure plot demonstrating population assignment
4. DISCUSSION
Ionizing radiation can cause mutations and thus supply genetic variation to populations (Adewoye et al., 2015; Parisot et al., 2017). This genetic variation is of fundamental importance, because it is the substrate for evolution and potentially adaptation within populations (Haldane, 1937; Kimura & Maruyama, 1966; Muller, 1927). On the contrary, radiation also has the potential to select against genotypes that are unable to cope with high radiation environments (Courtenay, 1965; Ellegren et al., 1997; Khodarev et al., 2004; Møller, 1993, 2002) and thus deplete variation. Previous work found that there was no evidence for phenotypic fitness differences among Daphnia populations that vary in contemporary radiation exposure across the CEZ (Goodman et al., 2019). However, this earlier finding does not mean radiation has no effect on populations. Other experimental studies have uncovered negative effects of ionizing radiation on fitness traits in both Daphnia pulex (Marshall, 1962) and Daphnia magna (Parisot et al., 2015). These works led us to hypothesize that evolutionary processes such as strong radiation‐mediated selection at the time of the accident (when dose rates were very high across the area) favoured the evolution of radiation‐tolerant Daphnia across populations, thus eliminating phenotypic differences between populations; the supply of de novo variation through mutation could subsequently be driving molecular evolution that is not (yet) visible at the level of the phenotype.
We found that lakes experiencing the highest radiation dose rates (Gluboke and Krasnyansky) contained Daphnia populations with the greatest population genetic diversity, consistent with radiation‐mediated supply of genetic variation (sensu Geras’kin & Volkova, 2014). This manifested as a positive relationship between dose rate and the average number of alleles per population (mean allelic richness; MAR; Figure 1a) and also expected heterozygosity (HE, Figure 1b). Moreover, our DAPC analysis uncovered that the lake populations that experience the highest radiation dose rates (Gluboke, Krasnyansky in particular) occupied the greatest volume of multivariate space, further supporting the idea of radiation‐mediated genetic variation (Figure 2a); these lakes also had the lowest likelihood of individuals being correctly assigned to their population based on genotype data alone (Figures 1e and 2b), with missassignment being most common between Krasnyansky and Gluboke populations. Other studies have revealed significantly higher mutation rates in microsatellite loci from samples within the CEZ experiencing contamination from the Chernobyl accident compared with local control sites (Baker et al., 2017; Dubrova et al., 1996; Ellegren et al., 1997; Kuchma et al., 2011). But ours is the first, to our knowledge, to show differences in population genetic diversity associated with continuous variation in dose rate across the Chernobyl area.
Microsatellites are neutral markers (Li et al., 2002), whereas selection acts directly on functional genes. As such, one must be careful not to over‐interpret patterns of selection using microsatellites. Nevertheless, since most mutations are deleterious, it is a reasonable assumption that radiation‐mediated selection, and thus genetic load, could correlate with the radiation dose experienced by populations. Any long‐term directional selection associated with chronic exposure would deplete genetic variation (Mort & Wolf, 1986), as genotypes with poor anti‐stress mechanisms are removed from high but not low dose populations (Diehn et al., 2009; Khodarev et al., 2004; Ramana et al., 1998). The positive relationship between radiation dose rate and genetic diversity (Figure 1a,b) demonstrates that any mutational supply likely outweighs genetic depletion due to radiation‐mediated selection in these Chernobyl Daphnia populations. This does not, of course, mean that radiation‐mediated selection is absent or weak. The positive relationships between population genetic diversity and radiation dose rate are driven by the two highest dose rate populations, and we cannot exclude a negative relationship between radiation dose rate and population genetic diversity at lower radiation levels (a limitation from having seven available populations). Further, it could be that radiation‐mediated selection for Daphnia with effective anti‐stress or DNA repair mechanisms was particularly strong at the time of the accident (Boubriak et al., 2008; Danchenko et al., 2009), acted on populations across the whole area, and that current radiation dose rates are below a threshold required to exert further selection. It is also possible that radiation‐mediated selection from lower dose chronic exposure is weaker and thus slower to act.
We next tested whether the Chernobyl accident may have indiscriminately culled genetic diversity within populations, driving genetic drift (Frankham et al., 2004), reducing the effective population size (Nei & Tajima, 1981). This is crucially important, as when effective population sizes are low, both beneficial and deleterious mutations behave as if they are neutral (Hartl & Clark, 1997), and there is little capacity for selection to drive adaptive evolutionary change (Hartl & Clark, 1997; Lande, 1993). As well as low diversity, genetic drift leads to increased differentiation among populations and strong population structure. We found little evidence for either. As discussed earlier, genetic diversity is surprisingly high (Figure 1a,b), and whereas populations are generally genetically distinct from each other (Table 2; Figure 2a), inter‐population differentiation is weak: only 12.5% of the overall genetic variation is due to between‐population differentiation (Table 2), overall F ST is low, and there is overlap in the genetic characteristics of each lake (Figure 2). Together, these results provide evidence that genetic drift is not a strong force among Chernobyl Daphnia populations.
We further uncovered evidence of significant within‐population structure (i.e. inbreeding) in two populations (of intermediate radiation dose rate, Figure 1c), and no evidence for heterozygote excess. This is in stark contrast to the related Daphnia magna, where heterozygote excess is the norm and systematic inbreeding is either rare or completely absent (Haag et al., 2006; Hebert, 1974a, 1974b; Hebert & Ward, 1976; Walser & Haag, 2012). One possible reason for our findings is that the sex ratios varied among lake populations. The production of males in Daphnia populations is known to be determined by environmental change (such as increased population density, light levels or high levels of toxins) (Eads et al., 2008; Hobaek & Larsson, 1990), and biased sex ratios are known to cause inbreeding, though this is more of a risk in small populations (Mills & Smouse, 1994). Alternatively, and perhaps more likely, there could have been multiple episodes of hatching from the Daphnia resting egg bank within these three lakes and within seasons; this could have led to matings and thus sexual reproduction occurring within subsets of genotypes within lakes, thus facilitating within‐population structure (termed the Wahlund effect; see also Thielsch et al., 2009). In any case, radiation is unlikely to be driving either of these possible scenarios, as within‐population structure was not linked to dose rates.
It is important to consider the complex reproductive biology of Daphnia when evaluating population genetic structure. Genetic recombination is followed by a period of asexual reproduction, and asexual reproduction is often accompanied by clonal selection, where selection on any one trait involves selection on the whole genome (Lynch, 1987). Clonal selection means the same MLG is represented in multiple individuals (Halkett, 2005), F IS values become negative within years as heterozygotes become overrepresented in the population and linkage disequilibrium can accumulate (this is in contrast to selection acting across bouts of sexual reproduction, which could generate positive F IS values). We found only one instance, in Gluboke lake (which had the highest radiation dose rate), where the same MLG was collected twice and, as discussed earlier, there were no significantly negative F IS values for that population. This could be because all the sampled lakes are sufficiently large to host very large Daphnia populations where the frequency of sex is high (Allen & Lynch, 2012). However, we did uncover signatures of linkage disequilibrium () and thus past clonal selection in Yampol, Buryakovka and Krasnyansky lakes. Importantly, the strength of this linkage disequilibrium was not associated with dose rate, suggesting that past bouts of clonal selection or sexual reproduction are caused by biotic or abiotic conditions that are unrelated to the nuclear accident.
Finally, we found evidence that gene flow from dispersal from neighbouring populations is reducing levels of population structure (in the form of strong isolation‐by‐distance: Figure 1d). High radiation environments could thus potentially supply genetic variation to other nearby populations as Daphnia resting stages disperse, fuelling within‐population evolution and adaptation in the manner of a metapopulations (Hanski, 1998). As such, it is plausible that the lack of phenotypic variation among contemporary Chernobyl Daphnia populations (Goodman et al., 2019) is likely concealing highly dynamic demographic and evolutionary processes that are, at least in part, fuelled by ionizing radiation.
It is important to note that this is a correlational study and not a common garden manipulation experiment, so we cannot completely exclude the idea that that an unmeasured variable could have shaped genetic diversity among these Chernobyl Daphnia populations. However, the relationships between radiation dose rate and measures of population genetic diversity are robust (Figure 1 and FigureS1), in spite of the low number of available populations, highly variable levels of linkeage disequilibrium and within‐population structure across lakes. This firmly points towards the explanation that Daphnia populations in high radiation lakes are experiencing greater mutation‐mediated supply of genetic variation than their low radiation lake counterparts. Of course, these findings provide the first insight into radiation‐mediated evolution in Chernobyl; the logical next step is to search for those mutations in coding regions and look for evidence of selection.
CONFLICT OF INTEREST
The authors declare no conflict of interest.
AUTHOR CONTRIBUTIONS
JG and SKJRA designed the research. JG conducted the Daphnia sampling. GL determined the radioisotope activities. JG calculated the radiation dose rates. JG and JB did the microsatellite genotyping. SKJRA contributed reagents and analytical tools. JG and SKJRA conducted the genetic analyses and wrote the manuscript. All authors approved the final version of the manuscript.
PEER REVIEW
The peer review history for this article is available at https://publons.com/publon/10.1111/jeb.13983.
Supporting information
Supplementary Material
ACKNOWLEDGEMENTS
We thank David Copplestone, Sergey Gashchak, Andrew Beckerman and André Gilburn for their contributions to discussions over the manuscript. We are also grateful for fieldwork assistance provided by Eugene Guliachenko. J.C. was supported by a NERC grant (NE/L000369/1), and S.K.J.R.A was supported by a NERC Fellowship (NE/L011549/1). This work was completed as part of the TREE (Transfer‐Exposure‐Effects) consortium under the RATE program (Radioactivity and the Environment), funded by the Environment Agency and Radioactive Waste Management Ltd.
Goodman, J. , Brand, J. , Laptev, G. , & Auld, S. K. J. R. (2022). Radiation‐mediated supply of genetic variation outweighs the effects of selection and drift in Chernobyl Daphnia populations. Journal of Evolutionary Biology, 35, 413–422. 10.1111/jeb.13983
DATA AVAILABILITY STATEMENT
Microsatellite marker data are published on Dryad: https://doi.org/10.5061/dryad.0cfxpnw48.
REFERENCES
- Adamack, A. T. , & Gruber, B. (2014). PopGenReport: Simplifying basic population genetic analyses in R. Methods in Ecology and Evolution, 5(4), 384–387. [Google Scholar]
- Adewoye, A. B. , Lindsay, S. J. , Dubrova, Y. E. , & Hurles, M. E. (2015). The genome‐wide effects of ionizing radiation on mutation induction in the mammalian germline. Nature Communications, 6, 6684. 10.1038/ncomms7684 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alekseev, V. , & Lampert, W. (2001). Maternal control of resting‐egg production in Daphnia. Nature, 414(6866), 899–901. 10.1038/414899a [DOI] [PubMed] [Google Scholar]
- Allen, D. E. , & Lynch, M. (2012). The effect of variable frequency of sexual reproduction on the genetic structure of natural populations of a cyclical parthenogen. Evolution, 66(3), 919–926. 10.1111/j.1558-5646.2011.01488.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Auld, S. K. J. R. , & Brand, J. (2017). Simulated climate change, epidemic size, and host evolution across host–parasite populations. Global Change Biology, 23(12), 5045–5053. 10.1111/gcb.13769 [DOI] [PubMed] [Google Scholar]
- Baker, R. J. , Dickins, B. , Wickliffe, J. K. , Khan, F. A. , Gaschak, S. , Makova, K. D. , & Phillips, C. D. (2017). Elevated mitochondrial genome variation after 50 generations of radiation exposure in a wild rodent. Evolutionary Applications, 10(8), 784–791. 10.1111/eva.12475 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnthouse, L. W. (1995). Effects of ionizing radiation on terrestrial plants and animals: A workshop report. [Google Scholar]
- Beresford, N. , Brown, J. , Copplestone, D. , Garnier‐Laplace, J. , Howard, B. J. , Larsson, C. M. , Oughton, D. , Pröhl, G. , & Zinger, I. (2007). D‐ERICA: An integrated approach to the assessment and management of environmental risks from ionising radiation. Description of purpose, methodology and application. EC project contract no. FI6R‐CT‐2004‐508847. www.erica‐project.org [Google Scholar]
- Boubriak, I. I. , Grodzinsky, D. M. , Polischuk, V. P. , Naumenko, V. D. , Gushcha, N. P. , Micheev, A. N. , McCready, S. J. , & Osborne, D. J. (2008). Adaptation and impairment of DNA repair function in pollen of betula verrucosa and seeds of oenothera biennis from differently radionuclide‐contaminated sites of chernobyl. Annals of Botany, 101(2), 267–276. 10.1093/aob/mcm276 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bougeard, S. , & Dray, S. (2018). Supervised multiblock analysis in R with the ade4 package. Journal of Statistical Software, 86(1), 1–17. 10.18637/jss.v086.i01 [DOI] [Google Scholar]
- Brown, A. H. , Feldman, M. W. , & Nevo, E. (1980). Multilocus structure of natural populations of Hordeum Spontaneum . Genetics, 96(2), 523–536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown, J. E. , Alfonso, B. , Avila, R. , Beresford, N. A. , Copplestone, D. , & Hosseini, A. (2016). A new version of the ERICA tool to facilitate impact assessments of radioactivity on wild plants and animals. Journal of Environmental Radioactivity, 153, 141–148. [DOI] [PubMed] [Google Scholar]
- Brown, J. E. , Alfonso, B. , Avila, R. , Beresford, N. A. , Copplestone, D. , Pröhl, G. , & Ulanovsky, A. (2008). The ERICA tool. Journal of Environmental Radioactivity, 99(9), 1371–1383. 10.1016/j.jenvrad.2008.01.008 [DOI] [PubMed] [Google Scholar]
- Courtenay, V. D. (1965). The response to continuous irradiation of the mouse lymphoma L 5178Y grown in vitro . International Journal of Radiation Biology and Related Studies in Physics, Chemistry and Medicine, 9, 581–592. [DOI] [PubMed] [Google Scholar]
- Danchenko, M. , Skultety, L. , Rashydov, N. M. , Berezhna, V. V. , Mátel, L. , Salaj, T. , Pret'ová, A. , & Hajduch, M. (2009). Proteomic analysis of mature soybean seeds from the Chernobyl area suggests plant adaptation to the contaminated environment. Journal of Proteome Research, 8(6), 2915–2922. 10.1021/pr900034u [DOI] [PubMed] [Google Scholar]
- Diehn, M. , Cho, R. W. , Lobo, N. A. , Kalisky, T. , Dorie, M. J. , Kulp, A. N. , Qian, D. , Lam, J. S. , Ailles, L. E. , Wong, M. , Joshua, B. , Kaplan, M. J. , Wapnir, I. , Dirbas, F. , Somlo, G. , Garberoglio, C. , Paz, B. , Shen, J. , Lau, S. K. , … Clarke, M. F. (2009). Association of reactive oxygen species levels and radioresistance in cancer stem cells. Nature, 458(7239), 780–783. 10.1038/nature07733 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dray, S. , & Dufour, A. B. (2007). The ade4 package: implementing the duality diagram for ecologists. Journal of Statistical Software, 22(4), 1–20. 10.18637/jss.v022.i04 [DOI] [Google Scholar]
- Dubrova, Y. E. , Nesterov, V. N. , Krouchinsky, N. G. , Ostapenko, V. A. , Neumann, R. , Neil, D. L. , & Jeffreys, A. J. (1996). Human minisatellite mutation rate after the Chernobyl accident. Nature, 380(6576), 683–686. 10.1038/380683a0 [DOI] [PubMed] [Google Scholar]
- Eads, B. D. , Andrews, J. , & Colbourne, J. K. (2008). Ecological genomics in Daphnia: Stress responses and environmental sex determination. Heredity, 100(2), 184–190. 10.1038/sj.hdy.6800999 [DOI] [PubMed] [Google Scholar]
- Edwards, A. W. F. (1971). Distances between populations on the basis of gene frequencies. Biometrics, 27(4), 873–881. 10.2307/2528824 [DOI] [PubMed] [Google Scholar]
- Ellegren, H. , Lindgren, G. , Primmer, C. R. , & Møller, A. P. (1997). Fitness loss and germline mutations in barn swallows breeding in Chernobyl. Nature, 389(6651), 593–596. 10.1038/39303 [DOI] [PubMed] [Google Scholar]
- Frankham, R. , Ballou, J. D. , & Briscoe, D. A. (2002). Introduction to conservation genetics. Cambridge University Press. [Google Scholar]
- Frankham, R. , Ballou, J. D. , & Briscoe, D. A. (2004). A primer of conservation genetics. Cambridge University Press. [Google Scholar]
- Geras'kin, S. A. , & Volkova, P. Y. (2014). Genetic diversity in Scots pine populations along a radiation exposure gradient. Science of the Total Environment, 496, 317–327. 10.1016/j.scitotenv.2014.07.020 [DOI] [PubMed] [Google Scholar]
- Gilpin, M. E. (1991). The genetic effective size of a metapopulation. Metapopulation dynamics: Empirical and theoretical investigations (pp. 165–175). Academic Press. [Google Scholar]
- Goodman, J. , Copplestone, D. , Laptev, G. V. , Gashchak, S. , & Auld, S. K. J. R. (2019). Variation in chronic radiation exposure does not drive life history divergence among Daphnia populations across the Chernobyl Exclusion Zone. Ecology and Evolution, 9, 2640–2650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goudet, J. , & Jombart, T. (2018). hierfstat: Estimation and tests of hierarchical F‐statistics. R Foundation for Statistical Computing. [Google Scholar]
- Haag, C. R. , Riek, M. , Hottinger, J. W. , Pajunen, V. I. , & Ebert, D. (2006). Founder events as determinants of within‐island and among‐island genetic structure of Daphnia metapopulations. Heredity, 96(2), 150–158. 10.1038/sj.hdy.6800774 [DOI] [PubMed] [Google Scholar]
- Haldane, J. B. S. (1937). The effect of variation on fitness. The American Naturalist, 71(735), 337–349. [Google Scholar]
- Halkett, F. , Simon, J. , & Balloux, F. (2005). Tackling the population genetics of clonal and partially clonal organisms. Trends in Ecology & Evolution, 20(4), 194–201. 10.1016/j.tree.2005.01.001 [DOI] [PubMed] [Google Scholar]
- Hanski, I. (1998). Metapopulation dynamics. Nature, 396, 41–49. 10.1038/23876 [DOI] [Google Scholar]
- Hartl, D. L. , & Clark, A. G. (1997). Principles of population genetics. Sinauer Associates. [Google Scholar]
- Hebert, P. D. N. (1974a). Enzyme variability in natural populations of Daphnia magna II. Genotypic frequencies in intermittent populations. Genetics, 77, 335–341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hebert, P. D. N. (1974b). Enzyme variability in natural populations of Daphnia magna II. Genotypic frequencies in permanent populations. Genetics, 77, 323–334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hebert, P. D. N. , & Ward, R. D. (1976). Enzyme variability in natural populations of Daphnia magna IV. Ecological differentiation and frequency changes of genotypes at audley end. Heredity, 36, 331–341. 10.1038/hdy.1976.40 [DOI] [PubMed] [Google Scholar]
- Hesterberg, T. , Monaghan, S. , Moore, D. S. , Clipson, A. , & Epstein, R. (2003). ‘Bootstrap methods and permutation tests’ in companion Chapter 18 to The practice of Business Statistics. [Google Scholar]
- Hinton, T. G. , Alexakhin, R. , Balonov, M. , Gentner, N. , Hendry, J. , Prister, B. , Strand, P. , & Woodhead, D. (2007). Radiation‐induced effects on plants and animals : Findings of the United Nations Chernobyl Forum. Health Physics, 93(5), 427–440. [DOI] [PubMed] [Google Scholar]
- Hobaek, A. , & Larsson, P. (1990). Sex determination in Daphnia magna . Ecological Society of America, 71(6), 2255–2268. 10.2307/1938637 [DOI] [Google Scholar]
- IAEA . (1992). Effects of ionising radiation on plants and animals at levels implied by current radiation protection standards. Technical Report Series No 332. [Google Scholar]
- IAEA . (2006). Environmental consequences of the chernobyl accident and their remediation: twenty years of experience (p. 167). IAEA. [Google Scholar]
- ICRP . (2009). Environmental protection: Transfer parameters for reference animals and plants. ICRP Publication 114. Annals of the ICRP, 39(6), 1–111. 10.1016/j.icrp.2011.08.009 [DOI] [PubMed] [Google Scholar]
- Intelligence Systems GEO . (2008). ‘Atlas Ukraine Radioactive Contamination’, on the order of the Ministry of Ukraine of Emergencies and Affairs of Population Protection from the Consequences of Chernobyl Catastrophe. Intelligence Systems GEO Ltd. [Google Scholar]
- Jansen, B. , Geldof, S. , De Meester, L. , & Orsini, L. (2011). Isolation and characterization of microsatellite markers in the water flea Daphnia magna . Molecular Ecology Resources, 11, 418–421.21429157 [Google Scholar]
- Jombart, T. (2008). adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics, 24, 1403–1405. 10.1093/bioinformatics/btn129 [DOI] [PubMed] [Google Scholar]
- Jombart, T. , & Ahmed, I. (2011). adegenet 1.3‐1: New tools for the analysis of genome‐wide SNP data. Bioinformatics, 27(21), 3070–3071. 10.1093/bioinformatics/btr521 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamvar, Z. N. , Tabima, J. F. , & Grünwald, N. J. (2014). Poppr: an R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ, 2, e281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khodarev, N. N. , Beckett, M. , Labay, E. , Darga, T. , Roizman, B. , & Weichselbaum, R. R. (2004). STAT1 is overexpressed in tumors selected for radioresistance and confers protection from radiation in transduced sensitive cells. Proceedings of the National Academy of Sciences of the United States of America, 101(6), 1714–1719. 10.1073/pnas.0308102100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimura, M. , & Maruyama, T. (1966). The mutational load with epistatic gene interactions in fitness. Genetics, 54(6), 1337–1351. 10.1093/genetics/54.6.1337 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuchma, O. , Vornam, B. , & Finkeldey, R. (2011). Mutation rates in Scots pine (Pinus sylvestris L.) from the Chernobyl exclusion zone evaluated with amplified fragment‐length polymorphisms (AFLPs) and microsatellite markers. Mutation Research/Genetic Toxicology and Environmental Mutagenesis, 725(1–2), 29–35. [DOI] [PubMed] [Google Scholar]
- Lande, R. (1993). Risks of population extinction from demographic and environmental stochasticity and random catastrophes. The American Naturalist, 142(6), 911–927. 10.1086/285580 [DOI] [PubMed] [Google Scholar]
- Lande, R. , & Shannon, S. (1996). The role of genetic variation in adaptation and population persistence in a changing environment. Evolution, 50(1), 434–437. 10.1111/j.1558-5646.1996.tb04504.x [DOI] [PubMed] [Google Scholar]
- Li, Y. , Korol, A. B. , Fahima, T. , Beiles, A. , & Nevo, E. (2002). Microsatellites: genomic distribution, putative functions, and mutational mechanism: A review. Molecular Ecology, 11(12), 2453–2465. [DOI] [PubMed] [Google Scholar]
- Lynch, M. (1987). The consequences of fluctuating selection for isozyme polymorphisms in Daphnia . Genetics, 115(4), 657–669. 10.1093/genetics/115.4.657 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marshall, J. S. (1962). The effects of gamma radiation on the intrinsic rate of natural increase of Daphnia pulex . Ecology, 43(4), 598–607. [Google Scholar]
- Mills, S. L. , & Smouse, P. E. (1994). Demographic consequences of inbreeding in remnant populations. The American Naturalist, 144(3), 412–431. 10.1086/285684 [DOI] [Google Scholar]
- Møller, A. P. (1993). Morphology and sexual selection in the barn swallow Hirundo rustica in Chernobyl, Ukraine. Proceedings of the Royal Society B: Biological Sciences, 252(1333), 3–19. [Google Scholar]
- Møller, A. P. (2002). Developmental instability and sexual selection in stag beetles from Chernobyl and a control area. Ethology, 108, 193–204. 10.1046/j.1439-0310.2002.00758.x [DOI] [Google Scholar]
- Møller, A. , & Mousseau, T. (2015). Strong effects of ionizing radiation from Chernobyl on mutation rates. Scientific Reports, 5, 8363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mort, M. A. , & Wolf, H. G. (1986). The genetic structure of large lake Daphnia populations. Evolution, 40, 756–766. [DOI] [PubMed] [Google Scholar]
- Muller, H. J. (1927). Artificial transmutation of the gene. Science, 66(1699), 84–87. 10.1126/science.66.1699.84 [DOI] [PubMed] [Google Scholar]
- Nazarov, A. , & Gudkov, D. (2009). Radiation monitoring of lake ecosystems within the Chernobyl accident exclusion zone. Presented at 13th World Lake Conference, Wuhan, China. [Google Scholar]
- Nei, M. , & Tajima, F. (1981). Genetic drift and estimation of effective population size. Genetics, 98, 625–640. 10.1093/genetics/98.3.625 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parisot, F. , Bourdineaud, J.‐P. , Plaire, D. , Adam‐Guillermin, C. , & Alonso, F. (2015). DNA alterations and effects on growth and reproduction in Daphnia magna during chronic exposure to gamma radiation over three successive generations. Aquatic Toxicology, 163, 27–36. 10.1016/j.aquatox.2015.03.002 [DOI] [PubMed] [Google Scholar]
- Ramana, C. V. , Boldogh, I. , Izumi, T. , & Mitra, S. (1998). Activation of apurinic/apyrimidinic endonuclease in human cells by reactive oxygen species and its correlation with their adaptive response to genotoxicity of free radicals. Proceedings of the National Academy of Sciences of the United States of America, 95(9), 5061–5066. 10.1073/pnas.95.9.5061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Real, A. , Sundell‐Bergman, S. , Knowles, J. F. , Woodhead, D. S. , & Zinger, I. (2004). Effects of ionising radiation exposure on plants, fish and mammals: Relevant data for environmental radiation protection. Journal of Radiological Protection, 24, A123–A137. [DOI] [PubMed] [Google Scholar]
- Schlotterer, C. , Vogl, C. , & Tautz, D. (1997). Polymorphism and locus‐specific effects on polymorphism at microsatellite loci in natural Drosophila melanogaster populations. Genetics, 146(1), 309–320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shestopalov, V. M. (1996). Atlas of the chernobyl exclusion zone. [Google Scholar]
- Slatkin, M. (1987). Gene flow and the geographic structure of natural populations. Science, 236(4803), 787–792. 10.1126/science.3576198 [DOI] [PubMed] [Google Scholar]
- Smirnov, D. A. , Brady, L. , Halasa, K. , Morley, M. , Solomon, S. , & Cheung, V. G. (2012). Genetic variation in radiation‐induced cell death. Genome Research, 22(2), 332–339. 10.1101/gr.122044.111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thielsch, A. , Brede, N. , Petrusek, A. , De Meester, L. , & Schwenk, K. (2009). Contribution of cyclic parthenogenesis and colonization history to population structure in Daphnia . Molecular Ecology, 18(8), 1616–1628. [DOI] [PubMed] [Google Scholar]
- Walser, B. , & Haag, C. R. (2012). Strong intraspecific variation in genetic diversity and genetic differentiation in Daphnia magna: the effects of population turnover and population size. Molecular Ecology, 21(4), 851–861. [DOI] [PubMed] [Google Scholar]
- Weir, B. S. , & Cockerham, C. C. (1984). Estimating f‐statistics for the analysis of population structure. Evolution, 38(6), 1358–1370. [DOI] [PubMed] [Google Scholar]
- Wright, S. (1951). The genetical structure of populations. Annals of Eugenics, 15(4), 323–354. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Material
Data Availability Statement
Microsatellite marker data are published on Dryad: https://doi.org/10.5061/dryad.0cfxpnw48.