

Abstract
The ecological dysbiosis of a biofilm includes not only bacterial changes but also changes in their metabolism. Related to oral biofilms, changes in metabolic activity are crucial endpoint, linked directly to the pathogenicity of oral diseases. Despite the advances in caries research, detailed microbial and metabolomic etiology is yet to be fully clarified. To advance this knowledge, a meta‐taxonomic approach based on 16S rRNA gene sequencing and an untargeted metabolomic approach based on an ultra‐high performance liquid chromatography‐quadrupole time‐of‐flight mass spectrometry analysis (UHPLC/Q‐TOF‐MS) were conducted. To this end, an in vitro biofilm model derived from the saliva of healthy participants were developed, under commensal and cariogenic conditions by adding sucrose as the disease trigger. The cariogenic biofilms showed a significant increase of Firmicutes phyla (p = 0.019), due to the significant increase in the genus Streptococcus (p = 0.010), and Fusobacter (p < 0.001), by increase Fusobacterium (p < 0.001) and Sphingomonas (p = 0.024), while suffered a decrease in Actinobacteria (p < 0.001). As a consequence of the shift in microbiota composition, significant extracellular metabolomics changes were detected, showed 59 metabolites of the 120 identified significantly different in terms of relative abundance between the cariogenic/commensal biofilms (Rate of change > 2 and FDR < 0.05). Forty‐two metabolites were significantly higher in abundance in the cariogenic biofilms, whereas 17 metabolites were associated significantly with the commensal biofilms, principally related protein metabolism, with peptides and amino acids as protagonists, latter represented by histidine, arginine, l‐methionine, glutamic acid, and phenylalanine derivatives.
Keywords: amino‐acids, biofilms, caries, metabolomics, metataxonomics, peptides, Streptococcus
1. INTRODUCTION
A vast range of bacteria are present in the mouth. As with other human body niches, the mouth provides various surfaces (e.g., teeth, gingival crevices, tongue, and keratinized and nonkeratinized mucosal surfaces) for the colonization of bacteria, which follow a tactic of structured and organized community building by means of a self‐produced matrix of extracellular polymeric substances. These biofilms favor the persistence of bacteria in the various oral locations, as well as the synergic and antagonistic interactions between individual species and between themselves, the environment, and the host, playing a key role in numerous human infections, conferring lower efficacy to antiseptic treatments and a high frequency of antibiotic resistance. Biofilm‐induced dental diseases, such as caries and periodontitis, are oral health concerns despite professional interventions and daily healthcare routines, with evidence implicating biofilms in several extraoral inflammatory diseases (Costerton et al., 1995; Hung et al., 2013; Marsh et al., 2011; Rosier et al., 2014; Sanz et al., 2017).
Teeth have a characteristic nonscaling rigid surface exposed to a medium rich in microorganisms and nutrients, which facilitates the formation of biofilms (the supragingival dental plaque) located in the area of the gingival margin, in proximal spaces, and in occlusal surfaces. These biofilms maintain a stable microbial composition with no significant fluctuations over time in the diverse niches, coexisting in homeostasis with the host (Marsh, 2006). In this environment, the interaction between species, biofilm, and host is reflected in the demineralization and remineralization process that, in most people, frequently takes place during the day (Selwitz et al., 2007). In homeostasis, several bacteria in the dental plaque, mainly Streptococcus spp. and Lactobacillus spp., produce weak organic acids as a by‐product of the metabolism of fermentable carbohydrates, leading to local tooth demineralization, which is however easily neutralized by the buffering capacity of saliva and other bacteria of the biofilm, maintaining the repair status quo and even reversing the possible damage (Simon‐Soro et al., 2013). Numerous host factors, such as dietary carbohydrate consumption, salivary flow and composition dysfunction, and poor oral hygiene (which can lead to plaque accumulation) can, however, disrupt this equilibrium and are directly linked to bacterial biofilm dysbiosis and thus to the onset and progression of dental tissue damage (Featherstone, 2004; Mira et al., 2017). As the microbiota shifts, there is a proliferation of aciduric representatives of Streptococcus, Lactobacillus, Bifidobacterium, Actinomyces, Veillonella, and certain yeasts, among other bacteria (Mira et al., 2017; Takahashi & Nyvad, 2011). The shift in microbiota composition results in increased time during which the oral pH is below the critical point for the demineralization process, resulting in dental tissue demineralization. However, the lesion progresses when the dysbiotic biofilm matures and remains on the teeth for prolonged periods, resulting in dental caries. If a cavity is allowed to develop, the site provides an ecological niche in which microorganisms progressively adapt to the reduced pH and offers protection against removal by the patient (e.g., through brushing) (Fejerskov, 2004; Kidd & Fejerskov, 2004; Selwitz et al., 2007).
For dental caries, the ecological dysbiosis includes not only bacterial changes in the biofilm but also changes in their metabolism (Fejerskov, 2004; Selwitz et al., 2007; Takahashi & Nyvad, 2011; ten Cate, 2009). Given that homeostasis disruption drives changes in species‐specific oral biofilm composition, it is unsurprising that the metabolites secreted to the local environment also change. Changes in bacterial metabolic activity are crucial endpoints, because they directly relate to the pathogenicity of oral diseases (e.g., organic acids to dental caries, short fatty acids and sulfides to periodontal diseases, and sulfides and ammonia to halitosis) (Kleinberg, 2002; Takahashi, 2015; Takahashi & Nyvad, 2008, 2011). In this context, bacterial omics approaches, including metagenomics and metabolomics, might provide information for understanding the role of the microbiome and the associated metabolome in conditions that favor the development of dental caries (Nascimento et al., 2017; Washio et al., 2016).
Despite the advances in this field, detailed microbial and metabolomic etiology remains the main focus of caries research and has yet to be fully clarified. To advance this knowledge, we conducted a meta‐taxonomic approach based on 16S rRNA gene sequencing and an untargeted metabolomic approach based on an ultra‐high performance liquid chromatography‐quadrupole time‐of‐flight mass spectrometry analysis (UHPLC/Q‐TOF‐MS). To this end, we used an in vitro biofilm model derived from the saliva of healthy participants, under commensal and cariogenic conditions by adding sucrose as the disease trigger. In addition to saliva, dietary sugars are one of the main sources of nutrients for dental plaque. Among them, sucrose is considered a highly cariogenic substrate, given that bacterial biofilms metabolize sucrose rapidly to generate large amounts of acids and produce intracellular and extracellular polysaccharides as additional virulent mechanisms (Sanz et al., 2017). In addition to the metabolites derived from fermentable carbohydrates, bacteria produce several metabolites that can also alter the conditions of the oral cavity, creating a potentially more pathogenic environment; however, their possible roles in caries have not been extensively researched (Nascimento et al., 2017; Washio et al., 2016). These approaches therefore provide insights into the dysbiosis in the composition and function of oral biofilms related to caries.
2. MATERIALS AND METHODS
2.1. In vitro saliva‐derived multispecies biofilm culturing
The study protocol was reviewed and approved by the Ethics Committee of the San Carlos Clinic Hospital, Madrid, Spain (C.I. 20/118‐E). All experiments and data collection were performed in accordance with relevant guidelines and regulations. Eight healthy participants between 25 and 30 years of age participated in the study after giving their written informed consent. All participants were included in the study based on careful periodontal and caries examinations, and none presented clinical symptoms of oral or systematic disease. The study's exclusion criteria included the presence of salivary gland or oral mucosa disease; diverse oral diseases (e.g., caries, gingivitis, periodontal disease, infectious processes); orthodontic treatment; severe systemic disease; diabetes; use in the last 3 months of antibiotics or medicine that affects oral health, salivary secretion or the periodontium; the use of psychotropic drugs; smoking more than 10 cigarettes/day; and pregnancy or lactation. Figure 1 depicts an overview of the experimental design.
FIGURE 1.

Scheme of the experimental tests carried out in the study. For abbreviations, see the text
2.1.1. Saliva collection
The participants were asked to refrain from eating and drinking 2 h before donating saliva; the saliva was donated 12 h after the last tooth brushing. Approximately 5 ml of spontaneous, unstimulated whole saliva was expectorated into sterile 50‐ml plastic tubes (Cellstar Tubes, Greiner Bio‐one, Frickenhausen, Germany). Saliva samples were pooled and centrifuged at 2500 rpm for 10 min to separate large debris and eukaryotic cells. The supernatant, referred to as the pooled saliva, was recovered and employed for two purposes: (1) as the biofilm inoculum; a portion of the pooled saliva was diluted in sterile glycerol (final concentration of 20%; Sigma, St. Louis, MO, USA), aliquoted, and stored at −80°C; and (2) to generate the acquired pellicle; 10‐ml aliquots were treated with 2.5 mM DL‐dithiothreitol (Sigma) for 10 min with stirring to reduce salivary protein aggregation. The treated saliva was then centrifuged (10 min, 4°C, 12,000 rpm), and the supernatant was diluted to 50% with phosphate buffered saline solution (PBS; pH, 7.4), filtered and sterilized through a 0.22‐μm pore size Millex GV low‐protein‐binding filter X50 (Millipore; Millipore Corp., Bedford, MA, USA) and stored at −20°C. The efficacy of this protocol was assessed by plating processed saliva samples onto supplemented blood agar plates (Blood Agar Oxoid No 2; Oxoid, Basingstoke, UK), supplemented with 5% (v/v) sterile horse blood (Oxoid), 5.0 mg/ml hemin (Sigma) and 1.0 mg/ml menadione (Merck, Darmstadt, Germany) for 72 h at 36 ± 1°C. No growth was observed either on the aerobically or the anaerobically incubated plates.
2.1.2. In vitro saliva‐derived biofilm model
An in vitro biofilm model was developed from the saliva of the healthy participants. The same model but compatible with a cariogenic biofilm was developed in parallel, using sucrose as a trigger for the cariogenic condition (based on unpublished results of biofilm development in both, commensal and cariogenic conditions, and acid production/final pH as virulence‐related outcomes; see Supporting Information S1). To this end, 24‐well plates (Greiner Bio‐one) were precoated with 200 μl/well of filtered bacterial‐free saliva, allowing for attached pellicle growth, and incubated at 36 ± 1°C for 2 h. The pooled saliva (20 μl), used as the biofilm inoculum, was then inoculated per well. For the commensal condition and to support the growth of different bacterial subpopulations, 2.0 ml of brain‐heart infusion medium (BHI; Becton, Dickinson and Company, Franklin Lakes, NJ, USA) was added to each well. For the disease condition and to simulate a cariogenic challenge, 2.0 ml of BHI supplemented with 0.5% of sucrose (Sigma) was added. In both cases, the plates were incubated at 36 ± 1°C under aerobic conditions with 5.5% CO2 (5.5% CO2 gasifying the atmosphere with ultra‐pure carbon dioxide; Carbon dioxide Premier‐X40S, Carburos, Air Products, Cornellá de LLobregat, Spain) without disturbance for 96 h to allow for biofilm formation.
After incubation, the cell‐free supernatants (CFSs) were carefully recovered and reserved for the metabolomics approach at −80°C. After withdrawing the supernatant, the wells were rinsed with PBS three times to remove nonadherent bacteria (10 s per rinse). After rinsing, the biofilm was detached by successive and vigorous pipetting of 1 ml of sterile PBS in the well. Once removed, the biofilm was dispersed by vortexing for 3 min. The samples were then centrifuged for 3 min at 13,000 rpm, and the pellets were kept at −20°C until use in the metataxonomic approach.
Each biofilm phenotype and the corresponding CFSs were developed in parallel in three separate trials with three repetitions per test (n = 9 for each biofilm and CFS). In total, 18 in vitro saliva‐derived biofilms and CFSs were developed, nine compatible with the commensal condition and nine with the caries disease.
2.2. Metataxonomic analysis
To study the possible shift in the supragingival bacterial communities related to caries disease, we conducted a comparative study of the bacterial composition of both in vitro biofilm phenotypes. To this end, we used the biofilms described in Section 2.1.2.
2.2.1. DNA extraction and 16S rRNA sequencing
For the metataxonomic analysis, the DNA of the two types of developed biofilms was extracted using the Ultra‐Deep Microbiome Prep Kit (Molzym Gmbh & Co. KG. Bremen, Germany), according to the manufacturer's protocol for bacteria. The DNA was eluted in 100 μl of sterile water (Roche Diagnostic GmbH; Mannheim, Germany) and frozen at −20°C for further analysis. A negative control blank, which included no sample, was subjected to all steps of the DNA extraction procedure described above.
The composition and structure of the microbial communities was assessed through the amplification and sequencing of the V3‐V4 variable regions of the 16S rRNA gene. The Illumina Miseq sequencing 300 × 2 approach was used. Amplification was performed after 25 polymerase chain reaction cycles. A negative control of the DNA extraction was included, as well as a positive Mock Community control to ensure quality control.
2.2.2. Bioinformatic analysis of the sequencing data
The sequences were clustered into operational taxonomic units (OTUs) at the 97% similarity level. Bioinformatics was performed using Dada2 (Callahan et al., 2016), and the phylogeny assessment was performed using Mafft (Katoh & Standley, 2013) and Fasttree (Price et al., 2009). We employed the phylotype data to calculate the following alpha diversity metrics: observed OTUs (community richness), evenness (or Pielou's Evenness; a measure of community evenness), and Shannon's diversity index (quantitative measure of community richness). We employed the phylotype and phylogenetic data to calculate the following beta diversity metrics: unweighted Unifrac distance (phylogenetic qualitative measure), weighted Unifrac distance (phylogenetic quantitative measure), Jaccard distance (qualitative measure), and Bray Curtis distance (quantitative measure).
The taxonomic assignment of phylotypes was performed using a Bayesian classifier trained with Silva database version 132 (99% OTUs full‐length sequences) (Q. Wang et al., 2007). The data were further analyzed as follows: Alpha diversity comparisons were performed using the Kruskal–Wallis nonparametric test. Beta diversity distance matrices were used to calculate the principal coordinate analysis (PCoA) and to create ordination plots using the R software package version 3.6.0. The significance of the groups present in the community structure was tested using permutational multivariate analysis of variance (PERMANOVA) and analysis of similarities (ANOSIM) tests. The Permdisp test was used to identify location versus dispersion effects (M. Anderson & Walsh, 2013). The significance threshold was set at 0.05. The differential relative abundance of taxa was tested using two methods: analysis of compositions of microbiomes (ANCOM; Mandal et al., 2015) and the Kruskal–Wallis nonparametric test. After employing the Kruskal–Wallis test, we used Conover's test with false discovery rate (FDR) Benjamini–Hochberg correction for the pairwise comparison. The significant threshold was set at 0.05. We employed BiodiversityR version 2.11‐1, PMCMR version 4.3, RVAideMemoire version 0.9‐7 and vegan version 2.5‐5 packages for the various statistical analyses.
2.3. Metabolomic analysis
To study the possible ecological dysbiosis, we conducted a comparative study of extracellular metabolites secreted to the environment by both biofilm phenotypes. After incubating the biofilms as described in Section 2.1.2 and before removing the biofilms for metataxonomic analysis, the CFS was obtained by centrifuging the supernatant at 1750 rpm, for 10 min at 4°C to collect the extracellular metabolites without bacterial cells. Following centrifugation, the CFS was stored at −80° until the analysis.
2.3.1. Ultra‐high performance liquid chromatography‐quadrupole time‐of‐flight mass spectrometry analysis
The metabolites were extracted from the CFS samples using methanol. Specifically, 200 μl of each CFS was mixed with 600 μl of methanol, agitated for 5 min at 2000 rpm at 4°C, placed in an ultrasonic bath for 10 min on ice, and incubated at −20°C for 30 min. After that, the samples were centrifuged for 15 min at 14,800 rpm at 4°C. We collected 400 μl of supernatant, which we then dried and suspended in 100 μl of 75% methanol (v/v H2O). Lastly, the samples were vortexed for 1 min and centrifuged at 14,800 rpm for 5 min at 4°C; the supernatants were then collected and stored at −80°C until ultra‐high performance liquid chromatography‐quadrupole time‐of‐flight mass spectrometry analysis (HPLC‐MS/MS) . An extraction blank was prepared following the same metabolite extraction protocol followed for the samples. The samples were injected randomly to eliminate any drift effect that the equipment, the analysis conditions, and so on could present. Blanks were intercalated after each injection to control the correct elution of each sample's components, to avoid contamination between samples (carry‐over), and to improve reproducibility. Aliquots of 2 μl were injected randomly into a liquid chromatography–tandem mass spectrometry (LC–MS/MS) system consisting of a quadrupole Q‐TOF series 6540 coupled to a HPLC (model 1290) both from Agilent Technologies (Germany), equipped with an Agilent Jet Stream (AJS) thermal orthogonal ESI source. MS control, data acquisition, and data analysis were carried out using the Agilent Mass Hunter Qualitative Analysis software (B.10.0). For the chromatographic separation, an Eclipe Plus C18 analytical column (100 × 2.1 mm, particle size 1.8 μm) and a C18 guard column (0.5 cm × 2.1 mm, particle size 1.8 μm), both from Agilent (Germany), were used. The column temperature was held at 40°C. Water (LC‐MS grade) was used as mobile phase (A) and ACN as mobile phase (B), and 0.1% formic acid was used as mobile phase modifier. The following mobile phase gradient was delivered at a flow rate of 0.5 ml min−1: start 0%–30% B, 7 min hold; linear gradient 30%−80% B in 9 min; hold 80%–100% B, 11 min; 100% B, 14 min. The mass spectrometer was operated using the following parameters: capillary voltage of 3000 V; mass range from 25 to 1100 m/z; nebulizer pressure of 40 psig; drying gas flow rate of 8 L/min and 300°C. The sheath gas flow was 11 L/min at 350°C. Tandem mass spectrometry (MS/MS) analyses were performed employing the auto MS/MS mode using five precursors per cycle, dynamic exclusion after two spectra (released after 0.5 min), and collision energies of 20 and 40 V. For proper mass accuracy, spectra were corrected using ions m/z 121.0509 (C5H4N4) and 922.0098 (C18H18O6N3P3F24), simultaneously pumped into the ionization source.
2.3.2. Statistical analysis
LC‐MS raw data files were converted to mzML and data processing was then performed using MS‐DIAL (v. 4.12) software for deconvolution, peak picking, alignment, and identification. The MSP file used for annotation was generated by combining MS/MS spectra from NIST20 MS/MS database, the LipidBLAST mass spectral library, and the MassBank of NorthAmerica database (MoNA, available at https://mona.fiehnlab.ucdavis.edu/downloads). Peak height calculation was performed by using the median of the three technical replicates, and combining data for different detected molecular species for each particular compound ([M+H]+, [M+NH4]+, [M+Na]+, [M+K]+, [2 M+H]+, [2 M+NH4]+, [2 M+Na]+, [2 M+K]+adducts).
The list of metabolites was then filtered removing unknown metabolites, metabolites with a maximum height below 1000 units, or metabolites with a maximum height below three times the average height in the extraction blanks. Missing values were imputed by half of the minimum height value.
Before carrying out the statistical study, the normal distribution of the data was verified and scaled using the “Auto‐Scaling” function (centered on the mean and divided by the standard deviation of each variable). The statistical analysis was carried out using the software implemented in the Metaboanalyst 5.0 tool. A principal component analysis (PCA) was carried out using all the samples from the two groups at the same time to detect possible outliers. Thereafter, a supervised partial least square‐discriminant analysis (PLS‐DA) was performed, and variable importance in projection (VIP) scores were obtained and considered significant when VIP scores > 1.0. Finally, fold changes between the two groups were calculated and evaluated by using the non‐parametric Mann–Whitney U test. Metabolites were considered significantly altered when FDR < 0.05 and fold change (FC) > 2.
3. RESULTS
3.1. Metataxonomic analysis
We employed a marker‐based approach using the rRNA16S gene to study the bacterial diversity of 18 in vitro saliva‐derived biofilm samples, nine compatible with commensal conditions and nine with caries disease. After quality filtering, denoising, paired‐ends joining, and chimera filtering, a total of 267,852 high‐quality reads for the 18 samples of both biofilm conditions could be assigned to 245 OTUs (97% similarity), with a mean amplicon length of the high‐quality reads of 420−460 bp. The rarefaction plots revealed how the achieved sequencing depth was enough to observe the complete diversity present in both microbial communities (data not shown). The reads were further classified into nine phyla, 13 classes, 21 orders, 31 families, 36 genera, and 52 species (Supporting Information S2). Table 1 summarizes the dominant bacteria in the commensal and cariogenic biofilms.
TABLE 1.
Dominant OTUs and its proportion (%) in the biofilm model in commensal condition and supplemented with sucrose (cariogenic group) at all taxonomic levels
| Biofilm model derived from saliva (commensal condition) | |||||
|---|---|---|---|---|---|
| Phylum | Class | Order | |||
| Firmicutes | 64.80% | Bacilli | 64.77% | Lactobacillales | 34.66% |
| Actinobacteriota | 34.22% | Actinobacteria | 34.22% | Staphylococcales | 30.11% |
| Proteobacteria | 0.56% | Gammaproteobacteria | 0.54% | Bifidobacteriales | 26.60% |
| Bacteroidota | 0.18% | Bacteroidia | 0.18% | Micrococcales | 7.39% |
| Fusobacteriota | 0.07% | Fusobacteriia | 0.07% | Pasteurellales | 0.53% |
| Patescibacteria | 0.11% | Saccharimonadia | 0.10% | Actinomycetales | 0.23% |
| Undeffined | 0.03% | Clostridia | 0.02% | Bacteroidales | 0.18% |
| Campilobacterota | 0.01% | Negativicutes | 0.01% | Fusobacteriales | 0.07% |
| Cyanobacteria | 0.01% | Alfaproteobacteria | 0.01% | Saccharimonadales | 0.10% |
| Campylobacteria | 0.01% | Selenomonadales | 0.03% | ||
| Family | Genus | Species | |||
|---|---|---|---|---|---|
| Streptococcaceae | 31.37% | Streptococcus | 31.38% | Alloscardovia_omnicolens | 26.60% |
| Bifidobacteriaceae | 26.80% | Alloscardovia | 26.80% | Staphylococcus sp. | 24.46% |
| Staphylococcaceae | 24.64% | Staphylococcus | 24.65% | Streptococcus_salivarius | 15.96% |
| Micrococcaceae | 7.45% | Rothia | 7.45% | Streptococcus sp. | 9.22% |
| Gemellaceae | 5.70% | Gemella | 5.70% | Rothia sp. | 7.39% |
| Carnobacteriaceae | 2.12% | Granulicatella | 2.12% | Gamella sp. | 5.65% |
| Lactobacillaceae | 1.43% | Lactobacillus | 1.43% | Streptococcus parasanguinis | 5.31% |
| Pasteurellaceae | 0.54% | Haemophilus | 0.54% | Granulicatella sp. | 2.11% |
| Actinomycetaceae | 0.23% | Actinomyces | 0.23% | Lactobacillus sp. | 1.42% |
| Porphyromonadaceae | 0.08% | Prevotella | 0.08% | Streptococcus anginosus | 0.65% |
| Sucrose saliva‐derived biofilm model (cariogenic condition) | |||||
|---|---|---|---|---|---|
| Phylum | Class | Order | |||
| Firmicutes | 96.47% | Bacilli | 96.29% | Lactobacillales | 85.48% |
| Bacteroidota | 0.69% | Actinobacteria | 1.60% | Staphylococcales | 10.80% |
| Proteobacteria | 0.60% | Bacteroidia | 0.69% | Bifidobacteriales | 1.02% |
| Actinobacteriota | 1.60% | Gammaproteobacteria | 0.55% | Micrococcales | 0.56% |
| Fusobacteriota | 0.42% | Fusobacteriia | 0.42% | Bacteroidales | 0.65% |
| Patescibacteria | 0.14% | Negativicutes | 0.14% | Pasteurellales | 0.51% |
| Campilobacterota | 0.06% | Saccharimonadia | 0.12% | Fusobacteriales | 0.42% |
| Clostridia | 0.08% | Veillonelalles | 0.10% | ||
| Campylobacteria | 0.06% | Saccharimonadales | 0.12% | ||
| Gracilibacteria | 0.03% | Campylobacterales | 0.06% | ||
| Family | Genus | Species | |||
|---|---|---|---|---|---|
| Streptococcaceae | 84.78% | Streptococcus | 84.83% | Streptococcus sp. | 36.47% |
| Bifidobacteriaceae | 1.04% | Gemella | 5.88% | Streptococcus_salivarius | 35.60% |
| Staphylococcaceae | 5.06% | Staphylococcus | 5.06% | Streptococcus parasanguinis | 11.60% |
| Micrococcaceae | 0.56% | Alloscardovia | 1.04% | Gemella sp. | 5.81% |
| Gemellaceae | 5.88% | Granulicatella | 0.82% | Staphylococcus sp. | 5.00% |
| Carnobacteriaceae | 0.82% | Lactobacillus | 0.98% | Alloscardovia_omnicolens | 1.02% |
| Lactobacillaceae | 0.98% | Rothia | 0.56% | Lactobacillus sp. | 0.96% |
| Pasteurellaceae | 0.51% | Haemophilus | 0.51% | Granulicatella sp. | 0.81% |
| Prevotellaceae | 0.36% | Fusobacterium | 0.36% | Rothia sp. | 0.56% |
| Fusobacteriaceae | 0.36% | Prevotella | 0.33% | Haemophilus sp. | 0.42% |
3.1.1. Commensal in vitro saliva‐derived multispecies biofilm model
We first established an in vitro saliva‐derived multispecies biofilm model and tested whether this model could reproduce the in vivo situation in oral health conditions. After the quality control, we included nine samples for the analysis in these conditions and assigned 129,744 reads (median/sample, 14,416; range, 12,275−18,299) into nine phyla and further classified them into 13 classes, 21 orders, 28 families, 29 genera, and 38 species. In these commensal conditions, two phyla were the most abundant (Table 1), representing more than 98% of the total sequences: Firmicutes (64.80%) and Actinobacteria (34.22%), followed by Proteobacteria (0.56%), although with low relative abundance. At the genus level, the five most abundant of the 29 detected genera in the commensal biofilms were Streptococcus (31.38%), Alloscardovia (26.80%), Staphylococcus (24.65%), Rothia (7.57%), and Gemella (5.70%) (Table 1). Table 1 shows the dominant species in the commensal biofilm group, highlighting the presence of Alloscardovia omnicolens (26.60%), Staphylococcus spp (24.46%), Streptococcus salivarius (15.96%), Streptococcus spp (9.22%), and Rothia spp (7.39%).
3.1.2. Cariogenic in vitro saliva‐derived multispecies biofilm model
We then demonstrated that the biofilms respond to the imbalance in sucrose levels. After preprocessing and filtering, nine samples were included in the analysis. We assigned 138,108 reads (median/sample, 15,345; range, 11,549−27,487) into seven phyla: Firmicutes (96.47%), clearly dominant and in relative abundance, Actinobacteria (1.60%), Bacteroidetes (0.69%), Proteobacteria (0.60%), Fusobacteria (0.42%), Patescibacteria (0.14%) and Campylobacteria (0.06%). These OTUs were further classified into 11 classes, 18 orders, 28 families, 33 genera, and 47 species. Among the 33 detected genera, the five most abundant in the cariogenic group were Streptococcus (84.83%), Gemella (5.88%), Staphylococcus (5.06%), Alloscardovia (1.04%), and Granulicatella (0.82%) (Table 1). Table 1 shows Streptococcus spp (36.47%), S. salivarius (35.60%), and Streptococcus parasanguinis (11.81%) as the dominant species, followed by Staphylococcus spp (5.00%) and Alloscardovia omnicolens (1.02%).
3.1.3. Taxonomic dysbiosis of the commensal biofilm toward a cariogenic phenotype
The analysis of the alpha diversity of the bacterial compositions of saliva‐derived biofilms, which were cultured in such a way as to have commensal or cariogenic phenotypes, did not reveal statistically significant differences between the groups in regard to microbial richness (p = 0.13) (Figure 2a). The evenness, which represents the uniformity of the communities, also showed no statistically significant differences among the commensal and cariogenic biofilms (p = 0.40) (Figure 2b). In terms of community diversity, Shannon's diversity index accounts for both the abundance and evenness of the species present; however, the index revealed no statistically significant differences between the two biofilm conditions (p = 0.76) (Figure 2c).
FIGURE 2.
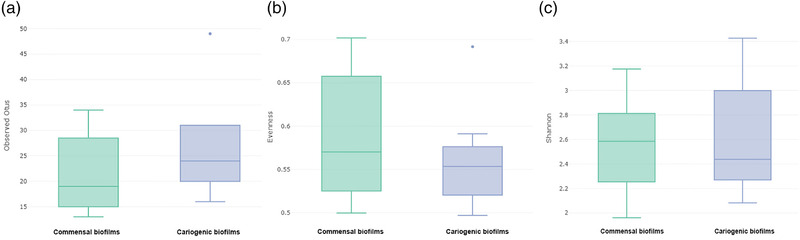
Box plots of microbial (a) richness as revealed by OTU number; (b) evenness, representing the uniformity of communities; (c) Shannon's diversity index, accounts for both abundance and evenness of the species present. No statistically differences were found in any analysis
To evaluate the similarity in bacterial composition among the samples and thereby explore the relationships between bacterial communities in the commensal and cariogenic conditions, we performed a PCoA from which two clear patterns emerged, indicating that the commensal and cariogenic biofilms could not be grouped into one cluster (Figure 3). The cariogenic group appeared to be more clustered, unlike the commensal biofilms, which appeared more dispersed. The confirmation of the statistically significant difference in the bacterial composition of the cariogenic biofilms compared with the commensal biofilms was revealed by the four distances used (PERMANOVA results). The results showed statistically significant differences between the two groups by the qualitative (unweighted UniFrac distance of p < 0.017) and quantitative (weighted Unifrac distance of p = 0.003) phylogenetic measures and by the Jaccard distance (p = 0.001) and Bray–Curtis distance (p = 0.001).
FIGURE 3.
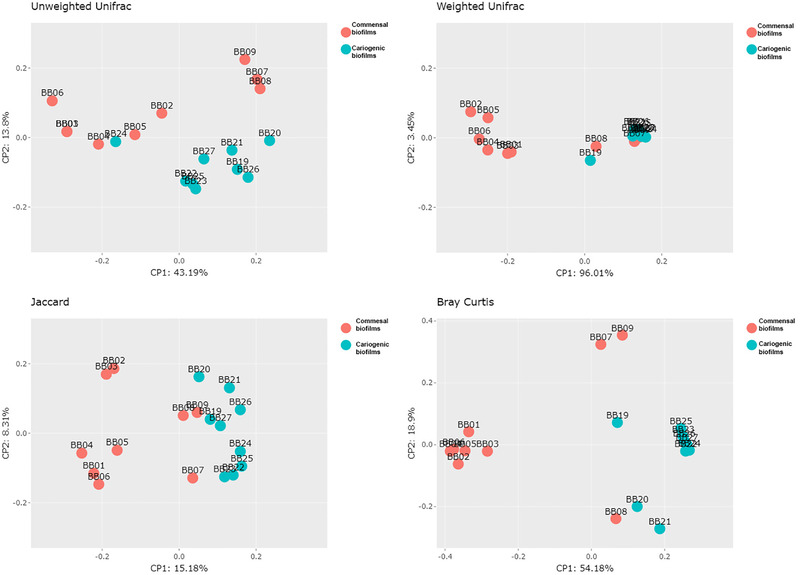
Caries biofilms group separation from commensal ones by principal coordinates analysis (PCoA), displaying the distribution among the 18 samples; ANOSIM, Permanova, and Permdisp tests were carried to test for significance of the a priori defined groups. The results show significant differences between both groups by phylogenetic qualitative (Permanova Analysis for Unweighted Unifrac p‐value = 0.017) and quantitative (Permanova Analysis for Weighted Unifrac distance p‐value = 0.003) measures, also showed by Jaccard distance (Permanova Analysis p‐value = 0.001) and Bray Curtis distance (Permanova Analysis p‐value = 0.001)
In terms of the beta‐diversity and relative abundance in the commensal and cariogenic biofilms, the Kruskal–Wallis tests showed changes in microbial community assembly after the sucrose treatments and revealed statistically significant differences (p ≤ 0.05) at different taxonomic levels, specifically in three phyla, four classes, five orders, six families, six genera, and nine species (Table 2). At the phylum level, there was a significant decrease in Actinobacteria in the cariogenic biofilms when compared with the commensal biofilms (p < 0.001), mainly due to the significant reduction in the genera Alloscardovia (p < 0.001) and Rothia (p < 0.001) (Figure 4a–c). In the cariogenic biofilms, the phylum Firmicutes increased significantly (p = 0.019), reflecting the significant increase in the genus Streptococcus (p = 0.010); however, there was a decrease in the genus Staphylococcus (p < 0.001) (Figure 4d–f). The phylum Fusobacter also increased significantly in the cariogenic biofilms (p < 0.001), reflecting the increase in the genera Fusobacterium (p < 0.001) and Sphingomonas (p = 0.024) (Figure 4g–i).
TABLE 2.
Changes in microbial community assembly after the sucrose treatments in terms of the beta‐diversity and relative abundance in the commensal and cariogenic biofilms. Statistically significant differences at p ≤ 0.05 at different taxonomic levels
| Phylum | p‐Value | Class | p‐Value | Order | p‐Value |
|---|---|---|---|---|---|
| Firmicutes | 0.019 | Bacilli | 0.019 | Lactobacillales | 0.004 |
| Actinobacteriota | <0.001 | Actinobacteria | <0.001 | Staphylococcales | 0.007 |
| Fusobacteriota | <0.001 | Fusobacteriia | <0.001 | Bifidobacteriales | <0.001 |
| Alfaproteobacteria | 0.024 | Micrococcales | <0.001 | ||
| Sphingomonadales | 0.024 |
| Family | p‐Value | Genus | p‐Value | Species | p‐Value |
|---|---|---|---|---|---|
| Streptococcaceae | 0.010 | Streptococcus | 0.010 | Streptococcus sp. | <0.001 |
| Bifidobacteriaceae | 0.001 | Alloscardovia | <0.001 | Streptococcus parasanguinis | 0.002 |
| Staphylococcaceae | <0.001 | Staphylococcus | <0.001 | Streptococcus anginosus | <0.001 |
| Micrococcaceae | <0.001 | Rothia | <0.001 | Staphylococcus sp. | <0.001 |
| Fusobacteriaceae | <0.001 | Fusobacterium | <0.001 | Rothia sp. | <0.001 |
| Sphingomonadaceae | 0.024 | Sphingomonas | 0.024 | Alloscardovia_omnicolens | <0.001 |
| Prevotella melaninogenica | 0.020 | ||||
| Fusobacterium periodonticum | <0.001 | ||||
| Sphingomonas sp. | 0.024 |
FIGURE 4.
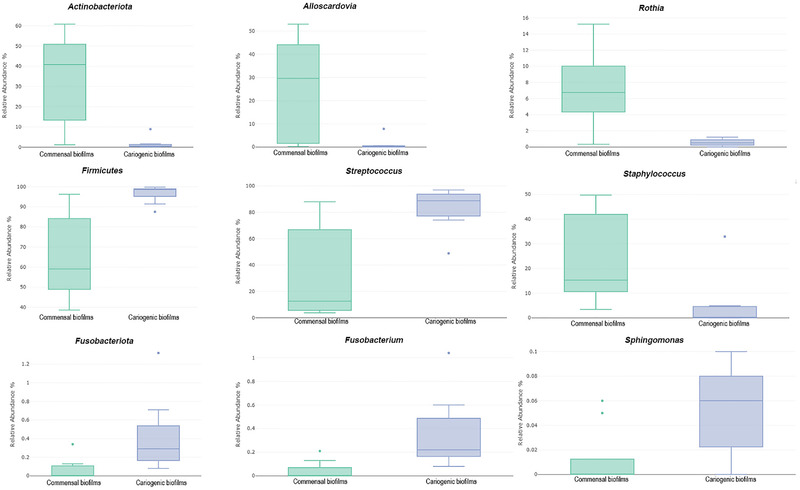
Representatives box plots of relative abundance in the commensal and cariogenic biofilms at different taxonomic levels
3.2. Metabolomic analysis
The CFSs from the biofilms developed under healthy and cariogenic environments were analyzed using UHPLC/Q‐TOF‐MS. We identified 120 known metabolites from all the metabolomics features observed (Supporting Information S3). The principal component analysis, which we employed to analyze the possible clustering of samples and determine the possible outliers, revealed that the cariogenic CFS biofilms appeared close together but separate from the commensal group samples (Figure 5a), according to the dysbiosis observed by the metataxonomic analysis. To highlight the differences between the metabolites of the CFSs from the biofilms evolved under commensal and cariogenic conditions, we then performed a PLS‐DA analysis. Once again, the cariogenic and commensal biofilms were clearly clustered (Figure 5b). The VIP score obtained from the PLS‐DA revealed the metabolites that best explain the grouping obtained in the PLS‐DA analysis (Figure 6).
FIGURE 5.
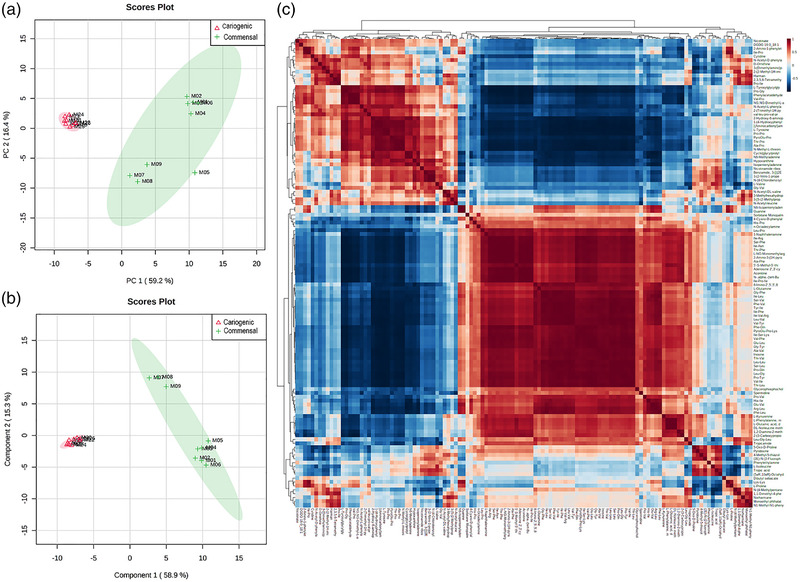
Representation of: (a) principal component analysis (PCA) of both cell free supernatants (CFSs) from cariogenic and commensal biofilms. As can be seen, cariogenic CFSs appear very close together and separated from the samples from the commensal group; (b) PLS‐DA of all CFSs samples (cariogenic vs. commensal); (c) correlation analysis of all CFSs samples
FIGURE 6.
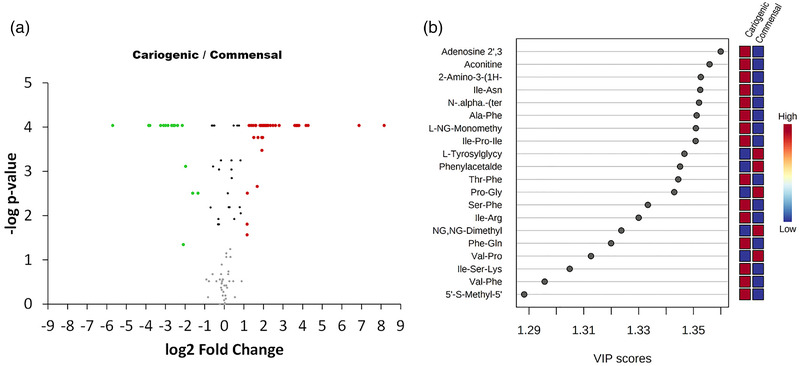
Representation of: (a) volcano plot (FC > 2; FDR < 0.05) of biofilm sample comparison (cariogenic/commensal); (b) representation of the 20 metabolites with the highest VIP
We constructed a heatmap to visualize all of the identified metabolites and the groupings of the CFS samples (Figure 5c). The univariate nonparametric Mann–Whitney analysis showed 59 metabolites that were significantly different in terms of relative abundance between the samples (cariogenic/commensal rate of change > 2 and FDR < 0.05). Figure 5 shows the volcano plot (compounds with fold changes > 2 and FDR < 0.05). Forty‐two metabolites were significantly higher in abundance in the CFSs from the cariogenic biofilms, whereas 17 metabolites were associated significantly with the CFSs from the commensal biofilms.
The metabolites with significantly elevated concentrations in the CFS cariogenic biofilm group were mostly peptides (64.3%), specifically dipeptides (54.8%) and tripeptides (9.5%), likely incomplete breakdown products of protein catabolism. Several amino acids were also abundant (14.3%), represented by histidine, arginine, l‐methionine, glutamic acid, and phenylalanine derivatives. Other secondary metabolites were also significantly present (with percentages < 5%), such as c‐di‐AMP (involved in the formation of bacterial biofilms), nucleosides (such as 5′methylthioadenosine and purine nucleosides), and the amine spermidine.
The metabolites found at significantly higher levels in the CFS commensal group included certain peptides (41.2%), purines (such as hypoxanthine, methyl‐adenine, and methyl‐threonine), and the nucleoside cytidine. Amino acids with significant abundance included d‐ornithine and dimethylarginine. Although the dipeptides in the cariogenic biofilm group were varied and included most amino acids except for proline, all dipeptides that were significantly abundant in the commensal biofilm group presented proline in their composition.
4. DISCUSSION
The present in vitro study evaluated and compared the bacterial composition and metabolic phenotype of a commensal and a dysbiotic oral biofilm model, the latter being compatible with a caries condition. The biofilms, which were derived from the saliva of healthy individuals, demonstrated that the bacterial community responds to imbalances in sucrose levels. The results correlate with those described for in vivo human oral microbiomes, with a consistent direction in the bacterial change, that is, from more to less abundant taxa when progressing from a healthy state to a disease state (Aas et al., 2008; Bradshaw & Lynch, 2013; Eriksson et al., 2018; Gross et al., 2010; Johansson et al., 2016; Lif Holgerson et al., 2015; Mason et al., 2018; Munson et al., 2004; Tanner et al., 2011; Wake et al., 2016; Ximenes et al., 2018; Zaura et al., 2009). The taxonomic analysis of both bacterial communities revealed known biomarkers for in vivo commensal and cariogenic states. Although the overall heterogeneity did not differ substantially between the two conditions, several OTUs were found to have significantly different relative abundances (Aas et al., 2008; Bradshaw & Lynch, 2013; Eriksson et al., 2018; Gross et al., 2010; Johansson et al., 2016; Lif Holgerson et al., 2015; Mason et al., 2018; Munson et al., 2004; Tanner et al., 2011; Wake et al., 2016; Ximenes et al., 2018; Zaura et al., 2009). As a consequence of the shift in microbiota composition, there were significant metabolomic changes when comparing the metabolites secreted to the environment by the commensal and cariogenic biofilms. The greatest differences appear to be related to protein metabolism, with peptides and amino acids as protagonists (Washio et al., 2016).
Mimicking in vitro the biofilm lifestyle of bacteria and biofilm‐related infections is difficult (Buskermolen et al., 2018; Exterkate et al., 2010; Filoche et al., 2007; Guggenheim et al., 2001; Klug et al., 2016; Lebeaux et al., 2013; Rudney et al., 2012; Sissons et al., 2007; Yu et al., 2017). Models used to replicate supragingival biofilms range from in vitro models, such as the Amsterdam Active Attachment model described by Exterkate et al. (2010), to complex in vivo systems, such as the splint system used by Anderson et al. (2018). The developed model, validated by two omics approximations, was representative of the complex oral microbiome in healthy and caries‐related conditions. We employed a closed microcosm system, mimicking in situ conditions that include more environmental parameters and taking into account the complexity and heterogeneity of natural settings through the use of material from the study environment (i.e., saliva), in contrast to the use of a defined consortium of bacteria. The advantages of this model have been discussed and reflected in numerous studies for numerous and varied applications, not only to describe the dynamics of biofilm evolution but also to study the effects of novel antiseptics, probiotics, prebiotics, dental material, and the influence of nutritional conditions (Buskermolen et al., 2018; Filoche et al., 2007; Janus et al., 2016; Klug et al., 2016; Pratten et al., 2000, 1998; Rudney et al., 2012; Sissons et al., 2007). According to a number of authors (Edlund et al., 2013), this closed model developed in culture plates allows for the simultaneous analysis of a large number of samples, with simple and reproducible handling under rigorously controlled environmental settings, unlike other complex systems such as artificial mouth systems, which are tedious to manage (Lebeaux et al., 2013; Pratten et al., 2000; Rudney et al., 2012; Sissons et al., 2007; Yu et al., 2017).
To induce the ecological dysbiosis of the dental plaque model in vitro, we selected sucrose, which has been attributed a central role in causing caries (Bradshaw & Lynch, 2013). Sucrose has been used in other in vivo and in vitro studies to induce changes in bacterial communities (Buskermolen et al., 2018; Filoche et al., 2007; Janus et al., 2016; Pratten et al., 2000; Sissons et al., 2007). According to the extended ecological plaque hypothesis, changes triggered by sucrose availability in the model's local environment stimulate certain members of the commensal bacteria in the oral biofilm model (Marsh, 2003; Rosier et al., 2014; Takahashi & Nyvad, 2008). There were no statistically significant differences in alpha diversity or richness when comparing the cariogenic and commensal biofilms, whereas the beta‐diversity analysis revealed that individual OTUs demonstrated statistically significant differential relative abundance when comparing the two groups, as occurs in natural teeth (Gross et al., 2010; Li et al., 2007; Peterson et al., 2013). This finding suggests that the acidic environment resulting from bacterial metabolism probably acts as the main selective force in microorganisms able to proliferate in this environment, confirming previous studies (Gross et al., 2012; Simon‐Soro et al., 2013).
The taxonomic assignment of the 16S rRNA gene reads identified Firmicutes as the major phylum in both biofilm conditions, although with different relative abundance in each case (64.80% for the commensal condition and 96.47% for the cariogenic). Firmicutes was followed by the phyla Actinobacteria (34.22% and 1.60%, respectively), Proteobacteria (0.56% and 0.60%), Bacteroidetes (0.18% and 0.69%), and Fusobacter (0.07% and 0.42%). These results are in line with previous reports that focused on comparing the initial oral plaque of young adults with and without carious lesions, using an in situ biofilm model (Rupf et al., 2018). The results also agree with the shift in microbiota in oral plaque in response to frequent sucrose consumption reported by Anderson et al. (2018) in an in vivo split system. We detected a significant increase in the relative abundance of Firmicutes (p = 0.019) and Fusobacter (p < 0.001), both more abundant in the cariogenic biofilms, compared with Actinobacteria (p < 0.001), which decreased significantly in the disease biofilms, results in line with those reported by Gross et al. (2012).
At the genus taxonomic level, the in vitro commensal biofilm model developed a greater abundance of Streptococcus, Staphylococcus, Rothia, Gemella, and Granulicatella (Table 1), known biomarkers for in vivo healthy supragingival plaque (Gross et al., 2012; Kumar et al., 2005). Nowadays, there remains considerable controversy as to whether Staphylococcus spp. play a role in the ecology of the normal oral microbiota or the oral cavity may represent a yet poorly recognized reservoir of staphylococci, some of which may, under appropriate conditions, cause local or systemic infection (Smith et al., 2001). There are several reports of S. aureus isolated from the healthy oral cavity and it intra‐oral distribution in saliva, plaque, both supragingival and subgingival, and in fissures of teeth (Ohara‐Nemoto et al., 2008; Percival et al., 1991; Smith et al., 2003; Theilade et al., 1982). These studies described oral carriage rates of 83%–94% in saliva of healthy adults, with similar isolation tendency observed in supragingival plaque, with a total occurrence rate around 73.2%. Therefore, its presence in the commensal biofilm is not surprising, although we cannot compare its relative abundance (24.46%; Table 1) since most reports are qualitative and do not provide abundance data with respect to the total bacterial population detected in the samples. Other major in vivo genera, such as Lactobacillus, Actinomyces, and Haemophilus, were abundantly present in the in vitro biofilm model (Kumar et al., 2005) (Table 1). In regard to the in vitro systems and compared with previous in vitro commensal biofilm models, the present findings correlate, among others, to the phenotypical composition observed by Janus et al. (2015) and with that described by Wake et al. (2016), who employed an in situ model to conduct a quantitative analysis and comprehensive identification of bacterial communities involved in the health of the human oral cavity. When commensal biofilm progresses to a cariogenic condition by adding sucrose as a disease trigger, some of the genera (mostly those related to oral disease) significantly increase in proportion, as is the case with the genera Streptococcus, Fusobacterium, and Prevotella, while other genera more related to oral health decrease in presence, such as Rothia and Granulicatella. The results of an in vivo study by Peterson et al. (2013) on dental plaque microbiomes in healthy and disease conditions support our results. The authors found that approximately 70% of the phyla described in dental plaque were present in their samples, with only four of these phyla having an abundance > 1% of the total. This in vitro dysbiosis reflects the in vivo situation, as described by Anderson et al. (2018) and Rupf et al. (2018), who applied in vivo biofilm models. The increase in the proportion of Prevotella, a proteolytic bacterium that has been described as saccharolytic and tolerant to the acidic environment, could also result in an acidogenic situation and actively affect the ecological dysbiosis (Simon‐Soro et al., 2013).
The study has also shown how the microbial community assembly significantly changed at the species level after sucrose treatments. The most notable results were the significant increase in the genus Streptococcus and the general non‐mutans streptococci. In particular, S. parasanguinis and S. anginosus increased significantly in the cariogenic biofilm. Studies have established that cariogenic biofilms often contain high levels of S. mutans, S. sobrinus, and various Lactobacillus species (Takahashi & Nyvad, 2008). Nevertheless, various other bacterial species have been shown to be associated with differing stages of caries and are thereby linked to the initiation of caries, involving non‐mutans streptococci such as S. salivarius and S. parasanguinis, as shown by the present cariogenic model (Aas et al., 2008; Gross et al., 2012; van Houte et al., 1996). Becker et al. (2002) found that the presence of S. parasanguinis was increased in progressed carious lesions in patients with early childhood caries. Gross et al. (2012) reported a similar significant association for S. parasanguinis in their 16S rDNA gene cloning study in children with early childhood caries. Aas et al. (2008) frequently detected S. parasanguinis in primary and secondary carious teeth, especially when S. mutans was not present.
The biofilm dysbiosis observed by the metataxonomic analysis, as evidenced by the patterns that emerged from the PCoA analysis with two clearly differentiated clusters (Figure 2), was also reflected in the metabolites secreted into the environment by the bacterial communities, with samples from the cariogenic group very close together but separate from the commensal group, which were more dispersed among them (Figure 3). To test the hypothesis that the functional profile of the cariogenic biofilm differs from that of the commensal biofilm and to determine the action of bacteria and their response to environmental fluctuations and metabolized determinate substrates when grown in biofilms (both in vivo and in vitro), we employed a second approach that consisted of a metabolomics analysis of the CFS based on UHPLC/Q‐TOF‐MS
The metabolomics analysis revealed how the bacterial consortium that constitutes the supragingival plaque was able to metabolize a wide range of substrates, including carbohydrates and proteins. The approach provides a real‐time view of the dynamic changes in the biofilm microbiome's derived metabolites. Contrary to our expectations, the degradation of carbohydrates (mainly through glycolysis and subsequent organic acids) showed no significant differences in relative abundance in the CFS of commensal biofilms compared with cariogenic biofilms. Conversely, protein metabolism was significantly affected in the bacterial biofilms, as reflected by the metabolites detected in the CFS of both biofilm phenotypes.
The metabolomic approach revealed that the VIP score of the metabolites in the CFSs for both biofilm phenotypes was dominated by peptides (Figure 6), representing 64.3% of the metabolites, with significant relative abundance in the cariogenic biofilms, and 41.2% for the commensal biofilms (Figure 5). Dipeptides predominated in both phenotypes. Certain dipeptides are known to have physiological or cell‐signaling effects, although most are simply short‐lived intermediates on their way to specific amino acid degradation pathways following further proteolysis. It is worth noting that in the CFS of the commensal biofilms, 100% of the dipeptides with significant relative abundance contained proline in their structure, compared with 9.7% in the CFS of the cariogenic biofilms.
Proline has recently been shown to play a role in pathogen and host interactions inside and outside the oral cavity by modulating cell signaling and osmotic stress and acting as an antibacterial molecule or prebiotic (Christgen & Becker, 2019; Cleaver et al., 2019, 2020). Slomka et al. (2017) investigated a proline‐containing prebiotic candidate based on methionine–proline, which after just three days of exposure shifted the composition of an oral biofilm model by reducing the pathogenic species to a predominantly beneficial species (Slomka et al., 2017, 2018). Drobni et al. (2006) verified that a proline peptide caused greater resistance in biofilms to sucrose‐induced decreases in pH (Drobni et al., 2006). Curiously, the tripeptide Ile‐Pro‐Ile (described as diprotin A, an inhibitor of the enzyme dipeptidyl peptidase IV, an enzyme that preferably hydrolyzes proteins with a penultimate proline residue) was significantly more abundant in the cariogenic CFS, which might explain the lower quantity of oligopeptides with proline in the cariogenic biofilms (Umezawa et al., 1984).
The study also revealed that the cariogenic CFSs showed a high abundance of the amino acids histidine, methionine, glutamic acid, and phenylalanine, results similar to those reported by several other metabolomics studies (Takahashi, 2015; Washio et al., 2016), while the commensal CFSs presented a significant abundance of d‐ornithine. Although the metabolic regulation of amino acid degradation has not yet been clearly elucidated, evidence supports the hypothesis that amino acids can be degraded by dental plaque bacteria to, among other substances, ammonia, organic acids, and amines, which are associated with acid neutralization, oral malodor, and tissue inflammation (Takahashi, 2015; Washio et al., 2016). It has been suggested that the arginine deiminase system is one of the main metabolic pathways used by several bacteria (such as diverse species of Streptococcus) to combat tooth demineralization by neutralizing acids in the dental plaque where arginine is deiminated via citrulline with the production of ammonia, ornithine, carbon oxides, and ATP (Takahashi, 2015; Washio et al., 2016). While l‐NG‐monomethyl arginine is significantly more abundant in cariogenic CFSs, NG, NG‐dimethyl‐L‐arginine, and d‐ornithine are present in significant quantities in commensal CFSs, indicating that commensal biofilms are significantly more active in neutralizing the environment's pH (Nascimento et al., 2017). In contrast, the polyamine spermidine was significantly more abundant in the cariogenic group, as was also observed by Washio et al. (2016), biosynthesized as the result of the decarboxylation of ornithine in putrescine and subsequent spermidine. The investigation of this polyamine function in bacteria has revealed that they are involved in bacterial growth and acid resistance and are essential in biofilm formation in certain bacterial species (Washio et al., 2016).
It has been noted that branched chain amino acids such as valine, leucine, and isoleucine are significantly abundant in the progressed cariogenic environment. These branched chain amino acids can be converted into isovalerate and isobutyrate, through sequential reactions of deamination, decarboxylation, and reduction, with the production of acetyl‐CoA and, subsequently, acetate, with relative abundance in the CFS of cariogenic biofilms (Takahashi, 2015). These pathways have been described for (among other bacteria) Prevotella (Takahashi, 2015). Although in relatively low proportions, Prevotella at genera level appeared among the 10 most abundant in both types of biofilm (Table 1), increasing its presence from 0.08% in commensal biofilms to 0.33% in cariogenic ones. This increase, although insignificant (p > 0.05), was largely due to the significant increase in the species Prevotella maleninogenica (p = 0.020; Table 2). Although discrete with respect to the increment experienced by other species in the cariogenic biofilm (Table 2) and which hardly seems a level of abundance that would mark large‐scale differences in the main concentrations of common metabolites, this increment could be relevant in the breakdown of biofilm homeostasis with drift toward a situation compatible with periodontal diseases, and is that among other capacities, P. maleninogenica has the ability to produce hemolysin, a compound responsible for the lysis of erythrocytes through the production of pores in the cytoplasmic membrane, thus releasing hemoglobin to the niche where it inhabits (Allison & Hillman, 1997). Hemoglobin may be used by different periodontal pathogenic species that require hemin as a source of iron and protoporphyrin IX to survive and initiate periodontal infection. Continuing in the line of the metabolism of amino acids, a significant amount of propionic acid [2‐amino‐3‐(1H‐pyrazole‐1‐yl) propanoic acid], which is secreted by certain bacterial species and has significant fungal and bacterial inhibitor potential, was detected in the cariogenic CFS.
The cyclic dimeric adenosine 3′,5′‐monophosphate (c‐di‐AMP), a recently identified secondary messenger in bacteria, was also significantly more abundant in the cariogenic biofilm (Konno et al., 2018). It has been reported that Gram‐positive bacteria mostly synthesize c‐di‐AMP, which appears to be essential for the viability of the bacterial cells that produce it, controlling important signaling cascades in bacteria, including the homeostasis of cell wall peptidoglycan architecture, size and envelope cell stress, drug resistance and biofilm formation, among others (Corrigan et al., 2011; Du et al., 2014; Konno et al., 2018; Luo & Helmann, 2012).
The nucleoside 5′‐methyl‐thioadenosine, also known as MTA or methylthioadenosine, was significantly abundant in the CFS of progressed cariogenic biofilms. MTA and spermidine (amines that were also significantly abundant in the cariogenic condition) can be biosynthesized from S‐adenosylmethioninamine and putrescine, respectively, through the action of the enzyme spermidine synthase. MTA is metabolized solely by MTA‐phosphorylase to yield 5‐methyl thioribose‐1‐phosphate and adenine, a crucial step in the methionine and purine salvage pathways, respectively. Both methionine and purine were increased in the CFS of the cariogenic biofilms. Within bacteria, MTA is involved in the quorum sensing circuit, which is highly relevant in controlling bacterial population density (Z. Wang et al., 2014).
In terms of nucleic acid synthesis, the commensal CFS presented a significant abundance of nitrogenous bases (N6‐methyladenine and 2‐hydroxy‐6‐aminopurine), the nucleotide cytidine, and hypoxanthine (a purine derivate), while the cariogenic CFS had a greater abundance of the nucleoside inosine, an intermediate of nucleic acid synthesis pathways that is formed when hypoxanthine binds to a ribose ring (also known as ribofuranose) through a β‐N9 glycosidic bond (Z. Wang et al., 2014). Hypoxanthine production has been reported in certain bacterial species, such as Escherichia coli (Z. Wang et al., 2014), which indicates a highly efficient activation of the biosynthesis of nucleic acid synthesis in cariogenic biofilms.
5. CONCLUSION
This study developed a complex and dynamic supragingival biofilm model comprising a representative bacterial diversity of the resident microbiome derived from saliva, responsible for plaque formation in the human oral cavity. In line with previous authors, the microcosm model was developed using inoculums from numerous donors to solve differences in species composition between participants. The taxonomic and metabolic potential of the model was addressed, responding with changes in taxa due to fluctuations in environmental conditions and facilitating its discovery and functional characterization through the metabolic pathways that define the transition from a healthy state to a disease state in the complex oral microbiome. Integration of microbial and metabolomics data paired with experimental validation are essential for understanding the links between microbial communities and disease. The study observations corroborate that exogenous environmental factors play a role in modulating not only the composition but also the activities of oral bacteria and factors such as diet with the intake of various macronutrients such as sucrose. The results reveal that we should focus on protein catabolism and, among other derived metabolites, on the amino acid proline, which could play a relevant role as a prebiotic in oral health. However, the results have to be interpreted with caution, given the study's limitations. We developed an in vitro model that was not subjected to all environmental conditions that occur in the oral cavity. The microcosms we obtained did not completely reproduce the microbiota of either plaque or saliva charge. There could be many reasons for this situation, such as incubation under aerobic conditions with 5% CO2, given that our goal was to simulate the ecological evolution of supragingival plaque. A number of anaerobes were therefore likely retained in the microcosms. The growth medium we employed (BHI) might also have caused the loss of certain species because it failed to fully satisfy their nutritional needs. Furthermore, the planktonic community present in the supernatant could have contributed to the metabolome that was measured, so some of the changes observed could be due to taxonomic changes produced in the supernatant, which were not examined here.
FUNDING INFORMATION
No funding was received for the research.
CONFLICT OF INTEREST
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
AUTHOR CONTRIBUTIONS
MCS. contributed to the design of the study, undertook experiments, analysed the data, and wrote the manuscript. AV., ALl‐P. and AV contributed to the design of the research, undertook experiments and analysed the data. AC, MJC and LC contributed to the design of the research, analysis of the data, supervision of the project, and the final draft of the manuscript. All authors contributed to the article and approved the submitted version.
PEER REVIEW
The peer review history for this article is available at https://publons.com/publon/10.1111/omi.12363.
Supporting information
Supporting information
Figure s1
{kind=link}
ACKNOWLEDGMENTS
DNA sequencing and analysis were performed at Microomics Systems S.L. (Barcelona, Spain). Metabolomic analysis were performed at the Metabolomic Platform of CIAL Institute, CSIC, Madrid. The authors would like to acknowledge the support and expertise of all of them. The authors also would like to acknowledge to the Microbiology teaching unit of the Department of Medicine of the UCM, and in particular Professor María Luisa Gómez‐Lus, for providing their facilities.
Sánchez, M. C. , Velapatiño, A. , Llama‐Palacios, A. , Valdés, A. , Cifuentes, A. , Ciudad, M. J. , & Collado, L. (2022). Metataxonomic and metabolomic evidence of biofilm homeostasis disruption related to caries: An in vitro study. Molecular Oral Microbiology, 37, 81–96. 10.1111/omi.12363
DATA AVAILABILITY STATEMENT
Metabolomics data have been deposited to the EMBL‐EBI MetaboLights database (DOI: 10.1093/nar/gkz1019, PMID:31691833) with the identifier MTBLS3982. The complete dataset could be accessed at “https://www.ebi.ac.uk/metabolights/MTBLS3982”. The metagenome data have been deposited in the SRA with accession number: PRJNA792086. The complete dataset could be accessed at https://www.ncbi.nlm.nih.gov/sra/PRJNA792086.
REFERENCES
- Aas, J. A. , Griffen, A. L. , Dardis, S. R. , Lee, A. M. , Olsen, I. , Dewhirst, F. E. , Leys, E. J. , & Paster, B. J. (2008). Bacteria of dental caries in primary and permanent teeth in children and young adults. Journal of Clinical Microbiology, 46(4), 1407–1417. 10.1128/JCM.01410-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allison, H. E. , & Hillman, J. D. (1997). Cloning and characterization of a Prevotella melaninogenica hemolysin. Infection and Immunity, 65(7), 2765–2771. 10.1128/iai.65.7.2765-2771.1997 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson, A. C. , Rothballer, M. , Altenburger, M. J. , Woelber, J. P. , Karygianni, L. , Lagkouvardos, I. , Hellwig, E. , & Al‐Ahmad, A. (2018). In‐vivo shift of the microbiota in oral biofilm in response to frequent sucrose consumption. Science Reports, 8(1), 14202. 10.1038/s41598-018-32544-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson, M. , & Walsh, D. (2013). PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? Ecological Monographs, 83, 557–574. 10.1890/12-2010.1 [DOI] [Google Scholar]
- Becker, M. R. , Paster, B. J. , Leys, E. J. , Moeschberger, M. L. , Kenyon, S. G. , Galvin, J. L. , Boches, S. K. , Dewhirst, F. E. , & Griffen, A. L. (2002). Molecular analysis of bacterial species associated with childhood caries. Journal of Clinical Microbiology, 40(3), 1001–1009. 10.1128/JCM.40.3.1001-1009.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradshaw, D. J. , & Lynch, R. J. (2013). Diet and the microbial aetiology of dental caries: New paradigms. International Dental Journal, 63, 39‐47. 10.1111/idj.12082 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buskermolen, J. K. , Janus, M. M. , Roffel, S. , Krom, B. P. , & Gibbs, S. (2018). Saliva‐derived commensal and pathogenic biofilms in a human gingiva model. Journal of Dental Research, 97(2), 201–208. 10.1177/0022034517729998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callahan, B. J. , McMurdie, P. J. , Rosen, M. J. , Han, A. W. , Johnson, A. J. , & Holmes, S. P. (2016). DADA2: High‐resolution sample inference from Illumina amplicon data. Nature Methods, 13(7), 581–583. 10.1038/nmeth.3869 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christgen, S. L. , & Becker, D. F. (2019). Role of proline in pathogen and host interactions. Antioxidants & Redox Signaling, 30(4), 683–709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cleaver, L. M. , Moazzez, R. , & Carpenter, G. H. (2019). Mixed aerobic‐anaerobic incubation conditions induce proteolytic activity from in vitro salivary biofilms. Journal of Oral Microbiology, 11(1), 1643206. 10.1080/20002297.2019.1643206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cleaver, L. M. , Moazzez, R. V. , & Carpenter, G. H. (2020). Evidence for proline utilization by oral bacterial biofilms grown in saliva. Frontiers in Microbiology, 11, 619968. 10.3389/fmicb.2020.619968 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corrigan, R. M. , Abbott, J. C. , Burhenne, H. , Kaever, V. , & Grundling, A. (2011). c‐di‐AMP is a new second messenger in Staphylococcus aureus with a role in controlling cell size and envelope stress. PLoS Pathogens, 7(9), e1002217. 10.1371/journal.ppat.1002217 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Costerton, J. W. , Lewandowski, Z. , Caldwell, D. E. , Korber, D. R. , & Lappin‐Scott, H. M. (1995). Microbial biofilms. Annual Review of Microbiology, 49, 711–745. 10.1146/annurev.mi.49.100195.003431 [DOI] [PubMed] [Google Scholar]
- Cuenca, M. , Sanchez, M. C. , Diz, P. , Martinez‐Lamas, L. , Alvarez, M. , Limeres, J. , Sanz, M. , & Herrera, D. (2021). In vitro anti‐biofilm and antibacterial properties of Streptococcus downii sp. nov. Microorganisms, 9(2), 450. 10.3390/microorganisms9020450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drobni, M. , Li, T. , Kruger, C. , Loimaranta, V. , Kilian, M. , Hammarstrom, L. , Jörnvall, H. , Bergman, T. , & Strömberg, N. (2006). Host‐derived pentapeptide affecting adhesion, proliferation, and local pH in biofilm communities composed of Streptococcus and Actinomyces species. Infection and Immunity, 74(11), 6293–6299. 10.1128/IAI.00068-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du, B. , Ji, W. , An, H. , Shi, Y. , Huang, Q. , Cheng, Y. , Fu, Q. , Wang, H. , Yan, Y. , & Sun, J. (2014). Functional analysis of c‐di‐AMP phosphodiesterase, GdpP, in Streptococcus suis serotype 2. Microbiological Research, 169(9‐10), 749–758. 10.1016/j.micres.2014.01.002 [DOI] [PubMed] [Google Scholar]
- Edlund, A. , Yang, Y. , Hall, A. P. , Guo, L. , Lux, R. , He, X. , Nelson, K. E. , Nealson, K. H. , Yooseph, S. , Shi, W. , & McLean, J. S. (2013). An in vitro biofilm model system maintaining a highly reproducible species and metabolic diversity approaching that of the human oral microbiome. Microbiome, 1(1), 25. 10.1186/2049-2618-1-25 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eriksson, L. , Lif Holgerson, P. , Esberg, A. , & Johansson, I. (2018). Microbial complexes and caries in 17‐year‐olds with and without Streptococcus mutans . Journal of Dental Research, 97(3), 275–282. 10.1177/0022034517731758 [DOI] [PubMed] [Google Scholar]
- Exterkate, R. A. , Crielaard, W. , & Ten Cate, J. M. (2010). Different response to amine fluoride by Streptococcus mutans and polymicrobial biofilms in a novel high‐throughput active attachment model. Caries Research, 44(4), 372–379. 10.1159/000316541 [DOI] [PubMed] [Google Scholar]
- Featherstone, J. D. (2004). The continuum of dental caries–evidence for a dynamic disease process. Journal of Dental Research, 83, C39‐42. 10.1177/154405910408301s08 [DOI] [PubMed] [Google Scholar]
- Fejerskov, O. (2004). Changing paradigms in concepts on dental caries: Consequences for oral health care. Caries Research, 38(3), 182–191. 10.1159/000077753 [DOI] [PubMed] [Google Scholar]
- Filoche, S. K. , Soma, K. J. , & Sissons, C. H. (2007). Caries‐related plaque microcosm biofilms developed in microplates. Oral Microbiology and Immunology, 22(2), 73–79. 10.1111/j.1399-302X.2007.00323.x [DOI] [PubMed] [Google Scholar]
- Gross, E. L. , Beall, C. J. , Kutsch, S. R. , Firestone, N. D. , Leys, E. J. , & Griffen, A. L. (2012). Beyond Streptococcus mutans: Dental caries onset linked to multiple species by 16S rRNA community analysis. PLoS One, 7(10), e47722. 10.1371/journal.pone.0047722 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gross, E. L. , Leys, E. J. , Gasparovich, S. R. , Firestone, N. D. , Schwartzbaum, J. A. , Janies, D. A. , Asnani, K. , & Griffen, A. L. (2010). Bacterial 16S sequence analysis of severe caries in young permanent teeth. Journal of Clinical Microbiology, 48(11), 4121–4128. 10.1128/JCM.01232-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guggenheim, B. , Giertsen, E. , Schupbach, P. , & Shapiro, S. (2001). Validation of an in vitro biofilm model of supragingival plaque. Journal of Dental Research, 80(1), 363–370. 10.1177/00220345010800011201 [DOI] [PubMed] [Google Scholar]
- Hung, C. , Zhou, Y. , Pinkner, J. S. , Dodson, K. W. , Crowley, J. R. , Heuser, J. , Chapman, M. R. , Hadjifrangiskou, M. , Henderson, J. P. , & Hultgren, S. J. (2013). Escherichia coli biofilms have an organized and complex extracellular matrix structure. mBio, 4(5), e00645‐00613. 10.1128/mBio.00645-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janus, M. M. , Crielaard, W. , Zaura, E. , Keijser, B. J. , Brandt, B. W. , & Krom, B. P. (2016). A novel compound to maintain a healthy oral plaque ecology in vitro. Journal of Oral Microbiology, 8, 32513. 10.3402/jom.v8.32513 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janus, M. M. , Keijser, B. J. , Bikker, F. J. , Exterkate, R. A. , Crielaard, W. , & Krom, B. P. (2015). In vitro phenotypic differentiation towards commensal and pathogenic oral biofilms. Biofouling, 31(6), 503–510. 10.1080/08927014.2015.1067887 [DOI] [PubMed] [Google Scholar]
- Johansson, I. , Witkowska, E. , Kaveh, B. , Lif Holgerson, P. , & Tanner, A. C. (2016). The microbiome in populations with a low and high prevalence of caries. Journal of Dental Research, 95(1), 80–86. 10.1177/0022034515609554 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katoh, K. , & Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Molecular biology and evolution, 30(4), 772–780. 10.1093/molbev/mst010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kidd, E. A. , & Fejerskov, O. (2004). What constitutes dental caries? Histopathology of carious enamel and dentin related to the action of cariogenic biofilms. Journal of Dental Research, 83, C35‐38. 10.1177/154405910408301s07 [DOI] [PubMed] [Google Scholar]
- Kleinberg, I. (2002). A mixed‐bacteria ecological approach to understanding the role of the oral bacteria in dental caries causation: An alternative to Streptococcus mutans and the specific‐plaque hypothesis. Critical Reviews in Oral Biology & Medicine, 13(2), 108–125. 10.1177/154411130201300202 [DOI] [PubMed] [Google Scholar]
- Klug, B. , Santigli, E. , Westendorf, C. , Tangl, S. , Wimmer, G. , & Grube, M. (2016). From mouth to model: Combining in vivo and in vitro oral biofilm growth. Frontiers in Microbiology, 7, 1448. 10.3389/fmicb.2016.01448 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Konno, H. , Yoshida, Y. , Nagano, K. , Takebe, J. , & Hasegawa, Y. (2018). Biological and biochemical roles of two distinct cyclic dimeric adenosine 3',5'‐monophosphate‐associated phosphodiesterases in Streptococcus mutans . Frontiers in Microbiology, 9, 2347. 10.3389/fmicb.2018.02347 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar, P. S. , Griffen, A. L. , Moeschberger, M. L. , & Leys, E. J. (2005). Identification of candidate periodontal pathogens and beneficial species by quantitative 16S clonal analysis. Journal of Clinical Microbiology, 43(8), 3944–3955. 10.1128/JCM.43.8.3944-3955.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lebeaux, D. , Chauhan, A. , Rendueles, O. , & Beloin, C. (2013). From in vitro to in vivo models of bacterial biofilm‐related infections. Pathogens, 2(2), 288–356. 10.3390/pathogens2020288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, Y. , Ge, Y. , Saxena, D. , & Caufield, P. W. (2007). Genetic profiling of the oral microbiota associated with severe early‐childhood caries. Journal of Clinical Microbiology, 45(1), 81–87. 10.1128/JCM.01622-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lif Holgerson, P. , Ohman, C. , Ronnlund, A. , & Johansson, I. (2015). Maturation of oral microbiota in children with or without dental caries. PLoS One, 10(5), e0128534. 10.1371/journal.pone.0128534 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo, Y. , & Helmann, J. D. (2012). Analysis of the role of Bacillus subtilis sigma(M) in beta‐lactam resistance reveals an essential role for c‐di‐AMP in peptidoglycan homeostasis. Molecular Microbiology, 83(3), 623–639. 10.1111/j.1365-2958.2011.07953.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mandal, S. , Van Treuren, W. , White, R. A. , Eggesbo, M. , Knight, R. , & Peddada, S. D. (2015). Analysis of composition of microbiomes: A novel method for studying microbial composition. Microbial Ecology in Health and Disease, 26, 27663. 10.3402/mehd.v26.27663 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marsh, P. D. (2003). Are dental diseases examples of ecological catastrophes? Microbiology (Reading, England), 149(Pt 2), 279–294. 10.1099/mic.0.26082-0 [DOI] [PubMed] [Google Scholar]
- Marsh, P. D. (2006). Dental plaque as a biofilm and a microbial community ‐ implications for health and disease. BMC Oral Health, 6, S14. 10.1186/1472-6831-6-S1-S14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marsh, P. D. , Moter, A. , & Devine, D. A. (2011). Dental plaque biofilms: Communities, conflict and control. Periodontology 2000, 55(1), 16–35. 10.1111/j.1600-0757.2009.00339.x [DOI] [PubMed] [Google Scholar]
- Mason, M. R. , Chambers, S. , Dabdoub, S. M. , Thikkurissy, S. , & Kumar, P. S. (2018). Characterizing oral microbial communities across dentition states and colonization niches. Microbiome, 6(1), 67. 10.1186/s40168-018-0443-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mira, A. , Simon‐Soro, A. , & Curtis, M. A. (2017). Role of microbial communities in the pathogenesis of periodontal diseases and caries. Journal of Clinical Periodontology, 44, S23‐S38. 10.1111/jcpe.12671 [DOI] [PubMed] [Google Scholar]
- Munson, M. A. , Banerjee, A. , Watson, T. F. , & Wade, W. G. (2004). Molecular analysis of the microflora associated with dental caries. Journal of Clinical Microbiology, 42(7), 3023–3029. 10.1128/JCM.42.7.3023-3029.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nascimento, M. M. , Zaura, E. , Mira, A. , Takahashi, N. , & Ten Cate, J. M. (2017). Second era of OMICS in caries research: Moving past the phase of disillusionment. Journal of Dental Research, 96(7), 733–740. 10.1177/0022034517701902 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohara‐Nemoto, Y. , Haraga, H. , Kimura, S. , & Nemoto, T. K. (2008). Occurrence of staphylococci in the oral cavities of healthy adults and nasal oral trafficking of the bacteria. Journal of Medical Microbiology, 57(Pt 1), 95–99. 10.1099/jmm.0.47561-0 [DOI] [PubMed] [Google Scholar]
- Percival, R. S. , Challacombe, S. J. , & Marsh, P. D. (1991). Age‐related microbiological changes in the salivary and plaque microflora of healthy adults. Journal of Medical Microbiology, 35(1), 5–11. 10.1099/00222615-35-1-5 [DOI] [PubMed] [Google Scholar]
- Peterson, S. N. , Snesrud, E. , Liu, J. , Ong, A. C. , Kilian, M. , Schork, N. J. , & Bretz, W. (2013). The dental plaque microbiome in health and disease. PLoS One, 8(3), e58487. 10.1371/journal.pone.0058487 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pratten, J. , Andrews, C. S. , Craig, D. Q. , & Wilson, M. (2000). Structural studies of microcosm dental plaques grown under different nutritional conditions. FEMS Microbiology Letters, 189(2), 215–218. 10.1111/j.1574-6968.2000.tb09233.x [DOI] [PubMed] [Google Scholar]
- Pratten, J. , Smith, A. W. , & Wilson, M. (1998). Response of single species biofilms and microcosm dental plaques to pulsing with chlorhexidine. Journal of Antimicrobial Chemotherapy, 42(4), 453–459. 10.1093/jac/42.4.453 [DOI] [PubMed] [Google Scholar]
- Price, M. N. , Dehal, P. S. , & Arkin, A. P. (2009). FastTree: Computing large minimum evolution trees with profiles instead of a distance matrix. Molecular Biology and Evolution, 26(7), 1641–1650. 10.1093/molbev/msp077 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosier, B. T. , De Jager, M. , Zaura, E. , & Krom, B. P. (2014). Historical and contemporary hypotheses on the development of oral diseases: Are we there yet? Frontiers in Cellular and Infection Microbiology, 4, 92. 10.3389/fcimb.2014.00092 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rudney, J. D. , Chen, R. , Lenton, P. , Li, J. , Li, Y. , Jones, R. S. , Reilly, C. , Fok, A. S. , & Aparicio, C. (2012). A reproducible oral microcosm biofilm model for testing dental materials. Journal of Applied Microbiology, 113(6), 1540–1553. 10.1111/j.1365-2672.2012.05439.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rupf, S. , Laczny, C. C. , Galata, V. , Backes, C. , Keller, A. , Umanskaya, N. , Erol, A. , Tierling, S. , Lo Porto, C. , Walter, J. , Kirsch, J. , Hannig, M. , & Hannig, C. (2018). Comparison of initial oral microbiomes of young adults with and without cavitated dentin caries lesions using an in situ biofilm model. Science Reports, 8(1), 14010. 10.1038/s41598-018-32361-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanz, M. , Beighton, D. , Curtis, M. A. , Cury, J. A. , Dige, I. , Dommisch, H. , Ellwood, R. , Giacaman, R. A. , Herrera, D. , Herzberg, M. C. , Könönen, E. , Marsh, P. D. , Meyle, J. , Mira, A. , Molina, A. , Mombelli, A. , Quirynen, M. , Reynolds, E. C. , Shapira, L. , & Zaura, E. (2017). Role of microbial biofilms in the maintenance of oral health and in the development of dental caries and periodontal diseases. Consensus report of group 1 of the Joint EFP/ORCA workshop on the boundaries between caries and periodontal disease. Journal of Clinical Periodontology, 44, S5‐S11. 10.1111/jcpe.12682 [DOI] [PubMed] [Google Scholar]
- Selwitz, R. H. , Ismail, A. I. , & Pitts, N. B. (2007). Dental caries. Lancet, 369(9555), 51–59. 10.1016/S0140-6736(07)60031-2 [DOI] [PubMed] [Google Scholar]
- Simon‐Soro, A. , Belda‐Ferre, P. , Cabrera‐Rubio, R. , Alcaraz, L. D. , & Mira, A. (2013). A tissue‐dependent hypothesis of dental caries. Caries Research, 47(6), 591–600. 10.1159/000351663 [DOI] [PubMed] [Google Scholar]
- Sissons, C. H. , Anderson, S. A. , Wong, L. , Coleman, M. J. , & White, D. C. (2007). Microbiota of plaque microcosm biofilms: Effect of three times daily sucrose pulses in different simulated oral environments. Caries Research, 41(5), 413–422. 10.1159/000104801 [DOI] [PubMed] [Google Scholar]
- Slomka, V. , Hernandez‐Sanabria, E. , Herrero, E. R. , Zaidel, L. , Bernaerts, K. , Boon, N. , Quirynen, M. , & Teughels, W. (2017). Nutritional stimulation of commensal oral bacteria suppresses pathogens: The prebiotic concept. Journal of Clinical Periodontology, 44(4), 344–352. 10.1111/jcpe.12700 [DOI] [PubMed] [Google Scholar]
- Slomka, V. , Herrero, E. R. , Boon, N. , Bernaerts, K. , Trivedi, H. M. , Daep, C. , Quirynen, M. , & Teughels, W. (2018). Oral prebiotics and the influence of environmental conditions in vitro. Journal of Periodontology, 89(6), 708–717. 10.1002/JPER.17-0437 [DOI] [PubMed] [Google Scholar]
- Smith, A. J. , Jackson, M. S. , & Bagg, J. (2001). The ecology of Staphylococcus species in the oral cavity. Journal of Medical Microbiology, 50(11), 940–946. 10.1099/0022-1317-50-11-940 [DOI] [PubMed] [Google Scholar]
- Smith, A. J. , Robertson, D. , Tang, M. K. , Jackson, M. S. , MacKenzie, D. , & Bagg, J. (2003). Staphylococcus aureus in the oral cavity: A three‐year retrospective analysis of clinical laboratory data. British Dental Journal, 195(12), 701–703; discussion 694. 10.1038/sj.bdj.4810832 [DOI] [PubMed] [Google Scholar]
- Takahashi, N. (2015). Oral microbiome metabolism: From “who are they?” to “what are they doing? Journal of Dental Research, 94(12), 1628–1637. 10.1177/0022034515606045 [DOI] [PubMed] [Google Scholar]
- Takahashi, N. , & Nyvad, B. (2008). Caries ecology revisited: Microbial dynamics and the caries process. Caries Research, 42(6), 409–418. 10.1159/000159604 [DOI] [PubMed] [Google Scholar]
- Takahashi, N. , & Nyvad, B. (2011). The role of bacteria in the caries process: Ecological perspectives. Journal of Dental Research, 90(3), 294–303. 10.1177/0022034510379602 [DOI] [PubMed] [Google Scholar]
- Tanner, A. C. , Kent, R. L. , Jr., Holgerson, P. L. , Hughes, C. V. , Loo, C. Y. , Kanasi, E. , Chalmers, N. I. , & Johansson, I. (2011). Microbiota of severe early childhood caries before and after therapy. Journal of Dental Research, 90(11), 1298–1305. 10.1177/0022034511421201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- ten Cate, J. M. (2009). The need for antibacterial approaches to improve caries control. Advances in Dental Research, 21(1), 8–12. 10.1177/0895937409335591 [DOI] [PubMed] [Google Scholar]
- Theilade, E. , Fejerskov, O. , Karring, T. , & Theilade, J. (1982). Predominant cultivable microflora of human dental fissure plaque. Infection and Immunity, 36(3), 977–982. 10.1128/iai.36.3.977-982.1982 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umezawa, H. , Aoyagi, T. , Ogawa, K. , Naganawa, H. , Hamada, M. , & Takeuchi, T. (1984). Diprotins A and B, inhibitors of dipeptidyl aminopeptidase IV, produced by bacteria. The Journal of Antibiotics, 37(4), 422–425. 10.7164/antibiotics.37.422 [DOI] [PubMed] [Google Scholar]
- van Houte, J. , Lopman, J. , & Kent, R. (1996). The final pH of bacteria comprising the predominant flora on sound and carious human root and enamel surfaces. Journal of Dental Research, 75(4), 1008–1014. 10.1177/00220345960750040201 [DOI] [PubMed] [Google Scholar]
- Wake, N. , Asahi, Y. , Noiri, Y. , Hayashi, M. , Motooka, D. , Nakamura, S. , Gotoh, K. , Miura, J. , Machi, H. , Iida, T. , & Ebisu, S. (2016). Temporal dynamics of bacterial microbiota in the human oral cavity determined using an in situ model of dental biofilms. NPJ Biofilms and Microbiomes, 2, 16018. 10.1038/npjbiofilms.2016.18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, Q. , Garrity, G. M. , Tiedje, J. M. , & Cole, J. R. (2007). Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Applied and Environmental Microbiology, 73(16), 5261–5267. 10.1128/AEM.00062-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, Z. , Wu, X. , Peng, J. , Hu, Y. , Fang, B. , & Huang, S. (2014). Artificially constructed quorum‐sensing circuits are used for subtle control of bacterial population density. PLoS One, 9(8), e104578. 10.1371/journal.pone.0104578 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Washio, J. , Ogawa, T. , Suzuki, K. , Tsukiboshi, Y. , Watanabe, M. , & Takahashi, N. (2016). Amino acid composition and amino acid‐metabolic network in supragingival plaque. Biomedical Research, 37(4), 251–257. 10.2220/biomedres.37.251 [DOI] [PubMed] [Google Scholar]
- Ximenes, M. , Armas, R. D. , Triches, T. C. , Cardoso, M. , & Vieira, R. S. (2018). Structure of the bacterial community in different stages of early childhood caries. Oral Health & Preventive Dentistry, 16(4), 369–373. 10.3290/j.ohpd.a39691 [DOI] [PubMed] [Google Scholar]
- Yu, O. Y. , Zhao, I. S. , Mei, M. L. , Lo, E. C. , & Chu, C. H. (2017). Dental biofilm and laboratory microbial culture models for cariology research. Dentistry Journal (Basel), 5(2), 21. 10.3390/dj5020021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaura, E. , Keijser, B. J. , Huse, S. M. , & Crielaard, W. (2009). Defining the healthy “core microbiome” of oral microbial communities. BMC Microbiology, 9, 259. 10.1186/1471-2180-9-259 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting information
Figure s1
Data Availability Statement
Metabolomics data have been deposited to the EMBL‐EBI MetaboLights database (DOI: 10.1093/nar/gkz1019, PMID:31691833) with the identifier MTBLS3982. The complete dataset could be accessed at “https://www.ebi.ac.uk/metabolights/MTBLS3982”. The metagenome data have been deposited in the SRA with accession number: PRJNA792086. The complete dataset could be accessed at https://www.ncbi.nlm.nih.gov/sra/PRJNA792086.